Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Exosomes in Diabetic Wound Healing: Mechanisms, Applications, and Perspectives
Authors Yi J ![]() , Tang Q, Sun S, Xie H, Wang L, Yin X
, Tang Q, Sun S, Xie H, Wang L, Yin X
Received 7 April 2025
Accepted for publication 5 August 2025
Published 21 August 2025 Volume 2025:18 Pages 2955—2976
DOI https://doi.org/10.2147/DMSO.S532885
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rebecca Baqiyyah Conway
Jialian Yi,1,2 Qisheng Tang,1,2 Shuifen Sun,1,2 Hongqing Xie,1,2 Linping Wang,1,2 Xin Yin1– 4
1The Regenerative Medicine Research Center, The Affiliated Hospital of Kunming University of Science and Technology, Kunming, People’s Republic of China; 2The Regenerative Medicine Research Center, The First People’s Hospital of Yunnan Province, Kunming, People’s Republic of China; 3The Ultrasound Medical Department, The First People’s Hospital of Yunnan Province, Kunming, People’s Republic of China; 4The NHC Key Laboratory of Healthy Birth and Birth Defect Prevention in Western China, Kunming, People’s Republic of China
Correspondence: Xin Yin, Email [email protected]
Abstract: Delayed wound healing is a major complication of diabetes, significantly impacting patients’ quality of life. Traditional treatments, such as wound debridement, dressing application, and growth factor therapy, often suffer from limited efficacy and significant side effects. In recent years, exosomes have emerged as a promising therapeutic strategy for promoting diabetic skin wound healing. Exosomes, nano-sized vesicles secreted by cells, are rich in bioactive components, including proteins, nucleic acids, and lipids, enabling them to participate in intercellular communication and modulate cellular functions. Studies have demonstrated that exosomes derived from stem cells can significantly enhance diabetic wound healing. This effect is attributed to their ability to modulate key wound healing processes, including shortening the inflammatory phase, promoting angiogenesis, facilitating cell migration and re-epithelialization, and regulating collagen remodeling. Moreover, various pre-treatment strategies, applied to either the exosome-secreting cells or the isolated exosomes themselves, can significantly augment their therapeutic efficacy. As natural nanocarriers with low immunogenicity and high targeting specificity, exosomes hold great promise in tissue repair and regenerative medicine. This review summarizes the latest research progress on exosomes in diabetic wound healing, exploring their mechanisms of action and potential clinical applications, thereby establishing a foundation for the clinical translation of exosomes in diabetic wound management.
Keywords: exosomes, diabetes, wound healing, angiogenesis, biomaterials, engineered exosomes
Graphical Abstract:

Introduction
Diabetes mellitus represents a prevalent chronic metabolic disorder. According to the International Diabetes Federation (IDF), approximately 537 million people worldwide are affected by diabetes, with a prevalence rate of 1 in every 11 individuals.1 Diabetes primarily arises from cellular resistance to insulin or insufficient insulin secretion, leading to hyperglycemia and disruption of physiological homeostasis. Among these, type 2 diabetes mellitus (T2DM) is the most common, accounting for approximately 90% of all cases. Delayed wound healing process is a major complication of T2DM, and treatment remains challenging.2 Impaired wound healing in diabetic patients stems from a multitude of factors, including compromised immune response to infection, impaired blood circulation, neuropathy, impaired protein synthesis, cellular dysfunction, and atherosclerosis.3,4 Conventional therapeutic approaches for diabetic wounds encompass wound debridement, dressing application, glycemic control combined with anti-infection measures, oxygen or other gas therapies, negative pressure wound therapy, and growth factor therapy.5–7 However, these conventional modalities possess inherent limitations. For instance, maintaining glycemic control frequently necessitates long-term insulin therapy, which can be associated with adverse effects such as lipohypertrophy, skin infections, and subcutaneous induration at the injection site. Moreover, insulin therapy can contribute to weight gain, thereby increasing the risk of cardiovascular and other complications.8,9 Improper timing and methodological choices in debridement, coupled with inter-individual variability, may lead to suboptimal outcomes.10 Consequently, there is an urgent clinical need for the development of novel technologies that promote wound healing in diabetic patients.
Recent studies have demonstrated the substantial efficacy of exosomes in promoting diabetic wound healing.11–14 Exosomes are extracellular vesicles generated by multivesicular bodies and can be isolated from various sources, including blood,15–17 breast milk,18 urine,19 tissues,20,21 and even plants.22 Serving as crucial mediators of paracrine signaling, exosomes encapsulate a diverse array of proteins, lipids, and nucleic acids. These bioactive molecules can be delivered to recipient cells, activating relevant signaling pathways23,24 or influencing target gene expression,25,26 thereby modulating gene and protein expression and affecting cellular functions. Exosomes derived from stem cells can exert regenerative effects analogous to those of their parent cells in tissue injury contexts, while circumventing the risks associated with stem cell transplantation.27 In recent years, the application of exosomes in diabetic wound healing has garnered considerable attention. Exosomes effectively promote wound healing through various mechanisms, including shortening the inflammatory phase, promoting angiogenesis, enhancing cell migration and proliferation, modulating collagen remodeling, and inhibiting scar formation.19,28 While previous reviews have discussed the general role of exosomes in wound healing, few have provided an integrated and up-to-date synthesis of natural exosomes, exosomes from pretreated parental cells, engineered exosomes, and exosome–biomaterial combinations in the specific context of diabetic wound healing. This review aims to fill these critical gaps by systematically summarizing the current understanding of exosome biology in diabetic wound healing, including their mechanisms of action, therapeutic effects, enhancement strategies, and integration with advanced biomaterials. By offering a comprehensive and mechanistically grounded review, this article aims to inform future research directions and support the development of effective exosome-based therapies for diabetic wounds.
Mechanisms Underlying Impaired Diabetic Wound Healing
Normal Wound Healing Process
Wound healing is a precisely orchestrated process comprising four overlapping phases: hemostasis, inflammation, proliferation, and remodeling.29 During hemostasis, immediately following injury, damaged blood vessels and surrounding microvessels constrict to stem blood flow. Vascular damage exposes subendothelial collagen, prompting rapid platelet adhesion. Under the influence of factors such as thrombin, platelets aggregate to form a hemostatic plug, occluding the wound. Concurrently, soluble fibrinogen in plasma is converted to fibrin by thrombin, forming a meshwork that further traps blood cells, reinforcing clot formation.30,31 In the inflammatory phase, the release of vasoactive substances and chemokines attracts inflammatory cells, including leukocytes, neutrophils, macrophages, and lymphocytes, to the wound site. These cells clear necrotic tissue, foreign debris, and bacterial products, while also releasing growth factors that prime subsequent healing phases.32,33 As necrotic tissue and bacteria are cleared, inflammation gradually resolves, marking the onset of the proliferative phase. Fibroblasts, endothelial cells, and keratinocytes begin to proliferate and migrate, generating granulation tissue. Fibroblasts proliferate extensively, producing extracellular matrix components such as collagen fibers and fibronectin.34 Endothelial cells form new blood vessels to provide essential nutrients to the wound. Keratinocytes, through re-epithelialization, cover the wound with a new epidermal layer, further promoting healing.35 In the remodeling phase, after the wound is filled with granulation tissue, the extracellular matrix undergoes remodeling by matrix metalloproteinases. Type III collagen is gradually replaced by type I collagen, forming a more robust scar tissue, completing the healing process.28,36–39
Mechanisms of Impaired Wound Healing in Diabetic Skin
While normal skin repair is rapid, several factors in diabetic patients, including the accumulation of advanced glycation end products (AGEs), vasculopathy, nerve damage, and aberrant inflammatory responses, contribute to delayed wound healing and the potential development of chronic ulcers (Figure 1).
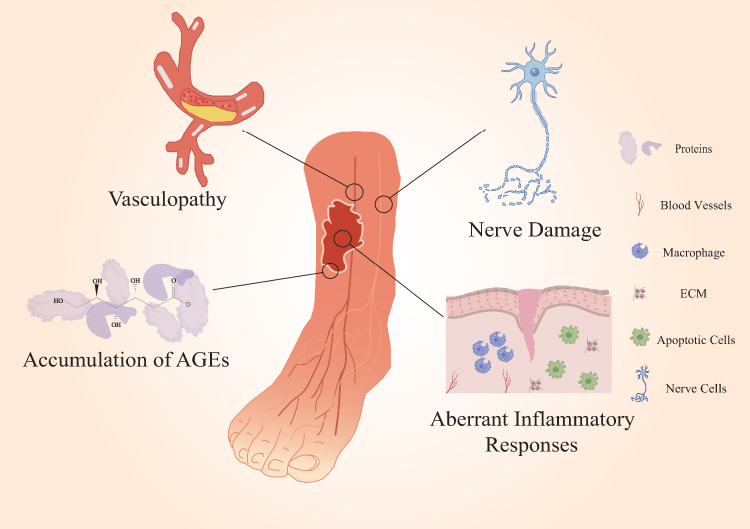 |
Figure 1 Key mechanisms impairing wound healing in diabetic skin. The schematic illustrates how hyperglycemia disrupts normal wound healing processes. Key factors such as hyperglycemia, chronic inflammation, angiopathy, and neuropathy create a hostile environment for repair. These pathologies disrupt essential cellular processes, impair immune responses, and stall the normal progression through the inflammatory, proliferative, and remodeling phases of healing. This cascade ultimately leads to the formation of chronic, non-healing wounds, a common complication of diabetes. |
Advanced Glycation End Products (AGEs)
In a hyperglycemic environment, excess glucose facilitates the non-enzymatic glycation of proteins and lipids, leading to the formation of Advanced Glycation End Products (AGEs).40 The accumulation of AGEs is a well-documented hallmark of diabetes and a major contributor to impaired wound healing due to their detrimental effects on cellular and extracellular matrix (ECM) homeostasis. Pathologically, AGEs exert their effects through multiple mechanisms: They perpetuate chronic inflammation by binding to the receptor for AGEs on immune cells, such as macrophages, promoting a sustained pro-inflammatory M1 phenotype and prolonging the inflammatory phase of wound healing. Furthermore, the interaction of AGEs with RAGE generates significant oxidative stress, leading to a surge in reactive oxygen species (ROS) that inflicts further cellular damage and inhibits the function of critical enzymes and growth factors necessary for effective repair.41–45 Collectively, these interwoven pathological processes create a hostile and non-permissive microenvironment that severely impedes the normal healing cascade.
Microvascular Disease and Endothelial Dysfunction
The dermal layer of the skin is richly vascularized with a dense capillary network, supplying nutrients and removing metabolic waste. In diabetic patients, persistent hyperglycemia increases the hemodynamic burden. Uncontrolled blood glucose leads to thickening of the microvascular basement membrane, narrowing or even occluding the vascular lumen, impeding normal blood flow.46 This ultimately compromises blood supply to skin tissues, affecting skin health. Due to impaired peripheral circulation, injured skin areas receive insufficient nutrients, leading to slow or stalled wound healing. Angiogenesis is a complex process precisely regulated by various cells and factors.47 During wound repair, endothelial progenitor cells and endothelial cells migrate and proliferate towards the injured site under the influence of angiogenic factors or stimuli. Through chemotaxis, they participate in new blood vessel formation and vascular network reconstruction, improving blood supply to the damaged area and accelerating wound healing.48 However, in diabetic wounds, the dysregulation of glucose and lipid metabolism as well as oxidative stress impairs the proliferation and migration of endothelial cells and even induces their apoptosis, directly impeding neovascularization.49 Moreover, the dysregulated glucose and lipid metabolism decreases the transcriptional activity of hypoxia-inducible factors, thereby weakening the cellular responses to hypoxia and subsequently reducing the activity of downstream proteins and various growth factors, which further hampers angiogenesis.50 Reduced endothelial cell activity and insufficient growth factor result in inadequate neovascularization in diabetic wounds, impeding healing.
Neuropathy
Diabetic neuropathy is a common and debilitating complication that profoundly impairs the wound healing process. In diabetic patients, persistent hyperglycemia induces metabolic and microvascular dysfunctions that lead to nerve damage, including reduced nerve blood flow and endoneurial hypoxia.51–53 This neuropathy manifests in three critical ways that compromise skin integrity and repair. First, damage to sensory nerves results in a loss of protective sensation (LOPS), diminishing the patient’s ability to perceive pain, pressure, and temperature. Consequently, minor mechanical traumas often go unnoticed, allowing them to escalate into severe, non-healing ulcerative wounds.54 Second, autonomic neuropathy disrupts sudoral (sweat gland) function, leading to anhidrosis. The resulting dry, brittle, and inelastic skin is prone to fissures and cracks, which serve as entry portals for pathogens and significantly increase the risk of infection, further compromising the skin’s barrier function.54,55 Finally, neuropathy creates a deficiency in crucial cutaneous neuropeptides and neurotrophic factors, such as Substance P (SP) and Nerve Growth Factor (NGF).56 These molecules are essential for orchestrating the healing cascade, as they promote keratinocyte and fibroblast proliferation, stimulate angiogenesis, and modulate local inflammation. Their depletion in diabetic skin impairs these vital signaling pathways, directly hindering re-epithelialization and neovascularization and contributing significantly to the wound’s chronic, non-healing state.57,58
Aberrant Inflammatory Response
Inflammation is an immune response initiated by the body in response to tissue injury or infection, typically characterized by redness, swelling, heat, pain, and loss of function.59,60 Normally, inflammation is a beneficial defense mechanism that helps clear pathogens, repair damaged tissue, and restore homeostasis. However, in diabetic wounds, hyperglycemia leads to persistent accumulation and infiltration of immune cells, accelerating the release of pro-inflammatory cytokines and chemokines,61–64 thereby exacerbating inflammation and slowing wound healing. Diabetic wounds exhibit a prolonged inflammatory phase, leading to a state of chronic inflammation. In the later stages of inflammation, hyperglycemia also hinders the transition of pro-inflammatory M1 macrophages to reparative M2 macrophages,65,66 perpetuating the inflammatory state. Furthermore, excessive neutrophil extracellular traps (NETs) released by neutrophils within the wound are capable of activating the NLRP3 inflammasome in macrophages and inducing the release of IL-1β. This, in turn, prolongs the residence time of inflammatory cytokines in diabetic wounds, further extends the inflammatory phase, and inhibits the formation of granulation tissue.67,68
Introduction of Exosomes
Origin, Definition, Biogenesis, and Isolation of Exosomes
In 1967, Wolf first identified a class of substances in plasma and named them “platelet-dust”,69 which were subsequently confirmed to be extracellular vesicles. In the 1980s, researchers elucidated the mechanism of extracellular vesicle secretion by reticulocytes of rats and sheep: inward budding of endosomes leads to the formation of multivesicular bodies (MVBs), which then fuse with the plasma membrane, releasing their contents into the extracellular environment.70,71 In 1987, Rose Johnstone et al, first used the term “exosome” to describe these cell-derived vesicles.72 Exosomes are a subtype of extracellular vesicles, ranging from 30 to 150 nm in diameter, with a density of 1.13–1.19 g/mL. They are released into the extracellular environment following fusion of MVBs with the plasma membrane.73,74 Under electron microscopy, exosomes exhibit a characteristic “cup-shaped” or “concave disc-shaped” morphology.75
Exosome biogenesis is precisely regulated by various proteins and cytokines, primarily involving the following steps: (1) inward budding of MVBs to form intraluminal vesicles (ILVs); (2) trafficking of various proteins, lipids, and nucleic acids to ILVs; (3) migration of MVBs to the plasma membrane; and (4) fusion of MVBs with the plasma membrane, releasing exosomes.76,77 ILV formation occurs via two main pathways: the endosomal sorting complex required for transport (ESCRT)-dependent and ESCRT-independent pathways, depending on the mechanisms of different exosome subpopulations.78,79
Exosome extraction and purification are crucial for exosome research, thus selecting an appropriate extraction technique is essential for obtaining high-quality exosomes. With technological advancements, exosome isolation methods have become increasingly diverse, with different methods significantly affecting exosome concentration, purity, and yield. Currently, commonly used isolation methods include ultracentrifugation,80,81 precipitation,82 immunoaffinity capture83 size-exclusion chromatography,20,66 and ultrafiltration.20 Although a universally accepted gold standard has not yet been established, ultracentrifugation remains the most widely used and recognized method, the comparison of different methods is shown in Table 1.
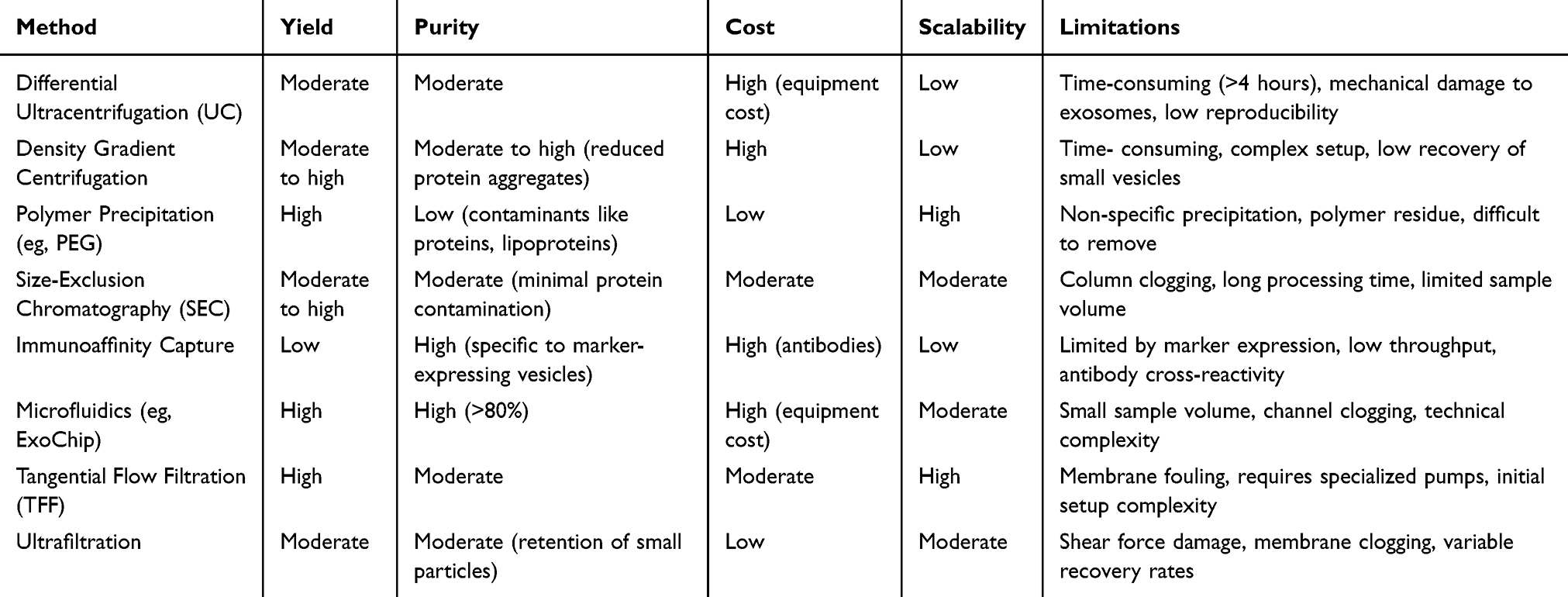 |
Table 1 Comparison of Exosome Isolation Methods |
Exosome Cargo
Exosomes contain a diverse array of nucleic acids, proteins, and lipids, the composition of which depends on the cell type and extracellular environment.84 Under specific conditions, they reflect the characteristics and physiological state of the donor cells, playing an important role in intercellular communication. Nucleic acids within exosomes include messenger RNA (mRNA), microRNA (miRNA), long non-coding RNA (lncRNA), circular RNA (circRNA), and DNA.85 These nucleic acids carry genetic information and are delivered to recipient cells via endocytosis, subsequently influencing protein synthesis. Proteins contained within exosomes play critical roles in their formation, transport, and interaction with recipient cells. These proteins encompass the tetraspanin family (eg, CD9, CD62, CD63, CD81), ESCRT proteins (eg, Alix, TSG101), integrins, heat shock proteins (eg, HSP70, HSP90), and actin.15,26,86–88 Among them, TSG101, HSP70, Alix, CD81, and CD63 are widely used as exosome marker proteins. The lipid composition of exosomes is similar to that of the cell membrane, mainly including sphingomyelin, cholesterol, glycerophospholipids, and ceramide.89,90 These lipids play important roles in maintaining exosome structure, protecting cargo, and participating in intercellular communication. In recent years, mitochondrial components, such as mitochondrial DNA (mtDNA), mtDNA-encoded mRNA, and mitochondrial-related proteins, have been detected in exosomes.91 Studies by Anisha et al have shown that cell-derived vesicles from human brain endothelial cells can transfer mitochondria under ischemic conditions and enhance endothelial cell survival.92
It should be noted that the influence of donor characteristics significantly impacts the cargo of exosomes, particularly miRNA profiles.93 Studies suggest that donor age can alter the exosomal miRNA signature, with exosomes from younger individuals potentially carrying miRNAs that promote cell proliferation, while those from older donors might contain miRNAs associated with senescence.94 Donor sex also appears to play a role in shaping exosomal miRNA content. Certain miRNAs have been found to be differentially expressed in exosomes based on the sex of the donor, highlighting potential sex-specific regulatory mechanisms mediated by these vesicles.95 Furthermore, diabetic status significantly affects exosomal miRNA profiles. Comparisons between diabetic and non-diabetic individuals have revealed distinct miRNA signatures in circulating exosomes.96 For example, specific miRNAs have been found to be upregulated in diabetic patients, suggesting that these alterations in exosomal miRNA cargo could contribute to the pathogenesis or progression of diabetes-related complications. These variations highlight the importance of considering donor profiles in studies utilizing exosomes for therapeutics and underscore the need for further research to fully elucidate the mechanisms and functional consequences of these alterations. Understanding how these factors shape exosomal content could pave the way for more personalized and effective applications of exosome-based technologies.
Biological Functions of Exosomes
Exosomes not only reflect the characteristics and physiological state of donor cells but also play a crucial role in intercellular communication. Donor cells release exosomes into the extracellular environment, and these exosomes are subsequently transported to recipient cells via body fluids. After exosomes fuse with the plasma membrane of recipient cells, they release their contents and interact with receptors on the recipient cell surface, thereby initiating downstream signaling cascades and triggering physiological or pathological responses in recipient cells.89,97 Studies have demonstrated that exosomes are capable of modulating the immune system’s function. Exosomes derived from bone marrow mesenchymal stem cells (BMSCs) affect the maturation and activation of dendritic cells (DCs) and reduce the release of inflammatory factors.98 Exosomes secreted by immune cells carry immunomodulatory molecules, such as miRNA, cytokines, and chemokines, participating in the activation, proliferation, and differentiation of immune cells. Exosomes derived from M2 macrophages (M2-Exos) are able to induce the polarization of pro-inflammatory M1 macrophages into anti-inflammatory M2 macrophages.66,99 Exosomes derived from regulatory T cells (Treg-exosomes) exert significant influence on peripheral blood mononuclear cells (PBMCs), modulating their phenotype and suppressing the production of the pro-inflammatory cytokine interleukin-6 (IL-6).81 Moreover, Treg-exosomes can induce dendritic cells (DCs) to release anti-inflammatory cytokines.100
Application of Exosomes in Wound Healing
Exosomes exhibit significant therapeutic potential in diabetic wound healing. Natural exosomes promote diabetic wound healing by modulating inflammation, promoting angiogenesis, restoring cellular function, and regulating the extracellular matrix (ECM). Furthermore, exosomes derived from parental cells pretreated with pharmacological, physical, or biological stimuli demonstrate enhanced therapeutic efficacy. Engineered exosomes, generated through endogenous or exogenous modification, enable precise delivery of specific therapeutic molecules. Moreover, combining exosomes with biomaterials such as hydrogels, nanofibers, or other dressings significantly improves exosome delivery efficiency and therapeutic outcomes at the wound site (Figure 2). The research progress of natural or modified exosomes in treating diabetic wounds is summarized in Table 2.
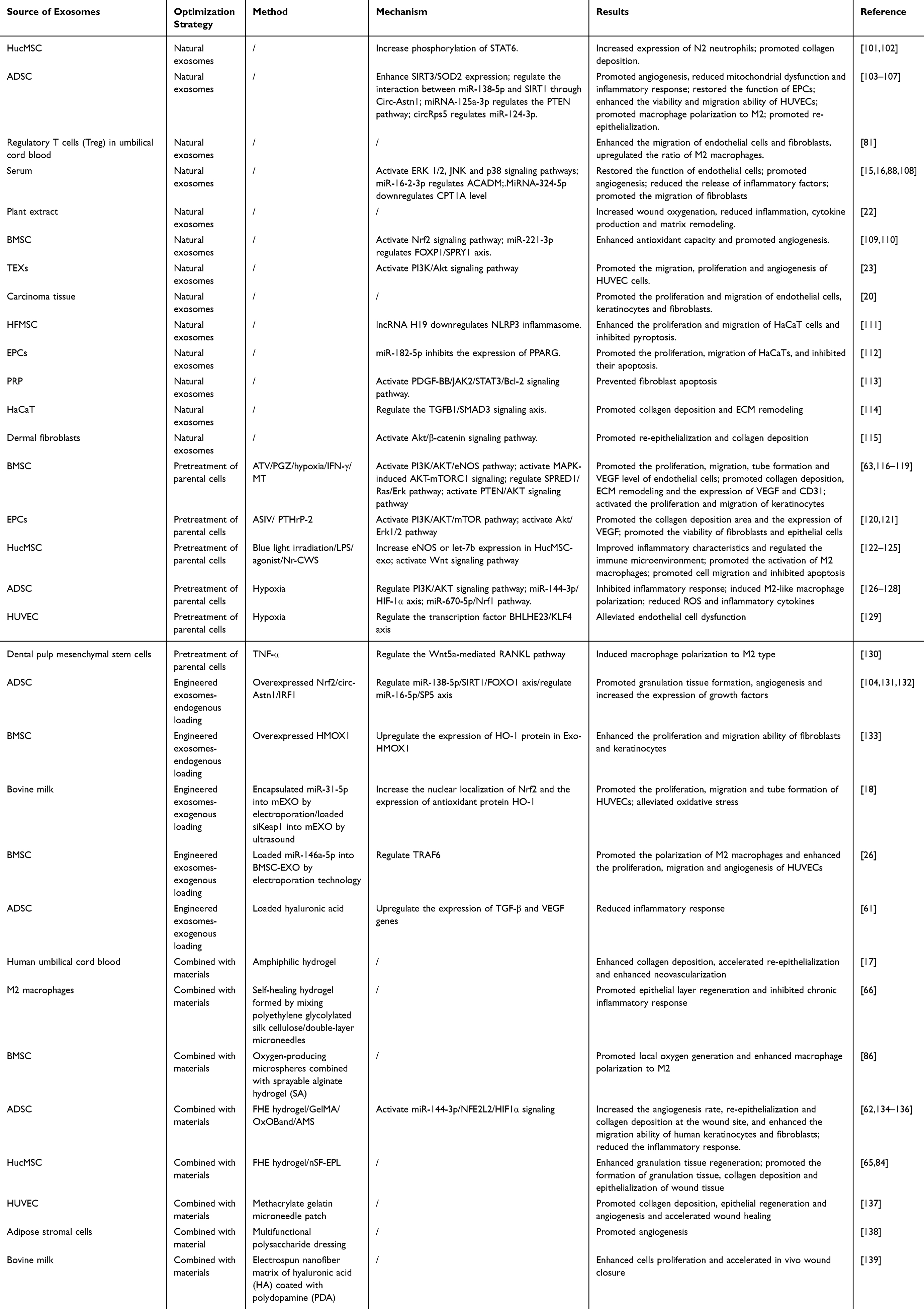 |
Table 2 Applications of Exosomes in the Treatment of Diabetic Skin Wounds |
 |
Figure 2 Therapeutic Applications of Exosomes in Wound Healing. The schematic illustrates the multifaceted roles of exosomes in promoting tissue regeneration. Derived from various sources, natural or modified exosomes carry therapeutic cargo to target cells in the wound microenvironment. Their therapeutic actions include enhancing angiogenesis, reducing inflammation, stimulating fibroblast proliferation and migration, promoting collagen deposition and extracellular matrix (ECM) remodeling, accelerating re-epithelialization, and reducing scarring. These coordinated effects accelerate wound healing. |
Role of Natural Exosomes in Wound Healing
Natural exosomes, derived from various sources, promote wound healing through multiple mechanisms, including inhibiting inflammation, promoting angiogenesis, restoring cellular function, and modulating the ECM.
Anti-Inflammatory Effects of Exosomes in Diabetic Wounds
Chronic inflammation triggered by diabetes is a critical factor contributing to impaired wound healing. Studies have shown that exosomes from various sources play a pro-healing role during the inflammatory phase of diabetic wounds.140 These exosomes are derived from a wide range of sources, including mesenchymal stem cells, immune cells, biological fluids, and plant extracts. Exosomes secreted by human umbilical cord mesenchymal stem cells (HucMSC-Exos) significantly increase the expression of N2 neutrophils, thereby shortening the inflammatory phase and increasing the release of angiogenic factors.101 Exosomes derived from adipose-derived mesenchymal stem cells (ADSC-Exos) act on human umbilical vein endothelial cells (HUVECs), increasing the expression of SIRT3, which enhances superoxide dismutase 2 (SOD2) activity and reduces the levels of inflammatory cytokines.103 Treg-Exos derived from umbilical cord blood target PBMCs, promoting their polarization to anti-inflammatory M2 macrophages and significantly reducing inflammatory cytokine secretion,81 an effect of significant importance in alleviating chronic inflammation in diabetic patients. Serum-derived exosomes carrying miRNA-324-5p counteract diabetes-related inflammatory responses by negatively regulating CPT1A gene expression and reducing inflammatory cytokine release.108 Plant-derived exosomes also exhibit anti-inflammatory effects, effectively reducing IL-6 levels in diabetic wounds, further highlighting the potential of exosomes in modulating inflammation in diabetic wounds.22
Role of Exosomes in Angiogenesis in Diabetic Wounds
Exosomes play a crucial role in promoting angiogenesis during diabetic wound healing. Serum exosomes laden with angiopoietin-like 6 (ANGPTL6) have the capacity to activate the ERK 1/2, JNK, and p38 signaling cascades, thereby facilitating the restoration of endothelial cell functionality.15 Furthermore, miRNA-16-2-3p in serum exosomes promotes endothelial cell repair by targeting ACADM.16 ADSC-Exos containing circ-Astn1 restore the function of endothelial progenitor cells (EPCs) by mediating the interaction between miR-138-5p and SIRT.104 miRNA-125a-3p in ADSC-Exos significantly enhances HUVEC viability and migration by inhibiting PTEN gene expression.105 Bone marrow mesenchymal stem cell-derived exosomes (BMSC-Exos) also demonstrate pro-angiogenic potential. Specifically, the circ-ITCH carried by them is capable of recruiting TAF15 protein and activating the Nrf2 signaling pathway, which consequently enhances both the cellular antioxidant capacity and the process of angiogenesis.109 miRNA-221-3p in BMSC-Exos promotes angiogenesis by regulating the FOXP1/SPRY1 axis, demonstrating its significant potential in treating diabetic wounds.110
Restoration of Cell Function Related to Diabetic Wound Healing by Exosomes
Exosomes play a key role in restoring the function of cells involved in skin wound healing, including endothelial cells, fibroblasts, keratinocytes (HaCaT), and macrophages. Previous studies have demonstrated that exosomes secreted by 4T1 mouse mammary cancer cells encompass miRNA-126a-3p. This miRNA has the ability to activate the PI3K/Akt signaling pathway, which in turn promotes cell migration, proliferation, and angiogenesis. Subsequently, this cascade of events leads to a significant acceleration of chronic wound healing.23 Similarly, exosomes from oral squamous cell carcinoma tissue show similar effects, promoting the proliferation and migration of endothelial cells, keratinocytes, and fibroblasts.20 Regarding cell protection, exosomes secreted by hair follicle mesenchymal stem cells (HFMSC-Exos) containing H19, a long non-coding RNA (lncRNA), which enhances HaCaT proliferation and migration and effectively inhibit their apoptosis by reversing NLRP3 inflammasome activation.111 Exosomes derived from endothelial progenitor cells (EPC-Exos) promote HaCaT proliferation and migration and inhibit their apoptosis by increasing miRNA-182-5p expression.112 Platelet-rich plasma-derived exosomes (PRP-Exos) prevent ROS-dependent apoptosis under high glucose conditions by activating the PDGF-BB/JAK2/STAT3/Bcl-2 signaling pathway,113 a mechanism crucial for protecting the survival of wound healing-related cells, such as fibroblasts. CircRps5 carried by ADSC-Exos promotes macrophage polarization towards the anti-inflammatory M2 phenotype by regulating miRNA-124-3p, further accelerating the proliferation and migration of wound healing-related cells.106
Regulation of the Extracellular Matrix by Exosomes
The ECM, composed of macromolecules such as collagen, elastin, proteoglycans, and glycosaminoglycans, forms a complex network structure surrounding cells. In diabetic patients, regulating the expression of ECM-related proteins is important for promoting wound healing. Recent studies indicate that metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) within exosomes derived from HaCaT cells promotes collagen deposition and extracellular matrix (ECM) remodeling, which are crucial for skin wound healing.114 Furthermore, HucMSC-Exos also have the ability to promote collagen deposition, indicating their potential application in promoting tissue repair and regeneration.102 ADSC-Exos are rich in matrix metalloproteinases MMP1 and MMP3, as well as type I and III collagen, the expression of these molecules promotes skin re-epithelialization.107 Exosomes from dermal fibroblasts promote skin re-epithelialization and collagen deposition by activating the Akt/β-catenin signaling pathway, further emphasizing the role of exosomes in promoting diabetic wound healing.115 Serum-derived exosomes highly express ECM proteins such as PCNA, Ki67, collagen α, and fibronectin, accelerating wound healing in diabetic mice by promoting the migration of mouse embryonic fibroblasts (NIH/3T3).88
These researches demonstrate that natural exosomes can promote chronic wound healing from multiple aspects, showing great clinical application potential in the treatment of diabetic-related chronic non-healing wounds.
Enhanced Diabetic Wound Healing by Exosomes from Pretreated Parental Cells
While natural exosomes offer several advantages in treating diabetic wound healing, challenges remain regarding their stability, targeting specificity, and widespread clinical applicability. To address these limitations, researchers are exploring various strategies to enhance the therapeutic potential of exosomes. The composition, content, and bioactivity of exosomes are closely related to the physiological state and microenvironment of their parental cells, suggesting that modulating parental cell status can optimize exosome function. For example, altering the culture conditions of parental cells or using genetic engineering to enhance the expression of certain beneficial components within exosomes can improve therapeutic outcomes. Accordingly, researchers employ parental cell pretreatment methods to enhance the function of secreted exosomes. These pretreatment methods include pharmacological pretreatment, physical factor intervention, and biological factor intervention. Exosomes secreted by pretreated parental cells can more precisely meet the specific requirements of diabetic wound healing, improving therapeutic efficacy.
Pharmacological Intervention
Direct drug administration during chronic wound healing may induce side effects, toxic reactions, and allergic reactions, thereby possibly impeding therapeutic efficacy and patient recovery. Therefore, using exosomes derived from pharmacologically pretreated parental cells to treat chronic wounds represents an effective strategy to avoid these adverse reactions and enhance therapeutic outcomes. Pharmacological pretreatment often involves treating parental cells with anti-inflammatory or pro-angiogenic drugs to optimize the secreted exosomes, making them more effective in promoting wound healing. For example, exosomes derived from BMSCs pretreated with atorvastatin (ATV) show significant effects on promoting full-thickness skin defect regeneration in streptozotocin (STZ)-induced diabetic rats. These ATV-pretreated exosomes (ATV-Exos) enhance endothelial cell proliferation, migration, and angiogenesis and increase VEGF levels by upregulating miRNA-221-3p and activating the AKT/eNOS signaling pathway, thereby promoting diabetic skin defect healing.116 In another study, BMSCs pretreated with pioglitazone (PGZ-Exos) accelerated diabetic wound healing by activating the PI3K/AKT/eNOS pathway, promoting collagen deposition, ECM remodeling, and VEGF and CD31 expression.117 Astragaloside IV (ASIV) treatment not only enhances EPC function (eg, proliferation) but also increases exosome secretion. Exosomes secreted by ASIV-pretreated EPCs (ASIV-Exos), by activating the PI3K/AKT/mTOR pathway, significantly improve the chronic wound healing rate, collagen deposition area, bromodeoxyuridine uptake, and VEGF expression in diabetic rats, while also increasing the number of CD31- and α-SMA-positive cells.120
Physical Factor Intervention
Physical factor interventions, such as hypoxia or mechanical stress, are also used to promote the expression of beneficial factors in exosomes. Low-intensity pulsed ultrasound (LIPUS) is a technique that delivers specific physical energy using low-intensity (less than 3 W/cm²) pulsed waves. When LIPUS is used to apply local mechanical stimulation to cells, it can significantly promote intracellular signal transduction, enhancing cell proliferation and increasing exosome secretion,141 while also accelerating the uptake efficiency of exosomes by recipient cells.87 HucMSCs exposed to blue light produce exosomes rich in eNOS. In vitro experiments show that these HucMSC-derived exosomes (HucMSC-Exos/eNOS) significantly improved the biological function of endothelial cells treated with high glucose, reduce inflammatory cytokine expression and cell apoptosis. In vivo studies demonstrated that these exosomes significantly accelerated wound healing in diabetic mice. This acceleration was associated with enhanced neovascularization and matrix remodeling, concurrent with an improved inflammatory profile at the wound site. These exosomes also modulated the local immune microenvironment, thereby promoting tissue repair.122 Under hypoxic conditions, ADSC-derived exosomes promote diabetic wound healing and attenuate inflammatory responses via modulation of the PI3K/AKT signaling pathway.126 ADSC-Exos also inhibit high glucose-induced endothelial cell damage by delivering circ-Snhg11 and induce M2 macrophage polarization through the miRNA-144-3p/HIF-1α axis.127 Furthermore, ADSC-Exos enriched with circ-Erbb2ip promote angiogenesis and reduce ROS and inflammatory cytokine expression by targeting the miRNA-670-5p/Nrf1 pathway, thereby promoting wound healing.128 BMSCs cultured under hypoxic conditions secrete exosomes rich in miRNA-4645-5p, which accelerates diabetic wound healing by promoting keratinocyte proliferation and migration through activation of the MAPK-AKT-mTORC1 signaling axis.118 HUVECs cultured under hypoxic conditions produce exosomes with high expression of lncHAR1B, which interacts with the transcription factor BHLHE23/KLF4 axis to promote endothelial cell function recovery and promote the M1-to-M2 transition of macrophages under high glucose conditions.129 These research findings demonstrate the potential of physical factor intervention in promoting exosome function for the treatment of diabetic wounds.
Biological Factor Intervention
Biological factors, such as the addition of cytokines and growth factors, can activate or enhance specific signaling pathways, thereby optimizing the content of proteins or miRNAs within exosomes. This method is particularly effective in enhancing the therapeutic efficacy of exosomes in specific diseases, especially in promoting skin wound healing. For example, IL-1β, IL-6, interferon-γ (IFN-γ), and TNF-α have been shown to effectively enhance exosome function.130,142 Exosomes derived from TNF-α-pretreated dental pulp mesenchymal stem cells have been shown to promote macrophage polarization towards the M2 phenotype, a process that plays a crucial role in resolving inflammation and accelerating wound healing.130 BMSC-secreted exosomes pretreated with IFN-γ are enriched in miR-126-3p, which significantly enhances wound healing in diabetic mice. This effect is mediated by the promotion of HUVEC proliferation, migration, and angiogenesis through the SPRED1/Ras/Erk signaling pathway.119 Similarly, exosomes derived from HucMSCs pretreated with lipopolysaccharide (LPS) promote M2 macrophage activation and alleviate inflammatory responses, promoting diabetic skin wound healing by upregulating the expression of anti-inflammatory cytokines through high expression of let-7b.123 After application of the Wnt signaling pathway agonist CHIR99021, the yield of HucMSC-derived exosomes (CHIR-Exos) increased 1.5-fold. In vitro experiments demonstrated that these exosomes promote cell migration, inhibit apoptosis, and alleviate cell cycle arrest, thereby effectively facilitating wound healing. In vivo studies further revealed that CHIR-Exos significantly accelerate wound healing in diabetic mice by enhancing local collagen deposition, promoting angiogenesis, and reducing chronic inflammation.124 Exosomes produced by HucMSCs pretreated with Nocardia rubra cell wall skeleton (Nr-CWS-Exos) promoted endothelial cell proliferation, migration, and angiogenesis.125 Furthermore, in vitro and in vivo studies observed that exosomes produced by parathyroid hormone-related peptide (PTHrP-2)-stimulated endothelial cells promote fibroblast and epithelial cell viability.121 Exosomes secreted by melatonin (MT)-pretreated BMSCs (MT-Exos) promote M2 macrophage polarization and inhibit M1 macrophage polarization by activating the PTEN/AKT signaling pathway, reducing inflammatory responses and thereby promoting diabetic wound healing.63
Enhanced Diabetic Wound Healing by Engineered Exosomes
Engineered exosomes can be primarily classified into two categories: endogenous modification and exogenous modification.143,144 These two techniques have their own unique methods and applications, making exosomes an efficient molecular delivery system. Endogenous modification involves bioengineering the parental cells that produce exosomes, enabling these cells to secrete engineered exosomes containing specific target molecules. The advantage of this method is that it can directly influence the exosome production process, ensuring that these target molecules are effectively packaged into exosomes by introducing specific RNA or proteins. Endogenous modification not only improves the encapsulation efficiency of molecules but also enhances the targeting specificity and therapeutic potential of exosomes. Exogenous modification encompasses the process of loading therapeutic molecules into purified exosomes following isolation. This approach utilizes various techniques, including co-incubation, sonication, electroporation, liposome fusion, and chemical conjugation. A key advantage of exogenous modification lies in its capacity for precise control over the exosome cargo without necessitating alterations to the parental cells. This enables flexible adjustment of the types and quantities of molecules encapsulated within the exosomes. Both approaches possess distinct advantages and are suitable for specific applications. Endogenous modification is suitable for situations requiring long-term or continuous secretion of specific molecules, while exogenous modification is beneficial for rapid and precise adjustment of exosome cargo.
Endogenous Modification of Exosomes
This approach utilizes the cell’s own production mechanisms to load and secrete molecules with specific therapeutic functions. For example, in a study by Xue Li et al, Nrf2 was overexpressed in adipose-derived stem cells (ADSCs). Subsequently, exosomes secreted by these genetically modified ADSCs were isolated and their therapeutic efficacy was compared to that of exosomes derived from wild-type ADSCs. The results demonstrated that the engineered exosomes exhibited enhanced efficacy in promoting granulation tissue formation and angiogenesis, increasing the expression levels of growth factors, and concurrently mitigating inflammation and oxidative stress.131 Bomin Cheng et al overexpressed the HMOX1 gene in bone marrow mesenchymal stem cells and extracted exosomes rich in heme oxygenase-1 (HO-1) (Exos-HMOX1). Experiments showed that compared with ordinary exosomes, Exos-HMOX1 significantly enhanced the proliferation and migration of fibroblasts and keratinocytes, as well as the angiogenic capacity of HUVECs. Injection of Exos-HMOX1 into a diabetic mouse wound model resulted in faster re-epithelialization, collagen deposition, angiogenesis, and wound closure compared with the control group.133 Furthermore, exosomes produced by adipose mesenchymal stem cells overexpressing circ-Astn1 can enhance wound healing in diabetic mouse models through the miRNA-138-5p/SIRT1/FOXO1 axis indicating that exosomes can influence complex cell signaling pathways and improve therapeutic efficacy through regulation by specific molecular axes.104 Another study has demonstrated that exosomes derived from rat adipose stem cells with overexpressed interferon regulatory factor 1 (IRF1) can effectively promote wound healing in diabetic rats. This therapeutic effect is mediated by the regulation of the miRNA-16-5p/SP5 axis.132 These studies demonstrate the application potential of endogenously modified exosomes in precision medicine.
Exogenous Modification of Exosomes
Researchers have developed various methods to directly encapsulate specific therapeutic molecules into already purified exosomes, thereby enhancing their therapeutic effects. By encapsulating miRNA-31-5p into milk exosomes using electroporation, the modified exosomes improved endothelial cell function and promoted angiogenesis. In vivo experiments further demonstrated that they accelerated wound healing in a diabetic mouse wound model.145 Xiang et al used sonication to load siRNA-Keap1 (siKeap1) into M-Exos. In vivo experiments showed that M-Exos-siKeap1 could promote HUVEC proliferation, migration and alleviate oxidative stress. In a mouse diabetic wound model, injection of M-Exos-siKeap1 significantly accelerated wound healing and enhanced collagen and neovascularization.18 Another study showed that using electroporation to load miRNA-146a-5p into BMSC-Exos (Exos-miRNA-146a) resulted in Exos-miRNA-146a not only promoting M2 macrophage polarization but also enhancing HUVEC proliferation, migration, and angiogenesis by regulating TRAF6 expression, improving refractory diabetic wound healing.26 In a diabetic rat model, exosomes derived from adipose stem cells and loaded with hyaluronic acid (HA+Exos) demonstrated the capacity to upregulate the expression of TGF-β and VEGF while simultaneously downregulating the expression of TNF-α and IL-1β and attenuating neutrophil infiltration. These findings collectively suggest the potential of HA+Exos in attenuating inflammation and promoting the healing of diabetic wounds.61 These studies indicate that exogenously modified exosomes enhance their efficiency in disease treatment by precisely loading specific therapeutic molecules.
Enhanced Diabetic Wound Healing by Combining Exosomes with Materials
In recent years, with advances in biomedical engineering and materials science, the application of biomaterials in disease treatment has garnered increasing attention. These biomaterials, whether synthetic or naturally derived, demonstrate substantial potential in the treatment of chronic wounds.146 Particularly, therapeutic strategies combining multifunctional biomaterials with exosomes have shown remarkable efficacy in chronic wound management.147 This combination leverages the unique properties of biomaterials, such as biocompatibility and plasticity, synergistically with the therapeutic effects of exosomes, creating a therapeutic synergy. Biomaterials can create a favorable microenvironment for exosomes, enhancing their in vivo stability. Additionally, biomaterials can efficiently deliver and retain exosomes at the wound site, significantly improving therapeutic efficiency.148,149 Therefore, combined therapies using biomaterials and exosomes offer more effective treatment avenues for refractory diseases such as chronic wounds.150
Combination of Exosomes with Hydrogels
The combination of exosomes and hydrogels holds considerable promise for treating chronic wounds, particularly those associated with diabetes. Hydrogels, due to their non-toxicity, hemostatic, antibacterial, highly adhesive, and excellent biocompatible properties, are widely used in the medical field. Combining these properties with the bioactivity of exosomes can address issues such as exosome degradation, poor flowability, and difficulty in accumulating at specific locations.134,151,152 For example, encapsulating umbilical cord blood-derived exosomes (UCB-Exos) within hydrogels, compared to using hydrogels alone, accelerates the healing process, promotes collagen deposition, and drives re-epithelialization and neovascularization.17 Furthermore, combining M2-Exos with oxidized hyaluronic acid and mixing with polyethylene glycolated silk fibroin forms a self-healing hydrogel that, in in vitro experiments, demonstrates the ability to modulate cytokine levels, stimulate fibroblast and keratinocyte interaction and migration, and promote angiogenesis and collagen deposition, thereby promoting epidermal regeneration.66 A sprayable alginate hydrogel (SA) dressing combined with oxygen-generating microspheres and BMSC-Exos has shown efficacy in promoting local oxygen generation, inducing M2 macrophage polarization, and promoting cell proliferation in diabetic wounds.86 Combining ADSC-Exos with a hydrogel composed of Pluronic F127 (PF127), oxidized hyaluronic acid (OHA), and an antimicrobial peptide (EPL) (FHE hydrogel) significantly improved wound closure rate, angiogenesis rate, re-epithelialization, and collagen deposition at the wound site in in vivo experiments, demonstrating healing effects unattainable with exosomes or FHE hydrogel alone.134 In a diabetic rat model, combined treatment with PF127 hydrogel and HucMSC-exos was more effective than using either hydrogel or exosomes alone. The combined therapy increased the expression of CD31 and Ki67, enhanced granulation tissue regeneration, and upregulated VEGF and TGFβ-1 expression, thereby effectively promoting wound healing.84 Embedding hypoxia-pretreated ADSC-Exos in methacrylated gelatin (GelMA) and applying them to diabetic mouse wounds also significantly improved wound closure rates.135 These studies indicate that optimizing the exosomal microenvironment through hydrogels can further enhance their therapeutic effects.
Combination of Exosomes with Nanofibers
Nanofiber technology, characterized by its diminutive fiber diameter, substantial surface area closely resembling the natural tissue extracellular matrix (ECM), and inherent capacity for facilitating oxygen and nutrient exchange, is widely regarded as a promising candidate for ideal wound dressing materials.153 These properties enable nanofibers to effectively load and stably release biomolecules, enhancing their application potential in wound treatment. Co-assembly of solvent-induced nanofibers with ε-poly-L-lysine nanoparticles (nSF-EPL) enables the fabrication of novel multifunctional composite nanoparticles. These composite nanoparticles were successfully employed to co-deliver polydeoxyribonucleotide (PDRN) and HucMSC-Exos. In vivo studies in diabetic wounds showed that these composite nanoparticles exhibit anti-inflammatory and pro-angiogenic capabilities and promote granulation tissue formation, collagen deposition, and wound tissue epithelialization, thus accelerating the skin healing process.65 Another study reported that fixing bovine milk-derived exosomes (M-Exos) on a polydopamine (PDA)-coated hyaluronic acid (HA)-based electrospun nanofiber matrix (M-Exos@PMAT) enhanced cell proliferation and accelerated wound closure, further confirming the effectiveness of nanofibers in promoting wound healing.139 These applications of nanofibers demonstrate their unique advantages in loading and delivering bioactive molecules such as exosomes.
Combination of Exosomes with Other Materials
The combination of exosomes with other materials has also opened new possibilities for wound treatment, especially in the application of complex diabetic-related wounds. These innovative combinations not only improve exosome delivery efficiency but also enhance therapeutic effects. The OxOBand, a dressing composed of antioxidant polyurethane (PUAO) with sustained oxygen release properties, has been shown to significantly enhance the migration of human keratinocytes and fibroblasts when loaded with ADSC-Exos, thereby promoting wound closure, collagen deposition, re-epithelialization, neovascularization, and reducing oxidative stress, effects that collectively promote diabetic wound healing.136,154 The combined use of HUVEC-derived exosomes and methacrylated gelatin microneedle patches accelerated wound healing in diabetic mouse models by promoting collagen deposition, epithelial regeneration, and angiogenesis.137 The use of this microneedle technology not only provides a stable carrier for exosomes but also more effectively delivers exosomes directly to the deeper layers of the wound through the penetration of the microneedles. Encapsulation of M2-Exos within a bilayer microneedle wound dressing system demonstrates the potential to effectively modulate macrophage polarization, attenuate chronic inflammatory responses, and stimulate angiogenesis, thereby accelerating the healing process in diabetic wounds.155 Multifunctional polysaccharide dressings combined with adipose matrix cell-derived exosomes also show potential for promoting angiogenesis and diabetic wound healing.138 The design of this dressing utilizes the natural moisturizing and biocompatible properties of polysaccharides, providing an ideal microenvironment for exosomes. In another study, ADSC-Exos were loaded onto bioengineered microporous three-dimensional amniotic membrane scaffolds (AMS). In vivo experimental results showed that this combination upregulated the gene expression of repair- and regeneration-related factors such as TGF-β, bFGF, and VEGF, while reducing the expression of TNF-α and IL-1β and the number of neutrophils, M1 macrophages, and mast cells, thereby effectively inhibiting inflammatory responses.62 This method not only increased the local concentration of exosomes but also leveraged the three-dimensional structural advantages of AMS, enhancing the interaction between cells and exosomes, further promoting wound repair and regeneration.
Challenges and Future Directions in Exosome-Based Therapeutics
Exosomes hold substantial promise as therapeutic agents for diabetic wound healing due to their multifaceted biological functions. However, the successful clinical translation of exosome-based therapies remains constrained by a number of technical, biological, and regulatory challenges. Overcoming these limitations is essential for the development of safe, effective, and scalable therapeutic strategies.
One of the primary bottlenecks is the low yield of exosomes from standard cell culture systems, which renders large-scale production both economically and technically challenging.156,157 Laboratory-scale isolation techniques—such as ultracentrifugation, size-exclusion chromatography and immunoaffinity capture—are labor-intensive, low-throughput, and often yield heterogeneous vesicle populations. To address this, future research must prioritize the optimization of scalable production systems. Approaches such as high-density bioreactors, stimulus-responsive culture environments, and three-dimensional culture models offer promising avenues for improving exosome output. Concurrently, advances in microfluidic isolation platforms and automated purification technologies may increase processing efficiency while reducing batch variability and cost.158–160
Another further challenge lies in the intrinsic heterogeneity of exosomes, which varies depending on the source cell type, culture conditions, and purification methodology. This variability affects not only therapeutic consistency but also complicates regulatory assessment and standardization. Establishing robust and standardized quality control protocols is imperative. These should include quantification of vesicle concentration and size distribution, profiling of surface markers (eg, CD9, CD63, CD81), and assessment of bioactive cargo such as miRNAs and proteins. Ensuring batch-to-batch reproducibility is a prerequisite for clinical reliability and regulatory compliance.
Exosomes are biologically labile particles, and their storage stability remains a major technical concern. Repeated freeze-thaw cycles and improper storage conditions may compromise membrane integrity and functional cargo, thereby reducing therapeutic efficacy.161,162 There is currently no universal consensus on optimal long-term storage protocols. Ongoing efforts to develop standardized cryopreservation techniques, lyophilization protocols, and stabilizing excipients are needed to maintain exosome bioactivity over extended periods, especially under Good Manufacturing Practice (GMP) conditions.
Another critical hurdle is the efficient and sustained delivery of exosomes to target tissues. In the context of wound healing, simple topical application or systemic administration can lead to rapid clearance, insufficient retention at the wound site, and off-target distribution.163 Integration of exosomes with biomaterial-based delivery systems, such as hydrogels, nanofibers or other materials has demonstrated improved localization, protection from degradation, and sustained therapeutic release in preclinical models. These platforms offer a compelling strategy for enhancing exosome stability and efficacy in vivo. However, biomaterial-exosome systems also face translational challenges, including degradation by protease-rich wound exudates (eg, MMP-9), shear stress-induced leakage, and batch-to-batch variability in release kinetics.36,164 For example, while hydrogels such as GelMA have been shown to retain over 80% of encapsulated exosomes at 72 hours, enzymatic activity and physiological forces can still lead to premature release.134 Additionally, the multi-step manufacturing processes required—encompassing exosome isolation, cargo loading, and biomaterial functionalization—significantly increase complexity and cost. Scaling these approaches under GMP conditions remains logistically demanding and financially prohibitive when compared to conventional therapies. Furthermore, temperature sensitivity, short shelf-life, and quality control inconsistencies in functionalized dressings present substantial barriers to commercialization.
While natural exosomes are widely regarded for their low immunogenicity and biocompatibility—key advantages in therapeutic contexts—the advent of engineered exosomes introduces a new spectrum of safety considerations. Both endogenous and exogenous modification strategies, aimed at enhancing therapeutic specificity or potency, can inadvertently alter the immunological profile of exosomes and increase the risk of adverse immune reactions. These modifications may transform otherwise inert biological vesicles into immunologically active agents, thereby complicating their clinical translation. Endogenous engineering, which involves genetically modifying parent cells to overexpress therapeutic proteins or signaling molecules (eg, Nrf2, HMOX1, or IRF1), can result in the presentation of neoantigens or non-self epitopes on the exosome surface.131–133 Such alterations may trigger immune responses through T-cell activation or neutralizing antibody production, undermining both therapeutic safety and efficacy. Furthermore, the long-term immunological consequences of administering genetically modified vesicles remain largely unknown and warrant thorough investigation. Similarly, exogenous loading techniques—including electroporation, sonication, and surfactant treatment—used to incorporate therapeutic cargos such as siRNA-Keap1 or miRNA-146a-5p can damage the exosomal membrane.18,26 This disruption may expose damage-associated molecular patterns (DAMPs), activating innate immune responses through Toll-like receptor (TLR) signaling pathways.165 Additionally, the cargo itself, particularly synthetic nucleic acids, may exhibit intrinsic immunogenicity. Surface modification strategies, such as hyaluronic acid conjugation, may also introduce new antigenic determinants, further increasing the potential for immune activation.
These safety concerns place engineered exosomes under intense regulatory scrutiny, often leading to their classification as advanced therapy medicinal products or combination products. This designation requires extensive preclinical toxicology, pharmacokinetics, and biodistribution studies, as well as validated assays to assess immunogenicity and ensure lot-to-lot consistency. Developing these regulatory frameworks and manufacturing pipelines represents a formidable challenge for industry stakeholders. Therefore, while engineering approaches unlock powerful opportunities for targeted and enhanced exosome therapeutics, they must be pursued with equally rigorous attention to safety, regulatory compliance, and manufacturing feasibility.
Although exosomes hold immense therapeutic potential, their clinical translation is contingent upon surmounting interconnected challenges in manufacturing, standardization, delivery, and safety. The field is now progressing from preclinical investigations into early-stage human trials, with an expanding cohort of Phase 1 and 2 studies evaluating the safety and efficacy of topically applied exosomes for chronic wounds, including diabetic foot ulcers (Table 3). The initiation of these trials represents a critical validation step for this therapeutic modality. While definitive outcomes are pending, the study designs incorporate innovative strategies, such as combination therapies and rigorous dose-escalation protocols. Nevertheless, for successful clinical translation, future efforts must focus on conducting larger, multicenter randomized controlled trials with standardized endpoints to definitively establish therapeutic efficacy.
 |
Table 3 Clinical Trials of Exosome-Based Therapies for Wound Healing |
Conclusion and Perspectives
Exosomes have introduced innovative approaches to diabetic wound treatment; their multifaceted roles in promoting wound healing have garnered significant attention. Exosomes demonstrate therapeutic potential in diabetes-related wound healing by modulating key biological processes, including the attenuation of inflammatory responses, promotion of angiogenesis, stimulation of cellular proliferation and migration, and enhancement of extracellular matrix deposition. Engineering exosomes further unlocks their therapeutic potential, enabling more precise targeting of key genes and promoting rapid wound closure. Moreover, combining exosomes with various materials enhances both their in vivo stability and therapeutic efficiency. These optimization strategies significantly enhance the applicability of exosomes in diabetic wound management.
However, despite their substantial therapeutic potential, the practical application of exosomes still faces several technical and production challenges. Future research should focus on optimizing exosome production processes and quality control to ensure the reliability of exosome-based therapies.158,159 Establishing standardized exosome preparation and evaluation procedures is also crucial for their clinical translation. With continued in-depth research on exosome biology and their role in disease treatment, more effective exosome-based therapeutic strategies are expected to emerge in the future, offering greater hope and treatment options for diabetic patients.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The author(s) declare that financial support was received for this research, authorship, and/or publication of this article. This research was supported by the Kunming University of Science and Technology & the First People’s Hospital of Yunnan Province Joint Special Project on Medical Research (Grant No. KUST-KH2022032Y), the Yunnan Provincial Department of Science and Technology - Kunming Medical University Joint Special Project on Applied Basic Research (Grant No. 202301AY070001-083), and the Yunnan Fundamental Research Projects (Grant No. 202501AT070041). We also would like to express our sincere gratitude to the editors and anonymous reviewers for their valuable comments, which have greatly improved this paper.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Sun H, Saeedi P, Karuranga S, et al. IDF diabetes atlas: global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diab Res Clin Pract. 2022;183:109119. doi:10.1016/j.diabres.2021.109119
2. Wang Y, Jing L, Lei X, et al. Umbilical cord mesenchymal stem cell-derived apoptotic extracellular vesicles ameliorate cutaneous wound healing in type 2 diabetic mice via macrophage pyroptosis inhibition. Stem Cell Res Ther. 2023;14(1). doi:10.1186/s13287-023-03490-6
3. Swoboda L, Held J. Impaired wound healing in diabetes. J Wound Care. 2022;31(10):882–885. doi:10.12968/jowc.2022.31.10.882
4. Wu S, Zhou Z, Li Y, Jiang J. Advancements in diabetic foot ulcer research: focus on mesenchymal stem cells and their exosomes. Heliyon. 2024 ;10(17).
5. Schaper NA-O, van Netten JA-O, Apelqvist J, Bus SA-O, Hinchliffe RJ, Lipsky BA. Practical Guidelines on the prevention and management of diabetic foot disease (IWGDF 2019 update). Diab/Metabol Res Rev. 2020;36(S1). doi:10.1002/dmrr.3266
6. Everett E, Mathioudakis NA-OX. Update on management of diabetic foot ulcers. Annals New York Acad Sci. 2018;1411(1):153–165. doi:10.1111/nyas.13569
7. Lim JZ, Ng NS, Thomas C. Prevention and treatment of diabetic foot ulcers. J Royal Soc Med. 2017;110(3):104–109. doi:10.1177/0141076816688346
8. Fau ND, Fau GS, Fau LJ, et al. The effect of intensive treatment of diabetes on the development and progression of long-term complications in insulin-dependent diabetes mellitus. New Eng J Med. 1993;329:977–86.
9. Hodson AE, Tippetts TS, Bikman BT. Insulin treatment increases myocardial ceramide accumulation and disrupts cardiometabolic function. Cardiovasc Diabetol. 2015;14(1):153.
10. Lenselink E, Holloway S, Eefting D. Outcomes after foot surgery in people with a diabetic foot ulcer and a 12-month follow-up. J Wound Care. 2017;26(5):218–227. doi:10.12968/jowc.2017.26.5.218
11. Bian D, Wu Y, Song GA-O, Azizi R, Zamani A. The application of mesenchymal stromal cells (MSCs) and their derivative exosome in skin wound healing: a comprehensive review. Stem Cell Res Ther. 2022;13(1). doi:10.1186/s13287-021-02697-9
12. Wu J, Chen LH, Sun SY, Li Y, Ran XW. Mesenchymal stem cell-derived exosomes: the Dawn of diabetic wound healing. World J Diab. 2022;13(12):1066–1095. doi:10.4239/wjd.v13.i12.1066
13. Wang Y, Zhu J, Chen J, Xu R, Groth TA-O. The signaling pathways induced by exosomes in promoting diabetic wound healing: a mini-review. Current Issues Mol Biol. 2022;44(10):4960–4976. doi:10.3390/cimb44100337
14. Ren S, Chen J, Guo J, et al. Exosomes from adipose stem cells promote diabetic wound healing through the eHSP90/LRP1/AKT axis. Cells. 2022;11:3229. LID. doi:10.3390/cells11203229
15. Wang W, Zhao Y, Zhu P, et al. Differential proteomic profiles of coronary serum exosomes in acute myocardial infarction patients with or without diabetes mellitus: ANGPTL6 accelerates regeneration of endothelial cells treated with rapamycin via MAPK pathways. Cardiovasc Drugs Ther. 2024;38(1):13–29. doi:10.1007/s10557-022-07365-5
16. Liu Y, Zhong C, Chen S, Xue Y, Wei Z. Circulating exosomal mir-16-2-3p is associated with coronary microvascular dysfunction in diabetes through regulating the fatty acid degradation of endothelial cells. Cardiovasc Diabetol. 2024;23(1). doi:10.1186/s12933-024-02142-0
17. Liu K, Gong B, Li T, et al. Bioactive self-healing umbilical cord blood exosomes hydrogel for promoting chronic diabetic wound healing. Biochem Biophys Res Commun. 2024;690:149241. doi:10.1016/j.bbrc.2023.149241
18. Xiang X, Chen J, Jiang T, et al. Milk-derived exosomes carrying siRNA-KEAP1 promote diabetic wound healing by improving oxidative stress. Drug Delivery Transl Res. 2023;13(9):2286–2296. doi:10.1007/s13346-023-01306-x
19. Chen CY, Rao SS, Ren L, et al. Exosomal DMBT1 from human urine-derived stem cells facilitates diabetic wound repair by promoting angiogenesis. Theranostics. 2018;8(6):1607–1623. doi:10.7150/thno.22958
20. Zhang M, Guo J, Xiang K, et al. Exosomes derived from oral squamous cell carcinoma tissue accelerate diabetic wound healing. Ame J Physiol -Cell Physiol. 2023;324(6):C1307–C1319. doi:10.1152/ajpcell.00541.2022
21. Liao Q, Su L, Pang L, et al. Natural exosome-like nanoparticles derived from ancient medicinal insect Periplaneta americana L. as a novel diabetic wound healing accelerator. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-01923-1
22. Miya MB, Chandra Gupta P, Pathak V, Mishra R. Therapeutic effects of OXY- Exo(Aloe) in diabetic wound injury. Biochem Biophys Res Commun. 2024;731:150398.
23. Zhang C, Xiao W, Wang H, et al. Exosomes derived from mouse breast carcinoma cells facilitate diabetic wound healing. Tissue Eng Regen Med. 2024;21(4):571–586. doi:10.1007/s13770-024-00629-1
24. Xu H, Yang H, Wang Z, et al. Epidermal stem cell derived exosomes alleviate excessive autophagy induced endothelial cell apoptosis by delivering miR200b-3p to diabetic wounds. J Investigative Dermatol. 2024;144(5):1134–1147.e2. doi:10.1016/j.jid.2023.08.030
25. Shi Y, Wang S, Wang K, et al. Relieving macrophage dysfunction by inhibiting SREBP2 activity: a hypoxic mesenchymal stem cells-derived exosomes loaded multifunctional hydrogel for accelerated diabetic wound healing. Small. 2024;20(25). doi:10.1002/smll.202309276
26. Zhou X, Ye C, Jiang L, et al. The bone mesenchymal stem cell-derived exosomal miR-146a-5p promotes diabetic wound healing in mice via macrophage M1/M2 polarization. Mol Cell Endocrinol. 2024;579:112089. doi:10.1016/j.mce.2023.112089
27. Lotfy A, AboQuella NM, Wang H. Mesenchymal stromal/stem cell (MSC)-derived exosomes in clinical trials. Stem Cell Res Ther. 2023;14(1):66.
28. Bharucha AE, Wald A, Enck P, Rao S. Functional anorectal disorders. Gastroenterology. 2006;5:1510–8.
29. Nourian Dehkordi A, Mirahmadi Babaheydari F, Chehelgerdi MA-O, Raeisi Dehkordi S. Skin tissue engineering: wound healing based on stem-cell-based therapeutic strategies. Stem Cell Res Ther. 2019;10(1). doi:10.1186/s13287-019-1212-2
30. Opneja A, Kapoor S, Stavrou EX. Contribution of platelets, the coagulation and fibrinolytic systems to cutaneous wound healing. Thrombosis Res. 2019;179:56–63. doi:10.1016/j.thromres.2019.05.001
31. Wilkinson HN, Hardman MJ. Wound healing: cellular mechanisms and pathological outcomes. Open Biol. 2020;10(9). doi:10.1098/rsob.200223
32. Louiselle AE, Niemiec SM, Zgheib C, Liechty KW. Macrophage polarization and diabetic wound healing. Transl Res. 2021;236:109–116. doi:10.1016/j.trsl.2021.05.006
33. Lopez TA-OX, Wendremaire MA-O, Lagarde J, Duquet O, Alibert L, Paquette B. Wound healing versus metastasis: role of oxidative stress. Biomedicines. 2022;10(11):2784.LID -. doi:10.3390/biomedicines10112784
34. Talbott HE, Mascharak S, Griffin M, Wan DC, Longaker MT. Wound healing, fibroblast heterogeneity, and fibrosis. Cell Stem Cell. 2022;29(8):1161–1180. doi:10.1016/j.stem.2022.07.006
35. Piipponen M, Li D, Landén NA-O. The immune functions of keratinocytes in skin wound healing. Int J Mol Sci. 2020;21(22):8790. LID - 8790. doi:10.3390/ijms21228790
36. Hade MD.Suire CN, Suo Z. Mesenchymal stem cell-derived exosomes: applications in regenerative medicine. Cells. 2021;10(8):1959. doi:10.3390/cells10081959
37. Lindley LE, Stojadinovic O, Pastar I, Tomic-Canic M. Biology and biomarkers for wound healing. Plastic Reconst Surg. 2016;138(3S):18S–28S. doi:10.1097/PRS.0000000000002682
38. Wang PH, Huang BS, Horng HC, Yeh CC, Chen YJ. Wound healing. J Chin Med Assoc. 2018;81(2):94–101. doi:10.1016/j.jcma.2017.11.002
39. Sharma S, Rai VK, Narang RK, Markandeywar TS. Collagen-based formulations for wound healing: a literature review. Life Sci. 2022;290:120096. doi:10.1016/j.lfs.2021.120096
40. Khalid MA-O, Petroianu G, Adem A. Advanced glycation end products and diabetes mellitus: mechanisms and perspectives. Biomolecules. 2022;12(4):542. LID - 542. doi:10.3390/biom12040542
41. Garay-Sevilla ME, Rojas A, Portero-Otin MA-O, Uribarri J. Dietary AGEs as exogenous boosters of inflammation. Nutrients. 2021;13(8):2802. LID - 2802. doi:10.3390/nu13082802
42. Yang K, Yang X, Gao C, Hua C, Hong C, Zhu L. Novel microfluidic device for the neutrophil functional phenotype analysis: effects of glucose and its derivatives AGEs. Micromachines. 2021;12(8):944. LID - 944. doi:10.3390/mi12080944
43. Peppa M, Fau SP, Raptis SA, Raptis SA. Advanced glycoxidation products and impaired diabetic wound healing. Wound Repair Regen. 2009;17(4):461–472. doi:10.1111/j.1524-475X.2009.00518.x
44. Mao QY, He SY, Hu QY, et al. Advanced glycation end products (AGEs) inhibit macrophage efferocytosis of apoptotic β cells through binding to the receptor for AGEs. J Immunol. 2022;208(5):1204–1213. doi:10.4049/jimmunol.2100695
45. Li M, Hou Q, Zhong L, Zhao Y, Fu X. Macrophage related chronic inflammation in non-healing wounds. Front Immunol. 2021;12. doi:10.3389/fimmu.2021.681710
46. Sharma SA-O, Schaper NA-O, Rayman G. Microangiopathy: is it relevant to wound healing in diabetic foot disease? Diab/Metab Res Rev. 2020;36(S1). doi:10.1002/dmrr.3244
47. Dudley AC, Griffioen AW. The modes of angiogenesis: an updated perspective. Angiogenesis. 2023;26(4):477–480. doi:10.1007/s10456-023-09895-4
48. Isidori AM, Venneri MA, Fiore D. Angiopoietin-1 and Angiopoietin-2 in metabolic disorders: therapeutic strategies to restore the highs and lows of angiogenesis in diabetes. J Endocrinol Investig. 2016;39(11):1235–1246. doi:10.1007/s40618-016-0502-0
49. Abdelsaid KA-O, Sudhahar V, Harris RA, et al. Exercise improves angiogenic function of circulating exosomes in type 2 diabetes: role of exosomal SOD3. FASEB J. 2022;36(3):e22177.
50. Zhang X, Jiang Y, Huang Q, et al. Exosomes derived from adipose-derived stem cells overexpressing glyoxalase-1 protect endothelial cells and enhance angiogenesis in type 2 diabetic mice with limb ischemia. Stem Cell Res Ther. 2021;12(1). doi:10.1186/s13287-021-02475-7
51. Obrosova IG. Update on the pathogenesis of diabetic neuropathy. Current Diab Rep. 2003;3(6):439–445. doi:10.1007/s11892-003-0005-1
52. Falanga V. Wound healing and its impairment in the diabetic foot. Lancet. 2005;366(9498):1736–1743. doi:10.1016/S0140-6736(05)67700-8
53. Elafros MA, Andersen H, Bennett DL, et al. Towards prevention of diabetic peripheral neuropathy: clinical presentation, pathogenesis, and new treatments. Lancet Neurol. 2022;21(10):922–936. doi:10.1016/S1474-4422(22)00188-0
54. Boulton AJ, Armstrong DG, Kirsner RS, et al. Diagnosis and management of diabetic foot complications. ADA Clin Compendia. 2018;2018(2):1–20. doi:10.2337/db20182-1
55. Deng LA-O, CA-O D, Song PA-O, et al. The role of oxidative stress and antioxidants in diabetic wound healing. Oxidative Med Cell Longevity. 2021;2021(1). doi:10.1155/2021/8852759
56. Volmer-Thole M, Lobmann R. Neuropathy and diabetic foot syndrome.Int J Mol Sci. 2016;17(6):917. doi:10.3390/ijms17060917
57. Ashrafi M, Baguneid M, Bayat A. The role of neuromediators and innervation in cutaneous wound healing. Acta Derm Venereol. 2016;96:587–594. doi:10.2340/00015555-2321
58. Bodman MA, Dreyer MA, Varacallo MA. Diabetic Peripheral Neuropathy. Treasure Island (FL) ineligible companies: StatPearls; 2025. Disclosure: Mark Dreyer declares no relevant financial relationships with ineligible companies. Disclosure: Matthew Varacallo declares no relevant financial relationships with ineligible companies.
59. Soares CLR, Wilairatana P, Silva LR, et al. Biochemical aspects of the inflammatory process: a narrative review. Biomed Pharmacother. 2023;168:115764. doi:10.1016/j.biopha.2023.115764
60. da Silva PA-O, Do Espírito Santo RF, Melo CA-O, et al. The compound (E)-2-Cyano-N,3-diphenylacrylamide (JMPR-01): a potential drug for treatment of inflammatory diseases. Pharmaceutics. 2022;14(1):188. LID - 188. doi:10.3390/pharmaceutics14010188
61. Huldani H, Kozlitina IA, Alshahrani M, et al. Exosomes derived from adipose stem cells in combination with hyaluronic acid promote diabetic wound healing. Tissue Cell. 2023;85:102252. doi:10.1016/j.tice.2023.102252
62. Khalatbary AA-O, Omraninava MA-O, Nasiry DA-O, et al. Exosomes derived from human adipose mesenchymal stem cells loaded bioengineered three-dimensional amniotic membrane-scaffold-accelerated diabetic wound healing. Arch Dermatol Res. 2023;315(10):2853–2870. doi:10.1007/s00403-023-02709-z
63. Liu W, Yu M, Xie D, Wang L, Ye C, Zhu Q. Melatonin-stimulated MSC-derived exosomes improve diabetic wound healing through regulating macrophage M1 and M2 polarization by targeting the PTEN/AKT pathway. Stem Cell Res Ther. 2020;11(1). doi:10.1186/s13287-020-01756-x
64. Sanders MA-O, Balaji S, Martin WB, et al. Protecting human amnion and chorion matrices during processing: performance enhancement in a diabetic mouse model and human co-culture system. Wound Repair Regen. 2023;31(4):475–488. doi:10.1111/wrr.13099
65. Li X, Qu S, Ouyang Q, et al. A multifunctional composite nanoparticle with antibacterial activities, anti-inflammatory, and angiogenesis for diabetic wound healing. Int J Biol Macromol. 2024;260:129531. doi:10.1016/j.ijbiomac.2024.129531
66. Banerjee A, Singh P, Sheikh PA, Kumar A, Koul V, Bhattacharyya J. A multifunctional silk-hyaluronic acid self-healing hydrogel laden with alternatively activated macrophage-derived exosomes reshape microenvironment of diabetic wound and accelerate healing. Int J Biol Macromol. 2024;270:132384. doi:10.1016/j.ijbiomac.2024.132384
67. Liu Y, Liu Y, Deng J, Li W, Nie X. Fibroblast growth factor in diabetic foot ulcer: progress and therapeutic prospects. Front Endocrinol. 2021;12. doi:10.3389/fendo.2021.744868
68. Worsley AL, Lui DH, Ntow-Boahene W, Song W, Good L, Tsui J. The importance of inflammation control for the treatment of chronic diabetic wounds. Int Wound J. 2023;20(6):2346–2359. doi:10.1111/iwj.14048
69. Wolf P. The nature and significance of platelet products in human plasma. Brit J Haematol. 1967;13(3):269–288. doi:10.1111/j.1365-2141.1967.tb08741.x
70. Pan BT, Teng K, Wu C, Adam M, Johnstone RM. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol. 1985;101(3):942–8.
71. Harding C, Heuser J, Stahl P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol. 1983;97(2):329–39
72. Johnstone RF, Adam M, Adam MF, et al. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–9420. doi:10.1016/S0021-9258(18)48095-7
73. Théry C, Amigorena S, Fau - Raposo G, Fau RG. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Current Protocols Cell Biol. 2006;30(1). doi:10.1002/0471143030.cb0322s30
74. Kowal J, Arras G, Colombo M, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proceed Nat Acad Sci. 2016;113(8). doi:10.1073/pnas.1521230113
75. Song L, Tang S, Han X, et al. KIBRA controls exosome secretion via inhibiting the proteasomal degradation of Rab27a. Nat Commun. 2019;10(1). doi:10.1038/s41467-019-09720-x
76. Villarroya-Beltri C, Baixauli F, Mittelbrunn M, et al. ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins. Nat Commun. 2016;7(1). doi:10.1038/ncomms13588
77. Hyenne VA-O, Labouesse M, Goetz JA-O. The Small GTPase Ral orchestrates MVB biogenesis and exosome secretion. Small GTPases. 2018;9(6):445–451. doi:10.1080/21541248.2016.1251378
78. Marie PP, Fan SJ, Mason J, et al. Accessory ESCRT-III proteins are conserved and selective regulators of Rab11a-exosome formation. J Extracellular Vesicles. 2023;12(3). doi:10.1002/jev2.12311
79. Read CB, Lind MCH, Chiarelli TJ, et al. The obligate intracellular bacterial pathogen anaplasma phagocytophilum exploits host cell multivesicular body biogenesis for proliferation and dissemination. mBio. 2022;13(6). doi:10.1128/mbio.02961-22
80. Graf S, Pesch H, Appleson T, Lei T, Breder A, Siewert I. Mechanistic analysis reveals key role of interchalcogen multicatalysis in photo-aerobic 3-pyrroline syntheses by aza-wacker cyclizations. ChemSusChem. 2024;17(9). doi:10.1002/cssc.202301518
81. Yang F, Cai D, Kong R, et al. Exosomes derived from cord blood Treg cells promote diabetic wound healing by targeting monocytes. Biochem Pharmacol. 2024;226:116413. doi:10.1016/j.bcp.2024.116413
82. Shi X, Li Y, Kang S, et al. Dual-functional gallium/chitosan/silk/umbilical cord mesenchymal stem cell exosome sponge scaffold for diabetic wound by angiogenesis and antibacteria. Int J Biol Macromol. 2024;274:133420. doi:10.1016/j.ijbiomac.2024.133420
83. Hildonen S, Skarpen E, Halvorsen TG, Reubsaet L. Isolation and mass spectrometry analysis of urinary extraexosomal proteins. Scientific Rep. 2016;6(1). doi:10.1038/srep36331
84. Yang J, Chen Z, Pan D, Li H, Shen J. Umbilical Cord-derived mesenchymal stem cell-derived exosomes combined pluronic f127 hydrogel promote chronic diabetic wound healing and complete skin regeneration. Int J Nanomed. 2020:5911–26. doi:10.2147/IJN.S249129
85. Han QF, Li WJ, Hu KS, et al. Exosome biogenesis: machinery, regulation, and therapeutic implications in cancer. Mol Cancer. 2022;21(1). doi:10.1186/s12943-022-01671-0
86. Zhang Y, Fang M, Xie W, et al. Sprayable alginate hydrogel dressings with oxygen production and exosome loading for the treatment of diabetic wounds. Int J Biol Macromol. 2023;242:125081. doi:10.1016/j.ijbiomac.2023.125081
87. Zhong F, Cao S, Yang L, et al. Low‑intensity pulsed ultrasound accelerates diabetic wound healing by ADSC‑derived exosomes via promoting the uptake of exosomes and enhancing angiogenesis. Int J Mol Med. 2024;53(3). doi:10.3892/ijmm.2024.5347
88. Qin L, Chen L, Chen C, Hu Q, Wang JA-O, Shen J. Serum exosomes accelerate diabetic wound healing by promoting angiogenesis and ECM formation. Cell Biol Int. 2021;45(9):1976–85.
89. Gurung S, Perocheau D, Touramanidou L, Baruteau JA-OX. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signaling. 2021;19(1). doi:10.1186/s12964-021-00730-1
90. Skotland T, Hessvik NP, Sandvig K, Llorente A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J Lipid Res. 2019;60(1):9–18. doi:10.1194/jlr.R084343
91. Wang X, Weidling I, Koppel S, et al. Detection of mitochondria-pertinent components in exosomes. Mitochondrion. 2020;55:100–110. doi:10.1016/j.mito.2020.09.006
92. D’Souza A, Burch A, Dave KM, et al. Microvesicles transfer mitochondria and increase mitochondrial function in brain endothelial cells. J Controlled Release. 2021;338:505–526. doi:10.1016/j.jconrel.2021.08.038
93. Abbasi Sourki P, Pourfathollah AA, Kaviani S, et al. The profile of circulating extracellular vesicles depending on the age of the donor potentially drives the rejuvenation or senescence fate of hematopoietic stem cells. Exp Gerontol. 2023;175:112142. doi:10.1016/j.exger.2023.112142
94. Hamdan Y, Mazini L, Malka G. Exosomes and Micro-RNAs in aging process. Biomedicines. 2021;9(8):968. doi:10.3390/biomedicines9080968
95. Kolhe R, Hunter M, Liu S, et al. Gender-specific differential expression of exosomal miRNA in synovial fluid of patients with osteoarthritis. Sci Rep. 2017;7(1):2029. doi:10.1038/s41598-017-01905-y
96. Lee H, Bae YU, Kim H, et al. Effect of diabetes on exosomal miRNA profile in patients with obesity. BMJ Open Diab Res Care. 2020;8:e001403. doi:10.1136/bmjdrc-2020-001403
97. Gurunathan S, Kang MA-O, Kim JA-O. A comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int J Nanomed. 2021;Volume 16:1281–1312. doi:10.2147/IJN.S291956
98. Reis M, Mavin E, Nicholson L, Green K, Dickinson AM, Wang XN. Mesenchymal stromal cell-derived extracellular vesicles attenuate dendritic cell maturation and function. Front Immunol. 2018;9. doi:10.3389/fimmu.2018.02538
99. Xia W, Liu Y, Jiang X, et al. Lean adipose tissue macrophage derived exosome confers immunoregulation to improve wound healing in diabetes. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-01869-4
100. Tung SL, Boardman DA, Sen M, et al. Regulatory T cell-derived extracellular vesicles modify dendritic cell function. Scientific Rep. 2018;8(1). doi:10.1038/s41598-018-24531-8
101. Yang J, Xie Y, Xia Z, et al. Hucmsc-Exo Induced N2 Polarization of Neutrophils: Implications for Angiogenesis and Tissue Restoration in Wound Healing. Int J Nanomed. 2024;2024:3555–75.
102. Teng L, Maqsood M, Zhu M, et al. Exosomes derived from human umbilical cord mesenchymal stem cells accelerate diabetic wound healing via promoting M2 macrophage polarization, angiogenesis, and collagen deposition. Int J Mol Sci. 2022;23(18):10421. doi:10.3390/ijms231810421
103. Zhang Y, Bai X, Shen K, et al. Exosomes derived from adipose mesenchymal stem cells promote diabetic chronic wound healing through SIRT3/SOD2. New Eng J Med. 1993;329(14):977–86. doi:10.3390/cells11162568
104. Wang Z, Feng C, Liu H, et al. Exosomes from circ-Astn1-modified adipose-derived mesenchymal stem cells enhance wound healing through miR-138-5p/SIRT1/FOXO1 axis regulation. World J Stem Cells. 2023;15(5):476.
105. Pi L, Yang L, Fang BR, Meng XX, Qian LA-O. Exosomal microRNA-125a-3p from human adipose-derived mesenchymal stem cells promotes angiogenesis of wound healing through inhibiting PTEN. Mol Cell Biochem. 2022;477(1):115–127. doi:10.1007/s11010-021-04251-w
106. Yin D, Shen GA-O. Exosomes from adipose-derived stem cells regulate macrophage polarization and accelerate diabetic wound healing via the circ-Rps5/miR-124-3p axis. Immun Inflammation Dis. 2024;12(6). doi:10.1002/iid3.1274
107. Zhao B, Zhang X, Zhang Y, et al. Human exosomes accelerate cutaneous wound healing by promoting collagen synthesis in a diabetic mouse model. Stem Cells Dev. 2021;30(18):922–33
108. Wu G, Zhang J, Fau - Fan GG, Fan Gg Fau - Zou ZY. MiRNA-324-5p inhibits inflammatory response of diabetic vessels by targeting CPT1A. Eur Rev Med Pharmacol Sci. 2020;24(24):1.
109. Chen J, Li X, Liu H, et al. Bone marrow stromal cell-derived exosomal circular RNA improves diabetic foot ulcer wound healing by activating the nuclear factor erythroid 2-related factor 2 pathway and inhibiting ferroptosis. Diab Med. 2023;40(7). doi:10.1111/dme.15031
110. Qiu ZY, Xu WC, Liang ZA-O. Bone marrow mesenchymal stem cell-derived exosomal miR −221-3p promotes angiogenesis and wound healing in diabetes via the downregulation of forkhead box P1. Diab Med. 2024;41(9). doi:10.1111/dme.15386
111. Yang H, Zhang Y, Du Z, Wu T, Yang C. Hair follicle mesenchymal stem cell exosomal lncRNA H19 inhibited NLRP3 pyroptosis to promote diabetic mouse skin wound healing. Aging. 2023;15(3):791–809. doi:10.18632/aging.204513
112. Li P, Hong G, Zhan W, Deng M, Tu C. Endothelial progenitor cell derived exosomes mediated miR-182-5p delivery accelerate diabetic wound healing via down-regulating PPARG. Int J Med Sci. 2023;20(4):468–481. doi:10.7150/ijms.78790
113. Cao W, Meng X, Cao F, Wang J, Yang M. Exosomes derived from platelet-rich plasma promote diabetic wound healing via the JAK2/STAT3 pathway. Iscience. 2023;26(11):108236. doi:10.1016/j.isci.2023.108236
114. Kuang L, Zhang CA-O, Li B, Deng H, Chen R, Li G. Human keratinocyte-derived exosomal MALAT1 promotes diabetic wound healing by upregulating MFGE8 via microRNA-1914-3p. Int J Nanomed. 2023;Volume 18:949–970. doi:10.2147/IJN.S399785
115. Han X, Wu P, Li L, et al. Exosomes derived from autologous dermal fibroblasts promote diabetic cutaneous wound healing through the Akt/β-catenin pathway. Cell Cycle. 2021;20(5–6):616–629. doi:10.1080/15384101.2021.1894813
116. Yu M, Liu W, Li J, Lu J, Lu H. Exosomes derived from atorvastatin-pretreated MSC accelerate diabetic wound repair by enhancing angiogenesis via AKT/eNOS pathway. Stem Cell Res Ther. 2020;11(1). doi:10.1186/s13287-020-01824-2
117. Hu Y, Tao R, Chen L, et al. Exosomes derived from pioglitazone-pretreated MSCs accelerate diabetic wound healing through enhancing angiogenesis. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-021-00894-5
118. Shi Y, Wang SA-O, Liu DA-O, et al. Exosomal miR-4645-5p from hypoxic bone marrow mesenchymal stem cells facilitates diabetic wound healing by restoring keratinocyte autophagy. Burns Trauma. 2024;12. doi:10.1093/burnst/tkad058
119. Lu W, Du XA-O, Zou S, et al. IFN -γ enhances the therapeutic efficacy of MSCs -derived exosome via miR −126-3p in diabetic wound healing by targeting SPRED1. J Diab. 2024;16(1). doi:10.1111/1753-0407.13465
120. Xiong W, Bai X, Zhang X, et al. Endothelial progenitor-cell-derived exosomes induced by astragaloside iv accelerate type I diabetic-wound healing via the PI3K/AKT/mTOR pathway in rats. Front Biosci Landmark. 2023;28(11). doi:10.31083/j.fbl2811282
121. Shen YF, Huang JH, Wang KY, et al. PTH Derivative promotes wound healing via synergistic multicellular stimulating and exosomal activities. Cell Commun Signaling. 2020;18(1):40
122. Zhao X, Fu L, Zou H, et al. Optogenetic engineered umbilical cord MSC-derived exosomes for remodeling of the immune microenvironment in diabetic wounds and the promotion of tissue repair. J Nanobiotechnol. 2023;21(1). doi:10.1186/s12951-023-01886-3
123. Ti D, Hao H, Tong C, et al. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J Transl Med. 2015;13(1). doi:10.1186/s12967-015-0642-6
124. Wang L, Chen J, Song J, et al. Activation of the Wnt/β-catenin signalling pathway enhances exosome production by hucMSCs and improves their capability to promote diabetic wound healing. J Nanobiotechnol. 2024;22(1). doi:10.1186/s12951-024-02650-x
125. Li Q, Guo L, Wang J, Tao S, Jin P. Exosomes derived from Nr-CWS pretreated MSCs facilitate diabetic wound healing by promoting angiogenesis via the circIARS1/miR-4782-5p/VEGFA axis. Chin J Nat Med. 2023;21(3):172–184. doi:10.1016/S1875-5364(23)60419-4
126. Wang J, Wu H, Peng Y, Qin Y, Zhao Y. Hypoxia adipose stem cell-derived exosomes promote high-quality healing of diabetic wound involves activation of PI3K/Akt pathways. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-021-00942-0
127. Shi R, Jin Y, Zhao S, Yuan H, Shi J, Zhao H. Hypoxic ADSC-derived exosomes enhance wound healing in diabetic mice via delivery of circ-Snhg11 and induction of M2-like macrophage polarization. Biomed Pharmacother. 2022;153:113463. doi:10.1016/j.biopha.2022.113463
128. Tang W, Du X, Wu Z, Nie Z, Yu C, Gao Y. circ-Erbb2ip from adipose-derived mesenchymal stem cell-derived exosomes promotes wound healing in diabetic mice by inducing the miR-670-5p/Nrf1 axis. Cell Signalling. 2024;121:111245. doi:10.1016/j.cellsig.2024.111245
129. Cheng P, Xie X, Hu L, et al. Hypoxia endothelial cells-derived exosomes facilitate diabetic wound healing through improving endothelial cell function and promoting M2 macrophages polarization. Bioactive Mat. 2024;33:157–173. doi:10.1016/j.bioactmat.2023.10.020
130. Nakao Y, Fukuda T, Zhang Q, et al. Exosomes from TNF-α-treated human gingiva-derived MSCs enhance M2 macrophage polarization and inhibit periodontal bone loss. Acta Biomaterialia. 2021;122:306–324. doi:10.1016/j.actbio.2020.12.046
131. Xie X, Li X, Lian W, et al. Exosomes from adipose-derived stem cells overexpressing Nrf2 accelerate cutaneous wound healing by promoting vascularization in a diabetic foot ulcer rat model. Exp Mol Med. 2018;50(4):1–4.
132. Wu M, Tu J, Huang J, Wen H, Zeng Y, Lu Y. Exosomal IRF1-loaded rat adipose-derived stem cell sheet contributes to wound healing in the diabetic foot ulcers. Mol Med. 2023;29(1). doi:10.1186/s10020-023-00617-6
133. Cheng B, Song X, Yin L, Lin J, Liu Z. HMOX1-overexpressing mesenchymal stem cell-derived exosomes facilitate diabetic wound healing by promoting angiogenesis and fibroblast function. Biochem Biophys Res Commun. 2024;690:149271. doi:10.1016/j.bbrc.2023.149271
134. Wang C, Wang M, Xu T, et al. Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration.Theranostics. 2019;9(1):65.
135. Hu N, Cai Z, Jiang X, et al. Hypoxia-pretreated ADSC-derived exosome-embedded hydrogels promote angiogenesis and accelerate diabetic wound healing. Acta Biomaterialia. 2023;157:175–186. doi:10.1016/j.actbio.2022.11.057
136. Shiekh PA, Singh A, Kumar A. Exosome laden oxygen releasing antioxidant and antibacterial cryogel wound dressing OxOBand alleviate diabetic and infectious wound healing. Biomaterials. 2020;249:120020. doi:10.1016/j.biomaterials.2020.120020
137. Yuan M, Liu K, Jiang T, et al. GelMA/PEGDA microneedles patch loaded with HUVECs-derived exosomes and Tazarotene promote diabetic wound healing. J Nanobiotechnol. 2022;20(1). doi:10.1186/s12951-022-01354-4
138. Wang M, Wang C, Chen M, et al. Efficient angiogenesis-based diabetic wound healing/skin reconstruction through bioactive antibacterial adhesive ultraviolet shielding nanodressing with exosome release. ACS Nano. 2019;13(9):10279–10293. doi:10.1021/acsnano.9b03656
139. Bui HD, You G, Lee M, et al. Milk exosome-infused fibrous matrix for treatment of acute wound. J Controlled Release. 2024;376:79–93. LID - S0168-3659(24)00663-1 [pii] LID -. doi:10.1016/j.jconrel.2024.09.053
140. Li S, Li Y, Zhu K, et al. Exosomes from mesenchymal stem cells: potential applications in wound healing. Life Sci. 2024;357:123066. doi:10.1016/j.lfs.2024.123066
141. Zhao Z, Qu L, Shuang T, et al. Low-intensity ultrasound radiation increases exosome yield for efficient drug delivery. J Drug Delivery Sci Technol. 2020;57:101713. doi:10.1016/j.jddst.2020.101713
142. Duan YR, Zhao YC, Song WY, Wang JX, Pei J, Wang XB. Research advances on improving the therapeutic efficacy of mesenchymal stem cell-derived exosomes in wound repair. Zhonghua Shao Shang za zhi= Zhonghua Shaoshang Zazhi= Chinese J Burns. 2023;39(7):695–700
143. Donoso-Quezada JA-O, Ayala-Mar S, González-Valdez J. State-of-the-art exosome loading and functionalization techniques for enhanced therapeutics: a review. Crit Rev Biotechnol. 2020;40(6):804–820. doi:10.1080/07388551.2020.1785385
144. Piffoux M, Volatron J, Cherukula K, et al. Engineering and loading therapeutic extracellular vesicles for clinical translation: a data reporting frame for comparability. Adv Drug Delivery Rev. 2021;178:113972. doi:10.1016/j.addr.2021.113972
145. Yan C, Chen J, Wang C, et al. Milk exosomes-mediated miR-31-5p delivery accelerates diabetic wound healing through promoting angiogenesis. Drug Delivery. 2022;29(1):214–228. doi:10.1080/10717544.2021.2023699
146. Chen M, Wang QA-O, Fan Y, Wang Y, Zhang X. Biomaterials-assisted exosomes therapy in osteoarthritis. Biomed Mat. 2022;17(2):022001. doi:10.1088/1748-605X/ac4c8c
147. Golchin AA-O, Shams F, Basiri A, et al. Combination therapy of stem cell-derived exosomes and biomaterials in the wound healing. Stem Cell Rev Rep. 2022;18(6):1892–1911. doi:10.1007/s12015-021-10309-5
148. Sareen N, Srivastava A, Alagarsamy KN, Lionetti V, Dhingra S. Stem cells derived exosomes and biomaterials to modulate autophagy and mend broken hearts. Biochimica Et Biophysica Acta. 2023;1869(7):166806. doi:10.1016/j.bbadis.2023.166806
149. Zhang X, Jiang W, Lu Y, Mao T, Gu Y. Exosomes combined with biomaterials in the treatment of spinal cord injury. Front Bioeng Biotechnol. 2023;11. doi:10.3389/fbioe.2023.1077825
150. Ojeda-Hernández DA-O, Hernández-Sapiéns MA-O, Reza-Zaldívar EE, et al. Exosomes and Biomaterials: In Search of a New Therapeutic Strategy for Multiple Sclerosis. Life. 2022 ;12(9):1417. doi:10.3390/life12091417
151. Wang C, Liang C, Wang R, et al. The fabrication of a highly efficient self-healing hydrogel from natural biopolymers loaded with exosomes for the synergistic promotion of severe wound healing. Biomat Sci. 2020;8(1):313–24.
152. Li D, Wu N. Mechanism and application of exosomes in the wound healing process in diabetes mellitus. Diab Res Clin Pract. 2022;187:109882. doi:10.1016/j.diabres.2022.109882
153. Blanco-Fernandez B, Castaño O, Mateos-Timoneda M, Engel E, Pérez-Amodio S. Nanotechnology approaches in chronic wound healing. Adv Wound Care. 2021 ;10(5):234–56.
154. Shiekh PA, Singh A, Kumar A. Data supporting exosome laden oxygen releasing antioxidant and antibacterial cryogel wound dressing OxOBand alleviate diabetic and infectious wound healing. Data in Brief. 2020;31:105671. doi:10.1016/j.dib.2020.105671
155. Zeng J, Sun Z, Zeng F, Gu C, Chen X. M2 macrophage-derived exosome-encapsulated microneedles with mild photothermal therapy for accelerated diabetic wound healing. Mat Today Bio. 2023;20:100649. doi:10.1016/j.mtbio.2023.100649
156. Chen J, Li P, Zhang T, et al. Review on strategies and technologies for exosome isolation and purification. Front Bioeng Biotechnol. 2021;9:811971. doi:10.3389/fbioe.2021.811971
157. Yamashita T, Takahashi Y, Takakura Y. Possibility of exosome-based therapeutics and challenges in production of exosomes eligible for therapeutic application. Biol Pharm Bull. 2018;41:835–842. doi:10.1248/bpb.b18-00133
158. Debbi L, Guo S, Safina D, Levenberg S. Boosting extracellular vesicle secretion. Biotechnol Adv. 2022;59:107983. doi:10.1016/j.biotechadv.2022.107983
159. Zhang Y, Bi J, Huang JA-O, Tang Y, Du S, Exosome LPA-O. A review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomed. 2020;Volume 15:6917–6934. doi:10.2147/IJN.S264498
160. Gao J, Li A, Hu J, Feng L, Liu L, Shen Z. Recent developments in isolating methods for exosomes. Front Bioeng Biotechnol. 2022;10:1100892. doi:10.3389/fbioe.2022.1100892
161. Sivanantham A, Jin Y. Impact of storage conditions on ev integrity/surface markers and cargos. Life. 2022;12(5):697. doi:10.3390/life12050697
162. Ma C, Ding R, Hao K, et al. Storage stability of blood samples for mirnas in glycosylated extracellular vesicles. Molecules. 2023;29(1):103. doi:10.3390/molecules29010103
163. Ye H, Wang F, Xu G, Shu F, Fan K, Wang D. Advancements in engineered exosomes for wound repair: current research and future perspectives. Front Bioeng Biotechnol. 2023;11:1301362. doi:10.3389/fbioe.2023.1301362
164. Boulton AJM, Whitehouse RW. The diabetic foot. 2000. Endotext.
165. Lener T, Gimona M, Aigner L, et al. Applying extracellular vesicles based therapeutics in clinical trials - an ISEV position paper. J Extracellular Ves. 2015;4(1). doi:10.3402/jev.v4.30087
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Loss of OTUD6B Stimulates Angiogenesis and Promotes Diabetic Atherosclerosis
Wang Z, Zhang L, Li L, Zhou M
Diabetes, Metabolic Syndrome and Obesity 2022, 15:3027-3038
Published Date: 28 September 2022
Carrier-Free Binary Self-Assembled Nanomedicines Originated from Traditional Herb Medicine with Multifunction to Accelerate MRSA-Infected Wound Healing by Antibacterial, Anti-Inflammation and Promoting Angiogenesis

Lu J, Wang Z, Cai D, Lin X, Huang X, Yuan Z, Zhang Y, Lei H, Wang P
International Journal of Nanomedicine 2023, 18:4885-4906
Published Date: 30 August 2023
Effect of Diabetes on Wound Healing: A Bibliometrics and Visual Analysis
Lang X, Li L, Li Y, Feng X
Journal of Multidisciplinary Healthcare 2024, 17:1275-1289
Published Date: 20 March 2024
Diverse-Origin Exosomes Therapeutic Strategies for Diabetic Wound Healing

Wang F, Yao J, Zuo H, Jiao Y, Wu J, Meng Z
International Journal of Nanomedicine 2025, 20:7375-7402
Published Date: 12 June 2025
Role of Extracellular Vesicles in Skin Barrier Repair: Applications in Atopic Dermatitis and Chronic Wounds
Khoo L, Chin YZ, Law JX
International Journal of Nanomedicine 2026, 21:598256
Published Date: 8 May 2026