Back to Journals » Journal of Inflammation Research » Volume 19
Regulatory T Cells in Diabetic Wound Healing: A Comprehensive Review
Authors Lin J ![]() , Liu Y, Min P, Zhao H, Xia Z, Zheng Y
, Liu Y, Min P, Zhao H, Xia Z, Zheng Y
Received 14 January 2026
Accepted for publication 9 May 2026
Published 22 May 2026 Volume 2026:19 594673
DOI https://doi.org/10.2147/JIR.S594673
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Quan Zhang
Jiezhi Lin,1,2,* Yingying Liu,1,* Peiyao Min,1,* Hui Zhao,2 Zhaofan Xia,1 Yongjun Zheng1
1Department of Burn Surgery, The First Affiliated Hospital of Naval Medical University, Shanghai, 200433, People’s Republic of China; 2Department of Burn Surgery, The 963rd Hospital of Joint Logistics Support Force of PLA, Jiamusi, Heilongjiang, 154007, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yongjun Zheng, Department of Burn Surgery, The First Affiliated Hospital of Naval Medical University, 168th Changhai Road, Yangpu District, Shanghai, 200433, People’s Republic of China, Email [email protected] Hui Zhao, Email [email protected]
Abstract: Patients with diabetes frequently face challenges associated with impaired wound healing, which is a serious clinical concern. Regulatory T cells (Tregs) are essential immune modulators that are integral to wound healing. While research is increasingly examining alterations in Tregs and their potential impacts on diabetic wound healing, a comprehensive review is lacking. This review employs a narrative approach to summarize the current knowledge regarding the mechanisms by which Tregs influence wound healing in diabetes, focusing on their impact on inflammatory responses, cellular regeneration, and angiogenesis. Furthermore, we investigated the potential therapeutic applications of Tregs in diabetic wound healing, highlighting recent advances in biomaterials, molecular pathways, and regulatory strategies targeting Tregs. Despite challenges such as cell acquisition and quality control, as well as risks like immunosuppression and infection, advancements in clinical studies indicate that Tregs could be a promising new treatment for diabetes. This review aims to provide novel insights and directions by analyzing the most recent literature on future therapeutic strategies for managing diabetic wounds.
Keywords: tregs, diabetes, wound healing, immune modulation
Introduction
Diabetes mellitus, a chronic metabolic disorder characterized by persistent hyperglycemia, is associated with numerous complications, including impaired wound healing.1,2 This impairment is particularly evident in patients with diabetes, in whom the healing of skin injuries, such as diabetic foot ulcers (DFU), is frequently severely compromised.3,4 The immune system is integral to wound healing, and regulatory T cells (Tregs) are pivotal in modulating inflammation and facilitating tissue repair. Tregs, a specialized subset of T cells, are essential for maintaining immune homeostasis and preventing excessive inflammatory responses, both of which are critical for effective wound healing.
Recent research suggests that Tregs play a complex role in wound healing, with particular emphasis on their function in diabetic contexts. Tregs are instrumental in modulating immune responses by suppressing pro-inflammatory activities, while promoting angiogenesis and tissue regeneration by secreting various cytokines and growth factors.4–6 Additionally, Tregs play a pivotal role in orchestrating the recruitment and activation of other immune cells, such as macrophages and neutrophils, which are essential for effective wound healing.7 Tregs in the wound microenvironment are correlated with enhanced healing outcomes because they contribute to the regulation of the inflammatory response and facilitate the transition from the inflammatory phase to the proliferative phase of wound healing.8,9
Treg cell functionality and stability may be compromised in diabetic wounds. Hyperglycemia and chronic inflammation can lead to Treg dysfunction, characterized by reduced numbers and impaired suppressive capabilities. This dysregulation contributes to the persistent inflammatory state observed in diabetic wounds, thereby impeding healing.10,11 Understanding the mechanisms by which Tregs influence wound healing in diabetes is essential for developing targeted therapeutic strategies to enhance Treg function or restore their numbers. This review aimed to elucidate the role of Tregs in diabetes-related wound healing by exploring their mechanisms of action and potential therapeutic implications for improving outcomes in patients with diabetes. By addressing the interplay between Tregs and wound healing, we can gain a better understanding of how to mitigate the complications associated with diabetes and enhance healing in affected individuals.
Main Body
Basic Characteristics and Functions of Tregs
Tregs are a specialized subset of T lymphocytes that are pivotal in sustaining immune homeostasis and tolerance, thereby averting the onset of autoimmune diseases and mitigating excessive inflammatory responses.12 Tregs are distinguished by the expression of specific markers, notably CD4, CD25, and the transcription factor forkhead box p3 (Foxp3), which is indispensable for their development and functional activity.12,13 Beyond their fundamental role in maintaining immune tolerance, Tregs are actively engaged in modulating immune responses during infectious processes and in oncological contexts, where they may either suppress inflammation or facilitate tumor progression, contingent upon the prevailing microenvironmental conditions.14
Classification and Markers of Tregs
Tregs are classified into two types: thymus-derived (tTregs) and peripherally induced (pTregs) (Figure 1). tTregs develop in the thymus via a two-step process involving high-affinity self-antigen presentation and maturation aided by cytokines, including IL-2, IL-15, and transforming growth factor-beta (TGF-β).15 In contrast, pTregs are formed in peripheral lymphoid tissues in response to environmental antigens, with their differentiation influenced by TGF-β, retinoic acid, and microbial metabolites, such as short-chain fatty acids. Tregs exhibit phenotypic plasticity, enabling them to maintain immunosuppressive functions across diverse inflammatory environments. These cells express the signature transcription factor Foxp3,16 and co-express T-box transcription factor (T-bet), GATA-binding factor 3 (GATA-3), B-cell lymphoma 6 (BCL6), and retinoid-related orphan receptor γ (RORγt), which are crucial for T helper 1 (Th1), Th2, T follicular helper (Tfh), and Th17 cell differentiation.17 This co-expression enables Tregs to suppress specific subsets of overactive CD4+ Th cells, ensuring precise immune regulation. Modulating Foxp3 can alter Treg functions; for example, mitogen-activated protein kinase kinase kinase kinase 3 (MAP4K3) can reduce Foxp3 expression by phosphorylating inhibitor of nuclear factor kappa-B kinase subunit beta (IKKβ), thereby inhibiting Treg differentiation.18 Inhibiting Foxp3 using hydrocarbon-stapled alpha-helical peptide inhibitors effectively suppresses Treg function. Tregs express CD4 and CD25, as well as Foxp3, which aid in their differentiation and peripheral induction, crucial for maintaining tolerance and preventing autoimmune diseases.19 Tregs also express cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4) and programmed cell death protein-1, important for their regulatory roles.20 tTregs have a stable phenotype, while pTregs are less stable due to inflammatory, epigenetic, and metabolic influences.21 Inflammatory cytokines, such as IL-6, can destabilize Foxp3 expression, causing pTregs to lose Foxp3 and gain effector functions.22 The DNA methylation status of the Foxp3 gene, especially in conserved non-coding sequences 2 (CNS2), is crucial for its expression stability; hypomethylation supports expression, while hypermethylation can silence it.23 Treg markers vary over time and location, and their presence in tissues has been detected using flow cytometry and immunohistochemistry, which has aided research on conditions, such as autoimmune diseases and cancer.24
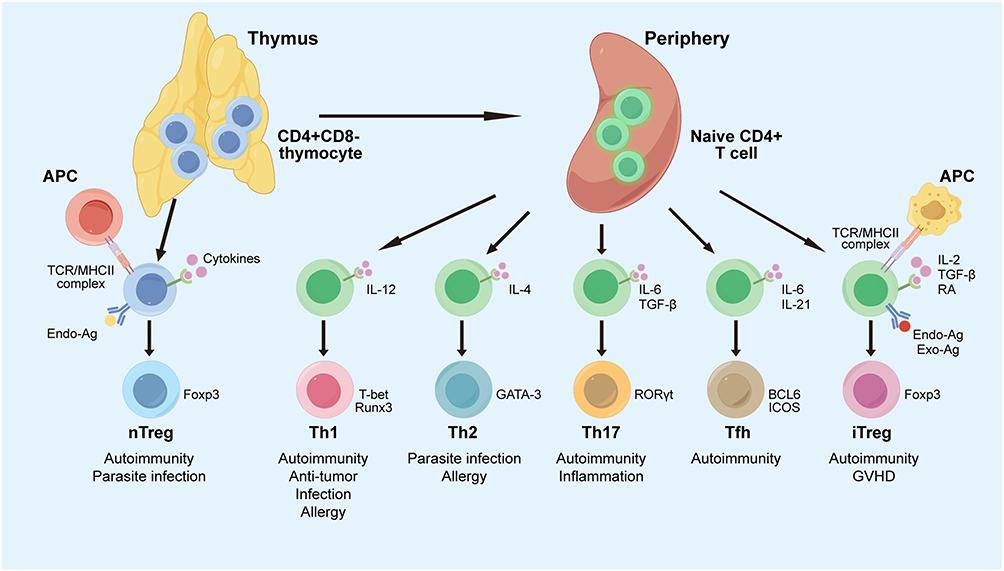 |
Figure 1 Classification, phenotypes, function, and characteristics of Tregs in the immune response. Naïve CD4+ T cells differentiate into Tregs or effector T cells. Naïve CD4+ T cells and nTregs are selected in the thymus. In peripheral tissues, naïve CD4+ T cells can differentiate into Th1, Th2, Th17, or iTregs, each with distinct immune functions. Cytokines regulate these differentiation pathways, requiring specific transcription factors: T-bet for Th1, GATA3 for Th2, RORγt for Th17, and Foxp3 for Tregs. Abbreviations: APC, antigen-presenting cell; BCL-6, B-cell lymphoma 6; CD, cluster of differentiation; Endo-Ag, endogenous antigen; Exo-Ag, exogenous antigen; Foxp3, Forkhead box p3; GATA-3, GATA-binding factor 3; GVHD, graft-versus-host disease; ICOS, inducible T Cell costimulator; IL, interleukin; iTreg, induced Treg; MHC, major histocompatibility complex; nTreg, natural Treg; RA, retinoic acid; RORγt, retinoid-related orphan receptor γ; Runx3, runt-related transcription factor 3; T-bet, T box transcription factor; TCR, T cell receptor; Tfh, follicular helper T cells; TGF-β, transforming growth factor-β; Th, T helper cell. |
Tregs in Immune Regulation
Tregs are crucial for immune balance by suppressing immune responses via CTLA-4-mediated inhibition of antigen-presenting cells (APC), IL-2 consumption, release of regulatory cytokines and metabolites, and direct suppression of effector cells.25
Tregs influence dendritic cells (DCs) via direct contact, cytokines, and exosomes.26 Tregs hinder the antigen-presenting ability of DC, primarily via the CTLA-4 receptor on Tregs.27 CTLA-4 signaling, especially in double negative (DN) Tregs, reduces CD80/CD86 on DCs.28 However, the exact mechanisms are complex and poorly understood. Studies in mice have revealed that blocking CTLA-4 negates the suppressive effects of Tregs in inflammatory bowel disease, autoimmune gastritis, and transplant rejection.29 Tregs also decrease nuclear factor kappa B (NF-κB)-related gene expression and increase wnt family member 5a (Wnt5a) in DCs, impairing their maturation and antigen presentation.30 Additionally, Tregs regulate DCs through cytotoxic mechanisms. DN Tregs can target and eliminate antigen-loaded DCs via the Fas-FasL pathway.28
Tregs suppress other immune cells and induce apoptosis by secreting anti-inflammatory cytokines, including IL-10, TGF-β,31 and IL-35, and producing cytolytic enzymes, including granzyme and perforin.32,33 They inhibit pro-inflammatory cytokine secretion by effector T cells, thereby preventing autoimmunity.34 Additionally, studies have demonstrated that Tregs can inhibit CD4+ T cell activation and proliferation via direct and indirect contact.35 They compete for IL-2 by expressing high levels of IL-2 receptors, thereby restricting its availability and limiting effector T cell expansion.36 When co-cultured with APC and antigen-specific responder T cells, Tregs suppress responder cell proliferation and decrease IL-2 production. Tregs also promote macrophage polarization toward an anti-inflammatory M2 phenotype for tissue repair.37 After co-culture with Tregs, monocytes exhibit M2 macrophage traits, such as increased CD206 and CD163 expression, and reduced pro-inflammatory responses, including lower IL-6 and TNF-β production and decreased NF-κB activation.38 In vitro-expanded Tregs promote a tolerogenic phenotype in monocytes, boost IL-10 production in macrophages by secreting IL-13, and activate the Vav1-Rac1 pathway to enhance phagocytosis and reduce inflammation.39
Neutrophil dysfunction may cause prolonged inflammation and autoimmune diseases.40 Tregs interact with neutrophils to reduce granulocyte infiltration by lowering chemokine expression. In vitro, Tregs co-cultured with neutrophils increase IL-10 and TGF-β1 production, which are linked to immunosuppression.41 Tregs also boost heme oxygenase-1 and indoleamine 2,3-dioxygenase expression, which play crucial roles in innate immune responses.41 According to Mathur et al, Treg cells enhance wound healing by guiding hair follicle stem cells (HFSCs) to become epithelial cells, reducing the release of the neutrophil chemoattractant C-X-C motif chemokine ligand 5 (CXCL5) by the injured epithelium, and preventing neutrophils from being drawn to the wound.42 Interestingly, research by Luan et al shows that migrating HFSCs activate immune regulatory programs, including CD80, MHCII, and CXCL5, which attract pTregs to the wound site. This reduces neutrophil infiltration and inflammation, thereby promoting healing. These findings appear to complement the research by Mathur et al, suggesting that the interaction between Tregs and HFSCs establishes a mutually reinforcing positive feedback loop, collectively sustaining the reduction of wound inflammation and promoting epithelialization.43
Tregs help resolve inflammation and tissue repair by modulating the immune response in chronic inflammatory and autoimmune diseases.44,45 However, they promote tumor growth by creating an immunosuppressive environment that hinders anti-tumor immunity.46 Tregs facilitate the progression of colorectal cancer, hepatocellular carcinoma, and triple-negative breast cancer (TNBC). In a liver tumor mouse model, changes in diacylglycerol O-acyltransferase 1 (DGAT1) and DGAT2 levels enhanced Treg recruitment and tumor progression.47 In TNBC, Tregs infiltrate the tumor microenvironment, promoting tumor growth and metastasis via immunosuppression.48 Thus, Treg cells act as a “double-edged sword”, with their balanced immunoregulatory functions crucially impacting immune disorders, such as tumors and autoimmune diseases.46 Understanding their dual role in tolerance and tumor growth is vital for developing therapies to manipulate their function, especially in cancer immunotherapy.
Pathophysiology of Diabetic Wounds
Diabetes mellitus impairs wound healing by disrupting hemostasis, inflammation, proliferation, and remodeling, leading to chronic wounds and an increased risk of infections and amputations.49 This is caused by altered cellular responses, impaired angiogenesis, and dysfunctional immune responses,50 with treatment-resistant biofilms further complicating the management of diabetic wounds.51,52
Diabetic wounds suffer from poor angiogenesis, neuropathy, and chronic inflammation due to high blood sugar levels, which cause the accumulation of advanced glycation end products (AGEs).50,53,54 These AGEs increase reactive oxygen species (ROS) and nitrogen species (RNS),55 which damage endothelial cells and reduce blood flow and oxygenation to wounds.56–58 Excessive ROS or RNS levels perpetuate tissue damage and inflammation.59 In diabetes, pathways, such as hexosamine60 and AGEs,61 elevate mitochondrial ROS production, causing oxidative stress and impaired antioxidant defenses, thereby delaying healing. High ROS levels disrupt collagen synthesis and re-epithelialization, and degrade the extracellular matrix and growth factors, exacerbating inflammation.62 Vascular defects cause hypoxia, increase ROS levels, and promote protein degradation, which damages the extracellular matrix and hinders wound healing.63 In the proliferation phase, fibroblasts in a hyperglycemic environment exhibit reduced growth and movement, resulting in less granulation tissue and slower wound contraction, delaying healing, and increasing complication risks.64 Oxidative stress also worsens neuropathy, and diabetic neuropathy can cause loss of sensation, increasing the risk of unnoticed injuries.65 The impact of ROS further extends to immune regulation. ROS negatively affects Treg proliferation and function. Research shows that oxidative stress from trauma increases the inhibitory protein guanine nucleotide-binding protein α subunit 3 (Gαi3) in Tregs, reducing key proliferation proteins and hindering Treg expansion.66 In chronic inflammation, ROS impairs Treg function, promoting inflammation and tissue damage.67 In diabetic wound healing, reducing ROS enhances Treg accumulation, fostering a regenerative immune environment and speeding up healing.68 Thus, removing excess ROS is crucial for restoring Treg recruitment and function, balancing inflammation and repair.
Hyperglycemia constricts capillaries near wounds, reducing nutrient and oxygen delivery. High glucose levels lower angiogenic factors, such as vascular endothelial growth factor (VEGF), and impair endothelial cells, resulting in poor blood flow.69–71 Glycosylated hemoglobin and altered red blood cell membranes exacerbate oxygen and nutrient deficiency, thereby disrupting macrophage function. This causes abnormal apoptosis of fibroblasts and keratinocytes, as well as excessive release of pro-inflammatory cytokines,72 which attracts more macrophages and perpetuates the inflammatory state of the wound.73,74 Macrophages in diabetic wounds produce excessive cytokines, including tumor necrosis factor-alpha (TNF-α) and interleukin-1 beta (IL-1β), delaying recovery.10,75 Chronic inflammation in patients with diabetes is characterized by excessive pro-inflammatory cytokine production, which hinders healing by prolonging the inflammatory response. This imbalance hampers pathogen clearance and tissue repair, complicating wound healing and increasing the risk of infection.76 Taken together, chronic inflammation and poor blood flow represent intertwined obstacles to diabetic wound healing. Functional Tregs, with their anti-inflammatory and immune-regulating abilities, help by reducing inflammation and converting macrophages from the inflammatory M1 type to the healing M2 type. They also promote angiogenesis, improving the microenvironment for tissue regeneration, thus potentially tackling the prolonged inflammation and impaired angiogenesis in diabetic wounds.
The wound’s pH plays a crucial role in wound healing, especially in diabetic wounds, by affecting cellular functions.77 Normal skin pH is slightly acidic, whereas chronic wounds become more alkaline due to microvascular leakage, leading to increased inflammation, delayed healing, and a higher risk of infection.78,79 Significant advancements have been achieved in developing pH-responsive hydrogels and controlled-release nanoplatforms tailored to the alkaline microenvironment of diabetic wounds.77,80 These materials help regulate wound pH and enhance Treg recruitment to accelerate healing, but the causal relationship between these effects has not been explored in these studies. Given that the recruitment, stability, and suppressive function of Tregs are known to be highly sensitive to metabolic cues within their microenvironment,81 the pH-driven alteration of the wound immune landscape presents a plausible yet unexplored regulatory axis. However, no studies have demonstrated an interaction between wound pH and Tregs, indicating a need for further research.
Tregs in Wound Healing
Chronic wounds, such as diabetic ulcers, are primarily characterized by persistent, excessive, and uncontrolled chronic inflammation, angiogenesis disorders, and consequent tissue hypoxia, nutrient deficiency, oxidative stress, and proliferation and migration disorders of skin-related cells.1,3 These multifaceted factors contribute to the recalcitrance of chronic wounds. Immunological disorders and chronic inflammation are driving factors that impede diabetic wound healing.74,82 However, chronic wound management struggles with effectively addressing the disordered immune microenvironment, as traditional treatments like surgery, dressings, and negative pressure suction mainly focus on infection control, blood supply restoration and tissue maintenance.83,84 This limitation leads to ongoing inflammation and non-healing wounds.65,82 In contrast, targeting Tregs, which regulate the immune environment, appears promising in addressing pathogenic factors in chronic wounds and seems to offer a comprehensive therapeutic advantage for diabetic wounds.
Although previous research has demonstrated that Tregs play a beneficial role in wound healing, the chemotactic ability and function of Tregs decline significantly in patients with diabetes and in murine wound models of diabetes.85–88 Depletion of Treg cells results in delayed re-epithelialization, reduced wound contraction, and impaired vascular maturation,89 but the precise mechanisms are unclear. Therefore, a more comprehensive understanding of the role of Tregs in wound healing can provide a robust therapeutic basis and theoretical foundation for treating chronic wounds, particularly those associated with diabetes. Tregs primarily facilitate wound healing by modulating inflammation, promoting cell regeneration and tissue remodeling, and enhancing angiogenesis (Figure 2).
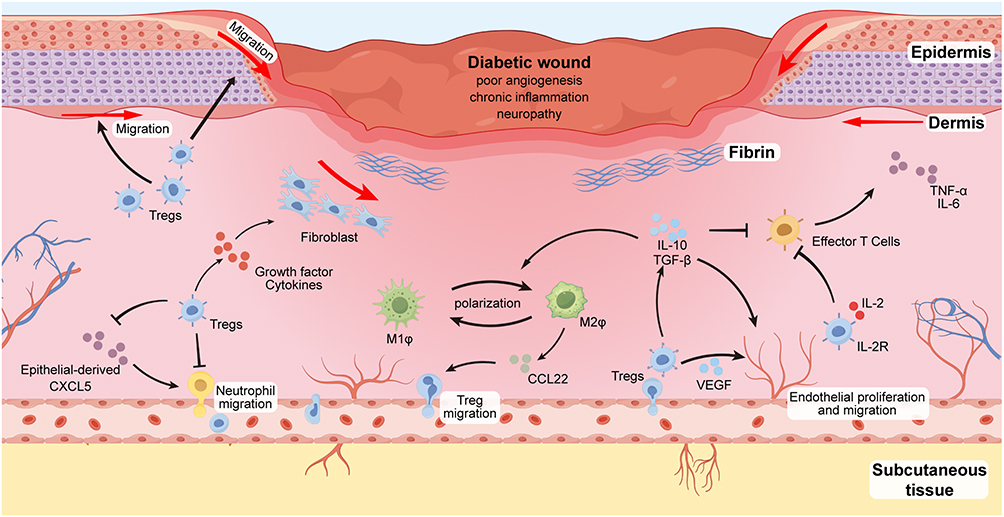 |
Figure 2 Regulatory function of Tregs in diabetic wound healing. Recruited Tregs in the wound area facilitate wound healing via various mechanisms. They exert anti-inflammatory effects, enhance angiogenesis, and stimulate cell proliferation and migration by secreting cytokines and growth factors. Additionally, Tregs modulate the immune microenvironment of diabetic wounds by directly or indirectly influencing the function of other immune cells. Abbreviations: CXCL5, C-X-C motif chemokine ligand 5; CCL22, C-C motif chemokine ligand 22; IL-2R, interleukin 2 receptor; M1φ, M1 macrophages; M2φ, M2 macrophages; VEGF, vascular endothelial growth factor. |
Inhibition of Inflammatory Responses
Tregs play a pivotal role in facilitating wound healing in patients with diabetes by mitigating excessive inflammatory responses.90 In diabetes, Tregs engage with various immune cells, a process that is critical for modulating the immune response and promoting wound repair.44 In non-obese diabetic (NOD) mice, C-C motif chemokine ligand 22 (CCL22) expression by β cells attracts Tregs and other immune cells, thereby establishing a tolerogenic milieu that safeguards β cells.91 In hyperglycemic murine models, Tregs accumulate in the liver and interact with other immune cells to regulate diabetes-associated inflammation, with C-C motif chemokine receptor 2 (CCR2) facilitating their migration.92 The extant literature indicates that Tregs primarily exert their regulatory influence on wound inflammation by modulating effector T cell activity, promoting polarization of macrophages towards the M2 phenotype, and inhibiting neutrophil recruitment.
Tregs produce anti-inflammatory cytokines, including IL-10 and TGF-β, which inhibit effector T cells. In type 1 diabetes, this suppressive mechanism aids in safeguarding pancreatic β cells from autoreactive T cell-mediated destruction and facilitates the re-establishment of normal wound healing processes.10,24 Excessive activation of effector T cells intensifies the inflammatory response, thereby impeding the healing of chronic wounds. In an experimental autoimmune encephalomyelitis model, the activation of effector CD4+ T cells, particularly the Th1 and Th17 subsets, has been demonstrated to enhance inflammation and tissue damage.93 This study suggests that mitigating effector T cell activation can reduce inflammation by modulating TGF-β signaling. Effector T cells sustain chronic inflammation by secreting pro-inflammatory factors, including IL-17 and interferon gamma (IFN-γ), which hinder wound healing processes, such as re-epithelialization and angiogenesis.94 In sepsis research, changes in lymphocyte subsets, including effector T cells, are linked to the prognosis.95 This underscores the dual role of effector T cells in inflammation regulation: moderate activation aids in pathogen elimination, while prolonged activation can cause immune dysregulation. In wound healing, Treg cells modulate inflammation by inhibiting effector T cells, as demonstrated in a mouse model.89 Depleting Treg cells increases IL-4 levels and the number of T-box transcription factor 21 (TBX21)+, RORγt+, and GATA-3+ αβ T cells, as well as IL-17A- and IFN-γ-producing T cells. The imbalance between effector T cells and Tregs is a critical determinant of wound healing outcomes, as effector T cells can resist Treg inhibition in chronic inflammation, hindering immune regulation and delaying healing.96
M2 macrophages are crucial for resolving wound inflammation and repairing tissue damage.31,97,98 Various interventions can facilitate the shift from pro-inflammatory M1 to anti-inflammatory M2 macrophages, thereby improving the inflammatory environment in chronic wounds, particularly diabetic wounds, and accelerating healing. This is widely recognized in the academic literature. Recent developments include lipid nanocomplexes with siRNA (cLpT@siRNA),99 sulfated glycosaminoglycan-based hydrogels,100 engineered exosomes,97,98 and activation of specific signaling pathways, such as the IL-27/IL-27Rα–STAT3 signaling axis,101 and dragon fruit stem polysaccharide/phosphoinositide-3-kinase (PI3K)/Akt pathway.102 These significantly enhanced the M1-to-M2 transition, reduced inflammation, promoted angiogenesis and collagen deposition, and accelerated diabetic wound healing. Previous research has presented that Tregs enhance M2 macrophage polarization at wound sites, thereby aiding healing. Nosbaum et al found that in a mouse skin defect model, highly activated Tregs accumulated in the skin, peaking at 7 days post-injury.103 These cells migrate from secondary lymphoid organs to wounds. Their removal increased IFN-γ-producing T cells and pro-inflammatory lymphocyte antigen 6 complex expression in macrophages, leading to reduced re-epithelialization, impaired wound closure, and increased granulation tissue and eschar formation. Nossbaum et al suggested that pro-inflammatory macrophages persist during wound closure, causing delayed healing. Mechanistically, Tregs produce cytokines, such as IL-10 and TGF-β, which promote M2 macrophage polarization, reduce inflammation, and repair tissues. Building on the interaction between Tregs and macrophages, Shen et al developed a sulfated chitosan (SCS) material to enhance M2 polarization via the IL-4/STAT6-PPARγ pathway and attract Tregs via CCL22, creating a “macrophage-Treg circuit” for sustained inflammation relief in diabetic wounds.7
Tregs are crucial in controlling inflammation by managing neutrophil activity at wound sites. Neutrophils, key players in the innate immune system, aid in acute wound healing by clearing pathogens and debris. However, in chronic wounds, such as DFU, dysfunction causes ongoing inflammation and tissue damage. The interaction between neutrophils and other immune cells contributes to persistent inflammation and impaired healing, which are central to chronic wound pathogenesis.83 Neutrophils exacerbate tissue damage and delay healing by releasing proteases and ROS. Specifically, neutrophils influence angiogenesis and inflammation via chemokines, including neutrophil-activating peptide-2 and IL-8, thereby affecting wound healing by altering chemokine levels. Increasing pro-angiogenic CXC chemokines and reducing neutrophil-related ones, such as interferon-γ inducible protein 10 and platelet factor-4, can shift wounds from chronic inflammation to healing.104 Moreover, neutrophil enzymes, such as elastase, can worsen tissue fibrosis, hindering repair in chronic wounds and lymphedema.105 Additionally, neutrophils can prolong inflammation by forming neutrophil extracellular traps (NETs), which are important for defense but can impede tissue healing if dysregulated.106 In neutrophilic asthma, NET accumulation is linked to hormone resistance and disease severity, indicating that excessive neutrophil activation may sustain inflammation and hinder tissue healing.107 Regulating neutrophil function is essential to reverse chronic wound inflammation. Studies suggest that Tregs play a crucial role in controlling neutrophil recruitment and activation. Mathur et al demonstrated that Treg depletion impairs skin barrier repair, triggers a Th17 response, and disrupts HFSC differentiation. Damaged epithelial cells express CXCL5, which attracts neutrophils; however, blocking CXCL5 or depleting neutrophils restores barrier function and stem cell differentiation.42 The regulation of Tregs on neutrophil chemotaxis and local inflammation aids skin stem cell differentiation and skin barrier regeneration, which are crucial for maintaining and repairing barrier tissues. Lui et al confirmed the positive role of Tregs in wound healing by regulating neutrophils, demonstrating that skin-resident Tregs are vital for skin immune homeostasis and repair.108 In a mouse wound model, they found that knocking out Jagged-1 in Tregs reduced lymphocyte antigen 6 family member G (Ly6G)+ neutrophil accumulation, suggesting that Tregs use the Jagged-1-Notch pathway to regulate neutrophil recruitment during injury, thereby maintaining skin immune balance and repair. Furthermore, a study found that using conditioned medium (CM) from low-oxygen-cultured amniotic-derived mesenchymal stem cells accelerated wound healing in mice with diabetes.109 This medium enhanced Treg chemotaxis, reduced local inflammation and chemokine gene expression (IL-1β, IL-6, CXCL1, and CXCL2), decreased neutrophil infiltration, and promoted capillary formation and epithelialization. These findings suggest that Tregs may facilitate wound healing in diabetes by reducing neutrophil activity and inflammation.
Promotion of Cellular Regeneration and Reconstruction
In addition to their role in modulating inflammation, Tregs promote cellular regeneration and tissue reconstruction during wound healing. They aid in stem cell differentiation for tissue repair and, in diabetic models, are linked to higher epithelial growth factors and keratinocyte proliferation, which are essential for wound re-epithelialization.21 Loretelli et al found that applying embryonic stem cell extracts (EXTs) to diabetic mouse wounds increased local Tregs and proliferating cells (Ki-67-positive) and reduced leukocyte infiltration (CD45+ cells).21 This highlights the importance of Tregs in creating a supportive environment for effective wound healing. Dongsoo et al cultured CD4+CD25+ Tregs from peripheral blood mononuclear cells (PBMCs) to obtain CM, which enhanced HaCaT keratinocyte migration and downregulated E-cadherin at wound edges.110 This CM also increased the levels of epithelial-mesenchymal transition (EMT)-related proteins, such as matrix metallopeptidase 1 (MMP-1), suggesting that Tregs aid in wound healing by promoting epithelial cell repair via cytokines and growth factors in CM. Trauma can trigger Tregs to express growth factors. In zebrafish, Tregs aid retinal repair by secreting insulin like growth factor 1 (IGF-1).111 Similarly, after lung injury, they express keratinocyte growth factor (KGF) to promote alveolar regeneration.112 Dendritic epidermal T cells also release IGF-1 and KGF to support skin regeneration. This suggests that Tregs may use these peptides to heal diabetic skin wounds, although this has not yet been confirmed. Additionally, Tregs aid in the differentiation of HFSCs for tissue repair,42 supporting hair regeneration and transforming into epithelial cells that migrate to the interfollicular epidermis, thereby enhancing skin wound healing. HFSCs help prevent infection and water loss by restoring the skin barriers. Tregs also protect HFSCs from damage by inflammatory neutrophils in wounds by inhibiting the CXCL5-IL-17 axis, further facilitating wound repair.
Furthermore, Tregs release growth factors and cytokines that promote fibroblast proliferation and migration, thereby aiding wound healing via collagen deposition and tissue remodeling.44,45,113,114 Umbilical cord blood Treg-derived exosomes enhance fibroblast migration, thereby accelerating wound closure.44 Tregs also secrete TGF-β, which has both anti-inflammatory and pro-fibrotic effects.115 Some studies suggest that Tregs help suppress scar formation, although opinions vary. Researchers have differing views on the role of Tregs in scar formation. Some evidence suggests that Tregs help prevent skin scarring.116,117 Kalekar et al found that removing Tregs in mice led to a five-fold increase in alpha smooth muscle actin (α-SMA)-expressing myofibroblasts and higher pro-fibrotic gene expression within 5 days. Chronic reduction in Tregs also results in increased numbers of fibrogenic myofibroblasts and increased pro-fibrotic gene activity.116 Another study revealed that co-culturing scar fibroblasts with Tregs increased IL-6 expression but reduced type I collagen and TGF-β mRNA levels.117 These findings indicate that Tregs play a protective and anti-fibrotic role in scar development. Nevertheless, some studies contradict this conclusion, indicating that scar tissue contains more Tregs and higher levels of Treg-related pro-fibrotic cytokines, including IL-10 and TGF-β1, than normal skin and peripheral blood.118,119 Additionally, Foxp3 expression in scar tissue is positively linked to type III collagen levels and the type III/type I collagen ratio. However, the causal link between Treg dysregulation and excessive collagen production remains unclear. Further research is needed to elucidate the relationship between Tregs, fibroblasts, and scars. Temporal and spatial variations in Treg numbers and activity may influence their roles in scar formation. Additionally, the role of Tregs in diabetic wound-related scar hyperplasia is underexplored and warrants investigation.
Promotion of Angiogenesis
Angiogenesis, new blood vessel formation, is crucial for wound healing, and Tregs significantly aid this process.120,121 They release factors, such as VEGF and IL-10, which encourage endothelial cell growth and movement, forming new capillaries.24 Tregs enhance neovascularization, improving blood supply and nutrient delivery to healing tissues.44,122 A study found that patients with diabetes have lower Treg levels in ischemic tissues than healthy individuals.123 CD4+Foxp3+ Tregs aid in vascular regeneration and promote new blood vessel formation in diabetic mice after ischemic injury. Reduced Tregs in chronic wound models are linked to impaired angiogenesis, underscoring their importance in vascular integrity and wound healing.6,124 Haertel et al found that Treg depletion increases IL-4 levels and expands αβT cells with Th1 and Th2 traits, hindering endothelial cell formation in wounds.89 This results in delayed re-epithelialization, reduced wound contraction, and impaired vascular maturation due to compromised epithelial cell proliferation and migration. Additionally, Nossbaum et al found that Tregs in inflamed skin significantly express epidermal growth factor receptor (EGFR),103 which is crucial for wound healing by aiding re-epithelialization, keratinocyte proliferation, and angiogenesis.125–127 In mice, Tregs can produce amphiregulin (AREG), an EGFR ligand, when stimulated by IL-18 and IL-33, promoting keratinocyte proliferation and restoring vascular integrity by activating TGF-β and AREG in mesenchymal stromal cells.128,129 These findings suggest that the AREG-EGFR pathway may enhance new blood vessel repair.130 Tregs also influence the extracellular matrix, which is crucial for stabilizing new vessels.89 Tregs help heal wounds and prevent chronic wound development by promoting angiogenesis.131
In summary, current research indicates that Tregs play a crucial role in wound healing by suppressing inflammation, aiding tissue regeneration, and promoting angiogenesis. However, evidence of their role in diabetic wound healing is limited, highlighting the need for further investigation to assess their potential as a therapeutic target.
Advancements in Studying Tregs in Diabetic Wound Healing
Tregs are key in managing immune responses during wound healing, especially in diabetes. They modulate inflammation and aid tissue repair, making them central to research on improving diabetic wound healing. Understanding Tregs can lead to new therapies for better healing in patients with diabetes.
Alterations in Tregs in Diabetes
Diabetes significantly affects immune cells, particularly Tregs, which are vital for immune balance and inflammation control.132 In type 1 diabetes mellitus (T1DM), Tregs in the blood remain largely unchanged,133–135 but their function, phenotype, and stability are compromised, leading to autoimmune tolerance failure and β cell destruction. In T1DM, disordered Treg function, especially CD8+ Tregs,86,136 contributes to disease progression and immune imbalance.87 Moreover, T1DM patients’ Tregs exhibit reduced effector T cell inhibition due to lower Foxp3 and CD39 expression, impacting their anti-inflammatory role.133 Additionally, pancreatic Tregs are impaired in T1DM onset, influenced by genetic/epigenetic factors, abnormal cytokine signaling, and the local metabolic environment.137 Moreover, in NOD mice, Tregs lose their suppressor functions with age due to impaired gap junction communication and decreased connexin 43 expression.88 This change may contribute to the development of diabetes and its complications. These variations in Treg functionality by demographics and age underscore the need to understand their influence on diabetes mellitus. Further research has demonstrated that the temporary depletion of Foxp3+ Tregs quickly leads to insulitis and hyperglycemia.138 Deepika Watts et al inserted the Foxp3BAC-DTR/GFP gene into autoimmune diabetes-prone NOD mice. They administered two diphtheria toxin (DT) injections, which eliminated more than 98% of the Tregs in the blood. DT treatment caused significant pancreatic damage, marked by CD3+ T-cell infiltration and islet disintegration, which worsened β-cell autoimmunity. The study observed increased Th1 cytokine levels of TNF-α and IFN-γ in the pancreas, as well as elevated levels of autoimmune molecules in the lymph nodes and pancreas. Over 50% of female mice developed diabetes within 5 days post-DT, suggesting that Treg cell removal accelerates β-cell autoimmunity and underscores their critical inhibitory role in diabetes development.
In type 2 diabetes mellitus (T2DM), unlike type 1, Tregs significantly change in quantity, linked to chronic inflammation, metabolic control, and disease complications. A systematic review and meta-analysis found that the percentage of CD4+CD25+Foxp3+ Tregs is notably lower in T2DM patients.139 In T2DM patients with diabetic nephropathy, peripheral blood Tregs are significantly lower and inversely related to the urinary albumin-creatinine ratio (UACR).140 Blood glucose variability is associated with reduced Tregs and may lead to nephropathy.141 T2DM patients also exhibit a pro-inflammatory shift with a higher Th17/Treg ratio due to increased Th17 levels and/or reduced Tregs.142,143 Similar to type 1 diabetes, Tregs in T2DM show impaired immunosuppressive function, linked to decreased Foxp3 expression and impaired IL-2 signaling, indicated by lower STAT5 phosphorylation.144,145
However, there are differing views on Treg changes in diabetes. In streptozotocin (STZ)-induced diabetic mice, CD4+CD25+Foxp3+ Tregs in peripheral tissues like the spleen and lymph nodes increased months after disease onset.146 However, these Tregs exhibit impaired immune suppression and display an activated/memory phenotype, possibly due to high sugar-induced oxidative stress and mitochondrial dysfunction. High sugar also enhances sp1 transcription factor (SP1) glycosylation, inhibits Na+/Ca2+/Li+ exchanger (NCLX) expression, and leads to mitochondrial calcium overload and oxidative damage in Tregs, thereby impairing their function.147 The literature on Treg number changes in diabetes is inconsistent, likely due to variations in diabetes subtypes, disease progression, species differences, and sample types. Nonetheless, all studies agree that Treg function is significantly impaired under diabetic conditions, regardless of blood sugar fluctuations.
High glucose levels make T cells more pro-inflammatory, disrupting the balance between pro- and anti-inflammatory responses.85 Although research is limited, some findings indicate an increased Th17/Treg cell ratio in diabetic patients. In diabetes, cytokines like IL-6, IL-21, and IL-23 impair Treg differentiation and migration while promoting pro-inflammatory Th17 cell differentiation and tissue infiltration, worsening inflammation and healing.143,148,149 IL-21, for instance, increases RORγt expression via the STAT3 pathway and decreases Foxp3 expression, enhancing Th17 differentiation and reducing Treg function.150 The balance between the STAT3 and STAT5 pathways is crucial for Th17/Treg outcomes, with STAT3 favoring Th17 and STAT5 supporting Tregs.151
Besides changes in circulating Tregs, diabetic wounds showed dysfunctional Tregs that failed to regulate effector T cell responses. Studies have revealed that Treg populations are often reduced and functionally impaired in diabetic wounds, leading to delayed recovery and a higher risk of infection.152–154 Focusing on the alterations of Tregs within human diabetic wounds, recent clinical and molecular studies provide a convergent evidence base. These studies highlight changes in Tregs in diabetic wounds. Zhang et al showed that maggot therapy increased Treg levels, improving healing in diabetic foot ulcers.155 Shi et al identified glutathione s-transferase mu 5 (GSTM5) as a key gene in disrupted immune pathways, providing genomic insight into impaired Treg function.156 Most recently, the high-resolution single-cell RNA sequencing study by Gan et al of human DFU granulation tissue definitively mapped the dysfunctional immune and fibroblast landscape, situating Treg alterations within a complex pathological cellular microenvironment.82 They revealed a higher abundance of monocytes/macrophages than in non-DFU tissues, which had more T cells and fibroblasts. DFU tissues exhibited increased expression of pro-inflammatory cytokines and chemokines, while non-DFU tissues had more repair-related processes. Effector T cells were more prevalent in DFU tissues, whereas Tregs were reduced, potentially impairing healing. These findings emphasize the increased inflammation in DFU granulation tissue and propose a therapeutic role for Tregs. The research journey from clinical observations to detailed cellular analysis confirms that Treg populations in diabetic wounds are dynamic and significantly altered, highlighting immune dysregulation’s role in impaired healing. However, direct clinical evidence connecting Treg dynamics to healing in diabetic wounds remains limited. More research is needed to validate these findings in larger patient groups, explore Treg population changes over time, and evaluate therapies to boost Treg function or numbers in diabetic wound care.
In summary, Treg dysfunction in diabetes may manifest as reduced numbers (eg, decreased frequency in high-risk T2DM patients),145 impaired function (eg, reduced IL-2 reactivity),144 or compromised differentiation and stability (eg, miR-142-3p affecting Treg development via ten-eleven translocation 2 (Tet2)).85 These issues likely stem from an “immune-metabolic vicious cycle”, in which immune dysregulation can worsen metabolic disorders, and high glucose levels can damage Tregs,138 diminishing their immunomodulatory capacity and exacerbating inflammation.147 It is noteworthy that research on Tregs in diabetes shows both consensus and disagreement, largely due to methodological differences. First, the choice of disease model significantly influences findings: studies in NOD mice highlight Tregs’ role in preventing autoimmune disease,88,138 while STZ models focus on Treg dysfunction due to high glucose.146,147 However, findings from NOD mice, such as Foxp3+ Treg depletion, do not fully explain human T1DM pathogenesis, whereas the STZ model’s insights into mitochondrial calcium overload may better apply to Treg metabolic dysfunction in high glucose, rather than autoimmune aspects. Second, there is a notable generation gap in research technology. Many studies still use traditional flow cytometry for phenotypic and functional analysis,133,140,142,144 despite its limitations, such as inconsistent definitions (eg, the inclusion of CD127low and CD39)133 and its limited capacity for causal inference, which is predominantly based on cross-sectional correlation studies. Meanwhile, advanced techniques like single-cell sequencing and live-cell imaging offer deeper insights but are expensive, complex, and not well integrated with animal models that mimic complex clinical scenarios.82,147 Third, in clinical studies, while it is agreed that Tregs are abnormal in diabetic patients, research groups differ on their causal role in diabetes. This is partly due to the heterogeneity of study populations. Some studies examine Tregs in T1DM within an autoimmune context,133,134 while most focus on T2DM and its associated metabolic inflammation.139–145 Even within T2DM studies, variations exist, such as differences in disease duration and the presence of complications like kidney disease or chronic obstructive pulmonary disease (COPD),141,143 which can affect immune status and potentially confound the observed Th17/Treg imbalance. Some studies highlight that the age of T1DM onset may influence Treg characteristics.134 However, many studies fail to match or detail patients’ glucose-lowering treatments, glycemic control levels, and other metabolic syndrome components, which can significantly impact immune cell function and lead to inconsistent results.147 Additionally, most research uses PBMCs, but it is unclear whether these accurately reflect the immune status in diabetic wounds. Single-cell studies suggest that the local wound environment has a distinct immune cell composition,82 indicating that peripheral blood tests might overlook crucial pathological details. Finally, most clinical studies on diabetes are cross-sectional, comparing patients with healthy controls at a single time point, which only shows associations rather than causality.133,134,140,141,143–145 Few studies have explored longitudinal effects like glycemic fluctuations, often with small sample sizes.141 Two key systematic reviews and meta-analyses compile multiple studies to offer stronger evidence, such as a decrease in Treg frequency and Th17/Treg imbalance in T2DM.139,142
Consequently, addressing the aforementioned challenges necessitates collaborative efforts in three pivotal areas to advance clinical research breakthroughs: 1) standardizing research methods and developing comprehensive models, such as those for diabetes and wound ischemia; 2) using a systems biology approach in mechanistic studies to integrate immune cells, metabolism, microbiota, and communication networks; and 3) designing longitudinal cohort studies to track Treg changes and disease progression, with detailed analyses of metabolic parameters, treatments, and complications to manage confounding factors.
Molecular Pathways Associated with Tregs
TGF-β is a versatile cytokine involved in development, wound healing, and immune regulation. Its dysfunction can cause inflammation, characterized by the ongoing release of pro-inflammatory cytokines. Tregs mediate immunosuppression during wound healing by activating TGF-β, secreting IL-10, and generating adenosine.130 Tissue-resident Tregs express AREG, which aids healing by promoting the release of bioactive TGF-β and promoting pericyte differentiation into myofibroblasts via the phospholipase C (PLC) signaling pathway. AREG also boosts the immunosuppressive function of Tregs, possibly via integrin activation.157 Vaz Emília et al used phage display technology to generate peptides that mimic TGF-β1 (the pm26TGF-β1 peptide). The pm26TGF-β1 peptide effectively downregulates TNF-α and upregulates IL-10 responses in PBMCs, thereby promoting the differentiation of Tregs.158 This finding suggests that the TGF-β pathway can be strategically targeted to enhance Treg-mediated wound healing, but its reliance on an ex vivo PBMC model may not reflect the complex tissue environment of diabetic wounds, where cellular interactions and metabolic stresses could alter this response. In contrast, in vivo models show that Tregs activate TGF-β, affecting keratinocytes and influencing wound healing. Joshua M. Moreau et al discovered that Tregs activate TGF-β via integrin αvβ8 to boost inflammation when the skin barrier is compromised.159 This activation affects nearby keratinocytes, not the Tregs themselves. Blocking TGF-β signaling speeds up skin repair and decreases neutrophil recruitment. While TGF-β activation aids in defending against Staphylococcus aureus by promoting inflammation and neutrophil recruitment, it also delays epithelial repair. This challenges the traditional view and highlights the complex role of Tregs in balancing inflammation and regeneration after skin injury. Moreau et al’s use of cell-specific knockout models provides strong evidence, unlike correlative in vitro studies.
Transitioning to another key pathway, the EGFR pathway also plays a crucial role in Treg-mediated healing, particularly during the transition from inflammation to proliferation in wounded skin.103 Nosbaum et al found that Tregs peaked at 20 times their baseline density on day 7 after skin injury, coinciding with a shift from the inflammatory to proliferative phase. These Tregs, recruited from secondary lymphoid organs, express high levels of CD25, CTLA-4, and inducible T cell costimulator (ICOS). Depleting Tregs delayed wound healing and increased granulation tissue and eschar. Early post-injury depletion increased the number of IFN-γ-producing T cells and pro-inflammatory macrophages. Tregs expressed high EGFR during the inflammatory phase, but not in draining lymph nodes. EGFR deletion delays wound healing, reduces Treg skin infiltration, and increases pro-inflammatory macrophages.103 Additionally, AREG, an EGFR ligand, may significantly influence Treg-mediated diabetic wound healing. Studies have indicated that AREG autocrine activation of EGFR helps T-cells preferentially express IL-33R, T1/ST2, allowing IL-33 to trigger the MAPK signaling pathway via EGFR, potentially promoting the proliferation of tissue-resident Tregs.130,160 These observations indicate that the EGFR pathway is crucial for Treg-mediated facilitation of wound healing, and targeting this pathway may offer a promising therapeutic strategy to enhance wound healing in patients with diabetes. However, a significant gap exists: most AREG evidence in Tregs comes from cancer or systemic immune models, leaving its role in diabetic wounds’ hypoxic, hyperglycemic, and inflamed environments largely unexplored, highlighting a crucial area for future research.
In summary, there is consensus that Tregs utilize TGF-β and EGFR pathways in wound healing, with EGFR signaling consistently linked to pro-reparative Treg accumulation. However, the main disagreement involves TGF-β’s role: it can either promote healing through immunosuppression and differentiation or hinder it by prolonging inflammation, depending on the cellular context and timing. Peptide interventions and ex vivo studies offer therapeutic insights but lack environmental complexity,158 while genetic in vivo models provide clarity but may oversimplify human disease.103,159 Future research should focus on in vivo diabetic models with temporal control of these pathways in specific cells, along with human studies, to resolve these issues and identify key therapeutic targets in the Treg signaling network for diabetic wound healing.
Tregs-Related Materials
Recent advancements in Treg-related materials display promise for diabetic wound healing (Table 1). Treg-derived exosomes enhance diabetic wound healing by modulating inflammation and tissue regeneration. Exosomes from cord blood Tregs (CB-Treg-Exos) target monocytes, promoting their differentiation into M2 macrophages and improving fibroblast and endothelial cell migration.44 Researchers successfully isolated CB-Treg-Exos and demonstrated their capacity to specifically target monocytes in wound tissues. These exosomes facilitate the differentiation of monocytes into M2 macrophages by delivering anti-inflammatory molecules, while concurrently enhancing the migratory capabilities of fibroblasts and endothelial cells. This was accompanied by a reduction in pro-inflammatory cytokines, including TNF-α and IL-6, which accelerated wound healing.
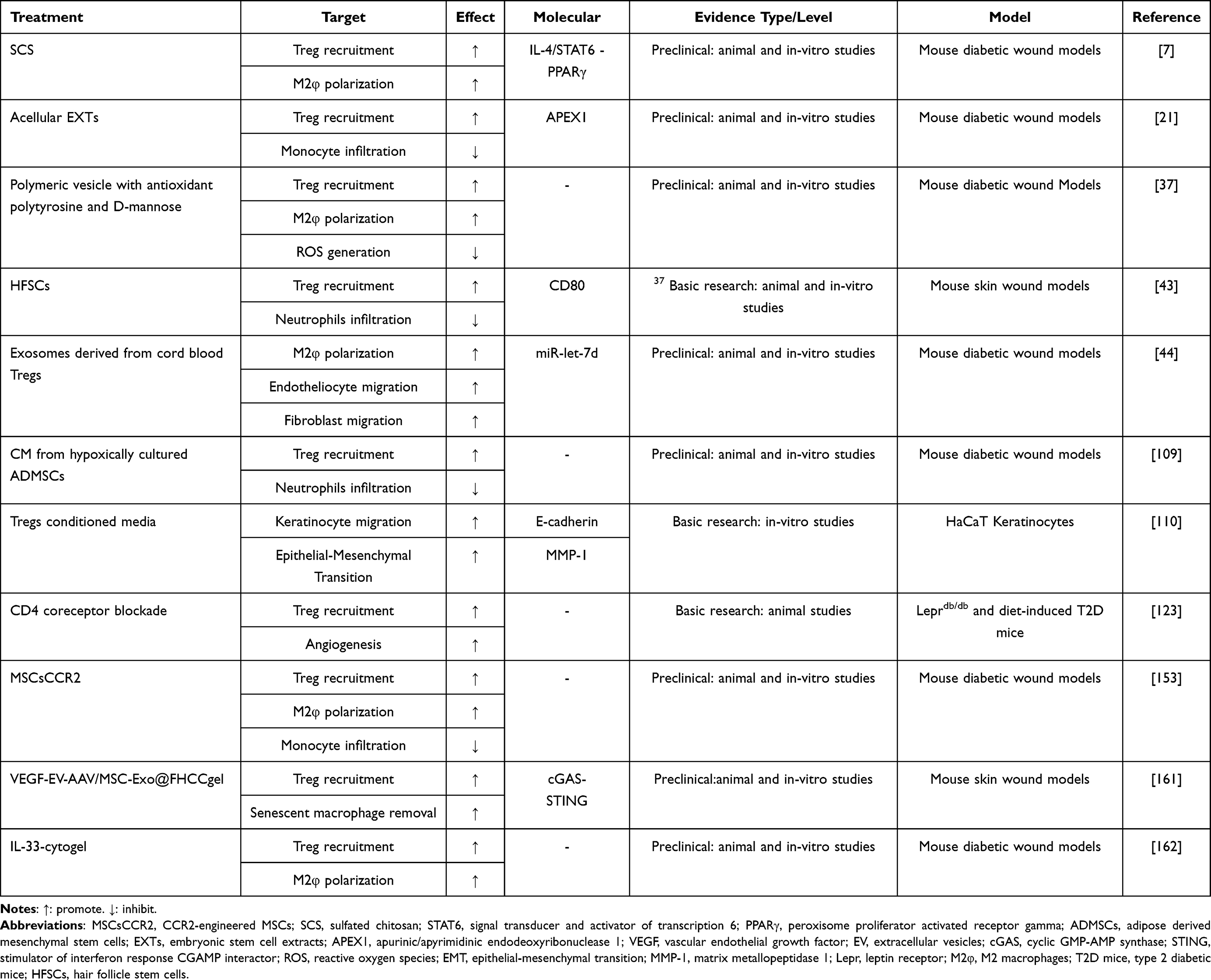 |
Table 1 Overview of Relevant Scientific Findings on Tregs in Diabetic Wound Healing |
Recent biomaterials targeting Tregs display promise in diabetic wound healing by promoting Treg aggregation, M2 macrophage polarization, and their interactions, thereby reducing inflammation and enhancing angiogenesis and epithelialization (Table 1). Shan et al found a significant improvement in the gene transfer efficiency of VEGF using an adeno-associated virus (AAV) encapsulated within extracellular vesicles (EV), termed EV-AAV. The researchers developed a thermosensitive hydrogel (VEGF-EV-AAV/MSC-Exo@FHCCgel) by integrating EV-AAV with exosomes derived from human umbilical cord mesenchymal stem cells (MSC-Exo). This hydrogel facilitated the accumulation of Tregs and the removal of senescent macrophages by inhibiting the cGAS-STING pathway, thereby reducing inflammatory cytokine levels and demonstrating considerable potential for treating chronic wounds.161 Wang et al created a biomaterial, IL-33-cytogel, by encapsulating IL-33 in a biodegradable DNA hydrogel, leveraging IL-33’s ability to enhance wound healing via immune regulation.162 Tregs co-express the IL-33 receptor T1/ST2, allowing IL-33 to activate the MAPK pathway, thereby enhancing Treg survival and expansion.160 This biomaterial attracts type 2 innate lymphoid cells and Tregs to the wound site, facilitates the conversion of macrophages from M1 to M2, reduces local inflammation, and promotes granulation tissue regeneration, thereby significantly accelerating wound healing in diabetic mice. Additionally, biomimetic tolerogenic artificial antigen-presenting cells (aAPCs) made from a mix of negatively charged biodegradable polymer (poly(lactic-co-glycolic acid), PLGA) and cationic biodegradable polymer (poly(beta-amino ester) have been studied for their ability to convert naïve T-cells into induced Tregs (iTregs).163 This non-cellular method of engineering Tregs offers promise as an “off-the-shelf” material for inducing tolerance, potentially aiding diabetic wound healing through improved Treg-mediated immune regulation. Moreover, biomaterials such as alginate-collagen cryogels have been used to deliver Treg inducers, such as eliglustat, to recruit and expand Tregs in subcutaneous tissues, thereby reducing inflammation and promoting tissue regeneration.164 This targeted approach aids in diabetic wound healing by modulating local immune responses, including macrophage polarization and Treg recruitment. Zhang et al found that chronic diabetic wounds are linked to high blood glucose and immune issues, causing increased ROS, immune dysfunction, prolonged inflammation, and delayed healing.37 To combat this, they created a polymeric vesicle with antioxidant polytyrosine and D-mannose to enhance Treg activation. In trials, mice treated with these vesicles exhibited a 23.7% increase in Treg cells and a 91.3% higher M2/M1 ratio than phosphate buffered saline (PBS)-treated mice. The vesicles accelerated wound healing, achieving full recovery of Staphylococcus aureus-infected diabetic wounds in 8 days. Furthermore, Kuang et al engineered mesenchymal stromal cells (MSCs) to overexpress CCR2, thereby improving their ability to target wounds.153 These modified MSCs (MSCsCCR2) boosted local Tregs by secreting anti-inflammatory cytokines, such as TGF-β and IL-10, thereby accelerating diabetic wound healing. This highlights the importance of restoring Treg numbers and function for effective wound healing in patients with diabetes. In conclusion, emerging evidence suggests that biomaterials designed to target Tregs significantly enhance diabetic wound healing by reprogramming the immune microenvironment.
Pharmaceutical Agents for Modulating Tregs
Therapies targeting Tregs or their precursors can improve wound healing by regulating the immune response in wounds.45 Modulating Foxp3 acetylation, a transcription factor for Tregs, can regulate their number and function.165 Histone deacetylase inhibitors have increased the number of functional Treg cells in mouse splenocytes, human PBMCs, T cell clones, and skin-derived T cells, suggesting this as a strategy to boost Treg function. Additionally, immunomodulatory agents, such as lipoic acid and embryonic stem cell extracts, have been found to increase Treg populations and improve wound healing in diabetic models.21,24 Research has demonstrated that lentinan treatment increases Foxp3+ Tregs in NOD mice, enhancing their immunosuppressive activity and potentially reversing hyperglycemia.166 Furthermore, research suggests that vitamin D supplementation can modulate Treg subsets, including ICOS+ Tregs.167 In patients with type 1 diabetes, low vitamin D levels are associated with an imbalance in the Th17/Treg cell ratio. Supplementation with vitamin D has demonstrated efficacy in restoring immune equilibrium by ameliorating this ratio.
Future Research Directions and Challenges
Exploring Tregs in diabetic wound management is promising, with future research focusing on targeted treatments and clinical applications. The primary challenge is to understand the Treg function and stability, especially in immune diseases, such as type 1 diabetes. Tregs are crucial for immune tolerance and preventing autoimmunity; thus, enhancing or restoring them can lead to new therapies. However, the diversity of Treg populations complicates treatment development, necessitating further research into specific markers and pathways.168,169
Targeted Therapy for Tregs
Targeting Tregs offers a promising approach for immunomodulation, as shown in autoimmune and cancer therapies. However, for diabetic wound healing, there is a need for tailored strategies rather than directly applying existing models. Recent research has focused on chimeric antigen receptor (CAR) Tregs, which improve the specificity and effectiveness of Treg therapies. CAR Tregs can be engineered to target specific receptors, selectively suppressing harmful T-cells in autoimmune diseases and restoring immune balance without broadly suppressing the immune system.170 They can also be tailored to target antigens in diabetic wound environments for precise Treg modulation.171 Enhancing Treg stability through metabolic or signaling pathways is also crucial.169,172 Yet, a major challenge remains: understanding how to expand and sustain functional Tregs in the unique, chronically inflamed environment of diabetic wounds, which is a significant barrier to progress in this field.
Future research should adopt a structured, multi-faceted approach: 1. **Treg Heterogeneity and Contextual Cues:** Focus on identifying and mapping the diverse phenotypes and functions of Treg subsets in diabetic wounds compared to autoimmune or cancer models. This will help pinpoint specific targets for precise manipulation instead of general Treg expansion. 2. **Systemic Influences:** Investigating the gut-skin axis is crucial, especially as fecal microbiota transplantation (FMT) shows potential in enhancing diabetic wound healing, although its immunomodulatory effects remain unclear. Peng et al connected FMT to IL-17A-induced keratinocyte activation through the mTOR-HIF1α pathway.173 With Tregs known to influence IL-17 responses,174,175 it is vital to understand how FMT alters Treg dynamics both systemically and at wound sites to aid healing. Clarifying this could lead to new microbiome-based treatments. 3. **Precision Engineering:** CRISPR/Cas gene-editing tools hold revolutionary promise for improving Treg function.136,176,177 Luo et al’s work, using nanoparticle-delivered CRISPR to edit dendritic cells and expand autoantigen-specific Tregs in type 1 diabetes, demonstrates this potential.176 The key opportunity is to adapt these platforms to create wound-sensing Tregs. Future efforts should focus on designing “smart” Tregs activated by wound-specific signals for localized immunomodulation, thereby reducing risks such as systemic suppression. This requires thorough validation in diabetic wound models for safety and efficacy.
To sum up, this review positions the future of Treg-based diabetic wound therapy as requiring context-specific innovation rather than merely applying current technologies. The essential way forward involves a structured research plan that addresses the particular instability of Tregs in diabetic wounds, incorporates systemic factors such as the gut microbiome, and develops precision-engineered cellular therapies customized for the wound’s pathological environment.
Challenges in Translating Treg Research to Clinical Practice
While basic studies have shown alterations and some functions of Tregs in diabetic wounds, direct clinical evidence for treating these wounds with Tregs is lacking. Fang’s and Gan’s teams found reduced Treg infiltration in the wounds of diabetic patients, indicating a potential role in poor healing.82,178 However, their studies focused on identifying disease-related genes and immune characteristics, not on clinical interventions or treatment efficacy. Further clinical research is needed to establish Tregs as a therapeutic option. However, translating Treg research into clinical practice is challenging due to the lack of standardized protocols for Treg isolation, expansion, and administration, leading to inconsistent results across studies. Additionally, the complex immune system and microenvironment in diabetic wounds complicate the understanding of Treg interactions and functions, hindering the development of precise therapeutic strategies. Bender et al conducted a randomized, double-blind, multicenter Phase 2 trial to assess factors affecting the efficacy of autologous polyclonal expanded Tregs (exTregs) in 110 children and adolescents with newly diagnosed type 1 diabetes.179 Participants received either a high dose (20 × 106 cells/kg) or a low dose (1 × 106 cells/kg) of exTregs or a placebo. A single dose of exTregs was safe and exhibited an activated phenotype with suppressive function in vitro. A temporary increase in circulating Tregs was observed in the high-dose group, indicating successful transfer. Lower in vitro fold expansion of exTregs, adjusted for age, correlated with better preservation of C-peptide levels, regardless of dose. These findings indicate that a single infusion of polyclonal exTregs does not significantly alter T1DM progression, and the functional quality of Tregs may be more crucial for therapeutic success. Further efforts are needed to improve the expansion, quality control, and transplantation of Tregs for treating diabetes.
Cell-based therapies, such as those involving Tregs, face substantial hurdles due to their high cost, time demands, and potential side effects.180 The use of polyclonal Tregs with unknown antigen-specificities raises concerns about systemic immunosuppression and increased infection risk.171 Diabetic patients, especially with poor blood sugar control, are more prone to infections due to a systemic immunosuppressive state.181 High blood sugar leads to increased methylglyoxal production, a compound that links high sugar levels to immunosuppression by forming AGEs and impairing immune cell function. This raises concerns about Tregs’ immunosuppressive role and infection risks in diabetics. A recent study found that excessive ROS activates the PI3K-AKT-mTOR pathway, leading to the differentiation of CD4+ T cells into Tregs and the apoptosis of effector T cells, thereby worsening immunosuppression and hindering infection control and tissue repair.182 This contradicts previous research suggesting ROS inhibits Treg differentiation, but highlights the risk of excessive Tregs causing immunosuppression. Nevertheless, a study has yielded promising results for using Tregs in diabetes treatment. Marek Trzonkowska et al reported a clinical trial involving 12 children with type 1 diabetes who received autologous, expanded Tregs. The participants received one or two infusions without serious side effects, and their antibody responses remained intact. Post-infusion, Treg counts increased, most patients exhibited positive outcomes, including higher C-peptide levels and reduced insulin needs, with two achieving insulin independence within a year.183 These results suggest that repeated Treg infusions are safe and may enhance β-cell survival in patients with type 1 diabetes. Another concern is fibrosis and excessive scarring. In other diseases, Tregs treatment can lead to fibrosis, as seen in metabolic-associated fatty liver disease, where increased Tregs promote fibrosis.184 This suggests Tregs’ role in certain conditions is complex, and their treatment may risk diabetic wound scar hyperplasia. Nevertheless, it is currently uncertain whether Tregs play a role in long-term fibrosis and scarring in diabetic wound healing, necessitating further research.
Conclusion
Recent research has unequivocally demonstrated the pivotal role of Tregs in the wound healing process associated with diabetes, underscoring their capacity to modulate inflammation, facilitate cellular regeneration, and enhance angiogenesis. Nonetheless, a comprehensive understanding of Treg functions necessitates an examination of their spatiotemporal specificity to effectively balance their anti-inflammatory and immunosuppressive effects while mitigating potential fibrotic risks.
Future research should focus on integrating mechanistic insights with clinical impacts by prioritizing key approaches. First, it is essential to quantify and identify Tregs at wound sites and to examine altered signaling pathways under hyperglycemic conditions. A major challenge is understanding the stability and plasticity of Tregs in chronic hyperglycemia and wound environments, particularly how they may become dysregulated or fibrotic. Second, clinical translation faces hurdles such as developing targeted delivery systems to regulate or recruit Tregs at wound sites without causing systemic immunosuppression or fibrosis, and identifying biomarkers to determine patient suitability for Treg-focused treatments.
Beyond reviewing existing literature, future research should: 1) enhance basic and preclinical research to address Treg instability in hyperglycemic conditions, potential immunosuppression, and fibrosis risk; 2) foster interdisciplinary collaboration among immunology, bioengineering, and clinical trial design. The clinical aim is to develop new immunomodulatory therapies, such as local drugs or cell therapies, to stabilize Tregs in wounds. These methods will close knowledge gaps, facilitate the application of Treg biology to the development of targeted treatments, and shift the focus from general wound care to precise immunomodulation. In essence, understanding Treg biology and applying this knowledge to research and clinical priorities is vital for creating safe, effective immune therapies to improve diabetic wound healing and patient quality of life.
Abbreviations
AAV, adeno-associated virus; AGEs, advanced glycation end products; APC, antigen-presenting cell; AREG, amphiregulin; BCL6, B-cell lymphoma 6; CAR, chimeric antigen receptor; CCL22, C-C motif chemokine ligand 22; CCR2, C-C motif chemokine receptor 2; CM, conditioned medium; CNS2, conserved non-coding sequences 2; COPD, chronic obstructive pulmonary disease; CTLA-4, cytotoxic T-lymphocyte-associated antigen 4; CXCL5, C-X-C motif chemokine ligand 5; DCs, dendritic cells; DFU, diabetic foot ulcers; DGAT1, diacylglycerol O-acyltransferase 1; DN Tregs, double negative Tregs; DT, diphtheria toxin; EGFR, epidermal growth factor receptor; EMT, epithelial-mesenchymal transition; EV, extracellular vesicles; Exo, exosome; EXTs, embryonic stem cell extracts; exTregs, expanded Tregs; FMT, fecal microbiota transplantation; Foxp3, forkhead box p3; GATA-3, GATA-binding factor 3; Gαi3, guanine nucleotide-binding protein α subunit 3; GSTM5, glutathione s-transferase mu 5; GVHD, graft-versus-host disease; HFSCs, hair follicle stem cells; ICOS, inducible T cell costimulator; IFN-γ, interferon gamma; IGF-1, insulin like growth factor 1; IKKβ, inhibitor of nuclear factor kappa-B kinase subunit beta; IL-1β, interleukin-1 beta; iTreg, induced Treg; KGF, keratinocyte growth factor; Ly6G, lymphocyte antigen 6 family member G; MAP4K3, mitogen-activated protein kinase kinase kinase kinase 3; MHC, major histocompatibility complex; MMP-1, matrix metallopeptidase 1; MSCs, mesenchymal stromal cells; NCLX, Na+/Ca2+/Li+ exchanger; NETs, neutrophil extracellular traps; NF-κB, nuclear factor kappa B; NOD, non-obese diabetic; nTreg, natural Treg; PBMCs, peripheral blood mononuclear cells; PBS, phosphate buffered saline; PI3K, phosphoinositide-3-kinase; PLC, phospholipase C; PLGA, poly(lactic-co-glycolic acid); pTregs, peripherally induced Tregs; RNS, reactive nitrogen species; RORγt, retinoid-related orphan receptor γ; ROS, reactive oxygen species; Runx3, runt-related transcription factor 3; SCS, sulfated chitosan; SP1, sp1 transcription factor; STAT3, signal transducer and activator of transcription 3; STZ, streptozotocin; TBX21, T-box transcription factor 21; T-bet, T-box transcription factor; TCR, T cell receptor; Tet2, ten-eleven translocation 2; Tfh, T follicular helper; TGF-β, transforming growth factor-beta; Th1, T helper 1; TNBC, triple-negative breast cancer; TNF-α, tumor necrosis factor-alpha; Tregs, regulatory T cells; tTregs, thymus-derived Tregs; T1DM, type 1 diabetes mellitus; T2DM, type 2 diabetes mellitus; UACR, urinary albumin-creatinine ratio; VEGF, vascular endothelial growth factor; Wnt5a, wnt family member 5a; α-SMA, alpha smooth muscle actin.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this study.
Acknowledgments
The schematic diagrams were drawn using Figdraw (www.figdraw.com).
Author Contributions
J. Lin, Y. Liu and P. Min: Investigation, Writing-original draft, Visualization, Writing-review and editing. H. Zhao: Conceptualization, Project administration, Supervision, Writing-review and editing. Z. Xia: Conceptualization, Funding acquisition, Supervision, Writing-review and editing. Y. Zheng: Conceptualization, Project administration, Funding acquisition, Supervision, Writing-review and editing. Each author has significantly contributed to the report and offered valuable feedback on the revisions of the article. All authors have approved the forthcoming version and collectively agreed on the journal for submission. They have also accepted responsibility for all aspects of the work.
Funding
This work was supported by the National Key R&D Program of China (2024YFA1108401); the National Natural Science Foundation of China (82372512, 81930057); Shanghai Municipal Health Commission Medical New Technology Research and Translation Seed Program Project(2025ZZ2069); Basic medical research project of Changhai Hospital (2023YQ02); Changhong talent plan of Changhai Hospital; Deep Blue Talent Project of Naval Medical University; Clinical Medical Research of Changhai Hospital (2024LYC06); Shanghai Top Priority Research Center Project (2023ZZ02013); Medical New Technology Research and Transformation Project of Shanghai Municipal Health Commission (2025ZZ2069).
Disclosure
The authors disclose that they have no identifiable conflicting financial interests or personal relationships that could potentially bias the findings presented in this paper.
References
1. Roep BO. Personalised prevention therapy in type 1 diabetes. Lancet. 2025;406(10519):2515–22. doi:10.1016/S0140-6736(25)02005-7
2. Kuhn C, Besançon A, Lemoine S, et al. Regulatory mechanisms of immune tolerance in type 1 diabetes and their failures. J Autoimmun. 2016;71:69. doi:10.1016/j.jaut.2016.05.002
3. Jeffcoate W, Boyko EJ, Game F, Cowled P, Senneville E, Fitridge R. Causes, prevention, and management of diabetes-related foot ulcers. Lancet Diabetes Endocrinol. 2024;12:472. doi:10.1016/S2213-8587(24)00110-4
4. Li Y, Elkord E. Targeting Tregs in T1DM: bridging heterogeneity, mechanisms, and clinical progress. Trends Pharmacol Sci. 2025.
5. Burgess JL, Wyant WA, Abujamra BA, Kirsner RS, Jozic I. Diabetic Wound-Healing Science. Medicina. 2021;57.
6. Knoedler S, Knoedler L, Kauke-Navarro M, et al. Regulatory T cells in skin regeneration and wound healing. Military Med Res. 2023;10:49. doi:10.1186/s40779-023-00484-6
7. Shen T, Xu H, Dai K, Wang J, Liu C. Sulfated polysaccharide facilitates macrophage-Treg crosstalk to mitigate chronic inflammation in diabetic wound healing. Bioact Mater. 2025;55:640. doi:10.1016/j.bioactmat.2025.09.044
8. Mgwenya TN, Abrahamse H, Houreld NN. Photobiomodulation studies on diabetic wound healing: an insight into the inflammatory pathway in diabetic wound healing. Wound Repair Regener. 2025;33:e13239. doi:10.1111/wrr.13239
9. Ding H, Wang Y, Bai R, et al. Exosomes From Adipose-Derived Stem Cells Inhibit Skin T-Cell Activation and Alleviate Wound Inflammation. Aest surg j. 2025;45:723. doi:10.1093/asj/sjaf040
10. Huang Q, Zhu J. Regulatory T cell-based therapy in type 1 diabetes: latest breakthroughs and evidence. Int Immunopharmacol. 2024;140:112724. doi:10.1016/j.intimp.2024.112724
11. Seltzer RR, Thompson BS. Pediatrician as advocate and protector: an approach to medical neglect for children with medical complexity. Pediatr Clin N Am. 2024;71:59. doi:10.1016/j.pcl.2023.08.006
12. Mikami N, Kawakami R, Sugimoto A, Arai M, Sakaguchi S. Generating functionally stable and antigen-specific T(reg) cells from effector T cells for cell therapy of inflammatory diseases. Sci, trans med. 2025;17:eadr6049. doi:10.1126/scitranslmed.adr6049
13. Nguyen VH. Balancing act for Treg immunotherapy. Blood. 2011;117:2751. doi:10.1182/blood-2011-01-328674
14. Liu S, Tao Z, Lou J, et al. CD4(+)CCR8(+) tregs in ovarian cancer: a potential effector tregs for immune regulation. J Transl Med. 2023;21:803. doi:10.1186/s12967-023-04686-3
15. Agidigbi TS, Fram B, Molloy I, Riedel M, Wiznia D, Oh I. CD177, MYBL2, and RRM2 are potential biomarkers for musculoskeletal infections. Clin Orthopaedics Related Res. 2025;483:1062. doi:10.1097/CORR.0000000000003402
16. Ohkura N, Hamaguchi M, Sakaguchi S. FOXP3+ regulatory T cells: control of FOXP3 expression by pharmacological agents. Trends Pharmacol Sci. 2011;32:158. doi:10.1016/j.tips.2010.12.004
17. Contreras-Castillo E, García-Rasilla VY, García-Patiño MG, Licona-Limón P. Stability and plasticity of regulatory T cells in health and disease. J Leukoc Biol. 2024;116:33. doi:10.1093/jleuko/qiae049
18. Chi JN, Yang JY, Hsueh CH, Tsai CY, Chuang HC, Tan TH. MAP4K3/GLK inhibits treg differentiation by direct phosphorylating IKKβ and inducing IKKβ-mediated FoxO1 nuclear export and Foxp3 downregulation. Theranostics. 2022;12:5744. doi:10.7150/thno.72148
19. Chen X, Du Y, Lin X, Qian Y, Zhou T, Huang Z. CD4+CD25+ regulatory T cells in tumor immunity. Int Immunopharmacol. 2016;34:244. doi:10.1016/j.intimp.2016.03.009
20. Cha J, Kim CG, Sim NS, et al. 4-1BB(+) Tregs and inhibitory progenitor exhausted T cells confer resistance to anti-PD-L1 and anti-CTLA-4 combination therapy. Cell Rep Med. 2025;6:102408. doi:10.1016/j.xcrm.2025.102408
21. Loretelli C, Nasr MB, Giatsidis G, et al. Embryonic stem cell extracts improve wound healing in diabetic mice. Acta diabetologica. 2020;57:883. doi:10.1007/s00592-020-01500-0
22. Arroyo-Olarte RD, Pérez-Noriega FA, Correa-Pérez MF, Mejía-Muñoz A, Terrazas LI, Leon-Cabrera S. STAT6 inhibition stabilizes induced regulatory T cells and enhances their therapeutic potential in inflammatory bowel disease. Immunol Res. 2025;73:158. doi:10.1007/s12026-025-09686-7
23. Iamsawat S, Daenthanasanmak A, Voss JH, et al. Stabilization of Foxp3 by targeting JAK2 enhances efficacy of CD8 induced regulatory T cells in the prevention of graft-versus-host disease. J Iimmunol. 2018;201:2812. doi:10.4049/jimmunol.1800793
24. Rhoiney ML, Alvizo CR, Jameson JM. Skin homeostasis and repair: a T lymphocyte perspective. J Iimmunol. 2023;211:1266. doi:10.4049/jimmunol.2300153
25. Ohue Y, Nishikawa H. Regulatory T (Treg) cells in cancer: can treg cells be a new therapeutic target? Cancer Sci. 2019;110:2080. doi:10.1111/cas.14069
26. Li R, Li H, Yang X, Hu H, Liu P, Liu H. Crosstalk between dendritic cells and regulatory T cells: protective effect and therapeutic potential in multiple sclerosis. Front Immunol. 2022;13:970508. doi:10.3389/fimmu.2022.970508
27. Kim KS, Habashy K, Gould A, et al. Fc-enhanced anti-CTLA-4, anti-PD-1, doxorubicin, and ultrasound-mediated blood-brain barrier opening: a novel combinatorial immunotherapy regimen for gliomas. Neuro-Oncology. 2024;26:2044. doi:10.1093/neuonc/noae135
28. Gao JF, McIntyre MS, Juvet SC, et al. Regulation of antigen-expressing dendritic cells by double negative regulatory T cells. Eur j immunol. 2011;41:2699. doi:10.1002/eji.201141428
29. Wing K, Sakaguchi S. Regulatory T cells exert checks and balances on self tolerance and autoimmunity. Nat Immunol. 2010;11:7. doi:10.1038/ni.1818
30. Mavin E, Nicholson L, Ahmed SR, Gao F, Dickinson A, Wang XN. Human regulatory T cells mediate transcriptional modulation of dendritic cell function. J Iimmunol. 2017;198:138. doi:10.4049/jimmunol.1502487
31. Li J, Lin X, Wang J, et al. Dendritic Cell repression by TNF-α-primed exosomes accelerate T2DM wound healing through miR-146a-5p/TXNIP/NLRP3 axis. Int J Nanomed. 2025;20:9963. doi:10.2147/IJN.S522994
32. Tang Q, Adams JY, Tooley AJ, et al. Visualizing regulatory T cell control of autoimmune responses in nonobese diabetic mice. Nat Immunol. 2006;7:83. doi:10.1038/ni1289
33. Iikuni N, Lourenço EV, Hahn BH, Cava AL. Cutting edge: regulatory T cells directly suppress B cells in systemic lupus erythematosus. J Iimmunol. 2009;183:1518. doi:10.4049/jimmunol.0901163
34. Wei X, Zhang J, Gu Q, et al. Reciprocal expression of IL-35 and IL-10 defines two distinct effector treg subsets that are required for maintenance of immune tolerance. Cell Rep. 2017;21:1853. doi:10.1016/j.celrep.2017.10.090
35. Goswami TK, Singh M, Dhawan M, et al. Regulatory T cells (Tregs) and their therapeutic potential against autoimmune disorders - advances and challenges. Hum Vaccines Immunother. 2022;18:2035117. doi:10.1080/21645515.2022.2035117
36. Sojka DK, Huang YH, Fowell DJ. Mechanisms of regulatory T-cell suppression - a diverse arsenal for a moving target. Immunology. 2008;124:13. doi:10.1111/j.1365-2567.2008.02813.x
37. Zhang W, Ge Z, Xiao Y, Liu D, Du J. Antioxidant and immunomodulatory polymer vesicles for effective diabetic wound treatment through ROS scavenging and immune modulating. Nano Lett. 2024;24:9494. doi:10.1021/acs.nanolett.4c01869
38. Tiemessen MM, Jagger AL, Evans HG, van Herwijnen MJ, John S, Taams LS. CD4+CD25+Foxp3+ regulatory T cells induce alternative activation of human monocytes/macrophages. Proc Natl Acad Sci USA. 2007;104:19446. doi:10.1073/pnas.0706832104
39. Proto JD, Doran AC, Gusarova G, et al. Regulatory T cells promote macrophage efferocytosis during inflammation resolution. Immunity. 2018;49:666. doi:10.1016/j.immuni.2018.07.015
40. Richards H, Williams A, Jones E, et al. Novel role of regulatory T cells in limiting early neutrophil responses in skin. Immunology. 2010;131:583. doi:10.1111/j.1365-2567.2010.03333.x
41. Lewkowicz N, Klink M, Mycko MP, Lewkowicz P. Neutrophil--CD4+CD25+ T regulatory cell interactions: a possible new mechanism of infectious tolerance. Immunobiology. 2013;218:455. doi:10.1016/j.imbio.2012.05.029
42. Mathur AN, Zirak B, Boothby IC, et al. Treg-cell control of a CXCL5-IL-17 inflammatory axis promotes hair-follicle-stem-cell differentiation during skin-barrier repair. Immunity. 2019;50:655. doi:10.1016/j.immuni.2019.02.013
43. Luan J, Truong C, Vuchkovska A, et al. CD80 on skin stem cells promotes local expansion of regulatory T cells upon injury to orchestrate repair within an inflammatory environment. Immunity. 2024;57:1071. doi:10.1016/j.immuni.2024.04.003
44. Yang F, Cai D, Kong R, et al. Exosomes derived from cord blood treg cells promote diabetic wound healing by targeting monocytes. Biochem Pharmacol. 2024;226:116413. doi:10.1016/j.bcp.2024.116413
45. Mohsin F, Javaid S, Tariq M, Mustafa M. Molecular immunological mechanisms of impaired wound healing in diabetic foot ulcers (DFU), current therapeutic strategies and future directions. Int Immunopharmacol. 2024;139:112713. doi:10.1016/j.intimp.2024.112713
46. Iglesias-Escudero M, Arias-González N, Martínez-Cáceres E. Regulatory cells and the effect of cancer immunotherapy. Mol Cancer. 2023;22:26. doi:10.1186/s12943-023-01714-0
47. Wang Y, Chen W, Qiao S, et al. Lipid droplet accumulation mediates macrophage survival and Treg recruitment via the CCL20/CCR6 axis in human hepatocellular carcinoma. Cell Mol Immunol. 2024;21:1120. doi:10.1038/s41423-024-01199-x
48. Malla RR, Vasudevaraju P, Vempati RK, Rakshmitha M, Merchant N, Nagaraju GP. Regulatory T cells: their role in triple-negative breast cancer progression and metastasis. Cancer. 2022;128:1171. doi:10.1002/cncr.34084
49. Dixon D, Edmonds M. Managing diabetic foot ulcers: pharmacotherapy for wound healing. Drugs. 2021;81:29. doi:10.1007/s40265-020-01415-8
50. Huelsboemer L, Knoedler L, Kochen A, et al. Cellular therapeutics and immunotherapies in wound healing - on the pulse of time? Military Med Res. 2024;11:23. doi:10.1186/s40779-024-00528-5
51. Crunkhorn S. Promoting efferocytosis heals diabetic wounds. Nat Rev Drug Discov. 2022;21:491. doi:10.1038/d41573-022-00097-z
52. Uberoi A, McCready-Vangi A, Grice EA. The wound microbiota: microbial mechanisms of impaired wound healing and infection. Nat Rev Microbiol. 2024;22:507. doi:10.1038/s41579-024-01035-z
53. Zhang W, Liu W, Leng F, Shen M, Xie J. Dietary non-starch plant polysaccharides: multi-mechanisms for managing diabetic microvascular complications. Carbohydr Polym. 2025;368:124074. doi:10.1016/j.carbpol.2025.124074
54. Falanga V, Isseroff RR, Soulika AM, et al. Chronic wounds. Nat Rev Dis Prim. 2022;8:50. doi:10.1038/s41572-022-00377-3
55. Bansal S, Siddarth M, Chawla D, Banerjee BD, Madhu SV, Tripathi AK. Advanced glycation end products enhance reactive oxygen and nitrogen species generation in neutrophils in vitro. Mol Cell Biochem. 2012;361:289. doi:10.1007/s11010-011-1114-9
56. Zhan QY, Xie LX, Wang C. Promoting critical care system and capacity building in pulmonary and critical care medicine subspecialties]. Zhonghua yi xue za zhi. 2023;103:3149. doi:10.3760/cma.j.cn112137-20230602-00919
57. Haalboom M. Chronic wounds: innovations in diagnostics and therapeutics. Curr Med Chem. 2018;25:5772. doi:10.2174/0929867324666170710120556
58. Morton LM, Phillips TJ. Wound healing and treating wounds: differential diagnosis and evaluation of chronic wounds. J Am Acad Dermatol. 2016;74:589. doi:10.1016/j.jaad.2015.08.068
59. Ahmad W, Ijaz B, Shabbiri K, Ahmed F, Rehman S. Oxidative toxicity in diabetes and Alzheimer’s disease: mechanisms behind ROS/ RNS generation. J Biomed Sci. 2017;24:76. doi:10.1186/s12929-017-0379-z
60. Kunkemoeller B, Bancroft T, Xing H, et al. Elevated Thrombospondin 2 Contributes to Delayed Wound Healing in Diabetes. Diabetes. 2016;2019:68.
61. Wan L, Bai X, Zhou Q, et al. The advanced glycation end-products (AGEs)/ROS/NLRP3 inflammasome axis contributes to delayed diabetic corneal wound healing and nerve regeneration. Int J Bio Sci. 2022;18:809. doi:10.7150/ijbs.63219
62. Yu J, Duan W, Zhang J, et al. Superhydrophobic ROS biocatalytic metal coatings for the rapid healing of diabetic wounds. Mater Today Bio. 2025;32:101840. doi:10.1016/j.mtbio.2025.101840
63. Zhao Y, Zhao Y, Xu B, Liu H, Chang Q. Microenvironmental dynamics of diabetic wounds and insights for hydrogel-based therapeutics. J Tissue Eng. 2024;15:20417314241253290. doi:10.1177/20417314241253290
64. Han YF, Sun TJ, Han YQ, Xu G, Liu J, Tao R. Clinical perspectives on mesenchymal stem cells promoting wound healing in diabetes mellitus patients by inducing autophagy. Eur Rev Med Pharmacol Sci. 2015;19:2666.
65. Swoboda L, Held J. Impaired wound healing in diabetes. J Wound Care. 2022;31:882. doi:10.12968/jowc.2022.31.10.882
66. Zhang SJ, Geng MM, Li QW, et al. Oxidative stress impairs the expansion of regulatory T cells in active vitiligo via dysregulated CGRP-RAMP1-Gαi3 signaling. Free Radic Biol Med. 2026;245:18. doi:10.1016/j.freeradbiomed.2025.12.041
67. Fenercioglu AK, Uzun H, Unal DO. The convergent immunopathogenesis of cigarette smoke exposure: from oxidative stress to epigenetic reprogramming in chronic disease. Int J Mol Sci. 2025;27:187. doi:10.3390/ijms27010187
68. Qi H, Shi J, Wei X, et al. Reactive oxygen species scavenging and thermosensitive smart release-stiffening integrated hydrogel for diabetic wound therapy. ACS nano. 2026;20:6034. doi:10.1021/acsnano.5c19613
69. Liu Z, Benard O, Syeda MM, Schuster VL, Chi Y. Inhibition of Prostaglandin Transporter (PGT) promotes perfusion and vascularization and accelerates wound healing in non-diabetic and diabetic rats. PLoS One. 2015;10:e0133615. doi:10.1371/journal.pone.0133615
70. Goldberg SR, Diegelmann RF. What makes wounds chronic. Surg Clin North Am. 2020;100:681. doi:10.1016/j.suc.2020.05.001
71. Li WW, Carter MJ, Mashiach E, Guthrie SD. Vascular assessment of wound healing: a clinical review. Int Wound J. 2017;14:460. doi:10.1111/iwj.12622
72. Davis FM, Kimball A, Boniakowski A, Gallagher K. Dysfunctional wound healing in diabetic foot ulcers: new crossroads. Curr Diabetes Rep. 2018;18:2. doi:10.1007/s11892-018-0970-z
73. Tricou LP, Al-Hawat ML, Cherifi K, Manrique G, Freedman BR, Matoori S. Wound pH-modulating strategies for diabetic wound healing. Adv Wound Care. 2024;13:446. doi:10.1089/wound.2023.0129
74. Zhao R, Liang H, Clarke E, Jackson C, Xue M. Inflammation in chronic wounds. Int J Mol Sci. 2016;17:2085. doi:10.3390/ijms17122085
75. Boon RA. Circulating MicroRNAs link inflammation to impaired wound healing in diabetes. Arteriosclerosis Thrombosis Vasc Biol. 2015;35:1296. doi:10.1161/ATVBAHA.115.305670
76. Kołodziej M, Saracyn M, Lubas A, et al. TSH stimulation before PET/CT as our frenemy in detecting thyroid cancer metastases-final results of a retrospective analysis. Cancers. 2024;16:3413. doi:10.3390/cancers16193413
77. Zhou Y, Dai F, Zhao S, et al. pH and glucose dual-responsive hydrogels promoted diabetic wound healing by remodeling the wound microenvironment. Adv Healthcare Mater. 2025;14:e2500810. doi:10.1002/adhm.202500810
78. Fu X, Wang J, Zhang H, Liu Z. Oxygen-vacancy-engineered self-regenerative nanozymes in dual-signal-responsive dynamic hydrogels: a closed-loop therapeutic strategy for diabetic wound microenvironment remodeling. Adv Healthcare Mater. 2025;15:e04199. doi:10.1002/adhm.202504199
79. Ali IH, Khalil IA, Hefnawy A, Chester A, Yacoub MH, El-Sherbiny IM. Exogenous and endogenous nitric oxide eluting polylactic acid-based nanofibrous scaffolds for enhancing angiogenesis of diabetic wounds. Int J Biol Macromol. 2024;261:129736. doi:10.1016/j.ijbiomac.2024.129736
80. Xue H, Zhang C, Lin D, et al. Isoliquiritigenin micellar microneedle for pH monitoring and diabetic wound healing. Mater Today Bio. 2025;35:102356. doi:10.1016/j.mtbio.2025.102356
81. Zeng H, Chi H. Metabolic control of regulatory T cell development and function. Trends immunol. 2015;36:3. doi:10.1016/j.it.2014.08.003
82. Gan J, Chen Y, Tang L, et al. Immune microenvironment and fibroblast subpopulation in diabetic wound healing. iScience. 2025;28:113440. doi:10.1016/j.isci.2025.113440
83. Riaz M, Iqbal MZ, Klar AS, Biedermann T. Immunomodulatory mechanisms of chronic wound healing: translational and clinical relevance. MedComm. 2025;6:e70378. doi:10.1002/mco2.70378
84. Almeida BM, Evans R, Kayssi A. Fundamentals of wound care for amputation prevention. Sem vascu surg. 2025;38:54. doi:10.1053/j.semvascsurg.2025.01.001
85. Scherm MG, Serr I, Zahm AM, et al. miRNA142-3p targets Tet2 and impairs treg differentiation and stability in models of type 1 diabetes. Nat Commun. 2019;10:5697. doi:10.1038/s41467-019-13587-3
86. Shimokawa C, Kato T, Takeuchi T, et al. CD8(+) regulatory T cells are critical in prevention of autoimmune-mediated diabetes. Nat Commun. 2020;11:1922. doi:10.1038/s41467-020-15857-x
87. Hull CM, Peakman M, Tree TIM. Regulatory T cell dysfunction in type 1 diabetes: what’s broken and how can we fix it? Diabetologia. 2017;60:1839. doi:10.1007/s00125-017-4377-1
88. Kuczma M, Wang CY, Ignatowicz L, Gourdie R, Kraj P. Altered connexin 43 expression underlies age-dependent decrease of regulatory T cell suppressor function in nonobese diabetic mice. J Iimmunol. 2015;194:5261. doi:10.4049/jimmunol.1400887
89. Haertel E, Joshi N, Hiebert P, Kopf M, Werner S. Regulatory T cells are required for normal and activin-promoted wound repair in mice. Eur j immunol. 2018;48:1001. doi:10.1002/eji.201747395
90. Hajam EY, Panikulam P, Chu CC, Jayaprakash H, Majumdar A, Jamora C. The expanding impact of T-regs in the skin. Front Immunol. 2022;13:983700. doi:10.3389/fimmu.2022.983700
91. Bischoff L, Alvarez S, Dai DL, Soukhatcheva G, Orban PC, Verchere CB. Cellular mechanisms of CCL22-mediated attenuation of autoimmune diabetes. J Iimmunol. 2015;194:3054. doi:10.4049/jimmunol.1400567
92. Lee YS, Eun HS, Kim SY, et al. Hepatic immunophenotyping for streptozotocin-induced hyperglycemia in mice. Sci Rep. 2016;6:30656. doi:10.1038/srep30656
93. Wang C, Jiang X, Lv J, et al. TPN10475 constrains effector T lymphocytes activation and attenuates experimental autoimmune encephalomyelitis pathogenesis by facilitating TGF-β signal transduction. J Neuroimmune Pharmacol. 2024;19:6. doi:10.1007/s11481-024-10109-x
94. Korn T, Oukka M, Kuchroo V, Bettelli E. Th17 cells: effector T cells with inflammatory properties. Semin Immunopathol. 2007;19:362. doi:10.1016/j.smim.2007.10.007
95. Yang J, Zhu X, Feng J. The changes in the quantity of lymphocyte subpopulations during the process of sepsis. Int J Mol Sci. 2024;25.
96. Kaskow BJ, Baecher-Allan C. Effector T cells in multiple sclerosis. Cold Spring Harbor Perspect Med. 2018;8:a029025. doi:10.1101/cshperspect.a029025
97. Wang T, Xue Y, Zhang W, Zheng Z, Peng X, Zhou Y. Collagen sponge scaffolds loaded with Trichostatin A pretreated BMSCs-derived exosomes regulate macrophage polarization to promote skin wound healing. Int J Biol Macromol. 2024;269:131948. doi:10.1016/j.ijbiomac.2024.131948
98. Chen C, Yang J, Shang R, et al. Orchestration of macrophage polarization dynamics by fibroblast-secreted exosomes during skin wound healing. J invest dermatol. 2025;145:171. doi:10.1016/j.jid.2024.05.007
99. Li Y, Du K, Peng D, et al. Local delivery of siRNA using lipid-based nanocarriers with ROS-scavenging ability for accelerated chronic wound healing in diabetes. Biomaterials. 2025;322:123411. doi:10.1016/j.biomaterials.2025.123411
100. Sun L, Wang X, Deng T, et al. Bionic sulfated glycosaminoglycan-based hydrogel inspired by snail mucus promotes diabetic chronic wound healing via regulating macrophage polarization. Int J Biol Macromol. 2024;281:135708. doi:10.1016/j.ijbiomac.2024.135708
101. Tao L, Wu S, Wang Q, et al. IL-27 accelerates diabetic wound healing by modulating macrophage polarization. Int Immunopharmacol. 2025;155:114575. doi:10.1016/j.intimp.2025.114575
102. Liu K, Cheong KL, He Y, et al. Pitaya stem polysaccharide promotes wound healing by modulating macrophage polarization via single-cell RNA sequencing evidence. Int J Biol Macromol. 2025;308:142653. doi:10.1016/j.ijbiomac.2025.142653
103. Nosbaum A, Prevel N, Truong HA, et al. Cutting edge: regulatory T cells facilitate cutaneous wound healing. J Iimmunol. 2016;196:2010. doi:10.4049/jimmunol.1502139
104. Fivenson DP, Faria DT, Nickoloff BJ, et al. Chemokine and inflammatory cytokine changes during chronic wound healing. Wound Repair Regener. 1997;5:310. doi:10.1046/j.1524-475X.1997.50405.x
105. Ryan TJ. Elephantiasis, elastin, and chronic wound healing: 19th century and contemporary viewpoints relevant to hypotheses concerning lymphedema, leprosy, erysipelas, and psoriasis--review and reflections. Lymphology. 2009;42:19.
106. Li Y, Yang T, Jiang B. Neutrophil and neutrophil extracellular trap involvement in neutrophilic asthma: a review. Medicine. 2024;103:e39342. doi:10.1097/MD.0000000000039342
107. Xiang J, Cui M, Zhang Y. Neutrophil extracellular traps and neutrophilic asthma. Respir Med. 2025;245:108150. doi:10.1016/j.rmed.2025.108150
108. Lui PP, Xu JZ, Aziz H, Sen M, Ali N. Jagged-1+ skin Tregs modulate cutaneous wound healing. Sci Rep. 2024;14:20999. doi:10.1038/s41598-024-71512-1
109. Takahashi H, Ohnishi S, Yamamoto Y, et al. Topical application of conditioned medium from hypoxically cultured amnion-derived mesenchymal stem cells promotes wound healing in diabetic mice. Plast Reconstr Surg. 2021;147:1342. doi:10.1097/PRS.0000000000007993
110. Kim D, Lo E, Kim D, Kang J. Regulatory T cells conditioned media stimulates migration in HaCaT Keratinocytes: involvement of wound healing. Clin Cosmet Invest Dermatol. 2020;13:443. doi:10.2147/CCID.S252778
111. Hui SP, Sheng DZ, Sugimoto K, et al. Zebrafish Regulatory T Cells Mediate Organ-Specific Regenerative Programs. Dev Cell. 2017;43:659. doi:10.1016/j.devcel.2017.11.010
112. Dial CF, Tune MK, Doerschuk CM, Mock JR. Foxp3(+) regulatory T cell expression of keratinocyte growth factor enhances lung epithelial proliferation. Am J Respir Cell Mol Biol. 2017;57:162. doi:10.1165/rcmb.2017-0019OC
113. Chen J, Wang C, Pan X, et al. Glycyrrhetinic acid mitigates radiation-induced pulmonary fibrosis via inhibiting the secretion of TGF-β1 by treg cells. Int J Radiat Oncol Biol Phys. 2024;118:218. doi:10.1016/j.ijrobp.2023.08.005
114. Prawira A, Xu H, Mei Y, et al. Targeting Treg-fibroblast interaction to enhance immunotherapy in steatotic liver disease-related hepatocellular carcinoma. Gut. 2025.
115. Short WD, Wang X, Keswani SG. The role of T lymphocytes in cutaneous scarring. Adv Wound Care. 2022;11:121. doi:10.1089/wound.2021.0059
116. Kalekar LA, Cohen JN, Prevel N, et al. Regulatory T cells in skin are uniquely poised to suppress profibrotic immune responses. Sci immunol. 2019;4. doi:10.1126/sciimmunol.aaw2910
117. Murao N, Seino K, Hayashi T, et al. Treg-enriched CD4+ T cells attenuate collagen synthesis in keloid fibroblasts. Exp dermatol. 2014;23:266. doi:10.1111/exd.12368
118. Chen Y, Jin Q, Fu X, Qiao J, Niu F. Connection between T regulatory cell enrichment and collagen deposition in keloid. Exp Cell Res. 2019;383:111549. doi:10.1016/j.yexcr.2019.111549
119. Jin Q, Gui L, Niu F, et al. Macrophages in keloid are potent at promoting the differentiation and function of regulatory T cells. Exp Cell Res. 2018;362:472. doi:10.1016/j.yexcr.2017.12.011
120. Shanmugam VK, Angra D, Rahimi H, McNish S. Vasculitic and autoimmune wounds. J Vasc Surg Venous Lymph Disord. 2017;5:280. doi:10.1016/j.jvsv.2016.09.006
121. Mustoe TA, O’Shaughnessy K, Kloeters O. Chronic wound pathogenesis and current treatment strategies: a unifying hypothesis. Plast Reconstr Surg. 2006;117:35s. doi:10.1097/01.prs.0000225431.63010.1b
122. Meng L, Tang Q, Zhou W, et al. Contributions of T cell signaling for wound healing. J Burn Care Res. 2024;45:1513. doi:10.1093/jbcr/irae151
123. Leung OM, Li J, Li X, et al. Regulatory T cells promote apelin-mediated sprouting angiogenesis in type 2 diabetes. Cell Rep. 2018;24:1610. doi:10.1016/j.celrep.2018.07.019
124. Astarita JL, Dominguez CX, Tan C, et al. Treg specialization and functions beyond immune suppression. Clin Exp Immunol. 2023;211:176. doi:10.1093/cei/uxac123
125. Repertinger SK, Campagnaro E, Fuhrman J, El-Abaseri T, Yuspa SH, Hansen LA. EGFR enhances early healing after cutaneous incisional wounding. J invest dermatol. 2004;123:982. doi:10.1111/j.0022-202X.2004.23478.x
126. Bodnar RJ. Epidermal growth factor and epidermal growth factor receptor: the Yin and Yang in the treatment of cutaneous wounds and cancer. Adv Wound Care. 2013;2:24. doi:10.1089/wound.2011.0326
127. Li B, Tang H, Bian X, et al. Calcium silicate accelerates cutaneous wound healing with enhanced re-epithelialization through EGF/EGFR/ERK-mediated promotion of epidermal stem cell functions. Burns Trauma. 2021;9:tkab029. doi:10.1093/burnst/tkab029
128. Delacher M, Imbusch CD, Weichenhan D, et al. Genome-wide DNA-methylation landscape defines specialization of regulatory T cells in tissues. Nat Immunol. 2017;18:1160. doi:10.1038/ni.3799
129. Minutti CM, Modak RV, Macdonald F, et al. A macrophage-pericyte axis directs tissue restoration via amphiregulin-induced transforming growth factor beta activation. Immunity. 2019;50:645. doi:10.1016/j.immuni.2019.01.008
130. Zaiss DM, Minutti CM, Knipper JA. Immune- and non-immune-mediated roles of regulatory T-cells during wound healing. Immunology. 2019;157:190. doi:10.1111/imm.13057
131. Lužnik Z, Anchouche S, Dana R, Yin J. Regulatory T Cells in Angiogenesis. J Iimmunol. 2020;205:2557. doi:10.4049/jimmunol.2000574
132. Takeuchi M, Nishio Y, Someya H, et al. Autoimmune uveitis attenuated in diabetic mice through imbalance of Th1/Th17 differentiation via suppression of AP-1 signaling pathway in Th cells. Front Immunol. 2024;15:1347018. doi:10.3389/fimmu.2024.1347018
133. Jin X, Zhang C, Gong L, et al. Altered expression of CD39 on memory regulatory T cells in type 1 diabetes patients. Jo diabet. 2019;11:440. doi:10.1111/1753-0407.12870
134. Paul M, Dayal D, Bhansali A, Sachdeva N. Characterization of proinsulin-specific regulatory T cells in type 1 diabetes at different ages of onset. Pediatr Diabetes. 2019;20:271. doi:10.1111/pedi.12813
135. Visperas A, Vignali DA. Are regulatory T cells defective in type 1 diabetes and can we fix them? J Iimmunol. 2016;197:3762. doi:10.4049/jimmunol.1601118
136. Qu G, Chen J, Li Y, Yuan Y, Liang R, Li B. Current status and perspectives of regulatory T cell-based therapy. J Genet Genome. 2022;49:599. doi:10.1016/j.jgg.2022.05.005
137. Zhou KJ, Rong SJ, Liu YC, et al. Regulatory T cell dysfunction and immunotherapeutic breakthroughs in type 1 diabetes. Front Endocrinol. 2025;16:1740102. doi:10.3389/fendo.2025.1740102
138. Watts D, Janßen M, Jaykar M, et al. Transient depletion of Foxp3(+) regulatory T cells selectively promotes aggressive β cell autoimmunity in genetically susceptible DEREG mice. Front Immunol. 2021;12:720133. doi:10.3389/fimmu.2021.720133
139. Qiao YC, Shen J, He L, et al. Changes of regulatory T cells and of proinflammatory and immunosuppressive cytokines in patients with type 2 diabetes mellitus: a systematic review and meta-analysis. J diabet res. 2016;2016:3694957. doi:10.1155/2016/3694957
140. Zhang C, Xiao C, Wang P, et al. The alteration of Th1/Th2/Th17/Treg paradigm in patients with type 2 diabetes mellitus: relationship with diabetic nephropathy. Hum Immunol. 2014;75:289. doi:10.1016/j.humimm.2014.02.007
141. Gu QW, Sun Q, Wang J, Gu WS, Wang W, Mao XM. Effects of glycemic variability on regulatory T cells in patients with type 2 diabetes and kidney disease. Diabetes Metab Syndr Obes. 2023;16:2365. doi:10.2147/DMSO.S413407
142. Zi C, He L, Yao H, Ren Y, He T, Gao Y. Changes of Th17 cells, regulatory T cells, Treg/Th17, IL-17 and IL-10 in patients with type 2 diabetes mellitus: a systematic review and meta-analysis. Endocrine. 2022;76:263. doi:10.1007/s12020-022-03043-6
143. Wang C, Wang H, Dai L, et al. T-helper 17 cell/regulatory T-cell imbalance in COPD combined with T2DM patients. Int J Chronic Obstr. 2021;16:1425. doi:10.2147/COPD.S306406
144. Sheikh V, Zamani A, Mahabadi-Ashtiyani E, Tarokhian H, Borzouei S, Alahgholi-Hajibehzad M. Decreased regulatory function of CD4(+)CD25(+)CD45RA(+) T cells and impaired IL-2 signalling pathway in patients with type 2 diabetes mellitus. Scand j immunol. 2018;88:e12711. doi:10.1111/sji.12711
145. Borzouei S, Mohamadtaheri M, Zamani A, Behzad M. Reduced frequency and functional potency of CD49d(-) T regulatory cells in patients with newly diagnosed type 2 diabetes mellitus. Immunobiology. 2021;226:152113. doi:10.1016/j.imbio.2021.152113
146. Zhen Y, Sun L, Liu H, et al. Alterations of peripheral CD4+CD25+Foxp3+ T regulatory cells in mice with STZ-induced diabetes. Cell Mol Immunol. 2012;9:75. doi:10.1038/cmi.2011.37
147. Hui Y, Kuang L, Zhong Y, Tang Y, Xu Z, Zheng T. High glucose impairs cognitive function through inducing mitochondrial calcium overload in treg cells. iScience. 2024;27:108689. doi:10.1016/j.isci.2023.108689
148. Holl J, Kowalewski C, Zimek Z, et al. Chronic diabetic wounds and their treatment with skin substitutes. Cells. 2021;10:655. doi:10.3390/cells10030655
149. Rehak L, Giurato L, Meloni M, Panunzi A, Manti GM, Uccioli L. The immune-centric revolution in the diabetic foot: monocytes and lymphocytes role in wound healing and tissue regeneration-a narrative review. J Clin Med. 2022;11:889. doi:10.3390/jcm11030889
150. Cai Y, Ji H, Zhou X, et al. Interleukin-21 modulates balance between regulatory T cells and T-helper 17 cells in chronic hepatitis B virus infection. BMC Infect Dis. 2023;23:719. doi:10.1186/s12879-023-08723-w
151. Wu YH, Zhang QL, Mai SY, et al. Strictosamide alleviates acute lung injury via regulating T helper 17 cells, regulatory T cells, and gut microbiota. Phytomedicine. 2024;128:155490. doi:10.1016/j.phymed.2024.155490
152. Zoheir KMA, Ali NI, Ashour AE, et al. Lipoic acid improves wound healing through its immunomodulatory and anti-inflammatory effects in a diabetic mouse model. J Diabetes Metab Disord. 2025;24:56. doi:10.1007/s40200-025-01559-y
153. Kuang S, He F, Liu G, et al. CCR2-engineered mesenchymal stromal cells accelerate diabetic wound healing by restoring immunological homeostasis. Biomaterials. 2021;275:120963. doi:10.1016/j.biomaterials.2021.120963
154. Geach T. Diabetes: new TREG gene signature for T1DM. Nat Rev Endocrinol. 2016;12:125. doi:10.1038/nrendo.2016.19
155. Zhang J, Chen J, Gao C, et al. Maggot treatment promotes healing of diabetic foot ulcer wounds possibly by upregulating Treg levels. Diabetes Res Clin Pract. 2022;184:109187. doi:10.1016/j.diabres.2021.109187
156. Shi H, Yuan X, Liu G, Fan W. Identifying and validating GSTM5 as an immunogenic gene in diabetic foot ulcer using bioinformatics and machine learning. J Inflamm Res. 2023;16:6241. doi:10.2147/JIR.S442388
157. Wang S, Zhang Y, Wang Y, et al. Amphiregulin confers regulatory T cell suppressive function and tumor invasion via the EGFR/GSK-3β/Foxp3 axis. J Biol Chem. 2016;291:21085. doi:10.1074/jbc.M116.717892
158. Vaz ER, Fujimura PT, Araujo GR, et al. A short peptide that mimics the binding domain of TGF-β1 presents potent anti-inflammatory activity. PLoS One. 2015;10:e0136116. doi:10.1371/journal.pone.0136116
159. Moreau JM, Dhariwala MO, Gouirand V, et al. Regulatory T cells promote innate inflammation after skin barrier breach via TGF-β activation. Sci immunol. 2021;6. doi:10.1126/sciimmunol.abg2329
160. Minutti CM, Drube S, Blair N, et al. Epidermal growth factor receptor expression licenses type-2 helper T Cells to function in a T cell receptor-independent fashion. Immunity. 2017;47:710. doi:10.1016/j.immuni.2017.09.013
161. He S, Li Z, Xie L, et al. Biomimetic gene delivery system coupled with extracellular vesicle-encapsulated AAV for improving diabetic wound through promoting vascularization and remodeling of inflammatory microenvironment. J Nanobiotechnology. 2025;23:242. doi:10.1186/s12951-025-03261-w
162. Wang Z, Li W, Gou L, et al. Biodegradable and antioxidant DNA hydrogel as a cytokine delivery system for diabetic wound healing. Adv Healthcare Mater. 2022;11:e2200782. doi:10.1002/adhm.202200782
163. Rhodes KR, Meyer RA, Wang J, Tzeng SY, Green JJ. Biomimetic tolerogenic artificial antigen presenting cells for regulatory T cell induction. Acta Biomater. 2020;112:136. doi:10.1016/j.actbio.2020.06.004
164. Vitner EB, Bovone G, Jung WH, et al. In vivo accumulation of regulatory t cells using eliglustat-loaded cryogels. Adv Healthcare Mater. 2025;14:e2501529. doi:10.1002/adhm.202501529
165. van Loosdregt J, Vercoulen Y, Guichelaar T, et al. Regulation of treg functionality by acetylation-mediated Foxp3 protein stabilization. Blood. 2010;115:965. doi:10.1182/blood-2009-02-207118
166. Wu T, Cai Z, Niu F, et al. Lentinan confers protection against type 1 diabetes by inducing regulatory T cell in spontaneous non-obese diabetic mice. Nut Diabetes. 2023;13:4. doi:10.1038/s41387-023-00233-7
167. Savastio S, Cadario F, D’Alfonso S, et al. Vitamin D supplementation modulates ICOS+ and ICOS- regulatory T cell in siblings of children with type 1 diabetes. J Clin Endocrinol Metab. 2020;105:e4767–e4777. doi:10.1210/clinem/dgaa588
168. Qin W, Sun L, Dong M, et al. Regulatory T cells and diabetes mellitus. Hum gene ther. 2021;32:875. doi:10.1089/hum.2021.024
169. Grebinoski S, Pieklo G, Zhang Q, et al. Regulatory T cell insufficiency in autoimmune diabetes is driven by selective loss of neuropilin-1 on intraislet regulatory T cells. J Iimmunol. 2024;213:779. doi:10.4049/jimmunol.2300216
170. Rafiqi SI, Aldasouqi A, Paparodis RD, et al. Conversion of T effector cells into t regulatory cells in type 1 diabetes/latent autoimmune diabetes of adults by inhibiting eIF5A and notch pathways. ImmunoTargets Ther. 2025;14:205. doi:10.2147/ITT.S504555
171. Zhang Q, Lu W, Liang CL, et al. Chimeric Antigen Receptor (CAR) treg: a promising approach to inducing immunological tolerance. Front Immunol. 2018;9:2359. doi:10.3389/fimmu.2018.02359
172. Zhang S, Zhao X, Zhang W, Wei X, Chen XL, Wang X. Zn-DHM nanozymes regulate metabolic and immune homeostasis for early diabetic wound therapy. Bioact Mater. 2025;49:63. doi:10.1016/j.bioactmat.2025.02.041
173. Peng C, Lei P, Qi H, et al. Effect of fecal microbiota transplantation on diabetic wound healing through the IL-17A-mTOR-HIF1α signaling axis. Appl Environ Microbiol. 2025;91:e0201924. doi:10.1128/aem.02019-24
174. Hefele F, Ditsch A, Krysiak N, et al. Trauma induces interleukin-17A expression on Th17 cells and CD4+ regulatory T cells as well as platelet dysfunction. Front Immunol. 2019;10:2389. doi:10.3389/fimmu.2019.02389
175. Ryu G, Bae JS, Yoo SH, Kim EH, Mo JH. Elevated IL-17A-Secreting Regulatory T Cells in Sinonasal Tissues of Chronic Rhinosinusitis with Nasal Polyps. Inflammation. 2025;48:2745. doi:10.1007/s10753-024-02227-8
176. Luo YL, Liang LF, Gan YJ, et al. An All-in-one nanomedicine consisting of CRISPR-Cas9 and an autoantigen peptide for restoring specific immune tolerance. ACS Appl Mater Interfaces. 2020;12:48259. doi:10.1021/acsami.0c10885
177. Hunt MS, Yang SJ, Mortensen E, et al. Dual-locus, dual-HDR editing permits efficient generation of antigen-specific regulatory T cells with robust suppressive activity. Mol Ther. 2023;31:2872. doi:10.1016/j.ymthe.2023.07.016
178. Fang S, Zhang H, Liu W, et al. Analysis and validation of mitophagy-related genes in diabetic foot ulcers. J Inflamm Res. 2025;18:4367. doi:10.2147/JIR.S504001
179. Bender C, Wiedeman AE, Hu A, et al. A phase 2 randomized trial with autologous polyclonal expanded regulatory T cells in children with new-onset type 1 diabetes. Sci, trans med. 2024;16:eadn2404. doi:10.1126/scitranslmed.adn2404
180. Ou K, Hamo D, Schulze A, et al. Strong expansion of human regulatory T cells for adoptive cell therapy results in epigenetic changes which may impact their survival and function. Front Cell Develop Biol. 2021;9:751590. doi:10.3389/fcell.2021.751590
181. Price CL, Knight SC. Methylglyoxal: possible link between hyperglycaemia and immune suppression? Trends Endocrinol Metab. 2009;20:312. doi:10.1016/j.tem.2009.03.010
182. Dai T, Zhao S, Qiu Y, et al. Pt@FeMOF-PEG nanozyme-mediated reactive oxygen species scavenging and reduction of treg cells to treat surgical trauma. ACS Appl Mater Interfaces. 2026;18:12838. doi:10.1021/acsami.5c24816
183. Marek-Trzonkowska N, Myśliwiec M, Dobyszuk A, et al. Therapy of type 1 diabetes with CD4(+)CD25(high)CD127-regulatory T cells prolongs survival of pancreatic islets - results of one year follow-up. Clin Immunol. 2014;153:23. doi:10.1016/j.clim.2014.03.016
184. Lu Y, Wang Y, Ruan T, et al. Immunometabolism of tregs: mechanisms, adaptability, and therapeutic implications in diseases. Front Immunol. 2025;16:1536020. doi:10.3389/fimmu.2025.1536020
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Accelerated Wound Healing in Diabetic Rat by miRNA-185-5p and Its Anti-Inflammatory Activity
Wang KX, Zhao LL, Zheng LT, Meng LB, Jin L, Zhang LJ, Kong FL, Liang F
Diabetes, Metabolic Syndrome and Obesity 2023, 16:1657-1667
Published Date: 7 June 2023
MicroRNA-221-3p Targets THBS1 to Promote Wound Healing in Diabetes
Hu K, Liu X, Chang H, Zhang Y, Zhou H, Liu L, Zhang X, Jiao Z, Shen B, Zhang Q
Diabetes, Metabolic Syndrome and Obesity 2023, 16:2765-2777
Published Date: 11 September 2023
Effect of Diabetes on Wound Healing: A Bibliometrics and Visual Analysis
Lang X, Li L, Li Y, Feng X
Journal of Multidisciplinary Healthcare 2024, 17:1275-1289
Published Date: 20 March 2024
Diverse-Origin Exosomes Therapeutic Strategies for Diabetic Wound Healing

Wang F, Yao J, Zuo H, Jiao Y, Wu J, Meng Z
International Journal of Nanomedicine 2025, 20:7375-7402
Published Date: 12 June 2025
Exosomes in Diabetic Wound Healing: Mechanisms, Applications, and Perspectives

Yi J, Tang Q, Sun S, Xie H, Wang L, Yin X
Diabetes, Metabolic Syndrome and Obesity 2025, 18:2955-2976
Published Date: 21 August 2025