Back to Journals » Journal of Inflammation Research » Volume 19
Integrated Bioinformatic Analysis of the Shared Molecular Mechanisms Between Diabetic Nephropathy and Atherosclerosis
Authors Zhang J ![]() , Wang Z, Cao H, Zhang H, Chen J
, Wang Z, Cao H, Zhang H, Chen J
Received 5 September 2025
Accepted for publication 6 January 2026
Published 20 January 2026 Volume 2026:19 560821
DOI https://doi.org/10.2147/JIR.S560821
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Quan Zhang
Juan Zhang,1,* Ziwei Wang,2,* Huimin Cao,1,* Hong Zhang,1 Juan Chen1
1Department of Endocrinology, The Affiliated Huai’an No.1 People’s Hospital of Nanjing Medical University, Huai’an, Jiangsu, 223300, People’s Republic of China; 2Department of General Practice, The Affiliated Huai’an No.1 People’s Hospital of Nanjing Medical University, Huai’an, Jiangsu, 223300, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Juan Chen, Email [email protected] Hong Zhang, Email [email protected]
Background: Increasing evidence suggests that individuals with diabetic nephropathy (DN) are at a significantly higher risk of developing atherosclerosis (AS) compared to the general population. However, the precise mechanisms underlying this association remain unclear. This study aims to explore the shared pathways and potential biomarkers implicated in this complication.
Methods: Microarray data downloaded from the Gene Expression Omnibus (GEO) database were used to identify differentially expressed genes (DEGs) in DN and AS. Weighted gene co-expression network analysis (WGCNA) was applied to identify co-expression modules relevant to DN and AS. We conducted functional pathway enrichment analysis on the shared genes. Support vector machine (SVM) and least absolute shrinkage and selection operator (LASSO) methods were utilized to identify and validate potential diagnostic markers. Additionally, immunoinfiltration analysis was performed to examine the relationship between the core markers of DN and AS and the expression of immunoinfiltrating cells. Finally, blood samples from patients were collected to assess the diagnostic efficacy of PRCP.
Results: The results of common genes analysis showed that immune and inflammatory response particularly cytokine-cytokine receptor interactions may be a common feature in the pathophysiology of DN and AS. Three potential shared diagnostic markers—ID4, RNF213, and PRCP—were identified and validated using SVM and LASSO. Immunoinfiltration analysis revealed that the expressions of ID4, RNF213, and PRCP were associated with variations in immune cell populations. Differential analysis of peripheral blood microarray data indicated that only PRCP was significantly overexpressed in both AS and DN samples. Moreover, qRT-PCR confirmed the increased expression of PRCP in peripheral blood mononuclear cells (PBMCs) samples from patients.
Conclusion: The co-expression of RNF213, PRCP, and ID4 may contribute to the development of AS and DN. PRCP, particularly, shows promise as a diagnostic marker due to its detectability as a protein in peripheral blood.
Keywords: diabetic nephropathy, atherosclerosis, bioinformatics, molecular mechanism, inflammation, immune
Introduction
The increasing prevalence of diabetes mellitus (DM) is an urgent public health concern on a global scale.1,2 By 2030, it is projected that 643 million people worldwide will be living with diabetes, a figure which is expected to rise to 783 million by 2045.2 Current research indicates that DM is a major risk factor for cardiovascular diseases (CVD), and CVD is also the main cause of onset and death in patients with type 2 diabetes mellitus (T2DM).3–7
DN is one of the main microvascular complications of DM.8 Due to long-term hyperglycemia leading to glomerular hyperfiltration and tubulointerstitial damage, it is manifested as increased proteinuria and progressive decline in glomerular filtration rate, which may result in end-stage renal failure.9,10 AS is a disease of the arterial wall characterized by lipid deposition, inflammatory response and fibrosis.11,12 Research has demonstrated a possible connection between DN and AS.3–7,13–16 Atherosclerotic dyslipidemia is a significant factor in the onset and progression of DN.6 Lowering atherosclerotic lipids and lipoproteins can diminish glomerular damage6,13 and slow or stop the decline in eGFR, which can cut down on CVD-related illnesses and deaths.3,14 Conversely, DN patients experience worsened plasma dyslipidemia,15 and the combination of proteinuria and renal dysfunction can raise the likelihood of atherosclerotic cardiovascular disease (ASCVD) in diabetic populations.16 These findings deepen our comprehension of the connection between blood lipids and renal dysfunction and could aid in the discovery of molecular mechanisms between two diseases.
Cytokines are a group of small molecules ranging from 8–80 kDa that act as mediators in various pathophysiological responses. This group includes interleukin, interferon, tumor necrosis factor, growth factor, colony-stimulating factor, and chemokine. Cytokines not only play a crucial role in regulating the immune response, but they are also involved in the pathogenesis of inflammatory disorders. Hemodynamic changes caused by some cytokines lead to an increase in intraglomerular pressure,17 followed by the induction of inflammatory cells to enter the kidney by chemokines, resulting in amplification of immune-mediated damage.18 Finally, factors such as TNF-a can cause direct damage to glomeruli, mesangium, and epithelial cells through apoptosis.18
ID4 is primarily located in the nucleus, RNF213 in the cytoplasm, and PRCP in vesicles, with the latter detectable in plasma as a protein. Previous studies have found that all three factors are involved in the process of vascular endothelial injury, inflammatory response and fibrosis caused by metabolic disorders.19–39 ID4 has been shown to act as a pro-differentiation or anti-differentiation factor in a cell-specific manner,19 regulating fatty acid uptake, triglyceride synthesis/storage, and MSC differentiation in mesenchymal stem cells.20 RNF213 intervenes in angiogenesis, affects fat metabolism and inflammation, and is involved in atherosclerotic plaque formation.21,22 Currently, PRCP is considered to be involved in metabolic disorders through its enzyme activity and peptide conversion, and leukocytes and plasma contribute the most to circulating PRCP activity, in which the close relationship between PRCP-dependent signaling and cytokines seems to play an important role in inflammation.23–26 PRCP regulates feeding and insulin receptor substrate 1 stability, and can induce renal oxidative stress through Ang 1-7-MAS receptor-mediated nitric oxide response.26 While MSH is present in atherosclerotic plaque, PRCP cleasts it to reduce macrophage cholesterol excretion, indirectly inhibit the activation of melanocortin receptor and form foam cells, promoting inflammation and plaque formation.
While there is ample evidence suggesting a connection between AS and DN,3–7,13–16 their co-pathogenesis and pathophysiology remain incompletely understood. As a result, identifying co-diagnostic markers of AS and DN is of immense clinical significance for early intervention in DN patients at risk of AS. To address this, we reviewed published gene expression data in the GEO database and utilized a systems biology approach to investigate shared gene pathways and diagnostic markers between DN and AS. Our goal is to use this data to uncover new potential diagnostic and therapeutic strategies for AS patients with underlying DN. The research flowchart of this research was shown in Figure 1.
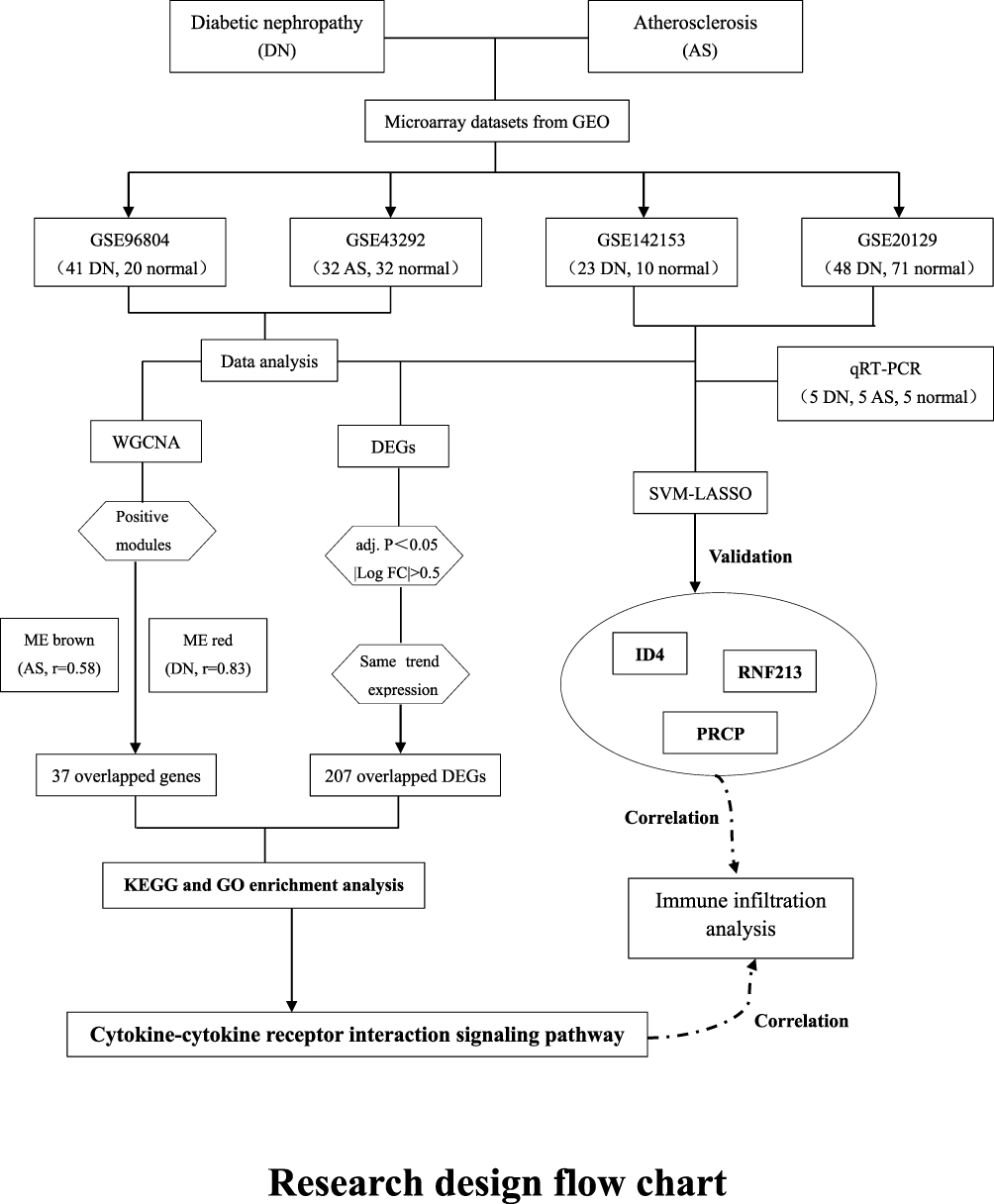 |
Figure 1 Research design flow chart. |
Materials and Methods
Transcriptome Data Preprocessing
We performed an extensive search of the GEO database using the keywords “diabetic nephropathy” and “atherosclerosis”. Our selection criteria required the use of PBMCs or tissues for sequencing. We identified a dataset for the DN group derived from glomeruli (GSE96804, 41 DN samples and 20 normal samples) and a dataset for the AS group derived from atherosclerotic plaques and adjacent tissues (GSE43292, 32 AS samples and 32 normal samples). To enhance our clinical prediction capabilities using the common markers identified, we included two additional datasets based on peripheral blood: GSE142153 (23 DN samples and 10 normal samples) and GSE20129 (48 AS samples and 71 normal samples). For the microarray data, we performed GeneSymbol mapping based on the respective platforms, using the median value when multiple matches were present. The data were then normalized using the log2 (X+1) method to obtain the final expression matrix.
Pre-Screening of Diagnostic Markers
The tissue datasets GSE96804 and GSE43292 underwent differential gene expression screening using the limma package. A truncation standard of P.adj.value <0.05 and |LogFC|>0.5 was applied. For the WGCNA, the entire expression profile of both datasets served as the input matrix, with a soft threshold range of 1 to 30 used for topological calculation to determine the optimal threshold. The topological overlap matrix was then utilized to classify relevant modules, each containing at least 50 genes, and similar modules were merged. Finally, the Pearson method was used to determine the correlation between the merged modules and disease occurrence, selecting the core module with the strongest positive correlation as the outcome.
Screening of Diagnostic Markers
By merging the common DEGs from the GSE96804 and GSE43292 datasets with the intersecting genes identified in the WGCNA, an expression matrix was formed. SVM is a machine learning method that uses feature vectors to identify optimal core genes. LASSO constructs a penalty function to refine the model for optimal gene selection. Disease occurrence was considered a categorical variable, and markers were selected using both SVM and LASSO.
Enrichment Analysis
The DEGs from the GSE96804 and GSE43292 datasets, as well as the intersecting genes identified in the WGCNA, were merged and further enriched using the clusterProfiler package. Gene Ontology (GO) enrichment analysis, a widely utilized bioinformatics technique, was employed to extract extensive insights from the genetic data. Additionally, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment assays were conducted to understand the biological functions and mechanisms. The aforementioned genes also underwent module enrichment analysis using the Metascape database to identify gene clusters with analogous functions.
Localization Analysis of Core Diagnostic Markers
We queried the Human Protein Atlas database for the genes ID4, RNF213, and PRCP to determine their intracellular localization and assess whether they are secreted proteins in U2-OS cells.
Immunoinfiltration Analysis
Utilizing the CIBERSORT algorithm, the ratio of various immune cell types can be determined by assessing the gene expression levels associated with immune cells. The resulting data yields a matrix of immune cell components, based on the output of 22 infiltrated immune cells, which can then be analyzed. To evaluate the correlation between core markers and immune-infiltrating cells, the Spearman method was employed.
Clinical Validation
In our study, 5 patients with DN, 5 patients with AS, and 5 healthy people were recruited from the above hospitals from April 2023 to May 2023, peripheral blood was collected, and PBMCs were extracted from the patients’ blood samples as described below. In this study, each patient signed an informed consent form in advance and was approved by the Ethics Committee of the First People’s Hospital of Huai’an Hospital. Total RNA was extracted using FastPure Cell/Tissue Total RNA Isolation Kit V2 (Vazyme, Nanjing, China) according to the manufacturer’s protocol. cDNA was synthesized from isolated RNA using the HiScript III All-in-one RT SuperMix Perfect for qPCR Kit (Vazyme, Nanjing, China). Taq Pro Universal SYBR qPCR Master Mix Kit (Vazyme, Nanjing, Nanjing, China performed qRT-PCR on the synthesized cDNA in the ABI7500 real-time PCR system (Applied Biosystems). The primer sequences used were PRCP-F: AGCTTCTGCCCCTATCTGG, PRCP-R: GCACTGATTACATTTCCC, GAPDH-F: CAATGTGGACCGCTTTTCCTA, GADPH-R: GGCTGTTGTCATACTTCTCATGG. The results were calculated by the 2−ΔΔCt method.editor.
Moreover, inclusion Criteria for DN group:1 Diagnosis of Type 2 Diabetes Mellitus according to ADA standards;2 Clinical diagnosis of diabetic kidney disease, defined as persistent albuminuria (UACR ≥ 30 mg/g) and/or decreased eGFR (< 60 mL/min/1.73m2) in the absence of other primary renal diseases. Inclusion Criteria for AS group: Patients with ultrasonography or angiography-confirmed carotid and/or coronary atherosclerosis. Inclusion Criteria for Healthy Control group: Individuals with normal glucose tolerance, no history of cardiovascular or renal disease, and normal routine blood and biochemical tests. Exclusion Criteria (applied to all groups):1 Age < 18 or > 80 years;2 Presence of active infection, autoimmune diseases, cancer, or other non-diabetic renal diseases;3 History of organ transplantation. (Table S1. Baseline Clinical Characteristics of the Study Participants).
Results
Differential Genetic Screening
In the DN dataset (Figure 2A), we identified 1199 DEGs. The accompanying volcano plot reveals 438 upregulated and 761 downregulated DEGs (Figure 2B). Similarly, the AS dataset produced 1071 DEGs, comprising 608 upregulated and 403 downregulated genes (Figure 2C and D). Ultimately, the AS and DN datasets revealed 207 overlapping DEGs (Figure 2E).
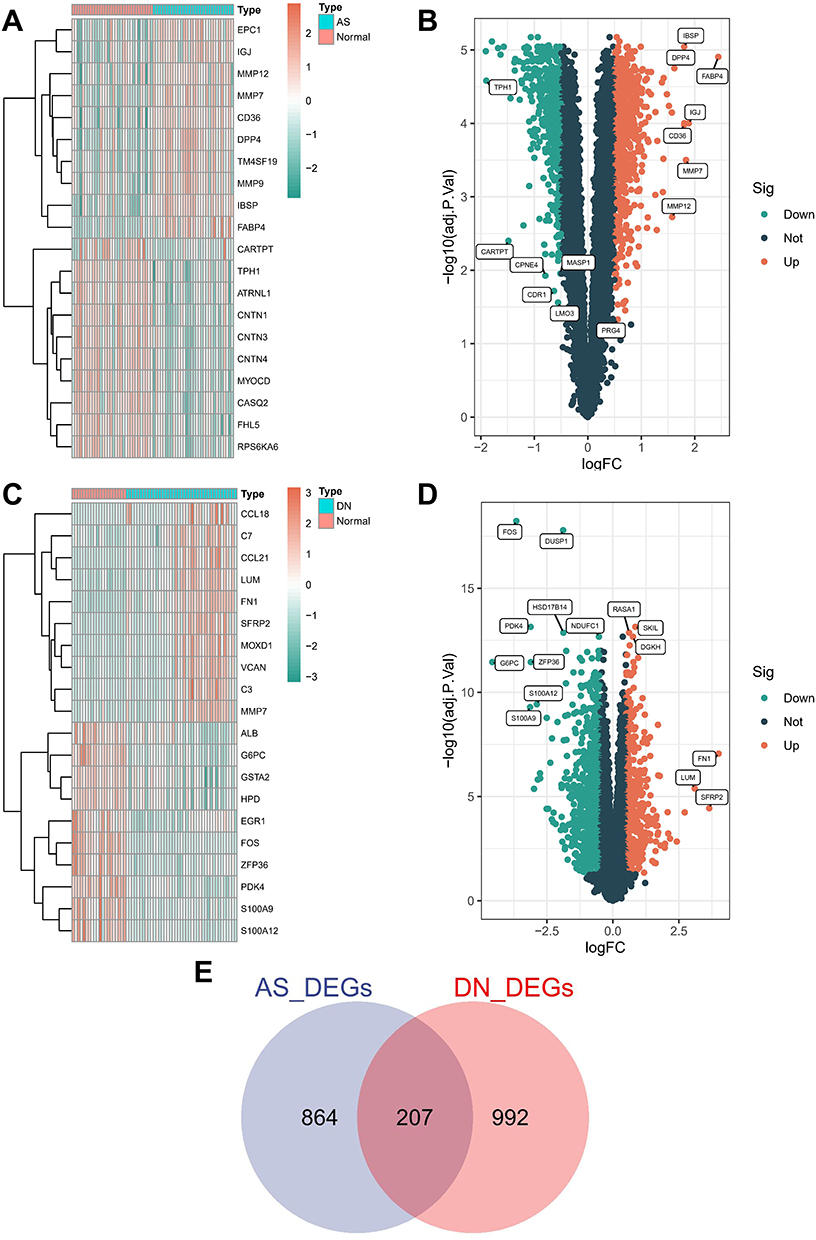 |
Figure 2 Identification of differentially expressed genes in DN and AS. (A) Heatmap of DEGs in the GSE96804 dataset. (B) Volcano plot of DEGs in GSE96804 (P < 0.05). (C) Heatmap of DEGs in the GSE43292 dataset. (D) Volcano plot of DEGs in GSE43292 (P < 0.05). (E) Venn diagram showing overlapping genes between DN and AS. Abbreviation: Sig, significant. |
The identification of 207 overlapping DEGs between the DN and AS datasets underscores the shared genetic landscape of these conditions.
Construction and Module Analysis of WGCNA
Using WGCNA, we explored the relationship between clinical traits and gene expression. Within the AS dataset, one significant outlier was identified and removed (Figure 3A), and two significant outliers were identified and removed from the DN dataset (Figure 3B). The optimal soft threshold for the AS dataset was determined to be 25 (Figure 3C), while the DN dataset’s optimal soft threshold was 5 (Figure 3D). By analyzing the modules’ similarities, we identified 16 modules in both the AS (Figure 3E) and DN datasets (Figure 3F). The brown module exhibited the most robust positive correlation with AS (r=0.58), while the red module had the strongest positive correlation with DN (r=0.83). Ultimately, we found 37 overlapping genes in the WGCNA results (Figure 3G).
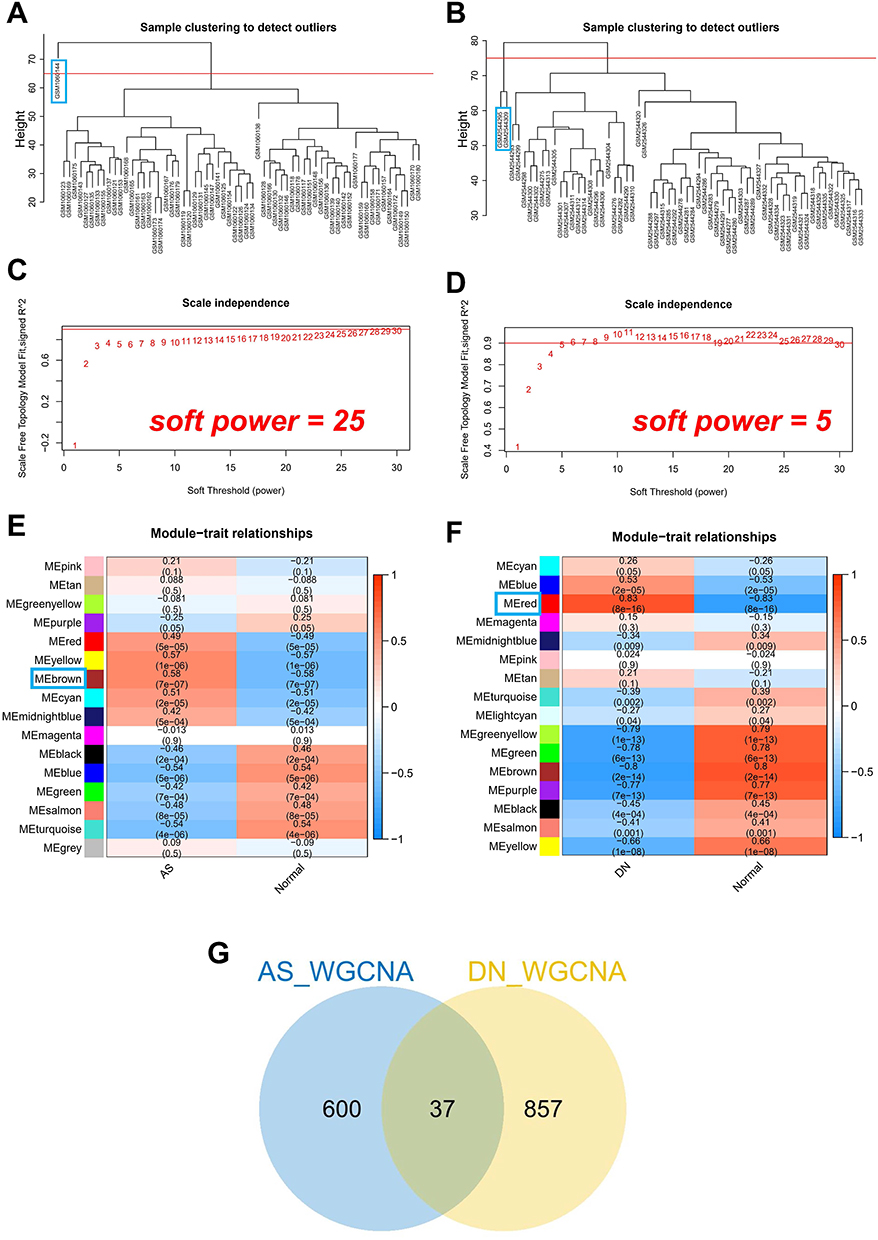 |
Figure 3 Weighted gene co-expression network analysis of DN and AS. (A) Cluster dendrogram of co-expression networks in AS. (B) Cluster dendrogram of co-expression networks in DN. (C) Soft thresholding power for AS. (D) Soft thresholding power for DN. (E) Module-trait correlations in AS. (F) Module-trait correlations in DN. (G) Venn diagram of shared genes between AS and DN. |
WGCNA revealed key gene modules associated with AS and DN, with 37 overlapping genes that may play pivotal roles in both diseases.
Identification of Shared Genes and Shared Pathways
We identified a total of 37 overlapping genes in the DN and AS modules and 207 overlapping DEGs. To ensure comprehensive analysis, we combined the DEGs and module genes for subsequent enrichment analysis. In the KEGG analysis, significant enrichment was observed in pathways such as cytokine-cytokine receptor interaction, cell adhesion molecules, and the Rap1 signaling pathway (Figure 4A). GO enrichment analysis revealed significant enrichment in biological processes such as leukocyte migration, positive regulation of cell adhesion, and leukocyte cell-cell adhesion (Figure 4B). In the cellular component category, significant enrichment was found in the collagen-containing extracellular matrix (ECM), secretory granule membrane, and focal adhesion (Figure 4C). For molecular functions, significant enrichment was observed in amide binding and signaling receptor activator activity (Figure 4D). Using the Metascape database, we enriched these genes into modules, revealing five distinct modules: regulation of cell growth, biological processes related to MHC class II molecules, regulation of plasma lipoprotein particle levels, IL1 and megakaryocytes in obesity, and NABA core matrisome (Figure 4E).
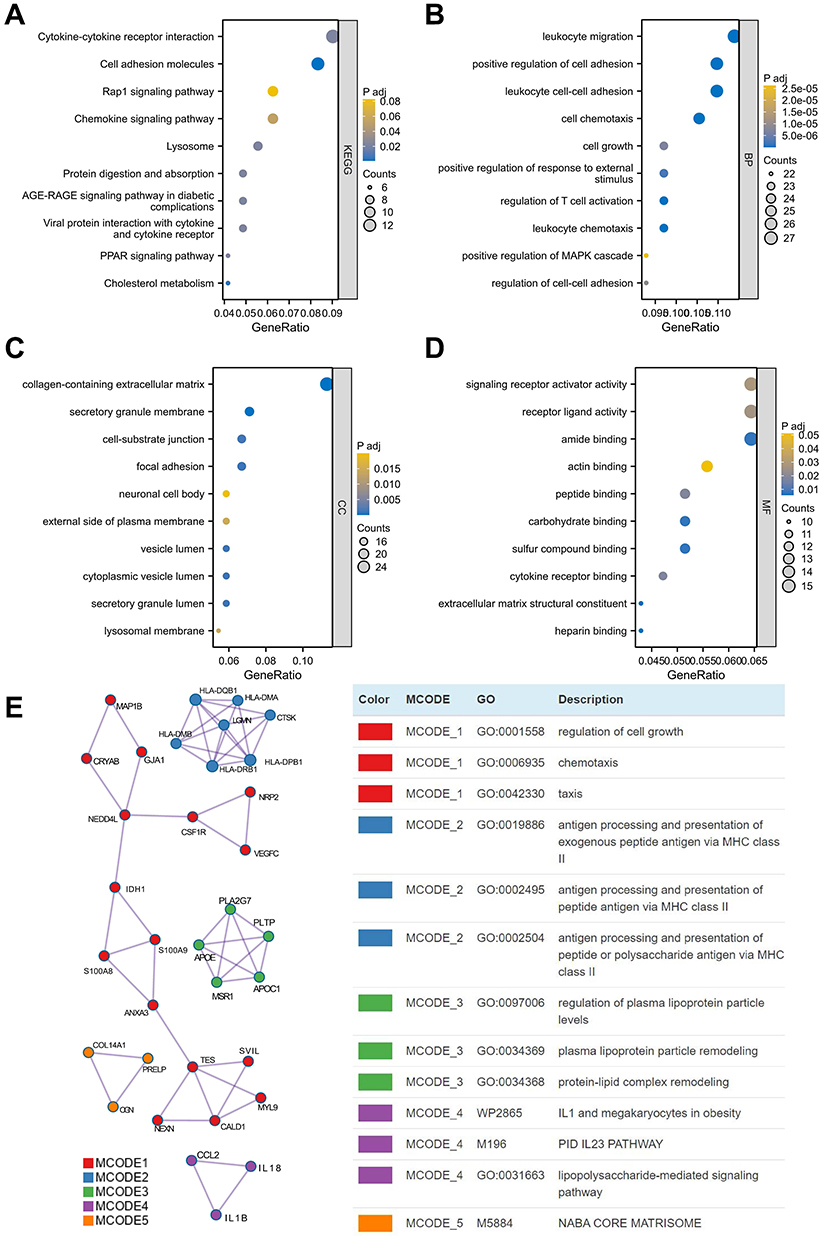 |
Figure 4 Pathway analysis reveals common mechanisms in AS and DN pathogenesis. (A) KEGG pathway enrichment analysis of 37 shared genes identified by WGCNA in AS and DN. (B) Biological process (BP) category of GO enrichment analysis for 37 shared genes. (C) Cellular component (CC) category of GO enrichment analysis for 37 shared genes. (D) Molecular function (MF) category of GO enrichment analysis for 37 shared genes. (E) Module enrichment analysis of DEGs. |
Pathways related to the immune system, particularly characterized by cytokine-cytokine receptor interaction and ECM changes, may represent shared mechanisms for the onset of DN and AS. This could provide insight into why individuals with DN may have an increased risk of developing AS.
Identification and Validation of Potential Shared Diagnostic Markers
To identify the hub genes with the greatest diagnostic value, we utilized machine learning algorithms to select the most significant features. We conducted SVM and LASSO regression analyses on the candidate genes. In the AS dataset, 13 genes were identified using the SVM method (Figure 5A), and 19 genes were identified in the DN dataset (Figure 5B). Additionally, using the LASSO method, we selected 5 genes in the AS dataset (Figure 5C) and 15 genes in the DN dataset (Figure 5D). Comparing the genes selected by different methods in each dataset, we discovered three common diagnostic markers: ID4, RNF213, and prolylcarboxypeptidase (PRCP) (Figure 5E). Localization studies revealed that ID4 is primarily found in the nucleoplasm, RNF213 in the cytosol, and PRCP in vesicles. Notably, PRCP is a detectable protein in plasma that can be utilized for clinical detection (Figure 5F).
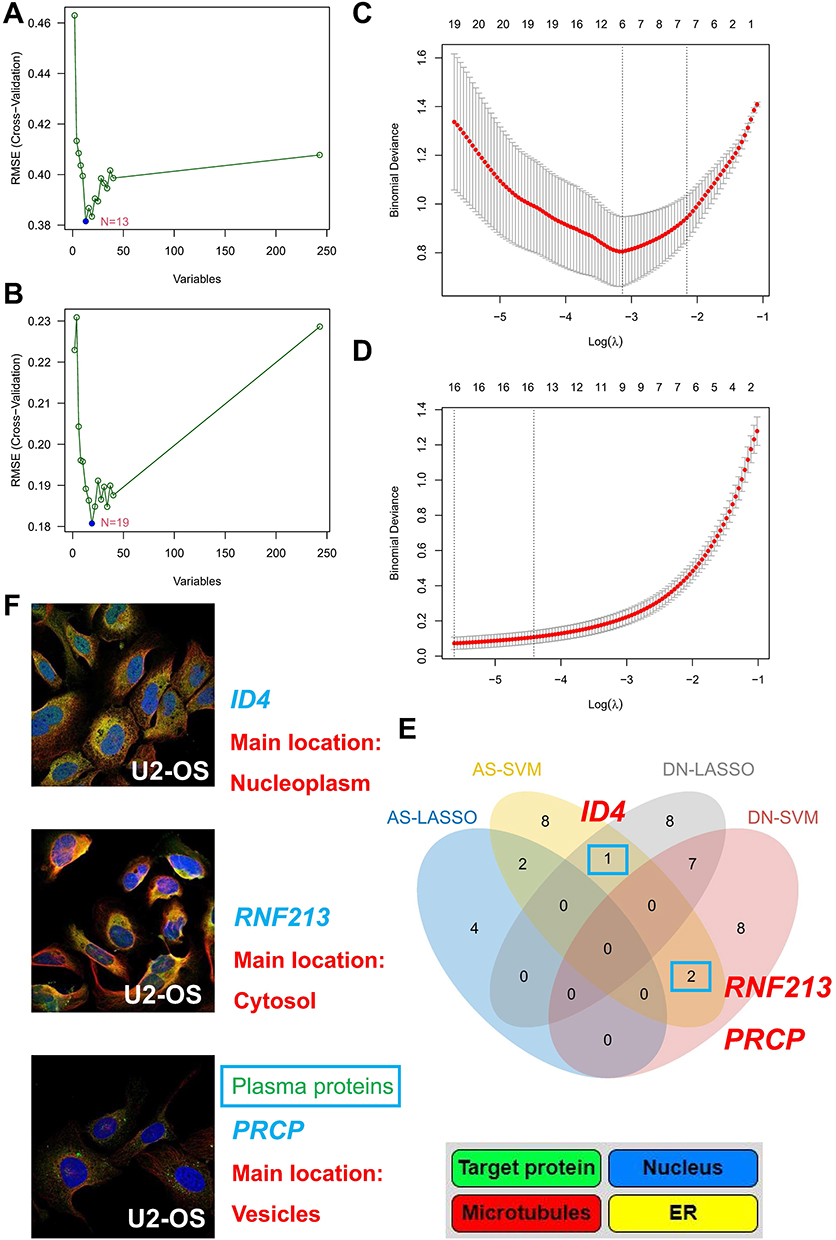 |
Figure 5 Identification and validation of shared diagnostic biomarkers using SVM and LASSO. (A) SVM recursive feature elimination (SVM-RFE) identifies diagnostic markers in AS. (B) SVM-RFE identifies diagnostic markers in DN. (C) LASSO regression identifies diagnostic markers in AS. (D) LASSO regression identifies diagnostic markers in DN. (E) Venn diagram of shared diagnostic biomarkers between DN and AS. (F) Localization of shared diagnostic markers. |
ID4, RNF213, and PRCP may serve as common diagnostic markers for AS and DN. Additionally, PRCP, as a detectable protein in plasma, holds potential for clinical application.
Immunocorrelation Analysis of Shared Diagnostic Markers
Enrichment analysis highlighted the critical role of immunity in the development of both diseases. Therefore, we further investigated the patterns of immune infiltration using the CIBERSORT method, which is based on 22 types of immune cells. Significant differences were found in B cells naive, B cells memory, T cells CD8, T cells CD4 memory activated, T cells regulatory (Tregs), and NK cells activated in AS and control samples, while dendritic cells resting showed notable differences (Figure 6A). In the DN dataset, significant changes were observed in B cells memory, T cells CD8, macrophages M1, macrophages M2, dendritic cells resting, mast cells resting, mast cells activated, and neutrophils (Figure 6B). To further understand the involvement of the three shared hub genes in immune infiltration, we conducted a correlation analysis. ID4 displayed a strong positive correlation with macrophages M0, PRCP was positively correlated with activated NK cells, and RNF213 was positively correlated with resting T cells CD4 in the AS dataset (Figure 6C). In the DN dataset, PRCP also showed positive correlations with plasma cells, resting NK cells, and more (Figure 6D).
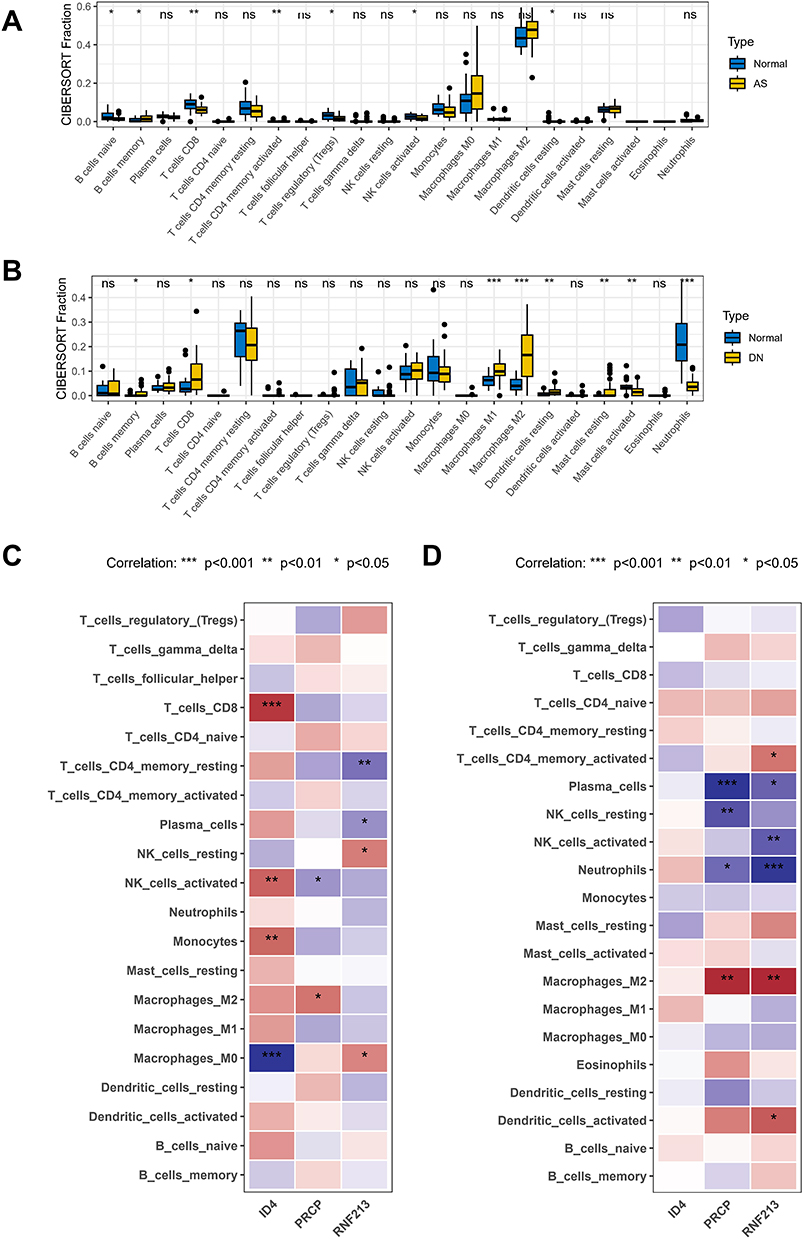 |
Figure 6 Immunocorrelation analysis of shared diagnostic markers in AS and DN. (A) Immune infiltration pattern in AS. (B) Immune infiltration pattern in DN. (C) Immunoinfiltration analysis of shared diagnostic markers in AS datasets. (D) Immunoinfiltration analysis of shared diagnostic markers in DN datasets. Note: All p-values are indicated where significant. *P < 0.05; **P < 0.01; ***P < 0.001. Non-significant results are marked as “ns” if applicable. |
Immune infiltration patterns and correlations with hub genes indicate that immune responses are integral to the pathogenesis of both DN and AS, with specific immune cell types linked to shared diagnostic markers.
Validation of Shared Diagnostic Markers
Analysis of shared markers revealed that ID4 was overexpressed in normal samples, while both RNF213 and PRCP were highly expressed in AS samples in the AS tissue dataset (Figure 7A). This expression trend was also observed in the DN tissue dataset (Figure 7B). To determine the diagnostic efficacy of these markers in peripheral blood sequencing, we performed a differential analysis in peripheral blood microarray data. The results indicated that PRCP was significantly higher in AS and DN samples (Figure 7C and D). ROC analysis in AS and DN cohorts confirmed the strong diagnostic ability of PRCP (Figure 7E and F). Furthermore, qRT-PCR tests on blood samples from our patients demonstrated that PRCP expression in AS samples (Figure 7G) and DN samples (Figure 7H) was significantly higher than in normal samples. According to our findings, the disruption in the co-expression of RNF213, PRCP, and ID4 could potentially contribute to the development of DN and AS. Moreover, results from peripheral blood tests indicate that PRCP may have promising diagnostic capabilities and could be utilized in clinical settings.
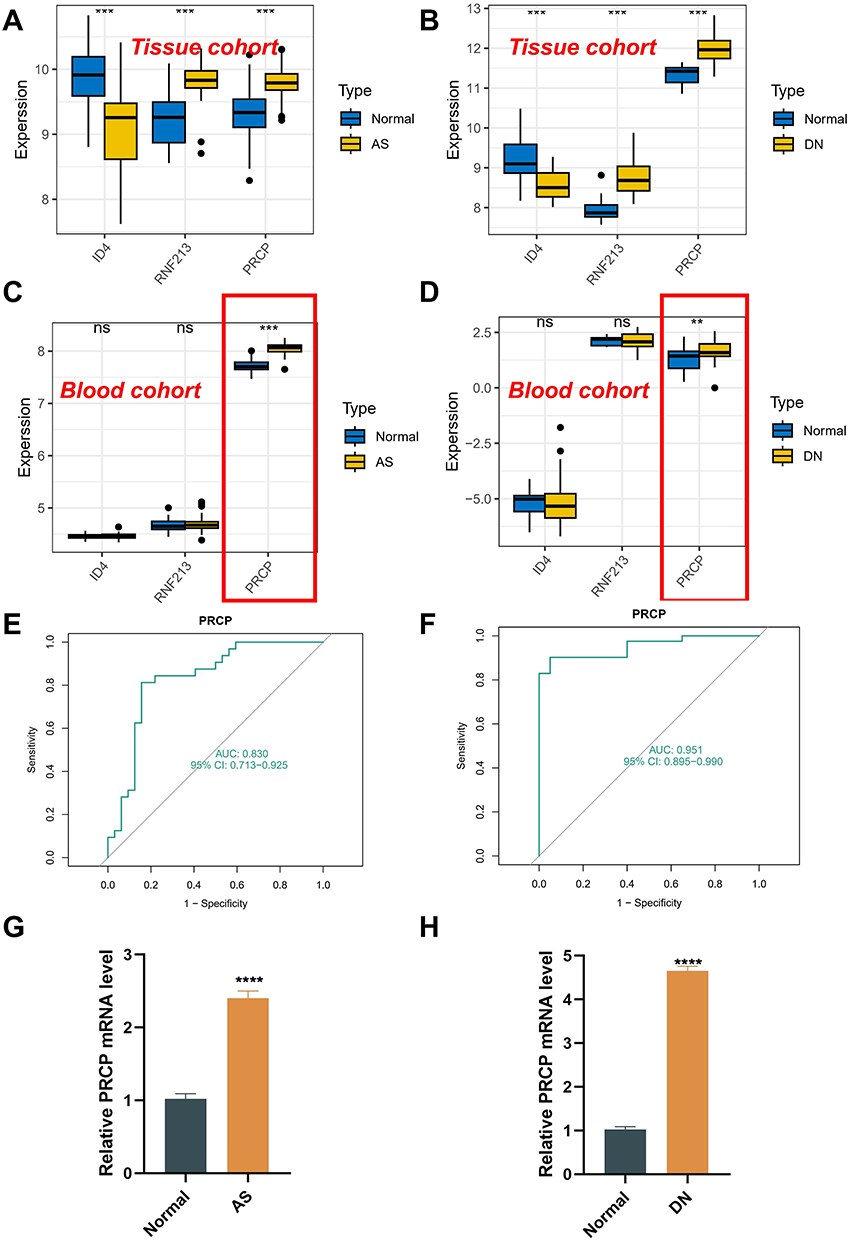 |
Figure 7 Validation of shared diagnostic markers in AS and DN. (A) Shared diagnostic markers show significant expression differences in GSE43292. (B) Shared diagnostic markers show significant expression differences in GSE96804. (C) Shared diagnostic markers show significant expression differences in GSE20129. (D) Shared diagnostic markers show significant expression differences in GSE142513. (E) ROC curve evaluating the diagnostic performance of PRCP in AS. (F) ROC curve evaluating the diagnostic performance of PRCP in DN. (G and H) Real-time PCR analysis of PRCP mRNA expression in PBMCs from patients vs healthy controls. Note: All p-values are indicated where significant. **P < 0.01; ***P < 0.001; ****P < 0.0001. Non-significant results are marked as “ns” if applicable. |
Discussion
This study delves deeper into the genetic level pathophysiological connection between DN and AS. Through the implementation of WGCNA and DEG analysis, we discovered that immune and inflammatory response-related functions may hold significant relevance in the development of DN and AS. Additionally, our enrichment analysis of common genes in public disease databases also revealed the enrichment of cell differentiation and inflammatory signaling pathways. We can infer that an inflammatory environment resulting from immune and inflammatory responses might be a shared characteristic of two diseases, with the Cytokine-cytokine receptor interaction signaling pathway potentially playing a vital role in their co-pathogenesis.
DN is essentially a metabolic disorder manifested by local inflammation of the kidney, characterized primarily by excessive deposition of ECM, resulting in basement membrane thickening, mesangial dilation, and tubulointerstitial fibrosis.7–10 Microinflammation and subsequent ECM dilation are common pathways for the progression of DN, and there is evidence that inflammation is the first promoter before the development of microvascular complications in DM.7,18 Transcription factors such as NF-κB (nuclear factor κB) can be activated by hyperglycemia to regulate genes associated with inflammation and ECM conversion. In DN, proteinuria itself is an important activator of NF-κB and an important pro-inflammatory stimulator of tubule cells.40 In addition, inflammatory cytokines such as IL-1, IL-6, IL-18 and TNF also play a crucial role in the pathogenesis of DN. Oxidative stress produces bioactive molecules, modifies low-density lipoprotein (LDL), and consumes nitric oxide, resulting in endothelial dysfunction, production of various oxidative reactants and free radical diffusion, and ultimately exacerbates DN vascular damage.40 Some immune cells including B and T lymphocytes, macrophages, dendritic cells, mast cells, and neutrophils are significantly altered and enter the kidney to participate in diabetic kidney damage.18 Their activation is usually associated with inflammatory mediators, such as pro-inflammatory cytokines and chemokines, which directly or indirectly affect glycolipid metabolism and promote the development of DN. In the innate immune system, macrophages are the main infiltrating cells in human DN. The adaptive immune system mainly includes T cells and B cells, especially the activation of T cells in the blood and the increase in the number of CD4+T cells in the kidney are closely related to the progression of DN, among which CD4+T cells can further differentiate into Th1, Th2, Th17, Treg cells and other cells involved in immune response and secretion of most cytokines.7–10,18,40
The researchers found that macrophages are present in the glomeruli and interstitium of all stages of DN through kidney biopsy analysis.18,40 This is closely related to serum creatinine and tissue fibrosis, and contributes to the progression of kidney disease by accumulating in the kidney. Macrophages also produce TNF-α, which has been shown to increase in the kidney of DN experimental animal models.18 The loss of macrophage TNF-α leads to a decrease in hypertrophy, proteinuria, and glomerular pathology. M1 macrophages secrete pro-inflammatory factors that exacerbate kidney cell damage, while M2 macrophages promote epithelial and vascular repair. The differentiation of M2 macrophages is key to immunology-based treatment of DN. Activated T cells can directly or indirectly cause damage to kidneys, further inducing inflammation and oxidative stress in DN through the secretion of factors such as IL-2 and IFN-γ. In response to ongoing immune injury, inflammatory mediators are produced by mesangial cells, endothelial cells, and podocytes, resulting in persistent chronic inflammation and leading to renal structure remodeling and tubulointerstitial fibrosis.7,18,40
Foam cells are the hallmark of AS, and atherosclerotic plaque is the main pathological basis of CVD. The accumulation of lipid molecules, specifically LDL, under the arterial wall’s endothelium is the initial stage of AS.11,12,41 This accumulation is taken up by macrophages, which differentiate into foam cells and accumulate intracellular cholesterol, resulting in the activation of innate immune and inflammatory mediators. Cytokines, such as interferon γ and IL-6, can also trigger an inflammatory response by binding to corresponding cytokine receptors, contributing to plaque formation.42 Monocytes are recruited into atherosclerotic plaques with the participation of CCR2 and CCR5, where they differentiate into dendritic cells and macrophages. Macrophages play a key role in all stages of atherosclerotic lesion progression, releasing cytokines, chemokines, and matrix metalloproteinases that break down ECM components and lead to the development of atherosclerotic plaques.43 Pro-inflammatory mediators such as IFN-γ, TNF-α, IL-2, and IL-3 produced by cells such as TH1 and TH2 activate macrophages and other plaque cells, accelerating the inflammatory response. Th1 releases TNF-α and IFN-γ to exert pro-atherosclerotic effects. Th2 cells predominantly interact with B cells and produce IL-4, IL-5, and IL-13. Th17 cells release IL-17A, F, and IL-22, which have pro-inflammatory effects. IL-4 and IL-5 regulate antibody production, while IL-10 inhibits the immune response. TGF-β inhibits both Th1 and Th2 cell activation and is required for Tregs cell differentiation. Its absence increases the entry of pro-inflammatory macrophages and T cells into the arterial wall, exacerbating AS.44 In conclusion, understanding the role of these pro-inflammatory mediators in AS can help us develop new ways to prevent or treat CVD.45
Current research suggests that chemokine signaling pathways, cytosolic DNA sensing pathways, peroxisome proliferator-activated receptor signaling pathways, NF-κB signaling pathways, and cytokine-cytokine receptor interactions all play a significant role in the occurrence, progression, and complications of AS,45 which aligns with our analysis. Upon discovering the abundance of both immune and inflammatory responses in DN and AS, we hypothesized that the immune-related pathway dominated by cytokine-cytokine receptor interaction, coupled with the alteration of the ECM, may be a common pathway for the development of AS and DN. Integrating the gene pool yielded three central genes: ID4, RNF213, and PRCP. The expression of ID4 was reduced, while the expression of the latter two was significantly higher in the DN and AS groups than in the control group, with PRCP expression in peripheral blood expected to be a potential marker for the clinical diagnosis of DN.
The location of ID4 in the 4Mb region of chromosome 6p22.3 and its status as the longest protein in the ID family with 161 residues have been established through studies.19 As a differentiation regulator, ID4 plays a crucial role in adipocyte differentiation, and its reduction can lead to impaired differentiation.20,27 Furthermore, ID4 is positively correlated with Macrophages M0, which accumulate with disease progression and can be differentiated into various forms such as M1, M2, TNF-α, and others, participating in the pathogenic processes of DN and AS. The pathophysiological pathway of Macrophages M0 is believed to cause co-expression dysregulation and pathogenesis. Moving on to RNF213, which is a 591 kDa E3 ubiquitin protein ligase consisting of 5207 amino acids and localized to chromosome 17q25.3.21 Studies have suggested that RNF213 is associated with inflammation and insulin pathways, specifically involved in TNFα-mediated macrophage inflammation and adipocyte inflammation-mediated insulin resistance.22,28 Additionally, RNF213 is commonly associated with the susceptibility gene for moyamoya disease, and its variants such as p. R4810K are known to cause arterial fragility and susceptibility to hemodynamic stress, altering AS and thereby increasing the risk of ASCVD.29–35 In endothelial cells, IFN-γ and TNF-α produced by Th1 and Th2 synergistically activate the transcription of RNF213 in vitro and in vivo. Our study found that RNF213 was positively correlated with T cells CD4 memory resting, and in this active process with unknown mechanism, mediators such as RNF213 and CD4+ T cells, Th1, Th2, and IFN-γ and TNF-α can trigger positive feedback loops, enhance kidney damage, and induce plaque formation, thereby promoting chronic inflammation.
Finally, we found that PRCP was significantly elevated in AS and DN samples and had diagnostic potential in peripheral blood, and was also involved in immune and inflammatory responses. It is a serine protease, located primarily in lysosomes, with the ability to cleat peptide substrates such as angiotensin II and alpha-MSH.23,36,37 Plasma levels of PRCP have been reported to be significantly elevated in patients with AS, inflammation, and DN and are strongly correlated with disease severity.23–26,36–39 Xu, Tabrizian et al suggest that PRCP plasma concentrations can be used to reflect the metabolic status of obese and diabetic individuals while playing a protective role in CVD.24,38,39 Recently, De Hert et al25 observed PRCP activity in human adipose tissue for the first time and confirmed that PRCP is involved in blood glucose and lipid metabolism through immune regulation. In addition, IL-1β and TGF-β1, key regulators of inflammation and fibrosis, have also been shown to be correlated with PRCP, and the reduction of circulating IGF1 is closely related to inflammation and TNF-α activation in endothelial cells. AS and DN data set analysis suggested that PRCP was correlated with immune cells such as NK cells activated, plasma cells and NK cells resting. Therefore, we speculated that PRCP is involved in the development of DN and AS during the differentiation of NK cells. Mψ can participate in NK cell activation either through soluble mediators or cytokines (IL-12, IL-18, TNF-α, CCR7, type I interferon) or through direct cell-cell interactions. Moreover, after M0 and M2 are polarized into M1, M1 will also enhance the activation of NK cells, thus inducing a strong pro-inflammatory response. At the same time, NK cells can also inhibit M1 polarization through IL-10, thereby regulating inflammatory response. It can be seen that PRCP expression is regulated by many cytokines related to inflammatory states and chemokine release, which damage endothelial cells and exacerbate oxidative stress response, thus contributing to DN and AS. Compared with traditional inflammatory indicators such as hs-CRP and IL-6, PRCP can simultaneously reflect the metabolic status of the body. Relying on its unique pathological mechanism, it has more obvious advantages in evaluating disease progression. Moreover, compared with common heart-kidney indicators (eg UACR, eGFR), PRCP integrates multiple factor information such as blood glucose, blood lipids, inflammatory response, and fibrosis, and can more sensitively indicate vascular endothelial injury, having dual significance for diagnosis and treatment. However, current research on PRCP is limited, and a standardized process for its peripheral blood detection protocol has not yet been established. Therefore, the quantitative relationship between its enzyme activity/concentration and disease status has not been established either. We will subsequently conduct in vitro experiments to verify the specific mechanism of action of PRCP, and further expand the sample size of clinical patients to determine its specific role in the cross-reaction between DN and AS.
Conclusion
This groundbreaking study utilized bioinformatics analysis to examine common pathways and genetic diagnostic markers for DN and AS. The findings indicate that the Cytokine-cytokine receptor interaction signaling pathway may play a role in the development of both DN and AS, and could potentially serve as a diagnostic marker for DN complicated by AS. Additionally, the study’s immunoinfiltration correlation analysis suggests that the pathogenesis of these conditions may be linked to an imbalance of ID4, RNF213, and PRCP expression. This research offers a new perspective on exploring the mechanisms underlying DN concurrent AS. However, considering the influence of various factors such AS individual differences, metabolic disorders, the application of hypoglycemic and lipid-lowering drugs and those protecting the cardiovascular and renal systems, as well as the presence of other complications, it remains quite challenging to identify candidates and shared pathways warranting validation in DN and AS, and to establish a multi-dimensional comprehensive evaluation system. Future studies will delve deeper into the Cytokine-cytokine receptor interaction pathway and the expression of ID4, RNF213, and PRCP, as well as conduct related in vitro experiments. At the same time, expand the clinical sample size, establish standardized detection techniques through multi-center cohort studies, and gradually promote its clinical transformation pathways.
Abbreviations
DN, diabetic nephropathy; AS, atherosclerosis; GEO, Gene Expression Omnibus; DEGs, differentially expressed genes; WGCNA, weighted gene co-expression network analysis; SVM, Support vector machine; LASSO, least absolute shrinkage and selection operator; PBMCs, peripheral blood mononuclear cells; DM, diabetes mellitus; CVD, Cardiovascular disease; T2DM, type 2 diabetes mellitus; ASCVD, atherosclerotic cardiovascular disease; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; ECM, extracellular matrix; Tregs, T cells regulatory; NF-κB, nuclear factor κB; LDL, low-density lipoprotein.
Data Sharing Statement
Data availability statement The datasets presented in this study can be found in online repositories (http://www.ncbi.nlm.nih.gov/geo/). The names of the repository/repositories and accession number(s) can be found in the article.
Ethics Statement
The studies involving human participants were reviewed and approved by The Ethics Committee of the Affiliated Huai’an No.1 People’s Hospital with Nanjing Medical University (KY-2023-031-01). The patients/ participants provided their written informed consent to participate in this study. Furthermore, the parts of this research involving humans (individuals, samples or data) were conducted strictly in accordance with the principles stipulated in the Declaration of Helsinki.
Author Contributions
Juan Zhang: Writing – original draft, Project administration, Data curation, Validation. Ziwei Wang: Conceptualization, Investigation. Huimin Cao: Formal analysis, Software – Programming. Hong Zhang: Writing – review and editing, Supervision, Funding acquisition, Methodology, Resources. Juan Chen: Writing – review and editing, Funding acquisition, Methodology, Project administration, Supervision. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was supported by the National Natural Science Foundation of China (Grant Award: 82000743), Medical Research General Program of Jiangsu Provincial Health Commission (Grant Award: M2024061), Northern Jiangsu Clinical Medicine Research Institute’ s 2024 Projects (Grant Award: HAKY20240000307), Key Research and Development Program of Jiangsu Provincial Department of Science and Technology - Social Development Project (Grant Award: BE2023745), General Project of the Health Commission of Jiangsu Province (Grant Award: H2023137) and General Project of Natural Science Foundation of Jiangsu Provincial Department of Science and Technology (Grant Award: BK20191213).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Bommer C, Sagalova V, Heesemann E, et al. Global economic burden of diabetes in adults: projections from 2015 to 2030. Diabetes Care. 2018;41(5):963–17. doi:10.2337/dc17-1962
2. GBD 2021 Diabetes Collaborators. Global Burden of Disease (GBD) Study 2021 analysis of diabetes prevalence, burden, and projections. Lancet. 2023. doi:10.1016/S0140-6736(23)01301-6
3. Taheri S. Heterogeneity in cardiorenal protection by Sodium glucose cotransporter 2 inhibitors in heart failure across the ejection fraction strata: systematic review and meta-analysis. World J Nephrol. 2023;12(5):182–200. doi:10.5527/wjn.v12.i5.182
4. Zhao BR, Hu XR, Wang WD, Zhou Y. Cardiorenal syndrome: clinical diagnosis, molecular mechanisms and therapeutic strategies. Acta Pharmacol Sin. 2025;46(6):1539–1555. doi:10.1038/s41401-025-01476-z
5. Zhu T, Du Y, Xuan M, Guo C, Rao X. Clinical characteristics and Chinese medicine therapy of chronic kidney disease combined with cardiovascular disease. Integrat Med Nephrol Androl. 2023;10(1):e00023. doi:10.1097/IMNA-D-22-00023
6. Zou TF, Liu ZG, Cao PC, et al. Fisetin treatment alleviates kidney injury in mice with diabetes-exacerbated atherosclerosis through inhibiting CD36/fibrosis pathway. Acta Pharmacol Sin. 2023;44(10):2065–2074. doi:10.1038/s41401-023-01106-6
7. Ren LL, Miao H, Wang YN, Liu F, Li P, Zhao YY. TGF-β as A master regulator of aging-associated tissue fibrosis. Aging and Disease. 2023;14(5):1633–1650. doi:10.14336/AD.2023.0222
8. Młynarska E, Buławska D, Czarnik W, et al. Novel insights into diabetic kidney disease. Int J Mol Sci. 2024;25(18):10222. doi:10.3390/ijms251810222
9. Gupta S, Dominguez M, Golestaneh L. Diabetic Kidney Disease: an Update. Med Clin North Am. 2023;107(4):689–705. doi:10.1016/j.mcna.2023.03.004
10. Wang Y, Jin M, Cheng CK, Li Q. Tubular injury in diabetic kidney disease: molecular mechanisms and potential therapeutic perspectives. Front Endocrinol. 2023;14:1238927. doi:10.3389/fendo.2023.1238927
11. Libby P. The changing landscape of atherosclerosis. Nature. 2021;592(7855):524–533. doi:10.1038/s41586-021-03392-8
12. Monaco C, McNamara CA, Slütter B, LeDucq International Network of Excellence on Immune Checkpoints in Atherosclerosis: “CHECKPOINT ATHERO”, et al. Immunotherapy for atherosclerosis. Physiol Rev. 2025;105(4):2141–2230. doi:10.1152/physrev.00016.2024
13. Sasaki T, Kurata H, Nomura K, Utsunomiya K, Ikeda Y. Amelioration of proteinuria with pravastatin in hypercholesterolemic patients with diabetes mellitus. Jpn J Med. 1990;29(2):156–163. doi:10.2169/internalmedicine1962.29.156
14. Athyros VG, Mitsiou EK, Tziomalos K, Karagiannis A, Mikhailidis DP. Impact of managing atherogenic dyslipidemia on cardiovascular outcome across different stages of diabetic nephropathy. Expert Opin Pharmacother. 2010;11(5):723–730. doi:10.1517/14656560903575654
15. Sacks FM, Hermans MP, Fioretto P, et al. Association between plasma triglycerides and high‐density lipoprotein cholesterol and microvascular kidney disease and retinopathy in type 2 diabetes mellitus: a global case‐control study in 13 countries. Circulation. 2014;129:999–1008.
16. Hirano T, Satoh N, Kodera R, et al. Dyslipidemia in diabetic kidney disease classified by proteinuria and renal dysfunction: a cross-sectional study from a regional diabetes cohort. J Diabetes Invest. 2022;13(4):657–667. doi:10.1111/jdi.13697
17. Tuttle KR, Agarwal R, Alpers CE, et al. Molecular mechanisms and therapeutic targets for diabetic kidney disease. Kidney Int. 2022;102(2):248–260. doi:10.1016/j.kint.2022.05.012
18. Navarro JF, Mora-Fernández C. The role of TNF-alpha in diabetic nephropathy: pathogenic and therapeutic implications. Cytokine Growth Factor Rev. 2006;17(6):441–450. doi:10.1016/j.cytogfr.2006.09.011
19. Patel D, Morton DJ, Carey J, Havrda MC, Chaudhary J. Inhibitor of differentiation 4 (ID4): from development to cancer. BBA. 2015;1855(1):92–103. doi:10.1016/j.bbcan.2014.12.002
20. Palatil M, Sharma BK, Satyanarayana A. Id transcription regulators in adipogenesis and adipose tissue metabolism. Front Biosci. 2014;19(8):1386–1397. doi:10.2741/4289
21. Pollaci G, Gorla G, Potenza A, et al. Novel multifaceted roles for RNF213 protein. Int J Mol Sci. 2022;23(9):4492. doi:10.3390/ijms23094492
22. Sarkar P, Thirumurugan K. New insights into TNFα/PTP1B and PPARγ pathway through RNF213- a link between inflammation, obesity, insulin resistance, and moyamoya disease. Gene. 2021;771:145340. doi:10.1016/j.gene.2020.145340
23. Nguyen BY, Zhou F, Binder P, et al. Prolylcarboxypeptidase alleviates hypertensive cardiac remodeling by regulating myocardial tissue angiotensin II. J Am Heart Assoc. 2023;12(12):e028298. doi:10.1161/JAHA.122.028298
24. Xu S, Lind L, Zhao L, Lindahl B, Venge P. Plasma prolylcarboxypeptidase (angiotensinase C) is increased in obesity and diabetes mellitus and related to cardiovascular dysfunction. Clin Chem. 2012;58(7):1110–1115. doi:10.1373/clinchem.2011.179291
25. De Hert E, Verboven K, Wouters K, Jocken JWE, De Meester I. Prolyl carboxypeptidase activity is present in human adipose tissue and is elevated in serum of obese men with type 2 diabetes. Int J Mol Sci. 2022;23(21):13529. doi:10.3390/ijms232113529
26. Rinne P, Lyytikäinen LP, Raitoharju E, et al. Pro-opiomelanocortin and its processing enzymes associate with plaque stability in human atherosclerosis - Tampere Vascular Study. Sci Rep. 2018;8(1):15078. doi:10.1038/s41598-018-33523-7
27. Murad JM, Place CS, Ran C, et al. Inhibitor of DNA binding 4 (ID4) regulation of adipocyte differentiation and adipose tissue formation in mice. J Biol Chem. 2010;285:24164–24173.
28. Kobayashi H, Yamazaki S, Takashima S, et al. Ablation of Rnf213 retards progression of diabetes in the Akita mouse. Biochem Biophys Res Commun. 2013;432(3):519–525. doi:10.1016/j.bbrc.2013.02.015
29. Ahmed S, Habu T, Kim J, et al. Suppression of RNF213, a susceptibility gene for moyamoya disease, inhibits endoplasmic reticulum stress through SEL1L upregulation. Biochem Biophys Res Commun. 2022;609:62–68. doi:10.1016/j.bbrc.2022.04.007
30. Liu M, Gutierrez J. Genetic risk factors of intracranial atherosclerosis. Curr Atherosclerosis Rep. 2020;22(4):13. doi:10.1007/s11883-020-0831-5
31. Kim HJ, Choi EH, Chung JW, et al. Luminal and wall changes in intracranial arterial lesions for predicting stroke occurrence. Stroke. 2020;51(8):2495–2504. doi:10.1161/STROKEAHA.120.030012
32. Kim HJ, Choi EH, Chung JW, et al. Role of the RNF213 variant in vascular outcomes in patients with intracranial atherosclerosis. J Am Heart Assoc. 2021;10(1):e017660. doi:10.1161/JAHA.120.017660
33. Ohara M, Yoshimoto T, Okazaki S, et al. RNF213 p.R4810K variant carriers with intracranial arterial stenosis have a low atherosclerotic burden. J Atherosclerosis Thrombosis. 2022;29(11):1655–1662. doi:10.5551/jat.63379
34. Iwanishi M, Azuma C, Tezuka Y, et al. Observation of p.R4810K, a polymorphism of the mysterin gene, the susceptibility gene for moyamoya disease, in two female Japanese diabetic patients with familial partial lipodystrophy 1. Internal Medicine. 2020;59(20):2529–2537. doi:10.2169/internalmedicine.4042-19
35. He XW, Zhao Y, Shi YH, et al. DNA methylation analysis identifies differentially methylated sites associated with early-onset intracranial atherosclerotic stenosis. J Atherosclerosis Thrombosis. 2020;27(1):71–99. doi:10.5551/jat.47704
36. Duan L, Calhoun SJ, Perez RE, et al. Prolylcarboxypeptidase promotes IGF1R/HER3 signaling and is a potential target to improve endocrine therapy response in estrogen receptor positive breast cancer. Cancer Biol Ther. 2022;23(1):1–10. doi:10.1080/15384047.2022.2142008
37. De Hert E, Bracke A, Pintelon I, et al. Prolyl carboxypeptidase mediates the C-terminal cleavage of (Pyr)-Apelin-13 in human umbilical vein and aortic endothelial cells. Int J Mol Sci. 2021;22(13):6698. doi:10.3390/ijms22136698
38. Tabrizian T, Hataway F, Murray D, Shariat-Madar Z. Prolylcarboxypeptidase gene expression in the heart and kidney: effects of obesity and diabetes. Cardiovasc Hematoll Agents Med Chem. 2015;13(2):113–123. doi:10.2174/1871525713666150911112916
39. Tabrizian T, Floyd L, Shariat-Madar Z. Altered prolylcarboxypeptidase expression and function in response to different risk factors of diabetes. Cardiovasc Hematoll Agents Med Chem. 2017;14(3):175–189. doi:10.2174/1871525714666161229130508
40. Wolf G. New insights into the pathophysiology of diabetic nephropathy: from haemodynamics to molecular pathology. Eur J Clin Invest. 2004;34(12):785–796. doi:10.1111/j.1365-2362.2004.01429.x
41. Bentzon JF, Otsuka F, Virmani R, Falk E. Mechanisms of plaque formation and rupture. Circ Res. 2014;114(12):1852–1866. doi:10.1161/CIRCRESAHA.114.302721
42. Verma S, Kumar A, Narang R, Bisoi AK, Mitra DK. Signature transcriptome analysis of stage specific atherosclerotic plaques of patients. BMC Med Genomics. 2022;15(1):99. doi:10.1186/s12920-022-01250-8
43. Kotlyarov S. Analysis of differentially expressed genes and signaling pathways involved in atherosclerosis and chronic obstructive pulmonary disease. Biomol Concepts. 2022;13(1):34–54. doi:10.1515/bmc-2022-0001
44. Tabares-Guevara JH, Villa-Pulgarin JA, Hernandez JC. Atherosclerosis: immunopathogenesis and strategies for immunotherapy. Immunotherapy. 2021;13(14):1231–1244. doi:10.2217/imt-2021-0009
45. Hridoy HM, Haidar MN, Khatun C, et al. In silico based analysis to explore genetic linkage between atherosclerosis and its potential risk factors. Biochem Biophys Rep. 2023;36:101574. doi:10.1016/j.bbrep.2023.1015746
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Sacubitril/Valsartan Improves Progression of Early Diabetic Nephropathy in Rats Through Inhibition of NLRP3 Inflammasome Pathway
Pan Y, Liu L, Yang H, Chen W, Chen Z, Xu J
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2479-2488
Published Date: 13 August 2022
Study on Relationship Between Carotid Intima-Media Thickness and Inflammatory Factors in Obstructive Sleep Apnea
Ji P, Kou Q, Zhang J
Nature and Science of Sleep 2022, 14:2179-2187
Published Date: 14 December 2022
α-Mangostin Inhibited M1 Polarization of Macrophages/Monocytes in Antigen-Induced Arthritis Mice by Up-Regulating Silent Information Regulator 1 and Peroxisome Proliferators-Activated Receptor γ Simultaneously

Wu YJ, Zhang SS, Yin Q, Lei M, Wang QH, Chen WG, Luo TT, Zhou P, Ji CL
Drug Design, Development and Therapy 2023, 17:563-577
Published Date: 23 February 2023
The Involvement of Glucose and Lipid Metabolism Alteration in Rheumatoid Arthritis and Its Clinical Implication
Luo TT, Wu YJ, Yin Q, Chen WG, Zuo J
Journal of Inflammation Research 2023, 16:1837-1852
Published Date: 26 April 2023
Transcriptomics Analysis Revealed Key Genes Associated with Macrophage Autophagolysosome in Male ApoE−/− Mice Aortic Atherosclerosis
Zhu M, Jin T, Wu D, Zhang S, Wang A
Journal of Inflammation Research 2023, 16:5125-5144
Published Date: 9 November 2023