Back to Journals » Breast Cancer: Targets and Therapy » Volume 17
Lactate Dehydrogenase-A-Forming LDH5 Promotes Breast Cancer Progression
Authors Fang T ![]() , Hu L, Chen T, Li F, Yang L, Liang B, Wang W, Zeng F
, Hu L, Chen T, Li F, Yang L, Liang B, Wang W, Zeng F
Received 8 November 2024
Accepted for publication 30 January 2025
Published 12 February 2025 Volume 2025:17 Pages 157—170
DOI https://doi.org/10.2147/BCTT.S502670
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Tianxing Fang,1– 4 Liyu Hu,5 Tianshun Chen,4 Fei Li,6 Liu Yang,4 Bin Liang,7 Wenjun Wang,8 Fancai Zeng4
1Department of Nuclear Medicine, Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 2Nuclear Medicine and Molecular Imaging Key Laboratory of Sichuan Province, Luzhou, Sichuan, People’s Republic of China; 3Institute of Nuclear Medicine, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 4Laboratory of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 5Oral & Maxillofacial Reconstruction and Regeneration of Luzhou Key Laboratory, The Affiliated Stomatological Hospital of Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 6Department of Pathology, The Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 7Department of General Surgery (Breast Surgery), the Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan, People’s Republic of China; 8Institute for Cancer Medicine and School of Basic Medical Sciences, Southwest Medical University, Luzhou, Sichuan, People’s Republic of China
Correspondence: Fancai Zeng; Wenjun Wang, Email [email protected]; [email protected]
Background: Breast cancer (BC) has become the main malignant tumor threatening the health of women worldwide. Previous studies have reported that Lactate dehydrogenase-A (LDHA) has critical roles in cancer development and progression. We aimed to explore the roles of LDHA and LDH5 isoenzyme activity in BC, which provides a new insight into LDHA for the treatment of BC.
Methods: The expression of LDHA in BC and its relationship with clinicopathological features were obtained from various databases including The Cancer Genome Atlas (TCGA), Human Protein Atlas (HPA), Breast Cancer-Gene Expression Miner (bc-GenExMiner), TNMplot, UALCAN. The Kaplan‒Meier Plotter was used to evaluate the prognostic value of LDHA. Western blot was performed to detect LDHA expression. Agarose gel electrophoresis was performed to detect the activities of LDH isoenzymes. The in vitro proliferation, migration and invasion potentials of BC cells were evaluated using MTT assays, colony formation, wound-healing assay, matrix metalloproteinase assays and transwell assays, respectively. The activities of LDH isoenzymes in serum and tissues were measured in patients with BC and healthy controls.
Results: Compared to normal tissues, LDHA expression was significantly higher in BC tissues. Patients’ nodal status, histological types, TP53 mutation status and PAM50 subtypes were significant factors influencing the LDHA expression. By overexpressing or silencing LDHA gene in BT549 cells, it was confirmed that LDHA promoted cell proliferation, migration and invasion. LDH5 isoenzyme activity in patients with BC was higher than healthy controls. The increased activity of LDH5 isoenzymes was induced by overexpression of LDHA in BC. High expression of LDHA was found to be associated with poor prognosis in BC.
Conclusion: LDHA plays a critical role in the progression of BC through the regulation of the activity of LDH5 isoenzyme, indicating that LDHA may serve as a valuable target for BC treatment.
Keywords: LDHA, LDH5 isoenzyme, breast cancer, progression, prognosis
Introduction
Breast cancer (BC) has become the main malignant tumor threatening the health of women worldwide.1 Due to the limited number of identified targets, chemotherapy, surgery, and hormone treatments are commonly used, but they often entail significant side effects.2 Therefore, it is urgent to develop novel molecular targets to help improve BC. Currently, increasing evidence has demonstrated that cancer is a metabolic disease.3,4 The “Warburg effect” indicates that under the condition of sufficient oxygen, tumor cells tend to rely on aerobic glycolysis for energy production. This seems to be an “uneconomic” way to obtain energy.5 Lactate dehydrogenase (LDH) is a crucial enzyme of glycolytic metabolic pathway. High LDH values is recognized as a major factor in the poor prognosis of BC.6
There are four subtypes of LDH: LDHA, LDHB, LDHC and LDHD. Among them, LDHA and LDHB are two significant isoforms of LDH, which are predominantly expressed in different types of cancer cells.7 LDHC is a typical cancer testicular antigen that can be used as a targetable cancer testis antigen for immunotherapy.8 LDHD promotes growth, migration, self-renewal, and tumorigenic potential in esophageal squamous cell carcinoma.9 However, many studies are still focused on LDHA, which favors the conversion of pyruvate to lactate.10 Acidic environment provides favorable conditions for tumor invasion.11 A recent study revealed that LDHA promoted the proliferation, invasion, and migration abilities of renal cell carcinoma cell via upregulating the expression of EPHA2 in exosomes.12 In addition, the up-regulation of LDHA in glioblastoma and pancreatic cancer has been confirmed to be correlated with poor prognosis of patients.13,14 Therefore, LDHA may be an important molecular target in BC.
LDH isoenzyme spectrum reflects the tissue-specific glucose metabolism status with the different contents of LDHA and LDHB subunits. There are five isoenzymes of LDH, corresponding to five possible combinations, named LDH1 (B4), LDH2 (B3A), LDH3 (B2A2), LDH4 (BA3), and LDH5 (A4) respectively.15 LDH5 is assembled only from subunit A. The activity of LDH5 isoenzyme in tumors is significantly increased and closely related to poor prognosis.16 Thus, it is interesting to investigate the role of LDHA and LDH5 isoenzyme in BC. It has been proposed that activity analysis of the LDH isoenzyme helps to reflect functions and energy requirements in different tissues.17 Changes in tissue or serum LDH isoenzymes are useful indicators of tissue damage in many diseases.18 Therefore, it is necessary to understand the composition and distribution of LDH isoenzymes in BC. However, the activities of LDH isoenzyme in BC remain unclear. In this study, we aim to explore the role of LDHA and LDH5 isoenzyme activity in BC, which provides a new insight into LDHA.
Materials and Methods
Gene Expression Analysis
The HPA database (https://www.proteinatlas.org) was used to explore the protein expression levels of LDHA in human pathology tissues. The RNA expression of LDHA in human mammary epithelial cell (HMLE) and various BC cell lines was also obtained through the HPA database. bc-GenExMiner (https://bcgenex.ico.unicancer.fr/) is a statistical mining tool. It was used to compare LDHA expression with clinical parameters such as nodal status, histological types, pathologic tumor stage and so on.19,20 The UALCAN database (http://ualcan.path.uab.edu) contains the gene expression data gathered from the TCGA database. We applied the website to analyze the differential expression of LDHA in BC tissues and normal tissues based on clinicopathological characteristics. The TNMplot online database (https://tnmplot.com/analysis/) was used to determine the expression of LDHA in pan-cancer tissues and normal tissues.21 The Kaplan–Meier plotter (https://kmplot.com/analysis/) was used to detect the correlation of LDHA protein expression and overall survival (OS) in BC patients.
Samples Collection
Human-tissue samples and serum samples were obtained from the Affiliated Hospital of Southwest Medical University from August 2021 to July 2022. Cancer and adjacent tissue samples were obtained from 12 patients with pathologically confirmed BC. A total of 69 serum samples of BC patients, as a case group, and 69 serum samples of healthy people, as a control group, the exclusion criterion were as follows: a history of malignancy in other organs, impairment of liver and kidney function. The study followed the 1964 helsinki Declaration and its later amendments or comparable ethical standards, all participants signed the informed consent.
Cell Lines, Culture and Transfection
The human BC cell lines MCF-7, BT-549, ZR-75-30, MDA-MB-231, SK-BR-3, T47D were obtained from the China center for type culture collection. MCF-7, MDA-MB-231, SK-BR-3, T47D were cultured in Dulbecco’s modified Eagle Medium (DMEM, Gibco, USA), BT-549, ZR-75-30 were cultured in RPMI-1640 medium (Gibco, USA), supplemented with 10% fetal bovine serum (Gibco), respectively. All cell lines cultured at 37 °C in humidified air with 5% CO2. Cells were transfected by Lipofectamine 3000 (Thermo Fisher Scientific) according to the manufacturer’s instructions.
RNA Interference
LDHA and control siRNAs were all purchased from Gene Pharma (Shanghai, China). Two independent LDHA siRNAs gave similar results and efficiently silenced LDHA: LDHA siRNA 1(5′-GGAGUGGAAUGAAUGUUGC(dTdT)-3′); LDHA siRNA 2 (5′-CUGGCAAAGACUAUAAUGU(dTdT)-3′).
Western Blot
Proteins from whole cells lysed in RIPA buffer, which were quantified by the BCA protein assay kit. Protein samples were separated by 10% SDS-PAGE electrophoresis and transferred onto PVDF membranes. Then membranes blocked with 5% nonfat milk in TBS Tween 20 at room temperature for 1h and incubated with each primary antibody overnight at 4 °C. LDHA polyclonal antibody was purchased from Santa Cruz, used at a dilution of 1:300 (sc-137243); anti-beta-actin was purchased from Bioworld, used at a dilution of 1:10000 (AP0060). Next day, membranes were washed three times with TBST and incubated with HRP-conjugated secondary antibodies for 1 h at room temperature (They were all purchased from Solarbio). Membranes were washed three times with TBST again. The immunoreactive protein bands were visualized by ECL detection system (ChemiScope 6000).
MTT for Cell Proliferation
Cells were seeded in a 96-well plate at a density of 5000 cells per well. After 4–6 h, cells were transfected and cultured for 48 h. Then, 20 μL MTT (5 g/L) (purchased from Solarbio) was added to each well at 37 °C. After 4–6 h, the media was removed and 150 μL DMSO was added to each well, absorbance values were obtained at 490 nm by using enzyme-labeled instrument (Bio Rad). Experiments were repeated three times and five parallel samples were measured each time.
Colony Formation
Transfected cells were seeded in 6-well plates at 1000 cells, 1500 cells, 2000 cells per well. The culture was replaced every three days. Three weeks later, the medium was removed, and 4% paraformaldehyde was added to each well for 20 min. Then the media was removed, cells were stained with 0.5% crystal violet for 30 min and we took pictures for record.
Wound Healing Assay
Cells were seeded in 24-well plates at a density of 2×105 /mL cells per well. After 24 h, cells were transfected and cultured to 80% density. Then the wound was created by 10 μL pipette tip. Detached cells were washed by PBS, took pictures at once. After 24 h, 48 h, took pictures again.
Transwell
Transfected cells diluted into 1×106 /mL serum-free RPMI-1640 medium at a volume of 200 μL for the transwell assay. Cells were seeded in the Matrigel-coated (BD Biosciences) upper chamber (Millicell), the lower chamber was loaded with RPMI-1640 medium containing 20% FBS at a volume of 600 μL. After 24 h, the medium was removed, 4% paraformaldehyde was added to each chamber for 20 min. Then the media was removed, cells were stained with 0.5% crystal violet for 30 min and we took pictures for record.
Matrix Metalloproteinase Assays
Cells were cultured to 80% density and transfected. After 48 h, cells were washed three times with serum-free RPMI-1640 medium, and continue to culture for 24 h with serum-free RPMI-1640 medium. Then cell supernatant was collected and proteins were quantified by the BCA protein assay kit. Protein samples were separated by 0.1% SDS-PAGE electrophoresis. The gels were washed four times with eluent, each time for 15 min; washed twice with floating lotion, each time for 20 min; placed in the water bath for 48 h at 37 °C; stained for 3 h. The gels were placed in the decolorizing liquid A, B and C successively, washed for 0.5 h, 1 h and 2 h respectively. Lastly, bands were visualized by gel image analysis system (Bio Rad).
LDH Agarose Gel Electrophoresis
Transfected cells were washed twice with PBS and terminated with 1‰ TritonX-100. Then cell supernatant was collected and placed in the water bath for 30 min at 37 °C. The supernatant was centrifuged for 30 minutes at 16000 r/min, retrieved for supernatant again. Samples were separated by agarose gel electrophoresis, dyed in the water bath for 30 min at 37 °C, Lastly, bands were visualized by gel image analysis system (Bio Rad).
Statistical Analysis
The expression of LDHA in pan-cancer tissues and normal tissues was analyzed by Mann–Whitney. Wilcoxon test was utilized to discuss LDHA expression and its relationship with clinical features from the TCGA database. Student’s t-test and ANOVA were used depending on experimental conditions. Survival analysis was performed using the Kaplan–Meier curve, and the differences were compared using the Log rank test. GraphPad Prism 7 and SPSS software (SPSS, version 19.0) was used for statistical analysis. All data presented as means ± SEM, P < 0.05 was considered statistically significant.
Results
LDHA Expression Was Higher in Breast Cancer Compared to Normal Tissues
To explore the potential roles of LDHA in tumors, we evaluated the gene expression of LDHA between cancer tissues and normal tissues by using TNMplot database. The results demonstrated that LDHA was highly expressed in most cancer tissues compared to the normal group (Figure 1A). Then we analyzed protein expression data of LDHA from the HPA databases. Our results confirmed LDHA exhibited strong or moderate expression levels across various types of malignancies including BC (Figure 1B). The results of UALCAN database analysis further confirmed that LDHA mRNA expression was higher in BC than that in normal tissues (Figure 1C). Collectively, these results suggested that LDHA was highly expressed in BC.
 |
Figure 1 LDHA expression was higher in Breast Cancer compared to normal tissues. (A), LDHA gene expression in pan-cancer tissues and normal tissues was analyzed by using TNMplot database. Significant differences are marked with red font; (B), Protein expression of LDHA in various human pathology tissues was analyzed by using HPA databases (antibody: CAB015336). For each cancer, purple bars indicated the percentage of patients (maximum 12 patients) with high and medium protein expression level; (C), LDHA expression in BC and normal samples from UALCAN database. * p<0.05, ** P < 0.01. |
Relationships Between LDHA and Clinical Characteristics of BC Patients
We further analyzed the expression of LDHA in BC patients and its relationship to clinical characteristics from the TCGA database. Notably, LDHA expression was associated with patients’ nodal status (p = 0.0010), histological types (p < 0.0001), TP53 mutation status (p < 0.0001) and PAM50 subtypes (p < 0.0001) in BC, as illustrated in Table 1. These results were consistent with that obtained by UALCAN database. We found that LDHA was highly expressed in BC patients without node metastasis (Figure 2A). Furthermore, BC patients with histological types of invasive lobular carcinoma (ILC) showed lower expression of LDHA compared to others (Figure 2B). Triple negative carcinoma (TNBC) is a more aggressive type of BC,22 which had higher LDHA expression compared to Luminal (Figure 2C). Moreover, LDHA expression was higher in patients with TP53 mutations compared to that with wild type status (Figure 2D). Existing studies have shown that TP53 mutations are involved in the progression of various tumors.23 This indicated higher expression of LDHA was associated with the function of TP53 mutations in BC.
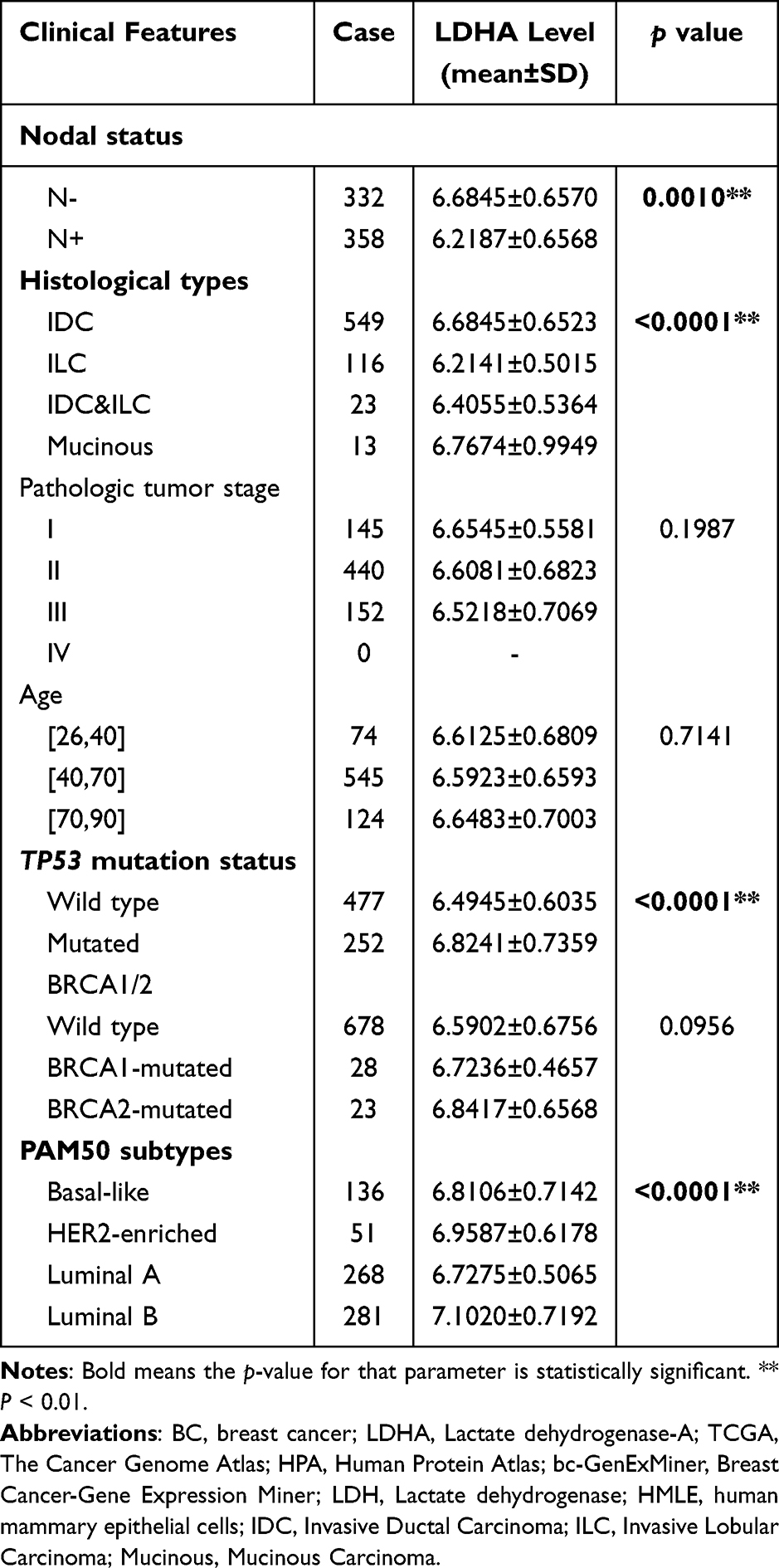 |
Table 1 Correlations Between the LDHA Expression and Clinical Characteristics of Patients with BC from the TCGA |
 |
Figure 2 Relationships between LDHA and clinical characteristics of BC patients. The expression of LDHA in BC patients and its relationship to clinical characteristics from the UALCAN database. (A), Expression of LDHA in BC based on nodal metastasis status; (B), Expression of LDHA in BC based on histological types; (C), Expression of LDHA in BC based on subclasses; (D), Expression of LDHA in BC based on TP53 mutation status. * P < 0.05, ** P < 0.01. |
LDHA is Highly Expressed in Most Breast Cancer Cells
To analyze the significance of LDHA in BC cells, we analyzed RNA-seq data from non-tumorigenic HMLE and BC cell lines available in the HPA databases. The HPA database revealed that BC cell lines exhibited higher levels of LDHA expression compared to HMLE (Figure 3A). Then we detected the basic LDHA protein expression of six types of BC cells by Western blot. Results showed that LDHA was expressed in the six types of cell lines. Meanwhile, there was relatively higher LDHA expression in ZR-75-30, SK-BR-3, MDA-MB-231 and MCF-7 compared to BT549 and T47D (Figure 3B, the detailed data are presented in Supplementary Figure 1). In addition, we detected the activities of LDH isoenzymes in six BC cells. Interestingly, LDH5 isoenzyme was found in six types of BC cells and occupied different proportions in different types of cells. BT549 had lower activity of LDH5 isoenzyme compared to other cell lines but with all five LDHs isoenzymes present (Figure 3C). Therefore, BT549 was chosen for further study.
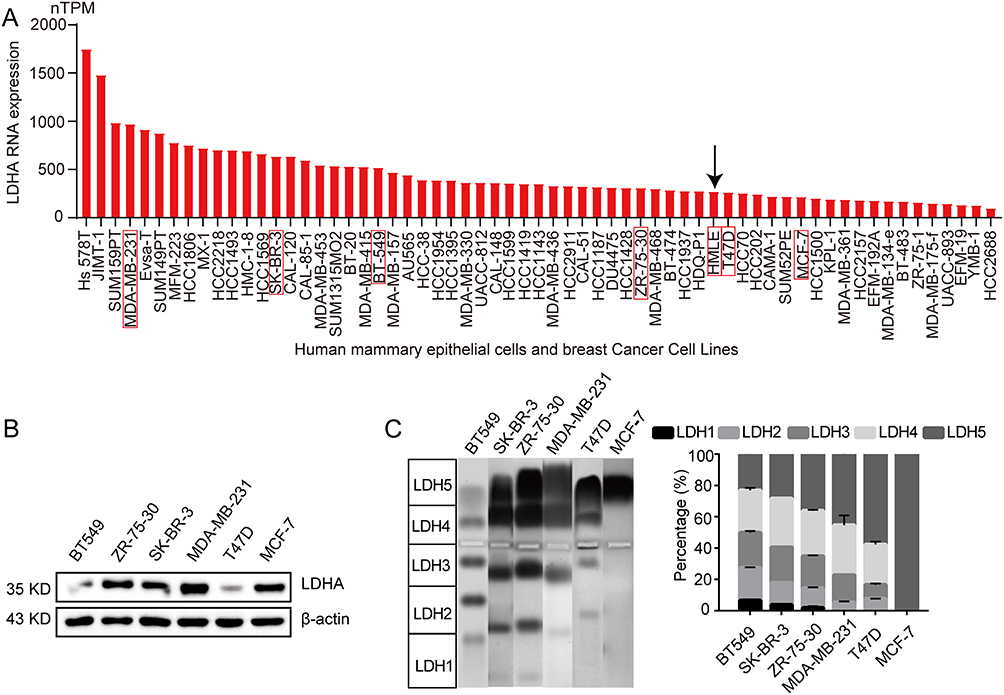 |
Figure 3 LDHA is highly expressed in most breast cancer cells. (A), RNA-seq data of LDHA in non-tumorigenic HMLE and various BC cell lines was analyzed by using HPA databases. The red box with the black arrow represented HMEL cells; (B), Western blot analysis of the expression of LDHA in six different BC cell lines; (C), Agarose gel electrophoresis assays of six different BC cell lines (n=3). Data are presented as the mean ± s.e.m. |
LDHA Overexpression Promotes Breast Cancer Cell Proliferation, Migration and Invasion
To explore the potential oncogenic roles of LDHA, we transfected BT549 cells with the LDHA-expressing plasmid. The result showed a significant increase in LDHA expression compared to the control group (Figure 4A, the detailed data are presented in Supplementary Figure 2). As we expected, overexpression of LDHA significantly increased cell proliferation and colony formation (Figure 4B and C). Wound healing is a process based on the interactions between cell proliferation and migration. Overexpression of LDHA significantly increased the capacity of wound healing in BT549 cells (Figure 4D). Additionally, MMP2 and MMP9 are part of a family of proteolytic enzymes responsible for remodeling the extracellular matrix. MMP2 and MMP9 can promote cancer cell migration and invasion by degradation of type IV collagen of basement membranes.24 It was found that overexpression of LDHA increased the activity of MMP-2 and MMP-9 indicating LDHA plays an important role in tumor migration and invasion (Figure 4E). Moreover, the number of migrating BT549 cells was significantly increased compared to the control group. (Figure 4F). Collectively, these findings demonstrated that the overexpression of LDHA promoted the progression of BC on cell proliferation, migration and invasion.
 |
Figure 4 LDHA overexpression promotes breast cancer cell proliferation, migration and invasion. (A), Western blot analysis of the expression of LDHA in BT549 cells transfected with LDHA-expressing plasmid; (B), Cell proliferation was determined by MTT assay after transfection 48 h (n=5); (C), Colony formation assay was performed to determine the colony forming ability in BT549 cells transfected with LDHA-expressing plasmid (n=3); (D), Cell proliferation and migration were assessed by wound healing assay; (E), The activity of MMP-2 and MMP-9 was assessed by gelatin zymography analysis; (F), Cell invasion capacity was assessed by transwell assay (n=3). Data are presented as the mean ± s.e.m. * P < 0.05, ** P < 0.01. |
Silencing of LDHA Inhibits Breast Cancer Cell Proliferation, Migration and Invasion
Then, we continued to evaluate the capacity of proliferation, migration and invasion in BC cells transfected with siRNAs (siLDHA-1 or siLDHA-2). The expression of LDHA in siRNAs groups was significantly reduced compared to the control group (Figure 5A, the detailed data are presented in Supplementary Figure 3). Silencing of LDHA significantly inhibited cell proliferation and colony formation (Figure 5B and C). In addition, silencing of LDHA significantly inhibited the capacity of wound healing in BT549 cells (Figure 5D). The activity of MMP-2 and MMP-9 were notably weakened by siRNAs transfection (Figure 5E). Moreover, transwell assay showed that the number of migrating BT549 cells was significantly reduced, and most of the cells were smaller compared to the control group (Figure 5F). Therefore, these data indicated that silencing of LDHA inhibited cell proliferation, migration and invasion.
 |
Figure 5 Silencing of LDHA inhibits breast cancer cell proliferation, migration and invasion. (A), Western blot analysis of the expression of LDHA in BT549 cells transfected with siRNA; (B), Cell proliferation was determined by MTT assay after siRNA transfection 48 h (n=5); (C), Colony formation assay was performed to determine the colony forming ability in BT549 cells transfected with siRNA (n=3); (D), Cell proliferation and migration were assessed by wound healing assay; (E), The activity of MMP-2 and MMP-9 was assessed by gelatin zymography analysis; (F), Cell invasion capacity was assessed by transwell assay (n=3). Data are presented as the mean ± s.e.m. ** P < 0.01. |
LDHA Expression is Associated with the Activity of LDH5 Isoenzyme
To further analyze the relationship between LDHA expression and LDH5 isoenzyme activity, we detected the activities of LDH isoenzymes. As we expected, overexpression or knockdown of LDHA expression mainly affected LDH5 isoenzymes. The LDH5 isoenzyme activity was prominently increased after LDHA overexpression (Figure 6A, the detailed data are presented in Supplementary Figure 4), while decreased after knockdown of LDHA (Figure 6B, the detailed data are presented in Supplementary Figure 5). LDH1 isoenzyme consisted of LDHB subunits. LDH2 isoenzyme contained three LDHB subunits and one LDHA subunit. Importantly, LDH1 and LDH2 were least affected when overexpression or knockdown of LDHA. However, the activities of LDH1 and LDH2 isoenzymes were increased after the knockdown of LDHA. We speculated that the relative increase in activities of LDH1 and LDH2 isoenzymes was due to the decrease of LDH5 isoenzyme. There were little changes in the activities of LDH3 and LDH4 isoenzymes. Therefore, these results indicated that LDHA expression determined the activity of the LDH5 isoenzyme in BC.
 |
Figure 6 LDHA expression is associated with the activity of LDH5 isoenzyme. (A), Agarose gel electrophoresis assay of BT549 cells transfected with LDHA-expressing plasmid (n=3); (B), Agarose gel electrophoresis assay of BT549 cells transfected with siRNAs (siLDHA-1 or siLDHA-2) (n=3). Data are presented as the mean ± s.e.m. * P < 0.05, ** P < 0.01. |
LDH5 Isoenzyme Activity is Higher in BC Patients
Once tissue was damaged, LDH is released into the circulation, contributing toward abnormal changes in the activities of LDH isoenzymes.25 However, the tissues and serum activities of LDH isoenzymes in BC remain unclear. To investigate the activities of LDH isoenzymes in BC patients, we used agarose gel electrophoresis assay to detect 12 pairs of BC tissues and adjacent normal breast tissues (Supplementary Figure 7A-J). The results showed that LDH isoenzyme in adjacent tissues consists of LDH1 to LDH4, and LDH5 isoenzyme was almost invisible. In contrast, LDH5 isoenzyme activity was detected in BC tissues and occupied a certain proportion (Figure 7A). Specifically, the activity of LDH5 isoenzyme in BC tissues increased to 18.84±8.07% compared to adjacent tissues (Figure 7B).
 |
Figure 7 LDH5 isoenzyme activity is higher in BC patients. (A), The activities of LDH isoenzymes in 12 pairs BC tissues and their adjacent normal breast tissues; (B), Comparison of LDH isoenzyme activity between BC tissues and adjacent normal breast tissues; (C), Representative Diagram of agarose gel electrophoresis assay of serum samples, all results (69 BC patients and 69 healthy controls) obtained from Supplementary Figure 7A-P; (D), Comparison of LDH isoenzyme activity and serum LDH levels between BC Patients and healthy controls; (E), The relative activity of LDH isoenzymes and serum LDH levels between BC Patients and healthy controls; (F), Kaplan–Meier Plotter database analysis of OS curves of patients with BC in relation to the LDHA levels with all subtypes (left) and with positive lymph node status (right). Data are presented as the mean ± s.e.m. ** P < 0.01. |
We further detected the activities of LDH isoenzymes in serum samples from 138 subjects (69 BC patients and 69 healthy controls) (Supplementary Figure 7A-P). The results demonstrated that serum LDH5 isoenzyme activity (13.41±3.05%) was higher in BC patients compared to healthy controls (12.07±2.13%), and the activity of the other isoenzymes had different degrees of decrease or increase (Figure 7C and D). Moreover, analysis of the statistical results showed that the increase of LDH5 isoenzyme activity was statistically significant (P<0.01) (Figure 7E). The experimental results of serum were similar to those of tissues. The above results indicated that LDHA played critical roles in BC, and the increased activity of LDH5 isoenzymes was induced by overexpression of LDHA in BC.
Compared to healthy controls, the serum LDH levels in BC patients were significantly increased (Figure 7E). However, the mean of serum LDH levels in BC patients was considered to be within the normal detection range (100–300 U/L) in clinical diagnosis, which suggested that LDH5 isoenzyme activity was a better biomarker than serum LDH levels for the diagnosis of BC. We also analyzed the Kaplan–Meier survival curves, BC patients with higher LDHA expression showed poor OS [hazard ratio (HR) = 2.14, p = 0.018]. In addition, in patients with positive lymph node status, high LDHA expression had a significantly negative effect on OS (HR = 3.38, p = 0.021) (Figure 7F). This indicated that LDHA was a risk factor for prognosis in patients with BC.
Discussion
Aerobic glycolysis is considered a key metabolic process involved in tumor growth.26 The molecular expression changes are closely related to metabolic reprogramming, which can be used to track the occurrence and development of tumors. Along this line, Much literature has revealed that LDHA is critically involved in tumor development.27,28 Our study showed that LDHA expression was significantly higher in BC tissues compared to normal tissues. The tumor subclass types of TNBC and patients with TP53 mutations tended to have higher expression of LDHA. TNBC is characterized by a high level of cell invasiveness and distant metastasis rates,29 which may indicate that high expression of LDHA is a poor prognostic factor in BC. Then we investigated the oncogenic role of LDHA in BC, the results showed that LDHA was highly expressed in most BC cells. Various small-molecule LDHA inhibitors and siRNA-mediated knockdown of LDHA have been reported to inhibit its LDH enzymatic activity, thereby suppressing glycolysis and tumor progression.30–33 Moreover, Juan Liu and colleagues reported that targeting LDHA activity and Rac1 activity simultaneously displays a promising therapeutic strategy for BC with LDHA overexpression.34 In our study, in vitro experiments demonstrated that overexpression of LDHA promoted cell proliferation, migration and invasion, while downregulation of LDHA suppressed proliferation migration and invasion in BC cells, indicated that LDHA has a tumor-promoting effect. Studies have demonstrated that LDH5 isoenzyme, encoded by LDHA, was considered as a vital therapeutic target for highly glycolytic human cancers. It was observed that overexpression of the LDH5 gene conferred the growth advantages of highly glycolytic cancer cells, which correlated with the aggressive phenotypes and poor prognosis in several tumors.35 However, few articles have reported on the role of LDH5 in BC. Our study revealed that the change in LDH5 isoenzyme activity was due to the expression of LDHA. Therefore, LDHA promotes the growth of BC cells by up-regulating the activity of LDH5 isoenzyme.
Moreover, the major type of LDH isoenzyme expressed in a particular tissue depends on its metabolic demands.36 It was the first time that the LDH5 isoenzyme activity was detected in BC tissues, and occupied nearly 19% proportion compare to adjacent tissues. Serum LDH5 isoenzyme activity was higher in BC patients compared to healthy controls. These findings intuitively demonstrated that the LDHA expression was significantly up-regulated in BC, and the increased activity of LDH5 isoenzymes was induced by overexpression of LDHA. Furthermore, the activities of LDH isoenzymes could help to identify the main source of tissue damage. LDH1 was predominantly expressed in myocardial damage, and LDH3 was predominantly expressed in pulmonary inflammation in both vascular and infectious pathologies.37 We found the change in the activity of LDH5 isoenzyme was most prominent, suggesting that LDH5 is useful for the diagnosis of BC.
LDH in the serum is widely used for the assessment of risk stratification and prognosis in many cancers compared to LDH5 isoenzyme activity.38–40 However, some studies have revealed serum LDH levels do not always reliably reflect serum LDH5 isoenzyme activity.41 In our study, we found some patients have been diagnosed with BC, but without abnormal LDH in the serum. However, serum LDH5 isoenzyme activity was significantly higher compared to healthy controls. These findings highlighted the importance of LDH5 isoenzyme in the BC diagnosis. In addition, Kaplan‒Meier analysis showed BC patients with high LDHA expression had poorer prognosis, which suggested that LDHA played an important role in the prognosis of BC.
Conclusions
In summary, we identified a new horizon to understand the role of LDHA in BC, suggesting the possibility that LDHA most likely promoted BT549 cell proliferation, migration and invasion by directly regulating LDH5 isoenzyme activity. The studies highlighted the importance of LDHA as an ideal target in BC, and a combination of LDHA expression and LDH5 isoenzyme activity can be used for early diagnosis and prognosis of BC.
Data Sharing Statement
The data that support the findings of this study are available on request from the corresponding author (Fancai Zeng, [email protected]).
Ethics Statement
The human samples used in this study conformed to national and institutional ethical guidelines and were approved by the Clinical Research Ethics Committee of Affiliated Hospital of Southwest Medical University (No. KY2021205). The studies were conducted in accordance with the local legislation and institutional requirements. All participants signed the informed consent.
Consent for Publication
All authors have consented to the publication of the manuscript.
Acknowledgments
All authors thank the TCGA databases for providing the useful data, and all the bioinformatics tools for data analysis. Thank you to the Breast Surgery Team of Affiliated Hospital of Southwest Medical University for supporting in the clinical work related to the article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Sichuan Science and Technology Program (Grant No. 2022YFS0623), the Youth Program of Southwest Medical University (Grant No. 2021ZKQN047) and the Undergraduate Training Program for Innovation and Entrepreneurship (Grant No. S202410632139, No. S202410632102).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Arnold M, Morgan E, Rumgay H, et al. Current and future burden of breast cancer: global statistics for 2020 and 2040. Breast. 2022;66:15–23. doi:10.1016/j.breast.2022.08.010
2. Montazeri Aliabadi H. Molecular targets for breast cancer therapy. Biomolecules. 2024;14(10):1219. doi:10.3390/biom14101219
3. Liu B, Qu X, Wang J, et al. LINC00365 functions as a tumor suppressor by inhibiting HIF-1α-mediated glucose metabolism reprogramming in breast cancer. Exp Cell Res. 2023;425(1):113514. doi:10.1016/j.yexcr.2023.113514
4. Wang X, Liu Z, Chu A, et al. Hsa_circ_0052611 and miR-767-5p guide the Warburg Effect, migration, and invasion of BRCA cells through modulating SCAI. J Bioenerg Biomembr. 2023;55(5):381–396. doi:10.1007/s10863-023-09985-4
5. Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–314. doi:10.1126/science.123.3191.309
6. Malhotra G, Gattani RG, Shinde RK, Gianchandani SG, Nayak K, Salwan A. Significance of serum lactate dehydrogenase as a prognostic marker and outcome predictor in patients with breast cancer. Cureus. 2024;16(3):e55932. doi:10.7759/cureus.55932
7. Urbańska K, Orzechowski A. Unappreciated role of LDHA and LDHB to control apoptosis and autophagy in tumor cells. Int J mol Sci. 2019;20(9):2085. doi:10.3390/ijms20092085
8. Thomas R, Shaath H, Naik A, Toor SM, Elkord E, Decock J. Identification of two HLA-A*0201 immunogenic epitopes of lactate dehydrogenase C (LDHC): potential novel targets for cancer immunotherapy. Cancer Immunol Immunother. 2020;69(3):449–463. doi:10.1007/s00262-020-02480-4
9. Lv M, Gong Y, Liu X, et al. CDK7-YAP-LDHD axis promotes D-lactate elimination and ferroptosis defense to support cancer stem cell-like properties. Signal Transduct Target Ther. 2023;8(1):302. doi:10.1038/s41392-023-01555-9
10. Sharma D, Singh M, Rani R. Role of LDH in tumor glycolysis: regulation of LDHA by small molecules for cancer therapeutics. Semin Cancer Biol. 2022;87:184–195. doi:10.1016/j.semcancer.2022.11.007
11. Yang Y, Gao Y, Xiong Y, et al. Research progress of Warburg effect in hepatocellular carcinoma. Front Biosci. 2024;29(5):178. doi:10.31083/j.fbl2905178
12. Gan X, Hu J, Pang Q, et al. LDHA-mediated M2-type macrophage polarization via tumor-derived exosomal EPHA2 promotes renal cell carcinoma progression. Mol, Carcinog. 2024;63(8):1486–1499. doi:10.1002/mc.23737
13. Khan F, Lin Y, Ali H, et al. LDHA-regulated tumor-macrophage symbiosis promotes glioblastoma progression. Res Sq. 2023;rs.3:rs–3401154. doi:10.21203/rs.3.rs-3401154/v1
14. Chen L, Xing X, Zhu Y, et al. Palmitoylation alters LDHA activity and pancreatic cancer response to chemotherapy. Cancer Lett. 2024;587:216696. doi:10.1016/j.canlet.2024.216696
15. Maeda M, Ko M, Mane MM, et al. Genetic and drug inhibition of LDH-A: effects on Murine Gliomas. Cancers. 2022;14(9):2306. doi:10.3390/cancers14092306
16. Augoff K, Hryniewicz-Jankowska A, Tabola R. Lactate dehydrogenase 5: an old friend and a new hope in the war on cancer. Cancer Lett. 2015;358(1):1–7. doi:10.1016/j.canlet.2014.12.035
17. Chen X, Liu L, Kang S, Gnanaprakasam JR, Wang R. The lactate dehydrogenase (LDH) isoenzyme spectrum enables optimally controlling T cell glycolysis and differentiation. Sci Adv. 2023;9(12):eadd9554. doi:10.1126/sciadv.add9554
18. Klein R, Nagy O, Tóthová C, Chovanová F. Clinical and diagnostic significance of lactate dehydrogenase and its isoenzymes in animals. Veterinary Med Int. 2020;2020:534648. doi:10.1155/2020/5346483
19. Jézéquel P, Campone M, Gouraud W, et al. bc-GenExMiner: an easy-to-use online platform for gene prognostic analyses in breast cancer. Breast Cancer Res Treat. 2012;131(3):765–775. doi:10.1007/s10549-011-1457-7
20. Jézéquel P, Frénel JS, Campion L, et al. bc-GenExMiner 3.0: new mining module computes breast cancer gene expression correlation analyses. Database. 2013;2013:bas060. doi:10.1093/database/bas060
21. Bartha Á, Győrffy B. TNMplot.com: a web tool for the comparison of gene expression in normal, tumor and metastatic tissues. Int J mol Sci. 2021;22(5):2622. doi:10.3390/ijms22052622
22. Battogtokh G, Obidiro O, Akala EO. Recent developments in combination immunotherapy with other therapies and nanoparticle-based therapy for triple-negative breast cancer (TNBC). Cancers. 2024;16(11):2012. doi:10.3390/cancers16112012
23. Su Y, Zhao J, Fu H, et al. TP53(R175H) mutation promotes breast cancer cell proliferation through CORO1A-P38 MAPK pathway regulation. Biochem Pharmacol. 2024;221:116047. doi:10.1016/j.bcp.2024.116047
24. Cabral-Pacheco GA, Garza-Veloz I, Castruita-De la Rosa C, et al. The roles of matrix metalloproteinases and their inhibitors in human diseases. Int J mol Sci. 2020;21(24). doi:10.3390/ijms21249739
25. Ergenc I, Capar E, Erturk SB, et al. Diagnostic performance of lactate dehydrogenase (LDH) isoenzymes levels for the severity of COVID-19. J Med Biochem. 2023;42(1):16–26. doi:10.5937/jomb0-37234
26. Nong S, Han X, Xiang Y, et al. Metabolic reprogramming in cancer: mechanisms and therapeutics. Med Comm. 2023;4(2):e218. doi:10.1002/mco2.218
27. Yang W, Wei Y, Wang T, et al. Cytoplasmic localization of SETDB1‑induced Warburg effect via c‑MYC‑LDHA axis enhances migration and invasion in breast carcinoma Cytoplasmic localization of SETDB1‑induced Warburg effect via c‑MYC‑LDHA axis enhances migration and invasion in breast carcinoma. Int J Mol Med. 2024;53(4):40. doi:10.3892/ijmm.2024.5364
28. Hou X, Ouyang J, Tang L, et al. KCNK1 promotes proliferation and metastasis of breast cancer cells by activating lactate dehydrogenase A (LDHA) and up-regulating H3K18 lactylation. PLoS Biol. 2024;22(6):e3002666. doi:10.1371/journal.pbio.3002666
29. Obidiro O, Battogtokh G, Akala EO. Triple negative breast cancer treatment options and limitations: future outlook. Pharmaceutics. 2023;15(7):1796. doi:10.3390/pharmaceutics15071796
30. Khajah MA, Khushaish S, Luqmani YA. Lactate Dehydrogenase A or B Knockdown Reduces Lactate Production and Inhibits Breast Cancer Cell Motility in vitro. Front Pharmacol. 2021;12:747001. doi:10.3389/fphar.2021.747001
31. Khajah MA, Khushaish S, Luqmani YA. The effect of lactate dehydrogenase inhibitors on proliferation, motility and invasion of breast cancer cells in vitro highlights a new role for lactate. Mol Med Rep. 2024;29(1):12. doi:10.3892/mmr.2023.13135
32. Feng Y, Xiong Y, Qiao T, Li X, Jia L, Han Y. Lactate dehydrogenase A: a key player in carcinogenesis and potential target in cancer therapy. Cancer Med. 2018;7(12):6124–6136. doi:10.1002/cam4.1820
33. Woodford MR, Chen VZ, Backe SJ, Bratslavsky G, Mollapour M. Structural and functional regulation of lactate dehydrogenase-A in cancer. Future Med Chem. 2020;12(5):439–455. doi:10.4155/fmc-2019-0287
34. Liu J, Zhang C, Zhang T, et al. Metabolic enzyme LDHA activates Rac1 GTPase as a noncanonical mechanism to promote cancer. Nat Metab. 2022;4(12):1830–1846. doi:10.1038/s42255-022-00708-4
35. Nadal-Bufí F, Chan LY, Mohammad HH, et al. Peptide-based LDH5 inhibitors enter cancer cells and impair proliferation. Cell mol Life Sci. 2022;79(12):606. doi:10.1007/s00018-022-04633-3
36. Comandatore A, Franczak M, Smolenski RT, Morelli L, Peters GJ, Giovannetti E. Lactate Dehydrogenase and its clinical significance in pancreatic and thoracic cancers. Semin Cancer Biol. 2022;86(Pt 2):93–100. doi:10.1016/j.semcancer.2022.09.001
37. Serrano-Lorenzo P, Coya ON, López-Jimenez A, et al. Plasma LDH: a specific biomarker for lung affectation in COVID-19? Pract Lab Med. 2021:
38. Fujibuchi T, Imai H, Kidani T, Morino T, Miura H. Serum lactate dehydrogenase levels predict the prognosis of patients with soft tissue sarcoma. Mol Clin Oncol. 2022;16(3):65. doi:10.3892/mco.2022.2498
39. Gu S, Yang C. Serum lactate dehydrogenase level predicts the prognosis in bladder cancer patients. BMC Urol. 2023;23(1):65. doi:10.1186/s12894-023-01239-0
40. Huang Q, Li S, Chen X, et al. Association between serum lactate dehydrogenase and lymph node metastasis in cervical cancer. Oncol Lett. 2023;26(5):482. doi:10.3892/ol.2023.14069
41. Koukourakis MI, Giatromanolaki A. Warburg effect, lactate dehydrogenase, and radio/chemo-therapy efficacy. Int J Radiat Biol. 2018;95(4):408–426. doi:10.1080/09553002.2018.1490041
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
A Novel lncRNA Panel for Risk Stratification and Immune Landscape in Breast Cancer Patients

Li C, Wang X, Chen T, Li W, Yang Q
International Journal of General Medicine 2022, 15:5253-5272
Published Date: 27 May 2022
Establishment and Validation of a Model for Disease-Free Survival Rate Prediction Using the Combination of microRNA-381 and Clinical Indicators in Patients with Breast Cancer
Shen J, Wang M, Li F, Yan H, Wang R, Zhou J
Breast Cancer: Targets and Therapy 2022, 14:375-389
Published Date: 30 November 2022
Chromobox Family Proteins as Putative Biomarkers for Breast Cancer Management: A Preliminary Study Based on Bioinformatics Analysis and qRT-PCR Validation
Tian H, Zhao T, Li Y, Sun N, Ma D, Shi Q, Zhang G, Chen Q, Zhang K, Chen C, Zhang Y, Qi X
Breast Cancer: Targets and Therapy 2022, 14:515-535
Published Date: 30 December 2022
Prognostic Significance of Preoperative Lactate Dehydrogenase to Albumin Ratio in Breast Cancer: A Retrospective Study
He J, Tong L, Wu P, Wu Y, Shi W, Chen L
International Journal of General Medicine 2023, 16:507-514
Published Date: 8 February 2023
The Prognostic Role of HuR Varies Between Different Subtypes of Breast Cancer Patients: Data Mining and Retrospective Analysis
Liao Y, Liao Y, Li J, Li Y, Fan Y
Breast Cancer: Targets and Therapy 2023, 15:135-146
Published Date: 11 February 2023