Back to Journals » Journal of Inflammation Research » Volume 19
Single-Cell and Spatial Transcriptomics Identify the Mono-S100A4 Subset Associated with Sepsis-Induced Lung Injury via the MIF-(CD74+CD44) Axis
Authors Wang A ![]() , Yin C, Jin Y, Yang L, Li S, Li J, Xu L, Shi R, Weng M, Liu W
, Yin C, Jin Y, Yang L, Li S, Li J, Xu L, Shi R, Weng M, Liu W ![]() , Wang X
, Wang X
Received 24 October 2025
Accepted for publication 27 May 2026
Published 10 July 2026 Volume 2026:19 575566
DOI https://doi.org/10.2147/JIR.S575566
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anh Ngo
Aihua Wang,1– 3,* Chengfen Yin,1– 3,* Yue Jin,4,5,* Lei Yang,6 Suiran Li,4,5 Jiaxin Li,1– 3 Lei Xu,1– 3 Ruoyu Shi,4,5 Maotong Weng,7 Wei Liu,4,5 Ximo Wang1– 3
1Central Hospital, Tianjin University, Tianjin, People’s Republic of China; 2Tianjin Key Laboratory of Extracorporeal Life Support for Critical Diseases, Tianjin, People’s Republic of China; 3Tianjin Artificial Cell Engineering Technology Research Center, Tianjin, People’s Republic of China; 4First Teaching Hospital of Tianjin University of Traditional Chinese Medicine, Tianjin, People’s Republic of China; 5National Clinical Research Center for Chinese Medicine, Tianjin, People’s Republic of China; 6Tianjin Nankai Hospital, Tianjin, People’s Republic of China; 7Santa Clara University, Santa Clara, California, USA
*These authors contributed equally to this work
Correspondence: Wei Liu, Email [email protected] Ximo Wang, Email [email protected]
Background: In sepsis-induced immune dysregulation, monocytes contribute critically through their dual pro-inflammatory and immunosuppressive functions. Leading to multi-organ injury, the lungs are the most vulnerable initial target, dictating clinical outcomes. Thus, studying monocyte dynamics is essential to understand sepsis-induced acute lung injury (ALI) pathophysiology.
Methods: We analyzed publicly available mouse-derived single-cell RNA sequencing (scRNA-seq) and spatial transcriptomic (ST) datasets for sepsis-induced ALI from the GEO database. Computational characterization of monocyte heterogeneity included cell-cell communication, single-cell trajectory analysis, and functional enrichment analyses to identify subtype-specific signaling and injury-related signatures. Spatial mapping further confirmed monocyte subtype localization. Experimental validation was performed in a mouse model of sepsis-induced ALI established by intraperitoneal LPS injection. Histological evaluation, immunohistochemistry, immunofluorescence, Quantitative real-time PCR, and Western blot were integrated to verify the spatial distribution of the Mono-S100A4 subset and its associated pathway molecules.
Results: Clustering analysis revealed a restructured immune landscape in septic ALI, characterized by both cellular heterogeneity and prominent monocyte accumulation. Spatial transcriptomics revealed that inflammatory infiltration mediated by monocytes and other immune cells predominantly localized to bronchial and bronchiole regions. Monocyte heterogeneity analysis identified a distinct Mono-S100A4 subset, which exhibited enhanced intercellular communication capacity, influencing other subclusters through the (MIF)-(CD74+CD44) axis. KEGG and GSEA analyses of Mono-S100A4 revealed significant enrichment in the IL-17 signaling pathway and Th17 cell differentiation, suggesting a potential role in exacerbating inflammatory responses. In vivo experiments revealed a significant upregulation in both the mRNA and protein expression levels of S100A4, IL-17A, MIF, and CD74 in the lung tissue of ALI mice.
Conclusion: The Mono-S100A4 subset is strongly associated with the activation of the Th17 signaling pathway via the MIF-(CD74+CD44) axis, and this correlative relationship supports a potential pathogenic role of Mono-S100A4 in sepsis-induced ALI, which forms a key hypothesis to be validated by subsequent functional experiments.
Keywords: sepsis, acute lung injury, monocyte, Mono-S100A4, scRNA-seq, spatial transcriptomics
Introduction
Sepsis is a life-threatening clinical syndrome characterized by organ dysfunction resulting from a dysregulated host response to infection.1 As a major global health challenge, it affects millions of patients worldwide annually.2 Despite advances in medical care and infection prevention efforts, the global age-standardized incidence rate remains high at 677.5 per 100000 population, with a mortality rate of 148.1 per 100000 population.3 These figures likely underestimate the true burden, as incomplete surveillance systems in low-income countries hinder accurate data collection. Recognized as one of medicine’s oldest yet most elusive syndromes,4 mitigating its global health burden and economic impact remains a critical medical challenge.
Sepsis represents the predominant infectious etiology of acute lung injury (ALI) and its more severe form, acute respiratory distress syndrome (ARDS), which represents the most frequent initial organ dysfunction in sepsis progression.5 Epidemiological studies indicate that approximately 50% of severe sepsis patients progress to develop ALI/ARDS.6 Sepsis-induced ALI exhibits greater overall severity, worse prognosis, and higher mortality rates compared to other etiologies.7 The core pathophysiology of ALI involves a dysregulated hyperinflammatory response, marked by extensive inflammatory cell infiltration, cytokine storm generation, pulmonary capillary leakage, and resultant alveolar edema.8 Crucially, the chemotaxis and maturation of monocytes are primarily driven by alveolar cell-derived chemokines, which are triggered by pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs).9 While their functional and phenotypic changes may predict post-traumatic complications or poor prognosis in sepsis, the precise underlying mechanisms remain unclear.10,11
Fuchou Tang et al12 elucidated the complexity of single-cell transcriptomes and has developed high-sensitivity single-cell RNA sequencing (scRNA-seq) technology capable of deconvoluting cellular heterogeneity, identifying rare cell populations, and delineating cell-cell interaction networks.13 Nevertheless, scRNA-seq faces a fundamental limitation: the requirement for tissue dissociation into single cells or nuclei inevitably sacrifices histological context, thereby precluding spatial interaction analysis.14 In contrast, spatial transcriptomics (ST) overcomes the loss of spatial information inherent in scRNA-seq15 by integrating DNA sequencing, fluorescence microscopy, and molecular barcoding to enable in situ capture and sequencing of genome-wide transcripts directly within the tissue context.16
While the molecular mechanisms underlying sepsis-associated ALI, especially in monocyte functional dynamics and their regulatory roles, remain not fully understood, our study employs integrated scRNA-seq and ST to comprehensively investigate monocyte-centered pathogenic mechanisms in this condition.
Materials and Methods
Data Collection
Mouse-derived scRNA-seq data (GSE264032, retaining only the control and extrapulmonary ALI groups) and spatial transcriptomic data (GSE224146, limited to lung tissue samples) were obtained from the Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/).
Single-Cell Transcriptome Analysis
All analyses were conducted using R software (v4.3.2), with scRNA-seq data processed through the Seurat package (v4.2).17 Cell quality control was performed by calculating mitochondrial gene content using PercentageFeatureSet, retaining only cells with <15% mitochondrial reads. For dimensionality reduction, the top 2000 highly variable genes were selected via FindVariableFeatures function in Seurat with the variance-stabilizing transformation method, followed by principal component analysis (PCA). Next, the RunHarmony function in Harmony package was used for performing sample batch correction. Cell clusters were identified through FindNeighbors and FindClusters, where the top 20 principal components (PCs) were used to construct a shared nearest-neighbor (SNN) graph, followed by modularity optimization-based clustering. Finally, UMAP (based on the top 30 PCs) was generated via RunUMAP for 2D visualization.
Spatial Transcriptome Analysis
For ST data analysis, we implemented the Seurat package using an analytical pipeline analogous to the scRNA-seq workflow. Data normalization was performed using SCTransform in Seurat, while spatial feature visualization was achieved through a customized analytical pipeline.18
Single-Cell Reference-Based Spatial Deconvolution
Raw spatial transcriptomics data were normalized and variance-stabilized with the SCTransform function, followed by principal component analysis (PCA) with the top 30 principal components selected for downstream analysis; the data were then clustered into 7 distinct cell clusters with a resolution parameter of 0.3. Our previously processed and annotated scRNA-seq data were used as the reference dataset, and the FindTransferAnchors function was applied to identify shared transcriptional features between the two datasets for the establishment of reliable cross-dataset anchors, thus achieving accurate transcriptomic alignment of scRNA-seq and spatial transcriptomics data. Cell type labels from the scRNA-seq reference were predicted and transferred to the spatial transcriptomics spots via the TransferData function, and the generated cell type prediction score matrix was extracted, transposed and integrated into the metadata of the spatial transcriptomics dataset for subsequent cell type deconvolution analysis.18
Cluster Marker Identification and Cell Type Annotation
Cell marker genes utilized for cell type annotation were primarily referenced from CellMarker 2.0,19 with the following markers used for each population: Mki67 for Alveolartype I epithelial cell (AT I); Ager, Sftpc for Alveolar type II epithelial cell (AT II); Cd79a, Pax5, Cd19 for B cell; Cd200r3, Fcer1a for Basophil; Cd209a for Dendritic cell (DC); Cd34, Cdh5, Cldn5 for Endothelial cell; Col1a1, Tcf21, Timp1 for Fibroblast; Ccl7, Mgl2 for M1 macrophage; Cd68, Chil3, Marco for M2 macrophage; F13a1, Treml4 for Monocyte; Aspn, Myh11, Tagln for Myofibroblast; Klrb1c, Ncr1, Nkg7 for Natural killer T cell (NKT); Arg2, Lcn2, Retnlg for Neutrophil; Higd1b, Vtn, Gucy1a1 for Pericyte; Il2ra for Regulatory T cell (Treg); Cd28, Cd4 for T helper cell (Th) and Slc4a1 for Erythrocytes.
Cell-Cell Communication Analysis
Cell-cell communication analysis was performed using CellChat (v1.1.3), a computational tool leveraging a curated ligand-receptor interaction database.20 Initial screening was conducted using the CellChatDB.human (Secreted Signaling) database to identify major signaling inputs and outputs across all cell clusters. For subsequent analysis, only ligand-receptor pairs expressed in ≥ 10 cells per cluster were considered. This threshold was set to filter out low-abundance, potentially spurious ligand-receptor expression events that may not reflect genuine intercellular communication.20
Single-Cell Trajectory Analysis
Cell state transitions were investigated using trajectory analysis with Monocle.21 Data from the Seurat object were extracted and converted into a CellDataSet object. Significant differentially expressed genes (DEGs) were selected as ordering genes. Following dimensionality reduction using the DDRTree method, cell ordering and trajectory construction were performed with default parameters.
GO and KEGG Analysis
DEGs were analyzed through GO and KEGG pathway enrichment analyses using the DAVID database.22 Genes with log2FC > 1 and log2FC < −1 were classified as up- and down-regulated, respectively. The top 30 most significantly enriched pathways (P < 0.05) showing the highest fold enrichment were selected for visualization.
Gene Set Enrichment Analysis (GSEA) Analysis
DEGs were identified using the “FindMarkers” function in Seurat. Pathway analysis enrichment analysis was performed with the “c2.cp.kegg.v7.5.1. symbols.gmt” gene set collection. GSEA was conducted using clusterProfiler package (v4.2.2), with visualization performed through the enrichplot package’s (v1.14.2) gseaplot2 function. Significant pathways were determined based on normalized enrichment scores (NES) and false discovery rate-adjusted p-values.23
Animals
12 male C57BL/6 mice aged 6–8 weeks were obtained from Beijing Huafukang Biotechnology Co., Ltd. (Beijing, China). The mice were acclimatized and maintained under specific pathogen-free conditions at the Animal Research Institute of Tianjin Nankai Hospital. Environmental parameters included a 12-hour light/dark cycle, controlled relative humidity, and a constant temperature of 23°C. The mice were provided with ad libitum access to food and water. The experimental protocol was reviewed and approved by the Medical Ethics Committee of Tianjin Nankai Hospital (NKYY-DWLL-2023-039), and all procedures were conducted in strict compliance with the institutional guidelines for animal welfare and ethical use. Random allocation of mice to the control and ALI groups (n=6 per group) was performed using a random number table, followed by the modeling procedure. Sepsis-induced ALI was induced via intraperitoneal injection of LPS (25 mg/kg).24 24 hours after modeling, mice were euthanized by isoflurane exposure followed by cervical dislocation. Lung tissue samples were collected and analyzed for subsequent analysis.
Histological Evaluation (HE)
The lung tissue samples were collected and rinsed with PBS, followed by fixation in 4% paraformaldehyde. After dehydration through an ethanol series, the tissues were embedded in paraffin, sectioned, and subjected to HE staining. The inflammatory severity in mouse lung tissues was assessed using the Szapiel scoring method.25
Immunohistochemistry (IHC)
Mouse lung tissues were sliced into 4 μm thick sections. Following deparaffinization, rehydration, antigen retrieval, and endogenous peroxidase blocking, the sections were incubated overnight at 4 °C with anti-CD74 (1:4000, 85,802-2-RR, Proteintech), anti-IL-17A (1:1000, ER1902-37, HUABIO) and anti-MIF (1:200, BA2058, BOSTER) antibodies. The anti-rabbit IgG detection kit (PV-9001, Zhongshan Jinqiao Biotechnology, Beijing, China) was then applied and incubated at 37°C for 30 minutes. Following DAB development and hematoxylin counterstaining, pathological changes in the lung tissues were examined under a light microscope. Images were captured at 400× magnification using optical microscopy.
Immunofluorescence (IF)
Sections following antigen retrieval were blocked with 1% BSA at room temperature for 1 hour and incubated with anti-S100A4 (1:200, ET1612-13, HUABIO) antibody at 4 °C overnight. After repeated washing, the sections were incubated in the dark with fluorescent secondary antibodies (1:200, A0562, Beyotime Biotechnology) for 1 hour at room temperature. Following DAPI (C0065, Solarbio, Beijing, China) counterstaining for 8 minutes, the samples were visualized under a fluorescence microscope.
Quantitative Real-Time PCR (RT-qPCR)
Total RNA was extracted from mouse lung tissue with an RNA extraction reagent (Q30704, TransGen Biotech). According to the manufacturer’s protocol, cDNA was synthesized from the extracted RNA using a reverse transcription kit (BSB40M1, BIOER). RT-PCR SYBR Green Mix (BSB25L1C, BIOER) was used to quantify gene transcripts and normalized to the HRPT expression. Primer sequences are provided in Table 1.
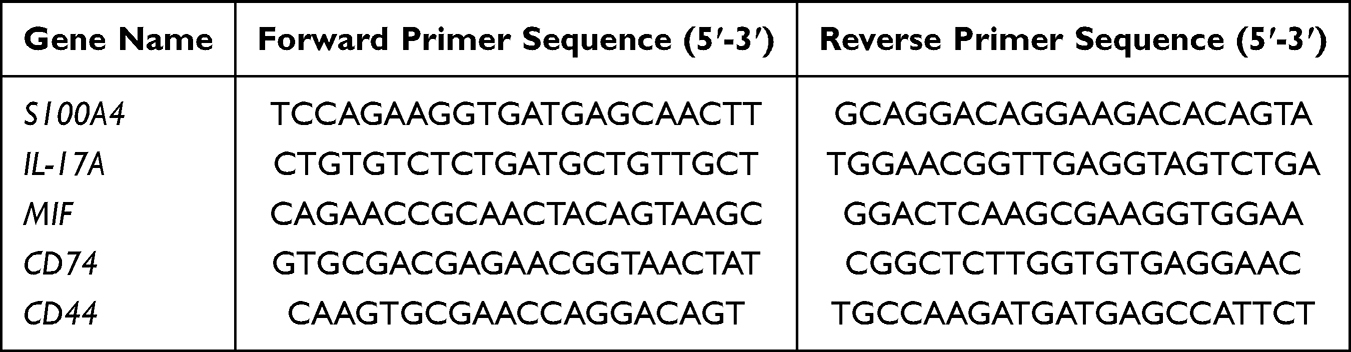 |
Table 1 Primer Sequence |
Western Blotting (WB)
Total protein of lung tissue was extracted using RIPA buffer supplemented with PMSF. After protein concentration determination, the protein was prepared with loading buffer and denatured at 100°C for 10 min, 50μg protein was electrophoretically transferred ontopolyvinylidene difluoride membranes from 12% SDS-PAGE gels. The membranes underwent blocking with 5% skim milk powder at ambient temperature for 1 h, and incubated overnight at 4°C with primary antibodies against S100A4 (1:1000, ET1612-13, HUABIO), IL-17A (1:500, ER1902-37, HUABIO), MIF (1:1000, BA2058, BOSTER) antibodies and GAPDH (1:200000, Proteintech, 60004-1-Ig) After being incubated with secondary antibodies for 1.5 h at room temperature, the membranes were visualized with the multi-function imager (Microline, ML8051), and the images obtained were then subjected to quantification through ImageJ software.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 9.0 software (San Diego, CA, USA). Continuous data are presented as mean ± standard deviation. The normality of all continuous datasets was assessed using the Shapiro–Wilk test. For comparisons between two groups, normally distributed continuous data were analyzed using an unpaired Student’s t-test, while non-normally distributed continuous data and ordinal data were analyzed using the non-parametric Mann–Whitney U-test. Statistical significance was defined as a two-tailed P value < 0.05.
Results
scRNA-Seq Reveals Specific Change in Pulmonary Cell Type During Sepsis
To establish a comprehensive immune cell transcriptional atlas of lung tissue for monitoring sepsis-induced alterations in pulmonary cellular composition, we reanalyzed publicly available scRNA-seq profiles from GSE264032, originally generated using the 10X Genomics platform (Figure 1A). Following scRNA-seq data preprocessing to remove doublets and non-viable cells, a total of 22308 cells met our quality criteria and were analyzed, consisting of 11452 cells from the control group and 10856 cells from pulmonary ALI.26 Using classical marker gene expression patterns, we classified and annotated the cells into 17 principal cell types, including AT I, AT II, B cell, Basophil, DC, Endothelial cell, Fibroblast, M1 macrophage, M2 macrophage, Monocyte, Myofibroblast, NKT, Neutrophil, Pericyte, Treg, Th and Erythrocytes. The classified data were visualized using UMAP dimensionality reduction (Figure 1B). Marker genes annotation confirmed the accuracy of our classification approach (Figure 1C), demonstrating that classification based on a curated set of marker genes achieved high concordance with both unsupervised clustering results and established cell-type annotations, thereby validating the robustness of our methodology. Cellular composition analysis revealed that Monocyte, Neutrophil, and B cell constituted the predominant populations in sepsis-induced ALI (Figure 1D). Comparative analysis revealed significant sepsis-induced alterations in pulmonary cellular composition, characterized by increased proportions of monocytes, neutrophils, and AT II cells, concomitant with decreased populations of B cells, Tregs, and Th cells, demonstrating a sepsis-induced immune response reprogramming in the lung tissue. Cell-cell communication analysis showed monocyte and B cell have notably enhanced communication potential (Figure 1E and F). Single-cell trajectory analysis revealed a dynamic pathological progression: at Marker 2, endothelial cell initiated differentiation, marking the onset of pulmonary injury; at Marker 3, a continuous fibroblast-to-myofibroblast transition emerged, indicating active fibrotic progression; Marker 4 showed late-pseudotime accumulation of monocyte, neutrophil and M1 macrophage, demonstrating persistent inflammatory responses; while at Marker 5, AT II exhibited terminal differentiation, likely representing irreversible damage induced by sustained inflammation (Figure 1G and H).
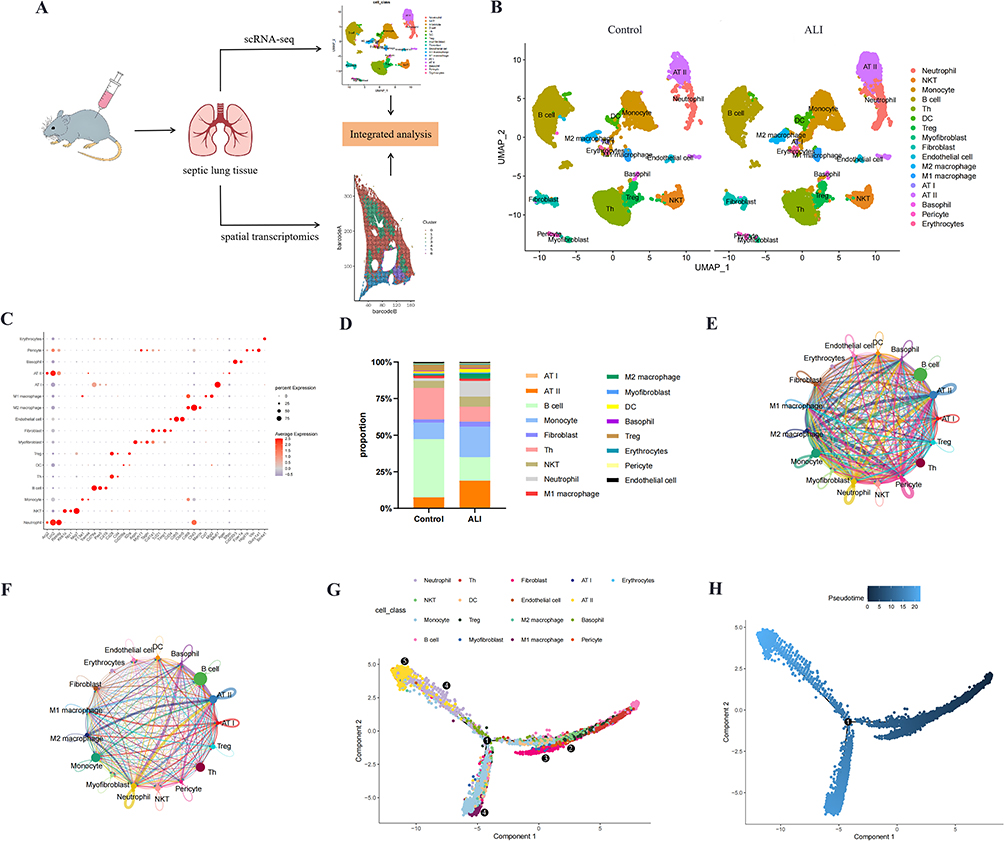 |
Figure 1 Identifying the cellular composition and biological characteristics in septic lung tissue. (A) Schematic overview of the study design, illustrating the integrated analysis of scRNA-seq and spatial transcriptomics data from septic lung tissue. (B) UMAP dimensionality reduction plot showing 17 distinct cell types were identified. (C) The bubble plot shows the expression of marker genes across different cell populations, with bubble size reflecting the number of expressing cells and color intensity indicating average expression. (D) Proportional graph showing the proportions of different cell types in control and ALI groups. (E and F) Network diagram of the number/intensity of cellular communication in the ALI group, with bubble size corresponding to the number of cells, line color denotes signal-sending cells, line thickness reflects the magnitude of the number/intensity, and arrow direction points to signal-receiving cells. (G and H) The developmental trajectory of ALI was predicted by mapping subcluster and pseudo-time information onto the visualization, where each point represents a single cell. |
ST Reveals the Spatial Arrangement of Various Cell Types in Sepsis-Induced ALI
While scRNA-seq reveals cellular diversity but lacks spatial resolution, ST overcomes this limitation by simultaneously capturing transcriptomic activity and precise cellular localization. ST analysis of 23375 tissue spots enabled examination of the spatial organization of cell types within lung tissue, which was anatomically categorized into alveolar, bronchial, bronchiolar, and other regions (Figure 2A). Unsupervised clustering of ST data identified seven distinct spatial domains (C0-C6) (Figure 2B and C). Using cell type-specific gene signatures derived from scRNA-seq analysis, we computed signature scores for each spatial spot (Figure 2D). We found that in addition to the Endothelial cell, Fibroblast and AT II having relatively high scores on most ST_Clusters, there were also high scores for monocyte in ST_clusters 3 and 5. These results demonstrated that monocyte-mediated inflammatory infiltration preferentially localizes to bronchial and bronchiolar regions.
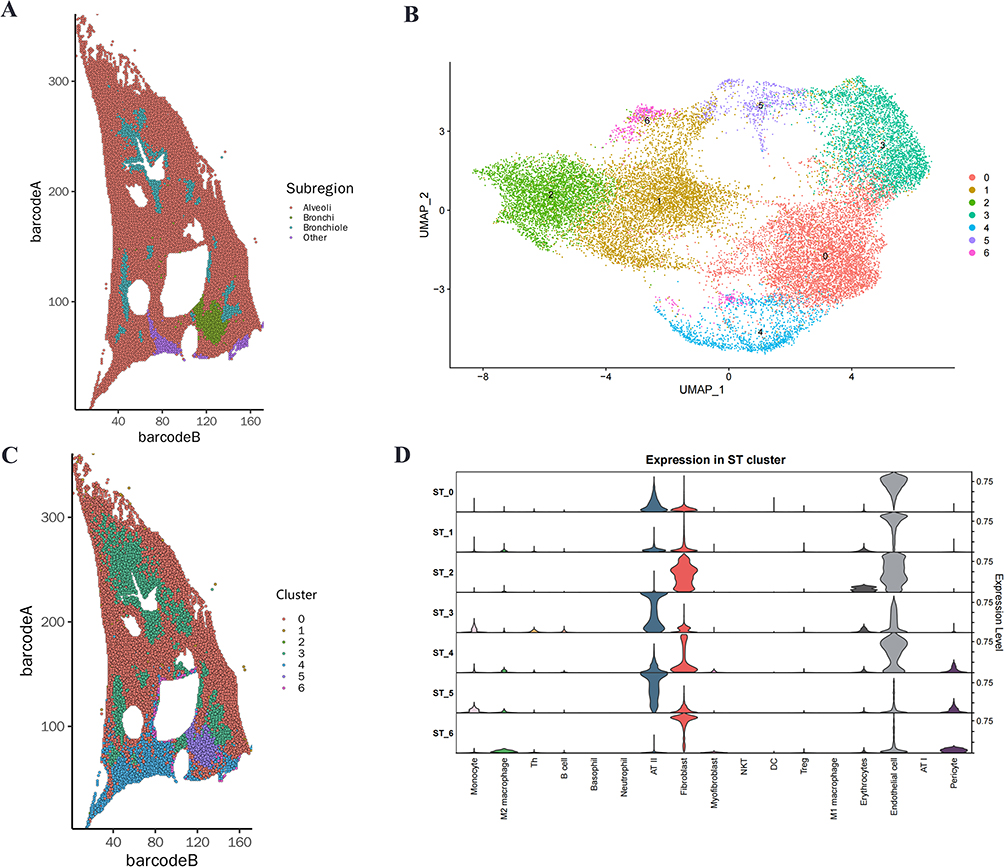 |
Figure 2 Spatial distribution patterns of heterogeneous cell populations in septic lung tissues. (A) ST plot displays the spatial distribution of annotated region. (B) UMAP plot of 23375 ST spots from septic ALI ST data, colored by clusters. (C) Spatial distribution of all spots within this sample color-coded by ST_Clusters. (D) Violin plots depicting heterogeneous cell population scores across ST_Clusters. |
Analysis of the Biological Functions of Monocytes in Sepsis-Induced ALI
To investigate the biological mechanisms of monocyte involvement in sepsis-induced ALI, we performed comprehensive bioinformatics analyses including GO, KEGG pathway and GSEA. GO enrichment analysis identified significantly enriched terms related to immune responses, including response to virus, defense response to virus, response to molecule of bacterial origin, response to lipopolysaccharide and regulation of innate immune response (Figure 3A). KEGG pathway analysis revealed marked enrichment in five key inflammatory pathways: Cytosolic DNA−Sensing Pathway, NOD-like receptor signaling pathway, TNF signaling pathway, Toll-like receptor signaling pathway, and IL-17 signaling pathway (Figure 3B). Subsequent GSEA validation confirmed significant activation of all these pathways (all NES > 1.5) (Figure 3C).
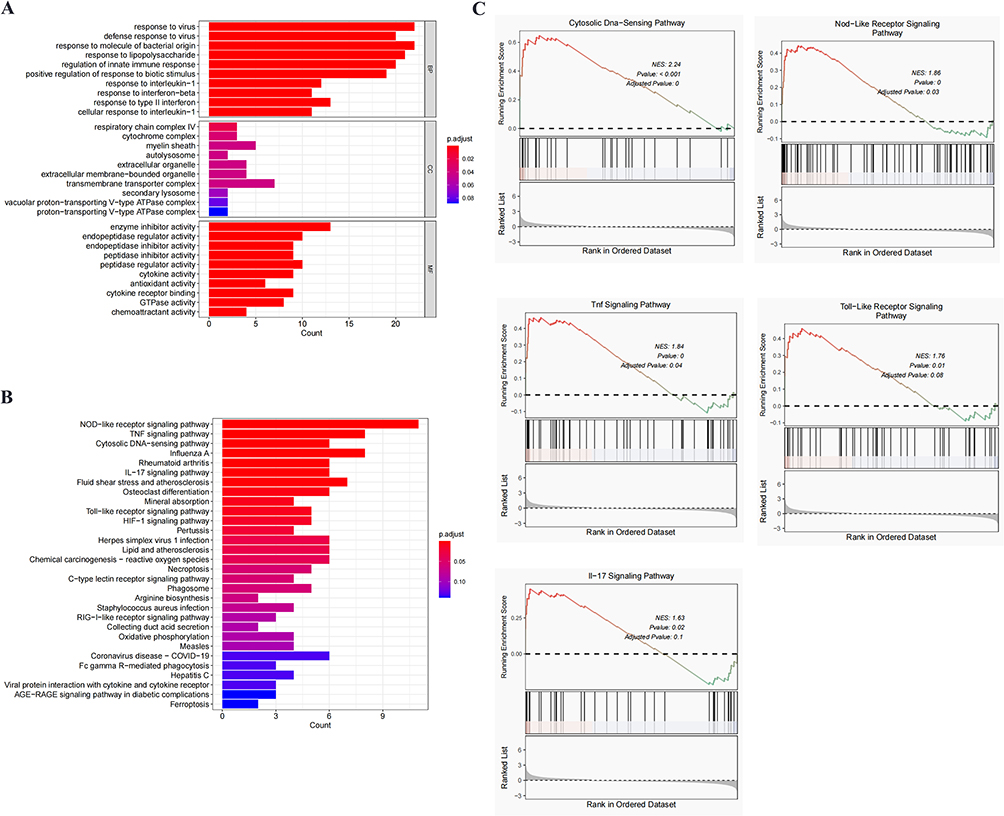 |
Figure 3 Functional characterization of monocytes in sepsis-induced ALI. (A) GO enrichment analysis of upregulated genes in monocytes from ALI versus control groups. (B) Top 30 significantly enriched KEGG pathways in monocytes from ALI versus control groups. (C) GSEA analysis of selected KEGG signaling pathways. |
scRNA-Seq Reveals Sepsis-Induced Remodeling of Pulmonary Monocyte Subpopulations and Their Interactive Signaling Networks
As primary frontline defenders against infection, monocytes play a pivotal role in bridging innate and adaptive immunity through antigen presentation, cytokine secretion, and costimulatory signal expression to activate adaptive immune responses.27 In sepsis, pathogen-induced monocyte activation leads to distinct functional and phenotypic changes reflecting their inherent heterogeneity. Through marker gene profiling, we identified five monocyte subpopulations: Mono-Lyz2, Mono-S100A4, Mono1, Macro-Grk3, and Macro-Cxcl3. One cluster lacking significant marker genes was designated as an unknown cell type (Figure 4A). The classification accuracy was validated by characterizing established marker genes (Figure 4B). Notably, Mono-S100A4 and Mono1 subpopulations were specifically enriched in sepsis-induced ALI, demonstrating significantly higher proportions compared to controls (Figure 4C). We identified the DEGs of Mono-S100A4 and Mono1 subpopulations, with S100A4, Atf3, Sirt1, Plk1, Hmgb1, and Ptx3 emerging as key regulators of septic lung injury (Figure 4D). In sepsis-induced ALI, cellular subsets function through coordinated intercellular communication rather than autonomously. Systematic cell-cell interaction analysis revealed significant heterogeneity in signaling strength across subpopulations (Figure 4E and F). Notably, the Mono-S100A4 subset exhibits heightened communication potential, with significantly amplified signaling toward both Macro-Grk3 and Macro-Cxcl3 compared to other cellular pairs. Analysis of ligand-receptor pairs showed that Mono-S100A4 predominantly influenced other subclusters through Macrophage migration inhibitory factor (MIF)-(CD74+CD44) axis (Figure 4G). These findings collectively demonstrate a strong association between the Mono-S100A4 subset and sepsis-induced ALI pathogenesis, which correlates with the upregulation of the pro-inflammatory MIF-(CD74+CD44) signaling pathway. Spatial transcriptomic profiling confirmed pan-pulmonary upregulation of S100A4, MIF, CD74, and CD44, albeit with varying expression intensities (Figure 4H).
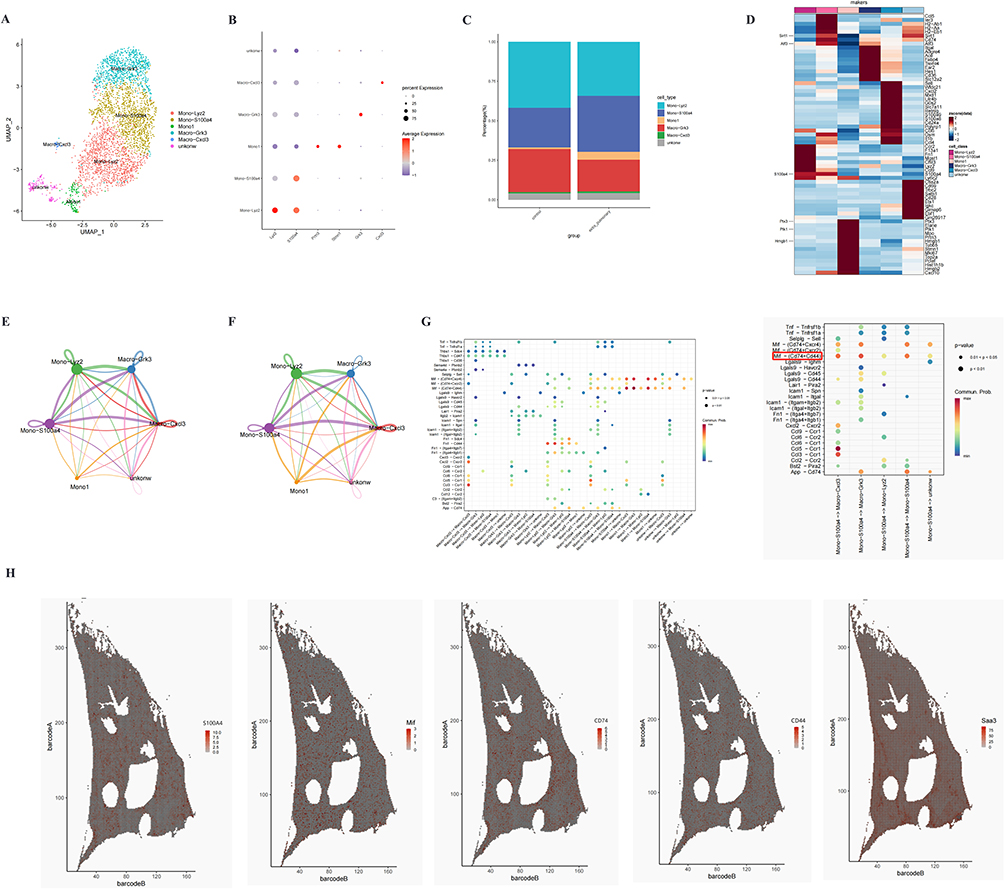 |
Figure 4 Cellular composition of monocyte in normal and septic lung tissues. (A) UMAP visualization identified six distinct monocyte clusters. (B) Bubble plot showing cell type-specific marker gene expression, with bubble size representing the proportion of expressing cells and color intensity indicating average expression levels. (C) Proportional graph showing the proportions of different monocyte subsets in control and ALI groups. (D) Heatmap displaying DEGs in Mono-S100A4 from the ALI group. (E and F) Network diagram of the number/intensity of cellular communication in the ALI group, with bubble size corresponding to the number of cells, line color denotes signal-sending cells, line thickness reflects the magnitude of the number/intensity, and arrow direction points to signal-receiving cells. (G) The interaction probabilities and associated p-values for ligand-receptor pairs between Mono-S100A4 and other monocytic subpopulations are shown, with p<0.05 indicating statistically significant interactions. (H) Spatial expression patterns of S100A4, MIF, CD74, and CD44 in septic lung tissue. |
Functional Characterization of the Pathogenic Mono-S100A4 Subset in Sepsis-Induced ALI
Our investigation focused on elucidating the pathogenic role of the Mono-S100A4 monocyte subset in sepsis-induced ALI. GO enrichment analysis identified leukocyte cell-cell adhesion and response to lipopolysaccharide as the most significantly enriched biological processes (Figure 5A). Leukocyte cell-cell adhesion is a critical immunological mechanism that coordinates cellular migration, activation, and effector functions through specific molecular interactions between leukocytes and vascular endothelial cells or other immune cells. These findings position Mono-S100A4 as a key regulator in the pathogenesis of sepsis-associated extrapulmonary organ injury. KEGG pathway analysis demonstrated significant enrichment in key inflammatory pathways, including the cytosolic DNA-sensing pathway, TNF signaling pathway, Toll-like receptor signaling pathway, NOD-like receptor signaling pathway, IL-17 signaling pathway, and Th17 cell differentiation (Figure 5B). GSEA further confirmed significant activation of the MAPK signaling pathway, Th17 cell differentiation, IL-17 signaling pathway, and PI3K-Akt signaling pathway (all NES > 1.5) (Figure 5C).
 |
Figure 5 Functional characterization of the Mono-S100A4 subset in septic ALI. (A) GO enrichment analysis of upregulated genes in Mono-S100A4 cells. (B) Top 30 significantly enriched KEGG pathways in Mono-S100A4. (C) GSEA of selected KEGG signaling pathways. |
Experimental Validation Confirms That the Mono-S100A4 Subset is Closely Associated with the Activation of the Th17 Signaling Pathway via the MIF-(CD74+CD44) Axis
Further in vivo studies were conducted to elucidate the pathogenesis of sepsis-induced ALI. As demonstrated, intraperitoneal injection of LPS in mice induced significant pulmonary pathological changes, including markedly thickened alveolar walls, interstitial congestion and edema, and extensive inflammatory cell infiltration. Consistently, histopathological scoring based on HE staining revealed a significant increase in the model group compared with the control group (P=0.034) (Figure 6A). By performing immunofluorescence labeling for S100A4, we evaluated the expression level and spatial distribution of the Mono-S100A4 subset in lung tissue during sepsis-induced ALI. The results demonstrated a significant increase in S100A4 protein expression, particularly in peribronchial areas (Figure 6B), which is consistent with the findings from our spatial transcriptomic analysis. IHC showed significantly higher protein levels of CD74, MIF and IL-17A in the lung tissues of ALI model mice (Figure 6C), and WB analysis confirmed the increased expression of MIF and IL-17A while also detecting a prominent upregulation of S100A4 protein in these mice (Figure 6D). In addition, RT-qPCR results demonstrated that the expression levels of S100A4, IL-17A, MIF, CD74, and CD44 mRNA were also significantly upregulated in the mice from the ALI model group (Figure 6E).
 |
Figure 6 In vivo experimental validation. (A) HE staining of mouse lung tissues at 20× (scale bar = 2000 μm) and 400× (scale bar = 100 μm) magnification, with corresponding Szapiel scoring (Mean ± SD, n = 3). (B) Representative immunofluorescence staining of S100A4 in mouse lung tissues at 30× (scale bar = 1000 μm) and 400× (scale bar = 100 μm) magnification, with corresponding quantitative analysis of relative fluorescence intensity (mean ± SD, n = 3). (C) Representative immunohistochemical staining of CD74, IL-17A, and MIF in mouse lung tissues at 400× magnification (scale bar = 100 μm), with corresponding quantitative analysis of relative protein levels (mean ± SD, n = 3). (D) Representative Western blot and quantitative analysis of IL-17A, MIF and S100A4 protein levels in mouse lung tissues, with GAPDH as the loading control (Mean ± SD, n = 3). (E) Quantitative analysis of S100A4, MIF, CD74, CD44 and IL-17A mRNA expression in mouse lung tissues (Mean ± SD, n = 4). |
Notably, all the above molecular detection experiments were conducted on whole lung tissue homogenates, and as such cannot precisely distinguish the specific cellular source of the upregulated molecules or definitively confirm that the increased expression of MIF, CD74 and IL-17A is uniquely derived from the Mono-S100A4 subset. Even so, combined with our single-cell and spatial transcriptomic findings, these results can preliminarily infer the potential functional mechanism by which the Mono-S100A4 subset exerts its biological effects in sepsis-induced acute lung injury.
Discussion
Our study identified a critical monocyte subtype, Mono-S100A4, and uncovered a highly consistent set of correlative evidence that Mono-S100A4 is closely linked to the activation of the Th17 signaling pathway through the MIF-(CD74+CD44) axis, which supports the hypothesis that Mono-S100A4 may potentially contribute to the progression of sepsis-induced ALI through this pathway.
The lung represents the primary and most commonly affected organ in multi-organ failure, with the development of ALI serving as a key determinant of mortality in septic patients.28,29 Therefore, understanding the underlying cellular mechanisms of sepsis-induced ALI is critically important. Our scRNA-seq data revealed consistently lower levels of Th cells, Treg cells, and B cells in ALI, suggesting that the systemic inflammatory response may trigger compensatory immune suppression.30 In contrast, monocytes, neutrophils, and AT II cells showed a marked increase. These alterations in inflammation-associated cell populations correlate with the elevated systemic inflammatory markers observed clinically in sepsis-related ALI, compared to pneumonia-induced ALI.31–33
Experimental data from animal studies reveal that depletion of peripheral blood monocytes mitigates ALI.34 The monocyte-macrophage system serves as a pivotal regulator in both the pathogenesis and evolution of ALI and its progression to ARDS.8 Macrophages originate from two distinct lineages:tissue resident macrophages (TR-Mφs) and monocyte-derived macrophages (Mo-Mφs). During pulmonary infection, circulating Ly6Chigh classical monocytes extravasate and undergo terminal differentiation into Mo-Mφs, ultimately displacing the resident macrophage population.35,36 These findings strongly suggest that Monocytes play a crucial role in ALI, consistent with our scRNA-seq results. Complementing the single-cell transcriptomic data, ST analysis revealed compartmentalized distribution patterns of monocytes in septic ALI, showing significant infiltration enrichment in peritracheal and peribronchial regions. Further characterization identified distinct monocyte subsets associated with sepsis-induced ALI pathogenesis, with the Mono-S100A4 subpopulation exhibiting unique spatial and molecular features.
S100A4, a core DEG in the Mono-S100A4 subpopulation, is a multifunctional calcium-binding protein with significant pathophysiological relevance. As a secreted factor at inflammatory sites, S100A4 promotes cell migration, extracellular matrix remodeling, autophagy, and angiogenesis.37 Under pathological conditions, extracellular S100A4 functions as a potent paracrine activator, stimulating robust secretion of pro-inflammatory cytokines and growth factors that amplify disease progression.38 Suzuki T et al39 demonstrated significantly increased S100A4 expression in isolated pulmonary vascular endothelial cells from LPS-treated mice. Li Y et al40 demonstrated that S100A4 mediates bleomycin-induced pulmonary fibrosis by activating fibroblast functionality. Xu Wu et al37 established the S100A4/STAT3 signaling axis as a critical pathogenic driver in sepsis-induced ALI, revealing its central role in orchestrating inflammatory cascades and tissue damage. In our study, spatial transcriptomics was used to visualize S100A4 in ALI tissues, showing a diffuse increase in lung tissues. Collectively, these findings position S100A4 as a master inflammatory trigger and a highly probable novel therapeutic target whose specific role in Mono-S100A4-mediated ALI requires further functional validation.
Intercellular communication mapping provides insights into the molecular dialogue between cellular subpopulations. Our CellChat analysis revealed that Monocytes orchestrate crosstalk with macrophages and AT II cells via the MIF-(CD74+CD44) signaling axis. Intriguingly, the Mono−S100A4 subset exhibits broader regulatory potential, modulating six other subclusters through this conserved pathway, suggesting that MIF is important for ALI, which is co-expressed with CD74 and CXCR4. Spatial transcriptomics confirmed pan-pulmonary upregulation of MIF, CD74, and CD44 expression in injured tissues. KEGG pathway analysis of Mono-S100A4 identified significant enrichment in key inflammatory pathways, including the cytosolic DNA-sensing pathway, TNF signaling pathway, Toll-like receptor signaling pathway, NOD-like receptor signaling pathway, and IL-17 signaling pathway. Collectively, these findings strongly suggest that Mono-S100A4 is closely correlated with the activation of downstream inflammatory pathways through the MIF-(CD74+CD44) axis, and this correlative pattern implies a potential critical role of Mono-S100A4 in sepsis-induced ALI, with the causal regulation yet to be directly verified by functional experiments.
MIF is a pleiotropic cytokine that enhances inflammatory responses through glucocorticoid antagonism, promotion of inflammatory cell survival, and modulation of angiogenic signaling and endocrine functions.41,42 The biological activity of MIF is mediated through autocrine/paracrine mechanisms involving multiple receptors including CD74, CXCR2, CXCR4, and CXCR7. While CD74 serves as a high-affinity binding site for MIF, it depends on the presence of CD44 or CXCR receptors to propagate intracellular signals.43 Therefore, the functional receptor for MIF signaling is a complex consisting of the ligand-binding protein CD74 and the signal transducer CD44, which triggers PI3K-Akt and ERK1/2 cascade activation.44,45 GSEA revealed significant activation of the PI3K-AKT signaling pathway (NES = 1.79), further corroborating the pivotal role of the MIF-(CD74+CD44) axis in sepsis-induced ALI. Fei Zhang et al46 propose that MIF engagement with the CD74/CD44 complex could trigger the p38 MAPK signaling cascade, subsequently inducing Th17 cell differentiation and increasing the level of IL-17 A. This is likely due to the nuclear translocation of p38 MAPK, which facilitates and sustains the Th17 cell transcription factor RORγt, thereby contributing to Th17 cell differentiation.47,48 Overactivation of the Th17 cell lineage may drive increased secretion of inflammatory mediators like IL-17A, exacerbating the overall inflammatory cascade.49 This finding is consistent with our GSEA results of Mono-S100A4. Furthermore, our in vivo murine experiments revealed that the expression levels of S100A4, IL-17A, MIF, CD74, and CD44 mRNA, as well as the protein levels of S100A4, CD74, MIF, and IL-17A, were significantly elevated in the lung tissues of mice in the model group. These results further substantiate our correlative hypothesis that Mono-S100A4 is associated with the activation of the Th17 signaling pathway via the MIF-(CD74+CD44) axis, which is a key unvalidated causal hypothesis requiring direct functional experimental confirmation. This also represents a highly promising novel therapeutic target.
Sepsis-induced ALI is characterized by complex immune dysregulation. Our previous work confirmed that Xuanfei Baidu Decoction (XFBD) protects against sepsis-induced ALI by activating GPR18 to suppress inflammatory signaling pathways, while simultaneously restoring gut microbial homeostasis.50 Although that study highlighted the therapeutic potential of traditional Chinese medicine in ALI, it primarily addressed the systemic effects of pharmacological intervention, without deeply exploring the functional heterogeneity of key immune cells such as monocytes and their spatial regulatory networks within the septic lung microenvironment. In contrast, the present study introduces innovations and extends our previous research through three core advances: First, we shift the focus from pharmacological mechanisms of therapeutic intervention to the intrinsic remodeling of immune cells in the septic lung, and apply single-cell RNA sequencing to dissect the heterogeneity of pulmonary monocytes in sepsis-induced ALI, thereby identifying a distinct Mono-S100A4 subset that is specifically enriched in the disease state. Second, we integrate ST to map the spatial localization of monocyte subsets, revealing that Mono-S100A4-mediated inflammatory infiltration preferentially occurs in bronchial and bronchiole regions—this spatial resolution complements the previous study’s lack of insights into the lung microenvironmental distribution of immune responses. Third, we clarify the unique molecular mechanism of Mono-S100A4 in promoting ALI progression via the MIF-(CD74+CD44) axis to activate the Th17 signaling pathway, which not only enriches the understanding of sepsis-induced ALI pathogenesis but also provides a more precise cellular target for future therapeutic interventions, complementing the previous study’s focus on broad inflammatory pathway regulation.
This study builds on our team’s prior work by further elucidating immune cell dynamics in sepsis-induced ALI, thereby laying a solid foundation for the development of targeted immunotherapies for this condition.
The present study has several limitations that need to be acknowledged and addressed in subsequent research. First, the public scRNA-seq and spatial transcriptomic datasets (GSE264032 and GSE224146) lack explicit sample size information in original annotations, which may compromise the statistical robustness of bioinformatic analyses; additionally, the small sample size of in vivo animal experiments (n=6 per group) reduces statistical power and may increase the risk of spurious molecular expression results. Second, all bioinformatic findings in this study are of a correlative nature; the inferences about the interaction between Mono-S100A4 and the MIF-(CD74+CD44)-Th17 axis are based on indirect evidence from cell communication and pathway enrichment analyses, without direct causal verification. Third, in vivo validation only confirmed key molecule upregulation in whole lung tissue, lacking cell-type-specific evidence to attribute the elevated MIF, CD74 and IL-17A specifically to the Mono-S100A4 subset. Fourth, the origin of the Mono-S100A4 subset remains unelucidated.
Conclusion
Analysis of scRNA-seq and spatial transcriptomics data (Figure 7), combined with in vivo experiments, reveals substantial heterogeneity in transcriptional profiles, cellular interactions, and spatial organization within ALI in mice. Notably, we identified a unique monocyte subtype, Mono-S100A4, and found a robust correlative relationship between Mono-S100A4 and the activation of the Th17 signaling pathway via the MIF-(CD74+CD44) axis; this series of consistent correlative evidence supports a potential pathogenic role of Mono-S100A4 in sepsis-induced ALI, yet the causal link between them remains to be confirmed. These correlative findings lay a foundation for translating molecular subphenotypes into tailored clinical approaches for ARDS by establishing a potential link between the Mono-S100A4 subset and a key injury pathway. Future studies will further verify the causal relationship between Mono-S100A4 and the MIF-(CD74+CD44)-Th17 axis via functional genetic experiments (eg., Mono-S100A4 knockout/knockdown, MIF/CD74/CD44 pathway inhibition) and explore the translational potential of these findings in patient-derived samples.
 |
Figure 7 This study employs a multi-omics approach to unravel the pivotal role of monocyte heterogeneity in sepsis-induced ALI, with further validation through in vivo experiments. Through scRNA-seq analysis, we identify distinct transcriptional reprogramming across pulmonary cell types and characterize a novel monocyte subset remodeling during sepsis. Spatial transcriptomics further deciphers the pathogenic spatial reconfiguration of cellular niches in ALI, providing a comprehensive view of monocyte-driven pathogenesis. Finally, the specific mechanisms of the novel monocyte subset were validated through in vivo experiments. |
Data Sharing Statement
The data presented in the study are deposited in the GEO-NCBI repository, accession numbers GSE264032 and GSE224146. Additional requests regarding data access may be addressed to the corresponding author.
Ethical Approval
This research followed the ARRIVE guidelines; all animal procedures were performed in compliance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and the experimental procedures were approved by the Medical Ethics Committee of Tianjin Nankai Hospital (NKYY-DWLL-2023-039).
Acknowledgments
The authors would like to thank the researchers who provided open access to the raw data.
Author Contributions
Aihua Wang: Writing - original draft, Writing - review and editing, Conceptualization, Methodology.
Chengfen Yin: Writing - original draft, Writing - review and editing, Conceptualization, Methodology.
Yue Jin: Writing - original draft, Writing - review and editing, Conceptualization, Methodology.
Lei Yang: Writing - review and editing, Investigation, Validation.
Suiran Li: Writing - review and editing, Investigation, Validation.
Jiaxin Li: Writing - review and editing, Data curation, Formal analysis.
Lei Xu: Writing - review and editing, Data curation, Formal analysis.
Ruoyu Shi: Writing - review and editing, Software, Visualization.
Maotong Weng: Writing - review and editing, Software, Visualization.
Wei Liu: Writing - review and editing, Funding acquisition, Project administration, Resources, Supervision.
Ximo Wang: Writing - review and editing, Funding acquisition, Project administration, Resources, Supervision.
All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by the Tianjin Key Research Project in Traditional Chinese Medicine (2025019) and the Research Project on the Integration of Traditional Chinese Medicine and Western Medicine by Tianjin Municipal Health Commission (2023221).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Singer M, Deutschman CS, Seymour CW, et al. The third international consensus definitions for sepsis and septic shock (Sepsis-3). JAMA. 2016;315(8):801–17. doi:10.1001/jama.2016.0287
2. Levy MM, Fink MP, Marshall JC, et al. 2001 SCCM/ESICM/ACCP/ATS/SIS international sepsis definitions conference. Crit Care Med. 2003;31(4):1250–1256. doi:10.1097/01.CCM.0000050454.01978.3B
3. Rudd KE, Johnson SC, Agesa KM, et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: analysis for the Global Burden of Disease Study. Lancet. 2020;395(10219):200–211. doi:10.1016/S0140-6736(19)32989-7
4. Angus DC, van der Poll T. Severe sepsis and septic shock. N Engl J Med. 2013;369(9):840–851. doi:10.1056/NEJMra1208623
5. Cohen J. The immunopathogenesis of sepsis. Nature. 2002;420(6917):885–891. doi:10.1038/nature01326
6. Sevransky JE, Levy MM, Marini JJ. Mechanical ventilation in sepsis-induced acute lung injury/acute respiratory distress syndrome: an evidence-based review. Crit Care Med. 2004;32(Supplement):S548–S553. doi:10.1097/01.CCM.0000145947.19077.25
7. Sheu -C-C, Gong MN, Zhai R, et al. Clinical characteristics and outcomes of sepsis-related vs non-sepsis-related ARDS. Chest. 2010;138(3):559–567. doi:10.1378/chest.09-2933
8. Meyer NJ, Gattinoni L, Calfee CS. Acute respiratory distress syndrome. Lancet. 2021;398(10300):622–637. doi:10.1016/S0140-6736(21)00439-6
9. Wang L, Tang Y, Tang J, et al. Endothelial cell-derived extracellular vesicles expressing surface VCAM1 promote sepsis-related acute lung injury by targeting and reprogramming monocytes. J Extracell Vesicles. 2024;13(3):e12423. doi:10.1002/jev2.12423
10. Janicova A, Becker N, Xu B, et al. Endogenous uteroglobin as intrinsic anti-inflammatory signal modulates monocyte and macrophage subsets distribution upon sepsis induced lung injury. Front Immunol. 2019;10:2276. doi:10.3389/fimmu.2019.02276
11. Santos SS, Carmo AM, Brunialti MKC, et al. Modulation of monocytes in septic patients: preserved phagocytic activity, increased ROS and NO generation, and decreased production of inflammatory cytokines. Intensive Care Med Exp. 2016;4(1):5. doi:10.1186/s40635-016-0078-1
12. Tang F, Barbacioru C, Wang Y, et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat Methods. 2009;6(5):377–382. doi:10.1038/nmeth.1315
13. Cheng C, Chen W, Jin H, et al. A review of single-cell RNA-Seq annotation, integration, and cell–cell communication. Cells. 2023;12(15):1970. doi:10.3390/cells12151970
14. Jovic D, Liang X, Zeng H, et al. Single-cell RNA sequencing technologies and applications: a brief overview. Clin Transl Med. 2022;12(3):e694. doi:10.1002/ctm2.694
15. Tian L, Chen F, Macosko EZ. The expanding vistas of spatial transcriptomics. Nat Biotechnol. 2023;41(6):773–782. doi:10.1038/s41587-022-01448-2
16. Rao A, Barkley D, Franca GS, et al. Exploring tissue architecture using spatial transcriptomics. Nature. 2021;596(7871):211–220. doi:10.1038/s41586-021-03634-9
17. Hao Y, Hao S, Andersen-Nissen E, et al. Integrated analysis of multimodal single-cell data. Cell. 2021;184(13):3573–3587. doi:10.1016/j.cell.2021.04.048
18. Yu X, Xie L, Ge J, et al. Integrating single-cell RNA-seq and spatial transcriptomics reveals MDK-NCL dependent immunosuppressive environment in endometrial carcinoma. Front Immunol. 2023;14:1145300. doi:10.3389/fimmu.2023.1145300
19. Yip SH, Sham PC, Wang J. Evaluation of tools for highly variable gene discovery from single-cell RNA-seq data. Brief Bioinform. 2019;20(4):1583–1589. doi:10.1093/bib/bby011
20. Jin S, Guerrero-Juarez CF, Zhang L, et al. Inference and analysis of cell-cell communication using CellChat. Nat Commun. 2021;12(1):1088. doi:10.1038/s41467-021-21246-9
21. Qiu X, Mao Q, Tang Y, et al. Reversed graph embedding resolves complex single-cell trajectories. Nat Methods. 2017;14(10):979–982. doi:10.1038/nmeth.4402
22. Li W, Wang S, Qiu C, et al. Comprehensive bioinformatics analysis of acquired progesterone resistance in endometrial cancer cell line. J Transl Med. 2019;17(1):58. doi:10.1186/s12967-019-1814-6
23. Yang R, Zheng T, Xiang H, et al. Lung single-cell RNA profiling reveals response of pulmonary capillary to sepsis-induced acute lung injury. Front Immunol. 2024;15:1308915. doi:10.3389/fimmu.2024.1308915
24. Zhao J, Zhen N, Zhou Q, et al. NETs promote inflammatory injury by activating cGAS-STING pathway in acute lung injury. Int J Mol Sci. 2023;24(6).
25. Tang J, Xu L, Zeng Y, et al. Effect of gut microbiota on LPS-induced acute lung injury by regulating the TLR4/NF-kB signaling pathway. Int Immunopharmacol. 2021;91:107272. doi:10.1016/j.intimp.2020.107272
26. Kang Z-Y, Huang Q-Y, Zhen N-X, et al. Heterogeneity of immune cells and their communications unveiled by transcriptome profiling in acute inflammatory lung injury. Front Immunol. 2024;15:1382449. doi:10.3389/fimmu.2024.1382449
27. Yang L, Gao Q, Li Q, et al. PD-L1 blockade improves survival in sepsis by reversing monocyte dysfunction and immune disorder. Inflammation. 2024;47(1):114–128. doi:10.1007/s10753-023-01897-0
28. Lagu T, Rothberg MB, Shieh M-S, et al. Hospitalizations, costs, and outcomes of severe sepsis in the United States 2003 to 2007. Crit Care Med. 2012;40(3):754–761. doi:10.1097/CCM.0b013e318232db65
29. Park I, Kim M, Choe K, et al. Neutrophils disturb pulmonary microcirculation in sepsis-induced acute lung injury. Eur Respir J. 2019;53(3):1800786. doi:10.1183/13993003.00786-2018
30. Nascimento DC, Melo PH, Pineros AR, et al. IL-33 contributes to sepsis-induced long-term immunosuppression by expanding the regulatory T cell population. Nat Commun. 2017;8(1):14919. doi:10.1038/ncomms14919
31. Wendel PD, Caccioppola A, Coppola S, et al. Latent class analysis to predict intensive care outcomes in Acute Respiratory Distress Syndrome: a proposal of two pulmonary phenotypes. Crit Care. 2021;25(1):154. doi:10.1186/s13054-021-03578-6
32. Khan YA, Fan E, Ferguson ND. Precision medicine and heterogeneity of treatment effect in therapies for ARDS. Chest. 2021;160(5):1729–1738. doi:10.1016/j.chest.2021.07.009
33. Wang F, Chen M, Ma J, et al. Integrating bulk and single-cell sequencing reveals the phenotype-associated cell subpopulations in sepsis-induced acute lung injury. Front Immunol. 2022;13:981784. doi:10.3389/fimmu.2022.981784
34. Dhaliwal K, Scholefield E, Ferenbach D, et al. Monocytes control second-phase neutrophil emigration in established lipopolysaccharide-induced murine lung injury. Am J Respir Crit Care Med. 2012;186(6):514–524. doi:10.1164/rccm.201112-2132OC
35. Mass E, Nimmerjahn F, KierdORF K, et al. Tissue-specific macrophages: how they develop and choreograph tissue biology. Nat Rev Immunol. 2023;23(9):563–579. doi:10.1038/s41577-023-00848-y
36. Park MD, Silvin A, Ginhoux F, et al. Macrophages in health and disease. Cell. 2022;185(23):4259–4279. doi:10.1016/j.cell.2022.10.007
37. Wu X, Xuan W, Yang X, et al. Ficolin A knockout alleviates sepsis-induced severe lung injury in mice by restoring gut Akkermansia to inhibit S100A4/STAT3 pathway. Int Immunopharmacol. 2023;121:110548. doi:10.1016/j.intimp.2023.110548
38. Li Z, Li Y, Liu S, et al. Extracellular S100A4 as a key player in fibrotic diseases. J Cell Mol Med. 2020;24(11):5973–5983. doi:10.1111/jcmm.15259
39. Suzuki T, Tada Y, Gladson S, et al. Vildagliptin ameliorates pulmonary fibrosis in lipopolysaccharide-induced lung injury by inhibiting endothelial-to-mesenchymal transition. Respir Res. 2017;18(1):177. doi:10.1186/s12931-017-0660-4
40. Li Y, Bao J, Bian Y, et al. S100A4+ macrophages are necessary for pulmonary fibrosis by activating lung fibroblasts. Front Immunol. 2018;9:1776. doi:10.3389/fimmu.2018.01776
41. Jankauskas SS, Wong D, Bucala R, et al. Evolving complexity of MIF signaling. Cell Signal. 2019;57:76–88. doi:10.1016/j.cellsig.2019.01.006
42. He Y, Li D, Zhang M, et al. Bioinformatic analysis reveals the relationship between macrophage infiltration and Cybb downregulation in hyperoxia-induced bronchopulmonary dysplasia. Scientific Reports. 2024;14(1):20089. doi:10.1038/s41598-024-70877-7
43. Shi X, Leng L, Wang T, et al. CD44 is the signaling component of the macrophage migration inhibitory factor-CD74 receptor complex. Immunity. 2006;25(4):595–606. doi:10.1016/j.immuni.2006.08.020
44. Amin MA, Haas CS, Zhu K, et al. Migration inhibitory factor up-regulates vascular cell adhesion molecule-1 and intercellular adhesion molecule-1 via Src, PI3 kinase, and NFκB. Blood. 2006;107(6):2252–2261. doi:10.1182/blood-2005-05-2011
45. Kang I, Bucala R. The immunobiology of MIF: function, genetics and prospects for precision medicine. Nat Rev Rheumatol. 2019;15(7):427–437. doi:10.1038/s41584-019-0238-2
46. Zhang F, Feng R, Meng T, et al. MIF regulates T helper 17 cell differentiation by activating the p38 MAPK signaling pathway to drive the pathogenesis of EAP. Int Immunopharmacol. 2025;159:114959. doi:10.1016/j.intimp.2025.114959
47. Di Mitri D, Sambucci M, Loiarro M, et al. The p38 mitogen-activated protein kinase cascade modulates T helper type 17 differentiation and functionality in multiple sclerosis. Immunology. 2015;146(2):251–263. doi:10.1111/imm.12497
48. Li W, Zhang Z, Zhang K, et al. Arctigenin suppress Th17 cells and ameliorates experimental autoimmune encephalomyelitis through AMPK and PPAR-γ/ROR-γt signaling. Mol Neurobiol. 2016;53(8):5356–5366. doi:10.1007/s12035-015-9462-1
49. Miossec P, Kolls JK. Targeting IL-17 and TH17 cells in chronic inflammation. Nat Rev Drug Discov. 2012;11(10):763–776. doi:10.1038/nrd3794
50. Yang L, Zhang S, Cui L, et al. Xuanfei baidu decoction alleviated sepsis-induced ALI by modulating gut microbial homeostasis and promoting inflammation resolution: bioinformatics and experimental study. ACS Omega. 2025;10(13):13105–13121. doi:10.1021/acsomega.4c10575
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Proteomics Combined with RNA Sequencing to Screen Biomarkers of Sepsis
Wang C, Li Y, Li S, Chen M, Hu Y
Infection and Drug Resistance 2022, 15:5575-5587
Published Date: 21 September 2022
Melatonin Attenuates Sepsis-Induced Acute Lung Injury via Inhibiting Excessive Mitophagy
Ling J, Yu S, Xiong F, Xu T, Li S
Drug Design, Development and Therapy 2023, 17:2775-2786
Published Date: 11 September 2023
The Emerging Roles of Ferroptosis in Pathophysiology and Treatment of Acute Lung Injury
Wang Y, Zhao Z, Xiao Z
Journal of Inflammation Research 2023, 16:4073-4085
Published Date: 14 September 2023
Synergistic Enhancement of Isoforskolin and Dexamethasone Against Sepsis and Acute Lung Injury Mouse Models
Fang Y, Xiao C, Wang L, Wang Y, Zeng J, Liang Y, Huang R, Shi Y, Wu S, Du X, Sun S, Li M, Zheng Y, Wu H, Guo Q, Yang W
Journal of Inflammation Research 2023, 16:5989-6001
Published Date: 7 December 2023
Moderate Hypothermia Alleviates Sepsis-Associated Acute Lung Injury by Suppressing Ferroptosis Induced by Excessive Inflammation and Oxidative Stress via the Keap1/GSK3β/Nrf2/GPX4 Signaling Pathway
Xu J, Tao L, Jiang L, Lai J, Hu J, Tang Z
Journal of Inflammation Research 2024, 17:7687-7704
Published Date: 25 October 2024