")
Back to Journals » Journal of Inflammation Research » Volume 16
The Emerging Roles of Ferroptosis in Pathophysiology and Treatment of Acute Lung Injury
Authors Wang Y, Zhao Z, Xiao Z
Received 9 May 2023
Accepted for publication 2 September 2023
Published 14 September 2023 Volume 2023:16 Pages 4073—4085
DOI https://doi.org/10.2147/JIR.S420676
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Ning Quan
Yufei Wang,1,2,* Zijun Zhao,2,3,* Zhaoyang Xiao1,2
1Department of Anesthesiology, The Second Affiliated Hospital of Dalian Medical University, Dalian City, Liaoning Province, People’s Republic of China; 2School of Graduates, Dalian Medical University, Dalian, Liaoning Province, People’s Republic of China; 3Department of Anesthesiology, The Affiliated Taizhou People’s Hospital of Nanjing Medical University, Taizhou School of Clinical Medicine, Nanjing Medical University, Taizhou City, Jiangsu Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhaoyang Xiao, Department of Anesthesiology, The Second Affiliated Hospital of Dalian Medical University, No. 467 Zhongshan Road, Shahekou District, Dalian, Liaoning Province, 116027, People’s Republic of China, Email [email protected]
Abstract: Ferroptosis, a programmed cell death discovered in recent years, is an iron-dependent lipid peroxidation accumulation. Unlike other modes of cell death (autophagy, necroptosis, pyroptosis, cuproptosis, etc.), ferroptosis has unique morphological characteristics and plays an important role in a variety of diseases. In recent years, there has been great progress in the study of ferroptosis. Studies have found that ferroptosis is associated with acute lung injury (ALI), a condition with a high mortality rate and limited treatment options. This paper summarizes the mechanism of ferroptosis from the perspectives of iron metabolism, lipid metabolism, amino acid metabolism, and glutathione metabolism. It also discusses the research progress of ferroptosis in ALI in order to find new directions for the prevention and treatment of this condition.
Keywords: ferroptosis, acute lung injury, ischemia, sepsis, targeted therapy
Introduction
Cell death plays an important role in promoting organ development and maintaining tissue and cell homeostasis. Cell death is divided into accidental cell death and programmed cell death.1 Programmed cell death is mainly divided into apoptotic and non-apoptotic forms. Ferroptosis is a programmed cell death different from necroptosis and autophagy discovered in recent years and is characterized by iron-dependent lipid peroxides (LPO) accumulation.2,3 ALI refers to alveolar epithelial cells and capillary endothelial cells damage caused by various intrapulmonary and extrapulmonary pathogenic factors, resulting in diffuse pulmonary interstitial and alveolar edema, leading to acute hypoxic respiratory insufficiency. Rapidly developing respiratory failure characterizes the acute, exudative phase of ALI (0–7 days). Within the first 12 to 24 hours following the onset of the disease, tachypnea, tachycardia, and respiratory alkalosis may occur. These symptoms may precede the discovery of diffuse bilateral infiltrates on an X-ray. These findings point to diffuse alveolar damage caused by the breakdown of the epithelial-endothelial barrier, which led to an excessive leakage of blood cells and protein-rich fluid into the interstitium and alveoli. Active neutrophils migrate in reaction to tissue damage, working with platelets, alveolar macrophages, other inflammatory and fixed lung cells to produce a number of substances that exacerbate inflammation but can also serve as biomarkers of the acute stage of the disease.4 ALI has a mortality rate of up to 40% and is a major disease endangering human life and health. The pathogenesis of ALI is very complex and is currently considered to be associated with cytotoxic mechanisms such as inflammation and oxidative stress, but there are no effective targeted therapeutic measures.5 As research has deepened, more evidence suggests that ferroptosis is involved in the pathogenesis of ALI. In recent years, Scientists have confirmed that ferroptosis is associated with ALI caused by ischemia-reperfusion,6–8 sepsis,9,10 radiation,11 drowning12 and oleic acid13 through animal and cell model experiments. However, their specific mechanisms, and potential treatment options remain to be further investigated. In this paper, the mechanism of ferroptosis and its research results in the pathogenesis and treatment of ALI are summarized. It is considered that ferroptosis can be used as a target for the prevention, diagnosis of ALI and shows great prospects in the treatment of ALI.
Materials and Methods
The following databases were searched for articles focusing on ferroptosis and ALI: EMBASE, PubMed, and Google Scholar. The retrieval period spanned from 2012, when ferroptosis was introduced, to January 2023. The papers were extracted using search keywords such as “acute lung injury” with “ischemia”, or “sepsis”, or “radiation”, or “drowning”, or “oleic acid”, and “ferroptosis”; studies written in English were extracted. We considered the manuscript suitable for further analysis if two of the three authors agreed that it met the eligibility criteria.
Mechanism of Ferroptosis
In 2012, Dixon et al13 found that the cause of death in some cells was related to iron ion concentration and named this novel mode of non-apoptotic cell death “ferroptosis” for the first time. Since then, the concept of ferroptosis has been gradually known by international scholars. In 2018, the Cell Death Nomenclature Committee defined ferroptosis as one of the programmed cell death patterns.1
Abnormal Iron Metabolism (Iron Overload)
Iron overload has been found to be one of the key links in ferroptosis. As an essential micronutrient for maintaining the health of the body, iron is mainly present in various cellular activities in the form of Fe2 + or Fe3 +. In normal human body, iron requires the help of transferrin (TF) and TF receptor (TFR) 1 to complete the process of operation, that is, one molecule of TF binds two molecules of Fe3 + to form a complex.14 When extracellular material enters the cell, Fe3 + released from the complex is reduced to Fe2 +, which is then stored in a variable iron pool by divalentmetal-iontransporter-1 (DMT1), and the remaining Fe2 + is oxidized out of the cell and circulates in the body to maintain iron homeostasis.15 When the imbalance of iron in the body initiates, the supply exceeds the demand for iron in the human body, and the excess active iron accumulates in the vital organs and tissue cells of the human body, resulting in iron overload. It produces a large number of cytotoxic hydroxyl radicals and a large number of reactive oxygen species (ROS) under the action of the Fenton reaction, which in turn breaks the molecular structure of genes and destroys vascular endothelial cell membranes, lipids and DNA. Superoxide radicals (O2•-), hydrogen peroxide ((H2O2), hydroxyl radicals (OH•), and singlet oxygen (1O2) are examples of non-free radical oxygen intermediates (peroxides) that are included in ROS. These compounds are produced by diverse cytosolic enzymes, including cyclooxygenases, lipid metabolism inside peroxisomes, and plasma membrane proteins, nicotinamide adenine dinucleotide phosphate (NADPH) oxidases. Despite the fact that all of these sources add to the overall oxidative burden, the majority of cellular ROS (about 90%) are produced in the mitochondria through oxidative phosphorylation.16,17 At the same time, iron also plays a non-negligible role in lipid peroxidation or ferroptosis.18,19 Therefore, ferroptosis is more likely to occur in response to stimulation by iron overload, and abnormal iron metabolism is an important manifestation of the cellular ferroptosis process.
Lipid Peroxidation
One of the most striking features of ferroptosis is plasma membrane damage mediated by lipid peroxide accumulation. Lipidomics have shown that long chain acyl-coenzyme A synthetase 4 (ACSL4) and lysophosphatidylcholine acyl transfer 3 (LPCAT3) play key roles in regulating lipid peroxidation.20,21 ACSL4 catalyzes the combination of acetyl-CoA and polyunsaturated fatty acids (PUFAs) such as arachidonic acid (AA) and adrenic acid (Adrenoyl, AdA) to generate AA-coenzyme A and AdA-coenzyme A, which are esterified to phosphatidylethanolamine (PE) to form AA-PE and AdA-PE under the action of LPCAT3, and LPO under the action of lipoxygenase. LPO is eventually metabolized into the toxic aldehydes 4-hydroxynonenal and malondialdehyde (MDA), causing irreversible damage to the cell membrane and plasma membrane, leading to cell death.22 Therefore, high expression of ACSL4 and LPCAT3 is considered as one of the important hallmarks of ferroptosis. Notably, Doll et al23 and Bersuker et al24 named ferroptosis suppressor protein 1 (FSP1), which uses NAD (P) H to catalyze coenzyme Q10 to promote the expression of lipophilic free radicals, inhibits the membrane lipid peroxidation cascade in the absence of GPX4 content, and prevents the occurrence of ferroptosis. In addition, ferroptosis inducer 56 (FIN56), a novel ferroptosis inducer, can inactivate GPX4 through the mevalonate pathway and inhibit the synthesis of lipophilic antioxidants and promote ferroptosis.25 Interestingly, autophagic degradation of lipid droplets promotes ferroptosis in hepatocytes induced by the ferroptosis activator RSL3, while RSL3-induced LPO with ferroptosis is prevented by promoting lipid storage or inhibiting lipid droplet degradation, suggesting that lipid droplets may play an antioxidant role during ferroptosis.26 The above studies showed that ferroptosis can be regulated by regulating enzymes related to PUFA membrane phospholipid biosynthesis.
Glutamate-Cystine Transport Dysfunction
Cystine/glutamate reverse transporter (systemXc-), which is blocked in glutamate-cystine transport as an important amino acid transporter, plays an important role in maintaining glutamate homeostasis, cystine transport, and glutathione (GSH) synthesis.27 Through this transporter, extracellular cystine can be transported into the intracellular space in healthy organisms to promote GSH synthesis. GSH acts as an antioxidant and detoxifying agent by transforming toxic substances in the body into nontoxic and harmless substances that eventually excrete them from the body. Substantial accumulation of ROS in the body is removed by GSH. GSH can assist GPX4 to complete the antioxidant effect.28,29 When cystine/glutamate reverse transporter is accidentally blocked, the influx of cystine is limited, the production of GSH decreases correspondingly with the decrease of cysteine, then GPX4 reactivity weakens, which further promotes the accumulation of ROS, and increases the probability of ferroptosis30 (Figure 1).
 |
Figure 1 The mechanisms of ferroptosis. Iron accumulation, excessive ROS production and overwhelming lipid peroxidation are hallmarks of ferroptosis. There are three main metabolic pathways that initiate and execute ferroptosis, amino-acid/GSH, lipid, and iron pathways. Moreover, ferroptosis sensitivity is also controlled by additional signaling pathways and regulators. Ferroptosis is illustrated here, demonstrating the key molecules and targets involved in regulating the peroxidation of iron and lipids. Abbreviations: TF, transferrin; TFR1, transferrin receptor 1; ACSL4, acyl-CoA synthetase long-chain family member 4; LPCAT, lysophosphatidylcholine acyltransferase; FIN 56, Ferroptosis-Inducer-56; DMT1, divalent metal transporter 1; FPN1, ferroportin 1; GSH, glutathione; GSSG, oxidized glutathione; GSS, glutathione synthetase; GCL, glutamate-cysteine ligase; GPX4, glutathione peroxidase 4; HO-1, haem oxygenase 1; HSPB1, heat shock protein beta-1; LOX, lipoxygenase; NCOA4, nuclear receptor coactivator 4; NRF2, nuclear factor E2-related factor 2; PUFA, polyunsaturated fatty acid; PE, phosphatidylethanolamine; ROS, reactive oxygen species; RSL3, Ras-selective lethal 3; STEAP3, six-transmembrane epithelial antigen of prostate 3 metalloreductase; SLC7A11, solute carrier family 7 member 11; SLC3A2, solute carrier family 3 member 2; VDAC2/3, voltage dependent-anion channel 2/3. |
Regulatory Mechanism of Ferroptosis
In recent years, scientists have made rapid progress in the study of ferroptosis regulation mechanism. Glutathione peroxidase 4 (GPX4), nuclear factor E2 related factor 2 (Nrf2), and Xc-system have been validated as important regulators of ferroptosis. GPX4 converts intracellular lipid hydrogen peroxide into lipid alcohols, promotes the breakdown of hydrogen peroxide (H2O2), repairs oxidative damage in lipid cells, and protects cell membranes from oxidative damage, and its inactivation will lead to the accumulation of intracellular lipid peroxides and ferroptosis.31 Nrf2 plays a very important role in ferroptosis by regulating anti-inflammatory, iron homeostasis, and lipid peroxidation.32 The Xc − system synthesizes GSH by transporting intracellular glutamate in exchange with extracellular cystine, scavenging intracellular free radicals, and inhibiting system Xc-can lead to a rapid decrease in intracellular GSH levels causing ferroptosis.33 Scientists have confirmed that a variety of compounds can induce and inhibit ferroptosis (Table 1). The above studies reveal that the occurrence and development of ferroptosis are regulated by a variety of cellular components and intracellular signaling pathways. It would be helpful to gain a deeper understanding of ferroptosis, as well as verify the agonists and antagonists that contribute to its onset and development, so that we can work towards developing effective treatment to ALI.
 |
Table 1 Common Ferroptosis Agonists, Antagonists and Their Regulatory Mechanisms |
ALI/ARDS and Its Clinically Relevant Biomarkers
ALI/acute respiratory distress syndrome (ARDS) is a common cause of respiratory failure in critically ill patients and often presents clinically with respiratory distress and progressive hypoxemia. The final severe stage of ALI is defined as ARDS. There are many risk factors inducing ALI/ARDS, common direct factors are severe pulmonary infection, gastric content aspiration, pulmonary contusion, etc., and indirect factors are severe extrapulmonary infection, severe non-chest trauma, severe acute pancreatitis, and massive blood transfusion, etc.57 At present, there is no specific clinical treatment for this disease. Therefore, how to effectively prevent the occurrence of ALI in the early stage and prevent ALI from further developing into ARDS is an important issue that urgently needs to be solved.58 Researchers have investigated blood, pulmonary edema fluid, and bronchoalveolar lavage fluid (BALF), exhaled air for biomarkers of the exudative and proliferative phases of ARDS. Despite their insufficient reliability if considered alone, combinations of biomarkers are recommended. The following table provides a list of the most important biomarkers associated with the exudative and fibroproliferative phases of ARDS4,59–61 (Table 2).
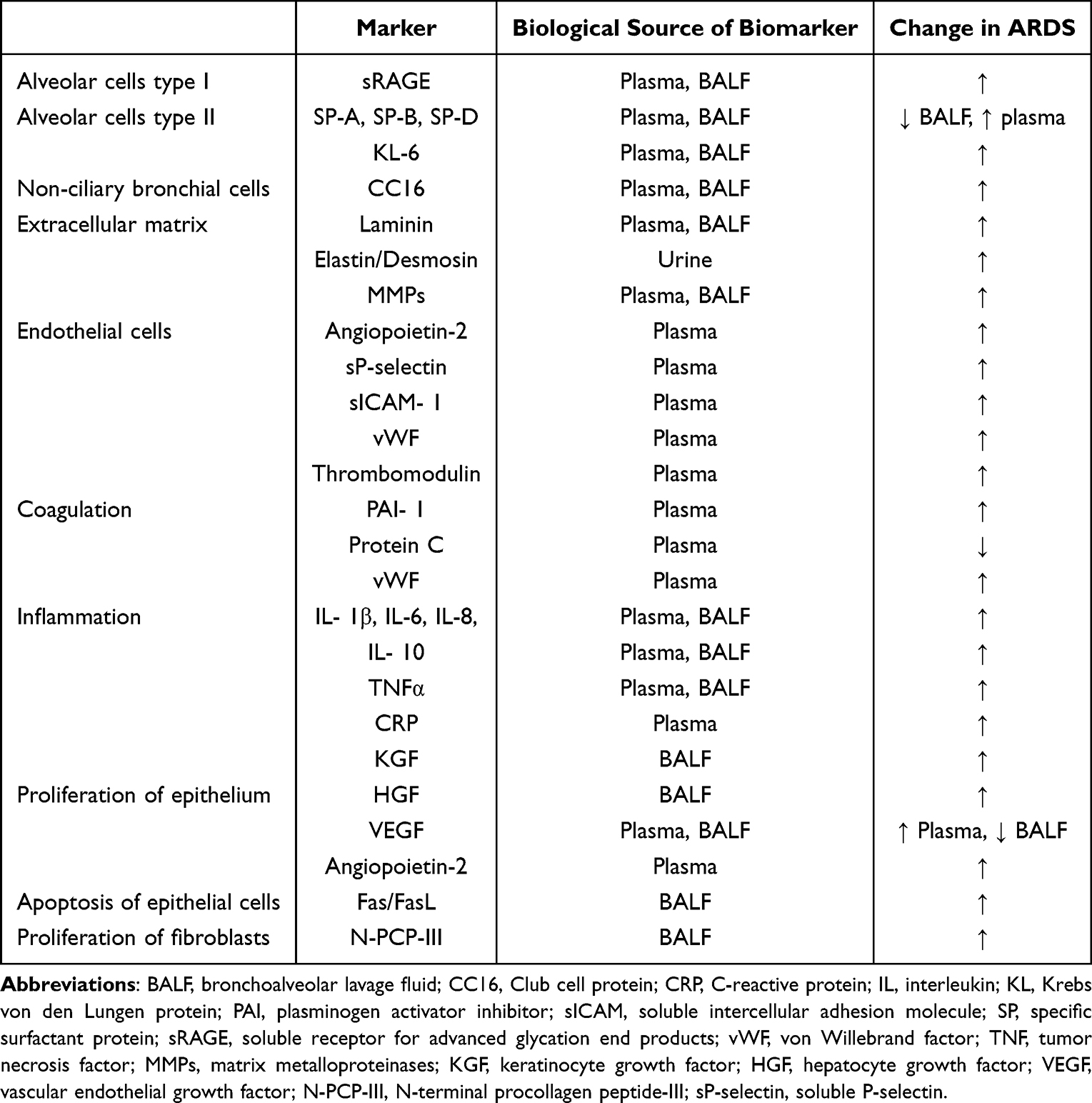 |
Table 2 Biomarkers of Acute (Exudative) and (Fibroproliferative) Phase of ARDS.4,59 |
Ferroptosis and ALI
Normally, iron homeostasis is maintained by phagocytosis of macrophages, transferrin secretion, antioxidant molecules on the epithelial surface of the respiratory tract, and the respiratory ciliary expectoration system in lung tissue. Once the protective mechanism is disrupted by endogenous or exogenous factors, deposition after disturbance of iron metabolism affects the normal function of cells. Iron deposition can be found in the lower respiratory tract of ALI patients, and iron accumulation can lead to inflammatory response, oxidative stress and mitochondrial dysfunction, further aggravating the degree of lung injury.62 Recent studies have shown that ferroptosis is associated with ALI caused by ischemia-reperfusion, sepsis, radiation, drowning, and oleic acid, etc. (Figure 2).
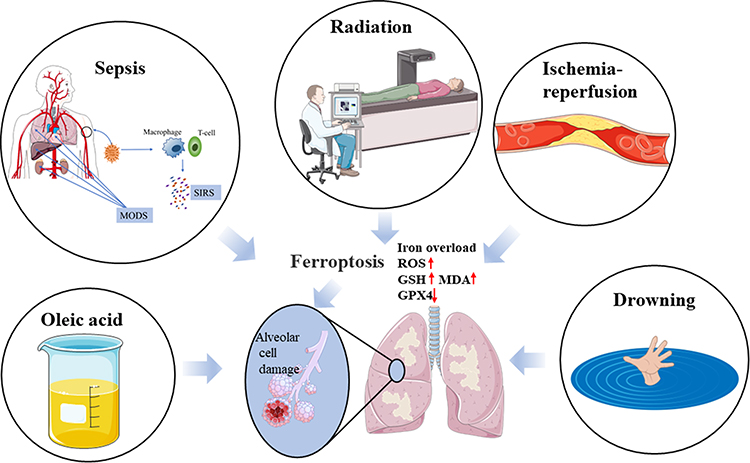 |
Figure 2 Ferroptosis is associated with ALI caused by ischemia-reperfusion, sepsis, radiation, drowning, and oleic acid. |
Ferroptosis Antagonists and ALI
The regulatory mechanism of ferroptosis in ALI is far more complex than previously understood, and although there have been many recent research advances and new findings, the detailed regulatory mechanism of ferroptosis remains unclear. Future studies should focus on the diagnostic and prognostic value of ferroptosis-related genes, and more in-depth studies on their regulatory mechanisms are also needed in order to improve our understanding of the pathogenesis and treatment of ALI. We believe that in the near future, the regulatory mechanisms of ferroptosis will become more and more thoroughly investigated, and ferroptosis will become a new therapeutic strategy for ALI (Table 3). Ferroptosis is expected to be an important target for the treatment of ALI, and its key is to inhibit iron metabolism and lipid peroxidation. Mechanism of various ALI and treatment of ferroptosis antagonists in typical ALI models were representatively elucidated (Table 4 and Figure 3).
 |
Table 3 Ferroptosis Antagonists and ALI |
 |
Table 4 Treatment of Ferroptosis Antagonists in Typical ALI Models |
 |
Figure 3 Mechanism of various ALI and treatment of ferroptosis antagonists in typical ALI models. Abbreviations: ALI, acute lung injury; IRRALI, Ischemia-reperfusion related ALI; RRALI, radiation related acute lung injury; SRALI, sepsis related acute lung injury; DRALI, drowning related acute lung injury; OARALI, oleic acid related acute lung injury. IL, interleukin; TNF, tumor necrosis factor; VEGF, vascular endothelial growth factor; SP, specific surfactant protein. |
Ischemia-Reperfusion Related ALI (IRRALI) and Ferroptosis
Recent studies have shown that ferroptosis is a major cause of ischemic injury. Ferroptosis has been demonstrated in animal models or cellular models of ischemia-reperfusion injury of the myocardium, kidney, liver, intestine, and brain.69–71 Damage away from lung tissue such as abdominal injury, infection, and surgery-induced intestinal ischemia-reperfusion (IR) can cause ALI.72 Li et al6 found that inhibitor of apoptosis stimulating p53 protein (iASPP), a p53 inhibitor, exerted a protective effect against ferroptosis by mediating Nrf2/HIF-1/TF and reduced lung tissue edema, atelectasis, necrosis, alveolar and interstitial inflammation in IR- ALI. p53 plays a critical role in ferroptosis in other disease models, while it is unknown whether iASPP acts by regulating p53 remains to be further investigated. Xu et al63 noticed iron content and lipid peroxidation accumulation in lung tissue and altered expression of key proteins (GPX4 and ACSL4) during reperfusion in a mouse model of IR-ALI. In addition, rosiglitazone, an ACSL4 inhibitor, administered before ischemia alleviated ferroptosis injury in lung tissue, which is consistent with the protective effect of ACSL4 gene knockout on lung epithelial cells. As a result, ACSL4 is an important ferroptosis regulatory gene. However, Nrf2, an antioxidant molecule, regulates downstream HO-1 and signal transducer and activator of transcription 3 (STAT3) phosphorylation expression and inhibits ferroptosis in IR- ALI thereby reducing tissue injury. In a recent study, possible mechanisms of IRRALI were clarified, and its effects on IR-induced ALI were explored. The study demonstrates that isoliquiritin apioside could protect against intestinal IR-induced ALI by reducing lung epithelial ferroptosis in a Hif-1α-dependent manner.7 But so far, the pathogenesis of IR-induced ALI has not been fully elucidated, and there are no specific drugs.32,73 In a study conducted by Dong et al,8 in their results, IR-ALI significantly reduced telomerase reverse transcriptase (TERT) in lung tissue of Nrf2-/-mice. Furthermore, MDA levels increased significantly in ALI models, while GSH and GPX4 levels decreased significantly. Additionally, type II alveolar epithelial cells in IR-ALI model showed typical structural changes. To examine Fe2+ levels and distribution within cells, they used scanning transmission X-ray microscopy (STXM). Their data suggest that Nrf2 can negatively regulate ferroptosis via modulation of TERT and solute carrier family 7, membrane 11 (SLC7A11) levels. As a result of this study, new candidates have been identified for IR-ALI treatment in the future.
Radiation Related ALI (RRALI) and Ferroptosis
RRALI is an important complication after radiotherapy for thoracic tumors, and its incidence is 16.7% ~ 50.3%, which increases the mortality and morbidity of cancer patients.74 Radiation induced a large amount of ROS production and is considered to be an important mechanism causing RRALI, and ferroptosis in lung tissue of mice is aggravated after radiation treatment. In addition, ferroptosis in alveolar endothelial cells in RRALI affects their barrier function, with increased expression of the mechanosensitive calcium channel. Piezo1 increased calcium influx, and increased calpain activity, and the degree of ferroptosis is relieved after the use of its specific inhibitor GsMTx4. Piezo1 regulates ferroptosis through the Ca2 +/calpain/VE- Cadherin pathway, and ferroptosis is associated with Ca2 + overload, but the mechanism remains to be further studied.64 It provides a new target for future mitigation of RRALI by revealing the previously unknown role of Piezo1 in modulating ferroptosis. Superoxide dismutase (SOD) is a typical endogenous antioxidant enzyme that directly inactivates ROS, and the clinical use of SOD and its analogues has been shown to protect the lung from radiation injury.75 Due to the characteristics of poor stability, low cellular uptake, high immunogenicity, and less circulation in the body, the clinical application of natural SOD as a therapeutic drug is extremely limited. Genetic engineering technology encapsulated SOD into a biomimetic nanoreactor composed of linear peptide ARA290-HBc to improve the stability and lung retention of ARA290 peptide, and further studies confirmed that it prevented radiation-induced alveolar epithelial cell apoptosis, ferroptosis, and oxidative stress.74 Although the feasibility of SOD@ ARA29- HBc in clinical application needs to be more widely and experimentally verified, it is undeniable that bio-nanotechnology has great therapeutic prospects.
Sepsis Related ALI (SRALI) and Ferroptosis
Sepsis is secondary to severe trauma, infection and surgery, and may develop into shock and multiple organ dysfunction syndrome after progression, and the lung is one of the organs easily involved. Lipopolysaccharide (LPS), a major component in Gram-negative bacteria, has been used to induce ALI. Liu et al53 further illustrated that ferroptosis is involved in LPS-ALI progression. By using ferrostatin- 1, an inhibitor of ferroptosis, they found that Nrf2/ARE signaling pathway may regulate ferroptosis to participate in LPS-induced ALI.76 An analysis of ferroptosis found that F-box and WD repeat domain containing 7 (FBXW7) mediates ubiquitination and degradation of AUF1. AUF1 inhibits ferroptosis through opposite regulation of NRF2 and activating transcription factor 3 (ATF3). Activating AUF1 pathway may be beneficial to the treatment of SRALI.9 Sevoflurane, a commonly used anesthetic, has a protective effect on ALI, and recent studies have found that its protective effect is partially achieved by inhibiting ferroptosis, but the specific mechanism remains to be further studied, and the practical feasibility for clinical use remains to be studied;65 another study elucidated that the Mucin 1 mechanism inhibits ferroptosis and sensitizes Vitamin E to ameliorate SRALI through GSK3 β/Keap1-Nrf2-GPX4 pathway.67 Tang et al found that ferulic acid may alleviate SRALI by activating the Nrf2/HO-1 pathway and inhibiting ferroptosis, suggesting a new approach for treating sepsis.68 While in terms of traditional Chinese medicine, Li et al66 isolated ginseng epoxyethynol from the roots of ginseng and found that it significantly inhibited ferroptosis and improved lung injury by up-regulating the level of the KEAP1- Nrf2-HO-1 pathway. In a study aiming to explore the regulatory role of itaconate on ferroptosis in SRALI, the researchers found that itaconate inhibits ferroptosis of macrophages via Nrf2 pathways in response to SRALI8.10 In summary, ferroptosis may be a potential therapeutic target for SRALI, and ferroptosis inhibitors have therapeutic potential.
Drowning Related ALI (DRALI) and Ferroptosis
Drowning is one of the main causes of accidental injury death, and ALI is one of the most common complications of drowning. Seawater has a chemical irritating effect on the respiratory tract and alveoli, and a large amount of protein and water exudate into the pulmonary interstitium and alveolar space after alveolar epithelial cells and pulmonary capillary endothelial cells are injured by seawater, causing non-cardiogenic pulmonary edema. Hypoxemia of varying degrees developed, resulting in heart failure and death.77 Because fluid enters the alveoli from the blood vessels, hemoconcentration, hypovolemia, hypoproteinemia, hypernatremia, hypercalcemia, and hypermagnesemia may occur. Hypermagnesemia can cause bradycardia, arrhythmias, conduction block, and even asystole.78 It also depresses central and peripheral nerves, dilates blood vessels, and lowers blood pressure. Using Nrf2-specific agonist (dimethyl fumarate), Nrf2 inhibitor (ML385), Nrf2-knockout mice and ferroptosis inhibitor (Ferrostatin-1), Qiu et al investigated the potential roles of Nrf2 on seawater DRALI and the underlying mechanisms. They found that Nrf2 inhibits ferroptosis, thus alleviating seawater DRALI. The effectiveness of ferroptosis inhibition by Nrf2 provides a novel therapeutic target for seawater DRALI.12
Oleic Acid Related ALI (OARALI) and Ferroptosis
Oleic acid, also called cis-9-octadecenoic acid, is an unsaturated fatty acid containing a double bond in natural oils and is present in natural animal and vegetable oils in the form of glycerides. Oleic acid, especially high purity oleic acid, is an important fine chemical product and can be widely used in paint inks, coatings, mineral flotation agents, thin film antistatic agents, smooth agents, textile additives, explosive emulsifiers and so on. Metal salts of oleic acid are widely used as surfactants and corrosion inhibitors. Oleic acid is modified by functional groups and can be used in lubricating oil, chemical analysis, pharmaceuticals and other industries. Recent study found that oleic acid could cause ALI, and there are three main mechanisms of oleic acid-induced respiratory failure: 1. Oleic acid is a highly toxic fatty acid, which strongly constricts pulmonary micro-vessels after intravenous injection, resulting in fat emboli blocking pulmonary capillaries and causing pulmonary microcirculatory disturbances. 2. Oleic acid can directly stimulate blood vessels to damage the vascular endothelium, increase vascular permeability, pulmonary interstitial edema, and thus causing dyspnea. 3. Oleic acid decreases alveolar surfactant, resulting in pulmonary edema.79,80 Ferroptosis has distinct morphological characteristics that differ from other cell death types. Existing studies have identified that ferroptosis is morphologically characterized by reduced mitochondria, ruptured mitochondrial membrane, reduced mitochondrial ridges, and reduced mitochondrial size. In the model of OARALI in mice, lipid peroxidation, iron overload, GSH depletion and MDA accumulation in lung tissue were observed, the protein expression levels of GPX4 and ferritin in lung tissue were down-regulated, mitochondrial atrophy in lung cells and the occurrence of ferroptosis were found suggesting that ferroptosis plays a potential role in the pathogenesis of OARALI.13,81 The above study attempts to investigate the mechanism of ferroptosis in ALI model induced by oleic acid, and expounds the typical changes of ferroptosis in OARALI model.
The importance of ferroptosis in the onset and development of ALI has been shown by an increasing amount of in vitro and in vivo research in recent years (Table 5). There were also review studies that mainly focused on the mechanism of ferroptosis in a general way, and briefly discussed the emerging role of ferroptosis in ALI,82,83 whereas we not only analyzed the mechanism of ferroptosis in a deeper way, but we also put much more time and effort specifically in better demonstrating the emerging roles of ferroptosis in pathophysiology and treatment of ALI. In summary, recent studies indicate that iron metabolism, oxidative stress, and glutathione potentially contribute to the pathogenesis of ALI. Ferroptosis antagonists are expected to be an important option for the treatment of ALI.
 |
Table 5 Ferroptosis and ALI |
Limitation
There are still many limitations in ferroptosis-related studies. First, the specific mechanism of ferroptosis in ALI has not been fully elucidated, more detailed signal transduction pathways are still being further explored; second, many studies still stay in the experimental stage at the cellular and animal levels, there is still a lack of large-sample, multicenter clinical randomized controlled studies and evidence; Lastly, more comprehensive, in-depth studies are needed to further investigate the relationship between ferroptosis and ALI, and intervention for ALI according to different signal transduction pathways is the direction of future exploration.
Conclusion
As novel programmed cell death, ferroptosis has been demonstrated to play an important role in a variety of systemic diseases. In recent years, scientists have demonstrated that ferroptosis plays an indispensable role in the pathogenesis of ALI from cellular or animal model studies such as IRRALI, SRALI, RRALI, DRALI and OARALI. GPX4 and Nrf2 are the key points in the occurrence and development of ferroptosis in ALI. Ferroptosis antagonists are expected to be an important option for the treatment of ALI. Relying on the latest technology to carry out a large number of basic and clinical research is helpful to further understand the occurrence and development of ALI, provide more reliable means for the prevention, and treatment of ALI.
Data Sharing Statement
All data analyzed were included in this paper; further requests can be consulted and data can be obtained from the correspondent author.
Acknowledgments
Yufei Wang and Zijun Zhao are co-first authors for this study. We highly appreciated Dr. Yunxiang Hu for his persistent support and encouragement, specifically in writing and revising our paper.
Funding
There is no funding to report.
Disclosure
The authors declare no competing interests in this work.
References
1. Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018;25(3):486–541. doi:10.1038/s41418-017-0012-4
2. Kopeina GS, Zhivotovsky B. Programmed cell death: past, present and future. Biochem Biophys Res Commun. 2022;633:55–58. doi:10.1016/j.bbrc.2022.09.022
3. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
4. Mokrá D. Acute lung injury - from pathophysiology to treatment. Physiol Res. 2020;69(Suppl 3):S353–s366. doi:10.33549/physiolres.934602
5. Mowery NT, Terzian WTH, Nelson AC. Acute lung injury. Curr Probl Surg. 2020;57(5):100777. doi:10.1016/j.cpsurg.2020.100777
6. Li Y, Cao Y, Xiao J, et al. Inhibitor of apoptosis-stimulating protein of p53 inhibits ferroptosis and alleviates intestinal ischemia/reperfusion-induced acute lung injury. Cell Death Differ. 2020;27(9):2635–2650. doi:10.1038/s41418-020-0528-x
7. Zhongyin Z, Wei W, Juan X, Guohua F. Isoliquiritin apioside relieves intestinal ischemia/reperfusion-induced acute lung injury by blocking Hif-1α-mediated ferroptosis. Int Immunopharmacol. 2022;108:108852. doi:10.1016/j.intimp.2022.108852
8. Dong H, Xia Y, Jin S, et al. Nrf2 attenuates ferroptosis-mediated IIR-ALI by modulating TERT and SLC7A11. Cell Death Dis. 2021;12(11):1027. doi:10.1038/s41419-021-04307-1
9. Wang Y, Chen D, Xie H, et al. AUF1 protects against ferroptosis to alleviate sepsis-induced acute lung injury by regulating NRF2 and ATF3. Cell Mol Life Sci. 2022;79(5):228. doi:10.1007/s00018-022-04248-8
10. He R, Liu B, Xiong R, et al. Itaconate inhibits ferroptosis of macrophage via Nrf2 pathways against sepsis-induced acute lung injury. Cell Death Discov. 2022;8(1):43. doi:10.1038/s41420-021-00807-3
11. Li X, Zhuang X, Qiao T. Role of ferroptosis in the process of acute radiation-induced lung injury in mice. Biochem Biophys Res Commun. 2019;519(2):240–245. doi:10.1016/j.bbrc.2019.08.165
12. Qiu YB, Wan BB, Liu G, et al. Nrf2 protects against seawater drowning-induced acute lung injury via inhibiting ferroptosis. Respir Res. 2020;21(1):232. doi:10.1186/s12931-020-01500-2
13. Zhou H, Li F, Niu JY, et al. Ferroptosis was involved in the oleic acid-induced acute lung injury in mice. Sheng Li Xue Bao. 2019;71(5):689–697.
14. Xu W, Deng H, Hu S, et al. Role of ferroptosis in lung diseases. J Inflamm Res. 2021;14:2079–2090. doi:10.2147/JIR.S307081
15. Liu J, Kang R, Tang D. Signaling pathways and defense mechanisms of ferroptosis. Febs j. 2022;289(22):7038–7050. doi:10.1111/febs.16059
16. Lambert AJ, Brand MD. Reactive oxygen species production by mitochondria. Methods Mol Biol. 2009;554:165–181.
17. Checa J, Aran JM. Reactive oxygen species: drivers of physiological and pathological processes. J Inflamm Res. 2020;13:1057–1073. doi:10.2147/JIR.S275595
18. Niu B, Liao K, Zhou Y, et al. Application of glutathione depletion in cancer therapy: enhanced ROS-based therapy, ferroptosis, and chemotherapy. Biomaterials. 2021;277:121110. doi:10.1016/j.biomaterials.2021.121110
19. Liu MY, Li HM, Wang XY, et al. TIGAR drives colorectal cancer ferroptosis resistance through ROS/AMPK/SCD1 pathway. Free Radic Biol Med. 2022;182:219–231. doi:10.1016/j.freeradbiomed.2022.03.002
20. Doll S, Proneth B, Tyurina YY, et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat Chem Biol. 2017;13(1):91–98. doi:10.1038/nchembio.2239
21. Ke P, Bao X, Liu C, et al. LPCAT3 is a potential prognostic biomarker and may be correlated with immune infiltration and ferroptosis in acute myeloid leukemia: a pan-cancer analysis. Transl Cancer Res. 2022;11(10):3491–3505. doi:10.21037/tcr-22-985
22. Kagan VE, Mao G, Qu F, et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat Chem Biol. 2017;13(1):81–90. doi:10.1038/nchembio.2238
23. Doll S, Freitas FP, Shah R, et al. FSP1 is a glutathione-independent ferroptosis suppressor. Nature. 2019;575(7784):693–698. doi:10.1038/s41586-019-1707-0
24. Bersuker K, Hendricks JM, Li Z, et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature. 2019;575(7784):688–692. doi:10.1038/s41586-019-1705-2
25. Sun Y, Berleth N, Wu W, et al. Fin56-induced ferroptosis is supported by autophagy-mediated GPX4 degradation and functions synergistically with mTOR inhibition to kill bladder cancer cells. Cell Death Dis. 2021;12(11):1028. doi:10.1038/s41419-021-04306-2
26. Bai Y, Meng L, Han L, et al. Lipid storage and lipophagy regulates ferroptosis. Biochem Biophys Res Commun. 2019;508(4):997–1003. doi:10.1016/j.bbrc.2018.12.039
27. Liu MR, Zhu WT, Pei DS. System Xc(-): a key regulatory target of ferroptosis in cancer. Invest New Drugs. 2021;39(4):1123–1131. doi:10.1007/s10637-021-01070-0
28. Yang W, Wang Y, Zhang C, et al. Maresin1 protect against ferroptosis-induced liver injury through ROS Inhibition and Nrf2/HO-1/GPX4 activation. Front Pharmacol. 2022;13:865689. doi:10.3389/fphar.2022.865689
29. Feng Z, Qin Y, Huo F, et al. NMN recruits GSH to enhance GPX4-mediated ferroptosis defense in UV irradiation induced skin injury. Biochim Biophys Acta Mol Basis Dis. 2022;1868(1):166287. doi:10.1016/j.bbadis.2021.166287
30. Forcina GC, Dixon SJ. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics. 2019;19(18):e1800311. doi:10.1002/pmic.201800311
31. Imai H, Matsuoka M, Kumagai T, Sakamoto T, Koumura T. Lipid peroxidation-dependent cell death regulated by GPx4 and ferroptosis. Curr Top Microbiol Immunol. 2017;403:143–170. doi:10.1007/82_2016_508
32. Dong H, Qiang Z, Chai D, et al. Nrf2 inhibits ferroptosis and protects against acute lung injury due to intestinal ischemia reperfusion via regulating SLC7A11 and HO-1. Aging. 2020;12(13):12943–12959. doi:10.18632/aging.103378
33. Maiorino M, Conrad M, Ursini F. GPx4, lipid peroxidation, and cell death: discoveries, rediscoveries, and open issues. Antioxid Redox Signal. 2018;29(1):61–74. doi:10.1089/ars.2017.7115
34. Louandre C, Marcq I, Bouhlal H, et al. The retinoblastoma (Rb) protein regulates ferroptosis induced by sorafenib in human hepatocellular carcinoma cells. Cancer Lett. 2015;356(2 Pt B):971–977. doi:10.1016/j.canlet.2014.11.014
35. Li Y, Zeng X, Lu D, Yin M, Shan M, Gao Y. Erastin induces ferroptosis via ferroportin-mediated iron accumulation in endometriosis. Hum Reprod. 2021;36(4):951–964. doi:10.1093/humrep/deaa363
36. Tang Z, Ju Y, Dai X, et al. HO-1-mediated ferroptosis as a target for protection against retinal pigment epithelium degeneration. Redox Biol. 2021;43:101971. doi:10.1016/j.redox.2021.101971
37. Zhang X, Yu K, Ma L, et al. Endogenous glutamate determines ferroptosis sensitivity via ADCY10-dependent YAP suppression in lung adenocarcinoma. Theranostics. 2021;11(12):5650–5674. doi:10.7150/thno.55482
38. Liu T, Jiang L, Tavana O, Gu W. The Deubiquitylase OTUB1 mediates ferroptosis via stabilization of SLC7A11. Cancer Res. 2019;79(8):1913–1924. doi:10.1158/0008-5472.CAN-18-3037
39. Mao C, Liu X, Zhang Y, et al. DHODH-mediated ferroptosis defence is a targetable vulnerability in cancer. Nature. 2021;593(7860):586–590. doi:10.1038/s41586-021-03539-7
40. Li S, He Y, Chen K, et al. RSL3 drives ferroptosis through NF-κB pathway activation and GPX4 depletion in glioblastoma. Oxid Med Cell Longev. 2021;2021:2915019. doi:10.1155/2021/2915019
41. Gai C, Yu M, Li Z, et al. Acetaminophen sensitizing erastin-induced ferroptosis via modulation of Nrf2/heme oxygenase-1 signaling pathway in non-small-cell lung cancer. J Cell Physiol. 2020;235(4):3329–3339. doi:10.1002/jcp.29221
42. Xu S, Wu B, Zhong B, et al. Naringenin alleviates myocardial ischemia/reperfusion injury by regulating the nuclear factor-erythroid factor 2-related factor 2 (Nrf2) /System xc-/ glutathione peroxidase 4 (GPX4) axis to inhibit ferroptosis. Bioengineered. 2021;12(2):10924–10934. doi:10.1080/21655979.2021.1995994
43. Gaschler MM, Andia AA, Liu H, et al. FINO(2) initiates ferroptosis through GPX4 inactivation and iron oxidation. Nat Chem Biol. 2018;14(5):507–515. doi:10.1038/s41589-018-0031-6
44. Shu X, Wu J, Zhang T, et al. Statin-induced geranylgeranyl pyrophosphate depletion promotes ferroptosis-related senescence in adipose tissue. Nutrients. 2022;14:20.
45. Wu W, Geng Z, Bai H, Liu T, Zhang B. Ammonium ferric citrate induced ferroptosis in non-small-cell lung carcinoma through the inhibition of GPX4-GSS/GSR-GGT axis activity. Int J Med Sci. 2021;18(8):1899–1909. doi:10.7150/ijms.54860
46. Eling N, Reuter L, Hazin J, Hamacher-Brady A, Brady NR. Identification of artesunate as a specific activator of ferroptosis in pancreatic cancer cells. Oncoscience. 2015;2(5):517–532. doi:10.18632/oncoscience.160
47. Cramer SL, Saha A, Liu J, et al. Systemic depletion of L-cyst(e)ine with cyst(e)inase increases reactive oxygen species and suppresses tumor growth. Nat Med. 2017;23(1):120–127. doi:10.1038/nm.4232
48. Zhou J, Zhang L, Wang M, et al. CPX Targeting DJ-1 Triggers ROS-induced cell death and protective autophagy in colorectal cancer. Theranostics. 2019;9(19):5577–5594. doi:10.7150/thno.34663
49. Zhong Y, Tian F, Ma H, et al. FTY720 induces ferroptosis and autophagy via PP2A/AMPK pathway in multiple myeloma cells. Life Sci. 2020;260:118077. doi:10.1016/j.lfs.2020.118077
50. Clemente LP, Rabenau M, Tang S, et al. Dynasore blocks ferroptosis through combined modulation of iron uptake and inhibition of mitochondrial respiration. Cells. 2020;9(10):2259. doi:10.3390/cells9102259
51. Xie Y, Song X, Sun X, et al. Identification of baicalein as a ferroptosis inhibitor by natural product library screening. Biochem Biophys Res Commun. 2016;473(4):775–780. doi:10.1016/j.bbrc.2016.03.052
52. Sun X, Ou Z, Xie M, et al. HSPB1 as a novel regulator of ferroptotic cancer cell death. Oncogene. 2015;34(45):5617–5625. doi:10.1038/onc.2015.32
53. Liu P, Feng Y, Li H, et al. Ferrostatin-1 alleviates lipopolysaccharide-induced acute lung injury via inhibiting ferroptosis. Cell Mol Biol Lett. 2020;25:10. doi:10.1186/s11658-020-00205-0
54. Rikhvanov EG, Fedoseeva IV, Varakina NN, Rusaleva TM, Fedyaeva AV. Mechanism of Saccharomyces cerevisiae yeast cell death induced by heat shock. Effect of cycloheximide on thermotolerance. Biochemistry. 2014;79(1):16–24. doi:10.1134/S0006297914010039
55. Baba Y, Higa JK, Shimada BK, et al. Protective effects of the mechanistic target of rapamycin against excess iron and ferroptosis in cardiomyocytes. Am J Physiol Heart Circ Physiol. 2018;314(3):H659–h668. doi:10.1152/ajpheart.00452.2017
56. Li Y, Yu P, Fu W, et al. Ginsenoside Rd inhibited ferroptosis to alleviate CCl(4)-induced acute liver injury in mice via cGAS/STING pathway. Am J Chin Med. 2023;51(1):91–105. doi:10.1142/S0192415X23500064
57. Meyer NJ, Gattinoni L, Calfee CS. Acute respiratory distress syndrome. Lancet. 2021;398(10300):622–637. doi:10.1016/S0140-6736(21)00439-6
58. Zaki A, Ali MS, Hadda V, Ali SM, Chopra A, Fatma T. Long non-coding RNA (lncRNA): a potential therapeutic target in acute lung injury. Genes Dis. 2022;9(5):1258–1268. doi:10.1016/j.gendis.2021.07.004
59. Bhargava M, Wendt CH. Biomarkers in acute lung injury. Transl Res. 2012;159(4):205–217. doi:10.1016/j.trsl.2012.01.007
60. Fujishima S. Pathophysiology and biomarkers of acute respiratory distress syndrome. J Intensive Care. 2014;2(1):32. doi:10.1186/2052-0492-2-32
61. Hussain M, Xu C, Ahmad M, et al. Acute respiratory distress syndrome: bench-to-bedside approaches to improve drug development. Clin Pharmacol Ther. 2018;104(3):484–494. doi:10.1002/cpt.1034
62. Ghio AJ, Carter JD, Richards JH, Richer LD, Grissom CK, Elstad MR. Iron and iron-related proteins in the lower respiratory tract of patients with acute respiratory distress syndrome. Crit Care Med. 2003;31(2):395–400. doi:10.1097/01.CCM.0000050284.35609.97
63. Xu Y, Li X, Cheng Y, Yang M, Wang R. Inhibition of ACSL4 attenuates ferroptotic damage after pulmonary ischemia-reperfusion. FASEB j. 2020;34(12):16262–16275. doi:10.1096/fj.202001758R
64. Guo XW, Zhang H, Huang JQ, et al. PIEZO1 ion channel mediates ionizing radiation-induced pulmonary endothelial cell ferroptosis via Ca(2+)/Calpain/VE-cadherin signaling. Front Mol Biosci. 2021;8:725274. doi:10.3389/fmolb.2021.725274
65. Liu X, Wang L, Xing Q, et al. Sevoflurane inhibits ferroptosis: a new mechanism to explain its protective role against lipopolysaccharide-induced acute lung injury. Life Sci. 2021;275:119391. doi:10.1016/j.lfs.2021.119391
66. Li J, Lu K, Sun F, et al. Panaxydol attenuates ferroptosis against LPS-induced acute lung injury in mice by Keap1-Nrf2/HO-1 pathway. J Transl Med. 2021;19(1):96. doi:10.1186/s12967-021-02745-1
67. Wang YM, Gong FC, Qi X, et al. Mucin 1 inhibits ferroptosis and sensitizes vitamin E to alleviate sepsis-induced acute lung injury through GSK3β/Keap1-Nrf2-GPX4 pathway. Oxid Med Cell Longev. 2022;2022:2405943. doi:10.1155/2022/2405943
68. Tang X, Liu J, Yao S, Zheng J, Gong X, Xiao B. Ferulic acid alleviates alveolar epithelial barrier dysfunction in sepsis-induced acute lung injury by activating the Nrf2/HO-1 pathway and inhibiting ferroptosis. Pharm Biol. 2022;60(1):2286–2294. doi:10.1080/13880209.2022.2147549
69. Li T, Tan Y, Ouyang S, He J, Liu L. Resveratrol protects against myocardial ischemia-reperfusion injury via attenuating ferroptosis. Gene. 2022;808:145968. doi:10.1016/j.gene.2021.145968
70. Yamada N, Karasawa T, Wakiya T, et al. Iron overload as a risk factor for hepatic ischemia-reperfusion injury in liver transplantation: potential role of ferroptosis. Am J Transplant. 2020;20(6):1606–1618. doi:10.1111/ajt.15773
71. Tuo QZ, Liu Y, Xiang Z, et al. Thrombin induces ACSL4-dependent ferroptosis during cerebral ischemia/reperfusion. Signal Transduct Target Ther. 2022;7(1):59. doi:10.1038/s41392-022-00917-z
72. Mura M, Andrade CF, Han B, et al. Intestinal ischemia-reperfusion-induced acute lung injury and oncotic cell death in multiple organs. Shock. 2007;28(2):227–238. doi:10.1097/01.shk.0000278497.47041.e3
73. Qiang Z, Dong H, Xia Y, Chai D, Hu R, Jiang H. Nrf2 and STAT3 alleviates ferroptosis-mediated IIR-ALI by Regulating SLC7A11. Oxid Med Cell Longev. 2020;2020:5146982. doi:10.1155/2020/5146982
74. Liu T, Yang Q, Zheng H, et al. Multifaceted roles of a bioengineered nanoreactor in repressing radiation-induced lung injury. Biomaterials. 2021;277:121103. doi:10.1016/j.biomaterials.2021.121103
75. Kang SK, Rabbani ZN, Folz RJ, et al. Overexpression of extracellular superoxide dismutase protects mice from radiation-induced lung injury. Int J Radiat Oncol Biol Phys. 2003;57(4):1056–1066. doi:10.1016/S0360-3016(03)01369-5
76. Yu JB, Shi J, Gong LR, et al. Role of Nrf2/ARE pathway in protective effect of electroacupuncture against endotoxic shock-induced acute lung injury in rabbits. PLoS One. 2014;9(8):e104924. doi:10.1371/journal.pone.0104924
77. Jin F, Li C. Seawater-drowning-induced acute lung injury: from molecular mechanisms to potential treatments. Exp Ther Med. 2017;13(6):2591–2598. doi:10.3892/etm.2017.4302
78. Topf JM, Murray PT. Hypomagnesemia and hypermagnesemia. Rev Endocr Metab Disord. 2003;4(2):195–206. doi:10.1023/A:1022950321817
79. Mishra P, Pandey R, Pandey N, Tripathi S, Tripathi YB. Prevention of mortality in acute lung injury induced by oleic acid: application of polyherbal decoction (bronco T). Front Cell Dev Biol. 2022;10:1003767. doi:10.3389/fcell.2022.1003767
80. de Oliveira Rodrigues S, Patricio de Almeida MA, Castro-Faria-Neto HC, Silva AR, Felippe gonçalves-de-albuquerque C. Mouse model of oleic acid-induced acute respiratory distress syndrome. J Vis Exp. 2022;2022:184.
81. Chen H, Ding Y, Chen W, Feng Y, Shi G. Glibenclamide alleviates inflammation in oleic acid model of acute lung injury through NLRP3 inflammasome signaling pathway. Drug Des Devel Ther. 2019;13:1545–1554. doi:10.2147/DDDT.S196040
82. Yin X, Zhu G, Wang Q, Fu YD, Wang J, Xu B. Ferroptosis, a new insight into acute lung injury. Front Pharmacol. 2021;12:709538. doi:10.3389/fphar.2021.709538
83. Liu X, Zhang J, Xie W. The role of ferroptosis in acute lung injury. Mol Cell Biochem. 2022;477(5):1453–1461. doi:10.1007/s11010-021-04327-7
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.