Back to Journals » International Journal of General Medicine » Volume 18
Integrated Proteomic and Metabolomic Profiling in Acute Promyelocytic Leukemia: Current Status and Perspectives
Authors Liu T, Qu M, Wang N, Xing X ![]()
Received 14 August 2025
Accepted for publication 14 December 2025
Published 25 December 2025 Volume 2025:18 Pages 7823—7838
DOI https://doi.org/10.2147/IJGM.S560730
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Dana Kristjansson
Ting Liu,1,* Mengqi Qu,2,* Ning Wang,1 Xue Xing1
1Department of Clinical Laboratory, The Second Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, 116021, People’s Republic of China; 2Department of Clinical Laboratory, The First Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, 116021, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xue Xing, Department of Clinical Laboratory, The Second Affiliated Hospital of Dalian Medical University, No. 467 Zhongshan Road, Dalian, Liaoning, 116021, People’s Republic of China, Email [email protected] Ning Wang, Department of Clinical Laboratory, The Second Affiliated Hospital of Dalian Medical University, No. 467 Zhongshan Road, Dalian, Liaoning, 116021, People’s Republic of China, Email [email protected]
Abstract: Acute promyelocytic leukemia(APL), a distinct subtype of acute myeloid leukemia(AML), has garnered significant attention in recent years regarding its pathogenesis and the molecular basis of its treatment response. With the rapid advancement of proteomics and metabolomics technologies, researchers can now delve deeper into revealing the molecular characteristics of acute promyelocytic leukemia and the regulatory role of its microenvironment. This review summarizes the latest research progress in proteomics and metabolomics within the acute promyelocytic leukemia field, focusing on analyzing the critical role of the the promyelocytic leukemia-retinoic acid receptor alpha (PML::RARα) fusion gene fusion protein in regulating cellular metabolism and protein expression. Furthermore, the article explores the importance of the immune system in acute promyelocytic leukemia treatment response and the impact of all-trans retinoic acid/arsenic trioxide therapy on the proteome and metabolome. By synthesizing existing research findings, this review aims to discuss how proteomic and metabolomic data elucidate the pathological mechanisms and therapeutic targets of acute promyelocytic leukemia, providing a theoretical basis for future precision medicine and translational research.
Keywords: acute promyelocytic leukemia, proteomics, metabolomics, pml::rarα fusion protein, immunoregulation, metabolites, treatment response
Introduction
Acute promyelocytic leukemia (APL) is a distinct subtype of acute myeloid leukemia defined by the presence of the PML::RARα fusion gene and characterized by unique clinical and molecular features.1 Although APL is relatively rare, it still accounts for a certain proportion of acute leukemia cases. According to the latest data from 2025, its annual incidence rate is 0.22 (95% CI: 0.21–0.23) per 100,000 people.2 The pathogenesis of this disease differs from that of conventional acute myeloid leukemia(AML), particularly in clinical manifestations and treatment response. The standard treatment regimen for APL includes all-trans retinoic acid (ATRA) and arsenic trioxide (ATO), which have significantly improved patient prognosis. However, their precise mechanisms and microenvironmental changes remain incompletely elucidated.3
Traditional cytogenetic and molecular biology studies have laid the macroscopic framework for understanding the pathogenesis of APL. However, the precise mechanisms by which the PML::RARα fusion protein remodels intracellular biological networks to drive the initiation, progression, and potential reversal of leukemia remain to be fully elucidated. In recent years, the rapid advancement of high-throughput technologies has given rise to proteomics and metabolomics, offering unprecedented insights into these processes. Proteomics enables the systematic analysis of protein expression, post-translational modifications, protein-protein interactions, and functional alterations under pathological conditions, thereby directly uncovering the executors of cellular activities. Metabolomics, on the other hand, captures the global profiles of endogenous small-molecule metabolites, providing a sensitive reflection of the cellular physiological and pathological states and their dynamic changes, and serving as a critical link between genotype and phenotype.
Despite significant progress in the treatment of APL, there remains a need to explore novel therapeutic strategies, particularly to overcome challenges related to drug resistance and disease relapse. The integration of proteomics and metabolomics provides powerful research tools4–7 that not only reveal the molecular basis of APL and alterations in the tumor microenvironment but also offer valuable references for future research and clinical applications.
Therefore, this review focuses on the cutting-edge applications of proteomics and metabolomics in APL research. We systematically summarize recent advances in the use of these omics technologies to investigate the downstream signaling networks of PML::RARα, mechanisms of drug resistance, discovery of disease biomarkers, and monitoring of treatment responses. By integrating multi-omics data, we can achieve a more comprehensive understanding of the interplay between aberrant signaling pathways and metabolic reprogramming in APL cells, and identify key molecular targets. This review aims to deepen the understanding of the molecular essence of APL and to discuss how these omics approaches may promote precise stratification, personalized treatment, and prognostic assessment, ultimately contributing to more effective and comprehensive therapeutic strategies for APL patients.
Literature Search Strategy
A systematic literature search was conducted to identify relevant publications on proteomics, metabolomics, and immunoregulation in APL. The primary electronic databases utilized were PubMed and Web of Science (WoS) Core Collection. The search was limited to articles published from the inception of each database up to November 30, 2025.
The search strategy was built around a combination of the following key concepts and their related terms:
Acute Promyelocytic Leukemia (Disease); Omics Technologies (Proteomics and Metabolomics); Molecular Driver (PML::RARα). Specific keywords and Medical Subject Headings (MeSH) terms were employed, including but not limited to: “acute promyelocytic leukemia”, “APL”, “proteomics”, “metabolomics”, “metabonomics”, “PML/RARA fusion gene”, “PML::RARα”, “immunoregulation”, “treatment response”, and “metabolites”. The Boolean operators “AND” and “OR” were used to logically combine these terms.
Results
Molecular Pathogenesis of Acute Promyelocytic Leukemia
APL is a specific type of acute myeloid leukemia whose pathogenesis is primarily associated with the formation of the PML::RARα fusion gene and its subsequent effects. This gene results from a translocation between chromosomes 15 and 17 (t(15;17)). It is well established that this genetic rearrangement not only fuses the PML and RARA genes into a functionally abnormal transcription factor but also leads to differentiation block and abnormal proliferation of myeloid cells.8 The expression of the PML::RARα fusion protein inhibits normal myeloid cell differentiation, promoting the abnormal accumulation of promyelocytes and thereby triggering the onset of APL.
At the molecular level, the pathogenesis of APL also involves the abnormal expression of multiple regulatory factors. For instance, studies have found that the overexpression of miR-125b in APL patients is closely related to disease progression; this microRNA accelerates PML::RARα-induced leukemogenesis, revealing its potential as a therapeutic target in APL.9 Furthermore, the activation of cellular signaling pathways in APL is closely linked to the regulation of cell proliferation, apoptosis, and differentiation. The abnormal production of cytokines, such as interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α), may further exacerbate disease progression and the occurrence of complications.10
Clinically, common symptoms in APL patients include bleeding tendencies and coagulation abnormalities, closely associated with coagulation factors released by tumor cells.10 These abnormalities are not only directly caused by the PML::RARα fusion protein but may also be related to alterations in the extracellular matrix and microenvironment. Studies indicate that elevated pro-inflammatory cytokines in the microenvironment of APL cells may alter the bone marrow microenvironment, further impacting normal hematopoietic function.11
While numerous molecular alterations are reported in APL, their functional necessity in the human disease context often remains speculative, requiring validation in physiologically relevant models. Variant forms like microgranular APL (M3v) often present with distinct clinical features, suggesting underlying biological differences, though the precise molecular determinants are not fully defined.12 Further research suggests that genomic analysis may reveal other gene mutations or deletions in APL patients, which could affect their response to standard therapy, indicating the need for individualized treatment strategies in clinical management.1
In summary, the molecular pathogenesis of acute promyelocytic leukemia is a complex network involving gene rearrangements, regulation of cellular signaling pathways, and microenvironmental changes, among other aspects. With in-depth research into the molecular mechanisms of APL, we are poised to develop more effective targeted therapeutic strategies to improve patient survival rates and quality of life.
Structure and Function of the PML::RARα Fusion Protein
The PML::RARα fusion protein is a central driver in the pathogenesis of APL. It disrupts normal myeloid differentiation, leading to the accumulation and aberrant proliferation of leukemic cells, thereby posing a major obstacle to anticancer therapy.10 A key finding from integrated analyses is that PML::RARα’s initial impact on the transcriptome and epigenome is surprisingly subtle, primarily involving the downregulation of secondary and tertiary granule genes—an effect hypothesized to represent the initial block in myeloid maturation. Furthermore, gene set enrichment analysis has shown that PML::RARα significantly enriches cell cycle-related genes in early promyelocytes, underscoring its subtle yet pivotal role in determining cell fate.13
The influence of PML::RARα on transcriptional regulation is mediated through its binding to specific DNA sequences, thereby modulating target gene expression. In mouse models, most genomic regions bound by PML::RARα are already chromatin-accessible in unmodified wild-type promyelocytes.14 This suggests that the oncoprotein does not primarily pioneer new open chromatin sites but rather hijacks pre-existing regulatory landscapes. It has been robustly demonstrated that PML::RARα binding enhances interactions with transcription factors like GATA2, activating transcriptional programs that promote leukemic self-renewal.
Currently, therapeutic strategies targeting PML::RARα—such as the combination of ATRA and ATO—have proven effective in inducing differentiation of leukemic cells and facilitating clinical remission. These findings not only provide novel insights for the treatment of APL but also offer valuable perspectives for research into other subtypes of acute myeloid leukemia.15,16
Regulation of Protein Expression Profiles by the Fusion Protein
Advances in proteomic technologies have enabled the systematic identification and quantification of differentially expressed proteins in patients with APL. For example, mass spectrometry-based approaches have revealed numerous proteins whose expression is altered in association with the PML::RARα fusion oncoprotein. Subsequent bioinformatic analyses indicate that these proteins are predominantly involved in cell cycle regulation, apoptosis, and metabolic processes.
PML::RARα expression drives the upregulation of certain cell cycle-related genes while suppressing pro-apoptotic genes. This dysregulation of transcriptional control represents a key component of APL pathogenesis.13
Maria et al demonstrated that the PML::RARα fusion protein suppresses the expression of heat shock protein 90 (HSP90), thereby affecting the stability of AKT protein in APL cells. Transcript levels of HSP90AA1 and HSP90AB1, as well as HSP90 protein expression, were significantly lower in APL patients compared to those with other AML subtypes. This suppression was associated with dysregulated AKT phosphorylation, indicating that HSP90 plays a critical role in APL cell survival and function. This presents a clear mechanistic link between the fusion oncoprotein and a crucial signaling pathway.17
Furthermore, the PML::RARα fusion protein interacts with multiple intracellular signaling pathways—particularly those involving adenylate cyclase (cAMP-producing enzyme) and the RTK/RAS axis—thereby complicating the cellular metabolic environment.
In studies using the PR9 cell line stably transfected with PML::RARα as an in vitro model, cAMP alone was found to slightly increase CD11b expression but failed to induce full cellular differentiation. Only when cAMP was combined with As2O3 treatment did the cells exhibit terminal differentiation morphology and significantly upregulated CD11b expression. Moreover, PML::RARα strongly suppressed the transcription of reporter genes containing cAMP response elements, indicating that the fusion protein substantially blocks cAMP signaling during myeloid differentiation.18
Another study revealed that in both murine and human hematopoietic cells, the PML::RARα fusion protein upregulates Gab2/GAB2 mRNA expression by modulating chromatin accessibility. GAB2 overexpression enhanced cellular signaling via increased phosphorylation of AKT and ERK1/2, whereas inactivation of Gab2 effectively suppressed the growth of APL cells in mice. These findings not only elucidate the functional interplay between PML::RARα and GAB2 within the RTK/RAS signaling network but also highlight GAB2 as a potential therapeutic target, offering new avenues for future APL treatment strategies.19 While these functional data are compelling, their translation to primary human APL cells requires further validation.
Impact of the Fusion Protein on Metabolic Pathways
The expression of the PML::RARα fusion protein in cells plays a critical regulatory role in pathways related to energy metabolism and substance metabolism. By influencing intracellular signaling pathways, it alters cellular bioenergetic metabolism, thereby impacting disease treatment.
Studies have revealed that NRF2 (NF-E2 p45-related factor 2) is pivotal in cellular responses to oxidative stress and metabolic regulation. Under conditions such as oxidative stress, inflammation, and growth factor stimulation, NRF2 activity is significantly enhanced. However, in APL cells, the presence of PML::RARα suppresses NRF2 function, impeding its translocation into the nucleus and promoting its degradation in the cytoplasm. This suppression alters the metabolic profile of APL cells and increases their sensitivity to high-dose ascorbate, suggesting a therapeutic vulnerability.20
Furthermore, RA-induced cell cycle arrest is closely associated with the degradation of PML::RARα and the activation of Trp53. Following high-dose RA treatment, PML::RARα-expressing APL cells exhibit characteristic senescence features, including cell cycle arrest, activation of Trp53 target genes, and alterations in nuclear morphology. Moreover, the combination of arsenic trioxide with all-trans retinoic acid (RA) demonstrates a notable synergistic effect in APL treatment. Arsenic trioxide not only promotes PML::RARα degradation through non-overlapping pathways but also directly targets normal PML protein, further enhancing the nuclear body remodeling mediated by PML::RARα.21 However, the precise metabolic consequences of this synergistic degradation remain an area of active research. The field is moving from observing metabolic changes to defining their direct mechanistic links to the oncoprotein.
An integrated overview of these PML::RARα-driven mechanisms is depicted in Figure 1.
 |
Figure 1 Schematic model of PML::RARα-mediated pathogenesis in APL. The schematic illustrates the central role of the PML::RARα fusion protein, resulting from the t(15;17) translocation, in reprogramming signaling transduction, metabolic pathways, and immune responses to drive APL pathogenesis. Signaling Reprogramming: PML::RARα transcriptionally upregulates GAB2, enhancing interactions within the RTK/RAS pathway (eg, with GRB2 and PI3K) to activate AKT and ERK1/2 phosphorylation, promoting cell survival and proliferation. Concurrently, it suppresses HSP90 expression, destabilizing AKT, and inhibits cAMP signaling and CRE-driven transcription, contributing to differentiation block. Metabolic Reprogramming: PML::RARα induces AKT degradation, suppressing glycolysis while enhancing the TCA cycle and fatty acid oxidation. It promotes abnormal lipid metabolism by interacting with TRIB3 to inhibit PPARγ activity. Furthermore, it impairs the oxidative stress response by binding to and suppressing the transcription factor NRF2. Immune Modulation: PML::RARα inhibits the transcription factor PU.1, leading to downregulation of immunoproteasome subunits (PSMBs) and impaired antigen presentation, facilitating immune evasion. It also induces the secretion of pro-inflammatory cytokines and chemokines (eg, IL-6, TNF-α), fostering a pro-inflammatory microenvironment. Key differentiation regulators like Irf8 are suppressed, maintaining cellular immaturity. Therapeutic Intervention: The combination of All-Trans Retinoic Acid (ATRA) and Arsenic Trioxide (ATO) directly targets the PML::RARα protein for degradation, reversing the oncogenic reprogramming and leading to differentiation and apoptosis of APL cells. Created with MedPeer (medpeer.cn). |
Application of Proteomics Technology in APL Research
Proteomic technologies are gaining increasing attention in APL research. By utilizing high-throughput mass spectrometry and other advanced methods, these approaches enable comprehensive and quantitative profiling of proteins in cells or tissues, thereby enhancing our understanding of APL biology and revealing potential therapeutic targets.
In clinical applications, proteomics has also demonstrated significant potential for early diagnosis and disease monitoring. Proteomic profiling of serum or bone marrow samples from APL patients can identify disease-specific proteins that may serve as biomarkers for early detection. For example, studies have shown that dysregulated expression of miR-125b actively contributes to APL progression and pathophysiology. Additionally, the inhibition of DDX5 by the 2F5 antibody suppresses APL cell proliferation and promotes differentiation through ROS induction. These findings illustrate that certain proteins involved in cell proliferation and differentiation are significantly upregulated in APL patients, highlighting their potential diagnostic value in early-stage disease.9,22
Protein-Protein Interaction Network Analysis
To better understand the molecular mechanisms of APL, the construction of a protein–protein interaction network (PPIN) related to APL is of great importance. Such a network can help reveal key regulatory nodes and provide a basis for identifying potential therapeutic targets.23
In constructing the PPIN, it is essential to first identify APL-associated proteins, such as PML, RARA, and other key proteins involved in cell proliferation and apoptosis. By analyzing the interactions among these proteins, researchers can pinpoint central players in APL progression—such as Skp2 and JunB—which are involved not only in cell proliferation but also in modulating treatment sensitivity.24
Another critical application of network analysis lies in the identification of potential therapeutic targets. Topological analysis of the PPIN can reveal proteins with high degree and betweenness centrality, which typically function as hub nodes and play essential roles in signal transduction and cellular function maintenance. Studies have shown that Skp2 overexpression is closely associated with APL progression and may promote APL cell proliferation by inhibiting PML::RARα degradation.24 This focus on central nodes offers important directions for novel drug development.
Furthermore, network analysis can be integrated with various bioinformatics tools to optimize treatment strategies. For instance, targeting key signaling pathways or modulating the expression of hub proteins may significantly improve therapeutic outcomes in APL patients.9
Correlation of Proteomic Data with Clinical Parameters
Integrating Proteomic Data with Clinical Parameters in Acute Promyelocytic Leukemia: Insights and Applications
Studies investigating the correlation between proteomic data and clinical parameters in acute promyelocytic leukemia (APL) have revealed close relationships between protein expression changes and clinicopathological features, as well as treatment responses in patients.
Different protein markers exhibit distinct expression patterns in APL patients, and these variations are closely associated with prognosis, disease severity, and therapeutic outcomes. A study investigating the anticancer effects of pioglitazone on APL cells demonstrated that pioglitazone activates PPARγ, significantly reduces viability of APL-derived NB4 cells, induces G1 phase cell cycle arrest, and promotes apoptosis. Its antileukemic effect is mediated through upregulation of PTEN and regulated by the PI3K signaling pathway. Moreover, pioglitazone exhibits synergistic anticancer effects when combined with arsenic trioxide.25
Quantitative proteomic analysis of serum samples from APL patients has revealed associations between specific protein expression levels and patient survival and relapse rates. A comparative study of protein expression in APL versus non-APL AML patients showed higher expression levels of most proteins in APL patients, with the exception of ZNF346, which was more highly expressed in AML. Additionally, APL patients demonstrated significant differences in three protein functional groups (PFGs): “Apoptosis Regulation”, “HIPPO”, and “TP53” signaling. Further analysis identified two distinct protein expression signatures (S1 and S2) using the “MetaGalaxy” method, with S1 associated with favorable prognosis and S2 correlated with disease relapse. The study also confirmed differential expression of HNRNPK in APL patients, supporting its potential as a prognostic biomarker.6
The potential application of protein markers in disease monitoring has also gained considerable attention. Research has shown that miR-146a plays an important role in APL development by suppressing Smad4 protein expression. As an oncogene, miR-146a may serve as a novel prognostic biomarker for APL.26 Similarly, miR-638 shows promise as a potential biomarker for APL diagnosis and targeted therapy response assessment, with peripheral blood miR-638 potentially enabling non-invasive minimal residual disease (MRD) monitoring.27 A constructed transcription factor-lncRNA-target gene network has revealed complex molecular mechanisms regulating APL differentiation, with key transcription factors including E2F1, E2F6, and EBF1 playing central roles. This provides new perspectives for understanding APL molecular mechanisms and establishes a foundation for future development of lncRNA-based diagnostic markers and therapeutic targets.28
The integration of proteomic data with clinical parameters provides substantial biomarker support for diagnosis, prognosis evaluation, and treatment monitoring in APL. These investigations not only enrich our understanding of APL biology but also contribute to enhanced personalization of clinical treatment, ultimately improving patient outcomes and quality of life.
Metabolomics Technology and Its Application in APL
Metabolomics is a scientific discipline focused on characterizing the composition and dynamic changes of small molecule metabolites within biological systems. Through qualitative and quantitative analysis of metabolites, it provides comprehensive information about the metabolic status of an organism, thereby revealing underlying biological mechanisms of disease.29,30 The primary analytical platforms in metabolomics include gas chromatography-mass spectrometry (GC-MS), liquid chromatography-mass spectrometry (LC-MS), and nuclear magnetic resonance (NMR) spectroscopy. These technologies enable efficient separation and detection of small molecule metabolites in biological samples, thereby generating rich datasets for research.31
In recent years, the application of metabolomics in APL has steadily expanded, providing novel approaches for diagnosis, treatment, and prognosis assessment. Metabolic profiling of APL cell lines has revealed distinctive alterations in amino acid metabolism and lipid metabolism. APL cells demonstrate significant increases in metabolites associated with proliferation and energy consumption, which are closely correlated with their high proliferation rates. These metabolic reprogramming features are likely intrinsically linked to the biological characteristics of APL cells, thus offering fresh perspectives for understanding the pathogenesis of this disease.
Dysregulation of Cysteine and Glutathione Metabolism in APL
In APL, aberrant amino acid metabolism—particularly involving cysteine and its metabolite glutathione—plays a critically important role in disease pathophysiology.
Cysteine, a sulfur-containing amino acid, not only participates in protein synthesis but also serves essential functions in cellular antioxidant defense. Studies indicate that cysteine metabolic pathways are significantly altered in APL cells. In the context of cell proliferation and survival, elevated cysteine levels are associated with cellular stress responses.32
During arsenic trioxide (As2O3) metabolism, treated cells exhibit increased S-adenosylhomocysteine (SAH) without significant changes in S-adenosylmethionine (SAM), reflecting cellular capacity to metabolize As2O3. The generation of intracellular methylated metabolites and associated DNA epigenetic modifications may contribute to the efficacy of As2O3 in APL treatment.33 However, this is still a hypothetical mechanism and direct evidence is needed to confirm this.
Glutathione (GSH), a major intracellular antioxidant whose synthesis depends on cysteine, shows level variations in APL that may correlate with resistance to chemotherapeutic agents. Compared with other AML subtypes, APL cases demonstrate significantly lower baseline mRNA expression of γ-glutamylcysteine synthetase (γ-GCS) at onset, with marked upregulation following treatment.34 Functionally, although NB4 cells (an APL cell line) exhibit high sensitivity to As2O3-induced apoptosis in vitro, increased intracellular GSH confers resistance to this agent.35
Disorders of Lipid Metabolism in APL
Disorders of lipid metabolism are recognized as a significant component of the pathophysiology of APL. APL patients frequently exhibit lipid abnormalities during treatment, which are closely associated with prognosis and clinical features. A study of 90 newly diagnosed APL patients revealed that elevated triglyceride (TG) levels correlated positively with high-risk APL, while reduced high-density lipoprotein cholesterol (HDL-C) was associated with low-risk disease. These findings highlight the potential role of lipid metabolism in APL, with low-density lipoprotein cholesterol (LDL-C) emerging as an independent prognostic marker—lower LDL-C levels were linked to poorer clinical outcomes.36 The PTK2 gene plays a crucial role in regulating lipid metabolism, suggesting that interventions targeting this gene may open new avenues for APL treatment.
Furthermore, APL cells can influence lipid metabolism in hepatocytes, leading to increased secretion of triglycerides, cholesterol, and other lipid metabolites. This process may result from the suppression of key transcription factors in hepatocytes through interactions with APL cells, ultimately disrupting hepatic lipid metabolism.37 Additionally, the interaction between PML::RARα and TRIB3 may promote abnormal lipid metabolism by inhibiting PPARγ activity, underscoring the important role of transcription factors in APL-associated lipid dysregulation.
Through in-depth metabolomic investigations, more lipid metabolites related to APL are expected to be identified. These metabolites could not only serve as disease biomarkers but also become novel therapeutic targets, offering new strategies for the treatment of APL patients.
Integrative Analysis of Proteome and Metabolome
In APL research, multi-omics integration of proteomic and metabolomic data provides novel insights into the disease’s complex regulatory networks. The introduction of modeling tools and analytical frameworks enables better elucidation of these networks. Through comprehensive analysis across different omics layers, we can gain deeper understanding of APL cell biology and treatment responses. The fusion of multi-omics data not only facilitates identification of potential biomarkers but also reveals complex mechanisms governing cell fate decisions, thereby advancing the development of personalized treatment strategies.
Association Between Metabolic Flux and Enzyme Abundance/Activity
Changes in metabolic flux are typically driven by alterations in key enzyme abundance or activity. Cytoscape and its plugin MetScape provide ideal platforms for constructing multi-omics networks that visually represent interactions between proteins and metabolites.38 Using the MetScape plugin with APL data, we dynamically visualized protein phosphorylation levels and metabolite concentrations by adjusting node colors and sizes, identifying key hub molecules such as AKT and CD36 regulated by PML::RARα. In the complex pathological state of APL, the PML::RARα fusion protein promotes AKT degradation, suppresses glycolysis, and enhances tricarboxylic acid (TCA) cycle and fatty acid oxidation, leading to metabolic reprogramming.39 In this study, proteomics effectively quantified the abundance of key glycolytic enzymes such as HK2 and PKM2, while metabolomics detected important metabolites including lactate and TCA intermediates. Using Cytoscape’s CytoKegg plugin, we further integrated signaling pathways (eg, RAS-MAPK pathway) with metabolic networks, providing more comprehensive biological insights.
MetaboAnalyst is another specialized software for metabolic pathway enrichment, topological analysis, and multi-omics data integration.40 We input lists of significantly differentially expressed proteins (eg, AKT, STAT3) and metabolites (eg, TCA intermediates, fatty acids) from APL into MetaboAnalyst for joint pathway analysis, such as KEGG pathway enrichment.
Integration of Protein Phosphorylation Patterns and Metabolic Dynamics
Phosphorylation modifications directly regulate enzyme activity, thereby influencing metabolic flux. We employed Multi-Omics Factor Analysis (MOFA) to integrate phosphoproteomic data (dynamic phosphorylation events), metabolomic data (metabolite concentrations), and clinical data (eg, drug resistance status), identifying shared factors such as the “oxidative phosphorylation activation factor”, which holds significant implications for understanding resistance mechanisms.
In APL research, this approach revealed how PML::RARα reprograms fatty acid synthesis flux through phosphorylation modifications (eg, phosphorylation of acetyl-CoA carboxylase), providing new insights into metabolic regulation.39 APL cells exhibit lower oxidative phosphorylation levels than normal promyelocytes and demonstrate vulnerability under OXPHOS inhibition, with metabolic reprogramming enhancing their dependence on mitochondrial function. Furthermore, the combination of venetoclax (a BCL-2 inhibitor) with azacitidine (a hypomethylating agent) shows significant synergistic killing effects on PML::RARα-positive cells.41
In summary, proteomic analyses suggest that PML::RARα may regulate metabolic enzyme activity by influencing the phosphorylation of specific kinases, such as acetyl-CoA carboxylase. Concurrently, metabolomic data reveal a corresponding accumulation of fatty acid synthesis precursors and a reduction in oxidation products. Integrative analysis can directly test the hypothesis that specific phosphorylation events drive quantitative changes in metabolic flux, providing a molecular mechanism for reprogramming. Furthermore, studies have identified a unique molecular network of proteins and metabolites in the plasma of APL patients, the features of which are closely associated with the proliferative and hypermetabolic state of leukemic cells.32 These integration-based molecular signatures not only deepen the understanding of APL pathogenesis but also provide new perspectives and targets for biomarker discovery and the development of combination therapies, such as the simultaneous targeting of metabolic enzymes and signaling pathway inhibitors.
Impact of ATRA/ATO Therapy on the Proteome and Metabolome of APL
In recent years, the combination therapy of ATRA and ATO has significantly improved outcomes in patients with APL, establishing itself as the standard treatment regimen. ATRA primarily exerts its antileukemic effects by inducing cellular differentiation and apoptosis, while ATO enhances therapeutic efficacy by promoting apoptosis and suppressing leukemia stem cell growth. Studies indicate that ATRA/ATO treatment substantially influences the proteomic and metabolomic profiles of leukemia cells,42 providing new insights into the therapeutic mechanisms in APL.
Effects of ATRA on APL
The impact of ATRA on the proteome has been validated in multiple studies. ATRA induces differentiation of APL cells, a process accompanied by expression changes in specific proteins. Research demonstrates that ATRA treatment upregulates key transcription factors such as PU.1, C/EBPβ, and C/EBPε, which play crucial roles in leukemic cell differentiation. Beyond promoting normal myeloid differentiation, ATRA also inhibits tumor cell proliferation and enhances cellular sensitivity to other treatments—for instance, combination with LSD1 inhibitors demonstrates enhanced differentiation effects.43
In APL treatment based on ATRA, integrated analysis of protein and metabolite expression changes reveals a coordinated regulatory relationship between them. ATRA promotes calcium influx in APL cells and enhances autophagy through activation of the CAMKK2/AMPK signaling pathway—a process demonstrated to be closely associated with cellular differentiation and death.44 Furthermore, ATRA-activated PML::RARα triggers a positive feedback gene regulatory circuit composed of the transcription factors SPI1 and CEBPE, driving further differentiation of NB4 cells and influencing APL cell fate.45
The interplay between APL cells and immune cells involves alterations in metabolic pathways. Studies elucidating the mechanisms underlying ATRA-induced granulocytic differentiation have revealed that ATRA treatment in NB4 cells drives the activation of aerobic glycolysis and reduces OXPHOS-dependent ATP production. Furthermore, ATRA drives a metabolic shift towards aerobic glycolysis in differentiating NB4 cells, revealing a potential targetable dependency.46
Effects of ATO on APL
Ceramide is known to induce apoptosis. Studies reveal that ATO exerts a more potent effect on ceramide levels in APL cells compared to ATRA. ATO downregulates neutral sphingomyelinase activity—contrary to ATRA’s action—and ATO-induced ceramide accumulation results not from induction of acid sphingomyelinase but rather from de novo ceramide synthesis and inhibition of glucosylceramide synthase activity.47
ATO also binds to cysteine residues in the PML fusion protein, promoting PML nuclear body formation and thereby activating cellular stress responses, cell cycle regulation, and apoptosis. Through enhanced SUMOylation and downregulation of PCGF2, ATO promotes PML::RARα degradation—a process involving the SUMO-dependent ubiquitin ligase activity of RNF4, ultimately leading to proteasomal degradation of PML::RARα.48
Furthermore, ATO-resistant NB4 cells exhibit an altered metabolic profile characterized by increased lipid accumulation. The combination of a CD36 inhibitor with ATO demonstrates potential synergistic effects, suggesting that targeting fatty acid metabolism may represent a strategy to overcome drug resistance.39
Effects of ATRA/ATO Combination Therapy
The combination of ATRA and ATO also significantly alters proteomic and metabolic characteristics of APL cells.34 Research shows that enhanced APL cell differentiation by the Src family kinase (SFK) inhibitor PP2, when combined with ATRA or ATO, involves differential activation of retinoic acid-induced genes.49 Moreover, Irf8 expression increases during spontaneous differentiation of APL progenitors in vivo, representing a PML::RARα- deregulated gene refractory to ATRA/ATO signaling. Irf8-driven monocytic maturation reduces APL leukemogenic potential.50 Thus, certain PML::RARα- deregulated genes unresponsive to ATRA/ATO signaling may serve as critical regulators of APL cell immaturity and leukemogenic potential, offering targets for novel therapeutic strategies in ATRA/ATO-resistant APL cases.
Role of the Immune System in APL Treatment and Its Proteomic/Metabolomic Characteristics
The role of the immune system in APL is multifaceted, encompassing both direct effects on leukemic cells and modulation of immune cell function. Integrated proteomic and metabolomic analyses provide a comprehensive approach to decipher the immune landscape of APL and its potential therapeutic implications.
Role of the Immune System in APL Treatment
In recent years, growing evidence has highlighted the critical role of the immune system in APL treatment. The immunological status of APL patients is closely linked to therapeutic outcomes, underscoring the importance of characterizing patient proteomic and metabolomic profiles to elucidate the immune system’s contribution.
First, significant alterations in immune cell populations occur in APL patients, particularly during treatment with ATRA and ATO.51 Yang et al demonstrated that the PML::RARα oncoprotein suppresses PU.1-dependent activation of immune subunits, potentially facilitating immune evasion by APL cells during leukemogenesis. ATRA treatment may counteract this by reactivating antigen processing and presentation mechanisms, thereby enhancing T cell-mediated recognition of leukemic cells.52 Such treatments not only induce differentiation and apoptosis of leukemic cells but also modulate immune cell functions to augment anti-tumor immunity.53 ATRA has been shown to upregulate specific cytokines, enhancing T cell activity and improving the overall immune milieu in patients.54,55 This process of immune remodeling is likely integral to the therapeutic efficacy of APL treatments.
Second, proteomic and metabolomic profiling of serum samples from APL patients has identified key immune-related proteins, such as cytokines and chemokines, implicated in APL pathophysiology. Dual induction therapy for APL can trigger Differentiation Syndrome (DS), a severe systemic inflammatory response mediated by increased expression of cytokines, chemokines, and adhesion molecules in differentiating blasts.56 Studies indicate that ATRA stimulation of the APL cell line NB4 induces the expression of various CC-chemokines (CCLs) and their receptors, leading to elevated chemokine levels and enhanced chemotaxis.57,58 Organs such as the kidneys and lungs can become targets for infiltrating differentiated blasts in DS, resulting in irreversible damage and contributing to adverse outcomes in APL. These findings provide insights into the immunologic mechanisms underlying APL induction therapy and highlight that prognosis management remains a clinical challenge, necessitating a deeper understanding of its pathogenesis.
Furthermore, another crucial aspect of the immune system in APL treatment involves its role in drug biotransformation. For instance, metabolites of Arsenic Trioxide (ATO) are believed to possess enhanced anti-tumor activity. Metabolomic analyses enable the tracking of ATO’s metabolic pathways and help delineate its role in immune responses.59 This suggests that metabolites are not merely reflections of therapeutic effect but may actively participate in immune modulation.
Collectively, these studies provide a theoretical foundation for personalized APL therapy and indicate future research directions, particularly in developing novel immunotherapies and assessing patient prognosis.
Proteomic Profiling of Immune Signaling Pathways
Proteomic characterization of immune signaling pathways in APL has garnered significant interest.60 Niu Qiao et al reported that IFN-γ activates macrophages and T cells, bolstering their tumoricidal capacity and thereby inhibiting the growth and metastasis of APL cells.32 This demonstrates IFN-γ’s role in promoting anti-tumor immunity. Studies also indicate enhanced IFN-γ signaling during the first complete hematologic remission, characterizing the tumor-suppressive function of the immune system. This enhancement may be triggered by therapy-induced cell death or senescence and subsequent supraphysiological levels of intracellular proteins. Consequently, activation of the IFN-γ signaling pathway, particularly in the later stages of APL treatment, may be closely associated with tumor suppression.
Simultaneously, altered expression of immune-related proteins correlates with treatment efficacy. Research has shown that culturing endothelial cells with high concentrations of IFN-γ alone, or low concentrations of IL-1β combined with IFN-γ, induces a robust apoptotic response via caspase-8 activation and DNA fragmentation. These retinoic acid-induced events may contribute to the pathogenesis of Retinoic Acid Syndrome (RAS) in APL patients.61 The mechanisms of action of Interferon-gamma (IFN-γ) and Interleukin-1β (IL-1β) reveal their potential applicability in APL treatment, and IFN-γ expression levels may serve as an important biomarker for evaluating therapeutic efficacy.
In summary, immune signaling pathways hold significant implications in the proteomic landscape of APL. IFN-γ activation is not only associated with tumor suppression but also closely linked to expression changes in immune-related proteins, highlighting its potential as a therapeutic biomarker.60 These findings offer novel perspectives for APL treatment and may guide future research towards more effective immunotherapeutic strategies.
Interaction Between the Immune Microenvironment and Tumor Cells
In APL, the immune microenvironment significantly influences tumor cell metabolism and protein secretion. Studies reveal that increased proliferation and energy consumption by APL blasts alter secretome and metabolome profiles. Furthermore, proteins enriched in APL plasma or metabolites altered by enzymatically catalyzed plasma changes are potential trans-regulatory targets of PML::RARα. It is established that APL cells can remodel the surrounding immune microenvironment, leading to changes in the distribution and function of immune cells.32
The mechanism by which microenvironmental regulation influences treatment response is equally critical. In APL therapy, standard regimens combining ATRA and ATO significantly improve patient outcomes. However, variability in treatment efficacy is often linked to alterations in the tumor microenvironment. Research indicates that pre- and post-treatment changes in the immune microenvironment of APL cells can impact therapeutic effectiveness and the emergence of drug resistance. During ATRA treatment, a strongly immunosuppressive tumor microenvironment can lead to inactivation of infiltrating Natural Killer (NK) cells. APL cells secrete various cytokines, such as NKG2D ligands (NKG2DLs), and other bioactive molecules, promoting the infiltration of tumor-associated macrophages and other immune cells, thereby facilitating tumor growth and metastasis.62 Conversely, following ATO treatment, levels of regulatory T cells (Tregs) in peripheral blood and serum IL-10 decrease significantly. ATO reduces Treg numbers and suppresses their function, consequently enhancing effector T cell (Teff) activity and overall anti-tumor immunity.63 Therefore, understanding how the immune microenvironment influences therapy response is paramount for optimizing APL treatment strategies.
These findings lead to a compelling yet still speculative hypothesis: that modulating the immune microenvironment could synergize with direct targeting of APL cells to improve outcomes. Future work must functionally validate this and determine the key molecular mediators.
Limitations and Future Perspectives
While this review synthesizes the current landscape of integrated proteomic and metabolomic profiling in APL, several limitations in the existing literature must be acknowledged (a summary of these key omics studies is provided in Table 1). The field remains in its nascent stages, dominated by proof-of-concept and small-scale studies. Technical constraints, such as limited proteome coverage by mass spectrometry, challenges in detecting low-abundance signaling proteins, and the dynamic range of metabolomic detection, often result in an incomplete molecular portrait. Furthermore, substantial heterogeneity in sample processing, analytical platforms, and data pipelines across studies complicates direct comparison and meta-analysis, limiting the generalizability of findings. The heavy reliance on cell line models over diverse primary patient samples also restricts insights into clinical heterogeneity, and correlations with detailed clinical outcomes remain underdeveloped. Finally, as a narrative review, our synthesis may be influenced by selection bias toward established pathways, potentially overlooking emerging aspects of APL biology.
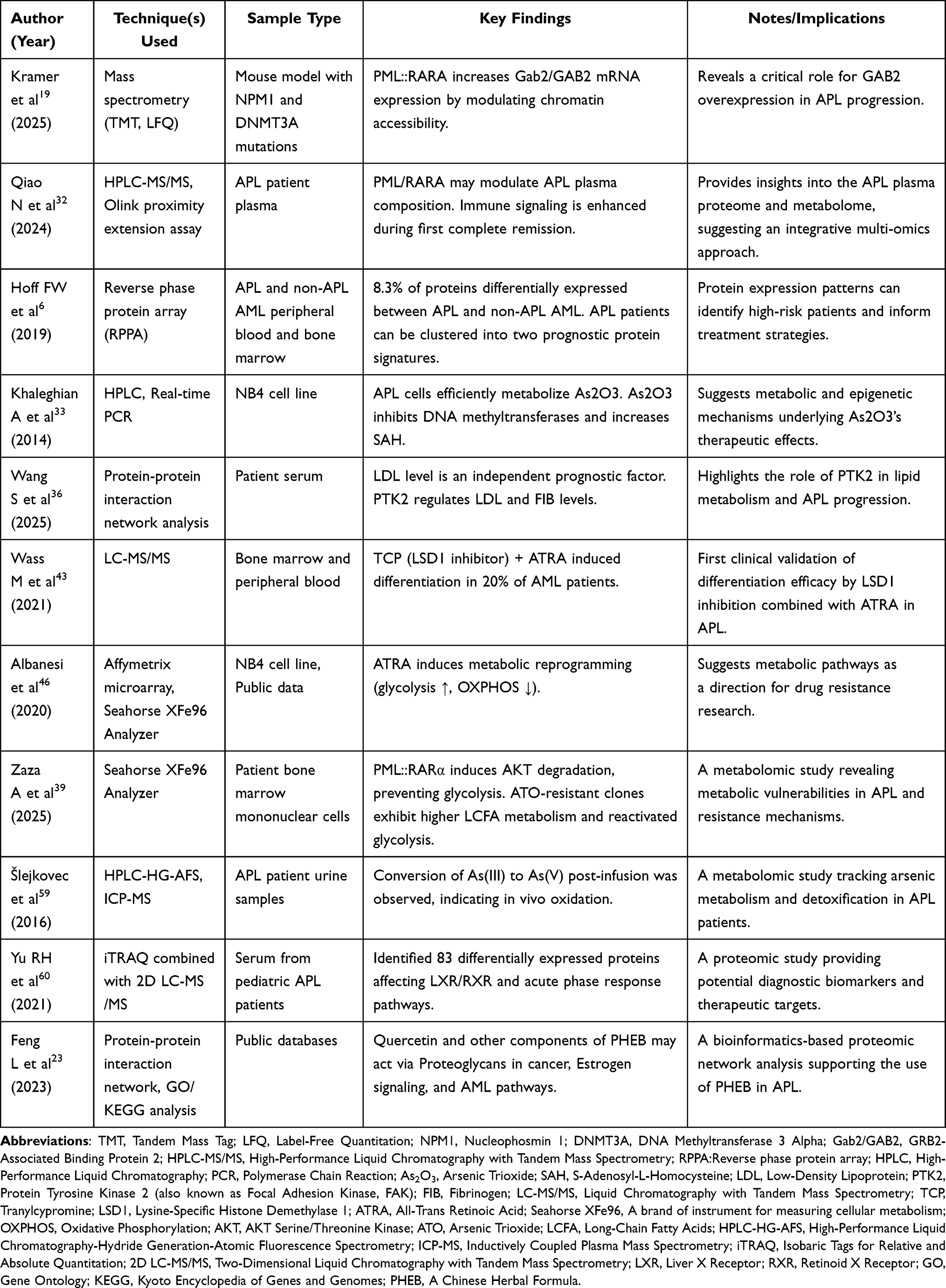 |
Table 1 Key Proteomic and Metabolomic Profiling Studies in Acute Promyelocytic Leukemia |
Addressing these limitations will be crucial to advancing the field. Future research should prioritize the following directions:
Advancing Multi-Omics Integration and Single-Cell Applications
The development and application of robust computational frameworks—such as MOFA, metabolic network tools (eg, Cytoscape with MetScape), and artificial intelligence (AI)—are essential for extracting biologically meaningful insights from complex datasets. Deep learning models can help identify non-linear patterns and predictive biomarkers from integrated proteomic and metabolomic data, improving disease classification and outcome prediction.64–67 Additionally, single-cell multi-omics technologies promise to resolve cellular heterogeneity and uncover subtype-specific metabolic-proteomic adaptations in APL, providing a deeper understanding of treatment resistance and relapse mechanisms.68
Biomarker Discovery and Clinical Translation
Future studies should focus on validating combined protein-metabolite biomarker panels to enhance diagnostic specificity, prognostic stratification, and therapy response monitoring. For example, overexpression of miR-125b has been linked to APL progression and may represent a promising therapeutic target.9 By systematically integrating proteomic and metabolomic data with clinical variables, such biomarker systems could form the basis for personalized treatment strategies.
Toward Personalized Therapy and Novel Therapeutics
While the ATO/ATRA regimen has revolutionized APL treatment, patient responses vary.69 Integration of proteo-metabolomic profiles may guide risk-adapted and individualized therapy. Simultaneously, functional validation of multi-omics-derived targets—such as ZDHHC3, whose inhibition impairs APL progression70—will enable the development of new targeted agents. Compounds like low-dose tetra-arsenic tetra-sulfide (As4S4), which enhances NK cell-mediated cytotoxicity against resistant APL cells,62 also underscore the therapeutic potential revealed by mechanistically-informed screening.
In summary, overcoming current technical and analytical challenges will require close collaboration among omics technologies, computational biology, and clinical research. The continued refinement of multi-omics integration holds significant promise for refining APL classification, uncovering novel therapeutic vulnerabilities, and ultimately improving patient outcomes.
Conclusion
In recent years, with the continuous advancement of proteomics and metabolomics technologies, our understanding of the molecular mechanisms of APL has deepened significantly. Particularly regarding the regulatory network of the PML::RARα fusion protein and its role in metabolic reprogramming, these studies have revealed potential pathogenic mechanisms and therapeutic targets in APL. The integration of proteomics and metabolomics not only provides a powerful tool for exploring tumor biology but also lays the foundation for optimizing clinical treatment strategies.
ATRA and ATO treatment in APL significantly changes protein and metabolite levels in plasma, impacting tumor cell behavior and the immune response, highlighting the tumor-host interaction’s importance.
Future research should focus on microenvironment regulation and immunotherapy combinations for better APL treatments, integrating diverse study findings to enhance understanding and support clinical practice. Despite promising results, ongoing exploration is necessary for personalized medicine, emphasizing the balance of omics data with clinical outcomes to innovate treatment strategies for APL patients.
Data Sharing Statement
The datasets used and analyzed in this study are available from the corresponding author on reasonable request.
Author Contributions
Co-first authors: Ting Liu and Mengqi Qu. They contributed equally to this work. Co-corresponding authors: Ning Wang and Xue Xing. They contributed equally to this work. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Cicconi L, Fenaux P, Kantarjian H, Tallman M, Sanz MA, Lo-Coco F. Molecular remission as a therapeutic objective in acute promyelocytic leukemia. Leukemia. 2018;32(8):1671–1678. doi:10.1038/s41375-018-0219-5
2. Yin W, Yan X, Cai J, et al. Incidence, mortality, and survival associated with acute leukaemia subtypes by age group in China: a population-based cancer registry analysis and cohort study. Lancet Haematol. 2025;12(10):e808–e822. doi:10.1016/S2352-3026(25)00236-4
3. Thomas X. Acute promyelocytic leukemia: a history over 60 years-from the most malignant to the most curable form of acute leukemia. Oncol Ther. 2019;7(1):33–65. doi:10.1007/s40487-018-0091-5
4. Moosavi MA, Djavaheri-Mergny M. Autophagy: new insights into mechanisms of action and resistance of treatment in acute promyelocytic leukemia. Int J Mol Sci. 2019;20(14):3559. doi:10.3390/ijms20143559
5. Gianni’ M, Goracci L, Schlaefli A, et al. Role of cardiolipins, mitochondria, and autophagy in the differentiation process activated by all-trans retinoic acid in acute promyelocytic leukemia. Cell Death Dis. 2022;13(1):30. doi:10.1038/s41419-021-04476-z
6. Hoff FW, Hu CW, Qutub AA, et al. Proteomic profiling of acute promyelocytic leukemia identifies two protein signatures associated with relapse. Proteomics Clin Appl. 2019;13(4):e1800133. doi:10.1002/prca.201800133
7. Fang J, Chen SJ, Tong JH, Wang ZG, Chen GQ, Chen Z. Treatment of acute promyelocytic leukemia with ATRA and As2O3: a model of molecular target-based cancer therapy. Cancer Biol Ther. 2002;1(6):614–620. doi:10.4161/cbt.308
8. Cho Y, Hyun J, Kim M, Han B, Lee YK. Acute promyelocytic leukemia with a BCR-ABL1 rearrangement in a minor clone. Lab Med. 2022;53(3):326–329. doi:10.1093/labmed/lmab097
9. Guo B, Qin R, Chen JJ, Pan W, Lu XC. MicroRNA-125b accelerates and promotes PML-RARa-driven murine acute promyelocytic leukemia. Biomed Environ Sci. 2022;35(6):485–493. doi:10.3967/bes2022.067
10. Belmahi S, Kajeiou Z, Yacoubi L, et al. Acute lower limb ischemia revealing hypo granular acute promyelocytic leukemia. Leuk Res Rep. 2024;23:100488. doi:10.1016/j.lrr.2024.100488
11. Jiang G, Bi K, Tang T, et al. Effect of arsenic trioxide on cytokine expression by acute promyelocytic leukemia cells. Chin Med J. 2003;116(11):1639–1643.
12. Su Z, Liu X. Comment on Geoffroy, M.-C.; de Thé, H. classic and variants APLs, as viewed from a therapy response. Cancers 2020, 12, 967. Cancers. 2021;13(23):5883. doi:10.3390/cancers13235883
13. Gaillard C, Tokuyasu TA, Rosen G, et al. Transcription and methylation analyses of preleukemic promyelocytes indicate a dual role for PML/RARA in leukemia initiation. Haematologica. 2015;100(8):1064–1075. doi:10.3324/haematol.2014.123018
14. Katerndahl CDS, Rogers ORS, Day RB, et al. PML::RARA and GATA2 proteins interact via DNA templates to induce aberrant self-renewal in mouse and human hematopoietic cells. Proc Natl Acad Sci U S A. 2024;121(18):e2317690121. doi:10.1073/pnas.2317690121
15. Jaffray EG, Tatham MH, Mojsa B, et al. The p97/VCP segregase is essential for arsenic-induced degradation of PML and PML::RARA. J Cell Biol. 2023;222(4). doi:10.1083/jcb.202201027
16. Testa U, Pelosi E. Function of PML-RARA in Acute Promyelocytic Leukemia. Adv Exp Med Biol. 2024;1459:321–339. doi:10.1007/978-3-031-62731-6_14
17. Piredda ML, Gaur G, Catalano G, et al. PML/RARA inhibits expression of HSP90 and its target AKT. Br J Haematol. 2019;184(6):937–948. doi:10.1111/bjh.15715
18. Jia PM, Dou AX, Zhang CL, et al. Effects of PML-RARalpha on cAMP-induced AML cell differentiation. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2008;16(6):1275–1278.
19. Kramer MH, Richardson SN, Li Y, et al. Overexpression of the signaling coordinator GAB2 can play an important role in acute myeloid leukemia progression. J Clin Invest. 2025;135(21):e195929. doi:10.1172/JCI195929
20. Banella C, Catalano G, Travaglini S, et al. PML/RARa interferes with NRF2 transcriptional activity increasing the sensitivity to ascorbate of acute promyelocytic leukemia cells. Cancers. 2019;12(1):95. doi:10.3390/cancers12010095
21. Ablain J, Rice K, Soilihi H, de Reynies A, Minucci S, de Thé H. Activation of a promyelocytic leukemia-tumor protein 53 axis underlies acute promyelocytic leukemia cure. Nat Med. 2014;20(2):167–174. doi:10.1038/nm.3441
22. Wu J, You YQ, Ma YX, et al. DDX5-targeting fully human monoclonal autoantibody inhibits proliferation and promotes differentiation of acute promyelocytic leukemia cells by increasing ROS production. Cell Death Dis. 2020;11(7):552. doi:10.1038/s41419-020-02759-5
23. Feng L, Zhu S, Ma J, et al. Integrated bioinformatics analysis and network pharmacology to explore the potential mechanism of Patrinia heterophylla Bunge against acute promyelocytic leukemia. Medicine. 2023;102(40):e35151. doi:10.1097/MD.0000000000035151
24. Dan W, Zhong L, Yu L, et al. Skp2 promotes APL progression through the stabilization of oncoprotein PML-RARα and the inhibition of JunB expression. Life Sci. 2022;289:120231. doi:10.1016/j.lfs.2021.120231
25. Esmaeili S, Safaroghli-Azar A, Pourbagheri-Sigaroodi A, et al. Stimulation of Peroxisome Proliferator-Activated Receptor-Gamma (PPARγ) using pioglitazone decreases the survival of acute promyelocytic leukemia cells through up-regulation of PTEN expression. Anticancer Agents Med Chem. 2021;21(1):108–119. doi:10.2174/1871520620666200817101533
26. Xu L, Zhong H, Wan H, et al. miR-146a expression level as a novel putative prognostic marker for acute promyelocytic leukemia. Dis Markers. 2014;2014:150604. doi:10.1155/2014/150604
27. Bai Y, Chen C, Guo X, et al. miR-638 in circulating leukaemia cells as a non-invasive biomarker in diagnosis, treatment response and MRD surveillance of acute promyelocytic leukaemia. Cancer Biomark. 2020;29(1):125–137. doi:10.3233/CBM-190899
28. Yu J, Guo XL, Bai YY, et al. Genome-wide profiling of lncRNA expression patterns in patients with acute promyelocytic leukemia with differentiation therapy. Oncol Rep. 2018;40(3):1601–1613. doi:10.3892/or.2018.6521
29. Sheikh KD, Khanna S, Byers SW, Fornace AJ, Cheema AK. Small molecule metabolite extraction strategy for improving LC/MS detection of cancer cell metabolome. J Biomol Tech. 2011;22(1):1–4.
30. Lin C, Tian Q, Guo S, et al. Metabolomics for clinical biomarker discovery and therapeutic target identification. Molecules. 2024;29(10):2198. doi:10.3390/molecules29102198
31. Wishart DS, Cheng LL, Copié V, et al. NMR and metabolomics-A roadmap for the future. Metabolites. 2022;12(8):678. doi:10.3390/metabo12080678
32. Qiao N, Lyu Y, Liu F, et al. Cross-sectional network analysis of plasma proteins/metabolites correlated with pathogenesis and therapeutic response in acute promyelocytic leukemia. Front Med. 2024;18(2):327–343. doi:10.1007/s11684-023-1022-x
33. Khaleghian A, Ghaffari SH, Ahmadian S, Alimoghaddam K, Ghavamzadeh A. Metabolism of arsenic trioxide in acute promyelocytic leukemia cells. J Cell Biochem. 2014;115(10):1729–1739. doi:10.1002/jcb.24838
34. Carnicer MJ, Bernardini S, Bellincampi L, et al. Role of gamma-glutamyl cysteine synthetase (gamma-GCS) gene expression as marker of drug sensitivity in acute myeloid leukemias. Clin Chim Acta. 2006;365(1–2):342–345. doi:10.1016/j.cca.2005.09.012
35. Zuo L, Li J, Yang Y, et al. Sodium selenite induces apoptosis in acute promyelocytic leukemia-derived NB4 cells by a caspase-3-dependent mechanism and a redox pathway different from that of arsenic trioxide. Ann Hematol. 2004;83(12):751–758. doi:10.1007/s00277-004-0920-5
36. Wang S, Wang Q, Lv S, Qin L. Prognostic value of serum lipids in newly diagnosed acute promyelocytic leukemia. Front Oncol. 2025;15:1522239. doi:10.3389/fonc.2025.1522239
37. Li K, Wang F, Yang ZN, et al. PML-RARα interaction with TRIB3 impedes PPARγ/RXR function and triggers dyslipidemia in acute promyelocytic leukemia. Theranostics. 2020;10(22):10326–10340. doi:10.7150/thno.45924
38. Saito R, Smoot ME, Ono K, et al. A travel guide to Cytoscape plugins. Nat Methods. 2012;9(11):1069–1076. doi:10.1038/nmeth.2212
39. Zaza A, Zardo G, Banella C, et al. PML::RARα+ myeloid cells display metabolic alterations that can be targeted to treat resistant/relapse acute promyelocytic leukemias. Leukemia. 2025;39(11):2708–2720. doi:10.1038/s41375-025-02738-9
40. Ewald JD, Zhou G, Lu Y, et al. Web-based multi-omics integration using the analyst software suite. Nat Protoc. 2024;19(5):1467–1497. doi:10.1038/s41596-023-00950-4
41. Milella M, Konopleva M, Precupanu CM, et al. MEK blockade converts AML differentiating response to retinoids into extensive apoptosis. Blood. 2007;109(5):2121–2129. doi:10.1182/blood-2006-05-024679
42. Wang HY, Gong S, Li GH, et al. An effective and chemotherapy-free strategy of all-trans retinoic acid and arsenic trioxide for acute promyelocytic leukemia in all risk groups (APL15 trial). Blood Cancer J. 2022;12(11):158. doi:10.1038/s41408-022-00753-y
43. Wass M, Göllner S, Besenbeck B, et al. A proof of concept Phase I/II pilot trial of LSD1 inhibition by tranylcypromine combined with ATRA in refractory/relapsed AML patients not eligible for intensive therapy. Leukemia. 2021;35(3):701–711. doi:10.1038/s41375-020-0892-z
44. Hu L, Li Q, Wang J, et al. The CDK4/6 inhibitor palbociclib synergizes with ATRA to induce differentiation in AML. Mol Cancer Ther. 2024;23(7):961–972. doi:10.1158/1535-7163.MCT-23-0528
45. Tian X, Zhang L, Xiang G, et al. Single-cell multiomics reveals a gene regulatory circuit driving leukemia cell differentiation. Oncogene. 2025;44(19):1350–1360. doi:10.1038/s41388-025-03309-z
46. Albanesi J, Noguera NI, Banella C, et al. Transcriptional and metabolic dissection of atra-induced granulocytic differentiation in NB4 acute promyelocytic leukemia cells. Cells. 2020;9(11):2423. doi:10.3390/cells9112423
47. Dbaibo GS, Kfoury Y, Darwiche N, et al. Arsenic trioxide induces accumulation of cytotoxic levels of ceramide in acute promyelocytic leukemia and adult T-cell leukemia/lymphoma cells through de novo ceramide synthesis and inhibition of glucosylceramide synthase activity. Haematologica. 2007;92(6):753–762. doi:10.3324/haematol.10968
48. Borges GSM, Sicard P, de Mello Gomides Loures C, et al. Tocotrienols-enriched self-nanoemulsifying drug delivery system enhances the antileukemic activity of all-trans retinoic acid but not electrocardiogram alterations evoked by its combination with arsenic trioxide. AAPS PharmSciTech. 2023;24(3):79. doi:10.1208/s12249-023-02531-w
49. Jung YS, Cheong HJ, Kim SJ, et al. Src family kinase inhibitor PP2 enhances differentiation of acute promyelocytic leukemia cell line induced by combination of all-trans-retinoic acid and arsenic trioxide. Leuk Res. 2014;38(8):977–982. doi:10.1016/j.leukres.2014.05.019
50. Liu X, Chen J, Yu S, et al. All-trans retinoic acid and arsenic trioxide fail to derepress the monocytic differentiation driver Irf8 in acute promyelocytic leukemia cells. Cell Death Dis. 2017;8(5):e2782. doi:10.1038/cddis.2017.197
51. Chen Y, Tong X, Lu R, Zhang Z, Ma T. All-trans retinoic acid in hematologic disorders: not just acute promyelocytic leukemia. Front Pharmacol. 2024;15:1404092. doi:10.3389/fphar.2024.1404092
52. Yang XW, Wang P, Liu JQ, et al. Coordinated regulation of the immunoproteasome subunits by PML/RARα and PU.1 in acute promyelocytic leukemia. Oncogene. 2014;33(21):2700–2708. doi:10.1038/onc.2013.224
53. Furugaki K, Pokorna K, Le Pogam C, et al. DNA vaccination with all-trans retinoic acid treatment induces long-term survival and elicits specific immune responses requiring CD4+ and CD8+ T-cell activation in an acute promyelocytic leukemia mouse model. Blood. 2010;115(3):653–656. doi:10.1182/blood-2007-08-109009
54. Park HY, Park JY, Kim JW, et al. Differential expression of dendritic cell markers by all-trans retinoic acid on human acute promyelocytic leukemic cell line. Int Immunopharmacol. 2004;4(13):1587–1601. doi:10.1016/j.intimp.2004.07.010
55. Sun X, Mu X, Li F, Wang Y, Yang X, Guo Q. Roles of PADI4 in the expression of cytokines involved in inflammation and adhesion in differentiated NB4 cells treated with ATRA. Exp Ther Med. 2023;25(3):118. doi:10.3892/etm.2023.11817
56. Shen K, Yang C, Huang J, Shuai X, Niu T, Ma H. The clinical characteristics and implications of acute kidney injury during induction therapy for acute promyelocytic leukemia. Front Pharmacol. 2025;16:1540409. doi:10.3389/fphar.2025.1540409
57. Luesink M, Pennings JL, Wissink WM, et al. Chemokine induction by all-trans retinoic acid and arsenic trioxide in acute promyelocytic leukemia: triggering the differentiation syndrome. Blood. 2009;114(27):5512–5521. doi:10.1182/blood-2009-02-204834
58. Shibakura M, Niiya K, Niiya M, et al. Induction of CXC and CC chemokines by all-trans retinoic acid in acute promyelocytic leukemia cells. Leuk Res. 2005;29(7):755–759. doi:10.1016/j.leukres.2005.01.005
59. Šlejkovec Z, Podgornik H, Černelč P, Falnoga I. Exceptions in patterns of arsenic compounds in urine of acute promyelocytic leukaemia patients treated with As2O3. Biometals. 2016;29(1):107–118. doi:10.1007/s10534-015-9901-5
60. Yu RH, Zhang JY, Liu YF, Zhu ZM. Screening serum differential proteins in children with acute promyelocytic leukemia based on iTRAQ technique. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2021;29(5):1462–1470. doi:10.19746/j.cnki.issn.1009-2137.2021.05.014
61. Gao Y, Camacho LH, Mehta K. Retinoic acid-induced CD38 antigen promotes leukemia cells attachment and interferon-gamma/interleukin-1beta-dependent apoptosis of endothelial cells: implications in the etiology of retinoic acid syndrome. Leuk Res. 2007;31(4):455–463. doi:10.1016/j.leukres.2006.07.004
62. Liu Y, Jia Y, Liu Y, Chen X, Zhang M. Tetra-arsenic tetra-sulfide enhances NK-92MI mediated cellular immunotherapy in all-trans retinoic acid-resistant acute promyelocytic leukemia. Invest New Drugs. 2022;40(6):1231–1243. doi:10.1007/s10637-022-01313
63. Xu W, Li X, Quan L, et al. Arsenic trioxide decreases the amount and inhibits the function of regulatory T cells, which may contribute to its efficacy in the treatment of acute promyelocytic leukemia. Leuk Lymphoma. 2018;59(3):650–659. doi:10.1080/10428194.2017.1346253
64. van Hilten A, van Rooij J, Consortium BIOS, et al. Phenotype prediction using biologically interpretable neural networks on multi-cohort multi-omics data. NPJ Syst Biol Appl. 2024;10(1):81. doi:10.1038/s41540-024-00405-w
65. Găman MA, Dugăeşescu M, Popescu DC. Applications of artificial intelligence in acute promyelocytic leukemia: an avenue of opportunities? A systematic review. J Clin Med. 2025;14(5):1670. doi:10.3390/jcm14051670
66. Cox AM, Kim D, García R, Fuda FS, Weinberg OK, Chen W. Automated prediction of acute promyelocytic leukemia from flow cytometry data using a graph neural network pipeline. Am J Clin Pathol. 2024;161(3):264–272. doi:10.1093/ajcp/aqad145
67. Ouyang N, Wang W, Ma L, et al. Diagnosing acute promyelocytic leukemia by using convolutional neural network. Clin Chim Acta. 2021;512:1–6. doi:10.1016/j.cca.2020.10.039
68. Poplineau M, Platet N, Mazuel A, et al. Noncanonical EZH2 drives retinoic acid resistance of variant acute promyelocytic leukemias. Blood. 2022;140(22):2358–2370. doi:10.1182/blood.2022015668
69. Sasijareonrat N, Jahn N, Ungprasert P, Owattanapanich W. Efficacy and the adverse effects of oral versus intravenous arsenic for acute promyelocytic leukemia: a meta-analysis of randomized-controlled studies. Technol Cancer Res Treat. 2020;19:1533033820937008. doi:10.1177/1533033820937008
70. Shao XJ, Wang W, Xu AX, et al. Palmitoyltransferase ZDHHC3 is essential for the oncogenic activity of PML/RARα in acute promyelocytic leukemia. Acta Pharmacol Sin. 2025;46(2):462–473. doi:10.1038/s41401-024-01371-z
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Vitamin B6 Metabolic Pathway is Involved in the Pathogenesis of Liver Diseases via Multi-Omics Analysis
Mei M, Liu D, Tang X, You Y, Peng B, He X, Huang J
Journal of Hepatocellular Carcinoma 2022, 9:729-750
Published Date: 11 August 2022
Metabolomics of Extracellular Vesicles: A Future Promise of Multiple Clinical Applications
Wu Y, Chen W, Guo M, Tan Q, Zhou E, Deng J, Li M, Chen J, Yang Z, Jin Y
International Journal of Nanomedicine 2022, 17:6113-6129
Published Date: 7 December 2022
A Novel Metabolic Score for Predicting the Acute Exacerbation in Patients with Chronic Obstructive Pulmonary Disease
Peng L, You H, Xu MY, Dong ZY, Liu M, Jin WJ, Zhou C
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:785-795
Published Date: 5 May 2023
In vitro Activity of Cefepime/Avibactam Against Carbapenem Resistant Klebsiella pneumoniae and Integrative Metabolomics-Proteomics Approach for Resistance Mechanism: A Single-Center Study
Wen L, Luo C, Chen X, Liu T, Li X, Wang M
Infection and Drug Resistance 2023, 16:6061-6077
Published Date: 11 September 2023
The Impact of Immune Cells, Metabolites, Inflammatory Factors, and Circulating Proteins on Atopic Dermatitis: Insights from a Mendelian Randomization Study
Zhou D, Gan G, Song S, Zi C, Bao Y, Hao W, Chen Q
Clinical, Cosmetic and Investigational Dermatology 2024, 17:2999-3011
Published Date: 21 December 2024