Back to Journals » Journal of Inflammation Research » Volume 18
Age-Stratified Immune-Inflammatory Profiles and Prognostic Implications of Old Patients in the ICU: A Prospective Study
Authors Lian H, Wang G, Zhang Q, Zhao H, Wang X
Received 21 July 2025
Accepted for publication 17 September 2025
Published 3 October 2025 Volume 2025:18 Pages 13727—13743
DOI https://doi.org/10.2147/JIR.S548582
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Hui Lian,1,* Guangjian Wang,1,* Qing Zhang,2 Hua Zhao,2 Xiaoting Wang2
1Department of Health Care, State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing City, People’s Republic of China; 2Department of Critical Care Medicine, State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing City, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hua Zhao; Xiaoting Wang, Department of Critical Care Medicine, State Key Laboratory of Complex Severe and Rare Diseases, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, No. 1 Shuaifuyuan, Dongcheng District, Beijing City, 100730, People’s Republic of China, Email [email protected]; [email protected]
Background: The aging population has advanced older patients (OPs) in intensive care units (ICUs). Critical illness and aging exacerbate immune-inflammatory dysregulation, impairing immune interactions and worsening prognosis. However, knowledge regarding immune-inflammatory profiles and prognostic implications for OPs, particularly in very old patients (VOPs), remains limited.
Methods: This single-center prospective study included patients aged ≥ 65 with hemodynamically confirmed shock, signifying severe critical illness admitted between August 2023 and February 2025. Demographic, medication, and laboratory data were collected from electronic medical records. The primary endpoint was in-hospital mortality; secondary endpoints included ICU and hospital stay duration. Participants aged 65– 74 were young old patients (YOPs), and those aged ≥ 75 as VOPs. Statistical analysis included χ² for categorical variables, Kolmogorov–Smirnov for non-normal continuous data, Principal component analysis (PCA) for inflammatory states, while generalized additive mixed models for statistical interactions.
Results: A total of 537 OPs were admitted, with no significant differences in inflammatory markers between YOPs and VOPs, except fibrinogen. Survivors had higher levels of hypersensitive C-reactive protein (hsCRP). Lymphocyte counts and subtypes were reduced in OPs. VOPs showed higher natural killer cells, CD8+CD38+, and CD8+DR+ counts versus YOPs. The median CD8+DR+ count was 82 in survivors versus 59 in non-survivors. Among VOPs, hsCRP, interleukin-8, and immunoglobulin G showed significant differences. B cell count was lower (median 104 vs 72), and CD8+ T cell activation declined in non-survivors. Mortality was higher in the low inflammatory state group. As B and T cell counts increased, mortality decreased in high inflammatory states. Higher CD8+DR+ counts reduced mortality.
Conclusion: OPs, especially VOPs, with hemodynamically confirmed shock and critical illness exhibit distinct immune-inflammatory characteristics affecting prognosis. A weakened immune response in low inflammation links to poor outcomes. Enhancing B cell and CD8+DR+ T cell responses may improve prognosis through precise immune regulation in ICU settings.
Keywords: old patient, critical illness, immune, inflammatory, interactions, lymphocyte subset, prognosis
Introduction
The global population is aging at an unprecedented rate, with an estimated increase of over 400 million individuals aged ≥65 years between 2010 and 2030.1,2 This demographic shift has led to a substantial rise in the proportion of older patients (OPs) admitted to intensive care units (ICUs). OPs with critical illnesses often present with multiple organ dysfunction, chronic comorbidities, and immune system impairments, resulting in significantly poorer clinical outcomes and prognosis compared to younger patients.3 Consequently, a deeper investigation into the underlying pathological mechanisms is urgently needed to improve treatment strategies for this vulnerable population.
Critical illness is frequently characterized by a dynamic imbalance in immune and inflammatory responses. During the acute phase, excessive inflammation can cause endothelial injury and multiple organ failure, while subsequent compensatory immunosuppression may lead to lymphocyte exhaustion and an increased risk of secondary infections.4,5 Aging further intensifies these processes through two key mechanisms: immunosenescence and inflammaging.
Immunosenescence refers to age-related alterations in the immune system that compromise both innate and adaptive immunity, resulting in weakened pro-inflammatory and autoimmune responses.6 These changes affect nearly all lymphocyte subsets, with T lymphocytes being particularly impacted.7 Previous research has shown that aging is associated with an inverted CD4/CD8 ratio and the loss of CD28 expression on T cells—markers of immunological aging and functional decline.8,9
Furthermore, immunosenescence increases susceptibility to infections, cancer, neurodegenerative diseases, and autoimmune disorders in the elderly population,10,11 thereby elevating mortality risk and drawing significant clinical attention. Inflammaging a phenomenon characterized by elevated baseline levels of pro-inflammatory cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) leads to a chronic low-grade inflammatory state. This persistent inflammation impairs tissue repair and exacerbates organ damage, further compromising recovery in older patients.12 As a result, aging not only alters the immune and inflammatory profiles of OPs with critical illnesses but also contributes to disease progression and poor prognosis.
Importantly, the immune and inflammatory systems do not operate independently but interact in complex and dynamic ways.13 For instance, IL-6 can amplify inflammation by activating specific T cell subsets while concurrently promoting immunosuppression through the expansion of regulatory T cells.14 Similarly, interferon-γ (IFN-γ), secreted by natural killer (NK) cells and T cells, plays a crucial role in antiviral defense but may induce tissue toxicity when excessively activated.15
In OPs with critical illnesses, the combination of immunosenescence and persistent low-grade inflammation results in intricate immune-inflammatory interactions, contributing to varied and often unpredictable treatment responses. However, most existing studies have focused on isolated immune or inflammatory markers, lacking a comprehensive assessment of their dynamic interplay. This gap is particularly evident in research involving very old patients (VOPs, ≥75 years), where systematic evaluations of immune-inflammatory interactions in critical illness remain limited.
Lymphocyte subsets play an important role in maintaining the function of the immune system and are also core indicators for evaluating immune function.16 Inflammatory markers such as hypersensitive C-reactive protein (hsCRP), procalcitonin (PCT), IL-6, IL-8, and IL-10 can quantify the inflammatory status. The combined detection of these indicators can effectively reflect the inflammatory and immune characteristics of OPs in the ICU, thereby laying a theoretical foundation for exploring the pathological mechanisms involved in the occurrence and progression of critical illnesses in this population and for identifying potential clinical treatment strategies. Furthermore, shock, a common complication in critically ill patients, leads to marked hemodynamic instability and reduced tissue perfusion, resulting in organ dysfunction. These physiological disturbances exacerbate immune-inflammatory dysregulation, reflecting the severity of the studied cohort.17 This prospective study aimed to: compare age-stratified differences in lymphocyte subsets and inflammatory markers between young old patients (YOPs, 65–74 years) and VOPs (≥75 years) in the ICU; screen the indicators that might be related to prognosis; and decipher immune-inflammatory interactions using principal component analysis (PCA), with a focus on their combined impact on prognosis.
Methods and Materials
Study Design and Research Population
This study is a single-center prospective study conducted at Peking Union Medical College Hospital, a top tertiary hospital in China. Patients admitted to the ICU aged 65 years or older with confirmed shock were included between August 2023 and February 2025. Patients without lymphocyte and subset analysis results or those who refused to provide written informed consent (either personally or via their relatives) were excluded. A total of 537 OPs admitted to the ICU were ultimately included in the study. Figure 1 shows the flowchart of the study procedure. All clinical data and analytical methods are well preserved. The corresponding author will provide the materials upon reasonable request. The study protocol was approved by the local ethics committee (Ethical Approval Number: I-23PJ1278).
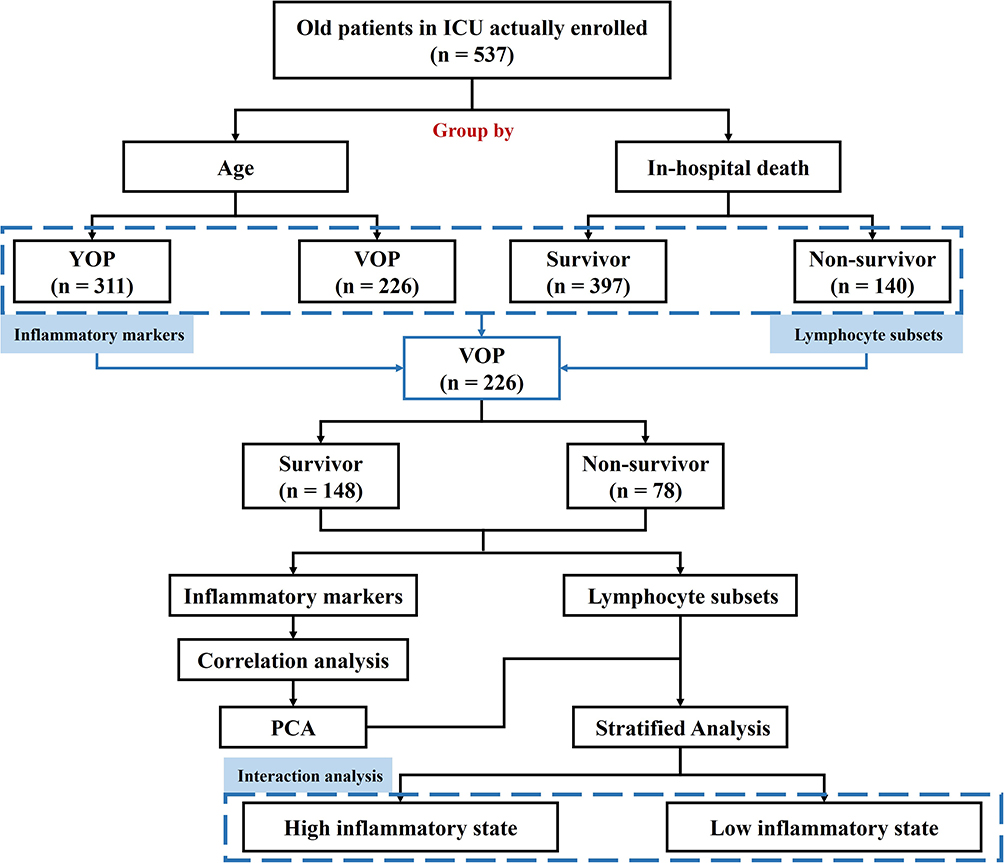 |
Figure 1 Flowchart of patient enrollment and study design. Abbreviations: ICU, intensive care unit; YOP, young old patient, 65–74 years; VOP, very old patients, ≥75 years; PCA, principal component analysis. |
Data Collection
We obtained demographic, medication, and laboratory data from the electronic medical record system. Medication data included past medical history, ICU parameters, and outcomes. Medical history encompassed information on surgery, hypertension, coronary heart disease, diabetes mellitus, autoimmune diseases, immunotherapy, and tumors. ICU parameters mainly included the Acute Physiology and Chronic Health Evaluation (APACHE) II score, ventilation time, and other relevant clinical indicators. Laboratory data included inflammatory biomarkers (IL-6, IL-8, IL-10, TNF-α, complement 3, complement 4, fibrinogen, D-dimer, PCT, hsCRP, and erythrocyte sedimentation rate18), immunoglobulins (immunoglobulin G [IgG], IgA, IgM), as well as lymphocyte counts and subset analysis results. The patient samples were all stored and sourced from the Clinical Biobank (ISO 20387), Peking Union Medical College Hospital, Chinese Academy of Medical Sciences. All the above parameters were recorded at the time of ICU admission.
The primary endpoint was in-hospital mortality, while the secondary endpoints included ICU stay duration and total length of hospital stay.
Statistical Analysis
All continuous data in this study were expressed using the minimum, 25th percentile, median, 75th percentile, and maximum values after confirming a skewed distribution. The distributions of continuous variables were assessed for normality using the Kolmogorov–Smirnov test. Categorical variables were presented as numbers (percentages). Participants were divided into two groups based on age: young old patients (YOPs, 65–74 years) and very old patients (VOPs, ≥75 years). The χ²-test was used to compare categorical variables. Continuous variables with a non-normal distribution were analyzed using the Kolmogorov–Smirnov test. Given the correlations among inflammatory markers, the principal component analysis (PCA) method was employed to identify principal components and evaluate inflammatory states. A generalized additive mixed model was used to construct smooth curves to quantify statistical interactions. Interaction tests were also performed to assess significance. A two-tailed P < 0.05 was considered statistically significant for all analyses. Statistical analyses were conducted using EmpowerStats (http://www.empowerstats.com) and R software, version 4.2.0 (http://www.R-project.org/).
Results
Baseline Characteristics and Clinical Outcomes
A total of 537 OPs were admitted to the ICU during the study period (Figure 1). The baseline characteristics of the participants are summarized in Table 1. The median age of the study population was 73 years, with 42.09% aged 75 years or older. The proportion of male patients was significantly higher than that of females. Overall, 43.8% of the participants underwent surgery; however, the proportion of surgical procedures was significantly lower among VOPs. The prevalence of hypertension increased significantly with age, while the prevalence of other chronic diseases did not differ significantly between age groups. The proportion of patients receiving immunotherapy was lower in VOPs. APACHE II scores were higher in VOPs, indicating more severe illness. The average length of hospital stay for all participants was 25.2 days, with VOPs having a significantly longer stay (30.0 days). ICU stays were also significantly prolonged in this group. The overall in-hospital mortality rate among OPs was 26.1%, with significantly higher mortality observed in VOPs, reaching 34.5%.
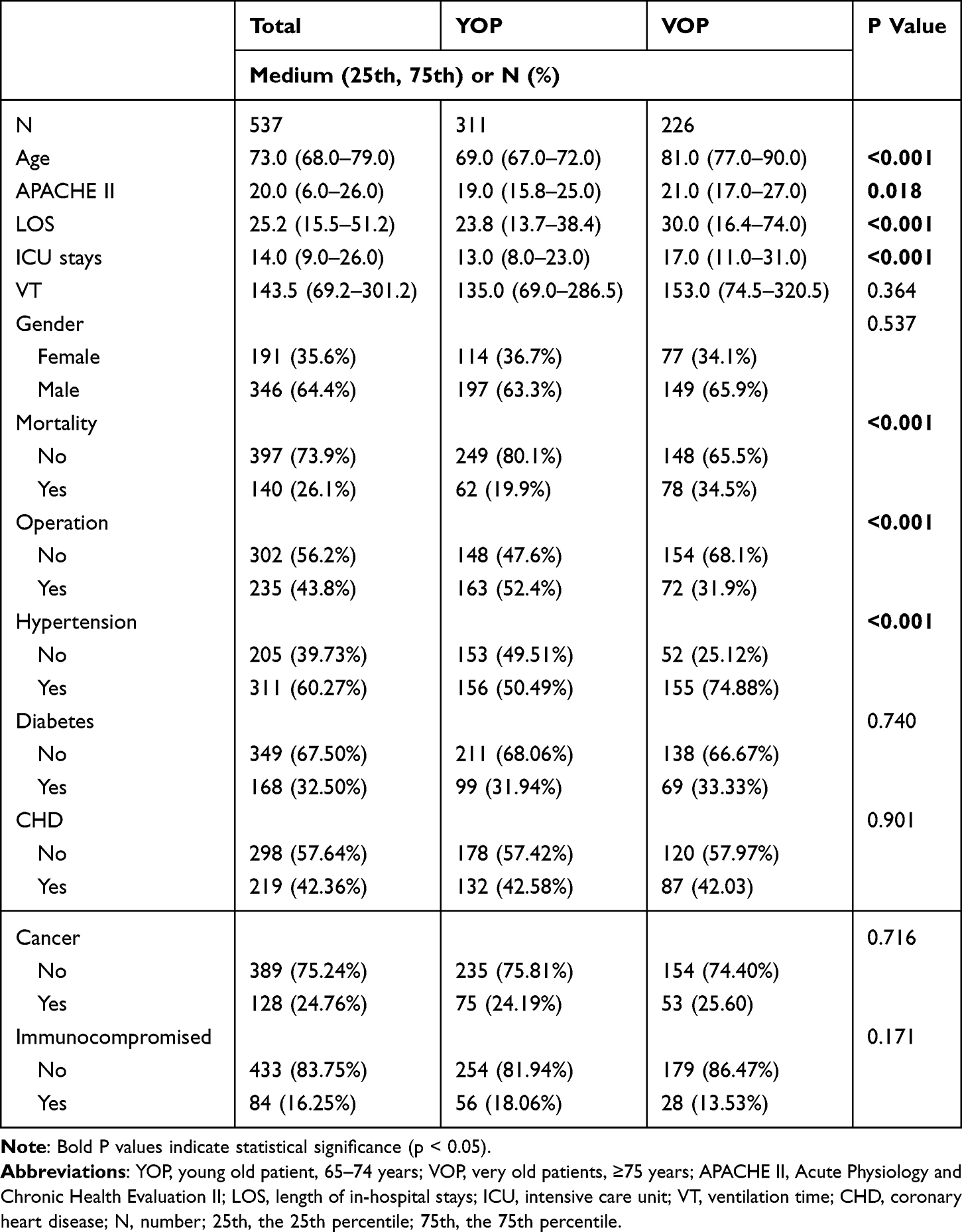 |
Table 1 Demographic Characteristics, Medical Data and Outcomes of the Study Population |
Inflammatory Biomarkers, Complements, and Immunoglobulins
Table 2 presents a comparison of inflammatory biomarkers, complement levels, and immunoglobulins between the YOP (65–74 years) and VOP (≥75 years) groups. Overall, there were no statistically significant differences in most inflammatory markers between the two groups. The levels of procalcitonin (PCT), erythrocyte sedimentation rate (ESR), hypersensitive C-reactive protein (hsCRP), D-dimer, IL-6, IL-8, IL-10, and TNF-α showed no significant intergroup differences (all P > 0.05). Similarly, complement components (C3 and C4) and immunoglobulin levels (IgG, IgA, and IgM) were also comparable between YOPs and VOPs. However, a statistically significant difference was observed in fibrinogen (FBG) levels. VOPs had higher median FBG levels [3.7 (2.5–4.7) g/L] compared to YOPs [3.0 (2.2–4.5) g/L], with a P value of 0.026, indicating a modest increase in fibrinogen levels with advancing age.
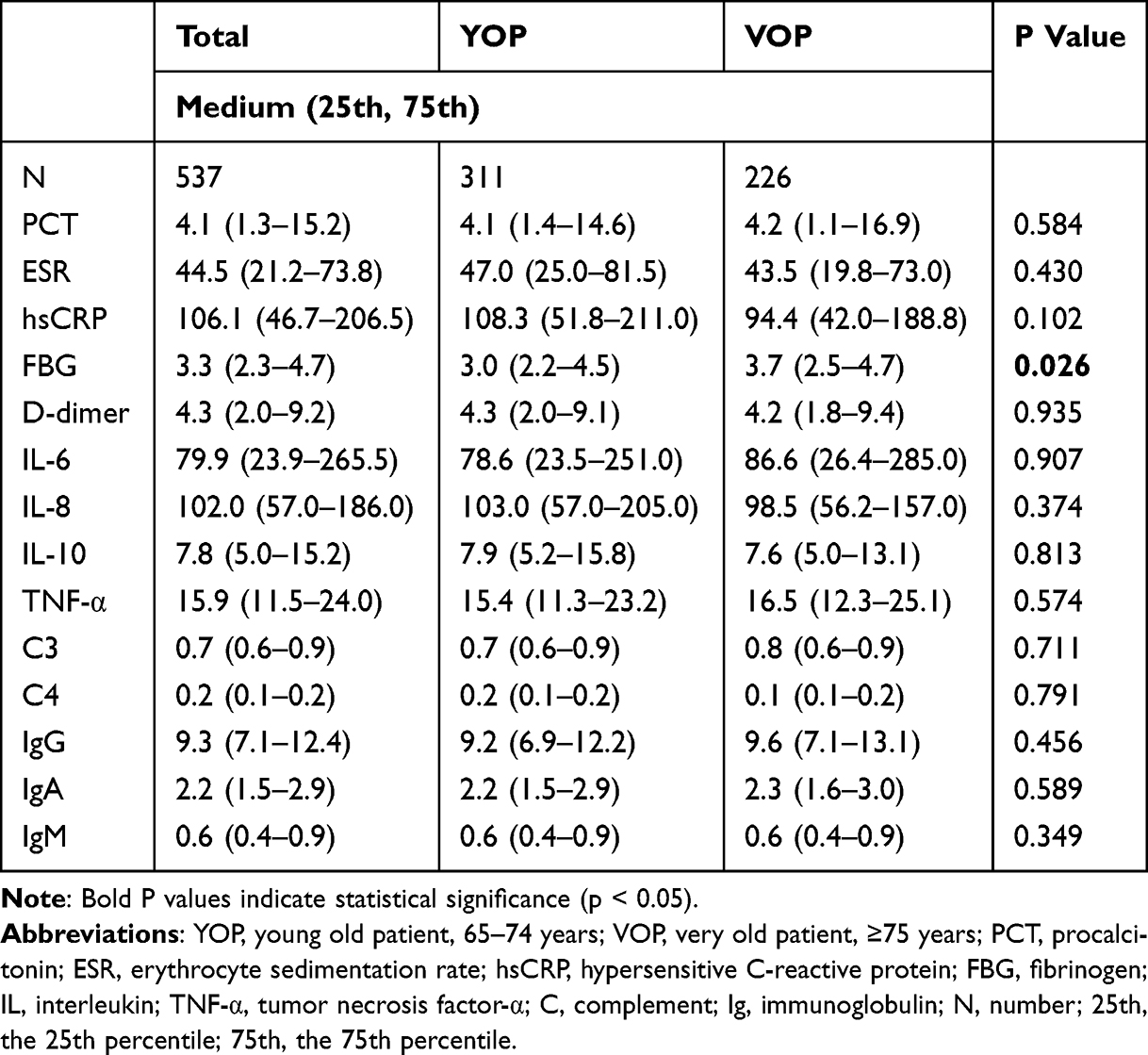 |
Table 2 Inflammatory Biomarkers, Complements and Immunoglobulins Between YOP and VOP |
Lymphocyte Counts and Subsets
Among the OPs, both total lymphocyte counts and their subsets—including B cells, natural killer (NK) cells, and T cells—were decreased (Table 3). The median lymphocyte count was 0.74 × 109/L, slightly below the lower limit of the normal reference range (0.8–4.0 × 109/L). The proportion of lymphocytes also showed a marked decline, with a median of 7.25%, compared to the normal range of 20%–40%. This lymphopenia was more pronounced in YOPs.
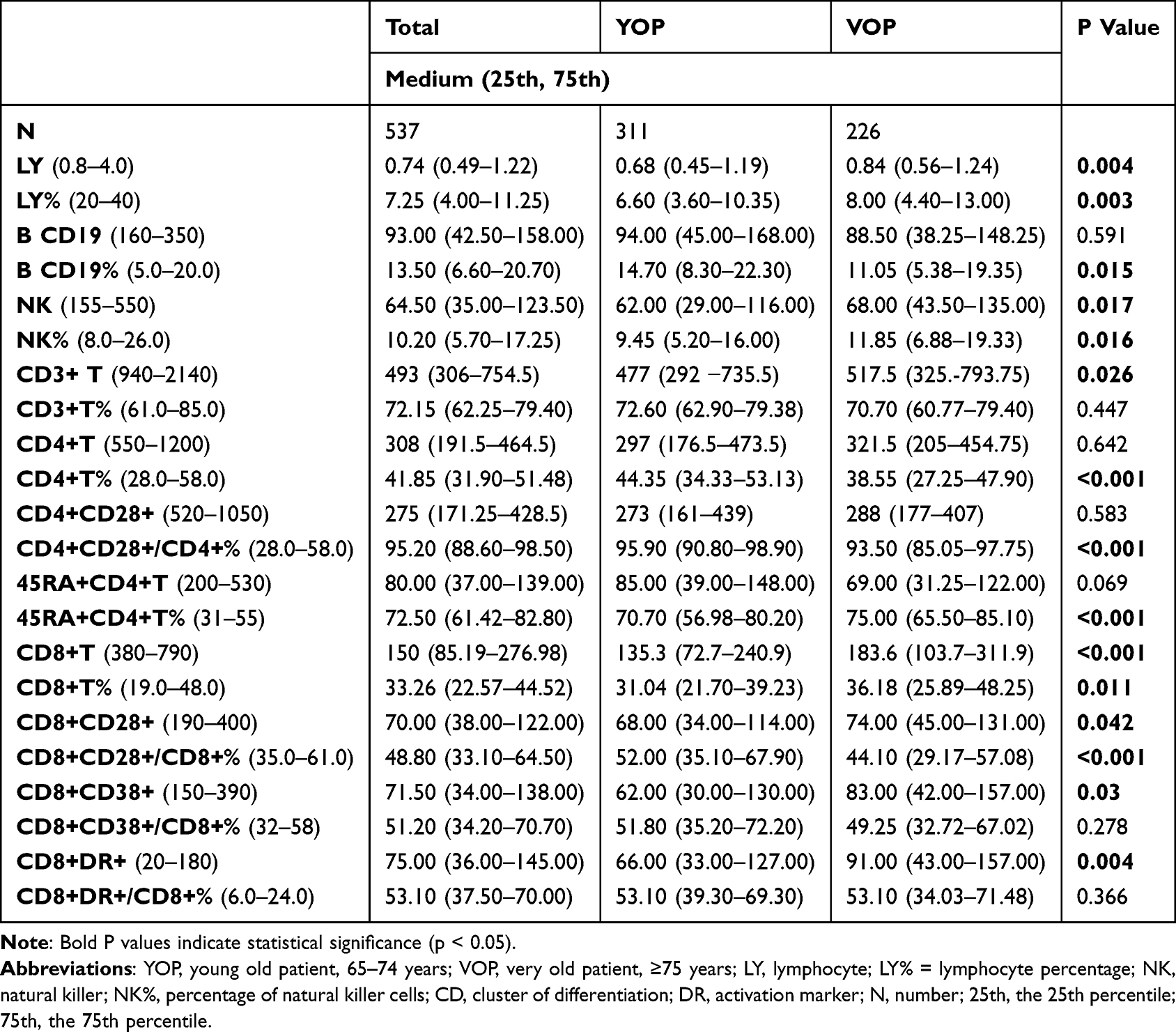 |
Table 3 Characteristics of Lymphocytes and the Subsets Between YOP and VOP |
Most lymphocyte subsets were within normal ranges, except for elevated proportions of CD4+CD28+ and CD8+DR+ T cells. In VOPs, these immune characteristics were more notable Both the absolute count and the proportion of NK cells were significantly higher in VOPs compared to YOPs. The activation subsets CD8+CD38+ and CD8+DR+ also showed significantly increased counts in VOPs, although the proportional differences were not statistically significant. While total lymphocyte counts were slightly higher in VOPs than in YOPs, B cell counts were reduced. NK cell counts were marginally higher, but a more substantial increase was observed in T cell counts. Further analysis of T cell subsets indicated that the rise in T cell counts was mainly driven by a significant increase in CD8+ T cells. Additionally, both functional (CD8+CD28+) and activation (CD8+CD38+ and CD8+DR+) subsets of CD8+ T cells were elevated in VOPs.
Inflammatory Markers in Survivor and Non-Survivor Groups
Based on in-hospital outcomes, patients were divided into a survivor group and a non-survivor group. Further analysis revealed notable differences in inflammatory markers between the two groups (Table 4). Survivors had significantly higher hsCRP levels compared to non-survivors. The median hsCRP level in the survivor group was 109.1 mg/L (interquartile range: 52.3–217.3), whereas the median level in the non-survivor group was 85.4 mg/L. Several other inflammatory markers also showed statistically significant differences between the groups, including tumor necrosis factor-α (TNF-α), D-dimer, immunoglobulin G (IgG), and immunoglobulin M (IgM). In contrast to hsCRP, the median levels of TNF-α, D-dimer, IgG, and IgM were slightly higher in the non-survivor group than in the survivor group.
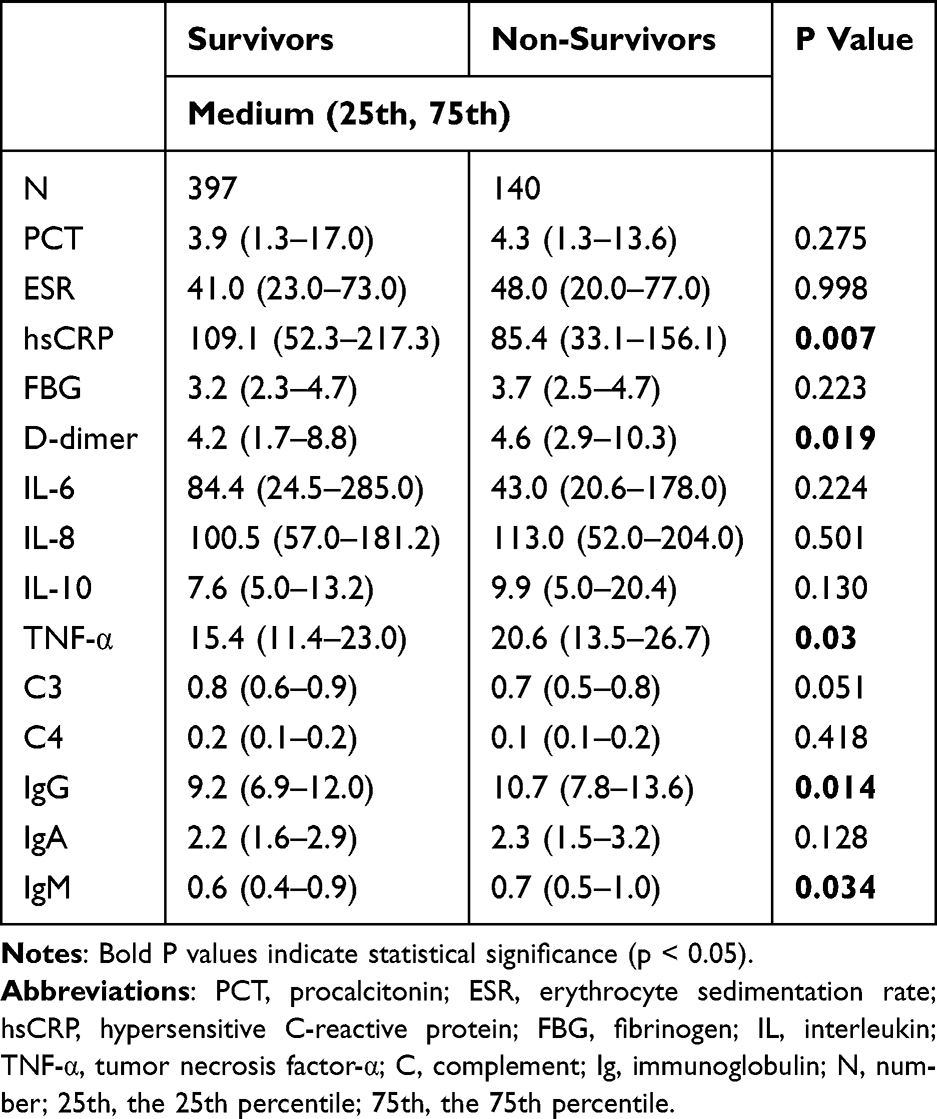 |
Table 4 Inflammatory Biomarkers, Complements and Immunoglobulins Between Survivors and Non-Survivors |
Lymphocyte Subsets in Survivors vs Non-Survivors
When comparing lymphocyte subsets between survivors and non-survivors, several significant differences were observed (Table 5). The B cell count was notably higher in the survivor group, with a median of 101.00 compared to 75.50 in the non-survivor group (P = 0.003). The B cell percentage also showed a statistically significant difference, with survivors having a median of 14.10% versus 11.00% in non-survivors (P = 0.009). Similarly, both NK cell count and T cell count were significantly reduced in non-survivors. The median NK cell count was 68.00 in survivors versus 50.00 in non-survivors (P = 0.006). T cell (CD3+) count was also higher in survivors (median: 513.50) than in non-survivors (419.50; P = 0.010). Further analysis of T cell subsets revealed no significant differences in the absolute counts of CD4+ and CD8+ T cells between the two groups. However, differences emerged in functional and activation markers. The CD4+ T cell percentage was significantly higher in survivors (median: 42.70%) compared to non-survivors (39.20%; P = 0.002). CD8+CD28+/CD8+% was also higher in the survivor group (median: 50.05%) than in non-survivors (45.20%; P = 0.033). Interestingly, while the absolute count of CD8+CD38+ cells was similar between groups, the proportion (CD8+CD38+/CD8+%) was significantly higher in non-survivors (P = 0.003), suggesting overactivation of this subset in poorer outcomes. The CD8+DR+ count was significantly greater in survivors (82.00) compared to non-survivors (59.00; P = 0.028), though the percentage difference was not statistically significant.
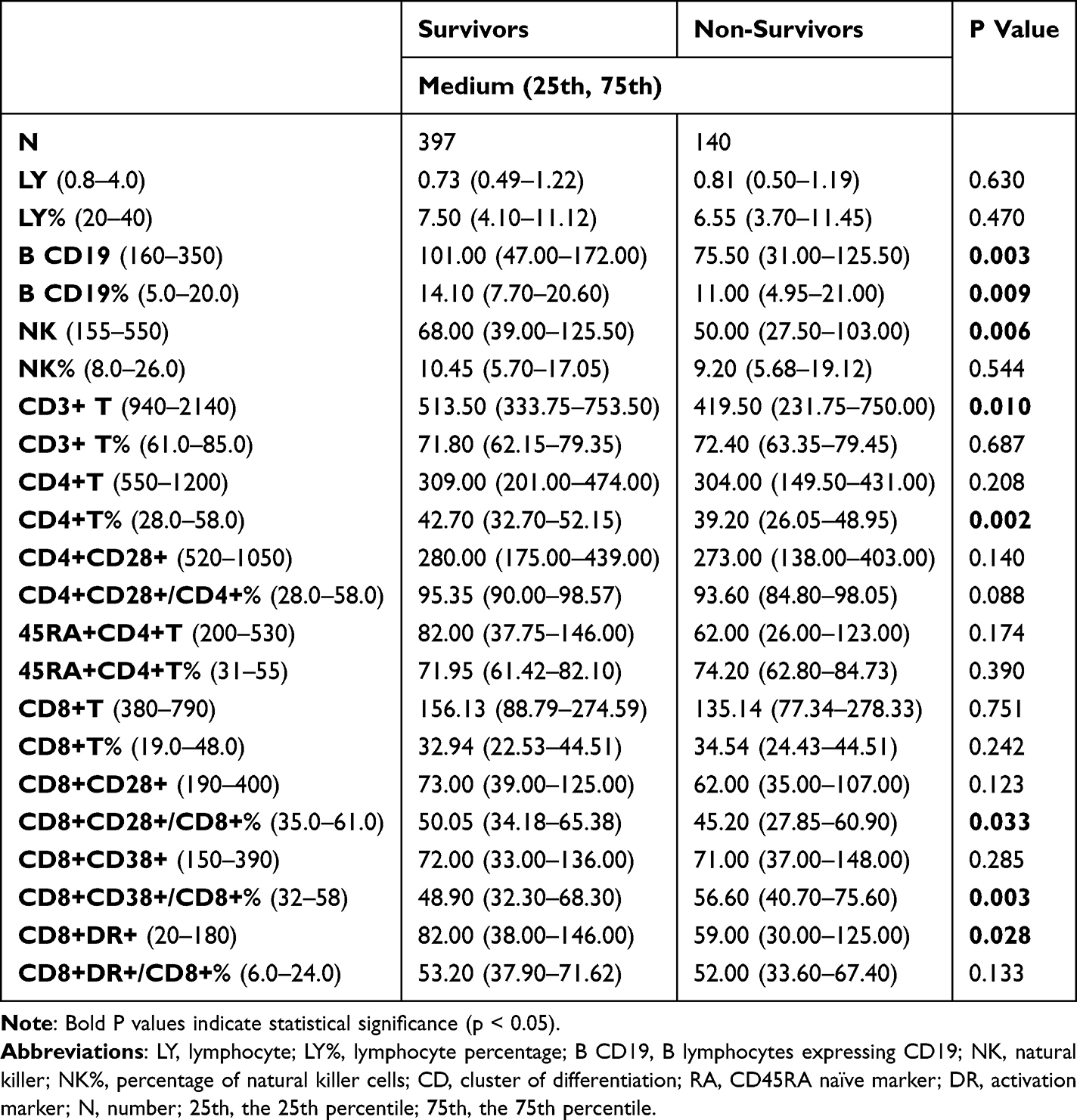 |
Table 5 Characteristics of Lymphocytes and the Subsets Between Survivors and Non-Survivors |
These findings indicate that better immune cell preservation and specific functional T cell subset profiles, particularly involving B cells and CD8+DR+ cells, are associated with improved in-hospital outcomes.
Inflammatory Markers in Survivors vs Non-Survivors Among VOPs
To further explore the influence of aging on critical illness, a subgroup analysis was conducted among VOPs (≥75 years). This analysis aimed to identify specific inflammatory and immune indicators associated with prognosis in this advanced age group. As shown in Table 6, only three markers hsCRP, IL-8, and IgG demonstrated statistically significant differences between survivors and non-survivors. Non-survivor VOPs exhibited lower hsCRP levels compared to survivors. The median hsCRP level in survivors was 107.1 mg/L, whereas it was reduced to 73.2 mg/L in non-survivors (P = 0.039). In contrast, IL-8 and IgG levels were significantly elevated in the non-survivor group. IL-8 had a median of 165.0 pg/mL in non-survivors versus 89.0 pg/mL in survivors (P = 0.004), while IgG levels were 12.1 g/L in non-survivors compared to 9.3 g/L in survivors (P = 0.001). No significant differences were observed for other inflammatory biomarkers, complements (C3, C4), or immunoglobulins (IgA, IgM) between the two groups. These findings suggest that in VOPs, specific dysregulations in inflammatory response particularly involving hsCRP, IL-8, and IgG may be closely associated with adverse outcomes.
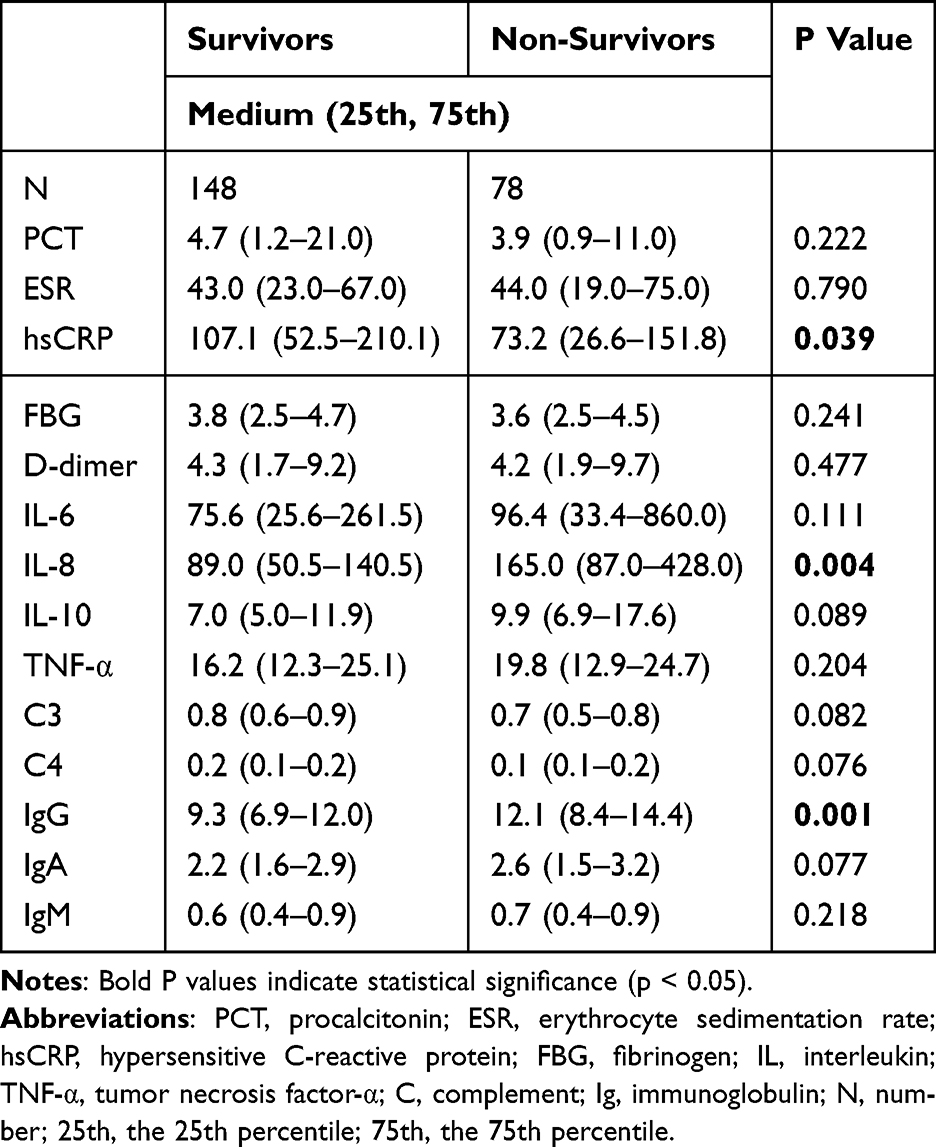 |
Table 6 Inflammatory Biomarkers, Complements and Immunoglobulins Between Survivors and Non-Survivors in VOPs |
Lymphocyte Subsets and Activation Markers in VOPs by Survival Status
To further elucidate the immune profile of very old patients (VOPs), T and B lymphocyte subsets were analyzed in relation to survival outcomes (Table 7). The results revealed important distinctions in immune status between survivors and non-survivors. B cell counts were significantly lower in non-survivors compared to survivors, with median values of 72.00 and 104.00, respectively (P = 0.020). The B cell proportion also differed significantly between the groups (7.00% vs 13.30%, P = 0.013), indicating a reduced humoral immune capacity in the non-survivor group.
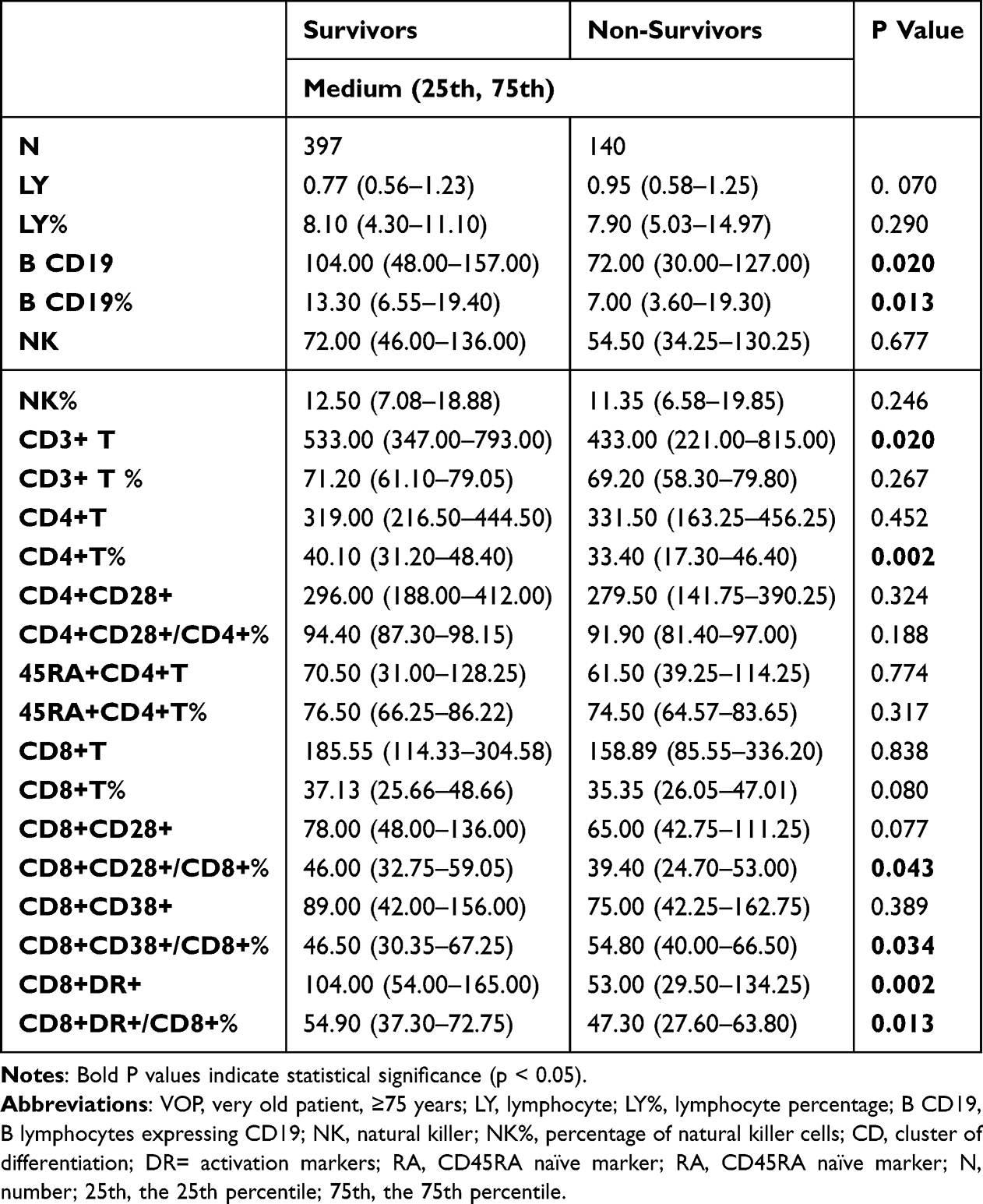 |
Table 7 Characteristics of Lymphocytes and the Subsets Between Survivors and Non-Survivors in VOPs |
Although NK cell and total lymphocyte counts were not significantly different, T lymphocyte (CD3+) counts were lower in non-survivors (median: 433.00) than in survivors (median: 533.00; P = 0.020). A marked reduction was observed in CD4+ T cell percentages in non-survivors (33.40%) compared to survivors (40.10%; P = 0.002). In terms of T cell function and activation, CD8+CD28+/CD8+% was significantly reduced in the non-survivor group (39.40%) versus the survivor group (46.00%; P = 0.043), suggesting weakened cytotoxic T cell function. Conversely, the CD8+CD38+/CD8+% was significantly higher in non-survivors (P = 0.034), indicating potential overactivation or exhaustion.
Importantly, CD8+DR+ T cell counts, an indicator of T cell activation, showed a dramatic decline in non-survivors (53.00) compared to survivors (104.00; P = 0.002). The proportion of CD8+DR+/CD8+% was also significantly reduced in non-survivors (P = 0.013), reinforcing the observation of impaired lymphocyte activation in poor prognosis. These findings suggest that reduced B cell numbers and impaired CD8+ T cell activation especially the CD8+DR+ subset—may be critical indicators of poor immune competence and are closely associated with higher in-hospital mortality in critically ill VOPs.
Immune-Inflammatory Sub-Phenotypes and Their Interaction in VOPs
To further investigate the sub-phenotypes of inflammatory and immune responses in VOPs with critical illness, we performed a two-step analysis. First, we conducted a Spearman correlation analysis to preliminarily assess the relationships among inflammatory biomarkers. As shown in Figure 2 and Table S1, the inflammatory markers demonstrated complex and interconnected correlations. Next, we performed principal component analysis (PCA) to integrate those inflammatory factors that showed significant correlations. This allowed us to categorize inflammatory status into high and low inflammatory states based on the principal components. Since Principal Component 1 (PC1) explained 95.85% of the total variance, it was selected to represent the overall inflammatory state. The equation for PC1 was as follows:
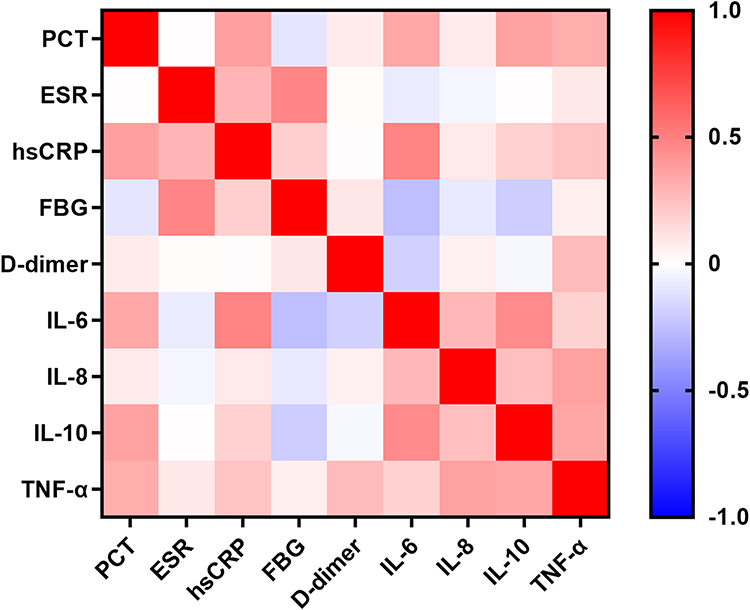 |
Figure 2 Heat map of the Spearman correlation coefficient of inflammatory biomarkers. Abbreviations: PCT, procalcitonin; ESR, erythrocyte sedimentation rate; hsCRP, hypersensitive C-reactive protein; FBG, fibrinogen; IL, interleukin; TNF-α, tumor necrosis factor-α; C, complement; Ig, immunoglobulin. |
PC1 = 0.11 × PCT − 0.005 × ESR − 0.009 × hsCRP + 0.11 × IL-6 + 0.994 × IL-8 + 0.006 × IL-10 + 0.009 × TNF-α.
Using this composite index, we classified patients into high or low inflammatory states and evaluated the impact of immune function particularly lymphocyte subsets on in-hospital mortality within each inflammatory category. Based on earlier findings among VOPs, where B cells and CD8+ T cell activation subsets (CD8+CD28+, CD8+CD38+, CD8+DR+) were significantly associated with prognosis, we specifically analyzed the interactions between these immune parameters and the inflammatory state to better understand their combined effects on patient outcomes.
Interaction Between Inflammatory State and Immune Cell Subsets
Interaction analysis revealed that in-hospital mortality was significantly higher in the low inflammatory state group compared to the high inflammatory state group. Moreover, as the counts of B cells and T cells increased, a significant decrease in mortality was observed across both inflammatory states. Notably, in the low inflammatory state, there was no clear association between activation subsets of CD8+ T cells and mortality. However, in the high inflammatory state, a clear downward trend in mortality was seen with increasing counts of the CD8+DR+ subset (Figure 3A–C), suggesting that CD8+DR+ T cell activity may be particularly protective under conditions of heightened inflammation.
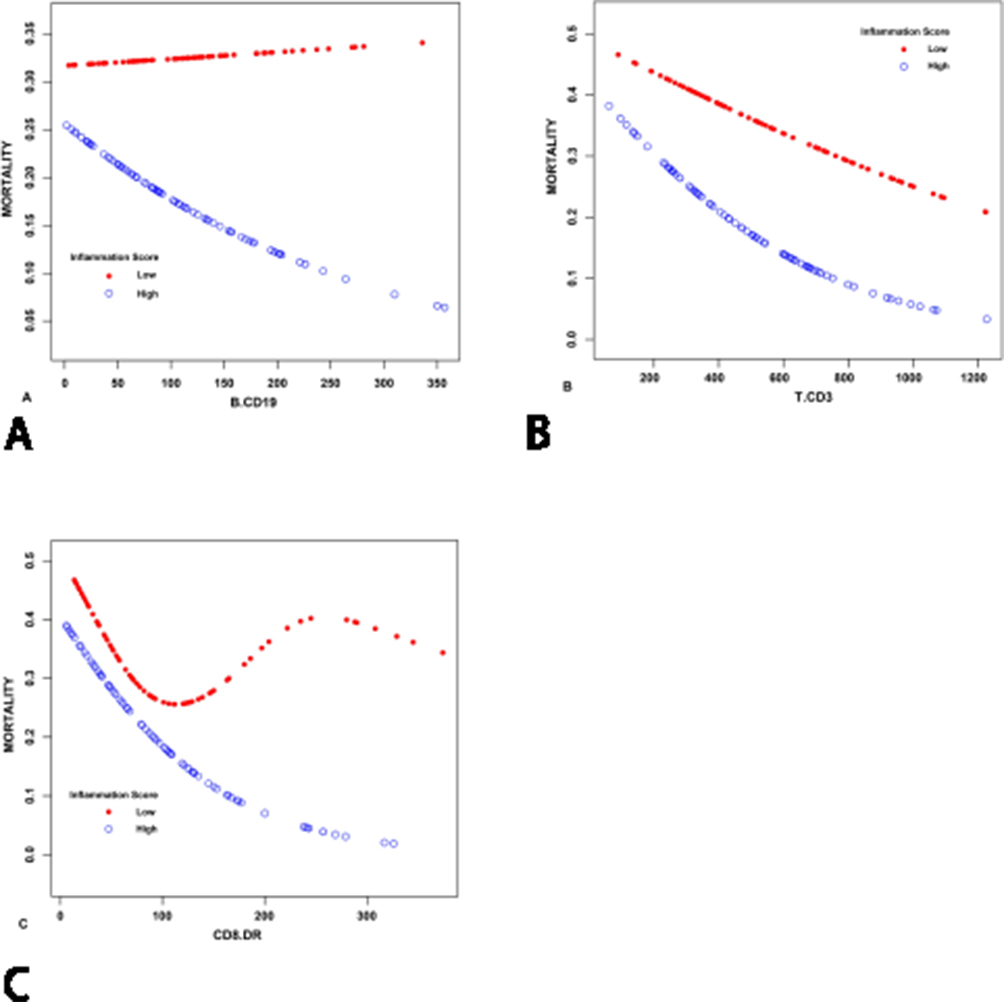 |
Figure 3 Smooth curves illustrating the association between lymphocyte subset counts and in-hospital mortality, stratified by inflammatory state, based on a multivariable-adjusted model. (A) Relationship between (B) cell count and mortality, (B) Relationship between T cell count and mortality and (C) Relationship between CD8+DR+ T cell subset count and mortality. Abbreviation: APACHE II, Acute Physiology and Chronic Health Evaluation II. Note: The model adjusts for age, gender and APACHE II score. |
Additional detailed results are available in Table S2.
Discussion
This prospective study systematically analyzed lymphocyte subsets and inflammatory markers in 537 older patients (OPs) with critical illnesses, revealing, for the first time, distinct immune-inflammatory profiles between young old patients (YOPs) and very old patients (VOPs) in the ICU, and their associations with prognosis. First, a decrease in lymphocyte count was common among OPs, accompanied by elevated proportions of CD8+DR+ and CD8+CD28+ T cells. Compared with YOPs, VOPs showed increased lymphocyte counts, primarily due to higher NK cells, T cells, and CD8+ T cell subsets, while B cells showed a decreasing trend. Second, compared to survivors, non-survivors had higher levels of D-dimer, TNF-α, IgG, and IgM, and lower hsCRP. Immune cell counts including B cells, NK cells, and T cells were reduced in non-survivors, although CD8+DR+ subset counts were elevated. Among VOPs, non-survivors had increased IL-8 and IgG but decreased hsCRP, B cells, T cells, and CD8+DR+ subsets. Lastly, in-hospital mortality was significantly higher in the low inflammation group. In contrast, among those with high inflammatory states, increased counts of B cells, T cells, and CD8+DR+ cells were associated with a significant reduction in mortality, suggesting protective immune roles under inflammatory stress.
This aligns with immunosenescence, the age-related decline in immune system structure and function, which affects immune cell quantity, diversity, and activity.19 Lymphocytes play a central role in immune regulation, and flow cytometry analysis of their subsets is considered the most accurate method to assess immunosenescence.20 Aging causes impaired lymphocyte development and function, resulting in reduced immune cell production.21 In our study, both the counts and proportions of lymphocytes in OPs were significantly below normal, confirming immunosenescence. This finding aligns with previous research showing age-related reductions in lymphocyte subsets.22,23 Critical illnesses further aggravate immune suppression. Conditions such as sepsis can accelerate lymphocyte apoptosis, contributing to lymphopenia.24 Adrie et al25 also found persistent lymphopenia in ICU shock patients, linked to increased infection risk and worse outcomes. Aging-associated thymic degeneration limits T cell generation, making T cells especially CD8+ highly susceptible to decline.26–28 Our results support this, as circulating T cell counts were generally reduced in OPs. Interestingly, we observed elevated proportions of CD4+CD28+ and CD8+DR+ subsets. CD4+CD28+ cells drive antigen-specific responses via inflammatory cytokines,29 while CD8+DR+ cells are activated cytotoxic T cells capable of eliminating infected targets.30 These findings may reflect a compensatory immune response. Still, such increases do not necessarily indicate functional enhancement; instead, they may signal functional exhaustion. Prior studies also show inconsistent trends in subset proportions with aging some report CD4+ declines, others CD8+.31,32 Overall, while immune cell proportions may shift, these changes do not always reflect preserved immune competence, especially in critically ill older adults.
Compared with YOPs, VOPs with critical illnesses display a distinct immune profile, characterized by increased lymphocyte counts, mainly driven by higher numbers of NK cells and CD8+ T cell subsets. While immunosenescence is commonly associated with weakened adaptive immunity and enhanced innate immune activity, this pattern is not absolute. NK cells, key components of innate immunity, have been shown in some studies to increase with age, consistent with our findings.22,31 However, other research, such as that by Valdiglesias et al,32 suggests that NK cell proportions remain stable during aging.
T cells and their subsets are essential to adaptive immunity, and their development is largely dependent on thymic function. Thymic degeneration typically reduces T cell production with age.33 Studies on healthy individuals confirm age-related declines in total T cells and CD4+ and CD8+ subsets.34,35 However, not all findings are consistent. For example, Qin et al22 found that T cells and CD8+ subsets declined significantly with age, while CD4+ subsets remained relatively unchanged. Importantly, the immune profiles of OPs with critical illnesses differ from those of healthy individuals. Xue et al36 reported that in sepsis, OPs had significantly lower levels of T cells, CD4+ subsets, and NK cells compared to younger patients, with a reduced CD4/CD8 ratio also observed.37 These differences may help explain our observation that VOPs with critical illnesses had higher counts of lymphocytes, NK cells, and CD8+ subsets compared to YOPs. This seemingly contradictory result may reflect the combined effects of aging and critical illness on immune activation. For example, VOPs are more prone to latent infections such as cytomegalovirus, and periodic reactivation may trigger expansion of CD8+ subsets, contributing to increased total lymphocyte counts.38
In the non-survivor group, D-dimer and TNF-α levels were elevated, while hsCRP was significantly decreased. This challenges the traditional view that higher hsCRP reflects more severe inflammation and worse prognosis. As an acute-phase protein, hsCRP plays a key role in innate immunity by activating complement and promoting phagocytosis.39 A higher hsCRP level may suggest a relatively intact immune response, while lower levels may reflect immune suppression. This could explain why non-surviving VOPs had lower hsCRP—indicating weaker immunity and lower disease tolerance. Additionally, the impact of treatment cannot be ruled out. Critically ill patients often receive glucocorticoids, which inhibit hsCRP synthesis and may suppress inflammatory markers despite ongoing inflammation.40 Moreover, since hsCRP production depends on IL-6-stimulated hepatocytes, liver dysfunction—a common complication in critically ill VOPs—may further reduce hsCRP levels. Thus, low hsCRP may result from both immune exhaustion and impaired hepatic function.41
This study found that although total lymphocyte counts did not differ significantly between survivors and non-survivors among OPs, the counts of B cells, NK cells, and CD8+DR+ T cell subsets were significantly reduced in non-survivors. This suggests that these specific lymphocyte subsets may play a critical role in prognosis. B cells are central to humoral immunity, producing antibodies that neutralize pathogens.42,43 A reduction in B cells leads to decreased IgG and IgM levels, impairing neutralization and increasing susceptibility to secondary infections. Moreover, B cells present antigens to CD4+ T cells, and their decline may indirectly impair T cell function.44 Prior studies confirm this link: Martin-Sanchez et al45 reported reduced B cells predicted mortality in COVID-19, while Wang et al46 found lower B cell counts in non-survivors of severe pneumonia. NK cells, essential for innate immunity, eliminate infected cells and recruit immune effectors.47 A decrease in NK cells reduces perforin and granzyme secretion, impairing viral and bacterial clearance,48,49 and is consistently linked to poor outcomes.36,50 CD8+DR+ is a key activation subset of CD8+ T cells involved in cytotoxic responses and viral control.51 It is associated with viral replication control in HIV patients,52 and Bobcakova et al53 found elevated CD8+DR+ counts in COVID-19 survivors. Thus, reduced CD8+DR+ counts in non-survivors may indicate immune exhaustion, offering a possible explanation for our findings.
In non-survivor VOPs, hsCRP levels were reduced, while IL-8 levels were significantly elevated. IL-8, secreted by monocytes, endothelial, and epithelial cells, is a strong chemokine that activates immune cells and recruits neutrophils, promoting degranulation and tissue damage.54–56 Elevated IL-8 has been linked to poor outcomes in traumatic brain injury, COVID-19, and sepsis.50,57,58 Additionally, B cells, T cells, and CD8+DR+ subsets were significantly decreased in non-survivors, reinforcing their prognostic value. These distinct immune-inflammatory profiles formed the basis for further exploration of immune-inflammation interactions and their impact on prognosis in critically ill VOPs.
Interaction analysis showed that a low inflammatory state is often associated with worse prognosis, likely due to a weakened immune response. In such states, decreased expression of human leukocyte antigen-DR on monocytes impairs pathogen clearance, allowing infections to spread.59 Reduced B and T cell counts lead to diminished antibody production and cytotoxic activity, increasing the risk of secondary infections, including fungi and drug-resistant bacteria.60 In contrast, higher B and T cell counts are linked to lower in-hospital mortality, especially in high inflammatory states. Notably, CD8+DR+ cells play a prominent protective role under high inflammation, likely due to effective pathogen clearance.61 However, in low inflammation, their prognostic impact weakens, possibly because immune suppression limits their activation. These findings confirm that the prognostic value of lymphocyte subsets varies with inflammatory status, supporting the concept of “precise immune regulation” through tailored immune interventions based on the patient’s inflammatory profile. Collectively, these findings support the concept of precise immune regulation and suggest that clinicians should adapt therapeutic strategies to the patient’s specific immune-inflammatory profile. This approach can facilitate the development of targeted immunomodulation for older ICU populations and guide the direction of future clinical research.
Firstly, this study is a single-center prospective study, which may limit the generalizability of the findings. Future multi-center studies are needed to further clarify the immune and inflammatory profiles of OPs with critical illnesses, particularly VOPs. Secondly, the single-center design results in a relatively limited sample size. It is essential to validate the current findings and conclusions in studies with larger and more diverse populations. Furthermore, this study measured lymphocyte subsets and inflammatory markers at only a single time point. Future research should incorporate dynamic monitoring of immune-inflammatory changes over time, allowing for the inclusion of time as a variable. This approach would provide a more comprehensive understanding of the immune response in critically ill OPs and help refine and strengthen the study’s conclusions.
Conclusion
This study highlights that critically ill older patients (OPs), particularly very old patients (VOPs), exhibit distinct immune-inflammatory profiles that significantly influence clinical outcomes. While aging and critical illness independently contribute to immune dysfunction, their combined effects result in complex immune-inflammatory interactions. Notably, lymphocyte counts and subsets, especially B cells and CD8+DR+ T cells, were found to be closely associated with prognosis. A lower immune response, particularly under conditions of low inflammation, was linked to higher in-hospital mortality. Conversely, increased counts of B cells and CD8+DR+ T cells were associated with improved outcomes, especially in high inflammatory states. These findings suggest that immune suppression and immune exhaustion play critical roles in the poor prognosis of this patient population. Precise immune regulation holds clinical value, suggesting that patients with low inflammatory responses may benefit from immune-enhancing strategies, while those exhibiting hyperinflammatory responses may achieve better outcomes with immunotherapeutic strategies. These findings support the development tailored therapeutic decisions based on individual immune profiles.
Data Sharing Statement
All datasets used and analyzed during the current study are available from the corresponding author (Xiaoting Wang) on reasonable request.
Ethics Approval and Consent to Participate
This study was approved by the ethics committee of Peking Union Medical College Hospital, Beijing, China (Approval No. I-23PJ1278). This study is conducted on a group of critically ill patients, and almost all of them have consciousness problems when they enter the ICU. Written informed consent was obtained from the next of kin of each patient. The study was conducted in accordance with the principles of the Declaration of Helsinki.
Author Contributions
Hui Lian and Hua Zhao conceived and designed the study. Qing Zhang acquired the data. Hui Lian and Guangjian Wang reorganized and independently verified the data; Hui Lian performed the statistical analyses, and Guangjian Wang prepared the figures. Hui Lian and Guangjian Wang co-drafted the manuscript, while Hua Zhao and Xiaoting Wang coordinated the work and critically revised the manuscript. Hui Lian and Guangjian Wang contributed equally to this work. All authors made substantial contributions to the conception, design, execution, data acquisition, analysis, and/or interpretation; participated in drafting, revising, or critically reviewing the article; approved the final version to be published; agreed on the journal of submission; and accepted accountability for all aspects of the work.
Funding
This work was supported by the Peking Union Medical College Hospital Talent Cultivation Program Category D (UHB11894), the Noncommunicable Chronic Diseases-National Science and Technology Major Project (2023ZD0517200), and CAMS Innovation Fund for Medical Sciences (CIFMS), 2024-I2M-CTT-B-003.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Jakovljevic M, Kumagai N, Ogura S. Editorial: global population aging – health care, social and economic consequences, volume II. Editorial. Front Public Health. 2023;11:84950. doi:10.3389/fpubh.2023.1184950
2. Crichton M, Craven D, Mackay H, Marx W, de van der Schueren M, Marshall S. A systematic review, meta-analysis and meta-regression of the prevalence of protein-energy malnutrition: associations with geographical region and sex. Age Ageing. 2019;48(1):38–48. doi:10.1093/ageing/afy144
3. Vallet H, Arai H, Jacobs JM. Bridging the gap: improving the long-term management of very old patients in intensive care. Eur Geriatric Med. 2025. doi:10.1007/s41999-025-01190-3
4. Liu D, Huang S-Y, Sun J-H, et al. Sepsis-induced immunosuppression: mechanisms, diagnosis and current treatment options. Military Med Res. 2022;9(1):56.
5. Jarczak D, Nierhaus A. Cytokine storm—definition, causes, and implications. Int J Mol Sci. 2022;23(19):11740.
6. Goyani P, Christodoulou R, Vassiliou E. Immunosenescence: aging and immune system decline. Vaccines. 2024;12(12):1314.
7. Lioulios G, Fylaktou A, Papagianni A, Stangou M. T cell markers recount the course of immunosenescence in healthy individuals and chronic kidney disease. Clin Immunol. 2021;225:108685. doi:10.1016/j.clim.2021.108685
8. Weinberger B, Lazuardi L, Weiskirchner I, et al. Healthy aging and latent infection with CMV lead to distinct changes in CD8+ and CD4+ T-cell subsets in the elderly. Hum Immunol. 2007;68(2):86–90. doi:10.1016/j.humimm.2006.10.019
9. Han S, Georgiev P, Ringel AE, Sharpe AH, Haigis MC. Age-associated remodeling of T cell immunity and metabolism. Cell Metab. 2023;35(1):36–55.
10. Batatinha HAP, Diniz TA, de Souza Teixeira AA, Krüger K, Rosa-Neto JC. Regulation of autophagy as a therapy for immunosenescence-driven cancer and neurodegenerative diseases: the role of exercise. Journal of Cellular Physiology. 2019;234(9):14883–14895. doi:10.1002/jcp.28318
11. Prcina M, Novak M, Cigankova V, Kontsekova E. Immunosenescence - the role in the immunotherapy of older population. Bratisl Lek Listy. 2018;119(4):217–220. doi:10.4149/bll_2018_040
12. Fulop T, Larbi A, Dupuis G, et al. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: friends or Foes? Review. Front Immunol. 2018;8:1960. doi:10.3389/fimmu.2017.01960
13. Standiford TJ. Anti-inflammatory cytokines and cytokine antagonists. Curr Pharm Des. 2000;6(6):633–649. doi:10.2174/1381612003400533
14. Hou W, Jin YH, Kang HS, Kim BS. Interleukin-6 (IL-6) and IL-17 synergistically promote viral persistence by inhibiting cellular apoptosis and cytotoxic T cell function. J Virol. 2014;88(15):8479–8489. doi:10.1128/jvi.00724-14
15. Dalskov L, Gad HH, Hartmann R. Viral recognition and the antiviral interferon response. EMBO J. 2023;42(14):e112907.
16. Sefik E, Qu R, Junqueira C. Inflammasome activation in infected macrophages drives COVID-19 pathology. Nature. 2022;606(7914):585–593. doi:10.1038/s41586-022-04802-1
17. Shirakabe A, Matsushita M, Shibata Y, et al. Organ dysfunction, injury, and failure in cardiogenic shock. J Intensive Care. 2023;11(1):26. doi:10.1186/s40560-023-00676-1
18. Dziedzic EA, Gąsior JS, Tuzimek A, et al. Investigation of the associations of novel inflammatory biomarkers—systemic inflammatory index (SII) and systemic inflammatory response index (SIRI)—with the severity of coronary artery disease and acute coronary syndrome occurrence. Int J Mol Sci. 2022;23(17):9553.
19. Liu Z, Liang Q, Ren Y, et al. Immunosenescence: molecular mechanisms and diseases. Signal Transduction Target Ther. 2023;8(1):200. DOI:10.1038/s41392-023-01451-2
20. Day J, Limaye V, Proudman S, Hayball JD, Hissaria P. The utility of monitoring peripheral blood lymphocyte subsets by flow cytometric analysis in patients with rheumatological diseases treated with rituximab. Autoimmunity Rev. 2017;16(5):542–547. doi:10.1016/j.autrev.2017.03.003
21. Pang WW, Price EA, Sahoo D, et al. Human bone marrow hematopoietic stem cells are increased in frequency and myeloid-biased with age. Proc Natl Acad Sci USA. 2011;108(50):20012–20017. doi:10.1073/pnas.1116110108
22. Qin L, Jing X, Qiu Z, et al. Aging of immune system: immune signature from peripheral blood lymphocyte subsets in 1068 healthy adults. Aging. 2016;8(5):848–859. doi:10.18632/aging.100894
23. Zhang K, Wang F, Zhang M, et al. Reference ranges of lymphocyte subsets balanced for age and gender from a population of healthy adults in Chongqing District of China. Cytometry Part B. 2016;90(6):538–542. doi:10.1002/cyto.b.21323
24. He Y, Liu Q, Wei L, et al. The value of peripheral blood leukocyte parameters in the early diagnosis and clinical prognosis of sepsis. Int J Analytical Chemi. 2023;2023(1):6052085.
25. Adrie C, Lugosi M, Sonneville R, et al. Persistent lymphopenia is a risk factor for ICU-acquired infections and for death in ICU patients with sustained hypotension at admission. Ann Intens Care. 2017;7(1):30. doi:10.1186/s13613-017-0242-0
26. Wertheimer AM, Bennett MS, Park B, et al. Aging and cytomegalovirus infection differentially and jointly affect distinct circulating T cell subsets in humans. J Immunol. 2014;192(5):2143–2155. doi:10.4049/jimmunol.1301721
27. Sun X, Nguyen T, Achour A, et al. Longitudinal analysis reveals age-related changes in the T cell receptor repertoire of human T cell subsets. J Clin Invest. 2022;132(17):1. doi:10.1172/jci158122
28. Liang Z, Dong X, Zhang Z, Zhang Q, Zhao Y. Age‐related thymic involution: mechanisms and functional impact. Aging Cell. 2022;21(8):e13671.
29. Carr TM, Wheaton JD, Houtz GM, Ciofani M. JunB promotes Th17 cell identity and restrains alternative CD4(+) T-cell programs during inflammation. Nat Commun. 2017;8(1):301. doi:10.1038/s41467-017-00380-3
30. Lodi L, Sarli WM, Ricci S, et al. CD38high/HLA‐DR+ CD8+ T lymphocytes display pathogen‐specific expansion regardless of hemophagocytic lymphohistiocytosis. Eur J Immunol. 2024;54(11):2451140.
31. Choi J, Lee SJ, Lee YA, Maeng HG, Lee JK, Kang YW. Reference values for peripheral blood lymphocyte subsets in a healthy Korean population. Immun net. 2014;14(6):289–295. doi:10.4110/in.2014.14.6.289
32. Valdiglesias V, Sánchez-Flores M, Maseda A, et al. Lymphocyte Subsets in a Population of Nonfrail Elderly Individuals. J Toxicol Environ Health Part A. 2015;78(13–14):790–804. doi:10.1080/15287394.2015.1051170
33. Sun H, Kang X, Chen X, et al. Immunosenescence evaluation of peripheral blood lymphocyte subsets in 957 healthy adults from 20 to 95 years old. Exp Gerontology. 2022;157:111615. doi:10.1016/j.exger.2021.111615
34. Hirokawa K, Utsuyama M, Hayashi Y, Kitagawa M, Makinodan T, Fulop T. Slower immune system aging in women versus men in the Japanese population. Immunity Ageing: I & A. 2013;10(1):19. doi:10.1186/1742-4933-10-19
35. Valdiglesias V, Sánchez-Flores M. Immune biomarkers in older adults: role of physical activity. J Toxicol Environ Health. 2017;80(13–15):605–620. doi:10.1080/15287394.2017.1286898
36. Xue L, Sun QM, Yu H, Liang YH, Li CS. Relationship between Cellular Immunity Changes and Prognosis in Elderly Patients with Sepsis. JCPSP. 2019;29(12):1144–1148. doi:10.29271/jcpsp.2019.12.1144
37. Rimmelé T, Payen D, Cantaluppi V, et al. IMMUNE CELL PHENOTYPE AND FUNCTION IN SEPSIS. Shock Augusta Ga. 2016;45(3):282–291. doi:10.1097/shk.0000000000000495
38. Reus B, Caserta S, Larsen M, et al. In-Depth Profiling of T-Cell Responsiveness to Commonly Recognized CMV Antigens in Older People Reveals Important Sex Differences. Front Immunol. 2021;12:707830. doi:10.3389/fimmu.2021.707830
39. Zhou HH, Tang YL, Xu TH, Cheng B. C-reactive protein: structure, function, regulation, and role in clinical diseases. Front Immunol. 2024;15:1425168. doi:10.3389/fimmu.2024.1425168
40. Denegri A, Boriani G. High sensitivity C-reactive protein (hsCRP) and its implications in cardiovascular outcomes. Curr Pharm Des. 2021;27(2):263–275.
41. Tang ZM, Yuan P, Gao N, et al. C-reactive protein attenuates CCl(4)-induced acute liver injury by regulating complement system activation. Mol Immunol. 2025;180:44–54. doi:10.1016/j.molimm.2025.02.008
42. Urbanczyk S, Baris OR, Hofmann J, et al. Mitochondrial respiration in B lymphocytes is essential for humoral immunity by controlling the flux of the TCA cycle. Cell Rep. 2022;39(10):1.
43. Wang T, Chen R, Ouyang R, et al. Peripheral lymphocyte phenotypic characteristics in healthy populations across the lifespan, from infancy to older adults. Lab Med. 2025;2025:117. doi:10.1093/labmed/lmae117
44. Akkaya M, Kwak K, Pierce SK. B cell memory: building two walls of protection against pathogens. Nat Rev Immunol. 2020;20(4):229–238. doi:10.1038/s41577-019-0244-2
45. Martín-Sánchez E, Garcés JJ, Maia C, et al. Immunological Biomarkers of Fatal COVID-19: a Study of 868 Patients. Front Immunol. 2021;12:659018. doi:10.3389/fimmu.2021.659018
46. Wang CM, Zhang Y, Xu HH, et al. B cell subsets were associated with prognosis in elderly patients with community acquired pneumonia. BMC Pulm Med. 2022;22(1):206. doi:10.1186/s12890-022-01985-1
47. Wang F, Cui Y, He D, Gong L, Liang H. Natural killer cells in sepsis: friends or foes? Front Immunol. 2023;14:1101918. doi:10.3389/fimmu.2023.1101918
48. Quatrini L, Della Chiesa M, Sivori S, Mingari MC, Pende D, Moretta L. Human NK cells, their receptors and function. Eur J Immunol. 2021;51(7):1566–1579.
49. Ramírez-Labrada A, Pesini C, Santiago L, et al. All About (NK Cell-Mediated) Death in Two Acts and an Unexpected Encore: initiation, Execution and Activation of Adaptive Immunity. Front Immunol. 2022;13:896228. doi:10.3389/fimmu.2022.896228
50. Kang Y, Lu S, Zhong R, et al. The immune inflammation factors associated with disease severity and poor prognosis in patients with COVID-19: a retrospective cohort study. Heliyon. 2024;10(1):e23583. doi:10.1016/j.heliyon.2023.e23583
51. Oh DY, Fong L. Cytotoxic CD4+ T cells in cancer: expanding the immune effector toolbox. Immunity. 2021;54(12):2701–2711.
52. Gonzalez SM, Taborda NA, Rugeles MT. Role of Different Subpopulations of CD8(+) T Cells during HIV Exposure and Infection. Front Immunol. 2017;8:936. doi:10.3389/fimmu.2017.00936
53. Bobcakova A, Barnova M, Vysehradsky R, et al. Activated CD8(+)CD38(+) Cells Are Associated With Worse Clinical Outcome in Hospitalized COVID-19 Patients. Front Immunol. 2022;13:861666. doi:10.3389/fimmu.2022.861666
54. Darif D, Hammi I, Kihel A, El Idrissi Saik I, Guessous F, Akarid K. The pro-inflammatory cytokines in COVID-19 pathogenesis: what goes wrong? Microb Pathogenesis. 2021;153:104799. doi:10.1016/j.micpath.2021.104799
55. Asiri A, Hazeldine J, Moiemen N. IL-8 Induces Neutrophil Extracellular Trap Formation in Severe Thermal Injury. Int J Mol Sci. 2024;25(13):7216. doi:10.3390/ijms25137216
56. Bernhard S, Hug S, Stratmann AEP, et al. Interleukin 8 elicits rapid physiological changes in neutrophils that are altered by inflammatory conditions. J Innate Immunity. 2021;13(4):225–241.
57. Gopcevic A, Mazul-Sunko B, Marout J, et al. Plasma interleukin-8 as a potential predictor of mortality in adult patients with severe traumatic brain injury. Tohoku J Exp Med. 2007;211(4):387–393. doi:10.1620/tjem.211.387
58. Zhang X, Wang J, Guo S. Predictive value of IL-8 for mortality risk in elderly sepsis patients of emergency department. Cytokine. 2024;184:156774. doi:10.1016/j.cyto.2024.156774
59. Chu C-M, Chung C-J, Huang C-Y, et al. Serial Increases in Human Leukocyte Antigen-DR Expression and Decreases in Interleukin-10 Expression in Alveolar Monocytes of Survivors of Pneumonia-Related Acute Respiratory Distress Syndrome. Biology. 2022;11(12):1793.
60. Tangye SG, Brink R, Goodnow CC, Phan TG. SnapShot: interactions between B Cells and T Cells. Cell. 2015;162(4):926–6.e1. doi:10.1016/j.cell.2015.07.055
61. Kiran S, Kumar V, Murphy EA, Enos RT, Singh UP. High fat diet-induced CD8+ T cells in adipose tissue mediate macrophages to sustain low-grade chronic inflammation. Front Immunol. 2021;12:680944.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
A Novel lncRNA Panel for Risk Stratification and Immune Landscape in Breast Cancer Patients

Li C, Wang X, Chen T, Li W, Yang Q
International Journal of General Medicine 2022, 15:5253-5272
Published Date: 27 May 2022
Construction and Identification of an NLR-Associated Prognostic Signature Revealing the Heterogeneous Immune Response in Skin Cutaneous Melanoma
Geng Y, Sun YJ, Song H, Miao QJ, Wang YF, Qi JL, Xu XL, Sun JF
Clinical, Cosmetic and Investigational Dermatology 2023, 16:1623-1639
Published Date: 26 June 2023
Significance of Pyroptosis in Immunoregulation and Prognosis of Patients with Acute Respiratory Distress Syndrome: Evidence from RNA-Seq of Alveolar Macrophages
Liu B, Li Y, Xiang J, Li Y, Zhou M, Ren Y, Fu Z, Ding F
Journal of Inflammation Research 2023, 16:3547-3562
Published Date: 21 August 2023
Research on the Association Between Periodontitis and COPD
Xiong K, Yang P, Cui Y, Li J, Li Y, Tang B
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:1937-1948
Published Date: 1 September 2023
Assessment of Immune Status in Patients with Mismatch Repair Deficiency Endometrial Cancer
Ma J, Lin J, Lin X, Ren Y, Liu D, Tang S, Huang L, Xu S, Mao X, Sun P
Journal of Inflammation Research 2024, 17:2039-2050
Published Date: 2 April 2024