Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Integrative Bioinformatics Analysis and Machine Learning Reveal TSPAN7 Regulates the Involution of Infantile Hemangioma via NK Cells
Received 29 April 2026
Accepted for publication 1 July 2026
Published 14 July 2026 Volume 2026:19 619344
DOI https://doi.org/10.2147/CCID.S619344
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Michela Starace
Jianxi Bai,1,2 Sheng Lin1,2
1Pediatric Surgery Department, Fujian Children’s Hospital (Fujian Branch of Shanghai Children’s Medical Center), Fuzhou, Fujian, People’s Republic of China; 2Pediatric Surgery Department, College of Clinical Medicine for Obstetrics & Gynecology and Pediatrics, Fujian Medical University, Fuzhou, Fujian, People’s Republic of China
Correspondence: Jianxi Bai, Pediatric Surgery Department, Fujian Children’s Hospital (Fujian Branch of Shanghai Children’s Medical Center), 966 Hengyu Road, Jinan District, Fuzhou, Fujian, People’s Republic of China, Tel +8615806032159, Email [email protected]
Background: The involution of infantile hemangioma (IH) involves complex lipid metabolism reprogramming and immune modulation, but the underlying molecular mechanisms remain incompletely understood. This study aimed to identify key lipid metabolism-related genes that may drive IH involution using a bioinformatic perspective.
Methods: The GSE127487 dataset of IH was analyzed. Time-series gene expression profiles linked to involution were identified using the R package Mfuzz. Differentially expressed genes (DEGs) between proliferation and involution phases were overlapped with involution-associated temporal genes to identify key involution-related genes. Lipid-associated genes with relevance scores > 2 were retrieved from the GeneCards database. The overlap of these genes with key involution-related genes yielded lipid-related involution genes. Machine learning algorithms were applied to further screen hub genes. WGCNA was conducted to identify co-expression modules of the target hub genes during the involution phase, followed by GO and KEGG enrichment analyses. Immune cell infiltration was quantified using MCP-counter. Mediation analysis was performed to explore potential relationships between hub genes and immune cells.
Results: Three hub genes, including CFH, TSPAN7 and MIR324, were finally identified. WGCNA revealed four modules co-expressed with TSPAN7 specifically during the involution phase. Functional enrichment analysis showed that these modules were mainly enriched in “natural killer cell-mediated cytotoxicity” and “leukocyte-mediated cell killing”. MCP-counter analysis showed that NK cells were enriched in the proliferation phase. Mediation analysis revealed the direct effect of TSPAN7 on IH involution was non-significant (P = 0.474), while its indirect effect mediated by NK cells was significant (P = 0.012).
Conclusion: Three key lipid metabolism-related genes associated with IH involution (CFH, TSPAN7, and MIR324) were identified. Among them, TSPAN7 may be associated with IH involution through NK cells. These findings provide clues for further investigations into the roles of lipid-related genes and immune cells in IH involution.
Keywords: bioinformatics, immune infiltration, infantile hemangioma, involution, lipid metabolism, NK cells, TSPAN7, WGCNA
Introduction
Hemangiomas are developmental vascular anomalies that may present as hamartomas, benign tumors, or vascular malformations, and are classified as infantile or congenital types. Infantile hemangiomas (IH) are GLUT1-positive lesions, unlike congenital types, that typically appear within the first two months of life, grow rapidly during the first year, and gradually regress, whereas congenital hemangiomas are present at birth and may become symptomatic due to growth, infection, hormonal changes, or trauma.1–3 While most IH lesions undergo spontaneous regression without functional sequelae, some may cause ulceration, disfigurement, or organ dysfunction, requiring medical intervention. Propranolol remains the first-line therapy, but some patients show an incomplete response or relapse after discontinuation, highlighting the need for a deeper understanding of the molecular mechanisms underlying IH involution.4–6
The involution phase is histologically defined by the gradual replacement of proliferation endothelial cells with fibrofatty tissue, a process driven by the differentiation of hemangioma stem cells (HemSCs) into adipocytes.7–9 Emerging evidence suggests that lipid metabolism plays a pivotal role in this transition. Transcriptomic analyses have revealed significant alterations in lipid-associated gene expression between proliferation and involution IH tissues.10,11 The activation of key adipogenic regulators such as peroxisome proliferator-activated receptor gamma (PPARγ) has been shown to promote hemangioma-derived mesenchymal stem cell (Hem-MSC) differentiation toward an adipocytic lineage while suppressing angiogenesis, thereby facilitating IH involution.12,13 Moreover, metabolic pathway analyses have indicated substantial metabolic reprogramming during IH involution, but the integration of lipid-related changes with the immune microenvironment remains poorly characterized.
Recent bioinformatics studies have used the GSE127487 dataset to explore gene expression dynamics across IH phases. Weighted gene co-expression network analysis (WGCNA) and machine learning approaches have identified candidate genes such as SH3KBP1 associated with disease progression, and immune infiltration analyses have suggested dynamic shifts in immune cell populations during proliferative and involutive stages.14 However, systematic screening for lipid metabolism-related genes that drive involution and elucidation of their interplay with immune cells have not been performed.
In this study, we integrated time-series expression analysis, differential expression profiling, and machine learning algorithms to identify key lipid metabolism-related genes associated with IH involution using the GSE127487 dataset. Through WGCNA co-expression analysis, immune infiltration assessment, and mediation analysis, we explored the potential relationships between hub genes and immune cells. These findings may provide clues for further investigations into the roles of lipid-related genes and immune cells in IH involution.
Methods
Data Processing and Differential Expression Analysis
Figure 1 illustrates the technical workflow of this study. The GSE127487 microarray dataset was downloaded from the Gene Expression Omnibus (GEO) database using the R package GEOquery. The dataset was generated on a GPL10558 platform (Illumina HumanHT-12 V4.0 expression beadchip). A total of 23 IH tissue samples and five normal skin tissue samples from Homo sapiens were included. Among these, five IH samples were obtained from patients who received propranolol treatment.15 For subsequent analyses, we selected six untreated proliferation-phase IH (PIH) samples and six untreated involution-phase IH (IIH) samples to avoid confounding factors caused by treatment.
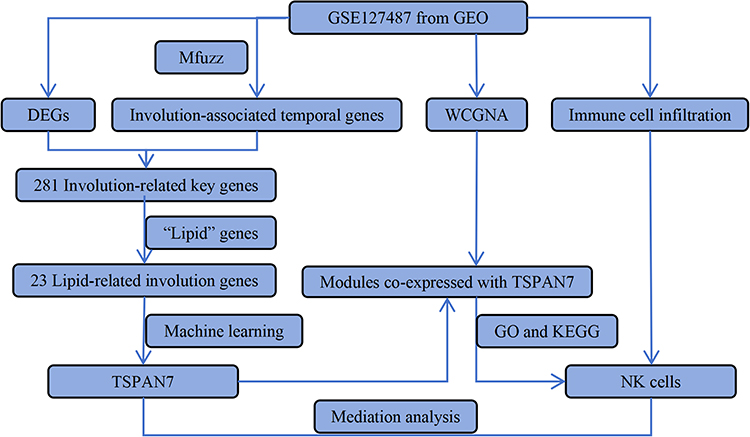 |
Figure 1 Diagram of the study work flow. |
Time-series gene expression patterns across 6, 12, and 24 months were analyzed using the Mfuzz package in R, which employs fuzzy c-means clustering to identify clusters with distinct temporal trajectories.
Differentially expressed genes (DEGs) between PIH and IIH samples were identified using the limma package with thresholds of |log2 fold change (logFC)| > 2 and P-value < 0.05. Key involution-related genes were identified from the overlap between involution-associated temporal genes and DEGs.
Lipid metabolism-related genes were retrieved from the GeneCards database using the keyword “lipid” and genes with a relevance scores > 2 were retained. Overlapping these lipid-associated genes with key involution-related genes generated lipid-related involution genes.
Machine Learning-Based Hub Gene Screening
Three machine learning algorithms were applied to identify hub genes from lipid-related involution genes.
Least absolute shrinkage and selection operator (LASSO) logistic regression was performed using the glmnet R package with 10-fold cross-validation. The optimal regularization parameter λ was selected as the value within one standard error of the minimum cross-validated deviance (λ.1se), balancing model sparsity and predictive performance. Genes with non-zero coefficients at the selected λ were retained as candidates.
Support vector machine with recursive feature elimination (SVM-RFE) was implemented using the e1071 and caret R packages with a linear kernel. Features were recursively eliminated based on the absolute values of the SVM weight vector, and the optimal feature subset was determined as the set that yielded the highest cross-validation accuracy.
Random forest (RF) was performed using the randomForest R package with 500 trees. Feature importance was computed by the mean decrease in Gini index, and the top 10 genes with the highest importance scores were retained.
To prevent overfitting and ensure model stability, 10-fold cross-validation repeated three times was employed for model tuning. Both LASSO and RF models used this cross-validation scheme. For SVM-RFE, feature selection was nested within cross-validation loops to avoid information leakage.
The genes identified by the three algorithms were considered hub genes. This intersection strategy reduces algorithm-specific bias and enhances the robustness of feature selection given the limited sample size.
Weighted Gene Co-Expression Network Analysis (WGCNA)
WGCNA was performed using the WGCNA package to construct co-expression networks specific to involution phase. Soft-thresholding power was selected based on scale-free topology fit. Modules with high similarity were merged and module eigengenes were calculated. Pearson’s correlation was used to identify modules significantly associated with TSPAN7 expression in involution phase. Modules with a correlation |r| > 0.5 and P < 0.05, were selected for further GO and KEGG enrichment analysis.
Functional Enrichment Analysis
To explore the biological functions and signaling pathways of the identified modules, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed using the clusterProfiler package in R. Enriched terms with P < 0.05 were considered statistically significant.
Immune Infiltration Analysis
The MCP-counter algorithm was applied to quantify the absolute abundance of ten immune cell populations and stromal components in each sample using the MCP-counter package. Differences in immune cell infiltration between the PIH and IIH groups were assessed using the Wilcoxon rank-sum test.
Mediation Analysis
A mediation analysis was performed to evaluate whether the effect of TSPAN7 on IH involution was mediated by immune cells. Linear regression models were constructed using the mediation package in R with bootstrapping (1000 simulations) to estimate the direct, indirect, and total effects. The significance of the mediation effect was determined by using a 95% confidence interval.
Statistical Analysis
All statistical analyses were performed using the R software (version 4.2.0). For comparisons between two groups, normally distributed variables were analyzed using the Student’s t-test, while non-normally distributed variables were compared using the Mann–Whitney U-test (Wilcoxon rank-sum test). Multiple testing corrections were applied using the Benjamini-Hochberg false discovery rate (FDR) method, where appropriate. A two-sided P < 0.05 was considered statistically significant.
Results
Identification of Involution-Related Lipid-Associated Hub Genes
Time-series expression analysis using Mfuzz clustering across 6, 12, and 24 months identified nine distinct expression clusters (Figure 2). Based on their temporal trajectories, clusters exhibiting gradually increasing expression (cluster 1), gradually decreasing expression (cluster 4), or stable expression during 6 and 12 months followed by specific downregulation during 24months (cluster 6) were selected as involution-associated temporal genes. Differential expression analysis of proliferation-phase IH (PIH) and involution-phase IH (IIH) samples identified 399 DEGs, including 241 upregulated and 158 downregulated genes (|logFC| > 2, P < 0.05). Intersection analysis of involution-associated temporal genes and DEGs identified 281 core genes linked to involution.
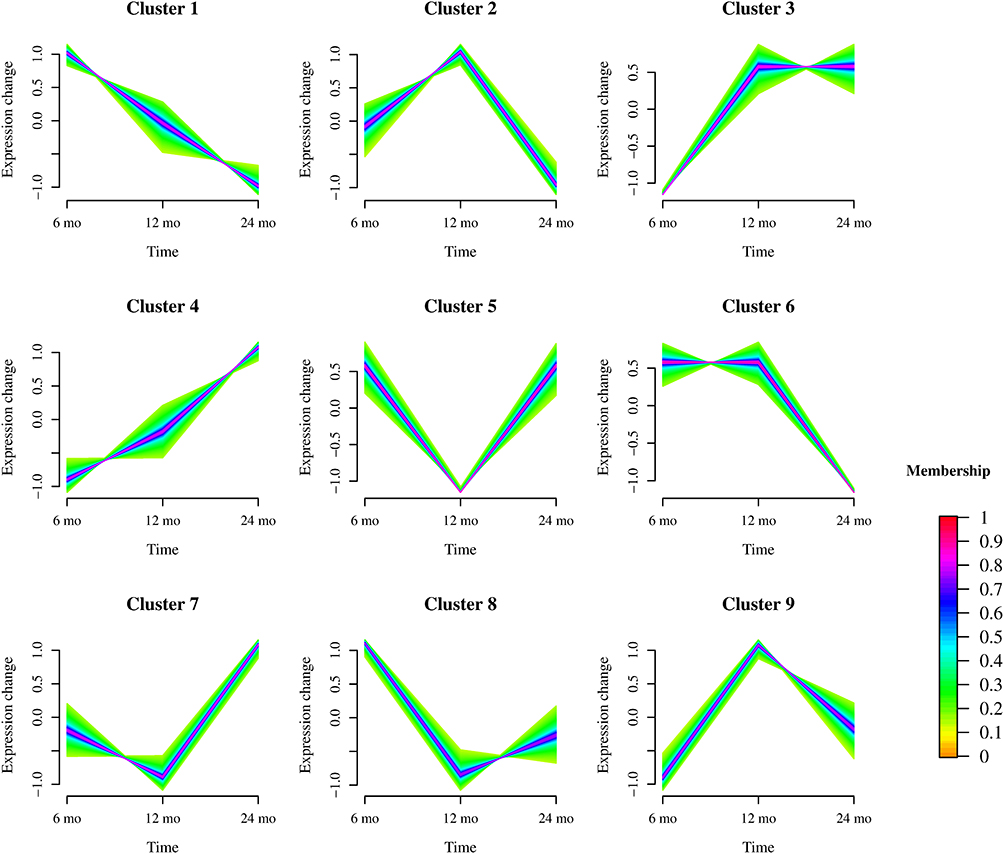 |
Figure 2 Time-series expression patterns of genes across infantile hemangioma phases (6, 12, and 24 months) revealed by Mfuzz clustering. The x-axis represents the time points (proliferation phase at 6 months, stable phase at 12 months, and involution phase at 24 months), and the y-axis represents relative gene expression levels. Nine distinct clusters (Cluster 1–Cluster 9) were identified, each exhibiting a unique temporal trajectory. Colors indicate the membership degree of each gene in the assigned cluster, with red representing high membership and yellow representing low membership. |
A total of 3773 lipid-related genes with relevance scores > 2 were retrieved from the GeneCards database. Overlapping these genes with 281 involution-related key genes identified 23 lipid-related involution genes (Figure 3A). Three machine learning algorithms, LASSO, SVM-RFE, and RF were applied to further refine these candidates. LASSO regression identified 12 genes with non-zero coefficients (Figure 3B and C). SVM-RFE selected seven genes with optimal feature importance (Figure 3D), and random forest analysis ranked the genes based on the mean decrease in accuracy and the Gini coefficient (Figure 3E). By intersecting candidate genes screened via the three algorithms, we identified three hub genes: CFH, TSPAN7, and MIR324 (Figure 3F). Among them, TSPAN7 and CFH displayed a gradually rising expression trend from the proliferation stage to the involution stage (cluster 4). In contrast, MIR324 maintained stable expression at 6 and 12 months, followed by distinct downregulation at 24 months.
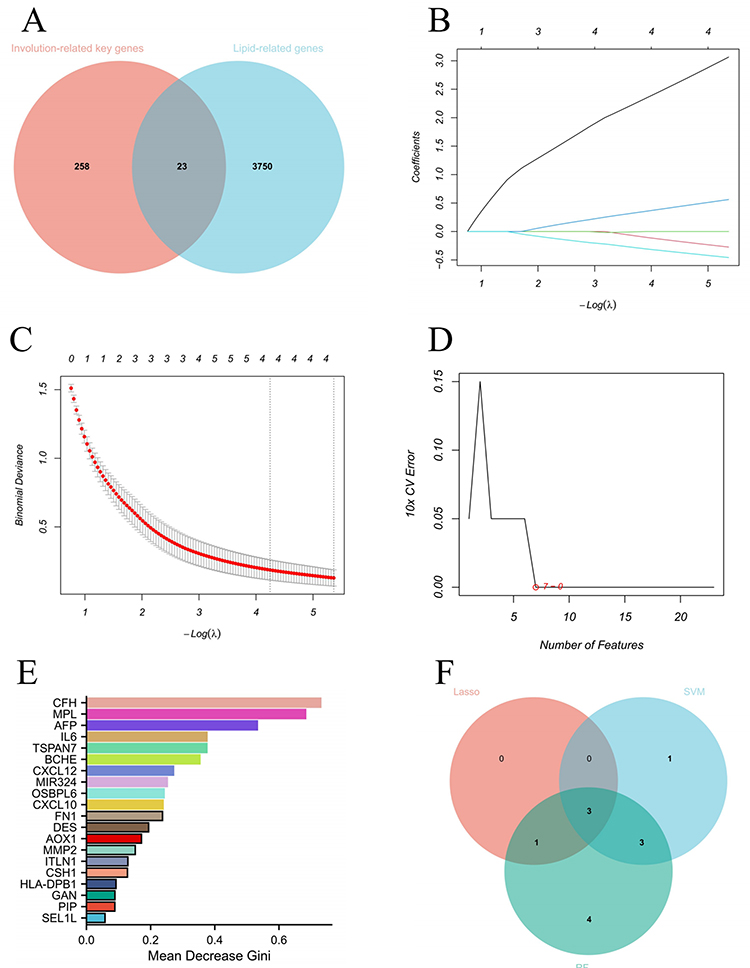 |
Figure 3 Identification of involution-related lipid-associated hub genes. (A) Venn diagram showing the intersection of involution-related key genes and lipid-associated genes, yielding 23 lipid-related involution genes. (B and C) LASSO regression analysis identified 12 genes with non-zero coefficients (B: Coefficient profile; (C) Binomial deviance). (D) SVM-RFE selected 7 genes with optimal feature importance. (E) Random forest analysis ranking genes based on Mean Decrease Gini (>0.2). (F) Venn diagram illustrating the overlap of genes selected by the three machine learning algorithms (LASSO, SVM-RFE, and random forest), identifying three hub genes (CFH, TSPAN7, MIR324). |
Weighted Gene Co-Expression Network Analysis Identifies TSPAN7-Associated Modules
To explore functional networks co-expressed with TSPAN7, WGCNA was performed. A soft-thresholding power of 8 was selected to achieve a scale-free topology fit (R2 = 0.85) (Figure 4A). Hierarchical clustering identified 20 co-expression modules (Figure 4B). Among these, four modules (blue, brown, turquoise, and yellow) were significantly correlated with TSPAN7 expression (correlation |r| > 0.5, P < 0.05) (Figure 5). These modules were selected for the functional enrichment analysis.
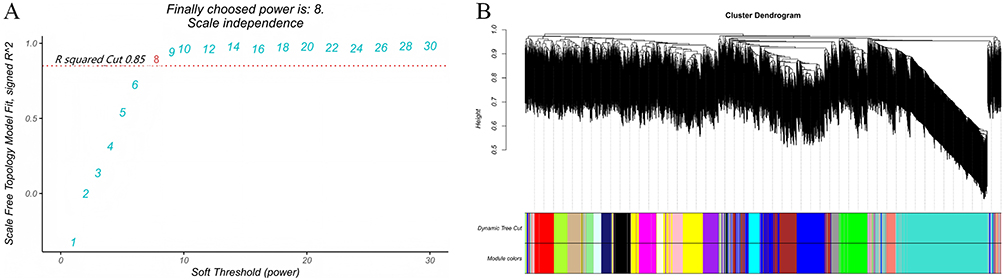 |
Figure 4 Weighted gene co-expression network analysis (WGCNA). (A) Analysis of scale-free topology fit. A soft-thresholding power of 8 was selected to achieve a scale-free topology fit (R2 = 0.85). (B) Hierarchical clustering dendrogram showing the identification of 20 co-expression modules, each labeled with a distinct color. |
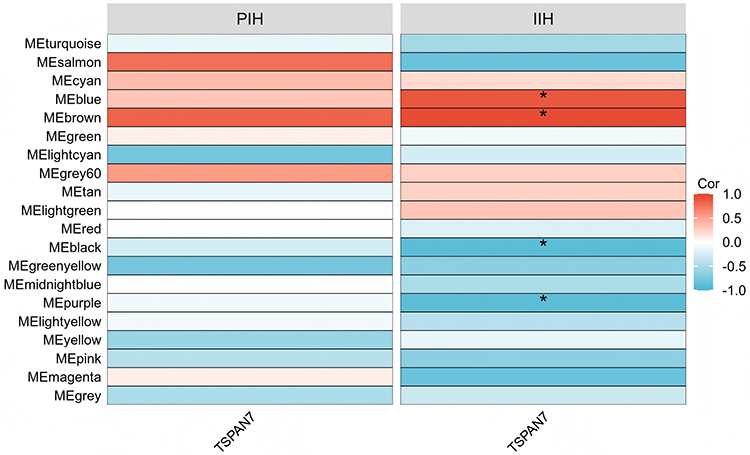 |
Figure 5 Correlation between TSPAN7 and module eigengenes (MEs) in PIH and IIH. Red indicates positive correlation, blue indicates negative correlation. *P < 0.05. |
GO and KEGG enrichment analyses of the four TSPAN7-associated modules revealed a striking convergence in immune-related functions. The most significantly enriched biological processes included “natural killer cell-mediated cytotoxicity”, “leukocyte-mediated cell killing”, “regulation of natural killer cell-mediated immunity”, and “adaptive immune response”. These findings suggest a potential association between TSPAN7 and NK cells during IH involution.
Immune Infiltration Landscape During IH Involution
To characterize immune cell dynamics, MCP-counter analysis was performed to estimate immune cell abundance in PIH and IIH samples. The results demonstrated a significant enrichment of NK cells in the proliferation phase compared to that in the involution phase. In contrast, myeloid dendritic cells were significantly more abundant during the involution phase (Figure 6).
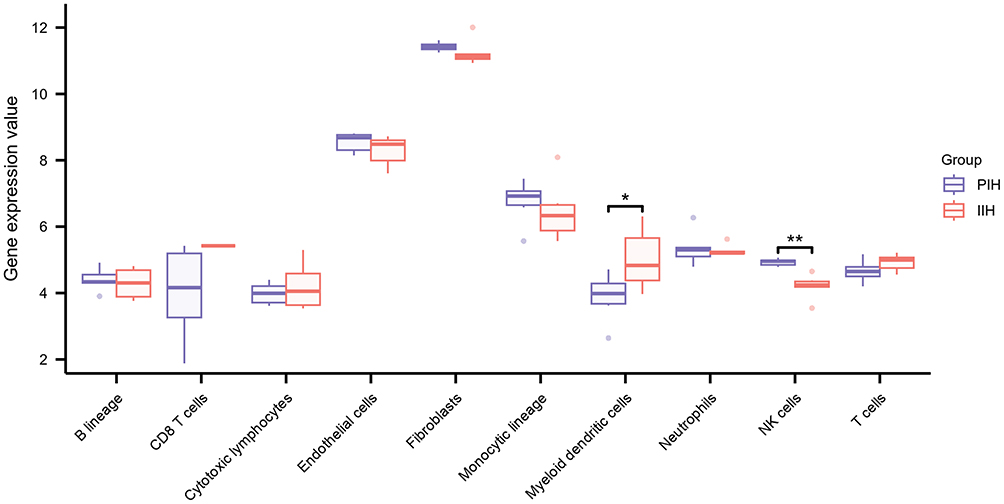 |
Figure 6 Immune cell infiltration landscape in proliferation and involution phases of IH assessed by MCP-counter. The x-axis represents immune cell populations, and the y-axis represents relative gene expression value. *P < 0.05; **P < 0.01. |
TSPAN7 Promotes Involution Through NK Cells
Given the functional enrichment of NK cells in TSPAN7-associated co-expression modules and the observed enrichment of NK cells during proliferation, we hypothesized that NK cells might mediate the effect of TSPAN7 on involution. Mediation analysis was performed to test this hypothesis, with TSPAN7 expression as the independent variable, NK cells abundance as the mediator, and involution status as the outcome variable.
The results revealed the existence of a complete mediation model. The direct effect of TSPAN7 on involution was not significant (Effect= 0.1113, P = 0.4740), whereas the indirect effect mediated by NK cells was highly significant (Effect= 0.1293, P = 0.0120). The total effect of TSPAN7 on involution was significant (Effect= 0.2406, P = 0) (Table 1). Path decomposition showed that TSPAN7 was negatively associated with NK cells abundance (path a = −0.2095, P < 0.01), and that, after controlling for TSPAN7, NK cells abundance was also negatively associated with involution (path b = −0.6174). The product of these two negative paths (a × b) yields a positive indirect effect.
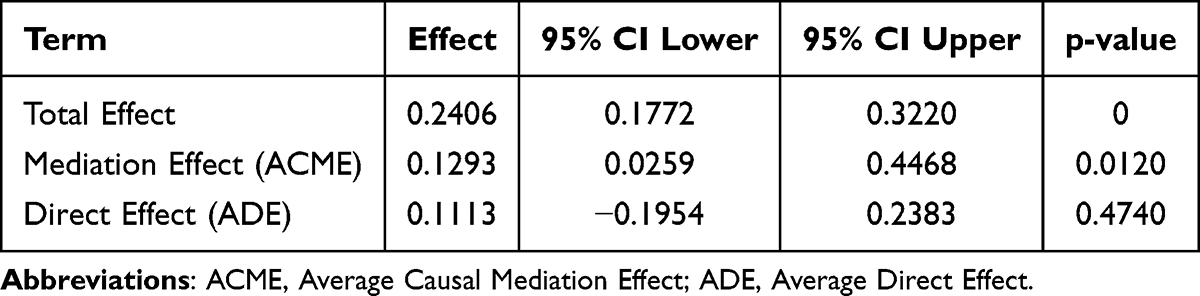 |
Table 1 Mediation Analysis Results of TSPAN7 and NK Cells |
Discussion
IH is distinguished by its unique capacity for spontaneous involution, but the precise molecular mechanisms orchestrating this regression remain unclear.16,17 In a previous bioinformatics study by our group, we identified IL6, EGFR, IRF1 and IL32 as key pyroptosis-related genes involved in the regression of IH using the same GSE127487 dataset.18 The present study extends that investigation by focusing on lipid-associated genes. Although lipid metabolism reprogramming has been implicated in the transition from the proliferative to involution phases,13,19,20 the interplay between lipid-associated genes and the immune microenvironment during IH involution has not been systematically characterized. By integrating time-series clustering and machine learning algorithms (LASSO, SVM-RFE, and random forest), we newly identified CFH, MIR324 and TSPAN7 as key lipid metabolism-related genes that may associated with IH involution. Importantly, WGCNA and mediation analysis revealed that the effect of TSPAN7 may be associated with NK cells during IH involution, a possibility not explored in our prior work.
Three hub lipid-related involution genes were identified: CFH, TSPAN7, and MIR324. CFH belongs to cluster 4, exhibiting a progressively increasing expression pattern during IH involution. CFH (Complement Factor H) is a key regulator of the alternative complement pathway and has been shown to participate in lipid metabolism. In human adipose tissue, high expression of the CFH gene is present in the stromal vascular fraction cells (comprising monocytes, fibroblasts, and endothelial cells), where it mediates the activation of the alternative complement pathway and participates in lipid metabolism reprogramming and insulin resistance.21 CFH can also bind to ox-LDL, directly participating in the clearance of oxidized lipids and affecting lipid deposition.22 The stromal vascular fraction cells of the involution phase of IH may highly express CFH, potentially participating in local lipid clearance, transport, or deposition, thereby directly or indirectly influencing adipose tissue formation following vascular regression. The MIR324 gene simultaneously encodes miR-324-5p and miR-324-3p. These two mature microRNA molecules originate from the same precursor hairpin structure but exhibit significant functional differences and have crucial impacts on the pathogenesis of a wide range of disorders.23 miR-324-3p is downregulated in proliferation-phase IH tissues and functions as a tumor suppressor by targeting the PDRG1 pathway.24 MIR324 plays different roles in different tumor microenvironments, and its role in IH involution warrants further investigation. TSPAN7 exhibited a progressively increasing expression pattern across IH developmental phases, suggesting its potential role in promoting involution. TSPAN7, a member of the tetraspanin family, regulate cell adhesion, migration, and immune synapse formation.25 Previous studies have demonstrated that tetraspanins can modulate immune cell function by organizing membrane microdomains that facilitate receptor clustering and signaling.26,27 However, its specific role in IH involution has not yet been reported.
The functional enrichment analysis of TSPAN7-associated co-expression modules revealed a striking convergence on immune-related pathways, particularly “natural killer cell-mediated cytotoxicity” and “leukocyte-mediated cell killing”, suggesting that TSPAN7 may participate in IH involution together with NK cells. The immune microenvironment of IH has been increasingly recognized as a critical regulator of disease progression and involution. Macrophages undergo a phenotypic switch from M2-predominant in the proliferative phase to M1-predominant in the involution phase.28 Single-cell RNA sequencing has further resolved the cellular heterogeneity of IH, confirming that macrophages, mast cells, and telocytes dynamically change during involution.10 Several immune infiltration analyses have revealed alterations in NK cells in IH,14,29 and accumulating evidence from tumor angiogenesis and vascular biology studies supports the possibility of NK cells-mediated vascular regression.30–32 However, the role of NK cells in IH has remained largely unexplored. Our immune infiltration analysis revealed that NK cells were significantly enriched in proliferation phase compared to involution phase IH tissues, indicating a phase-specific distribution of NK cells during IH progression. The precise role of NK cells in IH involution remains to be further explored.
Pan-cancer analysis revealed that TSPAN7 expression was positively associated with the infiltration of resting NK cells in renal clear cell carcinoma, supporting a potential link between TSPAN7 and NK cells.33 This finding prompted us to explore the potential relationship between NK cells and TSPAN7 during IH involution. The complete mediation model demonstrated that the effect of TSPAN7 on IH involution may depend on NK cells, as the direct effect was not significant, whereas the indirect effect mediated by NK cells was highly significant. This finding suggests that TSPAN7 may be associated with IH involution through NK cells. Although direct evidence for TSPAN7-mediated NK cells regulation is lacking, tetraspanins are known to induce cytoskeleton reorganization, which may modulates NK cells activation.34 It should be noted that the finding that TSPAN7 expression correlates negatively with NK cells abundance yet exerts its pro-involution effect through NK cells (complete mediation) may initially appear counterintuitive. We propose that this pattern reflects a quantitative-qualitative trade-off in NK cells biology during IH involution. In the context of IH, TSPAN7 upregulation on hemangioma endothelial cells or infiltrating immune cells may enhance the cytotoxic efficiency of each NK cells, leading to more effective clearance of proliferation endothelium. Following target elimination, activated NK cells may undergo activation-induced cell death (AICD).35,36 Consequently, the decline in NK cells number at the involution stage may be not a cause of reduced immune surveillance but rather a reflection of successful task completion. The complete mediation model thus indicates that the pro-involution effect of TSPAN7 is conditional upon the presence and activation of NK cells, even if their numerical abundance has started to wane by the time TSPAN7 peaks. However, we acknowledge that causal mediation analysis relies on strong assumptions that are unlikely to be fully satisfied in observational transcriptomic data, including no unmeasured confounding between the exposure and the mediator, no unmeasured confounding between the mediator and the outcome, no exposure-mediator interaction, and correct temporal ordering of variables. Given the cross-sectional nature of the GSE127487 dataset and the limited sample size, these assumptions cannot be rigorously verified. Therefore, the results of the mediation analysis should be interpreted exploratorily, as hypothesis-generating evidence rather than causal inference. We hypothesize that TSPAN7 may modulate NK cells function or turnover, but this requires experimental validation.
This study has several limitations. First, our findings were derived entirely from bioinformatics analyses of public microarray data and require validation through experimental approaches, including in vitro and in vivo models. Second, the precise molecular mechanisms by which TSPAN7 regulates NK cells function in IH remain to be determined. Third, although we excluded propranolol-treated samples to avoid confounding effects, the limited sample size may affect the robustness of our analyses, particularly for WGCNA and machine learning, which typically require larger cohorts. Therefore, our findings should be considered exploratory and hypothesis-generating, requiring validation in independent and larger datasets. Fourth, the roles of other lipid-related hub genes identified in this study, including CFH and MIR324, warrant further investigation.
In conclusion, this study identified three key lipid metabolism-related genes associated with IH involution (CFH, TSPAN7, and MIR324). Among them, TSPAN7 may be associated with IH involution through NK cells. The precise mechanisms of these molecules during IH involution require further experimental investigation. These findings provide clues for further investigations into the roles of lipid-related genes and immune cells in IH involution.
Data Sharing Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The research utilizes legally obtained public data, does not cause harm to the human body, does not involve sensitive personal information or commercial interests, and is therefore exempt from ethical review by the Ethics Committee of Fujian Children’s Hospital (IRB NO. 2024-002).
Consent for Publication
All authors approved the publication of the manuscript.
Acknowledgments
We would like to acknowledge the GEO databases for providing professional platforms.
Funding
Startup Fund for Scientific Research, Fujian Medical University (grant number: 2022QH1224). Fujian Provincial Natural Science Foundation (grant number: 2024J011133). Medicine Research Project of Fujian Children’s Hospital (grant number: ETK2025007).
Disclosure
The authors report no competing interests in this work.
References
1. Rodríguez Bandera AI, Sebaratnam DF, Wargon O, Wong LF. Infantile hemangioma. Part 1: epidemiology, pathogenesis, clinical presentation and assessment. J Am Acad Dermatol. 2021;85(6):1379–11. doi:10.1016/j.jaad.2021.08.019
2. Holm A, Mulliken JB, Bischoff J. Infantile hemangioma: the common and enigmatic vascular tumor. J Clin Invest. 2024;134(8). doi:10.1172/JCI172836
3. Hasan S, Khan A, Banerjee A, Ramalingam K. Infantile hemangioma of the upper lip: report of a rare case with a brief review of literature. Cureus. 2023;15(7):e42556. doi:10.7759/cureus.42556
4. Sebaratnam DF, Rodríguez Bandera AL, Wong LF, Wargon O. Infantile hemangioma. Part 2: management. J Am Acad Dermatol. 2021;85(6):1395–1404. doi:10.1016/j.jaad.2021.08.020
5. Li GX, Sebaratnam DF. Dermatology images: infantile hemangioma. J Am Acad Dermatol. 2026;94(2):720–729. doi:10.1016/j.jaad.2025.07.066
6. Chen Q, Yang L, Ni S, et al. Single-cell RNA sequencing discovered subtypes associated with angiogenesis and propranolol treatment in infantile hemangioma. Genomics Proteomics Bioinf. 2026. doi:10.1093/gpbjnl/qzag023
7. Chen Q, Zheng J, Bian Q. Cell fate regulation during the development of infantile hemangioma. J Invest Dermatol. 2025;145(2):266–279. doi:10.1016/j.jid.2024.06.1275
8. Yu Y, Fuhr J, Boye E, et al. Mesenchymal stem cells and adipogenesis in hemangioma involution. Stem Cells. 2006;24(6):1605–1612. doi:10.1634/stemcells.2005-0298
9. Roach EE, Chakrabarti R, Park NI, et al. Intrinsic regulation of hemangioma involution by platelet-derived growth factor. Cell Death Dis. 2012;3:e328. doi:10.1038/cddis.2012.58
10. Chen Q, Rong H, Zhang L, Wang Y, Bian Q, Zheng J. KLF2 orchestrates pathological progression of infantile hemangioma through hemangioma stem cell fate decisions. J Invest Dermatol. 2024;144(8):1850–1864.e9. doi:10.1016/j.jid.2024.01.029
11. Calicchio ML, Collins T, Kozakewich HP. Identification of signaling systems in proliferation and involution phase infantile hemangiomas by genome-wide transcriptional profiling. Am J Pathol. 2009;174(5):1638–1649. doi:10.2353/ajpath.2009.080517
12. Wang Y, Kong L, Sun B, Cui J, Shen W. Celecoxib induces adipogenic differentiation of hemangioma-derived mesenchymal stem cells through the PPAR-γ pathway in vitro and in vivo. Exp Ther Med. 2022;23(6):375. doi:10.3892/etm.2022.11303
13. Yuan SM, Guo Y, Xu Y, Wang M, Chen HN, Shen WM. The adipogenesis in infantile hemangioma and the expression of adipogenic-related genes. Int J Clin Exp Pathol. 2017;10(12):11596–11602.
14. Yin J, Gou H, Qi J, Xing W. Feature gene selection and functional validation of SH3KBP1 in infantile hemangioma using machine learning. Biochem Biophys Res Commun. 2025;752:151469. doi:10.1016/j.bbrc.2025.151469
15. Gomez-Acevedo H, Dai Y, Strub G, Shawber C, Wu JK, Richter GT. Identification of putative biomarkers for infantile hemangiomas and propranolol treatment via data integration. Sci Rep. 2020;10(1):3261. doi:10.1038/s41598-020-60025-2
16. Surlis T, De Sa Reilly H, Sadlier M, Nelson J. Infantile haemangiomas. BMJ. 2022;378:e068734. doi:10.1136/bmj-2021-068734
17. Zhu L, Xie J, Liu Z, et al. Pigment epithelium-derived factor/vascular endothelial growth factor ratio plays a crucial role in the spontaneous regression of infant hemangioma and in the therapeutic effect of propranolol. Cancer Sci. 2018;109(6):1981–1994. doi:10.1111/cas.13611
18. Liu L, Lin S, Bai J, Zhang B. Integrative bioinformatics analysis of pyroptosis-related genes and analysis of immune cell infiltration in infantile hemangioma regression. Clin Cosmet Invest Dermatol. 2025;18:291–302. doi:10.2147/CCID.S492535
19. Yuan SM, Guo Y, Wang Q, et al. Over-expression of PPAR-γ2 gene enhances the adipogenic differentiation of hemangioma-derived mesenchymal stem cells in vitro and in vivo. Oncotarget. 2017;8(70):115817–115828. doi:10.18632/oncotarget.23705
20. Li N, Gong X, Cao Z, et al. Myosin 1b inhibits the phenotype of HemECs to affect the progression of infantile hemangiomas. Clin Cosmet Invest Dermatol. 2025;18:989–998. doi:10.2147/CCID.S517750
21. Moreno-Navarrete JM, Martínez-Barricarte R, Catalán V, et al. Complement factor H is expressed in adipose tissue in association with insulin resistance. Diabetes. 2010;59(1):200–209. doi:10.2337/db09-0700
22. Shaw PX, Zhang L, Zhang M, et al. Complement factor H genotypes impact risk of age-related macular degeneration by interaction with oxidized phospholipids. Proc Natl Acad Sci U S A. 2012;109(34):13757–13762. doi:10.1073/pnas.1121309109
23. Kadkhoda S, Hussen BM, Eslami S, Ghafouri-Fard S. A review on the role of miRNA-324 in various diseases. Front Genet. 2022;13:950162. doi:10.3389/fgene.2022.950162
24. Zhao T, Zhang J, Ye C, Tian L, Li Y. lncRNA FOXD2-AS1 promotes hemangioma progression through the miR-324-3p/PDRG1 pathway. Cancer Cell Int. 2020;20:189. doi:10.1186/s12935-020-01277-w
25. Shao S, Bu Z, Xiang J, et al. The role of Tetraspanins in digestive system tumor development: update and emerging evidence. Front Cell Dev Biol. 2024;12:1343894. doi:10.3389/fcell.2024.1343894
26. Querol Cano L, Dunlock VE, Schwerdtfeger F, van Spriel AB. Membrane organization by tetraspanins and galectins shapes lymphocyte function. Nat Rev Immunol. 2024;24(3):193–212. doi:10.1038/s41577-023-00935-0
27. Rocha-Perugini V, Sánchez-Madrid F, Martínez Del Hoyo G. Function and dynamics of tetraspanins during antigen recognition and immunological synapse formation. Front Immunol. 2015;6:653. doi:10.3389/fimmu.2015.00653
28. Zhu Z, Jiang X, Wang F, Zhao L, Song Q. Decoding infantile hemangioma: cellular dynamics, molecular signals, and microenvironmental influences. Front Oncol. 2025;15:1675194. doi:10.3389/fonc.2025.1675194
29. Chen ZT, Cui MJ, Guo SK, et al. Single-cell RNA sequencing reveals bidirectional development of infantile hemangioma. J Invest Dermatol. 2026;146(5):1344–1353. doi:10.1016/j.jid.2025.10.593
30. Kurebayashi Y, Olkowski CP, Lane KC, et al. Rapid depletion of intratumoral regulatory t cells induces synchronized CD8 T- and NK-cell activation and IFNγ-dependent tumor vessel regression. Cancer Res. 2021;81(11):3092–3104. doi:10.1158/0008-5472.CAN-20-2673
31. Tran T, Galdina V, Urquidi O, et al. Assessment of NK cytotoxicity and interactions with porcine endothelial cells by live-cell imaging in 2D static and 3D microfluidic systems. Sci Rep. 2024;14(1):24199. doi:10.1038/s41598-024-75217-3
32. van Weel V, Toes RE, Seghers L, et al. Natural killer cells and CD4+ T-cells modulate collateral artery development. Arterioscler Thromb Vasc Biol. 2007;27(11):2310–2318. doi:10.1161/ATVBAHA.107.151407
33. Lu B, Liu Y, Yao Y, et al. Unveiling the unique role of TSPAN7 across tumors: a pan-cancer study incorporating retrospective clinical research and bioinformatic analysis. Biol Direct. 2024;19(1):72. doi:10.1186/s13062-024-00516-8
34. Ménager MM. TSPAN7, effector of actin nucleation required for dendritic cell-mediated transfer of HIV-1 to T cells. Biochem Soc Trans. 2017;45(3):703–708. doi:10.1042/BST20160439
35. Guilz NC, Ahn YO, Fatima H, et al. Replication stress in activated human NK cells induces sensitivity to apoptosis. J Immunol. 2024;213(1):40–51. doi:10.4049/jimmunol.2300843
36. Stanzer S, Janesch B, Resel M, Augustin T, Samonigg H, Bauernhofer T. The role of activation-induced cell death in the higher onset of spontaneous apoptosis of NK cells subsets in patients with metastatic epithelial cancer. Cell Immunol. 2010;261(2):99–104. doi:10.1016/j.cellimm.2009.11.006
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
DOCK4 as a Potential Biomarker Associated with Immune Infiltration in Stomach Adenocarcinoma: A Database Analysis
Lu Y, Yu J, Dong Q, Du Y, Liang Z
International Journal of General Medicine 2022, 15:6127-6143
Published Date: 11 July 2022
Lipid Metabolic-Related Signature CYP19A1 is a Potential Biomarker for Prognosis and Immune Cell Infiltration in Gastric Cancer

Wang N, Huang X, Long Q
Journal of Inflammation Research 2022, 15:5075-5088
Published Date: 5 September 2022
Identification of an Immune-Related Gene Diagnostic Model and Potential Drugs in Sepsis Using Bioinformatics and Pharmacogenomics Approaches
Chen P, Chen J, Ye J, Yang L
Infection and Drug Resistance 2023, 16:5665-5680
Published Date: 28 August 2023
Role of Kinetochore Scaffold 1 (KNL1) in Tumorigenesis and Tumor Immune Microenvironment in Pan-Cancer: Bioinformatics Analyses and Validation of Expression
Ding Y, Wang K, Zhao S, Li Y, Qiu W, Zhu C, Wang Y, Dong C, Liu J, Lu Y, Qi W
International Journal of General Medicine 2023, 16:4883-4906
Published Date: 31 October 2023
Screening of Lipid Metabolism-Related Genes as Diagnostic Indicators in Chronic Obstructive Pulmonary Disease
Jiang C, Peng M, Dai Z, Chen Q
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:2739-2754
Published Date: 28 November 2023