Back to Journals » Journal of Inflammation Research » Volume 15
Special Focus on the Cellular Anti-Inflammatory Effects of Several Micro-Immunotherapy Formulations: Considerations Regarding Intestinal-, Immune-Axis-Related- and Neuronal-Inflammation Contexts
Received 19 September 2022
Accepted for publication 24 November 2022
Published 13 December 2022 Volume 2022:15 Pages 6695—6717
DOI https://doi.org/10.2147/JIR.S389614
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Camille Jacques,* Ilaria Floris*
Preclinical Research Department, Labo’Life France, Nantes, France
*These authors contributed equally to this work
Correspondence: Camille Jacques, Preclinical Research Department, Labo’Life France, 1 rue François Bruneau, 44000 Nantes, France, Tel +33228444905, Email [email protected]
Introduction: Chronic inflammation is a pernicious underlying status, well-known for its contribution to the progressive development of various diseases. In this regard, Micro-immunotherapy (MI) might be a promising therapeutic strategy. MI employs low doses (LD) and ultra-low doses (ULD) of immune regulators in their formulations. In particular, as both IL-1β and TNF-α are often used at ULD in MI medicines (MIM), a special emphasis has been made on formulations that include these factors in their compositions.
Methods: Several in vitro models have been employed in order to assess the effects of two unitary MIM consisting of ULD of IL-1β and TNF-α (u-MIM-1 and u-MIM-2, respectively), and four complex MIM (c-MIM-1, − 2, − 3 and − 4) characterized by the presence of ULD of IL-1β and TNF-α amongst other factors. Thus, we first investigated the anti-inflammatory effects of u-MIM-1 and u-MIM-2 in a model of inflamed colon carcinoma cells. In addition, the anti-inflammatory potential of c-MIM-1, − 2, − 3 and − 4, was assessed in in vitro models of intestinal and neuronal inflammation.
Results: The results revealed that u-MIM-1 and u-MIM-2 both induced a slight decrease in the levels of IL-1β and TNF-α transcripts. Regarding the c-MIMs’ effects, c-MIM-1 displayed the capability to restore the altered transepithelial electrical resistance in inflamed-HCoEpiC cells. Moreover, c-MIM-1 also slightly increased the expression of the junction-related protein claudin-1, both at the mRNA and protein levels. In addition, our in vitro investigations on c-MIM-2 and c-MIM-3 revealed their immune-modulatory effects in LPS-inflamed human monocytes, macrophages, and granulocytes, on the secretion of cytokines such as TNF-α, PGE2, and IL-6. Finally, c-MIM-4 restored the cell viability of LPS/IFN-γ-inflamed rat cortical neurons, while reducing the secretion of TNF-α in rat glial cells.
Discussion: Our results shed the light on the potential role of these MIM formulations in managing several chronic inflammation-related conditions.
Keywords: cytokines, low-doses, ultra-low-doses, inflammation, immunotherapy, in vitro
Introduction
Inflammatory mechanisms are part of the normal body’s defense against aggressions, however, when they last for prolonged periods of time (months to years), they can induce a chronic inflammatory state that can be detrimental to the organism.1 The etiology of such slow and long-lasting inflammation is very heterogeneous, as multiple causes can trigger an over-activation of the immune system, inhibiting at the same time its normal return to homeostasis. Besides the context of auto-immune disorders, some of the reasons that can perpetuate a low-grade inflammation could be: (i) the organism’s inability to eliminate infectious agents such as bacteria, viruses or parasites, (ii) its prolonged exposure to low levels of chemicals, biochemicals or irritants, (iii) defects in the activation/inactivation mechanisms of the immune cells including an inadequate or insufficient resolution of inflammation or (iv), lifestyle stressors such as poor sleep quality or nutrition. If those triggers are maintained over a long period of time, they can lead to chronic inflammatory-related diseases that can involve many other organs and tissue of the body like the gut, driving inflammatory bowel diseases (IBD); the cardio-vascular system (stroke, atherosclerosis); the brain (depression, neurodegenerative diseases); the immune system (auto-immune diseases and cancer); the endocrine-metabolic system (diabetes and obesity).
Numerous treatment strategies have been employed in order to reduce inflammation and immunotherapies are one of them. For instance, the use of cytokine-based therapies has been reported in the management of sepsis, diabetes, or cardiovascular diseases with quite promising results. However, unwanted side effects still remain, probably due to the cytokines or immune factors dosages employed. A therapeutic approach that includes micro-immunotherapy (MI) could thus be of great interest in order to face such concerns. Indeed, MI medicines (MIM) are immune-factor-based medicines, which formulations either encompass a sole active substance (unitary preparations; referred to as u-MIM) or a distinctive combination of several ingredients (complex MIM; referred to as c-MIM). These medicines are manufactured in the form of sucrose-lactose pillules impregnated with the corresponding active ingredients, for oromucosal administration, and are intended to be taken in a fasted state. In this manner, the active substances can reach the oral-pharyngeal mucosal immune system,2 as well as the mucosal immune system of the gastrointestinal (GI) tract, with their associated lymphoid tissues and cell populations of the innate and adaptative immune responses.3 This route of administration will, in turn, elicit systemic and local effects, as we could observe in previous published preclinical in vivo studies.4–6
Through their exclusive preparation, MIM aim at gently modulating the expression and the related signaling pathways of cytokines and immune factors by providing to the organism these molecules at low doses (LD) and ultra-low doses (ULD) (Figure 1). These LD/ULD are expressed in Hahnemannian dilution (CH) or in Korsakovian dilutions (K). LD of bioactive molecules (ranging from 3 to 5 CH) are used in MI formulations to mimic the low doses occurring in the organism,7,8 and are employed to induce their own biological effects, stimulating their functions (Figure 1, green part). The recently published preclinical study on the unitary interferon-γ (4 CH) indeed shows how, at cellular levels, the drug elicits immunostimulatory and co-stimulatory effects on different cell types.7 On the other side, the use of ULD (from 6 CH up to 30 CH) employed in MI is intended to modulate (from 6 CH) and/or down-regulate (from 12 CH and beyond) the expression of the concerned active substances (Figure 1, orange part). Interestingly, we previously reported that the two unitary medicines (u-MIM) interleukin (IL) 1β (IL-1β) (27 CH) and tumor necrosis factor α (TNF-α) (27 CH) displayed anti-inflammatory properties, as they both exhibit the potential to inhibit the secretion of IL-1β and TNF-α, in a model of human primary monocytes as well as in THP-1 cells, after lipopolysaccharide (LPS) exposure (Figure 2A, blue box).9 Moreover, other in vitro and in vivo preclinical data have demonstrated the anti-inflammatory properties of a MI formulation employing ULD of TNF-α and IL-1β in association with other immune regulators,4,10,11 providing encouraging results about the potential of some MIM in slowing down the pro-inflammatory responses in the context of chronic inflammation. Based on these previous results, the first aim of this study was to assess the effects of these two ULD-based u-MIMs in another specific inflammatory context, an intestinal inflammation model (Figure 2A, red box).
 |
Figure 1 General scheme of the concept of low doses (LD) and ultra-low doses (ULD) in the context of MI supported by the two cited preclinical research studies referring to the immune-stimulatory/anti-inflammatory responses elicited by the LD or ULD, respectively. LD of cytokines, ranging from 3 centesimal Hahnemannian (CH) to 5 CH, aim at orienting the immune responses towards an immune stimulation, inducing their own biological effect through mimicking the physiological low doses employed in the body. The immune-stimulatory effects of IFN-γ (4 CH) have been reported by Jacques et al 2022.7 On the other side of the spectrum, ULD aim at modulating (from 6 CH)/down-regulating (from 12 CH and beyond) a response. For instance, the pro-inflammatory cytokines IL-1β and TNF-α employed at 27 CH, exerted anti-inflammatory responses (Floris et al 2020).9 The CH numbers represented on the x-axis are the ones employed in the MIM’s formulations studied in the current manuscript. The arrows represent the direction of the oriented biological response mediated by the LD/ULD employed in micro-immunotherapy medicines (MIM). Green arrow, Stimulation of a biological response; Red arrow, Inhibition of a biological response; Combined Green and Red arrow, Modulation of a biological response. |
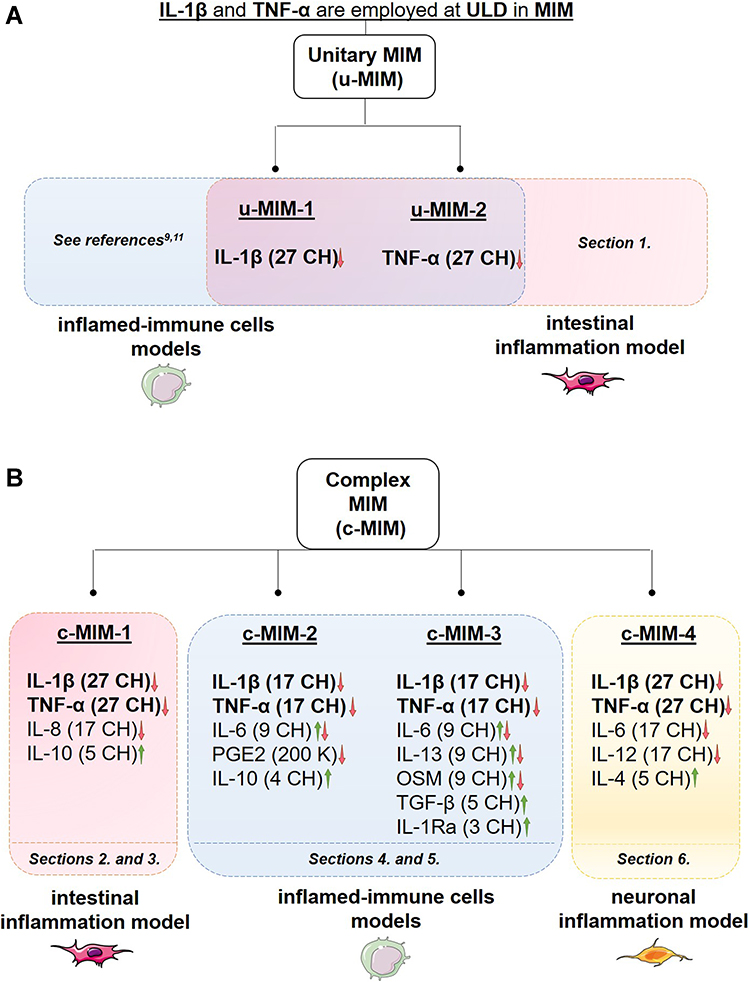 |
Figure 2 (A) The rationale of the present study is based on the fact that ULD of IL-1β and TNF-α are employed in the tested MI medicines (MIM). The anti-inflammatory role of these cytokines, when used at 27 CH, under the respective form of unitary MIM (u-MIM) has been reported by Floris et al 2020,9 and Jacques et al 2021,11 in a model of inflamed-immune cells (blue box) consisting of LPS-inflamed human primary monocytes and THP-1 cells. In the current study, the effects of these u-MIM were appraised in an intestinal model of inflammation (red box; see Results Section 1., for the results). (B) Scheme recapitulating the overall organization of the present manuscript regarding the four complex-MIM (c-MIM) studied. The formulation of each c-MIM is detailed. As each c-MIM formulation was tested in a particular context, the corresponding Results’ sections within this manuscript are reported at the bottom of each colored box (red box, intestinal inflammation model; blue box, inflamed-immune-cells model; yellow box, neuronal inflammation model). The arrows represent the direction of the oriented biological response mediated by the LD/ULD employed in micro-immunotherapy medicines (MIM). Green arrow, Stimulation of a biological response; Red arrow, Modulation/inhibition of a biological response; Combined Green and Red arrow, Modulation of a biological response; K, Korsakovian dilution. |
In contrast to unitary preparations, complex MI formulations consist of a unique association of active substances, thought to act synergistically in order to coherently channel a specific biological response. The second aim of this study was to conduct a complementary assessment of the effects of four c-MIM (referred to as c-MIM-1, c-MIM-2, c-MIM-3 and c-MIM-4 in this manuscript), as they all employ ULD of IL-1β and TNF-α in their formulations, in association with additional actives at LD/ULD. c-MIM-1 also includes ULD of IL-8 and LD of IL-10; c-MIM-2 includes ULD of prostaglandin E2 (PGE2) and IL-6, with LD of IL-10, too; c-MIM-3 uses ULD of IL-6, IL-13 and oncostatin M (OSM), as well as LD of tumor growth factor β (TGF-β) and IL-1 receptor antagonist (IL-1Ra). Finally, c-MIM-4 employs ULD of IL-6 and IL-12 as well as LD of IL-4 (Figure 2B).
It is now well-recognized that chronic inflammation triggers a whole-body response and the activation of many feedback loops with interconnected effects.12 Moreover, as an integrated and complex system, the different anatomical parts of the body communicate between them. Considering the particularly close relationship between the epithelial cells from the gut, the immunity and the nervous system,13 the main purpose of this study was indeed to assess the potential anti-inflammatory properties of four different complex MIM’s (c-MIM) formulations in several inflammatory contexts related to the (i) -intestinal tract (Figure 2B, red box), (ii) -immune axis (Figure 2B, blue box) and (iii) -nervous system (Figure 2B, yellow box), depending on their specific cytokines combination. The further paragraphs of this introduction will provide brief non-exhaustive detail about the role of each cytokine in the three above-mentioned contexts, thus explaining the deliberate choice to assess each c-MIM in a particular inflammatory model in the first place.
Recent literature delineated the importance of IL-8 and IL-10 in the molecular mechanisms driving GI inflammatory diseases.14,15 The fact that c-MIM-1 includes ULD of IL-8 and LD of IL-10 in its formulation thus motivated the choice of assessing the cellular effects of this medicine in an in vitro model of intestinal inflammation (Figure 2B, red box).
In a more general approach, c-MIM-2 and c-MIM-3 have been investigated in vitro, on the immune axis in different cell types and inflamed experimental contexts (Figure 2B, blue box). These two formulations employ ULD of IL-1β and TNF-α in association with ULD of PGE2, or ULD of IL-6, IL-13, and OSM, and LD of IL-10, or LD of TGF-β and IL-1Ra. Numerous publications, nicely put together in the review from Tsuge et al indeed reported the major role played by PGE2 in the mediation of inflammation and immune diseases, notably through its implication in mechanisms related to Th1 differentiation, and Th1/Th17/Th22 signaling.16 In the context of immunity, IL-10 is produced by a broad type of myeloid and lymphoid cells during infections, and suppresses macrophages and dendritic cells functions, thereby impeding Th1 and Th2 effector responses.17 In immune effector cell types, and in a wide range of established models of chronic inflammation, IL-10 is reported to play a central role in the regulation of homeostasis.18 On another side, TGF-β is important in maintaining the T cell tolerance to self or innocuous environmental antigens.19 IL-1 is known for its crucial involvement in the onset and/or the etiopathogenesis of several inflammatory-mediated diseases. Given its importance, several therapeutic approaches consisting of the use of IL-Ra have been used in clinical practice to block the activity of IL-1.20 OSM is a pleiotropic cytokine playing important roles in a wide range of homeostatic functions (such as hematopoiesis, bone/fat turnover, liver regeneration) and, under chronic inflammatory diseases, it may play pro-inflammatory roles, depending on the microenvironment and the interested tissue, without being overexpressed at systemic levels.21 IL-6 is also a pleiotropic cytokine playing important roles in hematopoiesis, bone metabolism embryonic development, immunity, and host responses. Its expression is finely regulated at multiple levels, and when a dysregulated and persistent IL-6 expression occurs, it may lead to the development of various inflammatory-mediated diseases.22,23 Similarly, IL-13 is known to exert pleiotropic effects and to be involved in inflammatory-mediated diseases.24
Finally, in the case of c-MIM-4, which employs ULD of both IL-6 and IL-12 in its formulation, as well as IL-4 at LD, one of the goals of this study was to assess its effects using an in vitro preclinical model of neuronal inflammation (Figure 2B, yellow box). The rationale behind the choice of the neuronal-inflammation context was supported by a rich bibliography, illustrated by the following examples. For instance, according to the results of a recent meta-analysis, patients suffering from depressive symptoms express higher levels of IL-6.25 Another study also reported that the IL-12 serum levels of patients with major depression were significantly higher than healthy controls, and that an 8-weeks treatment led to a reduction of these levels.26 On the other hand, it was suggested that IL-4 could modulate IL-1β-induced depressive behavior in rats, by inhibiting IL-1β-induced central glial activation and neurotransmitter alterations.27
Materials and Methods
Tested Item and Experimental Control
The tested MIMs are homeopathic medicinal products consisting of sucrose-lactose pillules impregnated with ethanolic preparations of cytokines. These MIMs were manufactured by Labo’Life España, as previously described,6,7 and have been provided for investigational purposes. The two u-MIM under investigation in this study (referred as u-MIM-1 and u-MIM-2) are presented as unitary medicines, while the four tested complex capsules (c-MIM-1, c-MIM-2, c-MIM-3, and c-MIM-4) are made of a specific combination of either four, five or seven different ingredients (Figure 2). Their composition is characterized by cytokines and immune factors and their amount is expressed in CH or in K.
The Veh. pillules used in the study as controls are manufactured by Labo’Life Belgium. Previous publications described how these vehicle controls are produced, in order to provide a suitable control for preclinical research.4,6,7,9,10 Regarding the in vitro experiments performed in the current study, u-MIM, c-MIM or the Veh. pillules were freshly diluted in 100 mL of culture medium to reach the final sucrose-lactose concentration of 11 mM.
Cell Lines
HT-29- cells, a human colorectal adenocarcinoma cell line with epithelial morphology was used in this study. This cell line was purchased from the ATCC (ATCC® HTB-38™, Molsheim, France) and was grown in Dulbecco’s Modified Eagle’s Medium (DMEM) with high glucose (4.5 g/L) containing 10% fetal calf serum (FCS) (DMEM 10% FCS, PAA Laboratories, Les Mureaux, France) and antibiotics (streptomycin 100 μg/mL and penicillin 100 units/mL, Sigma Aldrich, St. Quentin Fallavier, France). Cells were incubated at 37°C in 5% CO2 and 95% relative humidity.
To induce inflammatory conditions, a pre-stimulation of cells by TNF-α (ref # 39–8329-60/E08250-1633, eBioscience) at 10 ng/mL was performed for 3 hours. The tested Veh. and MIMs were then added to the culture medium containing the TNF-α for 24 more hours before an extensive wash, and harvested for mRNA extraction. Supernatants (SN) were collected to perform cytokine quantification by ELISA.
Evaluation of Gene Expression by RT-qPCR
Total mRNA was isolated from cells using Rneasy kit (Macherey Nagel, Hoerdt, France) according to the manufacturer’s instructions. RNA quantification was performed using spectrophotometry. After treatment at 37°C for 30 min with 20–50 units of RNase-free DNase I (Roche Diagnostics Corporation, Indianapolis, IN, USA), oligo-dT primers (Roche Diagnostics Corporation, Indianapolis, USA), the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA) was used to synthesize single-stranded cDNA. The mRNA was quantified using SYBR green Master Mix (Applera, Courtaboeuf, France) with human-specific oligonucleotides in a GeneAmp Abiprism 7000 (Applera). In each assay, calibrated and no-template controls were included. Each sample was run in triplicate. SYBR green dye intensity was analyzed using the Abiprism 7000 SDS software (Applera, Courtaboeuf, France). All results were normalized to the unaffected housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
All the results are expressed as a fold of the mean of activation of duplicate/mean of duplicate in medium alone or TNF-α alone. The primers sequences used are the following: GAPDH: Forward (F): 5’-GAC ACC CAC TCC TCC ACC TTT-3’; Reverse (R): 5’-TTG CTG TAG CCA AAT TCG TTG T-3’; TNF-α: F:5’-CAG GAC GTG GAG CTG GCC GAG GAG −3’; R: 5’- CAC CAG CTG GTT ATC TCT CAG CTC −3’; IL-1β: F: 5’- GAT GCA CCT GTA CGA TCA CT-3’; R: 5’-GAT GCA CCT GTA CGA TCA CT −3’.
Inflamed Colon Epithelial Intestinal Cells Model
Phytohemagglutinin-Stimulated Human Peripheral Blood Mononuclear Cell’ Supernatants Production
Human peripheral blood mononuclear cells (PBMC) from healthy adult volunteer donors were isolated by centrifugation of venous blood on Ficoll-Hypaque density gradients (Amersham Pharmacia Biotech, Piscataway, NJ, USA). Healthy volunteers were enrolled by the Blood Bank Center (Etablissement Français du Sang, EFS, Pays de Loire, France). All blood samples were approved by the Ethics committee of the EFS Blood Bank Center with written informed consent obtained for all the donors, in accordance with the Declaration of Helsinki. Cells (105/mL) were cultured in 24-well round-bottom plates in 2 mL of complete medium and stimulated with phytohemagglutinin (PHA) (5 µg/mL) for 48 hours. The SN were collected and stored at −80°C until use.
Trans-Epithelial Resistance Assessment in Human Colonic Epithelial Cells
Human Colonic Epithelial Cells (HCoEpiC) were purchased from Innoprot, Bizkaia, Spain. The cells were seeded at the density of 5×105 cells in 24-well plate inserts whose diameter is 6.5 mm and the membrane area 0.3 cm2 and grown until tight confluence. The cells were cultured in DMEM supplemented with 10% FCS, 2 mM L-glutamine, and antibiotics. The trans-epithelial resistance (TEER) measurements were obtained as Ω/cm2 and performed with a Millicell® ERS-2 device at different incubation times (24 hours and 72 hours), in the presence/absence of the PBMC SN, with/without c-MIM-4 or Veh. added 30 minutes before reading. The measurements were performed in HBSS without phenol red (Sigma-Aldrich, ref # H6648), adding 600 µL HBSS in the basolateral area and 100 µL of HBSS in the apical area for 30 minutes. After this time, readings in 3 individual wells (n = 3) were made. 10 mM of sodium caprate (C10) (Sigma-Aldrich, ref # C4151) was used as a positive control.
Claudin-1 Expression Assessment in Human Colonic Epithelial Cells
HCoEpiC cells were seeded at a density of 500.000 cells/well in 6 well plates and grown until confluence. HCoEpiC cells were treated with or without Veh. or c-MIM-1 for 24 h. After that, 400 μL of human PBMC SN previously activated with PHA during 48 h was added to each well for 30 minutes. After treatments, the cells were collected and divided into two parts, one part was used for mRNA isolation and the other part for protein extraction.
Evaluation of the Claudin-1 Expression by Real-Time qPCR
For quantitative reverse transcriptase-PCR assays, total RNA (1 µg) was retrotranscribed using the iScript cDNA Synthesis Kit (Ref #1708891, Bio-Rad, Hercules, CA, USA) and the cDNA was analyzed by real-time PCR using the iQTM SYBR Green Supermix (Ref #1708880, Bio-Rad) and a CFX96 Real-time PCR Detection System (Bio-Rad). GAPDH gene expression was used to standardize mRNA expression in each sample. Gene expression is quantified using the 2−ΔΔCt method and the percentage of relative expression against controls (untreated cells) was represented. Primers for use in real-time PCR analysis: Claudin-1: F: 5’-AGG TCT GGC GAC ATT AGT GG-3’; R: 5’-CGT GGT GTT GGG TAA GAG GT-3’; GAPDH: F: 3’-TGG CAA AGT GGA GAT TGT TGC C-3’; R: 5’-AAG ATG GTG ATG GGC TTC CCG-3’.
Evaluation of the Claudin-1 Expression by Western Blot
The cells were washed with phosphate-buffered saline (PBS) and proteins were extracted in 50 μL of lysis buffer (50 mM Tris–HCl pH 7.5, 150 mM NaCl, 10% glycerol, and 1% NP-40) supplemented with 10 mM NaF, 1 mM Na3VO4, 10 μg/mL leupeptin, 1 μg/mL pepstatin and aprotinin, and 1 μL/mL phenylmethyl sulfonyl fluoride (PMSF) saturated. Twenty micrograms of protein were boiled at 95°C in Laemmli buffer and electrophoresed in 10% SDS/PAGE gels. Separated proteins were transferred to polyvinylidene fluoride (PVDF) membranes (20V for 30 min) and blocked in tris-buffered saline (TBS) solution containing 0.1% Tween 20 and 5% non-fat dry milk for 1 hour at room temperature. Immunodetection of specific proteins was carried out by incubation with a primary antibody rabbit anti-Cldn-1 (1:200, Ref: #ab15098, Abcam, Cambridge, UK) and β-actin (1:10.000 dilution, Ref: #A5316, Sigma), overnight, at 4°C. After washing membranes, horseradish peroxidase-conjugated secondary antibody was added and detected by a chemiluminescence system (GE Healthcare Europe GmbH, Freiburg, Germany). The quantification of the Blot’s pictures, was performed thanks to the online version of the ImageJ software (downloaded on the 2nd of May 2022).
Immunomodulation in Immune Cells
Immunomodulation in Human Monocytes
Human primary monocytes were prepared from buffy coats of three healthy human blood donors. All blood samples were approved by the Ethics committee of the EFS Blood Bank Center with written informed consent obtained for all the donors. Cells were seeded in 24-well plates at a density of 2.200.000 cells/well for ELISA measurements. Monocytes were incubated with LPS (10 ng/mL, Salmonella typhimurium SL1181) for 30 min before treatment with either the Veh., or c-MIM-2 for an additional 24 hours. After 24 hours, SN were collected, centrifuged, and investigated for IL-1β, TNF-α and PGE2 concentrations in ELISA using the manufacturer’s protocol (n = 6, 3 different donors).
Immunomodulation in Human Macrophages
Human CD14+ monocytes were isolated from the blood of a healthy donor (67 years old man). The blood sample collection was approved by the Ethics committee of the EFS Blood Bank Center. After the mononuclear cells’ isolation by the Ficoll gradient centrifugation method, the CD14+ cells were selected by positive selection thanks to magnetic beads (MACS® Miltenyi Biotec B.V. and Co. KG, Bergisch Gladbach, Germany). Monocytes were then cultivated for 6 days in 24-well plates, with α-MEM medium supplemented with 10% fetal bovine serum (FBS) and 100 ng/mL macrophage colony-stimulating factor (M-CSF) (R&D Systems, Inc., Minneapolis, MN, USA, Ref: #216-MC). After these 6 days in culture, the medium was replaced with α-MEM supplemented with 10% FBS and 50 ng/mL M-CSF in addition to 100 ng/mL LPS (Sigma, Ref: #L4391) and either the Veh. or c-MIM-3. The treatments were applied for 24 hours and each condition was performed in triplicate. At the end of the incubation period, SN were collected, aliquoted, and stored at −80°C until ELISA assay (human IL-6 Quantikine, R&D Systems). Optic density was measured on a spectrophotometer Multiskan Go and the Skanlt 3.2 software was used to quantify the measured cytokine concentrations.
Immunomodulation in Human Granulocytes
Human primary granulocytes were freshly isolated from the blood of a healthy donor obtained from the EFS, and seeded in 48-well plates in optimized RPMI 1640 supplemented with 10% inactivated FBS. Cells were then pre-incubated with either Veh., or c-MIM-3 for one hour, before the addition of LPS (1 µg/mL) as the inflammation inducer within the media. Cells were then incubated for 18 hours in presence of the tested items and LPS. All the experimental conditions were performed in triplicate. At the end of the total incubation period, SN were collected and the assessed cytokines levels were evaluated. The IL-6 and IL-1β levels were evaluated by multiplex CBATM method following the provider recommendations (Human IL-6 Flex Set; BDTM Biosciences, A7 beads, Ref: #558276; dilution factor 1:5 and Human IL-1β Flex Set; BDTM Biosciences, B4 beads, Ref: #558279).
Immunomodulation in Neurons
Ethics Statement
All animal procedures were carried out in strict adherence to the European Community Council Directive of September 22, 2010 (2010/63/UE). All experiments and protocols were authorized and approved by the French Ministry of Research, as well as by the Animal Welfare Committees of Amylgen, Biotrial, QPS and Neuronexperts. All efforts were made to minimize the number of animals used.
Cortical Neurons Culture
Rat cortical neurons were cultured as described by Singer et al28 Briefly, a pregnant female rat of 15 days gestation was sacrificed by CO2 inhalation (Wistar; Janvier Lab, Le Genest-Saint-Isle, France) and the fetuses were removed from the uterus. The cortexes were removed and placed in ice-cold medium of Leibovitz (L15, Panbiotech GmbH, Aidenbach, Germany, Ref: #P04-27055 batch: 8940719) containing 2% of penicillin/streptomycin (PS, PanBiotech Ref: #P06-07100, batch: 2791019) and bovine serum albumin (BSA, Panbiotech Ref: #P06-1391100, batch: H190110). Cortexes were dissociated by trypsin-EDTA (Thermo Fisher Scientific, Illkirch-Graffenstaden, France Ref: #25300-054, batch: 2027082) for 20 min at 37°C. The reaction was stopped by the addition of DMEM (Panbiotech Ref: #P04-03600, batch:1220919) containing DNAase I grade II (0.1 mg/mL; PanBiotech, Ref: #P60-37780100, batch: H181015) and 10% FCS (Invitrogen, Waltham, MA, USA, Ref: #10270106, batch: 42G2082K). Cells were then mechanically dissociated by 3 passages through a 10 mL pipette. Cells were then centrifuged at 290 × g for 10 min at 4°C. The SN was discarded and the cell pellet was re-suspended in a culture medium for neurons consisting of Neurobasal (Invitrogen, Ref: #11570556, batch: 2161553) supplemented with 2% of B27 (Invitrogen, Ref: #11530536, batch: 2181375), 2 mM of L-glutamine (L-glut; PanBiotech, Ref: #P04-80100, batch: 1640518), 2% of PS solution, 1% of FCS and 10 ng/mL of brain-derived neurotrophic factor (BDNF, Peprotech France, Neuilly-sur-Seine, France, Ref #450-02, batch 31961). Viable cells were counted in a Neubauer cytometer using the trypan blue exclusion test. The cells were seeded at a density of 30.000 cells/well in 96 well-plates (Falcon, ref: 353072) pre-coated with poly-D-lysine (10 µg/mL; Sigma, ref: P7886, batch: SLBS8705) in H2O culture (PanBiotech ref: P04-991000, batch: 2900219) and were cultured at 37°C in a humidified air (95%)/CO2 (5%) atmosphere.
Preparation of Astrocytes and Microglial Cell and Culture with Cortical Neurons
One vial of rat glial cells (4.000.000 cells by vial at passage 2) was thawed and cells were seeded at the density of 12.000 cells per cm² on 75 cm² flask (Nunc ref: 055403) in DMEM supplemented with 2 mM of L-glutamine, 2% of PS solution and 10% of FCS. After 5 days of culture, the mix of astrocytes/microglial cells was dissociated by trypsin-EDTA for 20 min at 37°C. The reaction was stopped by the addition of DMEM, containing DNAase I grade II (0.1 mg/mL) and 10% FCS. Cells were then centrifuged at 290 × g for 10 min at 4°C temperature. The cell pellet was re-suspended in a culture medium for neurons. Glial cells were seeded at the same time as the neurons at a density of 20.000 cells per well.
Compound Incubation and LPS/IFN-γ Treatment
After 8 days of coculture, cells were treated with either Veh., c-MIM-2, or MK801 (Sigma ref: M107, batch: 105M4606V) at 20 µM in medium for neurons. After 9 days of coculture, cells were treated with or without the combination of LPS (100 ng/mL, Sigma ref: L2630, batch: 089M4016V)/IFN-γ (10 ng/mL, Peprotech ref: 315–05, batch: 061798) during 24 hours,29–31 and with or without the tested items or MK801 in 300 µL of medium for neuron. At the end of this 24 hours incubation, SN were recovered and frozen for quantification of TNF-α released. Cortical neurons and glial cells were incubated in a medium for neurons with and without the tested items for an additional 48 hours to analyze the survival of neurons. Six independent wells were done per culture condition.
Quantification of Neuronal Cells Survival After Glial Cells LPS/INF-γ Activation
After 72 hours of glial cells LPS/INF-γ activation, cells were fixed by a solution of 4% paraformaldehyde (Sigma, ref: 6148, batch: SZBE2390V) for 20 min at room temperature, and control conditions were also fixed according to the same procedure. The cells were then permeabilized and non-specific sites were blocked with a solution of PBS (PanBiotech; ref: P04-36500, batch: 5081219) containing 0.1% of saponin (Sigma; ref: S7900, batch: BCBL8667V) and 1% FCS for 15 min at room temperature. Cells were incubated overnight at 4°C, with a monoclonal anti-microtubule associated protein 2 antibodies produced in mice (MAP-2, 1/5000, Sigma; ref: M4403, batch: 063M4802), in a solution of PBS containing 1% FCS and 0.1% saponin. Antibody against MAP-2 stains specifically neuron cell body and neurite. This antibody was revealed with an Alexa Fluor 488 goat anti-mouse IgG (1/400, Molecular probe, ref: A11001, batch: 2140660) in PBS with 1% FCS and 0.1% saponin, for 1 hour, at room temperature. Nuclei of cells were labeled by a fluorescent marker (Hoechst solution, Sigma; ref: B1155, batch: 046M4048V) in the same solution. For each condition, 20 pictures per well were taken using InCell Analyzer 2200 (GE Healthcare) with 20x magnification. Images of each culture well were taken in the same conditions. Analysis of cell bodies of MAP-2 positive neurons was performed using Developer software (GE Healthcare). A total of 6 data per experimental condition were provided. All values are expressed as mean ± s.e.m.
Quantification of Cytokine Release by ELISA
The culture SN were thawed and 100 µL by well were used to quantified by ELISA the level of TNF-α (TNF-α rat ELISA kit, Fisher Scientific, ref: KRC3011, batch 218247–002) released by glial cells in the media, according to the manufacturer’s instructions. All values are expressed as mean ± SD.
Statistical Analysis
The graphs in the figures were performed with GraphPad Prism, Version 9.4 for Windows (GraphPad Software Inc., San Diego California USA, updated on 16/06/2022). Authors have followed the recent recommendations of D.L. Vaux that encourage performing descriptive statistics instead to make statistical inferences when the number of independent values is small.32 Indeed, no statistical inference has been performed to analyze the results of the in vitro studies presented here.
Results
IL-1β (27 CH) and TNF-α (27 CH), Both Unitary Micro-Immunotherapy Medicines, Slightly Reduced the Expression of IL-1β and TNF-α at Transcriptional Levels in TNF-α-Inflamed HT-29 Cells
Previous results reported the anti-inflammatory effects of the u-MIM IL-1β (27 CH) and TNF-α (27 CH) in human primary monocytes and in the THP-1 cell line.9 In this study, we wanted to assess the effect of these unitaries in a model of intestinal inflammation, by using a human epithelial colon cancer cell line exposed to TNF-α. Therefore, the effect of IL-1β (27 CH) and TNF-α (27 CH) was evaluated in HT-29 cells, inflamed by a 3 hours pre-incubation with 10 ng/mL TNF-α (Figure 3A). The cells were then treated with either the vehicle (Veh.), IL-1β (27 CH) or TNF-α (27 CH) for 24 hours before assessing the transcripts’ expression of IL-1β and TNF-α by RT-qPCR. The results are presented in Figure 3B and C, and the control (Ct) conditions showed that the mRNA expression of both of the cytokines was increased after TNF-α stimulation (Figure 3, white vs red histograms). A decrease of more than two-fold in IL-1β expression was obtained after IL-1β (27 CH) treatment compared with the Veh. (Figure 3, grey vs orange histograms). In addition, a decrease in TNF-α expression of about two-fold too was also observed in the same treatment conditions. Similar response patterns towards a down-regulation were observed after treatment with the other u-MIM, TNF-α (27 CH) (Figure 3, grey vs yellow histograms). Considering these interesting trends, the following sections of this study aimed at evaluating the anti-inflammatory properties of several complex MIM-containing ULD of IL-1β and TNF-α within their formulations.
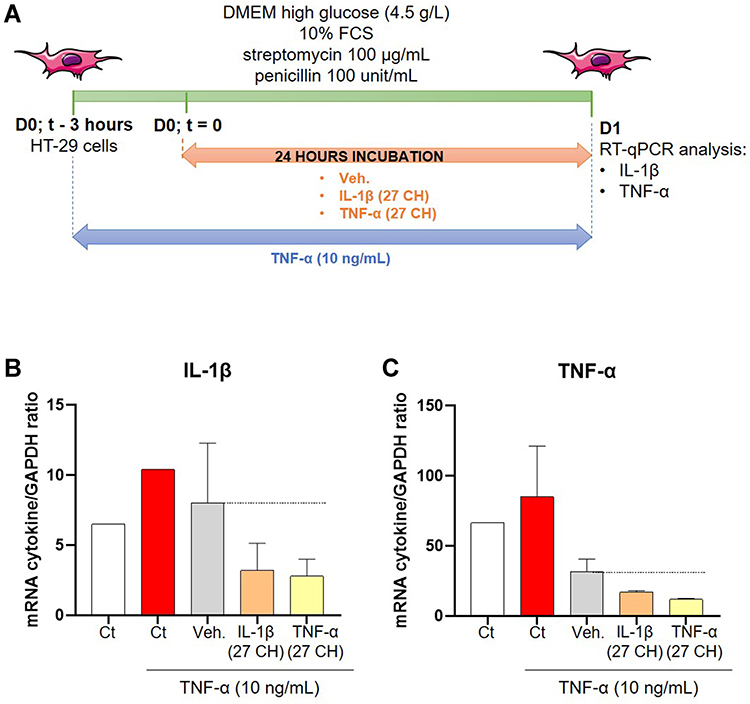 |
Figure 3 IL-1β (27 CH) and TNF-α (27 CH) tend to decrease the expression of both IL-1β and TNF-α at transcriptional levels, in a model of intestinal inflammation. (A) Representative scheme of the experimental protocol. HT-29 cells were pre-treated for 3 hours with 10 ng/mL TNF-α as an inflammatory trigger before the incubation with either the Veh., IL-1β (27 CH), or TNF-α (27 CH) during the next 24 hours. RT-qPCR analysis was performed at the end of the incubation period. (B and C) Expression of IL-1β and TNF-α at the transcriptional level, in presence of TNF-α (10 ng/mL) as an inflammation inducer and after a 24 hours incubation with either Veh., IL-1β (27 CH) or TNF-α (27 CH). The experiment has been done once and the means ± SD of three technical replicates are illustrated on the graph. The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal normalizer. The black dotted lines highlight the effect of the two tested u-MIM compared with the Veh. |
c-MIM-1 Could Restore the Transepithelial Resistance in a Cellular Model of Intestinal Inflammation
The integrity of the intestinal barrier is fundamental to maintain healthy gut homeostasis, as well as physiological intestinal permeability. It is widely known that, under chronic inflammation, intestinal homeostasis is altered, resulting in an impairment of the barrier functions and thus, an increase in intestinal permeability.33 In order to induce an intestinal inflammation status in vitro, human HCoEpiC cells were exposed to supernatants (SN) derived from phytohemagglutinin (PHA)-treated PBMCs, which have been reported to be concentrated in pro-inflammatory cytokines such as TNF-α, IL-8 and IL-17.34 The resistance of the epithelial barrier is one of the characteristics that reflects the intestinal permeability and the interactions’ strength between epithelial cells. As a consequence, any disruption within these epithelial interactions ultimately creates a so-called “leakage” in the barrier, leading to the symptoms related to inflammatory bowel diseases. Transepithelial electrical resistance (TEER) measurements, expressed in Ω/cm,2 were performed with a Millicell® ERS-2 device, after either a 24-hours or a 72-hours incubation period, with/without PHA-treated-PBMCs SN, with/without either the Veh. or c-MIM-1, added 30 minutes before the SN (Figure 4A). 10 mM of sodium caprate (C10) was used as a positive inducer of the epithelial barrier disruption and served as an internal control attesting the relevance of the model. The results in presence of PHA-treated PBMCs SN incubated for 24 and 72 hours are presented in Figure 4B and C, respectively.
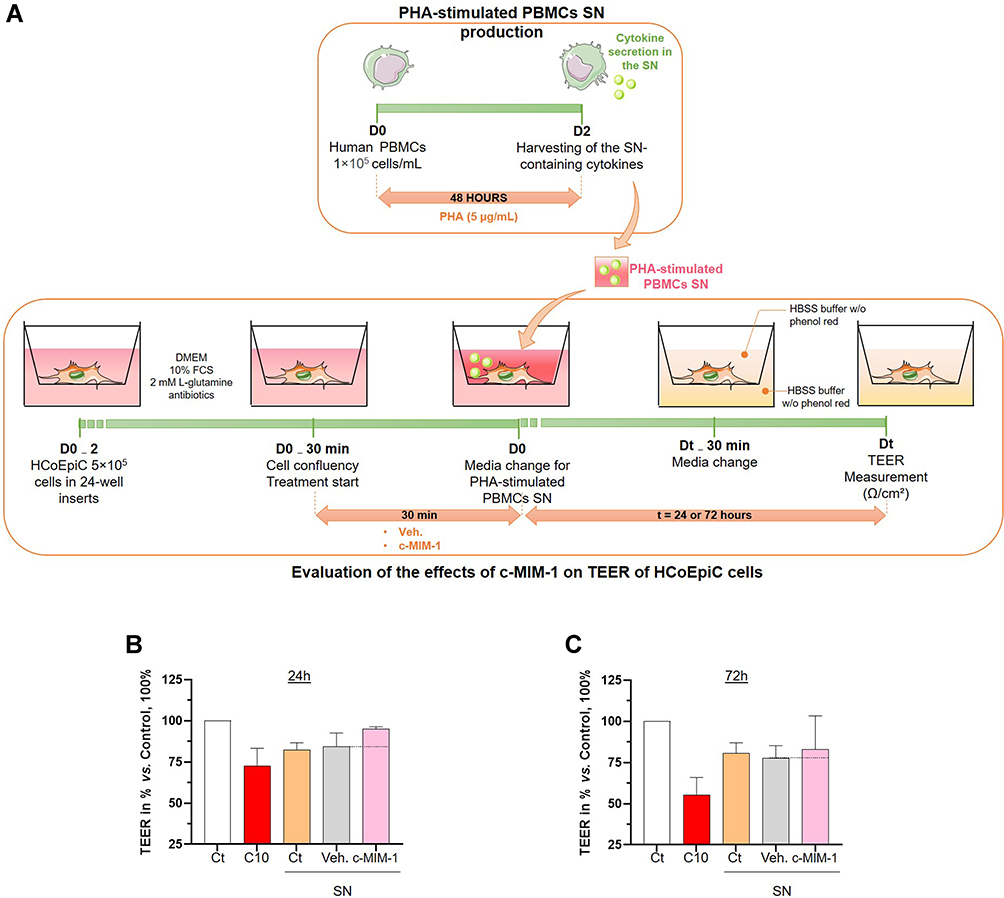 |
Figure 4 C-MIM-1 displays a protective effect against intestinal permeability in a model of intestinal cells inflamed with phytohemagglutinin (PHA)-stimulated PBMCs supernatants (SN). (A) Representative scheme of the experimental protocol. In the upper panel, the production of the PHA-stimulated peripheral blood mononuclear cells (PBMCs) supernatants (SN) is illustrated. The lower panel shows the evaluation of the effect of c-MIM-1 on the transepithelial electrical resistance (TEER) in a model of HCoEpiC cells. FCS, fetal calf serum; HBSS, Hanks’ balanced salt solution. Briefly, PBMCs from healthy adult volunteers’ donors were isolated by Ficoll-Hypaque density gradients and the cells were seeded in 24-well round-bottom plates and stimulated with PHA (5 µg/mL) for 48 hours. At the end of the incubation period, the SN containing pro-inflammatory cytokines were harvested. In parallel, HCoEpiC cells were grown in 24-well plate inserts until tight confluence. Cells were either not treated (Ct), treated with Veh. or treated with c-MIM-1 for 30 min before media replacement with either regular DMEM media or PHA-stimulated PBMCs SN during the next 24 hours or 72 hours. 30 minutes before the TEER measurement, the media was replaced with HBSS without phenol red alone in the basolateral area, in the apical chamber. Each condition has been measured in 3 individual wells (n = 3) and a condition in which C10 alone was incubated was used as a positive control for the induction of the epithelial disruption. (B and C) Effect of c-MIM-1 on HCoEpiC TEER after 24 or 72 hours of stimulation with PHA-induced PBMCs SN. Raw data were provided as the measures of the Ω/cm² for each triplicate, and were then expressed as percentages of the control conditions, set as 100%. Each histogram illustrating the mean ± SD obtained for each assessed condition, in 3 independent experiments. The black dotted lines highlight the effect of c-MIM-1 compared with Veh. |
As expected, the C10 stimulation alone decreased the TEER compared with the untreated control (Figure 4B and C, red histograms vs white histograms). Similarly, SN from PBMCs activated with PHA, also led to a decrease in TEER compared with the untreated HCoEpiC cells (Figure 4B and C, orange histograms vs white histograms), thus confirming here the relevance of these SN as a pro-inflammatory stimulus.
Interestingly, c-MIM-1 protected the epithelial cells against the PHA-stimulated PBMCs SN-induced disruption (Figure 4B, pink histogram vs yellow histogram), after 24 hours. Regarding the TEER results obtained at 72 hours, c-MIM-1 effects appear to be transient, as these protective effects, almost disappeared (Figure 4C pink histogram vs yellow histogram). The results derived from three independent experiments provide for the first time, indications of a potential effect of the c-MIM-1 formulation towards the protection of the epithelial barrier in an inflammatory context in vitro.
The effect of c-MIM-1 was assessed in human HCoEpiC cells in absence of inflammation, too. As shown in Supplementary Figure S1, c-MIM-1 does not alter the barrier permeability compared to the control (Ct), rather it seems that c-MIM-1 has restored the resistance and has strengthened the epithelial barrier at 24 hours in comparison with the Veh., which has induced a small TEER decrease.
c-MIM-1 Slightly Increased Claudin-1 Expression in a Cellular Model of Intestinal Inflammation
In light of the previous encouraging results, we wanted to investigate if the tested medicine could modulate the expression of one of the proteins present within the tight junctions, involved in the regulation of the epithelial barrier permeability. Indeed, claudin-1 is widely expressed in the intestinal epithelium and plays an important role in maintaining intestinal barrier integrity.35 Briefly, HCoEpiC cells were treated with either the Veh. or c-MIM-1 for 24 hours, and SN from PHA-treated PBMCs were then added for an additional 30 minutes incubation. The cells were then harvested and mRNA and protein samples were extracted for claudin-1 expression analysis. The results of the RT-qPCR are presented in Figure 5A and the results from the Western Blots are illustrated in Figure 5B. Overall, in inflammatory conditions, the Veh. did not modulate the claudin-1 expression at the mRNA level, compared with the SN from the PHA-treated PBMCs condition. However, when the cells were treated with c-MIM-1, a slight increase (of about 20%) in the claudin-1 expression was observed. In addition, according to the Western Blot quantization, this trend tended to be confirmed at the protein level, in a similar range of increase. Such results were also observed without any inflammatory stimulus (Supplementary Figure S2).
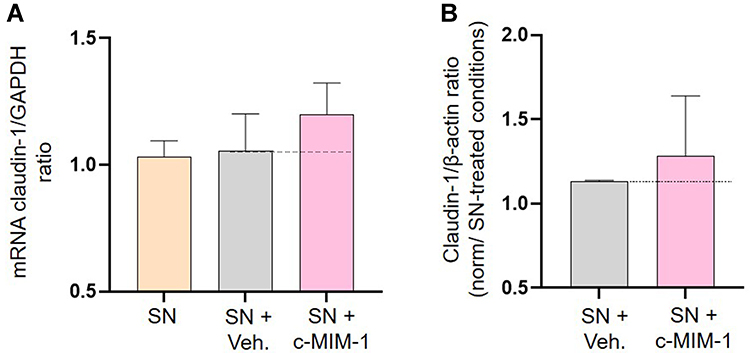 |
Figure 5 C-MIM-1 slightly increases the expression of claudin-1 in a cellular model of intestinal inflammation. (A) The expression of claudin-1 was assessed by RT-qPCR in the HCoEpiC cells, after 24 hours of treatment with either the Veh. or c-MIM-1 and stimulation with the supernatant (SN) from PBMCs treated with PHA (5µg/mL) for 30 min. The claudin-1 expression has been normalized by the one of the housekeeping gene GAPDH, and the control conditions, without any treatment, have been set at 1. The experiment has been done once; each condition was performed in triplicate. (B) The relative expression of claudin-1 at protein level has been assessed by Western Blot in the same cellular conditions as above. The expression of β-actin has been used as a housekeeping gene for the protein expression. The claudin-1/β-actin ratios are presented in PHA-stimulated PBMCs SN conditions, respectively, after quantization of the Blot’s pictures, using the online ImageJ software (downloaded on the 2nd of May 2022). The Western Blots were run twice and the mean ratios ± SD obtained from each Blot are shown. The black dotted lines highlight the effect of c-MIM-1 compared with Veh. |
c-MIM-2 Displays Anti-Inflammatory Effects in Human Primary Monocytes Exposed to Lipopolysaccharide
The role of TNF-α as a mediator of chronic inflammation,36 as well as the capacity of PGE2 in exacerbating the severity of the inflammatory-mediated diseases,16 both attest to the necessity for a better understanding of the effect of these factors when included in MI formulation at ULD. Interestingly, c-MIM-2 rightly contains PGE2 at the inhibitory dose of 200 K, altogether with TNF-α and IL-1β at 17 CH, IL-6 at 9 CH, and the anti-inflammatory cytokine IL-10 at 4 CH. In order to assess the ability of c-MIM-2 to act as an anti-inflammatory agent, and in particular, in reducing the secretion of TNF-α and PGE2, a cellular model of human primary monocytes inflamed with LPS was chosen. One of the reasons for this model’s selection relies on the fact that monocytes exposed to LPS produce high levels of TNF-α,37 and PGE2,38 making them a suitable model to test the capacity of c-MIM-2 to down-regulate their expression.
The followed protocol is illustrated in Figure 6A but, briefly, the cells were isolated from three different donors and then incubated with either Veh. or c-MIM-2. The inflammatory stimulus was delivered to the cells in the form of a 10 ng/mL LPS threat, 30 minutes after the initiation of the treatment. The treatments were applied for 24 hours and the secretion of the pro-inflammatory factors TNF-α and PGE2 was evaluated within the SN by ELISA assay. The results are presented in Figure 6B–E either as a percentage of the cytokine secretion in the untreated Ct (-) condition, or as a percentage of the cytokine secretion in the Veh. condition, these latter being set as 100% for each tested cytokine. As expected, LPS stimulation led to a drastic increase in TNF-α secretion compared with the untreated negative control Ct (-) (Figure 6B), thus validating our model selection. In these inflammatory conditions, c-MIM-2 led to a reduction in the TNF-α secretion of about 20% (Figure 6C) compared with the Veh. In the same manner, after LPS stimulation, which led to a high increase in the basal secretion of PGE2, c-MIM-2 induced a decrease in the secretion of PGE2 of about 30% (Figure 6D). Finally, as c-MIM-2 contains IL-1β at 17 CH and IL-10 at 4 CH, the secretion levels of these two other cytokines were also appraised in the same conditions. However, while c-MIM-2 did not affect IL-1β, the IL-10 levels were below the ELISA’s detection limit in all tested conditions (data not shown).
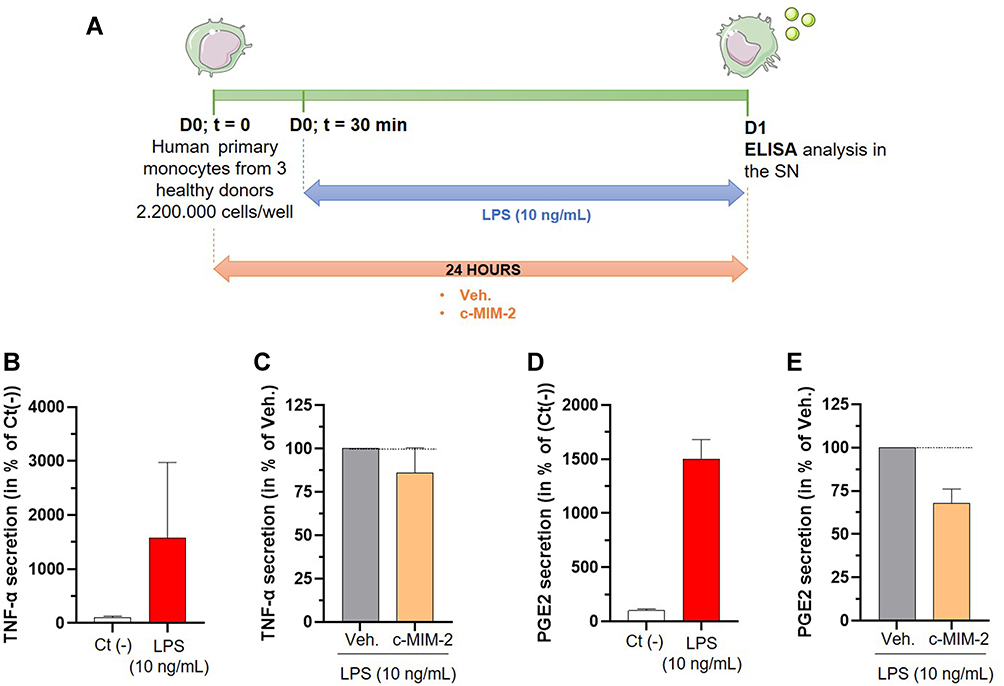 |
Figure 6 C-MIM-2 diminished the secretion of pro-inflammatory cytokines in human primary monocytes. (A) Experimental scheme for the cytokines secretion assessment in presence of c-MIM-2. Human primary monocytes were treated during 24 hours with either Veh. or c-MIM-2. LPS (10 ng/mL) was added to the medium 30 minutes after the treatment initiation, as an induced or the inflammatory conditions. The levels of secreted (B and C) TNF-α and (D and E) PGE2 were assessed in the culture media at the end of the incubation time by ELISA assay. The results are presented as the mean percentage ± SD of the values obtained for each donor. Each condition was performed in triplicate for each donor (3 donors) and in duplicate for the LPS conditions. The results are presented either as a percentage of the Ct (-) secretion or as a percentage of the Veh. secretion (set at 100%). The black dotted lines highlight the effect of c-MIM-2 compared with Veh. Abbreviation: SN, supernatants. |
c-MIM-3 Displays Modulatory Effects on Interleukin-6 Secretion in Lipopolysaccharide-Exposed Human Macrophages and Granulocytes
Human macrophages exposed to LPS markedly produce pro-inflammatory cytokines such as the pleiotropic cytokine IL-6.39 The next experiment thus aimed at assessing if c-MIM-3 could modulate the secretion of IL-6 in a model of LPS-inflamed human macrophages. The experimental scheme is illustrated in Figure 7A, but briefly, CD14+-cells were isolated from one healthy donor and were incubated for 6 days in a macrophage-differentiation medium supplemented with M-CSF 100 ng/mL. At day 6 (D6), the cells were treated with 100 ng/mL LPS concomitantly with either the Veh. or c-MIM-3 for a period of 24 hours or 48 hours. The amount of IL-6 released in the SN was quantified by ELISA at the end of the two tested incubation periods. As expected, 24 hours of LPS-stimulation induced a drastic increase in the secretion of IL-6 compared with the untreated control Ct (-) (Figure 7B). Interestingly, after 24 hours in presence of c-MIM-3, a slight decrease in the amount of IL-6 in the SN was noticed (about 10%) compared with the Veh. (Figure 7C). In the same manner, while at 48 hours, the induction of IL-6 secretion due to LPS exposure was even more important than the one observed after 24 hours (Figure 7D vs B), the reduction induced by c-MIM-3 was also slightly more pronounced (about 15%) after this longer incubation period (Figure 7E vs C).
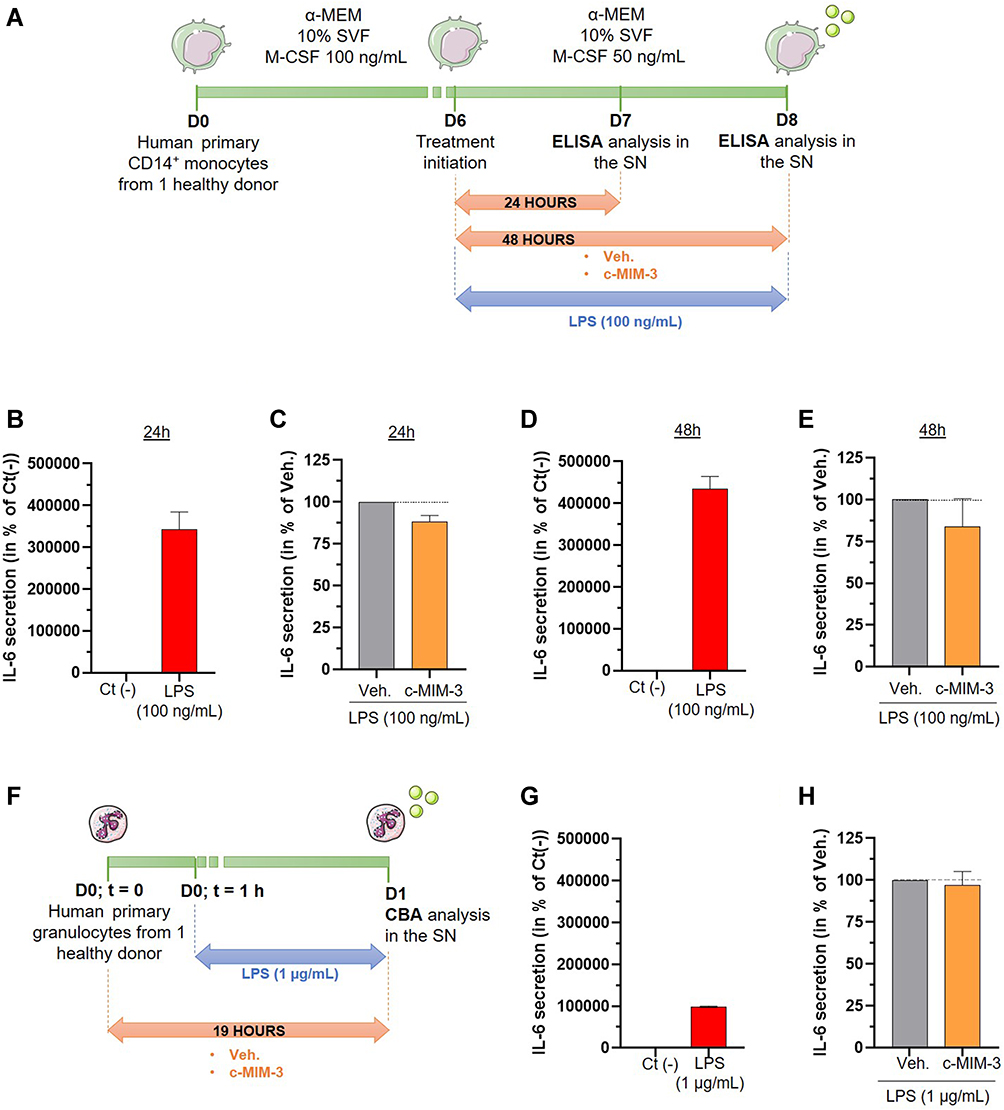 |
Figure 7 C-MIM-3 modulated the secretion of pro-inflammatory cytokines in immune cells. (A) Experimental scheme for the cytokine secretion assessment in presence of c-MIM-3. Human primary monocytes were differentiated into macrophages during 6 days and were then treated during 24 hours or during 48 hours with either Veh. or c-MIM-3. LPS (100 ng/mL) was added to the medium at the same time, as an inducer or the inflammatory conditions. The secreted levels of IL-6 were measured by ELISA assay after (B and C) 24 hours or (D and E) 48 hours. The results are presented as the mean percentage ± SD of the values obtained for each triplicate per condition (n = 6 for the LPS conditions). The untreated negative control (Ct (-)) or the Veh. conditions are set at 100%. (F) Experimental scheme for the IL-6 secretion assessment in presence of c-MIM-3. Human primary granulocytes freshly isolated from one healthy donor were seeded in 48-wells plates and pre-incubated during one hour in presence of either the Veh. or c-MIM-3. LPS (1 µg/mL) was then added to the medium, as an inducer of the inflammatory conditions. The secreted levels of IL-6 were measured by CBA 18 hours later. The secreted levels of IL-6 were evaluated after stimulation with LPS alone (G), and in presence of either the Veh. or c-MIM-3 (H). The results are presented as the mean percentage ± SD of the values obtained for each triplicate. For each panel, the black dotted lines highlight the effect of c-MIM-2 and c-MIM-3 compared with Veh. Abbreviation: SN, supernatants. |
Finally, the effect of c-MIM-3 on the IL-6 secretion was also assessed in a model of human primary granulocytes isolated from one healthy donor and incubated with either Veh. or c-MIM-3 for 19 hours (Figure 7F). The inflammatory stimulus was added one hour after treatment initiation, in the form of 1 µg/mL LPS. The levels of IL-6 were assessed by cytometric bead array (CBA) at the end of the incubation period (Figure 7G and H). As illustrated in Figure 7G, LPS alone elicited a strong IL-6 response when compared with the Ct (-). However, this response was weaker than the one observed in macrophages. In this model, the effect of c-MIM-3 on the secretion of this cytokine was barely any different from the Veh.
c-MIM-4 Displays Anti-Inflammatory Effects in a Cellular Model of Neuronal Inflammation
The following part of this study aimed at considering the effects of c-MIM-4 in a neuronal model of inflammation. In order to do so, rat cortical neurons and glial cells were cultured in presence of a combination of LPS (100 ng/mL)/IFN-γ (10 ng/mL), to provide an inflammatory stimulus, and treated during 96 hours with either the Veh., c-MIM-4 or MK801, an antagonist of N-methyl-D-aspartate (NMDA) receptor that has antidepressant-like effects, used as a reference compound for neuron protection.40 The cell survival was assessed after immune-staining with an anti-MAP-2 antibody, which is specific for neurons. The TNF-α release was investigated, too. The experimental scheme is illustrated in Figure 8A.
 |
Figure 8 C-MIM-4 displays anti-inflammatory effects in a cellular model of neuronal inflammation. (A) Representative scheme of the experimental protocol. Briefly, rat glial cells (astrocytes/microglial cells) and rat cortical neurons were co-cultured during 8 days before treatment initiation with either Veh., c-MIM-4, or MK801 (20 µM). The treatments were applied for 24 hours, before adding or not LPS (100 ng/mL)/IFN-γ (10 ng/mL) for an additional 24 hours. At this time, supernatants (SN) were harvested and frozen for further ELISA analysis. The tested items ± LPS/IFN-γ were incubated another 48 hours before neurons survival analysis. The latter was performed after cell fixation, permeabilization and staining with a monoclonal anti-microtubule associated protein 2 (MAP-2) antibody, specific towards neuron cell bodies and neurites. Images of each culture wells were taken and the fluorescence was quantified. BDNF, brain-derived neurotrophic factor; FCS, fetal calf serum; PS, penicillin – streptomycin. (B) Effect of a 96 hours c-MIM-4 treatment on rat primary cortical neurons survival, in presence of LPS (100 ng/mL)/IFN-γ (10 ng/mL), expressed in percentage of control medium. The results are presented as the mean ± SD of n = 6 technical replicates. (C) Effect of a 48 hours treatment by c-MIM-4 on TNF-α release by astrocytes and microglial cells, expressed as percentages of control medium (mean ± SD of n = 6 technical replicates). In this experiment, the cells were pre-incubated during 24 hours with either Veh., c-MIM-4 or MK801, then LPS (100 ng/mL)/IFN-γ (10 ng/mL) was added for an additional 24 hours. The black dotted lines highlight the effect of c-MIM-4 compared with the Veh. |
The results about neuronal viability are presented in Figure 8B. As illustrated, the inflammatory stimulus, applied during 72 hours, induced a decrease in cortical neurons survival compared with the control condition (about 40% of cellular death; Figure 8B, red histogram vs white histogram). As expected, the reference molecule, MK801, at 20 µM, applied during the entire 96 hours incubation period, restored the survival of neurons to a level that is comparable to the uninflamed, untreated control (about 95% of neuron survival, Figure 8B, white, red, and green histograms). Interestingly, while the Veh. did not seem to impact cellular viability (Figure 8B, grey histogram vs red histogram), treatment with c-MIM-4 was able to rescue neurons from cell death (about 90% of cell survival, Figure 8B, pink histogram).
The effect of c-MIM-4 on the release of TNF-α by reactive glial cells (astrocytes and microglia cells) was also appraised after 48 hours of c-MIM-4 treatment (Figure 8A). As observed in Figure 8C, the inflammatory stimulus induced an increase of about 40% in the TNF-α secretion by activated glial cells, compared to the control medium. Veh. and MK801 (20 µM), both displayed similar effects on TNF-α secretion, by slightly decreasing it compared to the inflammatory control medium. Interestingly, treatment with c-MIM-4 decreased the TNF-α release by activated glial cells by about 10% compared with the Veh. conditions (Figure 8C, grey histogram vs pink histogram). Taken together, these results illustrate for the first time, the anti-inflammatory potential of the complex MI formulation c-MIM-4, and its positive role in neuro-protection in a context of LPS/IFN-γ-induced threat.
The cell survival was finally assessed in absence of inflammation in the same cells. As showed in Supplementary Figure S3, the treatment did not affect the neuronal viability, thus suggesting that the tested MI formulation only acts in homeostatic-disrupted models of neuronal cells.
Discussion
In this study, we focused our attention on several MIMs which, based on their compositions, and according to previous published works,4,9,10 may exert anti-inflammatory effects in different specific inflammatory contexts (Figure 2). With this in mind, the body of data that we provide here particularly resonates with one of our works, in which we reported that the two unitary medicines TNF-α (27 CH) and IL-1β (27 CH) displayed the capacity to inhibit the secretion of IL-1β and TNF-α, in a model of human primary monocytes as well as in THP-1 cells, after LPS exposure.9
Here, the same unitary MI medicines IL-1β (27 CH) and TNF-α (27 CH) (referred to as u-MIM-1 and −2, respectively) were assessed for their anti-inflammatory capacities in a model of intestinal inflammation. The HT-29 cell line was thus used, due to the fact that these human colorectal adenocarcinoma cells of epithelial morphology are widely used as an in vitro model to study absorption, transport, and secretion by intestinal cells.41,42 Under standard culture conditions, these cells grow as a nonpolarized, undifferentiated multilayer. At basal levels, without inflammation, our results show that they express mRNA of both IL-1β and TNF-α, the amount of TNF-α being more than ten times higher than the one of IL-1β (Figure 3B and C). Surprisingly, in their study, Eckmann et al were only able to detect IL-1β mRNA but no TNF-α in these cells.43 Interestingly, in TNF-α-stimulated conditions, our model showed an increase in the transcription of TNF-α, thus suggesting an auto-amplificatory effect of this cytokine on its own expression. Even if such effect was already highlighted in TNF-α-treated 3T3-L1 adipocytes and in TNF-α-injected C57BL/6J mice,44 it is, to the best of our knowledge, the first time that such an auto-inducive effect of TNF-α was reported in a colonic epithelial cell model. In any event, in this model of inflamed HT-29 cells, both u-MIM-1 and u-MIM-2, when employed at 27 CH, were able to down-regulate mRNA levels of IL-1β and TNF-α (Figure 3B and C). These results let us assume that c-MIM formulations including these two factors at ULD could display interesting anti-inflammatory properties.
The intestine is the largest barrier to the external environment and preserving its integrity is crucial for the whole-body homeostasis. Cytokines are known to play an important role in maintaining intestinal homeostasis as they coordinate diverse functions of the intestinal epithelium (proliferation, viability, barrier permeability), as well as they orchestrate the communication between intestinal cells and resident immune cells.45 In particular, multiple studies have revealed the protective effects of IL-10 in maintaining a healthy intestinal epithelium and a good balance between permeability and barrier functions.46–48 The observed effects of c-MIM-1 in our in vitro intestinal model, which, alone, could limit the barrier disruption induced by the pro-inflammatory cocktail provided by SN from PHA-stimulated PBMCs (Figure 4), are in line with these pieces of evidence. It should also be noted that c-MIM-1 employs LD of IL-10 together with ULD of IL-1β, TNF-α, and IL-8. These three pro-inflammatory cytokines are involved in both the initiation and the progression of the inflammatory environment which leads to intestinal injury, increased intestinal permeability, and the development of intestinal inflammatory-related diseases.14,49,50 Counteracting the elevated levels of pro-inflammatory cytokines in the intestine may be a valid therapeutic solution in various intestinal inflammatory disorders.33,51 Keeping in mind that c-MIM-1’s formulation includes ULD of the pro-inflammatory factors TNF-α, IL-1β, and IL-8, it is important to consider that their ULD employ may have downregulated the physiological/biological effects of these cytokines, helping in restoring the epithelial permeability and the tight junctions’ proteins’ expression impaired by the inflammatory condition induced in vitro. Precisely, while additional studies are necessary to confirm the beneficial effects of c-MIM-1 and to understand its mode of action, our results revealed that it could act on claudin-1, as it slightly increased its expression, both at mRNA and at protein levels (Figure 5).
Previous pieces of evidence have indeed demonstrated that inflammatory stimuli, such as TNF-α exposure, lead to the reduction of claudin-1 expression and an altered epithelial permeability.52 Moreover, Maria-Ferreira et al also reported that IL-1β treatment led to a decrease in claudin-1 protein expression in Caco-2 cells.53 Although no direct evidence of the role of IL-8 in claudin-1 regulation has been reported yet, this cytokine was however demonstrated to exert an inhibitory effect towards the mRNA expression of claudin-5 and occludin, in a dose- and in a time-dependent fashion, in endothelial cells.54 On the other hand, c-MIM-1 also contains LD of the anti-inflammatory cytokine IL-10, with the intention to stimulate the conventional cellular responses to this cytokine. While the relationship between IL-10 and claudin-1 still needs to be refined, it was reported that claudin-1 localization was altered in liver cells from a model of IL-10-knocked-out mice, in which colitis was induced with dinitrobenzene sulfonic acid.55 Altogether, these data let us presume that the effect of c-MIM-1 on claudin-1 expression (Figure 5) could be attributable to the presence of TNF-α, IL-1β and IL-8 in ULD in its formulation, as well as IL-10 at LD, and highlight the fact that a deeper analysis of c-MIM-1’s global effect on the expression of tight junctions’ proteins could be envisioned in the future.
Regarding the interconnection between the gut and the immune axis, and taking into consideration the crucial involvement of pro-inflammatory cytokines in IBD, our results about the effects of c-MIM-2 on the secretion of TNF-α and PGE2 in an LPS-stimulated human primary monocyte model are quite encouraging (Figure 6C, E). It has indeed been reported that TNF initiated different and redundant cellular cascades participating in IBD development. In particular, TNF was shown to contribute to the generation of an IL-12 and IFN-γ-driven Th1-like response, and to the activation of a pathogenic CD8+ T cell compartment, in the TnfΔARE IBD mouse model, supporting the link between gut and immunity.56 Moreover, not only PGE2 is involved in the regulation of immune-related patterns,57 but it was also shown to be involved in IBD development in mice, especially through its regulatory effect towards the IL-17/IL-23 axis.58 The observed reduction in the secretion of these two factors, TNF-α and PGE2, after c-MIM-2 treatment, suggested the possible direct involvement of the two active substances, TNF-α (17 CH) and PGE2 (200 K), respectively. In addition, such secretion’s reduction also suggested the involvement of IL-1β (17 CH), as IL-1β is directly implicated in the TNF-α gene expression in a PKC-dependent manner,59 and in PGE2 expression through cAMP-PKA signaling pathway.60
Given the importance of IL-6 in inflammatory responses and inflammatory-mediated diseases, including IBD, our research focused on this cytokine. It has been reported that cultures of isolated lamina propria mononuclear cells and mucosal tissues from IBD patients exhibited higher IL-6 activity than healthy controls.61 This study also suggested that the local elevation of IL-6 activity could be a characteristic of active IBD and that, macrophages and colonic epithelial cells could be the responsible cells for this phenomenon. In this regard, our results showing a reduction of the levels of IL-6 secretion in c-MIM-3-treated inflamed-macrophages (Figure 7C, 7E), are quite encouraging and highlight the potential of c-MIM-3 to be used to decrease the inflammation in pathologies such as IBD. Moreover, preclinical and clinical research studies support the fact that circulating levels of IL-6 are also elevated in several chronic inflammation-mediated diseases other than IBD, like rheumatoid arthritis, systemic lupus erythematosus, ankylosing spondylitis, and psoriasis.62 Thus, even if more studies are needed, these data could open the anti-inflammatory action’s scope of c-MIM-3 to various pathological conditions related to inflammation in the future. On another hand, the different extents in IL-6 inhibition observed after c-MIM-3 treatment in macrophages and in granulocytes (Figure 7C, E, and H) confirmed the necessity of an in-depth understanding of how MIM’s formulations work, especially towards immune-populations. At the moment, it can be speculated that the effect of c-MIM-3 on the two assessed models could have depended on the degree of IL-6 release induced by the inflammatory stimulus. The results obtained in our study suggest that the inhibitory effect of c-MIM-3 on IL-6 occurs when the levels of IL-6 are very important (Figure B, D, and G). One of the limitations of the analysis here is that the experimental protocols were not exactly the same between the two models and further studies are definitely needed in order to compare the responses of the immune cells regarding c-MIM-3 treatment, at different extents of inflammation.
Finally, it is also important to highlight that c-MIM-3 employs IL-6 at 9 CH, and this dilution belongs to the modulatory dilutions of ULD-based MIM. While further studies aiming at assessing the cellular effects of unitaries MIM employing modulatory ULD are necessary, this study discusses for the first time the results of a complex MIM that uses a cytokine at 9 CH, on its own secretion.
In their work, Mizuno et al reported the direct effect of IFN-γ on neurons; as they observed the formation of dendritic beads containing proteins from collapsed cytoskeletal structures mediating neuronal dysfunctions, without increasing cell death.63 However, when IFN-γ is combined with pro-inflammatory cytokines, including TNF-α, the neurotoxicity is exacerbated and neurons viability decreases.64 Coherently with these previous pieces of evidence, the neuronal inflammatory model used in the present study, that combined LPS and IFN-γ, induced an important neurotoxicity (Figure 8). However, the tested c-MIM-4 formulation was able to almost rescue the neuronal viability and slightly reduce the release of TNF-α. Thus, it can reasonably be hypothesized that the observed c-MIM-4’s effect on neuronal cytotoxicity reduction could be due, at least partially, to the presence of ULD of TNF-α in its formulation, as this active ingredient may also be the direct mediator of the observed c-MIM-4’ effect on TNF-α secretion. Moreover, the ULD of IL-12 in its formulation may also have had an effect on the reduction in the TNF-α secretion, as it was reported that IL-12p40 monomer and homodimer induced TNF-α production in mouse BV-2 microglial cells, mouse primary microglia and peritoneal macrophages.65
The research at the frontiers of immunology and neuroscience has identified multiple points of convergence between the immune system and the nervous system. Thanks to the overall molecular mechanistic studies on this concern, the knowledge about the neuro-immune interactions has advanced greatly. Nowadays, a growing body of evidences supports that, pro-inflammatory cytokines such as TNF-α and IL-1β, and the clinical situation related to their uncontrolled overexpression, also referred as chronic inflammation, are extremely linked with the establishment of pathological/chronic pain.66 Pain is considered chronic and pathological when it lasts at least 3 months.67 It has been demonstrated that sensory neurons express cytokine receptors, including IL-1βR, TNF-αR, IL-6R, and G protein-coupled receptors for serotonin, histamine, and PGE2.68,69 Activation of these receptors expressed by nearby nociceptive neurons leads to enhanced membrane excitability and sensitization, which in turn, induce an over the responsiveness of those neurons, and the consequent development of pathological pain.69 Downregulating these pro-inflammatory mediators may represent the key strategy to improve chronic inflammation and the related pain. The therapeutic potentials of the MI formulations, by reducing the secretion of TNF-α and IL-1β, and/or PGE2, and/or IL-6, can indeed cover a very large spectrum of chronic-inflammatory mediated illness and conditions.
Visceral pain is one of the major impediments of IBD, as it highly affects patients’ quality of life. Interestingly, neutralization of TNF-α was reported to rapidly decrease pain perception in the brain of patients with Crohn's disease responding to anti-TNF therapy.70 Moreover, the role of TNF-α and satellite glial cells’ activation in neuron-glial communication has been highlighted in the context of inflammatory visceral hyperalgesia, in a model of rat-induced colonic inflammation in which intrathecal administration of anti-TNF-α (10 µg/kg/day) reduced visceral pain behavioral responses.71 With this data in mind, reducing TNF-α secretion through the use of the MI formulation c-MIM-4 could allow acting on TNF-α-mediated neuron-glial communication axis, which may result in antinociceptive effects in IBD.
As we developed all along the article, the three different inflammation contexts and models used are all interconnected together as the gut, the immune cells, and the nervous axis are constantly communicating between them. To cite some examples, several studies suggest that peripheral inflammation linked to IBD could also be a potential risk factor for certain neurodegenerative disorders. For instance, ulcerative colitis has been reported to exacerbate LPS-induced damage to the nigral dopaminergic system.72 As another example, a clinical study carried out in patients suffering from rheumatoid arthritis (a chronic inflammatory and autoimmune disease), demonstrated the efficacy of electrical vagus nerve stimulation in attenuating the symptoms and significantly inhibiting circulating levels of TNF, IL-1β, and IL-6.73
Another important consideration that can finally be developed is about the reported comorbidity of depression and chronic pain and the possibility that inflammation is the common mediator of this comorbidity.74 Preclinical studies demonstrated that intracerebroventricular injections of proinflammatory cytokines such as TNF-α and IL-1β were sufficient to produce depressive-like behaviors in rats,75 highlighting the crucial role of these two key cytokines, in neuronal inflammation and the related depression.
Conclusion
Overall, the present study highlighted the anti-inflammatory effects of several MIM preparations, which all include in their formulations ULD of IL-1β and TNF-α, in three different contexts of inflammation. The main results of this study are thus put together and illustrated in Figure 9 and could be summarized as this: MI formulations containing ULD of IL-1β and TNF-α displayed anti-inflammatory effects in several models of inflammation related to the (i) intestinal inflammation, (ii) immune-cells mediated-systemic inflammation and, (iii) neuronal inflammation, and could all be linked together in an attempt to control the inflammation within the body. In conclusion, it can be said that u-MIM-1 and −2 down-regulated the mRNA expression of both IL-1β and TNF-α in an in vitro model of intestinal inflammation. In the same cellular model, under inflammatory conditions mimicked by incubation with SN from PHA-treated PBMCs, c-MIM-1 allowed a restoration of the epithelial resistance, probably due to an increase in the expression of the tight junctions’ protein, claudin-1. In addition, MIM-2 and −3 were shown to decrease the secretion of the pro-inflammatory factors TNF-α, PGE2, and IL-6 in LPS-stimulated monocytes/macrophages. Finally, c-MIM-4 was able to restore the cell viability of murine primary neurons inflamed with a combination of LPS/IFN-γ, while reducing their TNF-α secretion. Even if more data are definitely needed, these results highlight for the first time that MI formulations may be a useful strategy to manage a large spectrum of inflammation-related diseases, including IBD or depression.
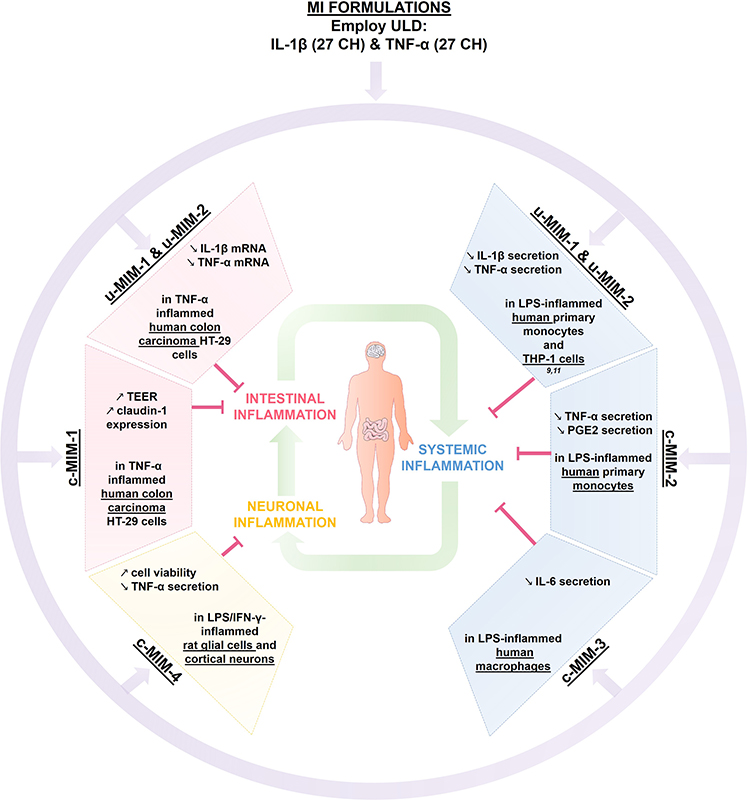 |
Figure 9 Recapitulative scheme about the anti-inflammatory effects of IL-1β and TNF-α, when employed at ULD in MI medicines formulations. The two unitary medicines IL-1β (27 CH) and TNF-α (27 CH) (u-MIM-1, and −2, respectively), as well as the four MI complex formulations (c-MIM-1, −2, −3, and −4) display anti-inflammatory properties in the context of (i) intestinal inflammation (red box), (ii) inflamed-immune cells related to systemic inflammation (blue box), and (iii) neuronal inflammation (yellow box) contexts. Purple arrows illustrate the fact that IL-1β and TNF-α are included into MI formulations, either in u-MIM or in c-MIM. Green arrows illustrate the interconnexions between the different inflammatory contexts (i), (ii) and (iii). Abbreviations: MI, micro-immunotherapy; ULD, ultra-low dose. |
Abbreviations
BDNF, brain-derived neurotrophic factor; CBA, cytometric bead array; CH, centesimal Hahnemannian; DNA, deoxyribonucleic acid; ELISA, enzyme-linked immunosorbent assay; EFS, Etablissement Français du Sang; F, forward; FBS, fetal bovine serum; FCS, fetal calf serum; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GI, gastro-intestinal tract; HBSS, Hank’s buffered saline solution; HLA, human leukocyte antigen; hr, human recombinant; IBD, inflammatory bowel diseases; IFN, interferon; ILs, interleukins; K, Korsakovian dilution; LD, low dose; LPS, lipopolysaccharide; MAP-2, microtubule-associated protein 2; MCH, major histocompatibility complex; M-CSF, macrophage colony-stimulating factor; MI, micro-immunotherapy; MIM, micro-immunotherapy medicine; NDMA, N-methyl-D-aspartate; OSM, oncostatin M; PBMCs, peripheral blood mononuclear cells; PBS, phosphate buffer saline; PHA, phytohemagglutinin; PMSF, phenylmethyl sulfonyl fluoride; P/S, penicillin/streptomycin; PVDF, polyvinylidene fluoride; R, reverse; RNA, ribonucleic acid; RT, room temperature; SD, standard deviation; SKP, serial kinetic process; SN, supernatant; SNA®, specific nucleic acids; TBS, tris-buffered saline; TEER, transepithelial electrical resistance; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-α; ULD, ultra-low dose; Veh, vehicle.
Acknowledgments
The authors are grateful to the staff of Intestinal Biotech Development (especially Pierre Desreumaux, Nghiep Truong Tan, and Christel Rousseaux), for their work in performing the experiments of the in vitro study on the intestinal inflammatory model. The authors would like to thank the staff of Atlantic Bone Screen, especially Mathieu Riffault for his work on the evaluation of the cytokine secretion in monocytes, Emilie Schol and Adrien Brulefert (QIMA Sciences), for having supervised the in vitro experiments on the evaluation of the cytokine secretion in granulocytes and Kurt Appel (VivaCell Biotechnology GmbH) for having supervised the in vitro experiments on the LPS-inflamed primary monocytes. The authors would also like to thank Neuron Experts, and especially Remy Steinschneider, for having supervised the experiments done in the neuronal inflammatory model as well as Jean-Claude Santiago Michea and Mourad Ouamer for having performed the experiments with neurons and the image analysis, respectively. The authors wish to thank Nicoletta Libera Lilli and Beatrice Lejeune for their substantial contribution to the conception and management of the in vitro studies on the intestinal and neuronal inflammatory models. The authors would like to thank Anne Naedts, Cédric Wolf, Coralie Lhots, Virginie Reynders, Sylvain D’Hooghe, and David Wilders for having prepared, provided and sent the vehicle and the tested MI medicines to the laboratories. The authors would also like to thank Cloé Azarias, Dr. Pascal Mensah, and Sofia Frau for their help in revising the manuscript. The authors finally thank Servier Medical Art (https://smart.servier.com/, accessed on 04/07/2022) for the items used in the figures.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. The data of the current study are available from the corresponding author on reasonable request.
Funding
This study was entirely funded by Labo’Life France.
Disclosure
The authors declared the following conflicts of interest with respect to the research, authorship, and/or publication of this article, Camille Jacques and Ilaria Floris work for Labo’Life France, the company service provider of Labo’Life, which is specialized in preclinical research and regulatory affairs. This professional relationship does not imply any misconduct on the part of the authors.
References
1. Pahwa R, Goyal A, Jialal I. Chronic Inflammation. In: StatPearls. StatPearls Publishing; 2022.
2. Wu R-Q, Zhang D-F, Tu E, Chen Q-M, Chen W. The mucosal immune system in the oral cavity—an orchestra of T cell diversity. Int J Oral Sci. 2014;6:125–132. doi:10.1038/ijos.2014.48
3. Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nat Rev Immunol. 2014;14:667–685. doi:10.1038/nri3738
4. Floris I, García-González V, Palomares B, Appel K, Lejeune B. The micro-immunotherapy medicine 2LARTH® reduces inflammation and symptoms of rheumatoid arthritis in vivo. Int J Rheumatol. 2020;2020:1594573. doi:10.1155/2020/1594573
5. Floris I, Chenuet P, Togbe D, Volteau C, Lejeune B. Potential role of the micro-immunotherapy medicine 2LALERG in the treatment of pollen-induced allergic inflammation. Dose-Response Publ Int Hormesis Soc. 2020;18:1559325820914092.
6. Jacques C, Chatelais M, Fekir K, et al. The micro-immunotherapy medicine 2LEID exhibits an immunostimulant effect by boosting both innate and adaptive immune responses. Int J Mol Sci. 2022;23:110. doi:10.3390/ijms23010110
7. Jacques C, Chatelais M, Fekir K, Brulefert A, Floris I. The unitary micro-immunotherapy medicine interferon-γ (4 CH) displays similar immunostimulatory and immunomodulatory effects than those of biologically active human interferon-γ on various cell types. Int J Mol Sci. 2022;23:2314. doi:10.3390/ijms23042314
8. Decker M-L, Gotta V, Wellmann S, Ritz N. Cytokine profiling in healthy children shows association of age with cytokine concentrations. Sci Rep. 2017;7:17842. doi:10.1038/s41598-017-17865-2
9. Floris I, Rose T, Rojas JA, et al. Pro-inflammatory cytokines at ultra-low dose exert anti-inflammatory effect in vitro, A possible mode of action involving sub-micron particles? Dose-Response Publ Int Hormesis Soc. 2020;18:1559325820961723.
10. Floris I, Appel K, Rose T, Lejeune B. 2LARTH®, a micro-immunotherapy medicine, exerts anti-inflammatory effects in vitro and reduces TNF-α and IL-1β secretion. J Inflamm Res. 2018;11:397–405. doi:10.2147/JIR.S174326
11. Jacques C, Floris I, Lejeune B. Ultra-low dose cytokines in rheumatoid arthritis, three birds with one stone as the rationale of the 2LARTH® micro-immunotherapy treatment. Int J Mol Sci. 2021;22:6717. doi:10.3390/ijms22136717
12. Bennett JM, Reeves G, Billman GE, Sturmberg JP. Inflammation-nature’s way to efficiently respond to all types of challenges. Implications for understanding and managing ‘the epidemic’ of chronic diseases. Front Med. 2018;5:316.
13. Jacobson A, Yang D, Vella M, Chiu IM. The intestinal neuro-immune axis, crosstalk between neurons, immune cells, and microbes. Mucosal Immunol. 2021;14:555–565. doi:10.1038/s41385-020-00368-1
14. Cotton JA, Platnich JM, Muruve DA, et al. Interleukin-8 in gastrointestinal inflammation and malignancy, induction and clinical consequences. Int J Interferon Cytokine Mediat Res. 2016;8:13–34.
15. Kumar S, Shukla R, Ranjan P, Kumar A. Interleukin-10, A compelling therapeutic target in patients with irritable bowel syndrome. Clin Ther. 2017;39:632–643. doi:10.1016/j.clinthera.2017.01.030
16. Tsuge K, Inazumi T, Shimamoto A, Sugimoto Y. Molecular mechanisms underlying prostaglandin E2-exacerbated inflammation and immune diseases. Int Immunol. 2019;31:597–606. doi:10.1093/intimm/dxz021
17. Couper KN, Blount DG, Riley EM. IL-10, the master regulator of immunity to infection. J Immunol. 2008;180:5771–5777. doi:10.4049/jimmunol.180.9.5771
18. Iyer SS, Cheng G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit Rev Immunol. 2012;32:23–63. doi:10.1615/CritRevImmunol.v32.i1.30
19. Li MO, Flavell RA. Contextual regulation of inflammation, a duet by transforming growth factor-β and interleukin-10. Immunity. 2008;28:468–476. doi:10.1016/j.immuni.2008.03.003
20. Dinarello CA, Simon A, van der Meer JWM. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat Rev Drug Discov. 2012;11:633–652. doi:10.1038/nrd3800
21. Richards CD. The enigmatic cytokine oncostatin M and roles in disease. ISRN Inflamm. 2013;2013:e512103. doi:10.1155/2013/512103
22. Hirano T. IL-6 in inflammation, autoimmunity and cancer. Int Immunol. 2021;33:127–148. doi:10.1093/intimm/dxaa078
23. Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014;6:a016295. doi:10.1101/cshperspect.a016295
24. Seyfizadeh N, Seyfizadeh N, Gharibi T, Babaloo Z. Interleukin-13 as an important cytokine, A review on its roles in some human diseases. Acta Microbiol Immunol Hung. 2015;62:341–378. doi:10.1556/030.62.2015.4.2
25. Valkanova V, Ebmeier KP, Allan CLCRP. IL-6 and depression, a systematic review and meta-analysis of longitudinal studies. J Affect Disord. 2013;150:736–744. doi:10.1016/j.jad.2013.06.004
26. Kim Y-K, Suh I-B, Kim H, et al. The plasma levels of interleukin-12 in schizophrenia, major depression, and bipolar mania, effects of psychotropic drugs. Mol Psychiatry. 2002;7:1107–1114. doi:10.1038/sj.mp.4001084
27. Park H-J, Shim H-S, An K, et al. IL-4 Inhibits IL-1 β -induced depressive-like behavior and central neurotransmitter alterations. Mediators Inflamm. 2015;2015:941413. doi:10.1155/2015/941413
28. Singer CA, Figueroa-Masot XA, Batchelor RH, Dorsa DM. The mitogen-activated protein kinase pathway mediates estrogen neuroprotection after glutamate toxicity in primary cortical neurons. J Neurosci off J Soc Neurosci. 1999;19:2455–2463. doi:10.1523/JNEUROSCI.19-07-02455.1999
29. Gresa-Arribas N, Viéitez C, Dentesano G, et al. Modelling neuroinflammation in vitro, a tool to test the potential neuroprotective effect of anti-inflammatory agents. PLoS One. 2012;7:e45227. doi:10.1371/journal.pone.0045227
30. Kim I-D, Lee J-K. HMGB1-binding heptamer confers anti-inflammatory effects in primary microglia culture. Exp Neurobiol. 2013;22:301–307. doi:10.5607/en.2013.22.4.301
31. Shu Z, Yang B, Zhao H, et al. Tangeretin exerts anti-neuroinflammatory effects via NF-κB modulation in lipopolysaccharide-stimulated microglial cells. Int Immunopharmacol. 2014;19:275–282. doi:10.1016/j.intimp.2014.01.011
32. Vaux DL. Research methods, Know when your numbers are significant. Nature. 2012;492:180–181. doi:10.1038/492180a
33. Friedrich M, Pohin M, Powrie F. Cytokine networks in the pathophysiology of inflammatory bowel disease. Immunity. 2019;50:992–1006. doi:10.1016/j.immuni.2019.03.017
34. Lin Z, Huang Y, Jiang H, et al. Functional differences and similarities in activated peripheral blood mononuclear cells by lipopolysaccharide or phytohemagglutinin stimulation between human and cynomolgus monkeys. Ann Transl Med. 2021;9:257. doi:10.21037/atm-20-4548
35. Saeedi BJ, Kao DJ, Kitzenberg DA, et al. HIF-dependent regulation of claudin-1 is central to intestinal epithelial tight junction integrity. Mol Biol Cell. 2015;26:2252–2262.
36. Popa C, Netea MG, Van Riel PL, Van Der Meer JW, Stalenhoef AF. The role of TNF-α in chronic inflammatory conditions, intermediary metabolism, and cardiovascular risk. J Lipid Res. 2007;48:751–762. doi:10.1194/jlr.R600021-JLR200
37. van der Bruggen T, Nijenhuis S, van Raaij E, Verhoef J, Sweder van Asbeck B. Lipopolysaccharide-induced tumor necrosis factor alpha production by human monocytes involves the Raf-1/MEK1-MEK2/ERK1-ERK2 Pathway. Infect Immun. 1999;67:3824–3829. doi:10.1128/IAI.67.8.3824-3829.1999
38. Lu Y, Wahl LM. Oxidative stress augments the production of matrix metalloproteinase-1, cyclooxygenase-2, and prostaglandin E2 through enhancement of NF-κB activity in lipopolysaccharide-activated human primary monocytes. J Immunol. 2005;175:5423–5429. doi:10.4049/jimmunol.175.8.5423
39. Liu X, Yin S, Chen Y, et al. LPS‑induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF‑κB, STAT3 or AP‑1 activation. Mol Med Rep. 2018;17:5484–5491. doi:10.3892/mmr.2018.8542
40. Rana P, Bagewadi H, Banerjee BD, Bhattacharya SK, Mediratta PK. Attenuation of oxidative stress and neurotoxicity involved in the antidepressant-like effect of the MK-801(dizocilpine) in bacillus Calmette-Guerin-induced depression in mice. J Basic Clin Physiol Pharmacol. 2020;31. doi:10.1515/jbcpp-2019-0016
41. Collett A, Sims E, Walker D, et al. Comparison of HT29-18-C1 and Caco-2 cell lines as models for studying intestinal paracellular drug absorption. Pharm Res. 1996;13:216–221. doi:10.1023/A:1016082829111
42. Martínez-Maqueda D, Miralles B, Recio I. HT29 Cell Line. in the Impact of Food Bioactives on Health, in vitro and ex vivo Models. eds. Verhoeckx K. Springer International Publishing; 2015:113–124. doi:10.1007/978-3-319-16104-4_11
43. Eckmann L, Jung HC, Schürer-Maly C, et al. Differential cytokine expression by human intestinal epithelial cell lines, regulated expression of interleukin 8. Gastroenterology. 1993;105:1689–1697. doi:10.1016/0016-5085(93)91064-O
44. Neels JG, Pandey M, Hotamisligil GS, Samad F. Autoamplification of tumor necrosis factor-α. Am J Pathol. 2006;168:435–444. doi:10.2353/ajpath.2006.050699
45. Andrews C, McLean MH, Durum SK. Cytokine Tuning of Intestinal Epithelial Function. Front Immunol. 2018;9:1270. doi:10.3389/fimmu.2018.01270
46. Kominsky DJ, Campbell EL, Ehrentraut SF, et al. IFN-γ-mediated induction of an apical IL-10 receptor on polarized intestinal epithelia. J Immunol Baltim Md. 2014;192:1267–1276.
47. Lorén V, Cabré E, Ojanguren I, et al. Interleukin-10 enhances the intestinal epithelial barrier in the presence of corticosteroids through p38 MAPK activity in caco-2 monolayers, A possible mechanism for steroid responsiveness in ulcerative colitis. PLoS One. 2015;10:e0130921. doi:10.1371/journal.pone.0130921
48. Zheng L, Kelly CJ, Battista KD, et al. Microbial-derived butyrate promotes epithelial barrier function through IL-10 receptor-dependent repression of claudin-2. J Immunol Baltim Md. 2017;199:2976–2984.
49. Gitter AH, Bendfeldt K, Schmitz H, et al. Epithelial barrier defects in HT-29/B6 colonic cell monolayers induced by tumor necrosis factor-alpha. Ann N Y Acad Sci. 2000;915:193–203. doi:10.1111/j.1749-6632.2000.tb05242.x
50. Al-Sadi RM, Ma TY. IL-1β causes an increase in intestinal epithelial tight junction permeability. J Immunol Baltim Md. 2007;178:4641–4649.
51. McLean MH, Neurath MF, Durum SK. Targeting interleukins for the treatment of inflammatory bowel disease—what lies beyond anti-TNF therapy? Inflamm Bowel Dis. 2014;20:389–397. doi:10.1097/01.MIB.0000437616.37000.41
52. Ye X, Sun M. AGR2 ameliorates tumor necrosis factor-α-induced epithelial barrier dysfunction via suppression of NF-κB p65-mediated MLCK/p-MLC pathway activation. Int J Mol Med. 2017;39:1206–1214. doi:10.3892/ijmm.2017.2928
53. Maria-Ferreira D, Nascimento AM, Cipriani TR, et al. Rhamnogalacturonan, a chemically-defined polysaccharide, improves intestinal barrier function in DSS-induced colitis in mice and human Caco-2 cells. Sci Rep. 2018;8:12261. doi:10.1038/s41598-018-30526-2
54. Yu H, Huang X, Ma Y, et al. Interleukin-8 regulates endothelial permeability by down-regulation of tight junction but not dependent on integrins induced focal adhesions. Int J Biol Sci. 2013;9:966–979. doi:10.7150/ijbs.6996
55. Mazzon E, Puzzolo D, Caputi AP, Cuzzocrea S. Role of IL-10 in hepatocyte tight junction alteration in mouse model of experimental colitis. Mol Med. 2002;8:353–366. doi:10.1007/BF03402016
56. Kontoyiannis D, Boulougouris G, Manoloukos M, et al. Genetic dissection of the cellular pathways and signaling mechanisms in modeled tumor necrosis factor-induced Crohn’s-like inflammatory bowel disease. J Exp Med. 2002;196:1563–1574. doi:10.1084/jem.20020281
57. Kalinski P. Regulation of immune responses by prostaglandin E2. J Immunol Baltim Md. 2012;188:21–28.
58. Sheibanie AF, Yen JH, Khayrullina T, et al. The proinflammatory effect of prostaglandin E2 in experimental inflammatory bowel disease is mediated through the IL-23-->IL-17 axis. J Immunol Baltim Md. 2007;178:8138–8147.
59. Bethea JR, Gillespie GY, Benveniste EN. Interleukin-1β induction of TNF-α gene expression, Involvement of protein kinase C. J Cell Physiol. 1992;152:264–273. doi:10.1002/jcp.1041520207
60. Park Y-G, Kang SK, Noh SH, et al. PGE2 induces IL-1beta gene expression in mouse osteoblasts through a cAMP-PKA signaling pathway. Int Immunopharmacol. 2004;4:779–789.
61. Kusugami K, Fukatsu A, Tanimoto M, et al. Elevation of interleukin-6 in inflammatory bowel disease is macrophage- and epithelial cell-dependent. Dig Dis Sci. 1995;40:949–959. doi:10.1007/BF02064182
62. Gabay C. Interleukin-6 and chronic inflammation. Arthritis Res Ther. 2006;8:S3. doi:10.1186/ar1917
63. Mizuno T, Zhang G, Takeuchi H, et al. Interferon-γ directly induces neurotoxicity through a neuron specific, calcium-permeable complex of IFN-γ receptor and AMPA GluRl receptor. FASEB J. 2008;22:1797–1806. doi:10.1096/fj.07-099499
64. Yu Z, Cheng G, Wen X, et al. Tumor necrosis factor alpha increases neuronal vulnerability to excitotoxic necrosis by inducing expression of the AMPA-glutamate receptor subunit GluR1 via an acid sphingomyelinase- and NF-kappaB-dependent mechanism. Neurobiol Dis. 2002;11:199–213. doi:10.1006/nbdi.2002.0530
65. Jana M, Dasgupta S, Saha RN, Liu X, Pahan K. Induction of tumor necrosis factor-α (TNF-α) by interleukin-12 p40 monomer and homodimer in microglia and macrophages. J Neurochem. 2003;86:519–528. doi:10.1046/j.1471-4159.2003.01864.x
66. Vanderwall AG, Milligan ED. Cytokines in pain, harnessing endogenous anti-inflammatory signaling for improved pain management. Front Immunol. 2019;10:3009.
67. Treede R-D, Rief W, Barke A, et al. Chronic pain as a symptom or a disease, the IASP classification of chronic pain for the international classification of diseases (ICD-11). PAIN. 2019;160:19–27. doi:10.1097/j.pain.0000000000001384
68. Chavan SS, Pavlov VA, Tracey KJ. Mechanisms and therapeutic relevance of neuro-immune communication. Immunity. 2017;46:927–942. doi:10.1016/j.immuni.2017.06.008
69. Scholz J, Woolf CJ. The neuropathic pain triad, neurons, immune cells and glia. Nat Neurosci. 2007;10:1361–1368. doi:10.1038/nn1992
70. Hess A, Roesch J, Saake M, et al. Functional brain imaging reveals rapid blockade of abdominal pain response upon anti-TNF therapy in crohn’s disease. Gastroenterology. 2015;149:864–866. doi:10.1053/j.gastro.2015.05.063
71. Song D, Li Y, Tang D, Huang L, Yuan Y. Neuron-glial communication mediated by TNF-α and glial activation in dorsal root ganglia in visceral inflammatory hypersensitivity. Am J Physiol Gastrointest Liver Physiol. 2014;306:G788–795. doi:10.1152/ajpgi.00318.2013
72. Villarán RF, Espinosa-Oliva AM, Sarmiento M, et al. Ulcerative colitis exacerbates lipopolysaccharide-induced damage to the nigral dopaminergic system, potential risk factor in Parkinson`s disease. J Neurochem. 2010;114:1687–1700. doi:10.1111/j.1471-4159.2010.06879.x
73. Koopman FA, Chavan SS, Miljko S, et al. Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. Proc Natl Acad Sci U S A. 2016;113:8284–8289. doi:10.1073/pnas.1605635113
74. Walker AK, Kavelaars A, Heijnen CJ, Dantzer R. Neuroinflammation and Comorbidity of Pain and Depression. Pharmacol Rev. 2014;66:80–101. doi:10.1124/pr.113.008144
75. Connor TJ, Song C, Leonard BE, Merali Z, Anisman H. An assessment of the effects of central interleukin-1β, −2, −6, and tumor necrosis factor-α administration on some behavioural, neurochemical, endocrine and immune parameters in the rat. Neuroscience. 1998;84(3):923–933. doi:10.1016/S0306-4522(97)00533-2
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Elevated CTSL Gene Expression Correlated with Proinflammatory Cytokines in Omental Adipose Tissue of Patients with Obesity
Zhou Q, Zhu Y, Li C, Li Z, Tang Z, Yuan B, Wang X, Zhang S, Wu X
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2277-2285
Published Date: 30 July 2022
May the Nitrosative and Carbonyl Stress Promote Inflammation in Patients with Colorectal Cancer?

Dorf J, Zaręba K, Matowicka-Karna J, Pryczynicz A, Guzińska-Ustymowicz K, Zalewska A, Maciejczyk M
Journal of Inflammation Research 2022, 15:4585-4600
Published Date: 11 August 2022
Variant Angina is Associated with Myocarditis
Xu X, Wang JJ, Zhao H, Miao K, Cui G, Zhang Y, Yang X, Wang L, Wu J, Wang DW
Journal of Inflammation Research 2022, 15:4939-4949
Published Date: 29 August 2022
An inflammatory Signature of Glucose Impairment in Cystic Fibrosis
Montemari AL, Manco M, Fiocchi AG, Bartoli M, Facchiano F, Tabolacci C, Scatigna M, Ciciriello F, Alghisi F, Montemitro E, Carsetti R, Lucidi V, Fiscarelli EV
Journal of Inflammation Research 2022, 15:5677-5685
Published Date: 10 October 2022
Pro-Inflammatory and Anti-Inflammatory Cytokines Levels are Significantly Altered in Cerebrospinal Fluid of Unruptured Intracranial Aneurysm (UIA) Patients
Kamińska J, Maciejczyk M, Ćwiklińska A, Matowicka-Karna J, Koper-Lenkiewicz OM
Journal of Inflammation Research 2022, 15:6245-6261
Published Date: 11 November 2022