Back to Journals » Breast Cancer: Targets and Therapy » Volume 17
Noncoding RNAs, Vital Players in Breast Cancer Metastasis
Authors Kang Y ![]() , Wang X, Liu Y, Cao J, Lin J, Qiao G
, Wang X, Liu Y, Cao J, Lin J, Qiao G
Received 24 July 2025
Accepted for publication 25 November 2025
Published 30 December 2025 Volume 2025:17 Pages 1463—1492
DOI https://doi.org/10.2147/BCTT.S556045
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Robert Clarke
Yujuan Kang,* Xingmiao Wang,* Yanqing Liu,* Jianqiao Cao,* Jun Lin, Guangdong Qiao
Department of Breast Surgery, The Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jun Lin, Department of Breast Surgery, The Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, People’s Republic of China, Email [email protected] Guangdong Qiao, Department of Breast Surgery, The Affiliated Yantai Yuhuangding Hospital of Qingdao University, Yantai, People’s Republic of China, Email [email protected]
Abstract: Breast cancer (BC) is the most common cause of deaths. Although recent medical advancements have improved survival of BC patients, the occurrence of metastasis increases mortality rate. Noncoding RNAs (ncRNAs) play important roles in BC metastasis (BCM). However, a detailed summary of ncRNAs involved in BCM is unavailable. Therefore, this review summarizes the key biological steps involved in BCM. We discussed the ncRNAs related to BCM according to “seed and soil” theory from four perspectives: (i) ncRNAs that make “breast cancer seed” preferable for BCM; (ii) ncRNAs that make “breast cancer seed” difficult for BCM; (iii) ncRNAs that make “breast cancer soil” preferable for BCM; (iv) ncRNAs that make “breast cancer soil” difficult for BCM. Lastly, we listed the ncRNAs and coding genes involved in BCM that act as initiators or suppressors. This review comprehensively overviews the biological mechanisms underlying BCM. The compiled evidence highlights the role of BCM-associated ncRNAs, which may serve as promising therapeutic targets for BCM.
Keywords: breast cancer, metastasis, noncoding RNAs, EMT, miRNA, circRNA
Introduction
BC is a major cause of cancer-related deaths.1,2 It is characterized by molecular heterogeneity,3,4 and the majority of deaths are associated with metastatic progression.5 Although advances have been made in treatment modalities, BCM remains the primary factor contributing to poor outcomes.6 All molecular types of BC are determined by the expression of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2).7 Triple-negative breast cancer (TNBC), characterized by the lack of ER, PR, and HER2 expression.8,9 The most common locations of MBC include the bone, lungs, liver and brain.10 The pattern of distant metastasis may vary depending on the patient’s age and disease stage at diagnosis.11,12 BC brain metastasis (BCBM) represents a severe clinical complication.13 Metastasis processes typically14 involve migration into adjacent tissues, adaptation to the new biological setting, cellular multiplication, and stimulation of blood vessel formation while avoiding programmed cell termination or defensive reactions from the immune system.15 Metastatic progression is driven by epithelial-mesenchymal transition (EMT).16,17 EMT consists of the transformation of epithelial cells into mesenchymal-like cells, along with changes in shape and function that increase their invasive abilities.18,19 Stem cells possess the dual capacity to maintain their population via self-replication while producing specialized tissue-specific cells through cellular maturation.20 BC stem cells (BCSCs) control BCM.21 The highly tumorigenic properties of stem cells are associated with BCM.22 The tumor microenvironment (TME), mainly consisting of cellular components and release agents such as cytokines and chemokines, stimulates metastasis.23,24 Extracellular matrix (ECM) was destroyed by matrix metalloproteinases (MMPs).25
About 75% of the genetic material is transcribed into RNA, but only around 3% encodes messenger RNAs (mRNAs),26 while the rest of the transcripts encode ncRNAs.27 The main types of ncRNAs include long noncoding RNAs (lncRNAs), circular RNAs (circRNAs), microRNAs (miRNAs), and PIWI-interacting RNAs (piRNAs).28 LncRNAs are transcripts longer than 200 nucleotides (nt)29 usually produced by RNA polymerase II and modified after transcription, resulting in multiple forms.30,31 These molecules involve different functions: epigenetic modifications, controlling transcription, post-transcription, and translation. The competing endogenous RNA (ceRNA) hypothesis, first introduced in 2011.32 It proposes that RNAs sharing microRNA response elements can compete for microRNA binding, affecting the availability of miRNAs and their target mRNAs.33 Interestingly, some lncRNAs can produce functional peptides.34–37 CircRNAs have closed-loop structures and lack 5′ caps and 3′ poly(A) tails.38 They are categorized into exonic, intronic, and exon-intron circRNAs.39 Moreover, miRNAs are short ncRNAs that control gene expression by interacting with 3′ untranslated regions (3′-UTRs) of target mRNAs.40,41 PiRNAs exhibit marginally greater lengths (24–31 nt), originating from single-stranded precursor molecules transcribed from genomic areas known as piRNA clusters.42,43
However, no studies have summarized the related ncRNAs that play vital roles in BCM. Herein, we summarize the key biological steps of BCM from two perspectives: Tumor cells and TME. Secondly, we summarized ncRNAs that function as BCM-promoting and BCM-inhibiting ncRNAs from the following four perspectives: (i) ncRNAs that make “breast cancer seed” preferable for BCM; (ii) ncRNAs that make “breast cancer seed” difficult for BCM; (iii) ncRNAs that make “breast cancer soil” preferable for BCM; (iv) ncRNAs that make “breast cancer soil” difficult for BCM. Finally, we summarized the ncRNAs and coding genes in BCM, and reviewed the related clinical trials and targeted medicines. This review offers an intuitive display of the landscape of ncRNAs involved in BCM. This review sheds light on the new directions for future BCM therapies.
BCM Mechanisms
Malignant cells proliferate at their original location, then penetrate the ECM and enter the bloodstream, subsequently migrating to distant tissues where they establish secondary growths.44 A hypothesis emphasized the crucial interplay between mobile tumor cells (“seeds”) and the organ microenvironments (“soil” or TME) was purposed by Stephen Paget.45 Metastasis depends on tumor cells (seed) and TME (soil).46,47 The TME can be divided into cellular, soluble, and physical components.48 In BC, TME comprises a network of cellular populations, including carcinoma-associated fibroblasts (CAFs), various immune cell subsets, pericytes, adipocytes, endothelial cells, and bone marrow-derived cells.49–51 Besides cellular mediators, the TME contains multiple non-cellular (soluble mediators), ie, chemokines, growth factors, cytokines, and MMPs, and insoluble factors, ie, exosomes and the ECM.52 CAFs are crucial abundant stromal cell types within TME.53 These fibroblasts are actively involved in the formation and rejuvenation of the ECM,54 and they contribute to ECM degradation through the secretion of proteolytic enzymes (MMPs and urokinase-type plasminogen activators).55,56 CAFs induce tumor metastasis through multiple mechanisms, such as the release of paracrine signaling molecules, ECM deposition and restructuring, and induction of metabolic changes within TME.57,58 Tumor-associated macrophages (TAMs) exert pro-tumorigenic functions.59,60 M1 and M2 macrophages occupying opposing ends.61,62 Classically activated M1 macrophages was activated by interferon-γ and tumor necrosis factor (TNF). They display anticancer properties by secreting inflammatory mediators (TNF and interleukin-2 [IL-2]) along with reactive nitrogen and oxygen species.63,64 Conversely, alternatively activated M2 macrophages respond to IL-4, IL-10, and IL-13, developing tumor-promoting biological behaviors.64 BC is characterized by a large proportion of M2 phenotype.65 M1/M2 macrophage polarization has tumor-promoting capabilities, including metastasis and immunosuppression.66 Neutrophils have anti-tumorigenic (“N1”) and pro-tumorigenic (“N2”) phenotypes.67 Extracellular vesicles (EVs) are divided into three types: exosomes (30–100 nm), apoptotic bodies (1000–5000 nm), and microvesicles (MVs; 100–1000 nm).68 The proposed mechanisms and functional importance of BCM are shown in Figure 1 and Table 1.
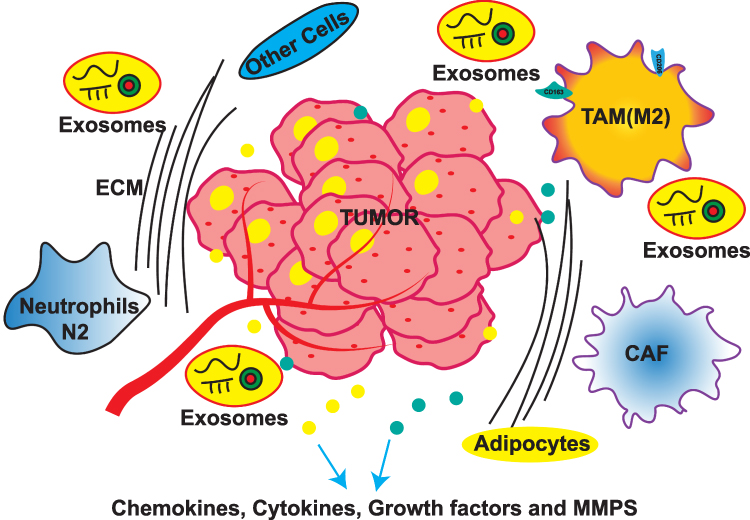 |
Figure 1 The biological composition of breast cancer metastasis (BCM). |
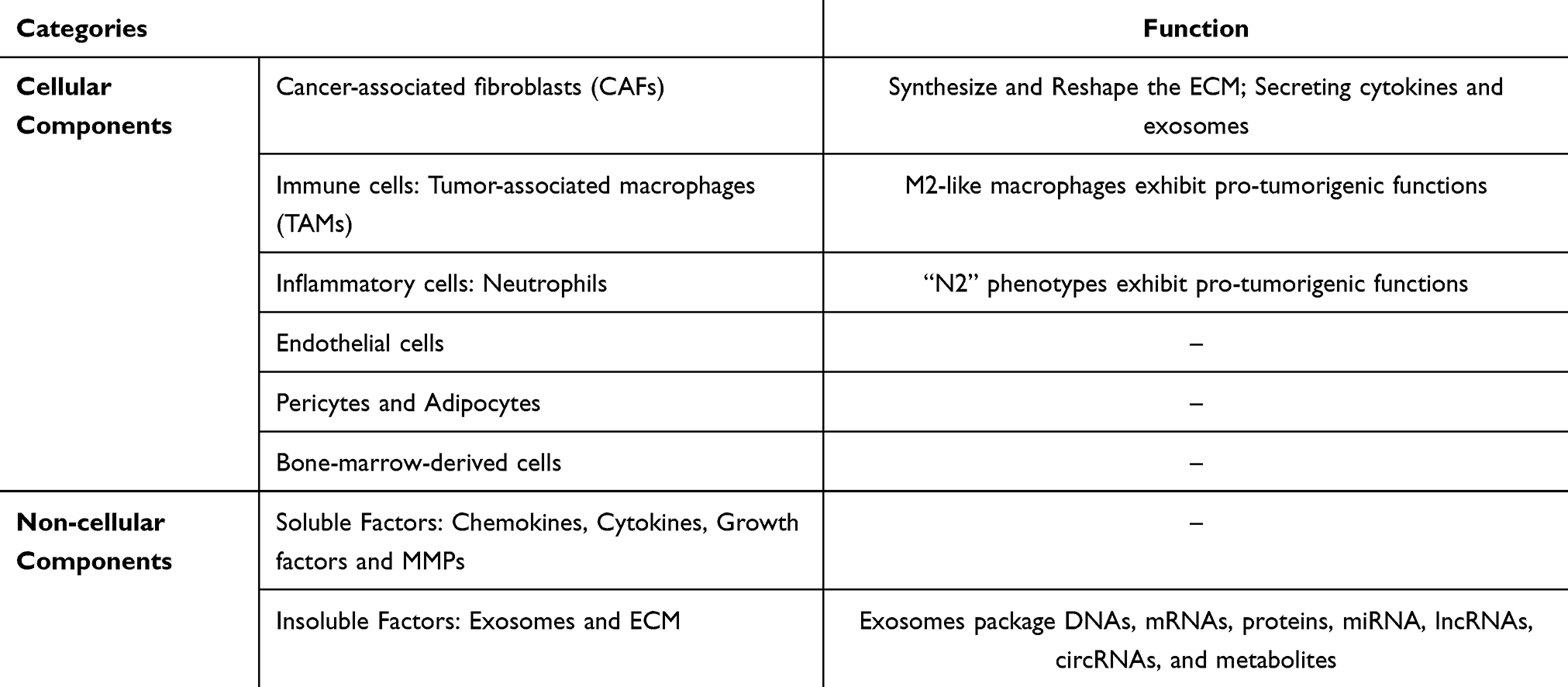 |
Table 1 The Components of the Breast Tumor Microenvironment (TME) |
NcRNAs Involved in BCM
NcRNAs affect the behavior of tumor cells and TME involved in BCM. We summarized the ncRNAs involved in BCM: (i) ncRNAs that make “breast cancer seed” preferable for BCM; (ii) ncRNAs that make “breast cancer seed” difficult for BCM; (iii) ncRNAs that make “breast cancer soil” preferable for BCM; (iv) ncRNAs that make “breast cancer soil” difficult for BCM.
NcRNAs that Make “Breast Cancer Seed” Preferable for BCM
NcRNAs promote “breast cancer seed” preferable for BCM through various means to promote BC cell migration and invasion, EMT, angiogenesis, and BCSCs malignant behavior (Figures 2 and 3, Table 2).
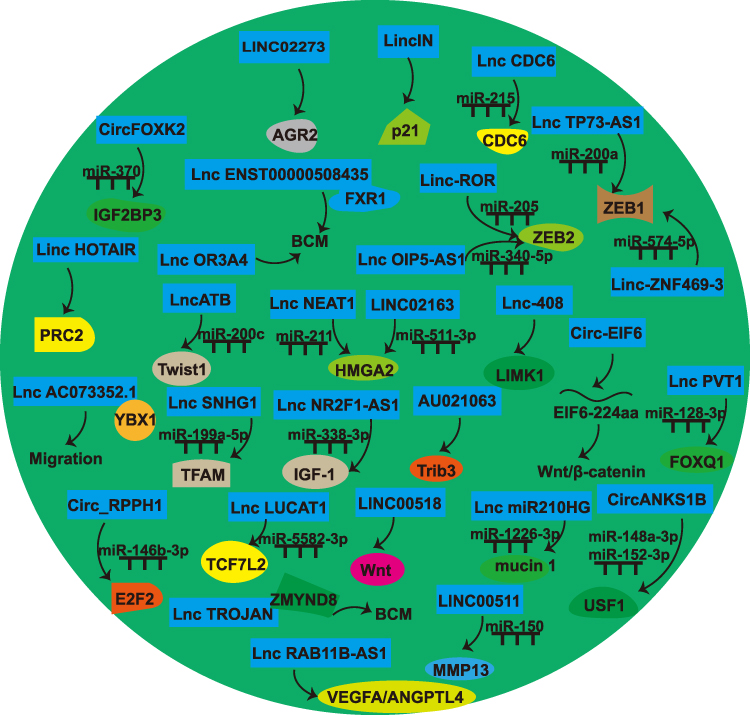 |
Figure 2 Noncoding RNAs (ncRNAs) [microRNAs (miRNAs) excluded] that make “breast cancer seed” preferable for BCM. |
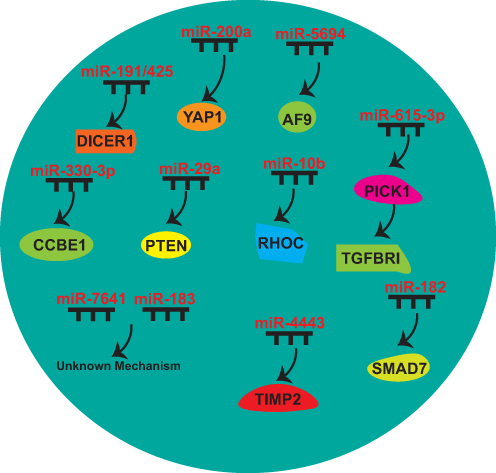 |
Figure 3 MiRNAs that make “breast cancer seed” preferable for BCM. |
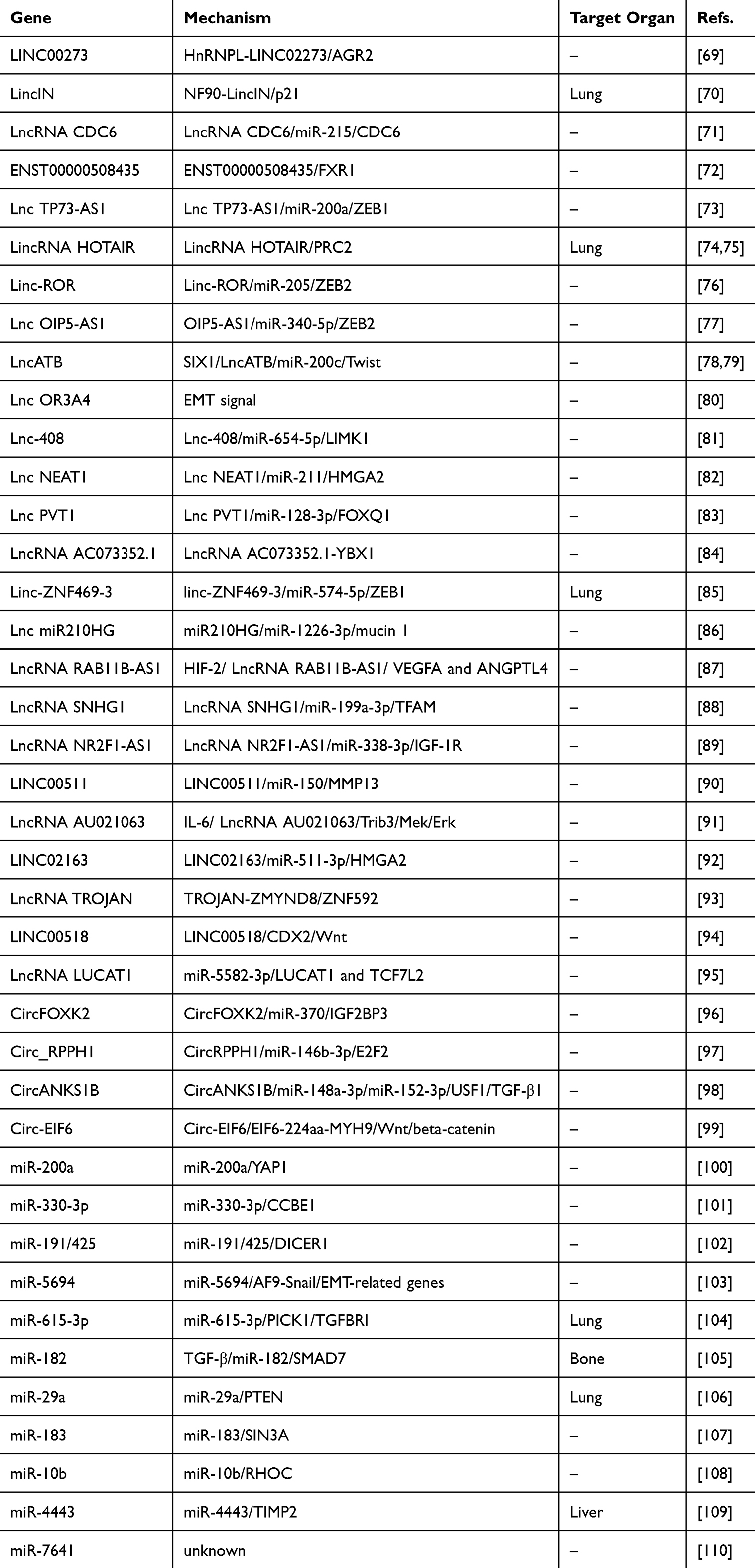 |
Table 2 Noncoding RNAs (ncRNAs) That Make “Breast Cancer Seed” Preferable for Breast Cancer Metastasis (BCM) |
LINC02273
HnRNPL-LINC02273 promotes BCM. HnRNPL-LINC02273 recruitment to the AGR2 promoter region upregulates AGR2 by augmenting H3K4me3 and H3K27ac expression.69
LincIN
LincIN knockdown reduces lung metastasis. Elevated lincIN suppress p21 protein synthesis through translational inhibition, while reduced lincIN expression correlates with increased p21 abundance.70
LncRNA CDC6
Overexpression of lncCDC6 has been shown to enhance migration. It acts as a ceRNA by interacting with miR‐215, thus preventing it from suppressing CDC6 mRNA.71
LncRNA ENST00000508435
LncRNAENST00000508435 substantially enhances migration. It directly interacts with FXR1, promoting BCM.72
LncRNA TP73-AS1
TP73-AS1 enhanced ZEB1 levels through molecular competition, specifically by binding to miR-200a instead of allowing it to interact with the 3’-UTR region of ZEB1 mRNA.73
LincRNA HOTAIR
HOTAIR promotes EMT lung metastasis.74 Elevated HOTAIR in malignant epithelial cells alter the genomic distribution patterns of polycomb repressive complex 2 (PRC2), while promoting tumor dissemination.75
Linc-ROR
Linc-ROR enhances BC migration and invasion. Linc-ROR functions as a ceRNA of miR-205. Linc-ROR prevents miR-205 target gene (ZEB2) degradation.76
LncRNA OIP5-AS1
LncRNA OIP5‑AS1 facilitates BCM by sponging miR‑340‑5p to enhance the ZEB2 mRNA transcription.77
LncATB
LncATB enhances EMT and migration. This molecule sequestered miR-200 cluster members while upregulating Twist1 transcription.78 SIX-1 promotes lncATB transcription, which plays a pro-metastatic role by directly interacting with miR-200c to promote BCM.79
LncRNA OR3A4
LncRNA OR3A4 was overexpressed in BC. OR3A4 silencing suppressed BC cell proliferation by repressing metastasis via EMT.80
Lnc-408
Lnc-408 acts as a ceRNA by sponging miR-654-5p, thus increasing LIMK1 levels. Overexpression of Lnc-408 increases LIMK1 levels, therefore enhancing the invasive potential of BC cells.81
LncRNA NEAT1
LncRNA NEAT1 promotes EMT. A bidirectional inhibitory relationship between NEAT1 and miR-211 was discovered. Studies identified HMGA2, a known EMT promoter, as directly regulated by miR-211.82
LncRNA PVT1
LncRNA PVT1 promotes EMT and BCM. It elevates FOXQ1 expression through the suppression of miR-128-3p and interacts with UPF1.83
LncRNA AC073352.1
LncRNA AC073352.1 promotes BC cell invasive behavior, contributing to BCM by directly interacting with YBX1.84
Linc-ZNF469-3
Linc-ZNF469-3 enhances lung metastasis. Mechanistically, it interacts with miR-574-5p, and restoring miR-574-5p levels decreases ZEB1 level.85
LncRNA miR210HG
LncRNA miR210HG involved in promoting BCM. It acts as a ceRNA, sequestering miR-1226-3p and thus increasing the level of mucin 1.86
LncRNA RAB11B-AS1
RAB11B-AS1 promotes metastatic potential by upregulating angiogenesis. Under hypoxic conditions, it increases the expression of vascular endothelial growth factor A (VEGFA) and angiopoietin-like 4 (ANGPTL4).87
LncRNA SNHG1
SNHG1 contributes to metastasis through the HIF-1α/SNHG1/miR-199a-3p/ mitochondrial transcription factor A (TFAM) signaling axis. It downregulates miR-199a-3p and simultaneously upregulates TFAM, supporting metastatic under hypoxic conditions.88
LncRNA NR2F1-AS1
LncRNA NR2F1-AS1 promotes BC angiogenesis. LncRNA NR2F1 sponges miR-338-3p to induce IGF-1, further activating the IGF-1R and ERK pathway.89
LINC00511
LINC00511 regulates MMP13 expression by sponging miR-150, promoting cell migration.90
LncRNA AU021063
IL-6 induces the level of lncRNA AU021063. AU021063 promotes BCM by stabilizing Trib3 and stimulating the Mek/Erk signaling pathway.91
LINC02163
LINC02163 knockdown inhibits migration. LINC02163 is a ceRNA of miR-511-3p and upregulates HMGA2 expression.92
LncRNA TROJAN
LncRNA TROJAN promotes invasion. TROJAN interacts with ZMYND8, accelerating its proteasomal breakdown via competitive displacement of ZNF592.93
LINC00518
LINC00518 induces CDX2 methylation by recruiting DNA methyltransferases, stimulating Wnt signaling pathway. Suppression of LINC00518 attenuates invasion and EMT.94
LncRNA LUCAT1
LUCAT1 was overexpressed in BCSCs. LUCAT1 promotes BCSC proliferation. miR-5582-3p directly binds to LUCAT1 and TCF7L2 and adversely regulates their level.95
CircFOXK2
CircFOXK2 is overexpressed in MBC. Its pro-metastatic function is related to regulating IGF2BP3 and miR-370.96
Circ_RPPH1
circRPPH1 enhances metastatic potential in BC by functioning as a ceRNA for miR-146b-3p, leading to E2F2 upregulation.97
CircANKS1B
CircANKS1B enhances BC invasion and EMT. It functions as a molecular sponge for miR-152-3p and miR-148a-3p, thus increasing USF1. USF1 induces transforming growth factor β1 (TGF-β1) transcription, stimulating the TGF-β1/Smad signaling cascade, which drives EMT.98
Circ-EIF6
Circ-EIF6 induces metastasis and encodes a new peptide, EIF6-224aa. This peptide binds to MYH9, inhibits its degradation, and stimulates the Wnt/β-catenin signaling pathway.99
miR-200a
miR-200a enhances BCM. Yes-associated protein 1 (YAP1) has been identified as a target of miR-200a. Its knockdown replicates the pro-metastatic effects observed with miR-200a overexpression, whereas YAP1 restoration reverses these effects in miR-200a-overexpressing BC cells.100
miR-330-3p
Elevated level of miR-330-3p in BC cells enhances their invasive capacity. CCBE1 has been detected as a target of miR-330-3p.101
miR-191/425
Overexpression of miR-191 or miR-425 promotes BC migration, relying on the miR-191/425-induced DICER1 suppression.102
miR-5694
miR-5694 binds to the 3′-UTR of AF9 mRNA, destabilizing it. AF9 interacts with Snail to suppress its transcriptional activity and facilitates the recruitment of CBP or GCN5, thus establishing an active chromatin state at the promoters of EMT-associated genes.103
miR-615-3p
miR-615-3p enhances lung metastasis. miR-615-3p targets the 3′-UTR of PICK1, resulting in the increased TGF-β type I receptor (TGFBRI).104
miR-182
TGF-β stimulates the expression of miR-182, which inhibits SMAD7. Elevated levels of miR-182 promote invasion and enhance TGF-β-induced osteoclastogenesis, contributing to bone metastasis.105
miR-29a
miR-29a promotes lung metastasis. PTEN serves as a primary miR-29a-regulated gene, triggering EMT through activation of AKT pathway signaling.106
miR-183
miR-183 facilitate migration and invasion depending on the existence of SIN3A, and ectopic miR-183 promotes BCM.107
miR-10b
Twist-induced miR-10b inhibited the translation of mRNA encoding homeobox D10, leading to increased RHOC levels.108
miR-4443
miR-4443 promotes BCM through tissue metalloproteinase 2 (TIMP2) suppression and MMPs activation. Coordinated TIMP2 reduction and MMP2 elevation following miR-4443 overexpression, establishing a mechanistic link to liver metastasis.109
miR-7641
miR-7641 promotes BCM, with its biological effects transmitted to target cells through exosomal transfer mechanisms.110
NcRNAs that Make “Breast Cancer Seed” Difficult for BCM
NcRNAs promote “breast cancer seed” difficult for BCM by various means, including inhibiting BC cell migration and invasion ability, EMT, and BCSC malignant behavior (Figure 4 and Table 3).
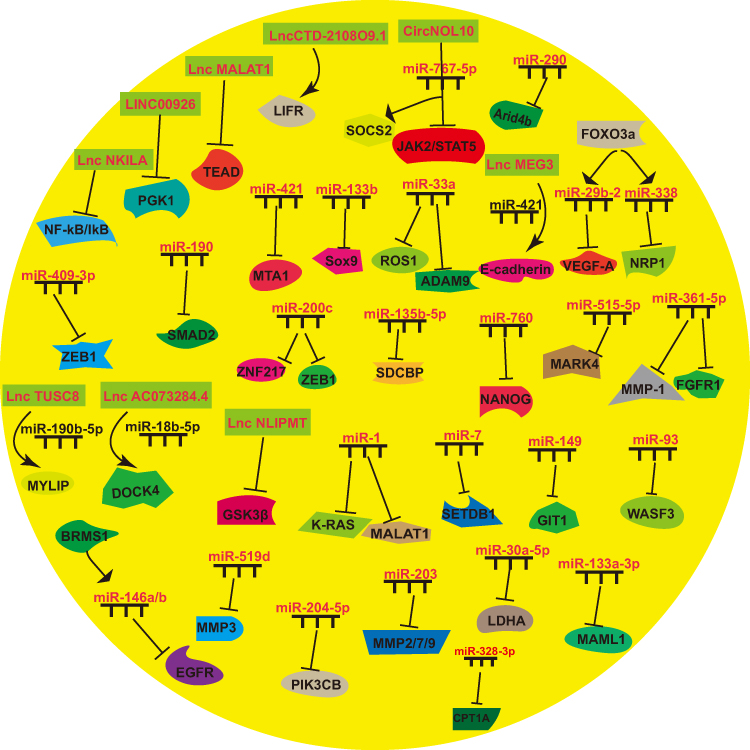 |
Figure 4 NcRNAs that make “breast cancer seed” difficult for BCM. |
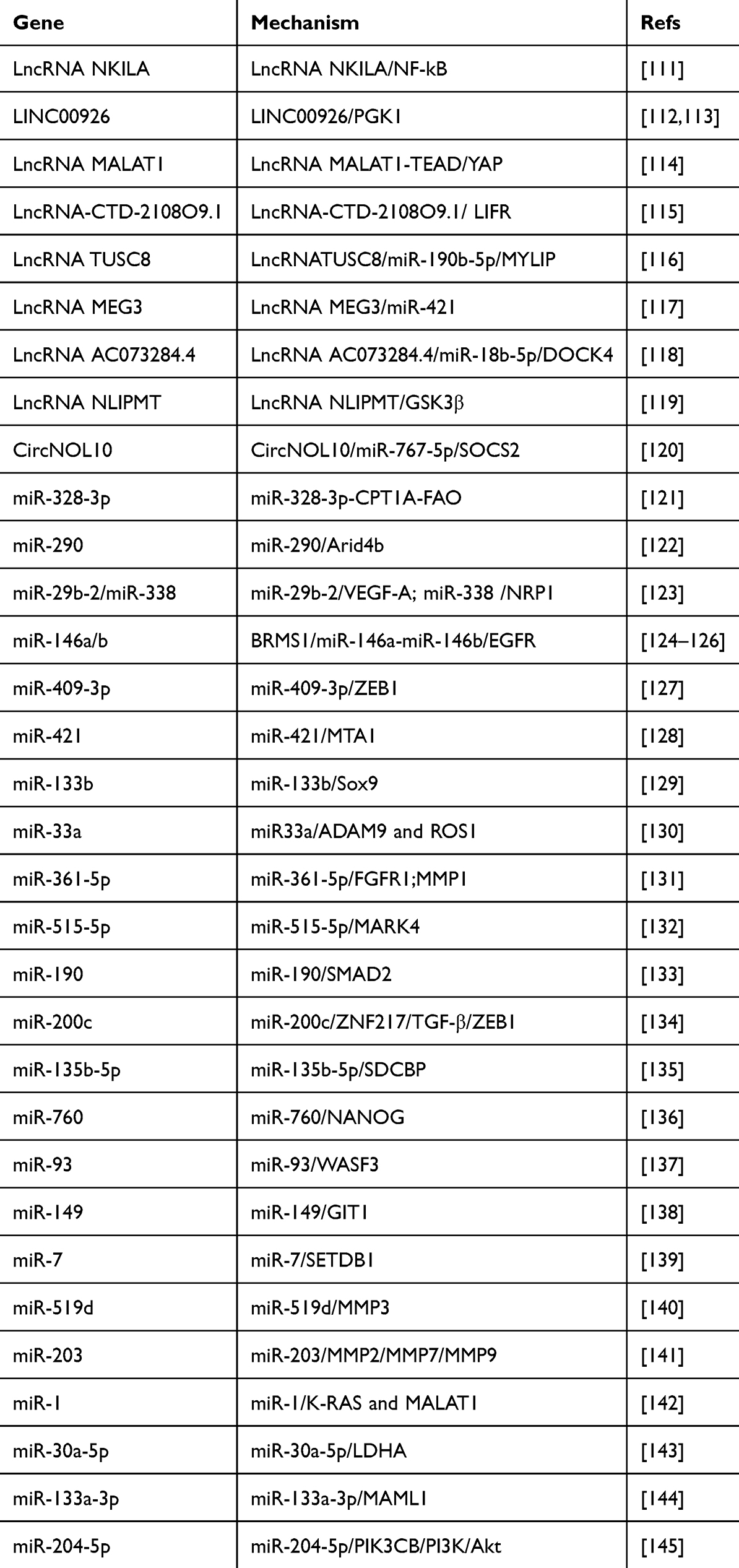 |
Table 3 NcRNAs That Make “Breast Cancer Seed” Difficult for BCM |
LncRNA NKILA
Low NKILA is associated with BCM. NKILA, transcriptionally activated by nuclear factor kappa B (NF-κB), interacts with the NF-κB/IκB complex and physically obstructs critical phosphorylation sites on IκB. This molecular interference prevents the phosphorylation of IκB by IKK, suppressing NF-κB signaling.111
LINC00926
PGK1 mediates glycolysis.112 LINC00926 inhibits BCM by inhibiting PGK1 expression.113
LncRNA MALAT1
MALAT1 inhibits BCM. MALAT1 interacts with the TEAD, effectively blocking its ability to form complexes with YAP.114
LncRNA-CTD-2108O9.1
LncRNA-CTD-2108O9.1 is a BCM inhibitor targeting the leukemia inhibitory factor receptor (LIFR), a BCM suppressor.115
LncRNA TUSC8
Silencing TUSC8 promotes BC cell migration. TUSC8 acts as a ceRNA for myosin regulatory light chain interacting protein (MYLIP) by sequestering miR-190b-5p, thus modulating the level of EMT-related markers and inhibiting BCM.116
LncRNA MEG3
MEG3 suppresses invasion. MEG3 inhibits miR-421 and E-cadherin expression by acting as a ceRNA, which could sponge miR-421.117
LncRNA AC073284.4
AC073284.4 functions as a molecular sponge for miR-18b-5p, thus enhancing the expression of dedicator of cytokinesis protein 4 (DOCK4). This regulatory interaction suppresses invasion, metastasis, and EMT.118
LncRNA NLIPMT
LncRNA NLIPMT inhibits cell motility. Glycogen synthase kinase 3β (GSK3β) is a targeted protein regulated by lncNLIPMT.119
CircNOL10
CircNOL10 suppresses EMT by acting as a molecular sponge for miR-767-5p. This interaction elevates suppressor of cytokine signaling 2 (SOCS2) and suppresses the JAK2/STAT5 signaling axis.120
miR-328-3p
miR-328-3pmiR-328-3p-CPT1A-FAO axis involved in BCM. miR-328-3p upregulation or interruption of this axis can be exploited to develop efficient strategies to decrease metastasismiR-328-3p.121
miR-290
miR-290 exerts its tumor-suppressive function by targeting Arid4b, promoting apoptotic pathways, ultimately inhibiting BC progression.122
miR-29b-2/miR-338
Overexpression of miR-29b-2 and miR-338 inhibits EMT. miR-29b-2 targets VEGFA, while miR-338 suppresses NRP1. These regulatory interactions mediate the inhibitory effect of FOXO3a on the VEGFA/NRP1 signaling axis, to reduce metastatic potential.123
miR-146a/b
BCM suppressor 1 (BRMS1) reduces BCM.124,125 BRMS1 upregulates the expression of miR-146a and miR-146b. These microRNAs suppress EGFR signaling, thus inhibiting BCM.126
miR-409-3p
miR-409-3p overexpression impairs BC cell migration and invasion. miR-409-3p interacts with the 3′-UTR of ZEB1.127
miR-421
miR-421 suppresses BCM. miR-421 suppress the 3′-UTR of metastasis-associated 1 (MTA1), and miR-421 suppresses BCM by inhibiting MTA1.128
miR-133b
miR-133b is a tumor suppressor. Downregulated miR-133b levels participate in BC progression by targeting Sox9, which regulates BCM.129
miR-33a
miR-33a overexpression decreases cell invasion and inhibits lung metastasis. ADAM9 and ROS1 are directly regulated by miR-33a.130
miR-361-5p
miR-361-5p suppresses BCM. Fibroblast growth factor receptor 1 (FGFR1) is a miR-361-5p target. Moreover, miR-361-5p inhibits invasion and metastasis by targeting MMP1.131
miR-515-5p
miR-515-5p inhibits MARK4 through direct 3′-UTR interaction involving migration. miR-515-5p/MARK4 signaling inhibits metastasis.132
miR-190
miR-190 inhibits BCM by directly targeting SMAD2.133
miR-200c
miR-200c impedes invasive ability by targeting ZNF217, a transcriptional enhancer of TGF-β and ZEB1, a downstream effector within the TGF-β signaling axis. miR-200c/ZEB1 and miR-200c/ZNF217/TGF-β/ZEB1 interactions that facilitate metastasis.134
miR-135b-5p
Syndecan-binding protein (SDCBP) has been identified as a downstream effector of miR-135b-5p. Suppression of miR-135b-5p is correlated with enhanced EMT and increased invasion.135
miR-760
miR-760 suppresses the BC stem cell population by inactivating the NANOG, thus restraining metastatic progression.136
miR-93
Elevated level of miR-93 suppresses invasiveness and metastatic potential. Wiskott-Aldrich syndrome protein family member 3 (WASF3), a key modulator of cytoskeletal reorganization and CSC characteristics, is a direct target of miR-93.137
miR-149
miR-149 plays an anti-metastatic role by negatively regulating G protein-coupled receptor kinase-interacting protein 1 (GIT1). miR-149 also disrupts fibronectin-induced focal adhesion assembly, an effect that can be reversed through the restoration of GIT1.138
miR-7
miR-7 is downregulated in BCSCs. It inhibits invasion and metastatic behavior, reduces the BCSC population, and partially counteracts EMT by directly targeting the SETDB1.139
miR-519d
miR-519d overexpression suppresses migration. MMP3 is a direct target of miR-519d.140
miR-203
miR-203 inhibits BC invasion. Furthermore, miR-203 demonstrates anti-metastatic properties by significantly reducing the production of MMP2, MMP7, and MMP9.141
miR-1
miR-1 acts as a tumor suppressor by targeting KRAS and MALAT1. Restoration of miR-1 suppresses metastasis.142
miR-30a-5p
miR-30a-5p negatively regulates LDHA by targeting its 3′-UTR. By suppressing LDHA-mediated glycolysis, miR-30a-5p attenuates BCM.143
miR-133a-3p
miR-133a-3p is epigenetically silenced due to DNA hypermethylation. Mastermind-like transcriptional co-activator 1 (MAML1) has been identified as a direct target of miR-133a-3p.144
miR-204-5p
miR-204-5p acts as a tumor suppressor and is involved in BCM progression. It directly targets PIK3CB and modulates downstream PI3K/Akt signaling pathways.145
NcRNAs that Make “Breast Cancer Soil” Preferable for BCM
NcRNAs promote “breast cancer soil” preferred for BCM by various means to promote M2 macrophage polarization, CAF signaling pathway, and N2 conversion (Figure 5 and Table 4).
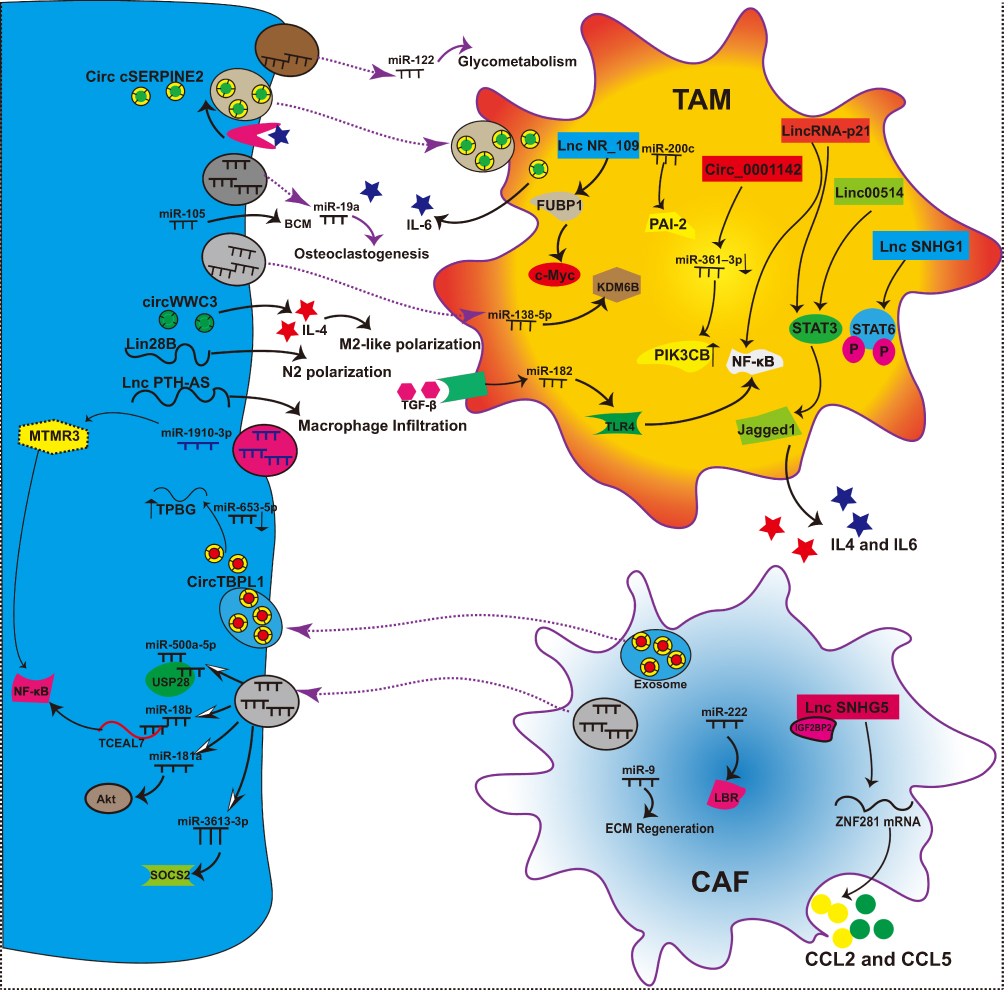 |
Figure 5 NcRNAs that make “breast cancer soil” preferable for BCM. |
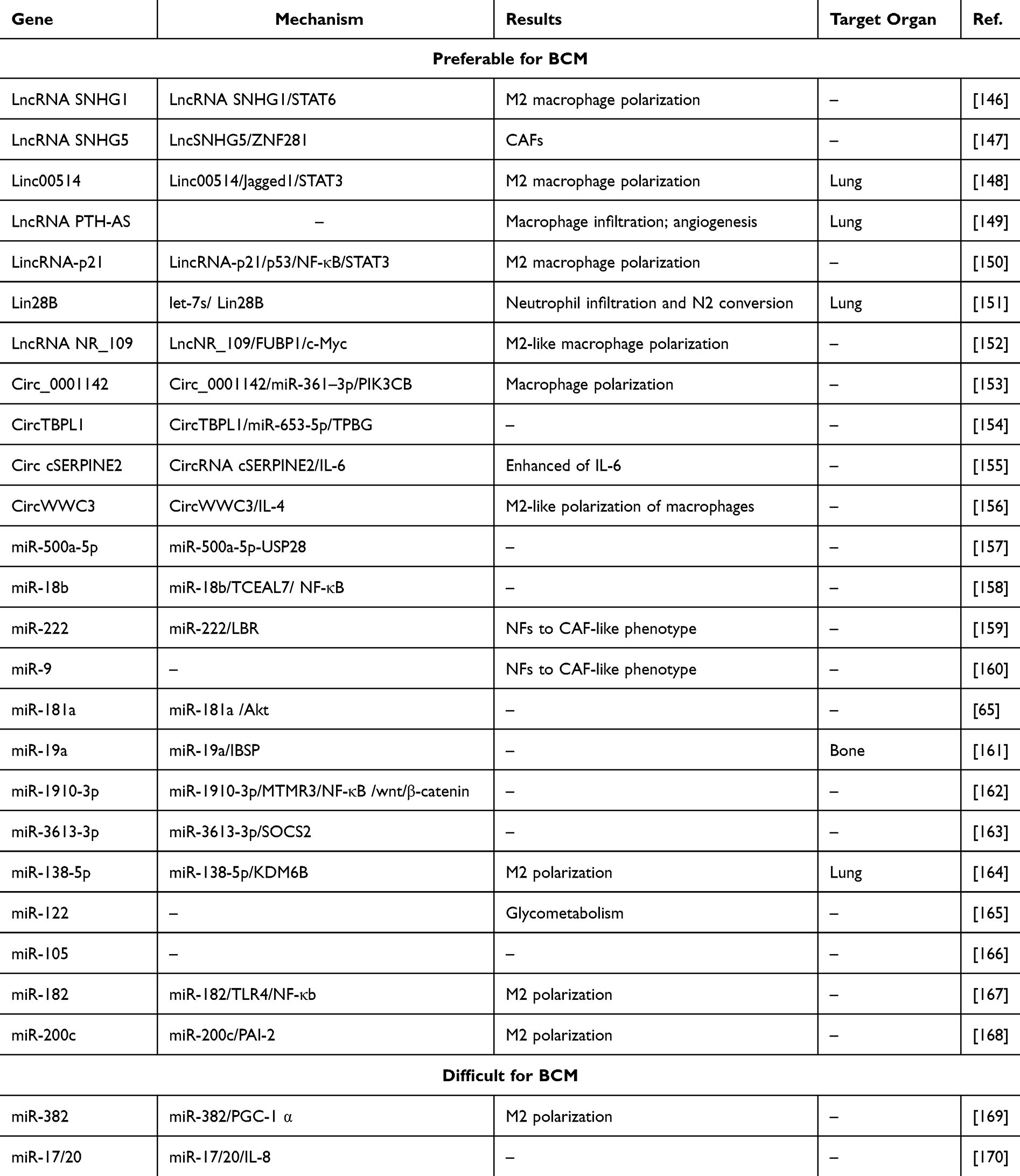 |
Table 4 NcRNAs That Make “Breast Cancer Soil” Preferable and Difficult for BCM |
LncRNA SNHG1
SNHG1 acts as a regulatory factor in M2 macrophage polarization. Its knockdown suppresses M2 polarization by inhibiting STAT6 phosphorylation and impairs the migratory ability.146
LncRNA SNHG5
SNHG5 is overexpressed in CAFs and plays a critical role in premetastatic niche formation by promoting angiogenesis. SNHG5 interacts with IGF2BP2, stabilizing ZNF281 mRNA. The upregulation of ZNF281 transcriptionally enhances CCL2 and CCL5, activating the P38 MAPK signaling cascade.147
Linc00514
Linc00514 promotes lung metastasis. Linc00514 increases the population of M2-polarized macrophages. Linc00514 upregulates Jagged1 by facilitating STAT3 phosphorylation, thus activating the Jagged1-mediated Notch. This activation elevated secretion of IL-4 and IL-6, and M2 macrophage polarization.148
LncRNA PTH-AS
LncRNA PTH-AS enhances invasion and lung metastasis. PTH-AS demonstrate elevated macrophage infiltration, accelerating angiogenesis.149
LincRNA-p21
LincRNA-p21 is highly expressed in macrophages. Its silencing promotes macrophage polarization toward the M1 phenotype, a process driven by MDM2-mediated proteasomal degradation of p53 and activating NF-κB and STAT3.150
Lin28B
Lin28B promotes an immunosuppressive pre-metastatic environment by stimulating neutrophil penetration and polarization toward the N2 phenotype, mediated by tumor-derived exosomes with reduced let-7s levels. It has been shown to facilitate BCM by modulating immune suppression within the metastatic lung microenvironment.151
LncRNA NR_109
NR_109 induces macrophage polarization toward an M2-like phenotype and participates in a positive feedback loop involving NR_109, FUBP1, and c-Myc, which drives its role in shaping a pro-tumorigenic TME.152
Circ_0001142
Endoplasmic reticulum stress facilitates exosome release and promotes the transfer of circ_0001142 into macrophages, thus modulating their polarization. Circ_0001142/miR-361-3p/PIK3CB axis is pivotal in controlling macrophage polarization.153
CircTBPL1
Exosomal circTBPL1, derived from CAFs, enhances migration. Mechanistically, circTBPL1 protects TPBG from miR-653-5p–mediated degradation. The CAF-derived exosomal circTBPL1/miR-653-5p/TPBG regulatory axis is crucial in BC advancement.154
CircRNA cSERPINE2
CircRNA cSERPINE2 is elevated in BC. Exosomal cSERPINE2 shuttles TAMs and enhances IL-6 secretion and invasion. Furthermore, IL-6 elevates intracellular concentrations of EIF4A3 and CCL2, thus amplifying cSERPINE2 production while simultaneously promoting macrophage infiltration.155
CircWWC3
CircWWC3 elevates IL-4 levels. The increased of IL-4 enhances the macrophages’ polarization into an M2-like phenotype within the TME, thus promoting migration.156
miR-500a-5p
miR-500a-5p is found at high levels in BC cells after treatment with exosomes released by CAFs. miR-500a-5p is passed from CAFs to BC cells, promoting metastasis by interacting with ubiquitin-specific peptidase 28 (USP28).157
miR-18b
miR-18b is elevated in exosomes secreted by CAFs, and exosomal miR-18b accelerates the metastatic behavior by targeting the 3′-UTR of transcription elongation factor A like 7 (TCEAL7). The miR-18b-TCEAL7 signaling enhances the nuclear translocation of Snail by activating the NF-κB pathway, consequently initiating EMT.158
miR-222
miR-222 is increased in CAFs relative to normal fibroblasts (NFs). Lamin B receptor (LBR) is a target of miR-222 and is involved in fibroblast reprogramming. Both miR-222 overexpression and LBR depletion convert NFs into a CAF-like phenotype.159
miR-9
miR-9, which is upregulated in BC cell. Tumor-derived miR-9 is transferred to NFs via exosomes, inducing their reprogramming into CAFs. NFs transiently transfected with miR-9 revealed altered expression of genes primarily involved in cell motility and ECM regeneration.160
miR-181a
Exosomal miR-181a, transferred by monocytes activated through interaction with CAFs, contributes to BC progression by partially activating the Akt.65
miR-19a
ER+ BC cells actively secrete high levels of exosomal miR-19a and integrin-binding sialoprotein (IBSP). IBSP establishes an osteoclast-rich microenvironment. This environment facilitates the targeted delivery of exosomal miR-19a to osteoclasts, thus promoting osteoclastogenesis.161
miR-1910-3p
Exosomes transfer miR-1910-3p to BC cells, inhibit myotubularin-related protein 3 expression (MTMR3), and activate the NF-κB and wnt/β-catenin, promoting BCM.162
miR-3613-3p
miR-3613-3p upregulation has been observed in CAF-secreted exosomes. Exosomal miR-3613-3p accelerates metastatic potential, whereas its downregulation in CAF-derived exosomes suppresses BCM by affecting SOCS2.163
miR-138-5p
Exosomal miR-138-5p promotes M2 polarization of macrophages by downregulating KDM6B. Macrophages exposed to exosomal miR-138-5p promote lung metastasis.164
miR-122
miR-122 promotes metastatic progression. miR-122 disrupts glucose processing in target niche cells through downregulation of pyruvate kinase, a pivotal enzyme in glycolysis.165
miR-105
Increased miR-105 are involved in metastasis. Therapeutic inhibition of miR-105 demonstrates potent suppression of metastatic progression.166
miR-182
miR-182 promotes TAM polarization toward the M2 phenotype. miR-182 further downregulates TLR4, thus inactivating NF-κB signaling and enhancing M2 polarization of TAMs.167
miR-200c
miR-200c/PAI-2 axis contributes to BCM by stimulating alternative activation of TAMs. This axis promotes M2 polarization via induction of IL-10 expression.168
NcRNAs that Make “Breast Cancer Soil” Difficult for BCM
NcRNAs promote “breast cancer soil” difficult for BCM by various means to inhibit M2 macrophage polarization (Table 4).
miR-382
Tumor cells suppress miR-382 in TAMs, leading to the inhibition of the target PGC-1α. The upregulation of PGC-1α alters the metabolic profile of TAMs, promoting their polarization into the M2 phenotype. These M2-polarized TAMs secrete TGF-β and IL-10, facilitating EMT and enhancing metastatic progression.169
miR-17/20
miR-17/20 is downregulated in BC. miR-17/20 directly targets the 3′-UTR of IL-8 mRNA, reducing IL-8 levels. miR-17/20 also downregulates cytokeratin 8 level via cyclin D1 modulation.170
NcRNAs and Coding Genes Involved in BCM
NcRNAs are involved in BCM through various mechanisms. As previously described, some of these are black sheep that promote BCM, including LINC02273, LincIN, Lnc CDC6, ENST00000508435, LncTP73-AS1, LincRNA HOTAIR, Lnc-ROR, Lnc OIP5-AS1, LncATB, Lnc OR3A4, Lnc-408, Lnc NEAT1, LncPVT1, CircFOXK2, Circ_RPPH1, miR-200a, miR-330-3p and so on. NcRNAs that inhibit BCM include LncRNA NKILA, LINC00926, LncRNA MALAT1, LncRNA-CTD-2108O9.1, LncRNA TUSC8, LncRNA MEG3, LncRNA AC073284.4, LncRNA NLIPMT, CircNOL10, miR-290, miR-29b-2/miR-338, miR-146a/b and so on (Table 5).
 |
Table 5 NcRNAs Involved in BCM |
We reviewed these ncRNA related clinical trials, however, there are currently no clinical trials available. In the vast majority of cases, ncRNAs often targeting downstream coding genes for certain regulating purposes. Then we summarized studies provide us with a clear understanding of the coding genes involved in “breast cancer seed” related to BCM which include: AGR2; P21; CDC6; FXR1; ZEB1; PRC2; ZEB2; Twist1; SIX1; LIMK1; HMGA2; FOXQ1; YBX1; mucin 1; VEGFA/ANGPTL4; TFAM; IGF-1; MMP13; Trib3; ZNF592; CDX2; LUCAT1/TCF7L2; IGF2BP3; E2F2; USF1; MYH9; YAP1; CCBE1; DICER1; AF9; PICK1; SMAD7; PTEN; CPT1A; SIN3A; RHOC; TIMP2; NF-Kb; PGK1; TEAD; LIFR; MYLIP; E-cadherin; DOCK4; GSK3β; SOCS2/JAK2/STAT5; Arid4b; VEGFA/NRP1; EGFR; MTA1; Sox9; ADAM9/ROS1; FGFR1/MMP1; MARK4; SMAD2; SDCBP; NANOG; WASF3; GIT1; SETDB1; MMP3; MMP2/7/9; K-RAS/MALAT1; LDHA; MAML1 and PIK3CB. Then we summarized studies provide us with a clear understanding of the coding genes involved in “breast cancer soil” related to BCM which include: STAT6; ZNF281; Jagged1/STAT3; NF-Κb/STAT3; let-7s; FUBP1; PIK3CB; TPBG; IL-6; IL-4; USP28; TCEAL7; LBR; Akt; IBSP; MTMR3; SOCS2; KDM6B; TLR4; PAI-2; PGC-1α; IL-8 (Figure 6 and Table 6).
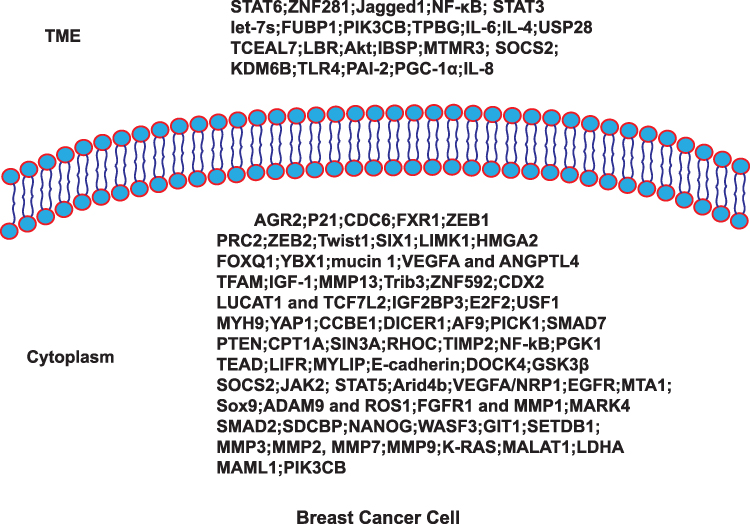 |
Figure 6 Coding genes involved in BCM. |
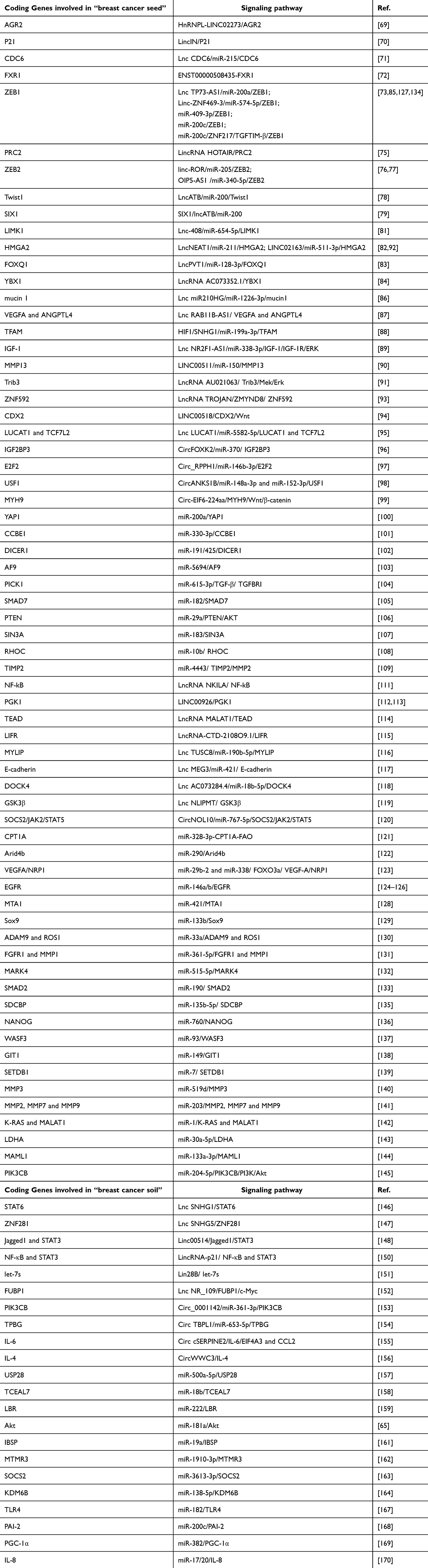 |
Table 6 Coding Genes Involved in “Breast Cancer Seed and Soil” Related to BCM |
We reviewed that the related clinical trials to these coding genes which include: JAK/STAT, mucin 1, VEGFA, IGF-1, GSK3β, EGFR, ROS1 and PIK3CB. The related targeted medicines in the clinical trials are as follows: JAK2 inhibitor: Ruxolitinib; STAT3 inhibitor: TTI-101 and silibinin; mucin 1 related vaccinia: Recombinant Vaccinia Virus That Expresses DF3/MUC1, HuMNC2-CAR44 CAR T cells or huMNC2-CAR22 CAR T cells, Recombinant vaccinia-MUC1 vaccine and so on; VEGFA inhibitor: BNT327, Aflibercept, Bevacizumab, BNT323, Apatinib, Lucitanib, erlotinib; IGF-1 inhibitor: [225Ac]-FPI-1434; GSK3β inhibitor: 9-ING-41; EGFR inhibitor: SCT200, TAS2940, Poziotinib, Lapatinib, SYS6010, AZD4547, Gefitinib; ROS1 inhibitor: Entrectinib, Taletrectinib; PIK3CB inhibitor: AZD8186 (Supplemental Table 1).
Conclusion
BC is the leading disease among women. Despite comprehensive treatment methods, metastasis still posing a serious threat to life. BCM is involved in various biological processes. The metastasis-related changes within the tumor play a vital role in BCM, but so do metastasis-related changes within the TME. NcRNAs play an indispensable role in the development of BCM. LncRNA, circRNA, and miRNAs are the primary ncRNAs that function through various mechanisms, including ceRNA and binding with other proteins and RNA. We first summarized the key biological steps of BCM from two perspectives: tumor cells and the TME. NcRNAs promote “breast cancer seed” preferable or difficult for BCM through various means to regulate BC cell migration and invasion, EMT procession, and BCSCs malignant behavior. NcRNAs promote “breast cancer soil” preferred or difficult for BCM by various means to promote M2 macrophage polarization, CAF signaling pathway, and N2 conversion. Certain ncRNAs, such as HOTAIR, can be involved in BCM through multiple mechanisms, and numerous ncRNAs can target the same downstream genes. For example, Lnc TP73-AS1 and Linc-ZNF469-3 can target ZEB1 to regulate BCM, and lncRNA NEAT1 and LINC02163 can target HMGA2 to regulate BCM. LncRNA SNHG1 promotes BC cells and TME preferred for BCM. Lastly, we summarized the coding-genes involved in BCM from two perspectives: tumor cells and TME. We reviewed these ncRNA related clinical trials, however, there are currently no clinical trials available. We reviewed that the related clinical trials and the medicines targeted these coding genes which include: JAK2-STAT3, mucin 1, VEGFA, IGF-1, GSK3β, EGFR, ROS1 and PIK3CB. A large number of targeted drugs have been introduced into clinical research.
Based on our article, it provides a new direction for inhibiting BCM in the future. Firstly, our future research could focus on upregulating the NcRNAs related to “breast cancer seed” difficult for BCM and “breast cancer soil” difficult for BCM; or downregulating NcRNAs related to “breast cancer seed” preferable for BCM and “breast cancer soil” preferred for BCM. In recent years, we noted that more and more researchers are committed to studying the reprogramming of TME. The reprogramming of TME include various aspects: metabolic, immune regulation, neutrophils, macrophage, CAFs and so on. In one word, our review provides novel therapeutic targets for improving the prognosis of BCM and highlights new avenues for future BCM therapies. In addition, what caught our attention was that VEGFA and EGFR having a large number of clinical trials and targeted drugs. According our review, ncRNA RAB11B-AS1 and miR-29b-2/miR-338 targeted VEGFA; miR-146a/b targeted EGFR. NcRNA RAB11B-AS1, miR-29b-2/miR-338/146a/b have the potential to be research subjects for the next clinical trial. Clinically, ncRNA cannot be used as a therapeutic target for breast cancer metastasis at present for lacking clinical trials. The biggest challenge is: clinically, it is difficult to obtain breast cancer samples, and its expression content in blood is low. It requires a large number of blood samples to detect the expression of ncRNA. In summary, our article provides research direction for future treatment of BCM.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Abbreviation
3′-UTR, 3′-untranslated regions; ANGPTL4, Angiopoietin-like 4; BC, Breast Cancer; BCM, Breast Cancer Metastasis; BCBM, BC brain metastasis; BCSCS, BC stem cells; BRMS1, BCM suppressor 1; CAFs, cancer-associated fibroblasts; ceRNA, competitive endogenous RNA; circRNA, circular RNA; ECM, Extracellular Matrix; ER, Estrogen Receptor; DOCK4, Dedicator of cytokinesis protein 4; EMT, Epithelial–mesenchymal transition; EVs, Extracellular vesicles; FGFR1, Fibroblast growth factor receptor 1; GIT1, G-protein-coupled receptor kinase-interacting protein 1; GSK3β, Glycogen synthase kinase 3β; HER2, Human Epidermal Growth Factor Receptor 2; IBSP, Integrin-Binding Sialoprotein; LBR, Lamin B receptor; LncRNA, Long noncoding RNA; LIFR, Leukemia Inhibitory Factor Receptor; MAML1, Mastermind-like transcriptional coactivator 1; miRNA, microRNA; MVs, Microvesicles; MMPs, matrix metalloproteinases; mRNAs, Message RNAs; MTA1, Metastasis Associated 1; MTMR3, Myotubularin-Related Protein 3; MYLIP, Myosin Regulatory Light Chain Interacting Protein; NcRNAs, Noncoding RNAs; NFs, Normal fibroblasts; NF-κB, Nuclear Factor Kappa B; nt, nucleotides; piRNA, PIWI interacting RNA; PR, Progesterone Receptor; PRC2, Polycomb Repressive Complex 2; SDCBP, Syndecan binding protein; SOCS2, Suppressors of Cytokine Signaling 2; TAM, Tumor-associated macrophages; TCEAL7, Transcription Elongation Factor A Like 7; TFAM, Mitochondrial Transcription Factor A; TIMP2, Tissue Inhibitors of Metalloproteinase 2; TGF-β1, Transforming Growth Factor β1; TGFBRI, TGF-β the type I Receptor; TME, Tumor Microenvironment; TNBC, Triple-Negative Breast Cancer; TNF, Tumor Necrosis Factor; USP28, Ubiquitin-specific peptidase 28; VEGFA, Vascular Endothelial Growth Factor A; WASF3, Wiskott-Aldrich syndrome protein family member 3; YAP1, Yes-associated protein 1.
Data Sharing Statement
No datasets were generated or analysed during the current study.
Acknowledgments
Acknowledgement all the authors.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Beijing Vlove Charity Foundation (ky1322); Yantai Yuhuangding Hospital Youth Launch Fund (kj0521) and Yantai Yuhuangding Hospital Doctoral Research Launch Fund (kj0570).
Disclosure
The authors declare no competing interests.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71:209–249. doi:10.3322/caac.21660
2. Desantis CE, Ma J, Gaudet MM, et al. Breast cancer statistics, 2019. CA Cancer J Clin. 2019;69:438–451. doi:10.3322/caac.21583
3. Sung H, Ren J, Li J, et al. Breast cancer risk factors and mammographic density among high-risk women in urban China. NPJ Breast Cancer. 2018;4(1):3. doi:10.1038/s41523-018-0055-9
4. Collaborative Group on Hormonal Factors in Breast Cancer. Menarche, menopause, and breast cancer risk: individual participant meta-analysis, including 118 964 women with breast cancer from 117 epidemiological studies. Lancet Oncol. 2012;13(11):1141–1151. doi:10.1016/S1470-2045(12)70425-4
5. Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331(6024):1559–1564. doi:10.1126/science.1203543
6. Harbeck N, Penault-Llorca F, Cortes J, et al. Breast cancer. Nat Rev Dis Primers. 2019;5(1):66. doi:10.1038/s41572-019-0111-2
7. Derakhshan F, Reis-Filho JS. Pathogenesis of triple-negative breast cancer. Annu Rev Pathol. 2022;17(1):181–204. doi:10.1146/annurev-pathol-042420-093238
8. Brenton JD, Carey LA, Ahmed AA, Caldas C. Molecular classification and molecular forecasting of breast cancer: ready for clinical application? J Clin Oncol. 2005;23(29):7350–7360. doi:10.1200/JCO.2005.03.3845
9. Foulkes WD, Smith IE, Reis-Filho JS. Triple-negative breast cancer. N Engl J Med. 2010;363(20):1938–1948. doi:10.1056/NEJMra1001389
10. Muller A, Homey B, Soto H, et al. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001;410(6824):50–56. doi:10.1038/35065016
11. Ernst MF, Van De Poll-Franse LV, Roukema JA, et al. Trends in the prognosis of patients with primary metastatic breast cancer diagnosed between 1975 and 2002. Breast. 2007;16(4):344–351. doi:10.1016/j.breast.2007.01.001
12. Ren Z, Li Y, Hameed O, Siegal GP, Wei S. Prognostic factors in patients with metastatic breast cancer at the time of diagnosis. Pathol Res Pract. 2014;210(5):301–306. doi:10.1016/j.prp.2014.01.008
13. Brufsky AM, Mayer M, Rugo HS, et al. Central nervous system metastases in patients with HER2-positive metastatic breast cancer: incidence, treatment, and survival in patients from registHER. Clin Cancer Res. 2011;17(14):4834–4843. doi:10.1158/1078-0432.CCR-10-2962
14. Al-Mehdi AB, Tozawa K, Fisher AB, Shientag L, Lee A, Muschel RJ. Intravascular origin of metastasis from the proliferation of endothelium-attached tumor cells: a new model for metastasis. Nat Med. 2000;6(1):100–102. doi:10.1038/71429
15. Hunter KW, Crawford NP, Alsarraj J. Mechanisms of metastasis. Breast Cancer Res. 2008;Suppl 10(S1):S2. doi:10.1186/bcr1988
16. Ren B, Cui M, Yang G, et al. Tumor microenvironment participates in metastasis of pancreatic cancer. Mol Cancer. 2018;17(1):108. doi:10.1186/s12943-018-0858-1
17. Lambert AW, Pattabiraman DR, Weinberg RA. Emerging biological principles of metastasis. Cell. 2017;168(4):670–691. doi:10.1016/j.cell.2016.11.037
18. Savagner P. The epithelial–mesenchymal transition (EMT) phenomenon. Ann Oncol. 2010;Suppl 21:vii89–vii92. doi:10.1093/annonc/mdq292
19. Smit MA, Peeper DS. Epithelial-mesenchymal transition and senescence: two cancer-related processes are crossing paths. Aging. 2010;2(10):735–741. doi:10.18632/aging.100209
20. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414(6859):105–111. doi:10.1038/35102167
21. Lanzardo S, Conti L, Rooke R, et al. Immunotargeting of antigen xCT attenuates stem-like cell behavior and metastatic progression in breast cancer. Cancer Res. 2016;76:62–72. doi:10.1158/0008-5472.CAN-15-1208
22. Lawson DA, Bhakta NR, Kessenbrock K, et al. Single-cell analysis reveals a stem-cell program in human metastatic breast cancer cells. Nature. 2015;526(7571):131–135. doi:10.1038/nature15260
23. Junttila MR, De Sauvage FJ. Influence of tumour micro-environment heterogeneity on therapeutic response. Nature. 2013;501(7467):346–354. doi:10.1038/nature12626
24. Entschladen F, Palm D, Drell TLT, Lang K, Zaenker KS. Connecting a tumor to the environment. Curr Pharm Des. 2007;13(33):3440–3444. doi:10.2174/138161207782360573
25. Kessenbrock K, Plaks V, Werb Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell. 2010;141(1):52–67. doi:10.1016/j.cell.2010.03.015
26. Kimura T. [Non-coding Natural Antisense RNA: mechanisms of Action in the Regulation of Target Gene Expression and Its Clinical Implications]. Yakugaku Zasshi. 2020;140(5):687–700. Danish. doi:10.1248/yakushi.20-00002
27. Furuno M, Pang KC, Ninomiya N, et al. Clusters of internally primed transcripts reveal novel long noncoding RNAs. PLoS Genet. 2006;2:e37. doi:10.1371/journal.pgen.0020037
28. Yan H, Bu P, Hon CC. Non-coding RNA in cancer. Essays Biochem. 2021;65(4):625–639. doi:10.1042/EBC20200032
29. Rinn JL, Chang HY. Genome regulation by long noncoding RNAs. Annu Rev Biochem. 2012;81(1):145–166. doi:10.1146/annurev-biochem-051410-092902
30. Cabili MN, Trapnell C, Goff L, et al. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011;25(18):1915–1927. doi:10.1101/gad.17446611
31. Guttman M, Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature. 2012;482(7385):339–346. doi:10.1038/nature10887
32. Salmena L, Poliseno L, Tay Y, et al. A ceRNA hypothesis: the rosetta stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
33. Tay Y, Rinn J, Pandolfi PP. The multilayered complexity of ceRNA crosstalk and competition. Nature. 2014;505(7483):344–352. doi:10.1038/nature12986
34. Bi P, Ramirez-Martinez A, Li H. Control of muscle formation by the fusogenic micropeptide myomixer. Science. 2017;356(6335):323–327. doi:10.1126/science.aam9361
35. D’Lima NG, Ma J, Winkler L. A human microprotein that interacts with the mRNA decapping complex. Nat Chem Biol. 2017;13(2):174–180. doi:10.1038/nchembio.2249
36. Huang J-Z, Chen M, Chen D. A peptide encoded by a putative lncRNA HOXB-AS3 suppresses colon cancer growth. Mol Cell. 2017;68(1):171–184e6. doi:10.1016/j.molcel.2017.09.015
37. Zhang Q, Vashisht AA, O’rourke J, et al. The microprotein minion controls cell fusion and muscle formation. Nat Commun. 2017;8(1):15664. doi:10.1038/ncomms15664
38. Nielsen AF, Bindereif A, Bozzoni I, et al. Best practice standards for circular RNA research. Nat Methods. 2022;19(10):1208–1220. doi:10.1038/s41592-022-01487-2
39. Liu J, Liu T, Wang X, He A. Circles reshaping the RNA world: from waste to treasure. Mol Cancer. 2017;16(1):58. doi:10.1186/s12943-017-0630-y
40. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
41. Mcguire A, Brown JAL, Kerin MJ. Metastatic breast cancer: the potential of miRNA for diagnosis and treatment monitoring. Cancer Metastasis Rev. 2015;34(1):145–155. doi:10.1007/s10555-015-9551-7
42. Siomi MC, Sato K, Pezic D, Aravin AA. PIWI-interacting small RNAs: the vanguard of genome defence. Nat Rev Mol Cell Biol. 2011;12(4):246–258. doi:10.1038/nrm3089
43. Vagin VV, Sigova A, Li C, Seitz H, Gvozdev V, Zamore PD. A distinct small RNA pathway silences selfish genetic elements in the germline. Science. 2006;313(5785):320–324. doi:10.1126/science.1129333
44. Talmadge JE, Fidler IJ. AACR centennial series: the biology of cancer metastasis: historical perspective. Cancer Res. 2010;70(14):5649–5669. doi:10.1158/0008-5472.CAN-10-1040
45. Paget S. The distribution of secondary growths in cancer of the breast. 1889. Cancer Metastasis Rev. 1989;8(2):98–101.
46. Lorusso G, Ruegg C. The tumor microenvironment and its contribution to tumor evolution toward metastasis. Histochem Cell Biol. 2008;130(6):1091–1103. doi:10.1007/s00418-008-0530-8
47. Bissell MJ, Radisky DC, Rizki A, Weaver VM, Petersen OW. The organizing principle: microenvironmental influences in the normal and malignant breast. Differentiation. 2002;70(9–10):537–546. doi:10.1046/j.1432-0436.2002.700907.x
48. Soysal SD, Tzankov A, Muenst SE. Role of the tumor microenvironment in breast cancer. Pathobiology. 2015;82(3–4):142–152. doi:10.1159/000430499
49. Pietras K, Ostman A. Hallmarks of cancer: interactions with the tumor stroma. Exp Cell Res. 2010;316(8):1324–1331. doi:10.1016/j.yexcr.2010.02.045
50. Arendt LM, Rudnick JA, Keller PJ, Kuperwasser C. Stroma in breast development and disease. Semin Cell Dev Biol. 2010;21(1):11–18. doi:10.1016/j.semcdb.2009.10.003
51. Yan X, Xie Y, Yang F, et al. Comprehensive description of the current breast cancer microenvironment advancements via single-cell analysis. J Exp Clin Cancer Res. 2021;40(1):142. doi:10.1186/s13046-021-01949-z
52. Kaushik N, Kim S, Suh Y, Lee S-J. Proinvasive extracellular matrix remodeling for tumor progression. Arch Pharm Res. 2019;42(1):40–47. doi:10.1007/s12272-018-1097-0
53. Gaggioli C, Hooper S, Hidalgo-Carcedo C. Fibroblast-led collective invasion of carcinoma cells with differing roles for RhoGTPases in leading and following cells. Nat Cell Biol. 2007;9(12):1392–1400. doi:10.1038/ncb1658
54. Hu D, Li Z, Zheng B, et al. Cancer-associated fibroblasts in breast cancer: challenges and opportunities. Cancer Commun. 2022;42(5):401–434. doi:10.1002/cac2.12291
55. Egeblad M, Werb Z. New functions for the matrix metalloproteinases in cancer progression. Nat Rev Cancer. 2002;2(3):161–174. doi:10.1038/nrc745
56. Dano K, Behrendt N, Hoyer-Hansen G, et al. Plasminogen activation and cancer. Thromb Haemost. 2005;93(04):676–681. doi:10.1160/TH05-01-0054
57. Wen S, Hou Y, Fu L, et al. Cancer-associated fibroblast (CAF)-derived IL32 promotes breast cancer cell invasion and metastasis via integrin β3–p38 MAPK signalling. Cancer Lett. 2019;442:320–332. doi:10.1016/j.canlet.2018.10.015
58. Xi L, Peng M, Liu S, et al. Hypoxia-stimulated ATM activation regulates autophagy-associated exosome release from cancer-associated fibroblasts to promote cancer cell invasion. J Extracell Vesicles. 2021;10(11):e12146. doi:10.1002/jev2.12146
59. Liu T, Han C, Wang S, et al. Cancer-associated fibroblasts: an emerging target of anti-cancer immunotherapy. J Hematol Oncol. 2019;12(1):86. doi:10.1186/s13045-019-0770-1
60. Gunaydin G. CAFs interacting with TAMs in tumor microenvironment to enhance tumorigenesis and immune evasion. Front Oncol. 2021;11:668349. doi:10.3389/fonc.2021.668349
61. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8(12):958–969. doi:10.1038/nri2448
62. Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002;23(11):549–555. doi:10.1016/S1471-4906(02)02302-5
63. Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11(10):889–896. doi:10.1038/ni.1937
64. Martinez FO, Gordon S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Reports. 2014;6:13. doi:10.12703/P6-13
65. Pakravan K, Mossahebi-Mohammadi M, Ghazimoradi MH, et al. Monocytes educated by cancer-associated fibroblasts secrete exosomal miR-181a to activate AKT signaling in breast cancer cells. J Transl Med. 2022;20(1):559. doi:10.1186/s12967-022-03780-2
66. Boutilier AJ, Elsawa SF. Macrophage polarization states in the tumor microenvironment. Int J Mol Sci. 2021;22(13):22. doi:10.3390/ijms22136995
67. Zhang W, Shen Y, Huang H. A rosetta stone for breast cancer: prognostic value and dynamic regulation of neutrophil in tumor microenvironment. Front Immunol. 2020;11:1779. doi:10.3389/fimmu.2020.01779
68. Huang T, Song C, Zheng L, et al. The roles of extracellular vesicles in gastric cancer development, microenvironment, anti-cancer drug resistance, and therapy. Mol Cancer. 2019;18(1):62. doi:10.1186/s12943-019-0967-5
69. Xiu B, Chi Y, Liu L, et al. LINC02273 drives breast cancer metastasis by epigenetically increasing AGR2 transcription. Mol Cancer. 2019;18(1):187. doi:10.1186/s12943-019-1115-y
70. Jiang Z, Slater CM, Zhou Y. LincIN, a novel NF90-binding long non-coding RNA, is overexpressed in advanced breast tumors and involved in metastasis. Breast Cancer Res. 2017;19(1):62. doi:10.1186/s13058-017-0853-2
71. Kong X, Duan Y, Sang Y. LncRNA–CDC6 promotes breast cancer progression and function as ceRNA to target CDC6 by sponging microRNA-215. J Cell Physiol. 2019;234(6):9105–9117. doi:10.1002/jcp.27587
72. Li L, Jin Y, Wang X, et al. Long noncoding RNA ENST00000508435 promotes migration of breast cancer via FXR 1. Cell Adh Migr. 2021;15(1):140–151. doi:10.1080/19336918.2021.1921402
73. Zou Q, Zhou E, Xu F, Zhang D, Yi W, Yao J. A TP73-AS1/miR-200a/ZEB1 regulating loop promotes breast cancer cell invasion and migration. J Cell Biochem. 2018;119(2):2189–2199. doi:10.1002/jcb.26380
74. Ma Q, Yang L, Tolentino K, et al. Inducible lncRNA transgenic mice reveal continual role of HOTAIR in promoting breast cancer metastasis. Elife. 2022;11:11. doi:10.7554/eLife.79126
75. Gupta RA, Shah N, Wang KC, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076. doi:10.1038/nature08975
76. Hou P, Zhao Y, Li Z. Correction: lincRNA-ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2025;16(1):340. doi:10.1038/s41419-025-07683-0
77. Meng L, Yue X, Zhou D, Li H. Long non coding RNA OIP5‑AS1 promotes metastasis of breast cancer via miR‑340‑5p/ZEB2 axis. Oncol Rep. 2020;44(4):1662–1670. doi:10.3892/or.2020.7724
78. Li R-H, Chen M, Liu J. Long noncoding RNA ATB promotes the epithelial−mesenchymal transition by upregulating the miR-200c/Twist1 axe and predicts poor prognosis in breast cancer. Cell Death Dis. 2018;9(12):1171. doi:10.1038/s41419-018-1210-9
79. Zhu L, Jiang S, Yu S. Increased SIX-1 expression promotes breast cancer metastasis by regulating lncATB-miR-200s-ZEB1 axis. J Cell Mol Med. 2020;24(9):5290–5303. doi:10.1111/jcmm.15185
80. Liu G, Hu X, Zhou G. Long non-coding RNA OR3A4 promotes proliferation and migration in breast cancer. Biomed Pharmacother. 2017;96:426–433. doi:10.1016/j.biopha.2017.10.011
81. Qiao Y, Jin T, Guan S, et al. Long non-coding RNA Lnc-408 promotes invasion and metastasis of breast cancer cell by regulating LIMK1. Oncogene. 2021;40(24):4198–4213. doi:10.1038/s41388-021-01845-y
82. Li X, Wang S, Li Z. The lncRNA NEAT1 facilitates cell growth and invasion via the miR-211/HMGA2 axis in breast cancer. Int J Biol Macromol. 2017;105:346–353. doi:10.1016/j.ijbiomac.2017.07.053
83. Liu S, Chen W, Hu H. Correction: long noncoding RNA PVT1 promotes breast cancer proliferation and metastasis by binding miR-128-3p and UPF1. Breast Cancer Res. 2023;25(1):98. doi:10.1186/s13058-023-01698-1
84. Kong X, Li J, Li Y, et al. A novel long non-coding RNA AC073352.1 promotes metastasis and angiogenesis via interacting with YBX1 in breast cancer. Cell Death Dis. 2021;12(7):670. doi:10.1038/s41419-021-03943-x
85. Wang P-S, Chou C-H, Lin C-H, et al. A novel long non-coding RNA linc-ZNF469-3 promotes lung metastasis through miR-574-5p-ZEB1 axis in triple negative breast cancer. Oncogene. 2018;37(34):4662–4678. doi:10.1038/s41388-018-0293-1
86. Li X-Y, Zhou L-Y, Luo H. The long noncoding RNA MIR210HG promotes tumor metastasis by acting as a ceRNA of miR-1226-3p to regulate mucin-1c expression in invasive breast cancer. Aging. 2019;11(15):5646–5665. doi:10.18632/aging.102149
87. Niu Y, Bao L, Chen Y, et al. HIF2-induced long noncoding RNA RAB11B-AS1 promotes hypoxia-mediated angiogenesis and breast cancer metastasis. Cancer Res. 2020;80(5):964–975. doi:10.1158/0008-5472.CAN-19-1532
88. Zuo Y, Qu C, Tian Y, et al. The HIF −1/ SNHG1 / miR −199a-3p/ TFAM axis explains tumor angiogenesis and metastasis under hypoxic conditions in breast cancer. Biofactors. 2021;47(3):444–460. doi:10.1002/biof.1702
89. Zhang Q, Li T, Wang Z, Kuang X, Shao N, Lin Y. lncRNA NR2F1-AS1 promotes breast cancer angiogenesis through activating IGF-1/IGF-1R/ERK pathway. J Cell Mol Med. 2020;24(14):8236–8247. doi:10.1111/jcmm.15499
90. Shi G, Cheng Y, Zhang Y, et al. Long non-coding RNA LINC00511/miR-150/MMP13 axis promotes breast cancer proliferation, migration and invasion. Biochim Biophys Acta Mol Basis Dis. 2021;1867:165957.
91. Nyati KK, Hashimoto S, Singh SK, et al. The novel long noncoding RNA AU021063, induced by IL-6/Arid5a signaling, exacerbates breast cancer invasion and metastasis by stabilizing Trib3 and activating the Mek/Erk pathway. Cancer Lett. 2021;520:295–306. doi:10.1016/j.canlet.2021.08.004
92. Qin C, Jin L, Li J, et al. Correction: long non-coding RNA LINC02163 accelerates malignant tumor behaviors in breast cancer by regulating the microRNA-511-3p/HMGA2 axis as a competing endogenous RNA. Oncol Res. 2024;32(9):1517–1522. doi:10.32604/or.2024.051893
93. Jin X, Xu X-E, Jiang Y-Z, et al. The endogenous retrovirus-derived long noncoding RNA TROJAN promotes triple-negative breast cancer progression via ZMYND8 degradation. Sci Adv. 2019;5(3):eaat9820. doi:10.1126/sciadv.aat9820
94. Wang H-B, Wei H, Wang J-S, et al. Down-regulated expression of LINC00518 prevents epithelial cell growth and metastasis in breast cancer through the inhibition of CDX2 methylation and the Wnt signaling pathway. Biochim Biophys Acta Mol Basis Dis. 2019;1865(3):708–723. doi:10.1016/j.bbadis.2019.01.003
95. Zheng A, Song X, Zhang L. Long non-coding RNA LUCAT1/miR-5582-3p/TCF7L2 axis regulates breast cancer stemness via Wnt/β-catenin pathway. J Exp Clin Cancer Res. 2019;38(1):305. doi:10.1186/s13046-019-1315-8
96. Zhang W, Liu H, Jiang J, et al. CircRNA circFOXK2 facilitates oncogenesis in breast cancer via IGF2BP3/miR-370 axis. Aging. 2021;13(14):18978–18992. doi:10.18632/aging.203347
97. Feng H, Sun S-Z, Cheng F, Zhang N-Q. Mediation of circ_RPPH1 on miR-146b-3p/E2F2 pathway to hinder the growth and metastasis of breast carcinoma cells. Aging. 2021;13(16):20552–20568. doi:10.18632/aging.203439
98. Zeng K, He B, Yang B, et al. The pro-metastasis effect of circANKS1B in breast cancer. Mol Cancer. 2018;17(1):160. doi:10.1186/s12943-018-0914-x
99. Li Y, Wang Z, Su P, et al. circ-EIF6 encodes EIF6-224aa to promote TNBC progression via stabilizing MYH9 and activating the Wnt/beta-catenin pathway. Mol Ther. 2022;30(1):415–430. doi:10.1016/j.ymthe.2021.08.026
100. Yu S-J, Hu J-Y, Kuang X-Y. MicroRNA-200a promotes anoikis resistance and metastasis by targeting YAP1 in human breast cancer. Clin Cancer Res. 2013;19(6):1389–1399. doi:10.1158/1078-0432.CCR-12-1959
101. Mesci A, Huang X, Taeb S. Targeting of CCBE1 by miR-330-3p in human breast cancer promotes metastasis. Br J Cancer. 2017;116(10):1350–1357. doi:10.1038/bjc.2017.105
102. Zhang X, Wu M, Chong Q-Y. Amplification of hsa-miR-191/425 locus promotes breast cancer proliferation and metastasis by targeting DICER1. Carcinogenesis. 2018;39(12):1506–1516. doi:10.1093/carcin/bgy102
103. Tian X, Yu H, Li D. The miR-5694/AF9/Snail axis provides metastatic advantages and a therapeutic target in basal-like breast cancer. Mol Ther. 2021;29(3):1239–1257. doi:10.1016/j.ymthe.2020.11.022
104. Lei B, Wang D, Zhang M, Deng Y, Jiang H, Li Y. miR-615-3p promotes the epithelial-mesenchymal transition and metastasis of breast cancer by targeting PICK1/TGFBRI axis. J Exp Clin Cancer Res. 2020;39(1):71. doi:10.1186/s13046-020-01571-5
105. Yu J, Lei R, Zhuang X. MicroRNA-182 targets SMAD7 to potentiate TGFbeta-induced epithelial-mesenchymal transition and metastasis of cancer cells. Nat Commun. 2016;7(1):13884. doi:10.1038/ncomms13884
106. Lu J, Zhao Q, Guo Y, et al. Regulation of ERalpha-dependent breast cancer metastasis by a miR-29a signaling. J Exp Clin Cancer Res. 2023;42:93. doi:10.1186/s13046-023-02665-6
107. Davenport ML, Davis MR, Davenport BN. Suppression of SIN3A by miR-183 promotes breast cancer metastasis. Mol Cancer Res. 2022;20(6):883–894. doi:10.1158/1541-7786.MCR-21-0508
108. Ma L, Teruya-Feldstein J, Weinberg RA. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature. 2007;449(7163):682–688. doi:10.1038/nature06174
109. Wang J, Zhang Q, Wang D. Microenvironment-induced TIMP2 loss by cancer-secreted exosomal miR-4443 promotes liver metastasis of breast cancer. J Cell Physiol. 2020;235(7–8):5722–5735. doi:10.1002/jcp.29507
110. Shen S, Song Y, Zhao B. Cancer-derived exosomal miR-7641 promotes breast cancer progression and metastasis. Cell Commun Signal. 2021;19(1):20. doi:10.1186/s12964-020-00700-z
111. Liu B, Sun L, Liu Q, et al. A cytoplasmic NF-kappaB interacting long noncoding RNA blocks IkappaB phosphorylation and suppresses breast cancer metastasis. Cancer Cell. 2015;27:370–381. doi:10.1016/j.ccell.2015.02.004
112. Daly EB, Wind T, Jiang XM, Sun L, Hogg PJ. Secretion of phosphoglycerate kinase from tumour cells is controlled by oxygen-sensing hydroxylases. Biochim Biophys Acta. 2004;169117–22. doi:10.1016/j.bbamcr.2003.11.004
113. Chu Z, Huo N, Zhu X. FOXO3A-induced LINC00926 suppresses breast tumor growth and metastasis through inhibition of PGK1-mediated Warburg effect. Mol Ther. 2021;29:2737–2753. doi:10.1016/j.ymthe.2021.04.036
114. Kim J, Piao HL, Kim BJ, et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat Genet. 2018;50:1705–1715. doi:10.1038/s41588-018-0252-3
115. Wang M, Wang M, Wang Z. Long non-coding RNA - CTD −2108O9.1 represses breast cancer metastasis by influencing leukemia inhibitory factor receptor. Cancer Sci. 2018;109(6):1764–1774. doi:10.1111/cas.13592
116. Zhao L, Zhou Y, Zhao Y, Li Q, Zhou J, Mao Y. Long non-coding RNA TUSC8 inhibits breast cancer growth and metastasis via miR-190b-5p/MYLIP axis. Aging. 2020;12:2974–2991. doi:10.18632/aging.102791
117. Zhang W, Shi S, Jiang J, et al. LncRNA MEG3 inhibits cell epithelial-mesenchymal transition by sponging miR-421 targeting E-cadherin in breast cancer. Biomed Pharmacother. 2017;91:312–319. doi:10.1016/j.biopha.2017.04.085
118. Wang -Y-Y, Yan L, Yang S. Long noncoding RNA AC073284.4 suppresses epithelial–mesenchymal transition by sponging miR-18b-5p in paclitaxel-resistant breast cancer cells. J Cell Physiol. 2019;234(12):23202–23215. doi:10.1002/jcp.28887
119. Jiang Y, Lin L, Zhong S. Overexpression of novel lncRNA NLIPMT inhibits metastasis by reducing phosphorylated glycogen synthase kinase 3β in breast cancer. J Cell Physiol. 2019;234(7):10698–10708. doi:10.1002/jcp.27738
120. Wang F, Wang X, Li J. Correction to: circNOL10 suppresses breast cancer progression by sponging miR-767-5p to regulate SOCS2/JAK/STAT signaling. J Biomed Sci. 2021;28(1):31. doi:10.1186/s12929-021-00723-9
121. Zeng F, Yao M, Wang Y, et al. Fatty acid beta-oxidation promotes breast cancer stemness and metastasis via the miRNA-328-3p-CPT1A pathway. Cancer Gene Ther. 2022;29:383–395. doi:10.1038/s41417-021-00348-y
122. Goldberger N, Walker RC, Kim CH, Winter S, Hunter KW. Inherited variation in miR-290 expression suppresses breast cancer progression by targeting the metastasis susceptibility gene Arid4b. Cancer Res. 2013;73(8):2671–2681. doi:10.1158/0008-5472.CAN-12-3513
123. Song Y, Zeng S, Zheng G, et al. FOXO3a-driven miRNA signatures suppresses VEGF-A/NRP1 signaling and breast cancer metastasis. Oncogene. 2021;40(4):777–790. doi:10.1038/s41388-020-01562-y
124. Phadke PA, Vaidya KS, Nash KT, et al. BRMS1 suppresses breast cancer experimental metastasis to multiple organs by inhibiting several steps of the metastatic process. Am J Pathol. 2008;172(3):809–817. doi:10.2353/ajpath.2008.070772
125. Hurst DR, Xie Y, Vaidya KS. Alterations of BRMS1-ARID4A interaction modify gene expression but still suppress metastasis in human breast cancer cells. J Biol Chem. 2008;283(12):7438–7444. doi:10.1074/jbc.M709446200
126. Hurst DR, Edmonds MD, Scott GK, et al. Breast cancer metastasis suppressor 1 up-regulates miR-146, which suppresses breast cancer metastasis. Cancer Res. 2009;69(4):1279–1283. doi:10.1158/0008-5472.CAN-08-3559
127. Ma Z, Li Y, Xu J, Ren Q, Yao J, Tian X. MicroRNA-409-3p regulates cell invasion and metastasis by targeting ZEB1 in breast cancer. IUBMB Life. 2016;68(5):394–402. doi:10.1002/iub.1494
128. Pan Y, Jiao G, Wang C, Yang J, Yang W. MicroRNA-421 inhibits breast cancer metastasis by targeting metastasis associated 1. Biomed Pharmacother. 2016;83:1398–1406. doi:10.1016/j.biopha.2016.08.058
129. Wang Q-Y, Zhou C-X, Zhan M-N. MiR-133b targets Sox9 to control pathogenesis and metastasis of breast cancer. Cell Death Dis. 2018;9(7):752. doi:10.1038/s41419-018-0715-6
130. Zhang C, Zhang Y, Ding W, Lin Y, Huang Z, Luo Q. MiR-33a suppresses breast cancer cell proliferation and metastasis by targeting ADAM9 and ROS1. Protein Cell. 2015;6(12):881–889. doi:10.1007/s13238-015-0223-8
131. Ma F, Zhang L, Ma L, Zhang Y, Zhang J, Guo B. MiR-361-5p inhibits glycolytic metabolism, proliferation and invasion of breast cancer by targeting FGFR1 and MMP-1. J Exp Clin Cancer Res. 2017;36(1):158. doi:10.1186/s13046-017-0630-1
132. Pardo OE, Castellano L, Munro CE. miR-515-5p controls cancer cell migration through MARK 4 regulation. EMBO Rep. 2016;17(4):570–584. doi:10.15252/embr.201540970
133. Yu Y, Luo W, Yang Z-J. miR-190 suppresses breast cancer metastasis by regulation of TGF-β-induced epithelial–mesenchymal transition. Mol Cancer. 2018;17(1):70. doi:10.1186/s12943-018-0818-9
134. Bai W-D, Ye X-M, Zhang M-Y. MiR-200c suppresses TGF-β signaling and counteracts trastuzumab resistance and metastasis by targeting ZNF217 and ZEB1 in breast cancer. Int J Cancer. 2014;135(6):1356–1368. doi:10.1002/ijc.28782
135. Pu T, Shen M, Li S, et al. Repression of miR-135b-5p promotes metastasis of early-stage breast cancer by regulating downstream target SDCBP. Lab Invest. 2019;99(9):1296–1308. doi:10.1038/s41374-019-0258-1
136. Han M-L, Wang F, Gu Y-T. MicroR-760 suppresses cancer stem cell subpopulation and breast cancer cell proliferation and metastasis: by down-regulating NANOG. Biomed Pharmacother. 2016;80:304–310. doi:10.1016/j.biopha.2016.03.024
137. Shibuya N, Kakeji Y, Shimono Y. MicroRNA-93 targets WASF3 and functions as a metastasis suppressor in breast cancer. Cancer Sci. 2020;111(6):2093–2103. doi:10.1111/cas.14423
138. Chan S-H, Huang W-C, Chang J-W. MicroRNA-149 targets GIT1 to suppress integrin signaling and breast cancer metastasis. Oncogene. 2014;33(36):4496–4507. doi:10.1038/onc.2014.10
139. Zhang H, Cai K, Wang J. MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem Cells. 2014;32(11):2858–2868. doi:10.1002/stem.1795
140. Chu C, Liu X, Bai X. MiR-519d suppresses breast cancer tumorigenesis and metastasis via targeting MMP3. Int J Biol Sci. 2018;14(2):228–236. doi:10.7150/ijbs.22849
141. Zhao S, Han J, Zheng L, Yang Z, Zhao L, Lv Y. MicroRNA-203 regulates growth and metastasis of breast cancer. Cell Physiol Biochem. 2015;37(1):35–42. doi:10.1159/000430331
142. Liu R, Li J, Lai Y, et al. Hsa-miR-1 suppresses breast cancer development by down-regulating K-ras and long non-coding RNA MALAT1. Int J Biol Macromol. 2015;81:491–497. doi:10.1016/j.ijbiomac.2015.08.016
143. Li L, Kang L, Zhao W, et al. miR-30a-5p suppresses breast tumor growth and metastasis through inhibition of LDHA-mediated Warburg effect. Cancer Lett. 2017;400:89–98. doi:10.1016/j.canlet.2017.04.034
144. Shi W, Tang T, Li X, et al. Methylation-mediated silencing of miR-133a-3p promotes breast cancer cell migration and stemness via miR-133a-3p/MAML1/DNMT3A positive feedback loop. J Exp Clin Cancer Res. 2019;38(1):429. doi:10.1186/s13046-019-1400-z
145. Hong BS, Ryu HS, Kim N. Tumor suppressor miRNA-204-5p regulates growth, metastasis, and immune microenvironment remodeling in breast cancer. Cancer Res. 2019;79(7):1520–1534. doi:10.1158/0008-5472.CAN-18-0891
146. Zong S, Dai W, Guo X, Wang K. LncRNA-SNHG1 promotes macrophage M2-like polarization and contributes to breast cancer growth and metastasis. Aging. 2021;13(19):23169–23181. doi:10.18632/aging.203609
147. Zeng H, Hou Y, Zhou X, et al. Cancer-associated fibroblasts facilitate premetastatic niche formation through lncRNA SNHG5-mediated angiogenesis and vascular permeability in breast cancer. Theranostics. 2022;12(17):7351–7370. doi:10.7150/thno.74753
148. Tao S, Chen Q, Lin C, Dong H. Linc00514 promotes breast cancer metastasis and M2 polarization of tumor-associated macrophages via Jagged1-mediated notch signaling pathway. J Exp Clin Cancer Res. 2020;39(1):191. doi:10.1186/s13046-020-01676-x
149. Akimoto M, Susa T, Okudaira N. A novel LncRNA PTH-AS upregulates interferon-related DNA damage resistance signature genes and promotes metastasis in human breast cancer xenografts. J Biol Chem. 2022;298(7):102065. doi:10.1016/j.jbc.2022.102065
150. Zhou L, Tian Y, Guo F. LincRNA-p21 knockdown reversed tumor-associated macrophages function by promoting MDM2 to antagonize* p53 activation and alleviate breast cancer development. Cancer Immunol Immunother. 2020;69(5):835–846. doi:10.1007/s00262-020-02511-0
151. Qi M, Xia Y, Wu Y, et al. Lin28B-high breast cancer cells promote immune suppression in the lung pre-metastatic niche via exosomes and support cancer progression. Nat Commun. 2022;13(1):897. doi:10.1038/s41467-022-28438-x
152. Zhang C, Wei S, Dai S, et al. The NR_109/FUBP1/c-Myc axis regulates TAM polarization and remodels the tumor microenvironment to promote cancer development. J Immunother Cancer. 2023;11:11.
153. Lu C, Shi W, Hu W. Endoplasmic reticulum stress promotes breast cancer cells to release exosomes circ_0001142 and induces M2 polarization of macrophages to regulate tumor progression. Pharmacol Res. 2022;177:106098. doi:10.1016/j.phrs.2022.106098
154. Ye F, Liang Y, Wang Y, et al. Cancer-associated fibroblasts facilitate breast cancer progression through exosomal circTBPL1-mediated intercellular communication. Cell Death Dis. 2023;14(7):471. doi:10.1038/s41419-023-05986-8
155. Zhou B, Mo Z, Lai G. Targeting tumor exosomal circular RNA cSERPINE2 suppresses breast cancer progression by modulating MALT1-NF-κB-IL-6 axis of tumor-associated macrophages. J Exp Clin Cancer Res. 2023;42(1):48. doi:10.1186/s13046-023-02620-5
156. Zheng Y, Ren S, Zhang Y. Circular RNA circWWC3 augments breast cancer progression through promoting M2 macrophage polarization and tumor immune escape via regulating the expression and secretion of IL-4. Cancer Cell Int. 2022;22(1):264. doi:10.1186/s12935-022-02686-9
157. Chen B, Sang Y, Song X. Exosomal miR-500a-5p derived from cancer-associated fibroblasts promotes breast cancer cell proliferation and metastasis through targeting USP28. Theranostics. 2021;11(8):3932–3947. doi:10.7150/thno.53412
158. Yan Z, Sheng Z, Zheng Y, et al. Cancer-associated fibroblast-derived exosomal miR-18b promotes breast cancer invasion and metastasis by regulating TCEAL7. Cell Death Dis. 2021;12(12):1120. doi:10.1038/s41419-021-04409-w
159. Chatterjee A, Jana S, Chatterjee S. MicroRNA-222 reprogrammed cancer-associated fibroblasts enhance growth and metastasis of breast cancer. Br J Cancer. 2019;121(8):679–689. doi:10.1038/s41416-019-0566-7
160. Baroni S, Romero-Cordoba S, Plantamura I, et al. Correction: exosome-mediated delivery of miR-9 induces cancer-associated fibroblast-like properties in human breast fibroblasts. Cell Death Dis. 2025;16(1):169. doi:10.1038/s41419-025-07382-w
161. Wu K, Feng J, Lyu F. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat Commun. 2021;12(1):5196. doi:10.1038/s41467-021-25473-y
162. Wang B, Mao J-H, Wang B-Y. Exosomal miR-1910-3p promotes proliferation, metastasis, and autophagy of breast cancer cells by targeting MTMR3 and activating the NF-κB signaling pathway. Cancer Lett. 2020;489:87–99. doi:10.1016/j.canlet.2020.05.038
163. Liu Y, Yang Y, Du J, et al. MiR −3613-3p from carcinoma-associated fibroblasts exosomes promoted breast cancer cell proliferation and metastasis by regulating SOCS2 expression. IUBMB Life. 2020;72(8):1705–1714. doi:10.1002/iub.2292
164. Xun J, Du L, Gao R, et al. Cancer-derived exosomal miR-138-5p modulates polarization of tumor-associated macrophages through inhibition of KDM6B. Theranostics. 2021;11(14):6847–6859. doi:10.7150/thno.51864
165. Fong MY, Zhou W, Liu L. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat Cell Biol. 2015;17(2):183–194. doi:10.1038/ncb3094
166. Zhou W, Fong M, Min Y. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell. 2014;25(4):501–515. doi:10.1016/j.ccr.2014.03.007
167. Ma C, He D, Tian P, et al. miR-182 targeting reprograms tumor-associated macrophages and limits breast cancer progression. Proc Natl Acad Sci U S A. 2022;119:119.
168. Meng Z, Zhang R, Wang Y, et al. miR-200c/PAI-2 promotes the progression of triple negative breast cancer via M1/M2 polarization induction of macrophage. Int Immunopharmacol. 2020;81:106028. doi:10.1016/j.intimp.2019.106028
169. Zhou H, Gan M, Jin X, et al. [Corrigendum] miR‑382 inhibits breast cancer progression and metastasis by affecting the M2 polarization of tumor‑associated macrophages by targeting PGC‑1alpha. Int J Oncol. 2023;62:62. doi:10.3892/ijo.2023.5510
170. Yu Z, Willmarth NE, Zhou J. microRNA 17/20 inhibits cellular invasion and tumor metastasis in breast cancer by heterotypic signaling. Proc Natl Acad Sci U S A. 2010;107(18):8231–8236. doi:10.1073/pnas.1002080107
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Comprehensive Genomic Profiling Identifies FAT1 as a Negative Regulator of EMT, CTCs, and Metastasis of Hepatocellular Carcinoma
Huang ZL, Zhang PB, Zhang JT, Li F, Li TT, Huang XY
Journal of Hepatocellular Carcinoma 2023, 10:369-382
Published Date: 7 March 2023
ITGB1BP1, a Novel Transcriptional Target of CD44-Downstream Signaling Promoting Cancer Cell Invasion
Ahmad SMS, Nazar H, Rahman MM, Rusyniak RS, Ouhtit A
Breast Cancer: Targets and Therapy 2023, 15:373-380
Published Date: 24 May 2023
The Progress of Platelets in Breast Cancer
Wang L, Zhang K, Feng J, Wang D, Liu J
Cancer Management and Research 2023, 15:811-821
Published Date: 11 August 2023
Identification and Validation of Novel Metastasis-Related Immune Gene Signature in Breast Cancer
Ma S, Hao R, Lu YW, Wang HP, Hu J, Qi YX
Breast Cancer: Targets and Therapy 2024, 16:199-219
Published Date: 12 April 2024
Pan-Cancer Screening and Validation of CALU’s Role in EMT Regulation and Tumor Microenvironment in Triple-Negative Breast Cancer
Chen SL, Hu D, Chen TZ, Shen SY, Zhao CF, Wang C, Tong SY, Liu Z, Lin SH, Jin LX, He YB, Zhang ZZ
Journal of Inflammation Research 2024, 17:6743-6764
Published Date: 25 September 2024