")
Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 18
Screening of Lipid Metabolism-Related Genes as Diagnostic Indicators in Chronic Obstructive Pulmonary Disease
Authors Jiang C , Peng M , Dai Z, Chen Q
Received 10 August 2023
Accepted for publication 11 November 2023
Published 28 November 2023 Volume 2023:18 Pages 2739—2754
DOI https://doi.org/10.2147/COPD.S428984
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Min Zhang
Chen Jiang,1,2,* Meijuan Peng,1,2,* Ziyu Dai,1,2 Qiong Chen1,2
1Department of Geriatrics, Respiratory Medicine, Xiangya Hospital, Central South University, Changsha, 410008, China; 2National Clinical Research Center for Geriatric Disorders, Xiangya Hospital, Central South University, Changsha, 410008, China
*These authors contributed equally to this work
Correspondence: Qiong Chen, National Clinical Research Center for Geriatric Disorders, Xiangya Hospital, Central South University, No. 87, Xiangya Road, Kaifu District, Changsha, Hunan, 410008, China, Email [email protected]
Objective: It has been observed that local and systemic disorders of lipid metabolism occur during the development of chronic obstructive pulmonary disease (COPD), but no specific mechanism has yet been identified.
Methods: The mRNA microarray dataset GSE76925 of COPD patients was downloaded from the Gene Expression Omnibus database and screened for differentially expressed genes (DEGs). Lipid metabolism-related genes (LMRGs) were extracted from the Kyoto Encyclopedia of Genes and Genomes database and Molecular Signature Database. The DEGs were intersected with LMRGs to obtain differentially expressed lipid metabolism-related genes (DeLMRGs). GO enrichment analysis and KEGG pathway analysis were performed on DeLMRGs, and protein-protein interaction networks were constructed and screened to identify hub genes. The GSE8581 validation set and further ELISA experiments were used to validate key DeLMRG expression.
Results: Differential analysis of dataset GSE76925 identified 587 DEGs, of which 62 genes were up-regulated and 525 were down-regulated. Taking the intersection of 587 DEGs with 1102 LMRGs, 20 DeLMRGs were obtained, including 1 up-regulated gene and 19 down-regulated genes. 10 hub genes were screened by cytohubba plugin, including 9 down-regulated genes PLA2G4A, HPGDS, LEP, PTGES3, LEPR, PLA2G2D, MED21, SPTLC1 and BCHE, as well as the only up-regulated gene PLA2G7. Validation of the identified 10 DeLMRGs using the validation set GSE8581 revealed that BCHE and PLA2G7 expression levels differed between the two groups. We further constructed the ceRNA network of BCHE and PLA2G7. Cell experiments also showed that PLA2G7 expression was up-regulated and BCHE expression was down-regulated in CSE-treated RAW264.7 and THP-1 cells.
Conclusion: Based on a comprehensive bioinformatic analysis of lipid metabolism genes, we identified BCHE and PLA2G7 as potentially significant biomarkers of COPD. These biomarkers may represent promising targets for COPD diagnosis and treatment.
Keywords: COPD, lipid metabolism, bioinformatics, BHCE, PLA2G7
Introduction
Chronic obstructive pulmonary disease (COPD) is a chronic respiratory disease characterized by irreversible and progressive airflow obstruction and chronic airway inflammation.1 The latest data show that an age-standardized incidence rate of COPD is 200.49 per 100,000 people worldwide, with smoking being a major environmental risk factor.2 The pathogenesis of COPD is still not fully understood and could be attributed to the overexpression of inflammatory mediators and cytokines, activation of inflammatory signaling pathways, protease/anti-protease imbalance and oxidation-antioxidant imbalance.3 An FEV1/FVC ratio less than 0.70 is considered diagnostic of COPD according to the Global Initiative for Obstructive Lung Disease (GOLD).4 It has been noted that the emergence of some novel COPD markers has improved clinical decision-making.5–8 The specific mechanisms that underlie the development and progression of COPD should therefore be investigated.
Increasing evidence indicates that lipid metabolic disorders are associated with COPD onset and progression. It has been reported that the levels of free alpha-linolenic acid, linoleic acid and eicosapentaenoic acid in the sputum of patients with stable COPD are significantly lower than those in the control group.9 The lipid metabolism profile during acute exacerbations of COPD exhibits a unique characteristic.10 It has been demonstrated that phospholipididylcholine 34:3 levels are positively correlated with COPD progression and lung function decline in FEV1.11 In COPD patients with acute exacerbations, a predictive model developed using the three lipid metabolites had excellent discriminatory power in distinguishing eosinophilic and non-eosinophilic inflammatory subtypes, with an area under the receiver operating characteristic (ROC) curve (AUC) of 0.834.12 Smoking and oxidative stress may contribute to the development of lipid-metabolism abnormalities in COPD, with smokers having elevated serum triglyceride concentrations and reduced plasma high-density lipoprotein and high-density lipoprotein cholesterol.13 A high level of high-density lipoprotein cholesterol is associated with an increased risk of COPD mortality.14 Exposure to cigarette smoke causes lipid peroxidation in bronchial epithelial cells and leads to the redistribution of various lipid components, resulting in the accumulation of lipids in the cytoplasm.15 During smoking, the continuous process of lipid metabolism in respiratory tract cells is disrupted.16 It also reduces the fluidity of the plasma membrane of alveolar macrophages, leading to changes in lipid composition.17 In addition, lipids can also act as a second messenger in cell signaling to modulate host inflammatory responses. Lipid molecules play a critical role in regulating inflammation generation and resolution by balancing pro-inflammatory and anti-inflammatory mediators.18
Lipid metabolism-related genes (LMRGs) are essential molecules involved in lipid production and metabolism, playing a vital role in tumors, immune disorders and inflammatory diseases.19,20 A growing body of research has also revealed the importance of LMRGs in COPD. It has been shown that cholesterol overload intensifies inflammation in cigarette smoke-treated bronchial epithelial cells and causes mitochondrial dysfunction mediated by StAR-related lipid transfer domain-3 (STARD3).21 In lung epithelial cells exposed to cigarette smoke, the family with sequence similarity 13 member A (FAM13A) induces the expression of carnitine palmitoyltransferase 1A (CPT1A) to enhance fatty acid oxidation, resulting in an accumulation of reactive oxygen species and cell death.22
In this study, the diagnostic characteristics of LMRGs in COPD were established by analyzing transcriptional profile data from multiple databases. Further cytological validation identified BCHE and PLA2G7 as potential COPD biomarkers associated with lipid metabolism.
Materials and Methods
Data Collection
The expression profiles of datasets GSE76925 and GSE8581 were downloaded from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/). The two datasets contain genome-wide information derived from lung tissue samples. R software (version 3.5.1) was used to preprocess the data, box plots were used to verify sample standardization, PCA plots and UMAP plots were used to check clustering between sample groups, and the “Limma” package was used to identify differentially expressed genes (DEGs). DEGs are defined as gene expression differences greater than twofold between two groups with adjusted p-values less than 0.05. Visualization of the data was accomplished using the ggplot2 package (version 3.3.3) and the ComplexHeatmap package (version 2.2.0).
Subject Characteristics
We included 38 control samples and 111 samples of COPD in GSE76925. According to GOLD criteria, COPD was diagnosed as FEV1/FVC<70%. Compared with healthy controls, FEV1/FVC values were significantly lower in the COPD group, as well as FEV1%pred. There were no statistically significant differences in gender and age between COPD patients and controls. Patients with COPD smoked significantly more pack-years and had a lower BMI compared with healthy controls, which is consistent with the clinical features of the disease. The demographic characteristics of dataset GSE76925 are shown in Table 1. In the validation set GSE8581, 18 control samples and 16 samples with COPD were included, and there was no statistical difference in age or gender between the two groups (Supplementary Table 1).
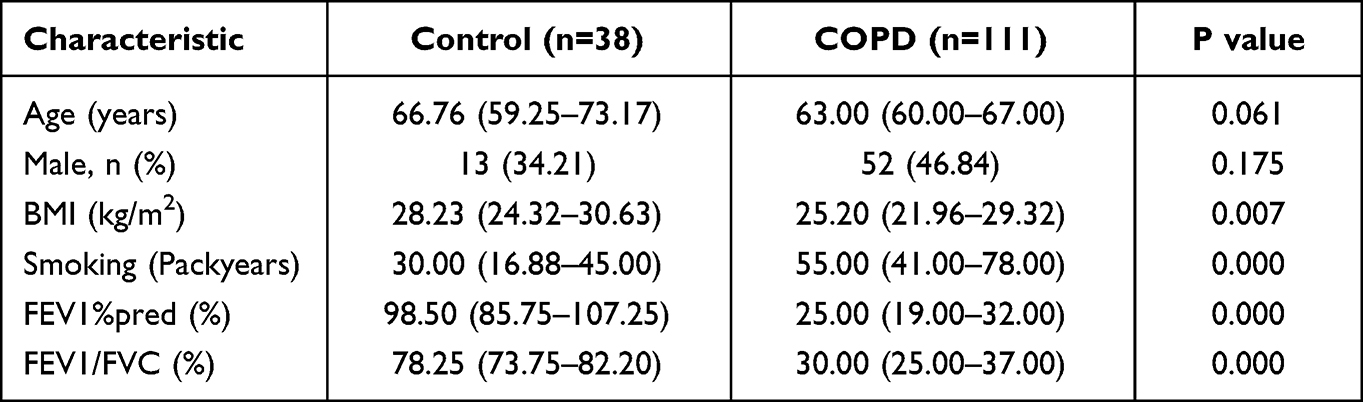 |
Table 1 Demographic and Baseline Characteristics of the Samples in GSE76925 |
Extraction of Genes in Lipid Metabolism
The Kyoto Encyclopedia of Genes and Genomes (KEGG) database (https://www.genome.jp/kegg/) and Molecular Signature Database (MSigDB, version 7.4) (https://www.gsea-msigdb.org/gsea/msigdb) were used to search for lipid metabolism pathways. We summarized major lipid metabolism-related gene sets from the KEGG database (including hsa00061, hsa00062, hsa00071 and other lipid metabolism-related gene sets) and the GSEA database (including Hallmark fatty acid metabolism, KEGG glycerophospholipid metabolism, and lipid raft), and extracted LMRGs from the main lipid metabolism-related gene sets. By removing duplicate, nonsense and ambiguous gene annotations, we identified a total of 1102 LMRGs for further analysis (Supplementary Table 2).
Functional Annotation of Differentially Expressed Genes
Gene ontology (GO) analysis and KEGG enrichment analysis of DEGs were performed using the “clusterProfiler” package in R language. GO enrichment analysis can explain the biophysical properties of the overall gene, mainly including three aspects: cellular components, molecular functions and biological processes. KEGG analysis provides an overview of metabolic pathways as well as functionally related databases of gene expression information in cells. P<0.05 was set as the cutoff criterion. Gene set enrichment analysis (GSEA) is a computational procedure that examines gene expression profiles at the level of entire gene sets to better understand candidate gene sets or pathways associated with disease. The clusterprofiler package was used to perform GSEA analysis, as well as normalized enrichment scores (|NES|>1) and adjusted p-values of 0.05 were set as threshold levels.
Protein-Protein Interaction (PPI) Network Construction
STRING (https://cn.string-db.org/) is an online database that holds information on all known and predicted proteins and can be used to construct functional protein association networks. To obtain the interaction information between differentially expressed lipid metabolism-related genes (DeLMRGs) and proteins, we used the STRING database and assigned a confidence score of 0.4 for the medium confidence score. PPI networks were constructed and visualized using Cytoscape software (version 3.9.1), a general open-source software platform for network biology analysis and visualization. With default parameters, the CytoHubba plug-in was used to identify hub genes with five core algorithms.
Competing Endogenous RNA (ceRNA) Regulatory Network Construction
To predict miRNA targets of selected DeLMRGs, online databases including TargetScan (http://www.targetscan.org/) and miRDB (http://www.mirdb.org/) databases were used. Using the VEEN graph, the targeted miRNAs were obtained by intersecting the miRNAs of the same DeLMRGS in the two databases. Starbase database (https://starbase.sysu.edu.cn/) was selected for predicting miRNA-targeted long non-coding RNAs (lncRNAs), with a screening condition of CLIP data high stringency (clipExpNum≥3). Data visualization was performed using Cytoscape.
Cigarette Smoke Extract (CSE) Preparation
Non-filtered cigarettes (tar: 11 mg, nicotine: 1.0 mg, carbon monoxide: 14 mg/cigarette; Furong Brand; Hunan China Tobacco Industrial Co., Ltd.) were burned, one end of the cigarette was connected to a 50mL centrifuge tube containing 5 mL PBS solution and the other end was connected to a 50 mL syringe. Take three cigarettes each time, and the smoke was directed through the tube and fully dissolved in 5 mL of PBS solution. OD values were detected at 320 nm and adjusted to 12±0.5 as the original CSE (100% concentration). Then the 100% CSE solution was filtered with a 0.22 μm filter (Merck-Millipore, USA) to remove bacteria and was used within 30 mins. Depending on the experimental group, 100% CSE was diluted to the required concentration using culture medium.
Cell Culture and Treatment
The Mouse macrophage cell line RAW264.7 and the Human mononuclear macrophage cell line THP-1 were obtained from the American Type Culture Collection (ATCC, USA). Among them, RAW264.7 cell line was cultured in DMEM medium (Gibco, China) containing 10% fetal bovine serum (Procell, Wuhan, China) and 1% penicillin/streptomycin (Ecotop, Guangzhou, China); THP-1 cell line in RPMI-1640 medium (Gibco, Shanghai, China) containing 10% fetal bovine serum (Procell, Wuhan, China) and 1% penicillin/streptomycin (Ecotop, Guangzhou, China), and cultured at 37°C in an incubator with 5% CO2. THP-1 monocytes were transformed into adherent macrophages in 6-well plates treated with 100 nM phobolol 12-myristate 13-acetate (PMA) final concentration for 48h. THP-1 cells and RAW264.7 cells were seeded to 6-well plates and stimulated with 0–3% (final concentration) CSE for 24h or 48h. Finally, cells were collected to extract proteins and RNA for subsequent analysis.
CCK8 Assay for Cell Viability
A cell viability assay was performed using the cell counting kit 8 (CCK-8; ECOTOP, Guangzhou, China) according to the manufacturer’s instructions. THP-1 cells (3×104 / well) and RAW264.7 cells (2×104 / well) were incubated with different concentrations of CSE in 96-well plates for 24h or 48h. Subsequently, the supernatant was collected, and 100ul DMEM or RPMI-1640 containing 10% CCK-8 solution was added. The cells were further incubated for 60 mins at 37°C. A microplate reader (Bio-TEK; USA) was used to measure the absorbance (Ab) value at 450 nm wavelength. Cell activity (%) = (Ab value of treated well – Ab value of blank well)/(Ab value of control well – Ab value of blank well) × 100 (Supplementary Figure 1).
Enzyme-Linked Immunosorbent Assay (ELISA)
Briefly, THP-1 cells and RAW264.7 cells were seeded in a 6-well culture plate at a concentration of 5×105 per well and treated according to established experimental grouping. The supernatant of each group was collected after 24h or 48h, followed by centrifugation at 1000 × g for 20 mins. PLA2G7 (LP-PLA2) and BCHE expression levels were measured according to the ELISA assay kit instructions (RenJie Biotechnology, Shanghai, China). Mouse (RJ23239, China) and human (RJ14515, China) PLA2G7, and Mouse (RJ17076, China) and human (RJ24172, China) BCHE ELISA kits.
Statistical Analysis
Statistical analysis was performed using R software (version 3.6.2) and SPSS 26.0. Data with a normal distribution were expressed as mean ± standard error of the mean (SEM), and Student’s t-test were used to compare the mean of two groups. Data that did not fit the normal distribution were expressed as interquartile ranges, and Mann–Whitney U-test were applied to compare the two groups. Comparisons between groups were made using the Chi-square test with count data expressed in n (%). Correlations between DeLMRGs expressions were analyzed using Pearson correlation analysis. Pearson’s correlation coefficient is between 0.5 and 1, indicating a strong correlation. The ROC curve analysis was used for determining the predictive value of the hub gene in Validation set for the diagnosis of COPD, with AUC>0.7 indicating a high diagnostic value. P -value < 0.05 was considered statistically significant.
Results
Identification of DEGs and DeLMRGs in COPD
According to the flow chart, the entire study was conducted (Figure 1). For gene expression profiling, 111 severe COPD cases and 38 smoking controls with normal lung function were selected from GSE76925 in the GEO database, all of whom underwent surgical removal for lung tissue sampling. The difference analysis between the two groups was performed and 587 DEGs were found with a 2-fold difference between the two groups and adjusted P values less than 0.05, including 62 genes upregulated and 525 genes downregulated (Figure 2A and B). The intersection of 587 DEGs with 1102 LMRGs led to the identification of 20 DeLMRGs, including 1 up-regulated and 19 down-regulated genes (Figure 2C).
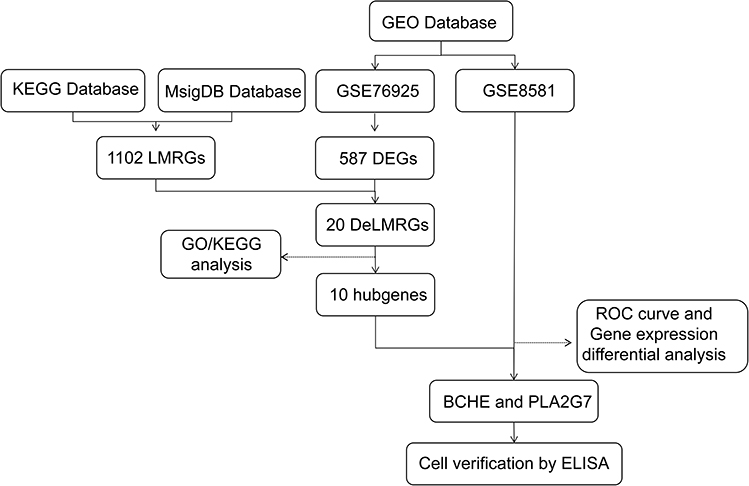 |
Figure 1 A flowchart of the study. Abbreviations: GEO, Gene Expression Omnibus; KEGG, Kyoto Encyclopedia of Genes and Genomes; MsigDB, Molecular Signatures Database; LMRGs, lipid metabolism-related genes; DEGs, differentially expressed genes; DeLMRGs, differentially expressed lipid metabolism-related genes; GO, Gene Ontology; ROC, receiver operating characteristic; BCHE, butyrylcholinesterase; PLA2G7, phospholipase A2 Group VII; ELISA, Enzyme linked immunosorbent assay. |
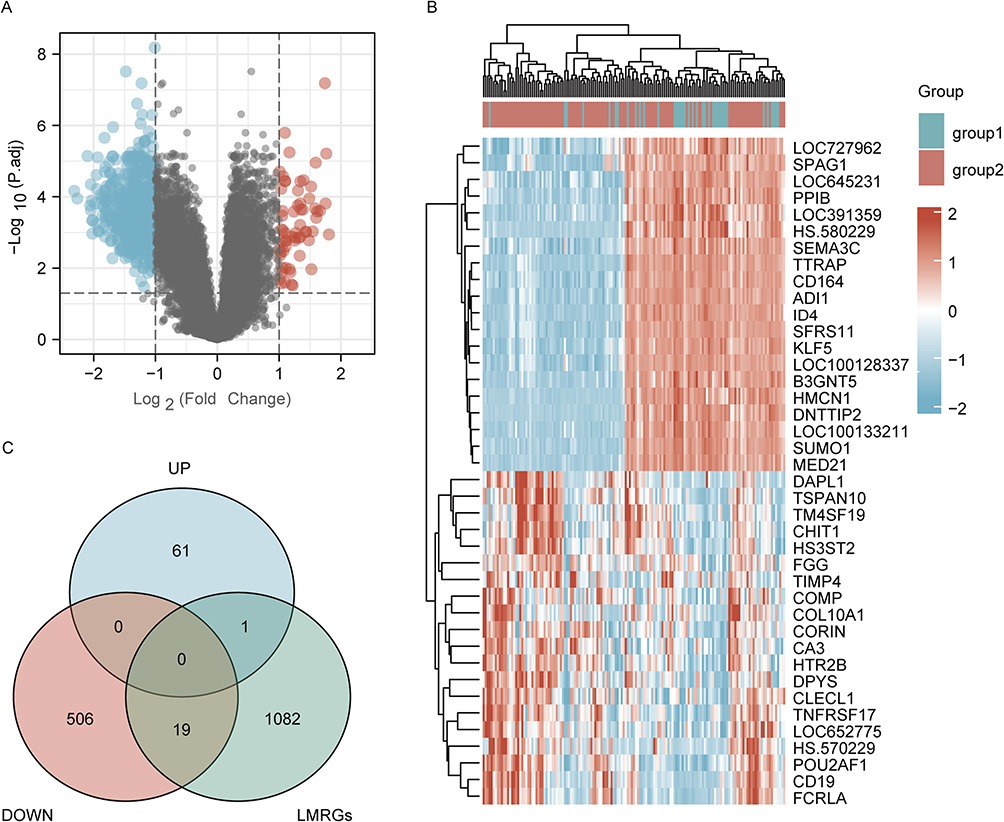 |
Figure 2 Identification of DeLMRGs in COPD patients. (A) Volcano plot of COPD-related DEGs, graphed with log2FoldChange on the abscissa and -log10 on the ordinate (adjusted P value): Nodes in red represent DEGs up-regulated, nodes in blue represent DEGs down-regulated, and nodes in gray represent genes not significantly differentially expressed. (B) Heat map of top 20 COPD-related DEGs: disease samples are indicated by group1, normal control samples by group2, and high gene expression is indicated by red with low gene expression by blue; (C) Venn diagram of DeLMRGs derived from DEGs and LMRGs intersection: green represents LMRGs, blue represents upregulated DEGs and pink represents downregulated DEGs. Abbreviations: DEGs, differentially expressed genes; LMRGs, lipid metabolism-related genes; DeLMRGs, differentially expressed lipid metabolism-related genes. |
Enrichment Analysis of DEGs and DeLMRGs in COPD
To better understand the biological functions and pathways underlying the 587 differential genes associated with COPD, we performed GO and KEGG enrichment analysis (Figure 3A). Biological processes associated with these DEGs include mitotic nuclear division, nuclear division, and sister chromatid segregation. Spindle, mitotic spindle, and centromeric regions are the major cell components associated with these DEGs. The main molecular functions of these DEGs include microtubule attachment, tubulin attachment, and microtubule motor activity. According to the KEGG pathway analysis, the cell cycle, oocyte meiosis, and cellular senescence are the most closely related pathways. Using GSEA analysis, concordant differences between disease and normal states were detected or the distribution trend of DEGs was examined. Results revealed that these DEGs were strongly associated with matrisome, extracellular matrix organization, membrane trafficking and vesicle mediated transport (Figure 3B).
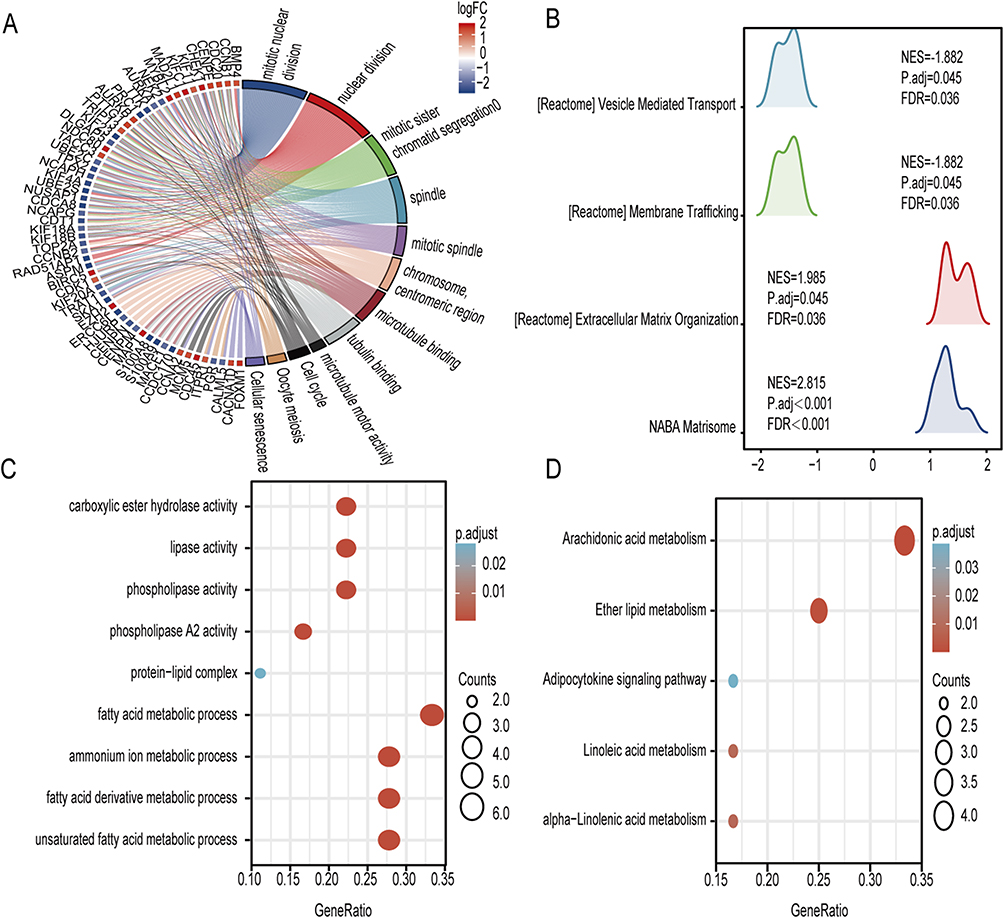 |
Figure 3 Enrichment analysis of DEGs and DeLMRGs in COPD patients. (A) GO and KEGG enrichment analysis of 587 DEGs. (B) KEGG analysis of 587 DEGs. (C) GO enrichment analysis of 20 DeLMRGs. (D) KEGG enrichment analysis of 20 DeLMRGs. Abbreviations: DEGs, differentially expressed genes; DeLMRGs, differentially expressed lipid metabolism-related genes; GO, gene ontology; KEGG, kyoto encyclopedia of genes and genomes. |
Similarly, we performed GO and KEGG enrichment analysis on these screened 20 DeLMRGs. GO analysis revealed that these DeLMRGs were primarily involved in biological processes such as unsaturated fatty acid metabolism, fatty acid derivative metabolism, fatty acid metabolism, ammonium ion metabolism, fatty acid derivative biosynthesis, protein-lipid complex, phospholipase activity, phospholipase A2 activity, lipase activity, carboxylate hydrolase activity and calcium-dependent phospholipase A2 activity (Figure 3C). KEGG pathway analysis showed that these DeLMRGs primarily function in signaling pathways such as arachidonic acid metabolism, ether lipid metabolism, α-linolenic acid metabolism, adipocytokine signaling pathway and linoleic acid metabolism (Figure 3D).
PPI Network Construction and Hubgene Screening
To investigate the relationship between DeLMRGs, these 20 DeLMRGs were imported into Cytoscape to construct a PPI network (Figure 4A) and 10 hub genes were obtained by screening with the cytohubba plug-in (details about the genes are provided in Table 2). Among them, PLA2G7 was the only gene that was up-regulated, followed by PLA2G4A, HPGDS, LEP, PTGES3, LEPR, PLA2G2D, MED21, SPTLC1, and BCHE, which were the nine genes down-regulated (Figure 4B).
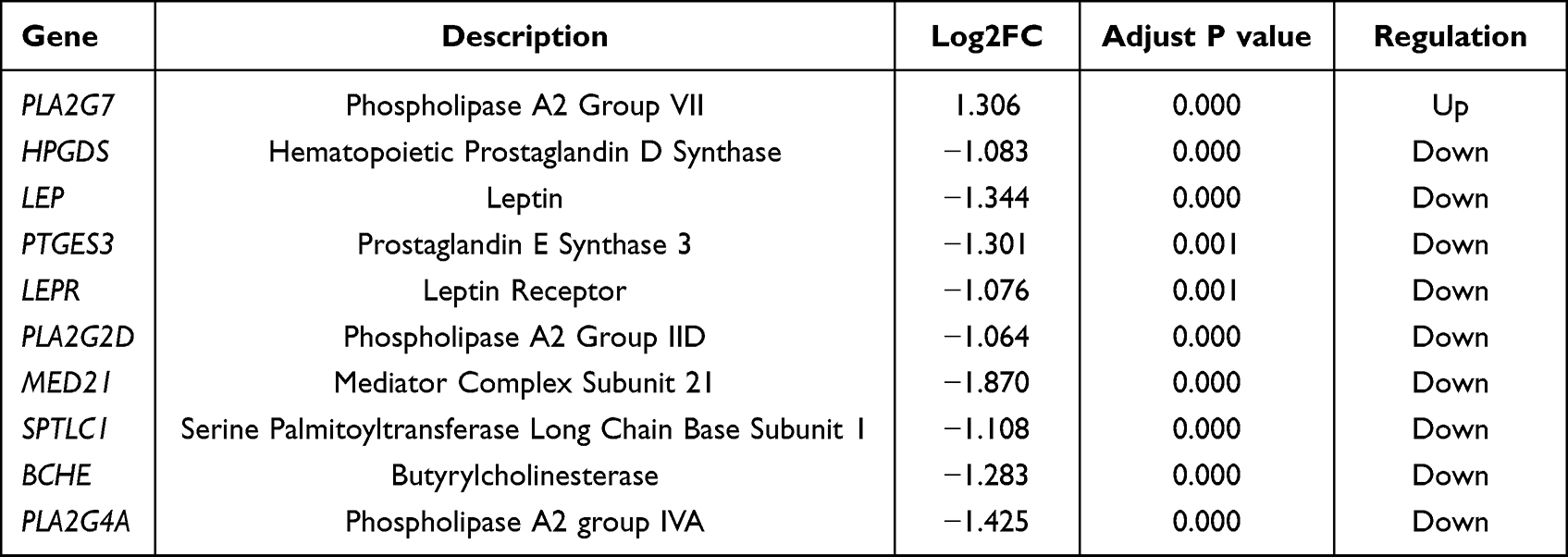 |
Table 2 Table of Detailed Information About 10 Hub DeLMRGs |
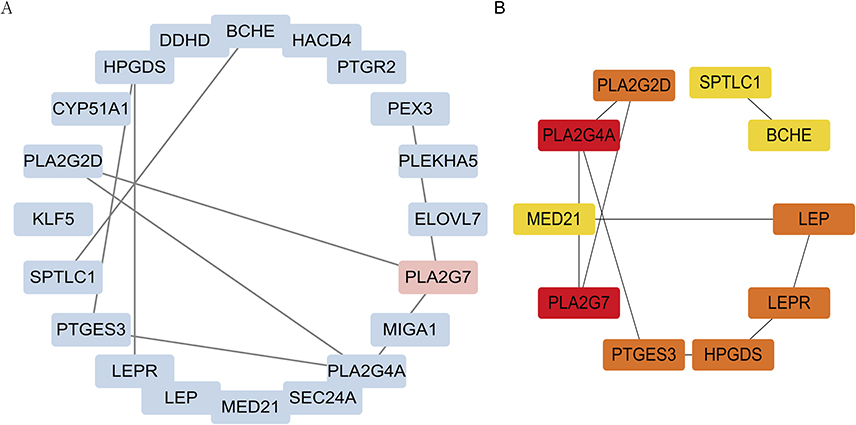 |
Figure 4 PPI network of DeLMRGs. (A) PPI network of 20 DeLMRGs: upregulated genes are shown in red, while downregulated genes are shown in blue. (B) PPI network of 10 hub DeLMRGs: The darker the color, the greater the contribution of genes to the PPI network. The black line indicates the presence of genes that interact. Abbreviations: DeLMRGs, differentially expressed lipid metabolism-related genes; PPI, Protein-Protein Interaction. |
Verification of Diagnostic Biomarkers
Validation of the identified 10 DeLMRGs was carried out using the dataset GSE8581 which includes gene expression profiles of lung tissues from 16 COPD patients and 18 controls with normal lung function. A total of 9 DeLMRGs (except MED21) were found in the validation set (Figure 5A–I), in which BCHE and PLA2G7 expression levels were statistically different between the two groups. In comparison with normal controls, patients with COPD had lower expressions of BCHE (P<0.01, Figure 5A), as well as higher expressions of PLA2G7 (P<0.01, Figure 5G). Meanwhile, we generated the ROC curves for 9 DeLMRGs. A high diagnostic value was found for both BCHE (with an AUC of 0.753) and PLA2G7 (with an AUC of 0.760) in predicting the development of COPD (Supplementary Figure 2).
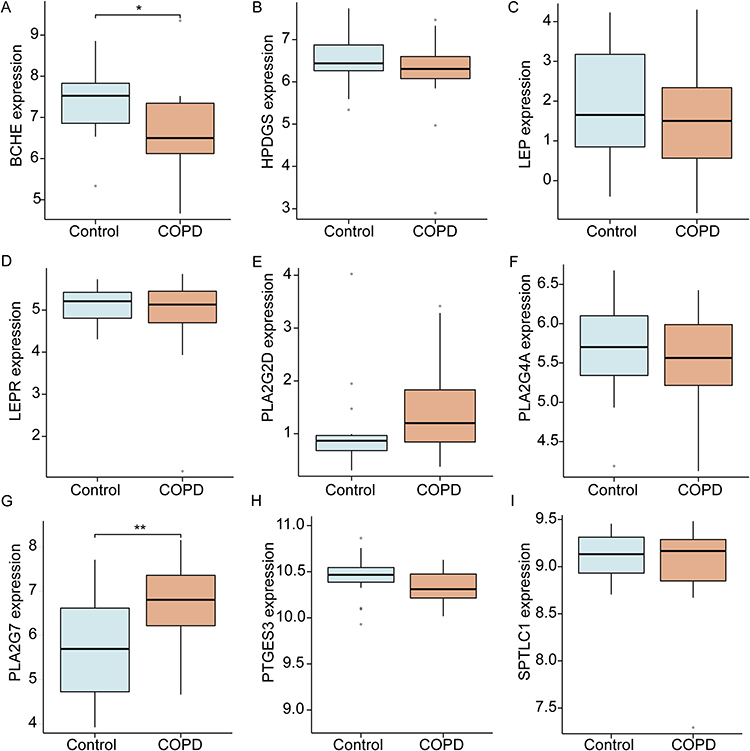 |
Figure 5 Validation of 9 hub DeLMRGs between COPD and controls in GSE8581. (A) The expressions of BCHE in validation sets. (B) The expressions of HPDGS in validation sets. (C) The expressions of LEP in validation sets. (D) The expressions of LEPR in validation sets. (E) The expressions of PLA2G2D in validation sets. (F) The expressions of PLA2G4A in validation sets. (G) The expressions of PLA2G7 in validation sets. (H) The expressions of PTGES3 in validation sets. (I) The expressions of SPTLC1 in validation sets. *P< 0.05, **P < 0.01. Abbreviations: DeLMRGs, differentially expressed lipid metabolism-related genes; BCHE, Butyrylcholinesterase; HPDGS, hematopoietic prostaglandin D synthase; LEP, leptin; LEPR, leptin receptor; PLA2G2D, phospholipase A2 group IID; PLA2G4A, phospholipase A2 group IVA; PLA2G7, phospholipase A2 group VII; PTGES3, prostaglandin E synthase 3; SPTLC1, serine palmitoyltransferase long chain base subunit 1. |
Construction of ceRNA Regulatory Network
CeRNA plays a crucial role in gene expression regulation, as ceRNA molecules (mRNA, lncRNA, pseudogene, etc.) can modulate gene expression by competing with miRNA response elements. To explore possible interactions among lncRNAs, miRNAs, and DeLMRGs in COPD, BCHE and PLA2G7 ceRNA regulatory networks were constructed. We collected 57 miRNAs in this study, including hsa-miR-342-3p, hsa-miR-3145-3p, and hsa-miR-218-5p (Figure 6A and B). The starbase database was used to map the above 57 miRNAs and find their target lncRNAs. A total of 20 lncRNAs were examined for interactions with seven of the 57 miRNAs in the Starbase database. The ceRNA regulatory network was then constructed and visualized by CytoScape (Figure 6C).
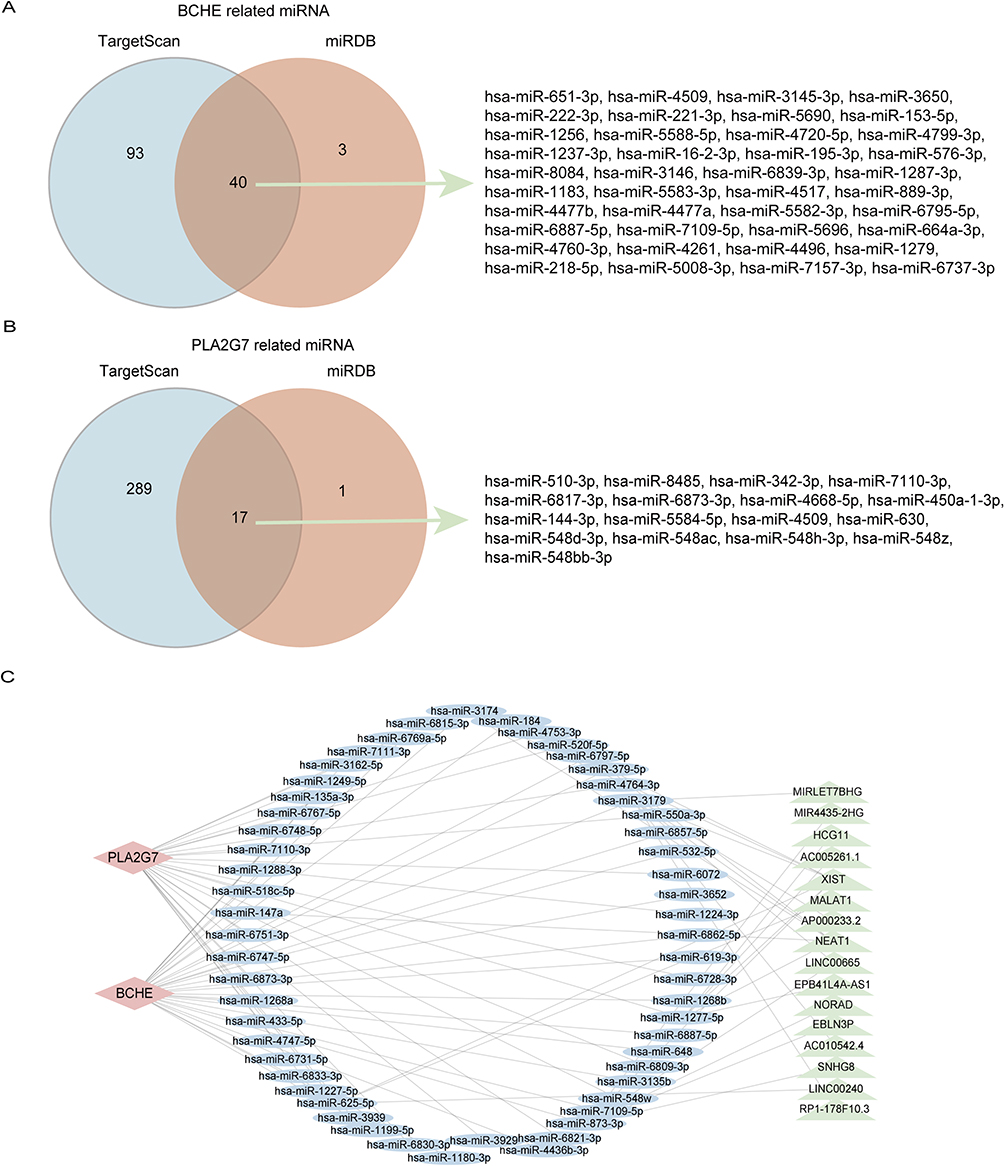 |
Figure 6 CeRNA regulatory network of lncRNA-miRNA-DeLMRG. (A) Venn diagram showing that 40 miRNAs from TargetScan and miRDB interact with BCHE. (B) Venn diagram showing that 17 miRNAs from TargetScan and miRDB interact with PLA2G7. (C) CeRNA network of BCHE and PLA2G7: red diamonds represent protein-coding genes, blue circles represent miRNAs, green rectangles represent lncRNAs, and black lines represent the interactions between lncRNAs, miRNAs, and mRNAs. Abbreviations: BCHE, butyrylcholinesterase; PLA2G7, phospholipase A2 Group VII; ceRNA, competing endogenous RNA; lncRNA, long non-coding RNA; miRNA, microRNA; DeLMRG, differentially expressed lipid metabolism-related genes. |
Verification of BCHE and PLA2G7 Expression in Cell Experiment
To verify the bioinformatics results, we detected the expression of PLA2G7 and BCHE in macrophages treated with different concentrations of CSE. CSE-induced groups showed significant decreases in the expression of BCHE (Figure 7) and increases in the expression of PLA2G7 (Figure 8), indicating the analysis was reliable and PLA2G7 and BCHE may contribute to the development of COPD.
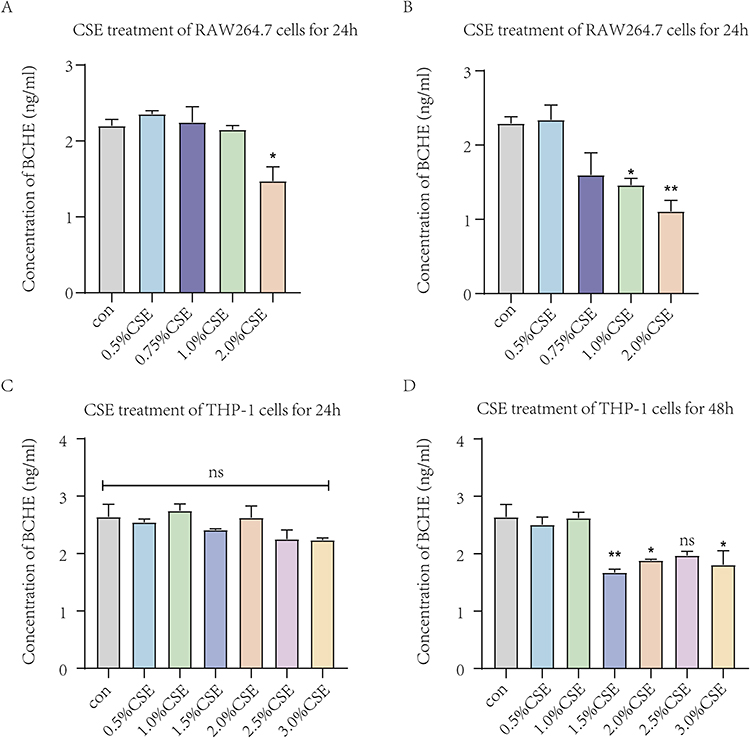 |
Figure 7 The expression levels of BCHE in macrophages treated with different concentrations of CSE by ELISA. (A) Expression levels of BCHE after different concentrations of CSE treated RAW264.7 cells for 24h. (B) Expression levels of BCHE after different concentrations of CSE treated RAW264.7 cells for 48h. (C) Expression levels of BCHE after different concentrations of CSE treated THP-1 cells for 24h. (D) Expression levels of BCHE after different concentrations of CSE treated THP-1 cells for 48h. Data are presented as mean±sem and differences between groups were assessed by one-way ANOVA. *Indicates the comparison between different CSE administration (0.5–3%) with the blank control group, and *p<0.05; **p<0.01; NS: no significant (P > 0.05). Abbreviations: BCHE, butyrylcholinesterase; CSE, cigarette smoke extract. |
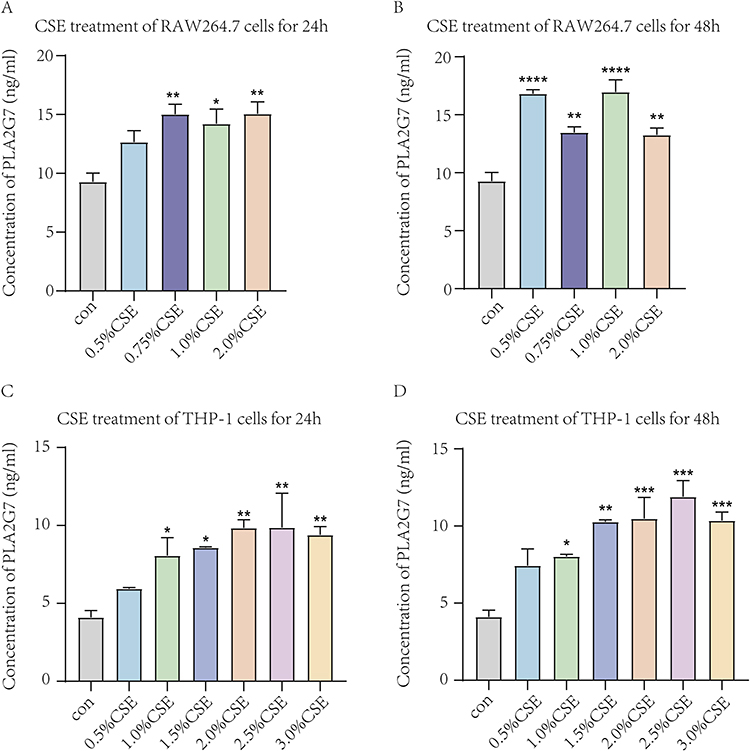 |
Figure 8 The expression levels of PLA2G7 in macrophages treated with different concentrations of CSE by ELISA. (A) Expression levels of PLA2G7 after different concentrations of CSE treated RAW264.7 cells for 24h. (B) Expression levels of PLA2G7 after different concentrations of CSE treated RAW264.7 cells for 48h. (C) Expression levels of PLA2G7 after different concentrations of CSE treated THP-1 cells for 24h. (D) Expression levels of PLA2G7 after different concentrations of CSE treated THP-1 cells for 48h. Data are presented as mean±sem and differences between groups were assessed by one-way ANOVA. *Indicates the comparison between different CSE administration (0.5–3%) with the blank control group, and *p<0.05; **p<0.01; ***p<0.001; ****p < 0.0001. Abbreviations: PLA2G7, phospholipase A2 Group VII; CSE, cigarette smoke extract. |
Discussion
Advances in proteomics and metabolomics have greatly contributed to the understanding of disease. The metabolic alteration of the lungs and even the exhaled breath can be used as indicators of lung disease.23,24 As a common metabolic process, lipid metabolism is not only significant for energy production, but also for biosynthesis, redox homeostasis, and the regulation of intercellular communication.25 Numerous studies have demonstrated that lipid metabolism disorders contribute to the development of diseases, including cancer,26 autoimmune diseases27 and chronic inflammatory conditions.28 Lipid metabolism is active in the lungs, particularly in the alveoli. In the lung, surfactants are typical lipid complexes that regulate the homeostasis of each respiratory cycle.29 Lipids are not only important surfactants and energy storage compounds in the lungs, but they also contribute to the development of diseases such as pulmonary fibrosis and COPD by acting as signaling molecules in a variety of physiological and pathophysiological processes.30,31 As a common chronic inflammatory airway disease, COPD has been found to be associated with lipid metabolic disorders.32 Tobacco smoking is a major risk factor for COPD due to its significant effects on lung cell function, surfactant composition and lung lipid composition (particularly phospholipids, cholesterol, and fatty acids).33
Supported by the close association of altered lipid metabolism with disease, specific lipid profiles are emerging as unique disease biomarkers with diagnostic, prognostic and predictive potential. Our study explored this specific lipid profile between COPD patients and normal ones. The results identified PLA2G4A, HPGDS, LEP, PTGES3, LEPR, PLA2G2D, MED21, SPTLC1, PLA2G7, and BCHE as lipid molecules that strongly associated with COPD development. The hematopoietic prostaglandin D synthase (HPGDS) is a σ-like glutathione transferase that participates in the arachidonic acid metabolic pathway and catalyzes the production of prostaglandin D2, which is an important mediator of inflammation and malignant tumor growth. According to Shao et al34 mutations of HPGDS promote lung cancer cell migration by upregulating the expression of ACSL1 and ACC, important enzymes involved in lipid metabolism. In type 2 diabetic mice, HPGDS was significantly downregulated in wounds, and its deficiency delayed normal wound healing.35 Overexpression of HPGDS in adipose-derived mesenchymal stem cells reduced inflammation and improved wound healing. Leptin (LEP) is a product of the obesity gene, synthesized and secreted by white adipocytes. Besides regulating food intake and body weight, LEP and leptin receptors (LEPRs) also play an integral role in fetal growth, pro-inflammatory immune responses, angiogenesis and lipolysis.36–38 Accordingly, elevated levels of LEP during lung inflammation and the presence of functional LEPR in the lungs suggest that LEP/LEPR play an important role in respiratory immune responses and the development of inflammatory respiratory diseases.39 Among patients with moderate COPD, Broekhuizen et al40 found that leptin levels were detected in induced sputum and were significantly correlated with C-reactive protein and total TNF-α. Hansel et al41 found that genetic variants in the LEPR gene were significantly associated with reduced lung function in COPD patients who smoked, by genotyping 36 single nucleotide polymorphisms in LEPR. Wang et al42 suggested that the mutations of the LEPR gene Gln223Arg site may worsen chronic bronchitis by inhibiting the biological effects of leptin, although not directly affecting LEP levels. The molecular chaperone P23, also known as prostaglandin E synthase 3 (PTGES3), is an important component of the Hsp90 molecular chaperone machinery. Gao et al43 found that PTGES3 was highly expressed in lung, cholangiocarcinoma, and breast invasive carcinoma samples, and overexpression was associated with poor overall survival in lung adenocarcinomas. Adekeye et al44 showed that PTGES3 can be a prognostic marker in breast cancer. Mediator Complex Subunit 21 (MED21) is a subunit of the Arabidopsis Mediator, which can interact with the human RNA polymerase II holoenzyme and participate in the transcriptional regulation of the RNA polymerase II transcriptional gene.45 Tan et al46 have demonstrated that all members of the MED family, including MED21, are highly expressed in hepatocellular carcinoma, and that expression levels correlate with pathological stages. Nikas et al47 found that MED21 may be a diagnostic marker for prostate cancer. Serine palmitoyl transferase (SPT) is the rate-limiting enzyme in the de novo synthesis of sphingolipids (SL) and is essential for embryonic development, physiological homeostasis, and stress response.48 SPT long chain subunits 1 (SPTLC1) is a key subunit in the enzymatic activity of the SPT complex and is closely associated with vascular development and systemic sphingolipid homeostasis.49 Jiang et al50 found that inhibition of SPTLC1 expression in lung microvascular endothelial cells significantly reduced SPT expression and inhibited LPS-induced ceramide production, thereby inducing endothelial barrier dysfunction. Medler et al51 found that inhibition of SPTLC1 expression in lung endothelial cells inhibited TNF-α (plus actinomycin) induced apoptosis. Gorshkova et al52 found that inhibition of SPTLC1 downregulated sphingosine kinase 1 expression and thereby delayed the onset of radiation-induced pulmonary fibrosis. Accordingly, these findings support our conclusion that lipid-related molecules are predictive of COPD development.
The results of the enrichment analysis suggest that the biological processes in which these genes are mainly involved include polyunsaturated fatty acids (PUFA) metabolism, fatty acid derivative metabolism and fatty acid metabolism as the molecular pathways in which these genes are mainly involved. PUFA metabolism plays a crucial role in COPD development. Cigarette smoke may contribute directly to an alteration in epithelial PUFA metabolism, promoting the remodeling of airway epithelium. Significant differences in PUFA metabolism have been observed in COPD patients, with further changes occurring during acute exacerbations.9 Increasing the intake of PUFA may reduce the risk of COPD due to its antioxidant and anti-inflammatory properties.30,53 Rutting et al54 found that supplementation with the ω-6 polyunsaturated fatty acid arachidonic acid resulted in impaired cytokine release from fibroblasts and inhibition of extracellular matrix protein expression in COPD patients. Atlantis et al55 found that supplementation with omega-3 fatty acids (eicosapentaenoic acid, docosahexaenoic acid or alpha-linolenic acid) significantly improved exercise capacity in COPD patients. According to Lee-Sarwar et al56 dietary intake and plasma levels of PUFA were negatively associated with asthma and recurrent wheezing at age three. Many respiratory diseases are associated with the metabolism of fatty acids and their derivatives. The impaired metabolism of fatty acids in cystic fibrosis, characterized by an increase in arachidonic acid-derived metabolites and a decrease in docosahexaenoic acid-derived metabolites, may be partially corrected by supplementation with docosahexaenoic acid.57 Linoleic acid-derived lipid mediators are associated with an increased risk of COPD in women, including the cytochrome P450-derived epoxide product of linoleic acid (leukotoxin) and its corresponding soluble epoxide hydrolase (sEH)-derived product (leukotoxin diol).58 Cigarette smoke may be involved in COPD development by reducing fatty acid catabolism in pulmonary microvascular endothelial cells, leading to apoptosis of pulmonary endothelial cells.59 There is some evidence that exogenous supplementation of fatty acids and their derivatives may have a beneficial effect on lung disease. When short-chain fatty acids are supplemented, inflammation and oxidative stress are reduced in the lungs of aging mice, as well as the increased inflammatory signal in acute lung injury in these mice.60 A fatty acid derivative of quercetin-3-O-glucoside prevents cigarette smoke extract-induced cell death and membrane lipid peroxidation in human fetal lung fibroblasts.61 As a result of these studies, lipids and their derivatives have been implicated as a critical pathogenic factor in COPD and other lung diseases.
The ceRNA network regulatory mechanism also plays a crucial role in the regulation of COPD onset and progression. Liu et al62 showed that overexpression of LncRNA CASC2 is involved in the development of COPD by targeting the miR-18a-5p/IGF1 axis to inhibit bronchial epithelial cell apoptosis and inflammation. Shen et al63 found that LncRNA SNHG5 attenuated the pro-apoptotic and pro-inflammatory effects of cigarette smoke extracts on bronchial epithelial cells via the miR-132/PTEN axis. Comprehensive analysis of COPD-related coding and non-coding RNA transcriptome expression profiles and the construction of competing endogenous RNA networks facilitates visualization of the interactions between these RNAs, providing a more comprehensive insight into the pathology of COPD.64,65 Our study constructs a ceRNA network comprising lipid metabolism-related molecules, which partially serves as a foundation for exploring therapeutic targets associated with COPD-related lipid metabolism.
We identified PLA2G7 and BCHE as lipid metabolism diagnostic molecules in COPD. Lp-PLA2, formerly known as plasma platelet-activating factor acetylhydrolase, is encoded by the PLA2G7 gene located on chromosome 6p12-21. In respiratory diseases, PLA2G7 is secreted by macrophages and circulates in the blood as a complex with lipoproteins.66 Deng et al67 found that Lp-PLA2 levels were upregulated in COPD patients and that significantly associated with FEV1/FVC, CAT scores, mMRC scores and 6-minute walk test in COPD patients. Zhao et al68 found that PLA2G7 promotes the progression of COPD by promoting the expansion of myeloid-derived suppressor cells and inhibiting their function. Woodruff et al69 found increased expression of PLA2G7 in alveolar macrophages of cigarette smoke-treated mice. Butyrylcholinesterase (BCHE), also known as plasma cholinesterase, is synthesized in the liver and secreted into the plasma as an enzyme that catalyzes the hydrolysis of the neurotransmitter acetylcholine to acetate, thus restoring resting cholinergic neurons.70 Halu et al71 suggest that BCHE may be involved in the common pathogenesis of COPD and idiopathic pulmonary fibrosis. Sicinska et al72 observed a significant reduction in plasma BCHE activity in COPD patients and that this reduction was associated with increased lipid peroxidation and reduced total antioxidant capacity. However, Ben et al73 showed that BCHE activity was increased in COPD patients compared to healthy controls, and in COPD smokers, plasma BCHE activity was positively correlated with levels of several protein oxidative damage biomarkers, including total protein carbonyl compounds and advanced oxidative protein products These findings imply a potential role of PLA2G7 and BCHE in the pathogenesis of COPD, however, further investigations are warranted to validate their pathological mechanisms.
By developing a ceRNA network of lipid metabolism molecules, our study provides comprehensive investigation of lipid metabolism-related molecular markers associated with COPD. We aim to establish a foundation for therapeutic strategies targeting lipid metabolism pathways in COPD. However, there are some limitations to our study. Firstly, we were unable to analyze the relationship between these key genes and patient prognosis due to the lack of prognostic information in the public databases. A second factor that may have influenced our experimental results was the lack of nutrition-related data such as obesity level and weight information for these patients.
Conclusions
In summary, through bioinformatics analysis, we have identified two lipid metabolism-related factors closely associated with the occurrence of COPD. PLA2G7 expression was up-regulated while BCHE expression was down-regulated. Our study also involved the lipid metabolism molecules PLA2G7 and BCHE in the construction of a comprehensive ceRNA network. Our findings provide valuable insights into the pathophysiological mechanisms underlying COPD at the level of lipid metabolism, offering potential therapeutic targets and paving new avenues for disease prediction and intervention.
Data Sharing Statement
The underlying data support the results of the present study and can be obtained from the corresponding author upon reasonable request.
Funding
This study was supported by National Key R&D Program of China (2020YFC2008605 and 2020YFC2008600).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Labaki WW, Rosenberg SR. Chronic obstructive pulmonary disease. Ann Intern Med. 2020;173(3):Itc17–Itc32. doi:10.7326/aitc202008040
2. Y LH, Gao TY, Fang W, et al. Global, regional and national burden of chronic obstructive pulmonary disease over a 30-year period: estimates from the 1990 to 2019 Global Burden of Disease Study. Respirology. 2023;28(1):29–36. doi:10.1111/resp.14349
3. Lange P, Ahmed E, Lahmar ZM, et al. Natural history and mechanisms of COPD. Respirology. 2021;26(4):298–321. doi:10.1111/resp.14007
4. Agustí A, Celli BR, Criner GJ, et al. Global initiative for chronic obstructive lung disease 2023 report: GOLD executive summary. Eur Respir J. 2023;61(4):2300239. doi:10.1183/13993003.00239-2023
5. S VS, Hartl S, Barnes N, et al. Blood eosinophil counts in the general population and airways disease: a comprehensive review and meta-analysis. Eur Respir J. 2022;59(1):2004590. doi:10.1183/13993003.04590-2020
6. B VA, Subramoniam M, Mathew L. Detection of COPD and lung cancer with electronic nose using ensemble learning methods. Clin Chim Acta. 2021;523:231–238. doi:10.1016/j.cca.2021.10.005
7. Mouronte-Roibás C, Leiro-Fernández V, Ruano-Raviña A, et al. Predictive value of a series of inflammatory markers in COPD for lung cancer diagnosis: a case-control study. Respir Res. 2019;20(1):198. doi:10.1186/s12931-019-1155-2
8. Binson VA, Subramoniam M, Mathew L. Discrimination of COPD and lung cancer from controls through breath analysis using a self-developed e-nose. J Breath Res. 2021;15(4). doi:10.1088/1752-7163/ac1326
9. Van der Does AM, Heijink M, Mayboroda OA, et al. Dynamic differences in dietary polyunsaturated fatty acid metabolism in sputum of COPD patients and controls. Biochim Biophys Acta. 2019;1864(3):224–233. doi:10.1016/j.bbalip.2018.11.012
10. Liu X, Zhang H, Si Y, et al. High-coverage lipidomics analysis reveals biomarkers for diagnosis of acute exacerbation of chronic obstructive pulmonary disease. J Chromatogr B Analyt Technol Biomed Life Sci. 2022;1201–1202:123278. doi:10.1016/j.jchromb.2022.123278
11. Ben Anes A, Ben Nasr H, Tabka Z, et al. Plasma lipid profiling identifies phosphatidylcholine 34:3 and triglyceride 52:3 as potential markers associated with disease severity and oxidative status in chronic obstructive pulmonary disease. Lung. 2022;200(4):495–503. doi:10.1007/s00408-022-00552-z
12. Wang Y, Chang C, Tian S, et al. Differences in the lipid metabolism profile and clinical characteristics between eosinophilic and non-eosinophilic acute exacerbation of chronic obstructive pulmonary disease. Front Mol Biosci. 2023;10:1204985. doi:10.3389/fmolb.2023.1204985
13. Vicol C, Buculei I, Melinte OE, et al. The lipid profile and biochemical parameters of COPD patients in relation to smoking status. Biomedicines. 2022;10(11):2936. doi:10.3390/biomedicines10112936
14. Mørland JG, Magnus P, Vollset SE, et al. Associations between serum high-density lipoprotein cholesterol levels and cause-specific mortality in a general population of 345 000 men and women aged 20–79 years. Int J Epidemiol. 2023;52(4):1257–1267. doi:10.1093/ije/dyad011
15. Ween MP, White JB, Tran HB, et al. The role of oxidised self-lipids and alveolar macrophage CD1b expression in COPD. Sci Rep. 2021;11(1):4106. doi:10.1038/s41598-021-82481-0
16. Agudelo CW, Kumley BK, Area-Gomez E, et al. Decreased surfactant lipids correlate with lung function in chronic obstructive pulmonary disease (COPD). PLoS One. 2020;15(2):e0228279. doi:10.1371/journal.pone.0228279
17. Hannan SE, Harris JO, Sheridan NP, et al. Cigarette smoke alters plasma membrane fluidity of rat alveolar macrophages. Am Rev Respir Dis. 1989;140(6):1668–1673. doi:10.1164/ajrccm/140.6.1668
18. Leuti A, Fazio D, Fava M, et al. Bioactive lipids, inflammation and chronic diseases. Adv Drug Deliv Rev. 2020;159:133–169. doi:10.1016/j.addr.2020.06.028
19. Liu L, Mo M, Chen X, et al. Targeting inhibition of prognosis-related lipid metabolism genes including CYP19A1 enhances immunotherapeutic response in colon cancer. J Exp Clin Cancer Res. 2023;42(1):85. doi:10.1186/s13046-023-02647-8
20. Luo T, Peng J, Li Q, et al. GDPD5 related to lipid metabolism is a potential prognostic biomarker in neuroblastoma. Int J Mol Sci. 2022;23(22):13740. doi:10.3390/ijms232213740
21. Li L, Liu Y, Liu X, et al. Regulatory roles of external cholesterol in human airway epithelial mitochondrial function through STARD3 signalling. Clin Transl Med. 2022;12(6):e902. doi:10.1002/ctm2.902
22. Jiang Z, Knudsen NH, Wang G, et al. Genetic control of fatty acid β-oxidation in chronic obstructive pulmonary disease. Am J Respir Cell Mol Biol. 2017;56(6):738–748. doi:10.1165/rcmb.2016-0282OC
23. B VA, Subramoniam M, Mathew L. Noninvasive detection of COPD and lung cancer through breath analysis using MOS sensor array based e-nose. Expert Rev Mol Diagn. 2021;21(11):1223–1233. doi:10.1080/14737159.2021.1971079
24. Binson VA, Subramoniam M, Sunny Y, et al. Prediction of pulmonary diseases with electronic nose using SVM and XGBoost. IEEE Sens J. 2021;21(18):20886–20895. doi:10.1109/JSEN.2021.3100390
25. Butler LM, Perone Y, Dehairs J, et al. Lipids and cancer: emerging roles in pathogenesis, diagnosis and therapeutic intervention. Adv Drug Deliv Rev. 2020;159:245–293. doi:10.1016/j.addr.2020.07.013
26. Li H, Feng Z, He ML. Lipid metabolism alteration contributes to and maintains the properties of cancer stem cells. Theranostics. 2020;10(16):7053–7069. doi:10.7150/thno.41388
27. Yi M, Li J, Chen S, et al. Emerging role of lipid metabolism alterations in cancer stem cells. J Exp Clin Cancer Res. 2018;37(1):118. doi:10.1186/s13046-018-0784-5
28. Badmus OO, Hillhouse SA, Anderson CD, et al. Molecular mechanisms of metabolic associated fatty liver disease (MAFLD): functional analysis of lipid metabolism pathways. Clin Sci. 2022;136(18):1347–1366. doi:10.1042/cs20220572
29. Agudelo CW, Samaha G, Garcia-Arcos I. Alveolar lipids in pulmonary disease. A review. Lipids Health Dis. 2020;19(1):122. doi:10.1186/s12944-020-01278-8
30. Kotlyarov S, Kotlyarova A. Anti-inflammatory function of fatty acids and involvement of their metabolites in the resolution of inflammation in chronic obstructive pulmonary disease. Int J Mol Sci. 2021;22(23):12803. doi:10.3390/ijms222312803
31. Suryadevara V, Ramchandran R, Kamp DW, et al. Lipid mediators regulate pulmonary fibrosis: potential mechanisms and signaling pathways. Int J Mol Sci. 2020;21(12):4257. doi:10.3390/ijms21124257
32. Chen H, Li Z, Dong L, et al. Lipid metabolism in chronic obstructive pulmonary disease. Int J Chron Obstruct Pulmon Dis. 2019;14:1009–1018. doi:10.2147/copd.S196210
33. Lugg ST, Scott A, Parekh D, et al. Cigarette smoke exposure and alveolar macrophages: mechanisms for lung disease. Thorax. 2022;77(1):94–101. doi:10.1136/thoraxjnl-2020-216296
34. Shao F, Mao H, Luo T, et al. HPGDS is a novel prognostic marker associated with lipid metabolism and aggressiveness in lung adenocarcinoma. Front Oncol. 2022;12:894485. doi:10.3389/fonc.2022.894485
35. Ouyang L, Qiu D, Fu X, et al. Overexpressing HPGDS in adipose-derived mesenchymal stem cells reduces inflammatory state and improves wound healing in type 2 diabetic mice. Stem Cell Res Ther. 2022;13(1):395. doi:10.1186/s13287-022-03082-w
36. Obradovic M, Sudar-Milovanovic E, Soskic S, et al. Leptin and obesity: role and clinical implication. Front Endocrinol. 2021;12:585887. doi:10.3389/fendo.2021.585887
37. Mei L, Li M, Zhang T. MicroRNA miR-874-3p inhibits osteoporosis by targeting leptin (LEP). Bioengineered. 2021;12(2):11756–11767. doi:10.1080/21655979.2021.2009618
38. Zhang KW, Jia Y, Li YY, et al. LEP and LEPR are possibly a double-edged sword for wound healing. J Cell Physiol. 2023;238(2):355–365. doi:10.1002/jcp.30936
39. Vernooy JH, Ubags ND, Brusselle GG, et al. Leptin as regulator of pulmonary immune responses: involvement in respiratory diseases. Pulm Pharmacol Ther. 2013;26(4):464–472. doi:10.1016/j.pupt.2013.03.016
40. Broekhuizen R, Vernooy JH, Schols AM, et al. Leptin as local inflammatory marker in COPD. Respir Med. 2005;99(1):70–74. doi:10.1016/j.rmed.2004.03.029
41. Hansel NN, Gao L, Rafaels NM, et al. Leptin receptor polymorphisms and lung function decline in COPD. Eur Respir J. 2009;34(1):103–110. doi:10.1183/09031936.00120408
42. Wang B, Fu E, Cao Y, et al. Effect of leptin receptor mutation on the development of chronic bronchitis. Asia Pac J Public Health. 2013;25(4 Suppl):80s–87s. doi:10.1177/1010539513497218
43. Gao P, Zou K, Xiao L, et al. High expression of PTGES3 is an independent predictive poor prognostic biomarker and correlates with immune infiltrates in lung adenocarcinoma. Int Immunopharmacol. 2022;110:108954. doi:10.1016/j.intimp.2022.108954
44. Adekeye A, Agarwal D, Nayak A, et al. PTGES3 is a putative prognostic marker in breast cancer. J Surg Res. 2022;271:154–162. doi:10.1016/j.jss.2021.08.033
45. Sato S, Tomomori-Sato C, Tsai KL, et al. Role for the MED21-MED7 hinge in assembly of the mediator-RNA polymerase II holoenzyme. J Biol Chem. 2016;291(52):26886–26898. doi:10.1074/jbc.M116.756098
46. Tan W, Peng S, Li Z, et al. Identification of therapeutic targets and prognostic biomarkers among genes from the mediator complex family in the hepatocellular carcinoma tumour-immune microenvironment. Comput Math Methods Med. 2022;2022:2021613. doi:10.1155/2022/2021613
47. Nikas JB, Mitanis NT, Nikas EG. Whole exome and transcriptome RNA-sequencing model for the diagnosis of prostate cancer. ACS Omega. 2020;5(1):481–486. doi:10.1021/acsomega.9b02995
48. Lowther J, Naismith JH, Dunn TM, et al. Structural, mechanistic and regulatory studies of serine palmitoyltransferase. Biochem Soc Trans. 2012;40(3):547–554. doi:10.1042/bst20110769
49. Kuo A, Checa A, Niaudet C, et al. Murine endothelial serine palmitoyltransferase 1 (SPTLC1) is required for vascular development and systemic sphingolipid homeostasis. Elife. 2022;11:e78861. doi:10.7554/eLife.78861
50. Jiang J, Ouyang H, Zhou Q, et al. LPS induces pulmonary microvascular endothelial cell barrier dysfunction by upregulating ceramide production. Cell Signal. 2022;92:110250. doi:10.1016/j.cellsig.2022.110250
51. Medler TR, Petrusca DN, Lee PJ, et al. Apoptotic sphingolipid signaling by ceramides in lung endothelial cells. Am J Respir Cell Mol Biol. 2008;38(6):639–646. doi:10.1165/rcmb.2007-0274OC
52. Gorshkova I, Zhou T, Mathew B, et al. Inhibition of serine palmitoyltransferase delays the onset of radiation-induced pulmonary fibrosis through the negative regulation of sphingosine kinase-1 expression. J Lipid Res. 2012;53(8):1553–1568. doi:10.1194/jlr.M026039
53. Pizzini A, Lunger L, Sonnweber T, et al. The role of omega-3 fatty acids in the setting of coronary artery disease and COPD: a review. Nutrients. 2018;10(12):1864. doi:10.3390/nu10121864
54. Rutting S, Papanicolaou M, Xenaki D, et al. Dietary ω-6 polyunsaturated fatty acid arachidonic acid increases inflammation, but inhibits ECM protein expression in COPD. Respir Res. 2018;19(1):211. doi:10.1186/s12931-018-0919-4
55. Atlantis E, Cochrane B. The association of dietary intake and supplementation of specific polyunsaturated fatty acids with inflammation and functional capacity in chronic obstructive pulmonary disease: a systematic review. Int J Evid Based Healthc. 2016;14(2):53–63. doi:10.1097/xeb.0000000000000056
56. Lee-Sarwar K, Kelly RS, Lasky-Su J, et al. Dietary and plasma polyunsaturated fatty acids are inversely associated with asthma and atopy in early childhood. J Allergy Clin Immunol Pract. 2019;7(2):529–538.e528. doi:10.1016/j.jaip.2018.07.039
57. Teopompi E, Risé P, Pisi R, et al. Arachidonic acid and docosahexaenoic acid metabolites in the airways of adults with cystic fibrosis: effect of docosahexaenoic acid supplementation. Front Pharmacol. 2019;10:938. doi:10.3389/fphar.2019.00938
58. Balgoma D, Yang M, Sjödin M, et al. Linoleic acid-derived lipid mediators increase in a female-dominated subphenotype of COPD. Eur Respir J. 2016;47(6):1645–1656. doi:10.1183/13993003.01080-2015
59. Gong J, Zhao H, Liu T, et al. Cigarette smoke reduces fatty acid catabolism, leading to apoptosis in lung endothelial cells: implication for pathogenesis of COPD. Front Pharmacol. 2019;10:941. doi:10.3389/fphar.2019.00941
60. Hildebrand CB, Lichatz R, Pich A, et al. Short-chain fatty acids improve inflamm-aging and acute lung injury in old mice. Am J Physiol Lung Cell Mol Physiol. 2023;324(4):L480–L492. doi:10.1152/ajplung.00296.2022
61. Warnakulasuriya SN, Rupasinghe H. Novel long chain fatty acid derivatives of quercetin-3-O-glucoside reduce cytotoxicity induced by cigarette smoke toxicants in human fetal lung fibroblasts. Eur J Pharmacol. 2016;781:128–138. doi:10.1016/j.ejphar.2016.04.011
62. Liu P, Zhang H, Zeng H, et al. LncRNA CASC2 is involved in the development of chronic obstructive pulmonary disease via targeting miR-18a-5p/IGF1 axis. Ther Adv Respir Dis. 2021;15:17534666211028072. doi:10.1177/17534666211028072
63. Shen Q, Zheng J, Wang X, et al. LncRNA SNHG5 regulates cell apoptosis and inflammation by miR-132/PTEN axis in COPD. Biomed Pharmacother. 2020;126:110016. doi:10.1016/j.biopha.2020.110016
64. Liu P, Wang Y, Zhang N, et al. Comprehensive identification of RNA transcripts and construction of RNA network in chronic obstructive pulmonary disease. Respir Res. 2022;23(1):154. doi:10.1186/s12931-022-02069-8
65. Feng X, Dong H, Li B, et al. Integrative analysis of the expression profiles of whole coding and non-coding RNA transcriptomes and construction of the competing endogenous RNA networks for chronic obstructive pulmonary disease. Front Genet. 2023;14:1050783. doi:10.3389/fgene.2023.1050783
66. Dua P, Mishra A, Reeta KH. Lp-PLA2 as a biomarker and its possible associations with SARS-CoV-2 infection. Biomark Med. 2022;16(10):821–832. doi:10.2217/bmm-2021-1129
67. Deng M, Yin Y, Zhang Q, et al. Identification of inflammation-related biomarker Lp-PLA2 for patients with COPD by comprehensive analysis. Front Immunol. 2021;12:670971. doi:10.3389/fimmu.2021.670971
68. Zhao X, Yue Y, Wang X, et al. Bioinformatics analysis of PLA2G7 as an immune-related biomarker in COPD by promoting expansion and suppressive functions of MDSCs. Int Immunopharmacol. 2023;120:110399. doi:10.1016/j.intimp.2023.110399
69. Woodruff PG, Ellwanger A, Solon M, et al. Alveolar macrophage recruitment and activation by chronic second hand smoke exposure in mice. COPD. 2009;6(2):86–94. doi:10.1080/15412550902751738
70. Gok M, Cicek C, Sari S, et al. Novel activity of human BChE: lipid hydrolysis. Biochimie. 2023;204:127–135. doi:10.1016/j.biochi.2022.09.008
71. Halu A, Liu S, Baek SH, et al. Exploring the cross-phenotype network region of disease modules reveals concordant and discordant pathways between chronic obstructive pulmonary disease and idiopathic pulmonary fibrosis. Hum Mol Genet. 2019;28(14):2352–2364. doi:10.1093/hmg/ddz069
72. Sicinska P, Bukowska B, Pajak A, et al. Decreased activity of butyrylcholinesterase in blood plasma of patients with chronic obstructive pulmonary disease. Arch Med Sci. 2017;13(3):645–651. doi:10.5114/aoms.2016.60760
73. Ben Anes A, Ben Nasr H, Garrouch A, et al. Alterations in acetylcholinesterase and butyrylcholinesterase activities in chronic obstructive pulmonary disease: relationships with oxidative and inflammatory markers. Mol Cell Biochem. 2018;445(1–2):1–11. doi:10.1007/s11010-017-3246-z
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.