Back to Journals » Drug Design, Development and Therapy » Volume 19
Glycolytic Dysfunction in Granulosa Cells and Its Contribution to Metabolic Dysfunction in Polycystic Ovary Syndrome
Authors Cao Z, Zhou Q, An J ![]() , Guo X, Jia X, Qiu Y
, Guo X, Jia X, Qiu Y
Received 1 March 2025
Accepted for publication 6 June 2025
Published 18 June 2025 Volume 2025:19 Pages 5255—5270
DOI https://doi.org/10.2147/DDDT.S525651
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Yan Zhu
Zhenzhen Cao,1,* Qin Zhou,1,* Jie An,1,* Xiaojing Guo,1 XiaoFang Jia,1 Yuena Qiu2
1Department of Gynecology, Kunshan Hospital of Traditional Chinese Medicine, Kunshan, Jiangsu, 215300, People’s Republic of China; 2Department of Reproductive Medicine, The Second Affiliated Hospital of Fujian Medical University, Quanzhou, 362000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yuena Qiu, Email [email protected]
Abstract: Polycystic ovary syndrome (PCOS) is a common endocrine-metabolic disorder in women of reproductive age, marked by hyperandrogenism, ovulatory dysfunction, and insulin resistance, accompanied by significant metabolic disturbances, including glycolytic dysfunction, mitochondrial impairment, and increased oxidative stress. In granulosa cells (GCs), disrupted glycolysis impairs follicular development and compromises oocyte quality, exacerbating reproductive and metabolic abnormalities. At the molecular level, dysregulated energy-sensing pathways, such as AMPK and mTOR, reduce glucose uptake, lower ATP generation, and enhance oxidative stress, fueling disease progression. Epigenetic changes and non-coding RNAs further modulate glycolytic enzyme expression, destabilizing metabolic homeostasis within ovarian follicles. Therapeutically, restoring glycolytic balance using agents like metformin, resveratrol, mogroside V, and nicotinamide mononucleotide (NMN) has shown promise in improving glycolysis, insulin sensitivity, and ovarian function in various models. This review synthesizes current evidence on glycolysis’s critical role in PCOS pathophysiology, its influence on follicular energetics and oocyte quality, and highlights metabolic targets for future therapies, offering a foundation for novel mechanism-driven interventions in PCOS management.
Keywords: polycystic ovary syndrome, glycolysis, metabolic dysfunction, insulin resistance, granulosa cells, oxidative stress, metabolic therapy
Introduction
Polycystic Ovary Syndrome (PCOS) is a prevalent endocrine-metabolic disorder that primarily affects women of reproductive age, characterized by menstrual irregularities, ovulatory dysfunction, and hyperandrogenemia. According to the Rotterdam criteria, the diagnosis of PCOS relies on meeting at least two of the following three criteria: polycystic ovarian morphology, ovulatory dysfunction, and clinical or biochemical hyperandrogenism.1,2 Epidemiological studies indicate that the prevalence of PCOS varies among different populations, with a global incidence ranging from approximately 5% to 10%, predominantly affecting women aged 20 to 35 years.3 The distribution of PCOS exhibits certain disparities across ethnicities and geographical regions, suggesting that both genetic and environmental factors play a crucial role in its pathogenesis.3,4
PCOS not only impacts female reproductive health, leading to infertility and ovarian cyst formation, but is also closely associated with various endocrine disorders. Patients frequently present with elevated androgen levels, abnormal estrogen metabolism, and insulin resistance, manifesting as hirsutism, acne, and adipose tissue deposition.2,5 Additionally, PCOS significantly increases the risk of developing type 2 diabetes, cardiovascular diseases, and metabolic syndrome.5,6 These endocrine disturbances impose a substantial physiological burden, exacerbating lipid metabolism dysfunction and impairing blood pressure regulation, thereby elevating cardiovascular risk.2,5 Moreover, PCOS may contribute to osteoporosis due to its complex interactions within the endocrine system, further complicating disease management and increasing long-term health risks.5,6
Among the pathological mechanisms of PCOS, energy metabolism dysfunction is considered a key characteristic. Glycolysis, a fundamental pathway for cellular energy production, plays a critical role in PCOS pathogenesis.1,2 This metabolic process converts glucose into pyruvate, generating a limited amount of ATP to meet immediate cellular energy demands.5,7 Also, evidence shows that mitochondrial bioenergetics are compromised in PCOS ovaries: granulosa-cell mitochondria display a lower membrane potential, diminished oxidative‐phosphorylation capacity, and excessive reactive-oxygen‐species production.6,7 This defective mitochondrial state short-circuits ATP output, feeds back to restrain glycolytic flux, and activates redox-sensitive pathways that aggravate insulin resistance and hyper-androgenism, thereby amplifying the metabolic and reproductive derangements characteristic of PCOS.1,6 In summary, as a complex endocrine-metabolic disorder, the etiology and pathophysiology of PCOS involve multiple biological processes. A deeper understanding of the role of glycolysis and mitochondrial energy metabolism in PCOS is essential for elucidating its pathological mechanisms and developing novel therapeutic strategies.2,5
Clinical Manifestations of Polycystic Ovary Syndrome
PCOS is one of the most prevalent disorders among women of reproductive age, with a global prevalence reaching up to 15%.8 PCOS is a hormonal disorder that manifests through a variety of symptoms, including hair loss, hirsutism (excessive hair growth), pelvic pain, infertility, and irregular menstrual cycles.9,10 Additionally, it is frequently associated with symptoms such as overweight, fatigue, and acne, which are often linked to hormonal imbalances, particularly elevated testosterone levels. Many women with PCOS exhibit high androgen levels, contributing to hair thinning and acne.9,11 These symptoms, along with irregular menstrual cycles and ovulatory dysfunction, serve as the clinical foundation for PCOS diagnosis.
In PCOS patients, ovarian structure and function undergo significant alterations. The ovaries often exhibit multiple small cysts, typically measuring 2–9 mm in diameter, which represent immature follicles that fail to undergo normal maturation and ovulation.12,13 This leads to increased ovarian volume and a characteristic “string of pearls” appearance on ultrasound imaging.14 Functionally, ovarian activity is disrupted by hormonal imbalances, particularly elevated androgen levels, which impair normal follicular development and ovulation. Disrupted folliculogenesis and anovulation contribute to the irregular menstrual cycles commonly observed in PCOS. Additionally, alterations in ovarian steroidogenesis and androgen levels further impact both ovarian morphology and function (Figure 1).
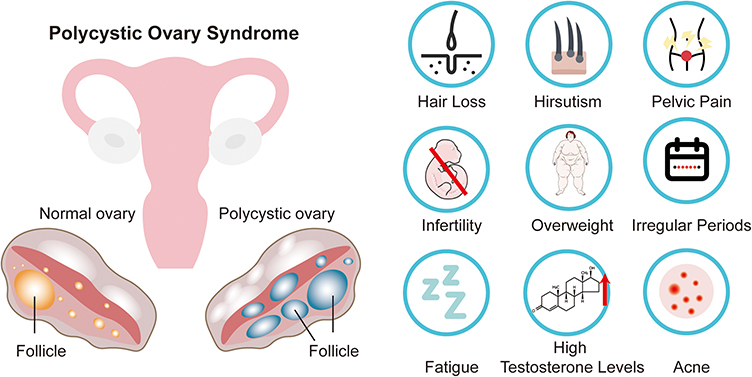 |
Figure 1 Ovarian changes and clinical symptoms in PCOS patients. |
Moreover, PCOS is frequently associated with insulin resistance and obesity, which contribute to reproductive and metabolic abnormalities across the lifespan, leading to severe conditions such as anovulatory infertility and type 2 diabetes (T2D).15,16 Despite decades of research, the precise etiology of PCOS remains unclear. Familial clustering suggests that genetic factors play a crucial role in its pathogenesis.17,18 PCOS typically follows a non-Mendelian inheritance pattern, reflecting a complex genetic architecture similar to that of T2D and obesity, involving interactions between susceptibility genes and environmental factors.19 Genome-wide association studies (GWAS) have identified 19 PCOS-associated genetic loci in East Asian and European populations, with 12 of these loci replicated across multiple studies. These loci include genes such as FSHB, LHCGR, FSHR, INSR, and DENND1A, which are involved in gonadotropin secretion, androgen biosynthesis, metabolic regulation, and follicular development.19–24 Furthermore, genetic correlations between PCOS and various metabolic and reproductive traits, including body mass index (BMI), insulin levels, T2D, high-density lipoprotein (HDL) levels, triglyceride levels, depression, and age at menarche, have been reported.23,25–29 These findings suggest that BMI and insulin resistance play causal roles in PCOS development, whereas PCOS itself does not directly increase the risk of T2D or cardiovascular diseases.23,25,27,28,30
Epigenetic studies have revealed alterations in DNA methylation and gene expression in ovarian granulosa cells, adipose tissue, and skeletal muscle of PCOS patients. These changes may be driven by a combination of genetic variants and environmental exposures.31–36 Furthermore, PCOS is closely associated with glycolytic dysregulation in granulosa cells, characterized by reduced glucose uptake and utilization due to insulin resistance.37 Altered expression of glucose transporters (GLUT) further restricts glucose availability,38 while mitochondrial dysfunction leads to energy deficits and oxidative stress.1 Transcriptomic sequencing of ovarian granulosa cells and follicular fluid-derived exosomes has revealed significant alterations in the expression of energy metabolism-related genes and non-coding RNAs, suggesting that ovarian cellular metabolism undergoes substantial changes in PCOS.39
Glycolysis
Glycolysis is one of the most fundamental and essential metabolic pathways in cells, responsible for converting glucose into pyruvate while generating a small amount of energy in the form of ATP. This process occurs in the cytoplasm and does not require oxygen, making it an anaerobic metabolic pathway. However, under aerobic conditions, glycolysis serves as the initial step in a broader aerobic metabolic network, linking the metabolism of carbohydrates, lipids, and proteins40,41 (Figure 2).
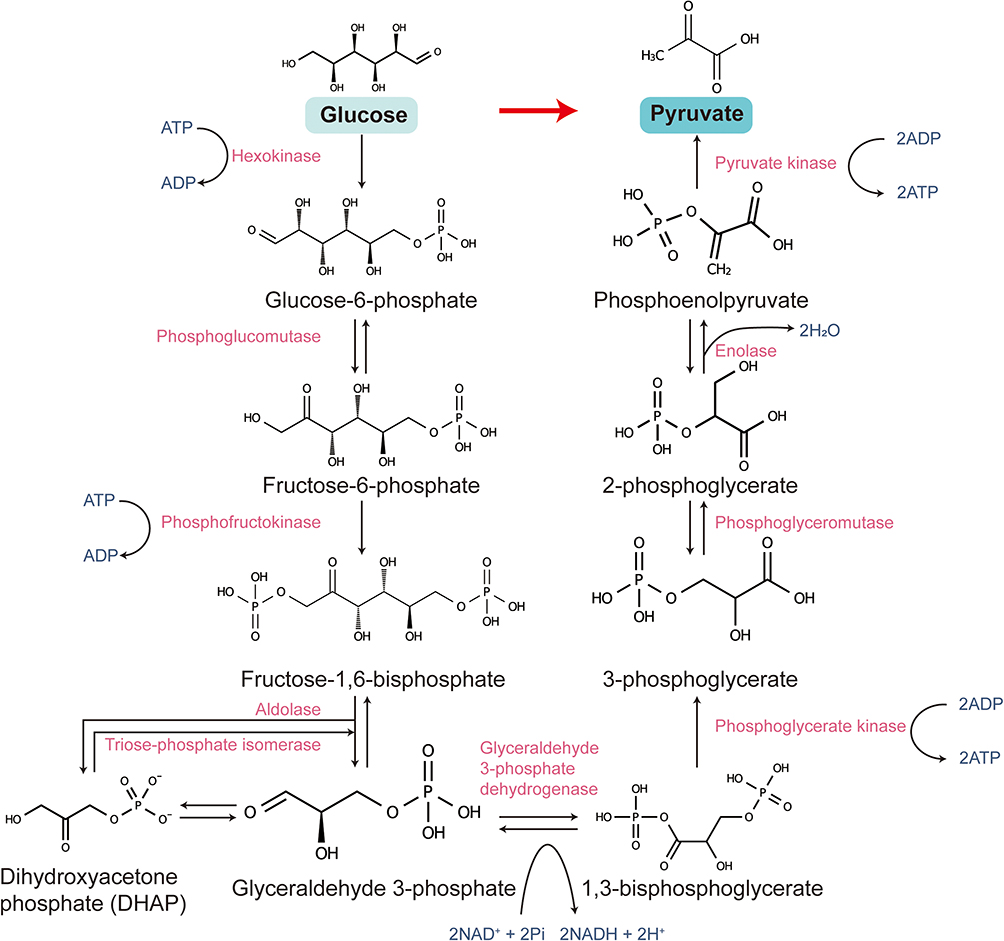 |
Figure 2 Biochemical pathway of glycolysis. |
Glycolysis consists of ten consecutive enzymatic reactions, which can be divided into the energy investment phase and the energy payoff phase. In the energy investment phase, glucose is first phosphorylated by hexokinase (HK) to form glucose-6-phosphate, which then undergoes isomerization to generate fructose-6-phosphate (F6P). Subsequently, phosphofructokinase-1 (PFK-1) catalyzes the conversion of F6P to fructose-1,6-bisphosphate (F1,6BP), a rate-limiting step in glycolysis.42,43 During the energy payoff phase, F1,6BP is cleaved by aldolase into dihydroxyacetone phosphate and glyceraldehyde-3-phosphate (G3P). G3P then undergoes a series of reactions, leading to its oxidation into 1,3-bisphosphoglycerate (1,3-BPG), ultimately producing pyruvate along with ATP and NADH.40,41
The regulation of glycolysis is complex and diverse, primarily achieved through the modulation of enzyme activity. PFK-1, as a key regulatory point of glycolysis, is controlled by various metabolic factors. For instance, AMP and fructose-2,6-bisphosphate act as activators, enhancing PFK-1 activity, whereas ATP and citrate serve as inhibitors, suppressing its function.42,44 Additionally, pyruvate kinase (PK) is another crucial regulatory enzyme in glycolysis, with its activity positively regulated by acetyl-CoA and F1,6BP, while ATP and pyruvate exert negative feedback inhibition.41,45
Beyond its critical role in energy metabolism, glycolysis also plays a significant role in biosynthesis and cellular signaling. The pyruvate produced can enter the mitochondria and participate in the tricarboxylic acid (TCA) cycle and oxidative phosphorylation to generate large amounts of ATP.41,46 Moreover, glycolytic intermediates such as G3P and 1,3-BPG contribute to the biosynthesis of fatty acids and amino acids, meeting the cellular demand for metabolic precursors.41,47 Although glycolysis operates under both aerobic and anaerobic conditions, its activity and regulatory mechanisms vary significantly among different cell types and physiological states. Particularly in ovarian granulosa cells and oocytes, glycolytic activity is crucial for oocyte development and maturation, highlighting its essential role in female reproductive physiology.41,46
Microenvironment of Polycystic Ovary Syndrome
The disease microenvironment in PCOS displays complex pathological features resulting from the combined effects of insulin resistance, metabolic abnormalities, and chronic low-grade inflammation. These pathological features not only intertwine but also influence each other through various signaling pathways and molecular mechanisms, creating an environment conducive to the onset and progression of PCOS.
PCOS patients are often accompanied by hyperandrogenism and insulin resistance, which not only disrupt the hormonal balance within the ovaries but also directly affect the function of granulosa cells and the support provided to oocytes. Specifically, BMP15 and GDF9 secreted by oocytes normally activate the TGFβ signaling pathway, promoting granulosa cell proliferation, differentiation, and functional maintenance, thereby supporting oocyte maturation and normal follicular development.48–50 However, in the PCOS microenvironment, the expression of these growth factors may be suppressed or dysfunctional, leading to increased granulosa cell apoptosis, which weakens their support for oocytes.51 Additionally, the expression and function of gap junction proteins such as Cx43 in granulosa cells of PCOS follicles may be negatively regulated by elevated androgen levels. This not only weakens the nutrient and signaling exchange between oocytes and granulosa cells but also hinders normal metabolic processes and gene expression in oocytes, thus affecting their maturation.52,53 Furthermore, the elevated androgen levels in the PCOS microenvironment may directly act on granulosa cells, altering their hormone secretion profile, increasing androgen secretion while suppressing estrogen synthesis, disrupting the hormonal balance within the follicle. This hormonal imbalance not only suppresses the action of FSH but also interferes with the expression of FSH receptors, hindering the transformation of follicles from primordial to secondary and dominant stages.54–56 Additionally, under insulin resistance, abnormalities in the insulin and IGF signaling pathways further affect granulosa cell proliferation and function, leading to impaired follicular development and reduced oocyte quality.57,58
In the follicular fluid (FF) of PCOS patients, the levels of inflammatory cytokines such as interleukin-6 (IL-6), interleukin-1β (IL-1β), and tumor necrosis factor-α (TNFα) are significantly elevated, while the levels of the anti-inflammatory cytokine interleukin-10 (IL-10) are reduced. This imbalance in inflammatory cytokines leads to the formation of a pro-inflammatory and immune-suppressive microenvironment within the follicle, which in turn affects follicular development and oocyte quality.59–61 Among these, IL-6 not only plays a key role in the inflammatory response but also regulates cell proliferation and differentiation within the follicle. IL-1β, by activating the inflammasome, promotes the maturation and secretion of pro-inflammatory cytokines, further exacerbating the local inflammatory state. In addition, overexpression of TNFα can lead to increased cell apoptosis, inhibiting the expression of follicular development-related genes, ultimately affecting oocyte maturation and quality.62,63 Moreover, inflammasomes and their products IL-1β and IL-18 are significantly elevated in the FF of PCOS patients, suggesting that inflammasomes may play an important role in the pathogenesis of PCOS.62,63 Inflammasomes are intracellular multi-protein complexes that primarily promote the maturation and secretion of IL-1β and IL-18 through the activation of caspase-1, thereby enhancing the inflammatory response. In PCOS patients, excessive activation of inflammasomes may result in prolonged inflammation within the follicular microenvironment, further disrupting normal follicular development and oocyte maturation. Additionally, inflammasome activation may induce cell apoptosis and autophagy, further damaging the cellular structure and function within the follicle, leading to a decline in oocyte quality and limited embryonic developmental potential.62,63 Therefore, the role of inflammasomes in PCOS is not limited to promoting the secretion of pro-inflammatory cytokines but also directly or indirectly affects follicular development and oocyte function through various pathways. This pro-inflammatory environment not only directly acts on oocytes but also affects the function of granulosa cells and surrounding support cells, leading to impaired follicular development and ovulation disorders.
Insulin resistance, as one of the core pathological features of PCOS, further exacerbates the disruption of inflammation. Elevated insulin levels not only promote androgen synthesis but also interfere with glucose metabolism pathways within the follicle, disrupting the energy supply and maturation process of oocytes.64 Specifically, insulin resistance leads to hyperinsulinemia, and elevated insulin levels increase androgen production by promoting the expression of androgen synthesis enzymes in ovarian granulosa cells. These excess androgens not only directly affect follicular development but also activate downstream signaling pathways by binding to androgen receptors, further exacerbating metabolic and inflammatory disruptions. Furthermore, insulin resistance also activates inflammatory pathways, such as the NF-κB signaling pathway, further promoting the expression of pro-inflammatory cytokines and exacerbating the inflammatory state within the follicle.63,65 NF-κB, as a key transcription factor, can lead to the overexpression of various pro-inflammatory genes with sustained activation, further enhancing local inflammation, creating a vicious cycle, and ultimately leading to abnormal follicular development and oocyte dysfunction.
In terms of metabolism, the levels of branched-chain amino acids (BCAAs), such as leucine, isoleucine, and valine, are significantly elevated in the FF of PCOS patients, which is closely related to insulin resistance, hyperandrogenism, and menstrual irregularities.66 Moreover, glucose metabolism in PCOS patients is significantly affected, with reduced glucose levels in the FF and complex changes in lactate and pyruvate levels, reflecting impaired glycolysis and abnormal regulation of gluconeogenesis.64,66 Specifically, glucose is the main energy source for both oocytes and granulosa cells, and pyruvate and lactate from glycolysis are crucial metabolic products for oocyte energy supply. However, in PCOS patients, due to insulin resistance and abnormal activation of the mTOR pathway, glycolysis is inhibited, leading to reduced glucose utilization efficiency, which subsequently affects the energy supply and maturation of oocytes. In addition, the abnormal regulation of gluconeogenesis may lead to an imbalance of intracellular glucose metabolites, further exacerbating metabolic stress and cellular dysfunction.64,66 This metabolic disorder not only affects the energy supply of oocytes but also alters the balance of intracellular metabolic products, affecting cellular signaling and gene expression, ultimately leading to abnormal follicular development and a decline in oocyte quality. Furthermore, oxidative stress levels are significantly elevated in the FF of PCOS patients, as indicated by an increase in reactive oxygen species (ROS) products such as malondialdehyde (MDA) and 8-oxodeoxyguanosine (8-OHdG), while the activities of antioxidant enzymes such as superoxide dismutase (SOD) and glutathione peroxidase (GPx) are significantly reduced.67,68 Oxidative stress is caused by excessive ROS generation and a decline in antioxidant defense system function, leading to an imbalance in the cellular redox state. In PCOS patients, excessive ROS not only directly damages the cellular structure and DNA of oocytes but also activates multiple signaling pathways, inducing cell apoptosis and autophagy, which further reduce oocyte survival and developmental potential. Additionally, oxidative stress is closely linked to inflammation, as ROS can act as signaling molecules to promote the expression of inflammatory cytokines and the recruitment of inflammatory cells, exacerbating the local inflammatory state.69–71 Specifically, ROS promote the secretion of pro-inflammatory cytokines such as IL-6 and TNFα by activating NF-κB and other pro-inflammatory transcription factors, creating a vicious cycle of inflammation and oxidative stress that further disrupts the follicular microenvironment, leading to a decline in oocyte quality and impaired embryonic.
Abnormal Glycolysis in PCOS
During follicular development, the glycolytic activity in granulosa cells is crucial (Figure 3). The enhancement of glycolysis in granulosa cells can promote the activation of primordial follicles via the mTOR signaling pathway.72 Correspondingly, during the transition of primordial follicles to primary follicles in mice, the levels of glycolysis-related proteins in granulosa cells, such as GLUT 4, HK 1, PFKL, and PKM 2, are significantly increased.72 However, in granulosa cells (GCs) of PCOS patients, significant metabolic changes occur, with decreased mTOR pathway activity and reduced Akt phosphorylation levels.72,73 Granulosa cells also show notable changes in mitochondrial morphology, quantity, and localization, accompanied by reduced membrane potential, decreased ATP synthesis, increased mitochondrial ROS and oxidative stress, insufficient oxidative phosphorylation (OXPHOS), and reduced glycolysis.74
 |
Figure 3 How the disruption of the ovarian microenvironment in PCOS patients affects the glycolytic activity of granulosa cells: Increased DHT levels in follicular fluid may affect the changes in BMP4 levels within granulosa cells, thereby influencing the transcriptional activity of YAP1 and inhibiting glycolytic activity. Fluctuations in BCAA levels significantly inhibit glycolytic activity and oxidative phosphorylation in granulosa cells, shifting towards the pentose phosphate pathway. Additionally, during PCOS, the reduced activity of the mTOR pathway, decreased levels of deacetylases SIRT1/3 in granulosa cells, and the binding of YTHDF6 with MSS51 mRNA significantly increase mitochondrial ROS accumulation and decrease oxidative phosphorylation levels. Furthermore, granulosa cell-derived (miR-143-3P, miR-155-3P) and exosome-derived (miR-34a-5p) miRNAs may epigenetically suppress the expression of glycolysis-related genes, thereby affecting glycolytic activity. |
Studies indicate that the low expression of HIF-1α and the reduction in glycolysis levels in granulosa cells may be important factors contributing to the lower oocyte quality in PCOS.1 In vitro studies show that glycolytic activity in granulosa cells is time-dependently inhibited by DHT stimulation.73
Direct metabolic profiling of follicular fluid has shown that the levels of pyruvate and lactate are significantly decreased during the development of PCOS.75 However, the exact reasons for the altered metabolic profile in granulosa cells of PCOS patients remain unclear. During the development of PCOS, the levels of branched-chain amino acids (BCAAs) in both plasma and follicular fluid are significantly elevated.66,76,77 Studies on female mice with a Ppm1k deficiency have shown increased BCAA levels and PCOS-like characteristics, and these changes are linked to the conversion of glycolysis to the pentose phosphate pathway and the inhibition of mitochondrial oxidative phosphorylation (OXPHOS).66
In granulosa cells of PCOS patients, significant changes in acetylation levels are observed, mainly in the form of increased lysine acetylation modifications. These changes are associated with abnormal expression of acetyltransferases and deacetylases. Several sirtuin family proteins, including Sirt1-3, show significant alterations in expression during PCOS development, with SIRT3 expression being notably decreased.74 As a mitochondrial-localized deacetylase, SIRT3 removes acetyl groups from mitochondrial proteins, regulating mitochondrial function and metabolic homeostasis. The downregulation of SIRT3 leads to reduced mitochondrial membrane potential, increased ROS levels, and decreased OXPHOS efficiency. These changes not only impair cellular energy production but also exacerbate oxidative stress, further inhibiting glycolytic activity.74,78 Moreover, multiple studies have shown that manipulating Sirt2 in granulosa cells can effectively improve insulin resistance and activate glycolytic pathways, promoting lactate metabolism in granulosa cells.79,80 This is consistent with earlier studies indicating that Sirt2 expression is related to glycolytic activity in cells. Inhibition of SIRT2 using small-molecule inhibitors like AGK2 significantly decreases lactate levels and increases pyruvate accumulation.79,81
Similarly, SIRT1 expression is significantly downregulated in PCOS, resulting in decreased expression of downstream genes like PGC1α and mtTFA, which impairs mitochondrial function and energy metabolism. The reduction in SIRT1 inhibits AMPK activity, further suppressing glycolysis and leading to insufficient energy supply and metabolic imbalance. These changes exacerbate oxidative stress and mitochondrial dysfunction, ultimately affecting granulosa cell survival and follicular development.82,83 The expression of SIRT1 in granulosa cells may be influenced by advanced glycation end-products (AGEs), while the expression of ACAT1 (Acetyl-CoA Acetyltransferase 1) is significantly upregulated.84 This upregulation is accompanied by significant changes in acetylation levels across glycolysis, fatty acid degradation, the TCA cycle, tryptophan metabolism, and BCAA degradation pathways, which correlate negatively with oocyte quality and embryo development efficiency.84
Transcriptome sequencing of granulosa cells and follicular fluid exosomes has revealed changes in the levels of various non-coding RNAs during the development of PCOS.85,86 These changes may significantly impact the glycolytic activity in granulosa cells. While the roles of some non-coding RNAs in PCOS are still not fully understood,87 studies have found that miR-143-3p and miR-155-5p exhibit differential expression in exosomes from granulosa cells of PCOS patients.85,88 Specifically, the upregulation of miR-143-3p significantly inhibits glycolytic activity in granulosa cells by directly or indirectly targeting key enzymes in the glycolysis pathway, reducing the production of pyruvate and lactate. In contrast, silencing miR-155-5p weakens the activation of glycolysis, suggesting that miR-155-5p plays a positive role in promoting glycolysis.85,88 Non-coding RNAs in follicular fluid exosomes may also play a communicative role in PCOS pathogenesis. Studies have shown that miR-34a-5p directly targets lactate dehydrogenase A, inhibiting glycolysis and promoting granulosa cell apoptosis,88 thus affecting GC proliferation and follicular growth and development.
Additionally, studies suggest that the regulation of glycolytic activity in granulosa cells may be influenced by transcriptional and post-translational modifications. Bone Morphogenetic Protein 4 (BMP4) expression is significantly upregulated in a high androgen environment.89 BMP4 regulates glycolysis and autophagy pathways in granulosa cells by affecting the nuclear translocation of Yes-associated protein 1 (YAP1).89 Furthermore, the expression of YTHDF2 (YTH N6-methyladenosine RNA binding protein 2) is significantly upregulated in granulosa cells of PCOS patients.90 YTHDF2 regulates the stability and translation efficiency of MSS51 mRNA, leading to decreased MSS51 protein expression, which in turn affects the assembly and function of mitochondrial respiratory chain complexes and reduces the expression of glycolysis-related genes such as LDHA, PFKP, and PKM.90
Glycolytic Dysregulation and Metabolic Pathway Alterations in PCOS
AMPK Pathway
AMPK (AMP-activated protein kinase) plays a central role in regulating cellular energy balance, metabolism, proliferation, and apoptosis. Its role in PCOS (Polycystic Ovary Syndrome) has been widely studied, as it is a key enzyme that responds to the energy status of cells. Research indicates that AMPK activity is significantly reduced in PCOS patients, leading to metabolic imbalance, which in turn affects ovarian and uterine function.91 In PCOS animal models, silencing AMPK exacerbated polycystic ovarian changes and disrupted ovulation.91 AMPK regulates metabolism by sensing cellular energy levels, and its reduced activity leads to abnormal glycolysis in granulosa cells, disrupting cellular energy metabolism.
AMPK pathway dysfunction in PCOS is primarily linked to altered glycolytic activity, which impacts downstream signaling pathways such as mTOR and Akt. Specifically, the reduction in AMPK activity contributes to abnormal androgen levels, which further impairs follicular development.92 Additionally, AMPK activity is suppressed by abnormal glucose metabolism and mitochondrial dysfunction, particularly in the ovaries and granulosa cells of PCOS patients.74 AMPK also regulates autophagy through SIRT1, and disruption of AMPK signaling in PCOS can lead to autophagic dysfunction, further compromising ovarian function and fertility.91,92 The relationship between AMPK and glycolysis in PCOS is central to metabolic disturbances, particularly in granulosa cells. Disruption of glycolysis and oxidative phosphorylation (OXPHOS) impairs AMPK signaling. Glycolysis, being a core metabolic pathway, directly influences ATP synthesis and energy status within the cell. AMPK, as an energy sensor, activates under energy stress to regulate metabolic pathways, including enhancing glycolysis and inhibiting fatty acid synthesis to restore energy balance.92 However, in PCOS, changes in glycolysis hinder AMPK activation. When glycolysis is impaired, ATP levels decrease, activating AMPK. Yet, due to the continued suppression of glycolysis, AMPK’s activity cannot be effectively restored, creating a vicious cycle that exacerbates metabolic dysfunction in PCOS.92
Conversely, increasing glycolysis may help improve metabolic abnormalities in PCOS by enhancing AMPK activation. Studies show that short-term pyruvate deprivation can increase glycolytic protein levels in mouse ovaries and activate the AMPK pathway, promoting the restoration of ovarian energy metabolism.72 By enhancing glycolysis, AMPK activation can normalize energy balance and regulate downstream mTOR signaling to promote granulosa cell proliferation and follicular development.72 Additionally, AMPK activation can increase the expression of glycolytic enzymes like LDHA and PKM2, further promoting glycolysis and correcting metabolic dysregulation in PCOS.91
mTOR Pathway
The mTOR (mechanistic target of rapamycin) pathway plays a significant role in the regulation of follicular development, and its relationship with glycolysis is particularly relevant during the transition of primordial follicles to primary follicles. Studies on mice have shown that glycolysis-related proteins such as GLUT4, HK1, PFKL, and PKM2 are significantly upregulated in granulosa cells during this process, indicating a positive correlation between glycolytic activity and mTOR signaling.72 Acute fasting and pyruvate deprivation in vivo and in vitro increase glycolytic gene and protein expression in the ovaries, decrease AMPK activity, and enhance mTOR activation, which in turn leads to elevated phosphorylation of downstream proteins like Akt and FOXO3a, promoting primordial follicle activation.72 Blocking glycolysis with 2-deoxyglucose significantly inhibits the mTOR pathway activation caused by pyruvate deprivation, while KIT inhibitor ISCK03 does not affect the communication between granulosa cells and oocytes, further reducing mTOR activity.72 However, the relationship between PCOS and the mTOR pathway is complex and contradictory. For example, in the DHEA-induced PCOS mouse model, phosphorylation levels of mTOR (ser2448 and ser2481) in granulosa cells are significantly increased compared to normal mice, suggesting activation of mTORC1/2. However, downstream kinase S6K1 expression is reduced.93 Conversely, in granulosa cells during the luteal phase of PCOS patients, although basal mTOR protein levels are similar to those of healthy women, insulin stimulation leads to a marked decrease in mTOR expression,94 along with downregulation of downstream kinase S6K1. This suggests that PCOS may be associated with reduced mTOR pathway activity in granulosa cells. Meanwhile, aberrant Akt/mTOR pathway regulation in PCOS granulosa cells affects their proliferation and apoptosis.94 At the same time, studies have shown that short-term administration of the mTOR inhibitor rapamycin not only fails to protect DHEA-induced non-cyclical and PCO morphology, but also hinders DHEA-induced mouse follicle development and increased serum testosterone levels, which is related to the inhibition of Hsd3b1, Cyp17a1 and Cyp19a1 expression and the effect on mitochondrial function,95 This also shows the complexity of the mTOR pathway in the pathogenesis of PCOS, so precise regulation may be crucial for the treatment of PCOS. Several studies have indicated that activation of the PI3K/Akt/mTOR pathway is crucial for ovarian function,96 and inactivation of this pathway is linked to inhibited granulosa cell proliferation and increased apoptosis97 Table 1.
Potential Strategies for Targeting Glycolysis in PCOS Treatment
Metformin, a classic drug for treating polycystic ovary syndrome (PCOS), has been widely used. Additionally, adolescent PCOS patients, especially those who are overweight or obese, often face a higher risk of metabolic syndrome.98,99 The use of metformin (1700 mg/day for 6 months) in these patients can improve menstrual cycles and promote ovulation. Moreover, metformin can reduce patients’ body mass index (BMI) and symptoms of hyperandrogenism, such as hirsutism and acne, while also lowering total testosterone, androstenedione, and free testosterone levels.100,101 The regulatory effects of metformin on hormone and metabolic status in PCOS patients are closely related to improvements in ovulation, pregnancy rates, and hyperandrogenism symptoms.102 The primary mechanism of action of metformin is the improvement of insulin resistance. It is believed that metformin improves ovulation and hormone changes by activating the AMPK/SIRT1 signaling pathway in the ovaries.103 Dysfunction of the AMPK/SIRT1 signaling pathway in granulosa cells is associated with insulin resistance and hyperandrogenemia. By activating the AMPK/SIRT1 pathway, metformin effectively regulates insulin resistance and improves hormonal status.104,105 Furthermore, by activating AMPK, metformin inhibits the mTOR signaling pathway and reduces the expression of MMP-2 and MMP-9 in PCOS animal models.106 Recent studies have also revealed that metformin improves the metabolic state and ovarian function of PCOS patients by activating the AMPK/SIRT1 signaling pathway and regulating the expression of glycolytic enzymes. Glycolysis is a crucial metabolic pathway in cellular energy metabolism, and its dysfunction plays a key role in the metabolic disturbances of PCOS.80,83 Metformin can restore energy balance by activating AMPK, inhibit the mTOR signaling pathway, and repair the glycolytic dysfunction, improving the physiological state of the ovaries. Studies have shown that metformin significantly increases the expression of lactate dehydrogenase A (LDH-A), pyruvate kinase M2 (PKM2), and SIRT1 in the ovaries of PCOS rats.80,83 LDH-A and PKM2 are key rate-limiting enzymes in the glycolytic pathway, and their increase not only promotes the glycolytic reaction but also improves the follicular development defects caused by abnormal glucose metabolism in PCOS patients.80,83 Recent studies have shown that metformin, by activating the AMPK–SIRT2 axis, restores the expression of key glycolytic enzymes suppressed by insulin, thereby enhancing glycolytic flux in ovarian granulosa cells.107 At the same time, metformin downregulates insulin-like growth factor 1 (IGF-1) and upregulates its receptor (IGF-1R), further ameliorating insulin resistance and hyperandrogenism to collectively correct the energy metabolic imbalance characteristic of polycystic ovary syndrome (PCOS).107 Moreover, combination therapy of metformin with other agents has been reported to improve glycolytic activity and overall metabolic function in granulosa cells from PCOS models.108 The therapeutic effects of metformin also manifest in its ability to inhibit the mTOR signaling pathway, reduce excessive energy accumulation in the ovaries, restore ovarian function, and improve follicle quality and fertility.92 Through these mechanisms, metformin effectively alleviates the metabolic disturbances and ovarian dysfunction of PCOS, providing significant therapeutic benefits for PCOS patients. However, it is worth noting that, although metformin is generally well-tolerated in the treatment of PCOS and no serious adverse events or hepatic or renal dysfunction were observed during the clinical period, the most commonly reported side effects are short-term gastrointestinal disturbances—including nausea, bloating, and diarrhea—with roughly one quarter of adult participants experiencing these symptoms yet continuing the medication, and a small number of adolescents discontinuing treatment after gastrointestinal reactions persisted for more than two weeks.100,101
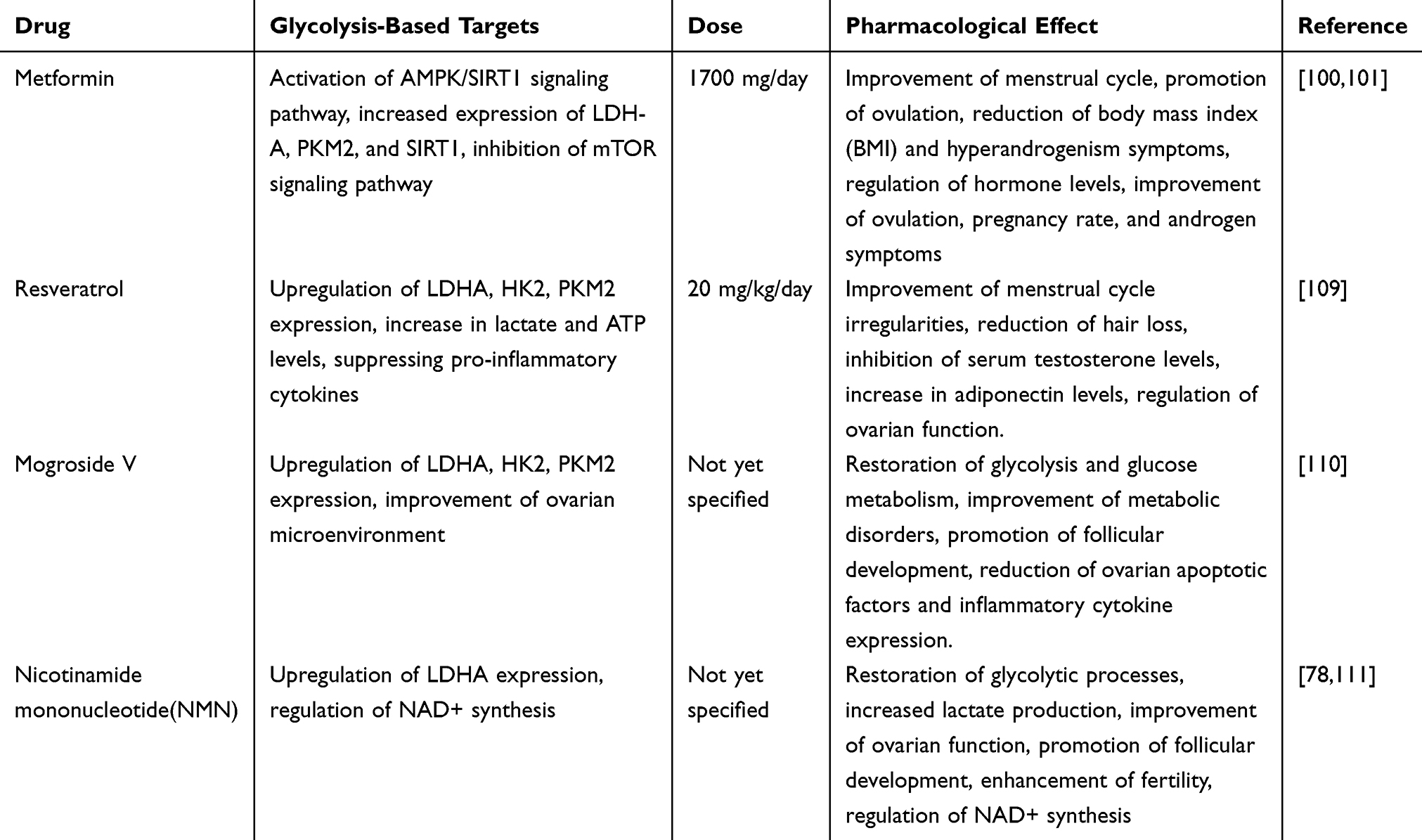 |
Table 1 Potential Drugs Targeting Glycolysis for the Treatment of PCOS |
Resveratrol is a natural polyphenolic compound widely found in grape skins, berries, and peanuts. Its antioxidant and anti-inflammatory properties make it a potential treatment for PCOS. In rat experiments, resveratrol (20 mg/kg/day) combined with metformin (300 mg/kg/day) significantly reduced the levels of LH, LH/FSH ratio, TNF-α, and AMH in the ovaries of PCOS rats.109 Additionally, resveratrol possesses anti-inflammatory, antioxidant, and anti-apoptotic properties. Several studies have shown that resveratrol can modulate the NF-κB pathway by inhibiting IκB kinase activity,112 suppressing pro-inflammatory cytokines.113,114 Clinical studies have demonstrated that resveratrol can improve menstrual cycle irregularities in PCOS and reduce hair loss in PCOS patients.115 In combination with myo-inositol (1000 mg twice daily for 12 weeks), resveratrol significantly reduced serum testosterone levels, increased adiponectin levels, and resulted in a decrease in serum LH and FSH levels as well as ovarian volume.116 From the perspective of glycolysis, resveratrol can restore the glycolytic process by upregulating the expression of key enzymes (such as LDHA, HK2, and PKM2), significantly increasing lactate and ATP levels, and thus improving cellular energy metabolism.80 The mechanism of action of resveratrol primarily involves activating the AMPK/SIRT2 signaling pathway, promoting normal glucose metabolism. AMPK is a key kinase for cellular energy sensing, and its activation helps regulate metabolic balance within cells, restoring ovarian dysfunction caused by glucose metabolism disorders in PCOS patients.80 In PCOS rat models, resveratrol significantly improved insulin resistance and regulated the expression of glycolytic enzymes by enhancing AMPK activity, suggesting its potential in the treatment of PCOS.80 These studies suggest that resveratrol, as a natural glycolysis regulator, can alleviate metabolic disturbances in PCOS and may improve ovarian function and fertility. However, clinical studies indicate that resveratrol’s teratogenicity remains controversial, often leading to an increased abortion rate in assisted reproduction.117 Clinical studies have shown that, although combination therapy with resveratrol does not present obvious side effects and is generally well tolerated, a double-blind trial demonstrated an increase in fat mass and a decrease in lean body mass after 3 months (p = 0.03 and 0.004),116 and previous research indicates that high-dose resveratrol treatment may be associated with elevated TSH levels and an increased risk of thyroid enlargement.118
Mogroside V (MV) is a natural compound extracted from the fruit of Siraitia grosvenorii, known for its antioxidant and anti-inflammatory properties, making it a promising drug for the treatment of PCOS. MV regulates the glycolytic pathway to protect ovarian function in PCOS rats. Studies have shown that after MV treatment in PCOS rats, the levels of D-glucose-6-phosphate, lactate, and GTP were significantly increased, while pyruvate levels were reduced, indicating that MV can improve metabolic disturbances in PCOS patients by restoring glycolysis and glucose metabolism.110 Glycolysis is one of the important pathways for cellular energy production, and a normal glycolytic process can effectively provide ATP for cellular metabolism, improving cellular energy supply. In PCOS patients, the ovaries often exhibit glycolytic insufficiency, which leads to impaired follicular development.72 MV enhances the expression of rate-limiting enzymes in glycolysis, such as LDHA, hexokinase 2 (HK2), and PKM2, increasing lactate and ATP production, thereby improving the ovarian microenvironment, promoting follicular development and ovulation.110 These effects suggest that MV can not only improve ovarian function by regulating the glycolytic pathway but also support follicular development by providing energy, and it may reduce the expression of ovarian stress factors (NLRP3, Caspase-1, GSDMD) and inflammatory cytokines (IL-1β, IL-18), thereby lowering insulin resistance and apoptotic markers.119 However, the clinical research on MV for the treatment of PCOS is still lacking.
Nicotinamide mononucleotide (NMN) is a precursor molecule that can regulate cellular energy metabolism by supplementing NAD+. NAD+ is an important coenzyme in cellular metabolism, involved in glycolysis, oxidative phosphorylation, and many other metabolic pathways. The levels of NAD+ and the NAD+/NADH ratio in granulosa cells (GC) decrease in response to LPS-induced PCOS, while NMN can mitigate LPS-induced GC inflammation. NMN supplementation restores NAD+ levels in GCs and significantly reduces the expression of pro-inflammatory markers at both mRNA and protein levels, downregulates pro-apoptotic markers such as Caspase-3, Caspase-9, and Bax, and upregulates anti-apoptotic markers like Bcl-2 to reduce GC apoptosis.120 Moreover, NMN intervention can alleviate hyperandrogenemia, ovarian abnormalities, and bile acid reduction.111 Studies have shown that NMN can restore the glycolytic pathway in granulosa cells, promote follicular development, and improve fertility.78 NMN activates the glycolytic process in the ovaries, increasing lactate production and enhancing LDHA expression, improving ovarian function in PCOS rats.78 Normalizing the glycolytic pathway provides sufficient energy to support follicular development and maturation, thus improving fertility in PCOS patients. NMN also enhances the NAD+ synthesis pathway, improving cellular energy metabolism and promoting normal ovarian function.78 These studies suggest that NMN has potential in the treatment of PCOS, especially in improving ovarian function and follicular development. Meanwhile, research indicates that NMN supplementation may address the decline in NAD+ levels associated with PCOS in skeletal muscle, normalizing insulin sensitivity and preventing compensatory hyperinsulinemia associated with obesity and hepatic lipid deposition.121 However, clinical studies on NMN for PCOS treatment are still lacking.
Conclusion
PCOS remains a complex endocrine disorder with a multifaceted pathophysiology, involving reproductive and metabolic dysfunctions. Among the key features of PCOS, the aberrant regulation of energy metabolism, particularly glycolysis, plays a pivotal role in the disease’s progression. Glycolysis, a fundamental metabolic pathway responsible for converting glucose into pyruvate while generating ATP, is crucial for ovarian cell function and follicular development. In PCOS, the glycolytic pathway is disrupted, leading to reduced glucose utilization, altered mitochondrial function, and compromised energy supply in granulosa cells and oocytes, which are vital for follicular growth and oocyte maturation. These metabolic disturbances contribute to the hallmark features of PCOS, including insulin resistance, hyperandrogenism, and impaired ovarian function. As such, glycolytic dysregulation is a central feature of PCOS pathogenesis and a potential therapeutic target.
Recent research has underscored the significance of glycolysis in PCOS, particularly in granulosa cells, where changes in the expression of key enzymes like HK2, LDHA, and PKM2 can impact cellular energy metabolism. These enzymes are critical for the proper functioning of glycolysis and its relationship with mitochondrial function. A marked reduction in glycolytic activity, often due to altered signaling pathways like AMPK, mTOR, and insulin signaling, results in decreased ATP production, contributing to follicular dysfunction and impaired oocyte quality. In particular, the interaction between glycolysis and AMPK activation is crucial for maintaining metabolic homeostasis in ovarian cells. Reduced AMPK activity, commonly observed in PCOS, disrupts glycolysis and further exacerbates the metabolic abnormalities associated with the syndrome.
Given the critical role of glycolysis in PCOS pathophysiology, therapeutic interventions targeting metabolic pathways offer promising treatment strategies. Drugs that modulate glycolytic activity and improve cellular energy balance may provide significant benefits to PCOS patients. Metformin, a widely used drug for managing insulin resistance in PCOS, has shown efficacy in improving metabolic profiles and restoring ovarian function. By activating AMPK and regulating glycolysis-related enzymes, metformin enhances energy metabolism, reduces hyperandrogenism, and improves follicular development, thereby addressing both the metabolic and reproductive aspects of PCOS. Moreover, natural compounds like resveratrol, which modulate glycolysis via AMPK and SIRT1 pathways, have also demonstrated potential in improving metabolic dysfunction in PCOS. These treatments highlight the therapeutic potential of targeting glycolytic pathways and energy metabolism to ameliorate the symptoms and underlying causes of PCOS.
Other emerging therapies, such as NMN (Nicotinamide Mononucleotide), have shown promise in restoring NAD+ levels and improving energy metabolism in granulosa cells. NMN supplementation has been demonstrated to enhance glycolysis and mitochondrial function, offering a novel approach for addressing the metabolic dysregulation in PCOS. Similarly, natural compounds like Mogroside V (MV), which have antioxidant and anti-inflammatory properties, can improve glycolytic activity and metabolic homeostasis in the ovaries, potentially improving ovulatory function and fertility outcomes in PCOS patients. Despite these promising therapeutic options, much remains to be understood regarding the precise mechanisms linking glycolysis to PCOS and how these metabolic alterations can be effectively targeted for therapeutic intervention. Future research should focus on further elucidating the molecular pathways that regulate glycolysis and energy metabolism in PCOS. In particular, the role of non-coding RNAs, epigenetic modifications, and metabolic enzymes in the pathogenesis of PCOS warrants further investigation. Additionally, clinical trials are needed to validate the therapeutic potential of drugs and natural compounds that target glycolysis and energy metabolism in improving the reproductive and metabolic health of PCOS patients.
In conclusion, glycolysis is central to the pathophysiology of PCOS, and understanding its regulation and dysfunction in ovarian cells could provide key insights into novel therapeutic approaches. As research continues to unfold, targeted metabolic therapies, including those focused on enhancing glycolytic pathways, may offer new hope for patients with PCOS, particularly in addressing the complex interplay between metabolic and reproductive dysfunctions that define this disorder.
Data Sharing Statement
The current study was based on the results of relevant published studies.
Author Contributions
Zhenzhen Cao contributed to the study conception and manuscript drafting. Qin Zhou assisted in data interpretation and critical revision. Jie An participated in data collection and manuscript editing. Xiaojing Guo contributed to reference management and formatting. Yuena Qiu assisted in data acquisition. XiaoFang Jia supervised the study and finalized the manuscript. All authors made a significant contribution to the work reported, took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This review was funded by Suzhou Science and Technology Development Plan Project, Project No. SYWD2024282, Suzhou Major Diseases Multi-center Clinical Research Project, Project No. DZXYJ202421 and Xia Guicheng Gynecology Expert Team of Jiangsu Provincial Hospital of Traditional Chinese Medicine (01201804).
Disclosure
The authors declared that they have no competing interests in this work.
References
1. Wang J, Wu X. The effects of mitochondrial dysfunction on energy metabolism switch by HIF-1alpha signalling in granulosa cells of polycystic ovary syndrome. Endokrynol Pol. 2020;71(2):134–145. doi:10.5603/EP.a2020.0002
2. Yan H, Wang L, Zhang G, et al. Oxidative stress and energy metabolism abnormalities in polycystic ovary syndrome: from mechanisms to therapeutic strategies. Reprod Biol Endocrinol. 2024;22(1):159. doi:10.1186/s12958-024-01337-0
3. Meng Y, Zhao T, Zhang R, Zhu X, Ma C, Shi Q. Global burden of polycystic ovary syndrome among women of childbearing age, 1990-2021: a systematic analysis using the global burden of disease study 2021. Front Public Health. 2025;13:1514250. doi:10.3389/fpubh.2025.1514250
4. Sendur SN, Yildiz BO. Influence of ethnicity on different aspects of polycystic ovary syndrome: a systematic review. Reprod Biomed Online. 2021;42(4):799–818. doi:10.1016/j.rbmo.2020.12.006
5. Siemers KM, Klein AK, Baack ML. Mitochondrial dysfunction in PCOS: insights into reproductive organ pathophysiology. Int J Mol Sci. 2023;24(17):13123. doi:10.3390/ijms241713123
6. Zhang J, Bao Y, Zhou X, Zheng L. Polycystic ovary syndrome and mitochondrial dysfunction. Reprod Biol Endocrinol. 2019;17(1):67. doi:10.1186/s12958-019-0509-4
7. Cozzolino M, Seli E. Mitochondrial function in women with polycystic ovary syndrome. Curr Opin Obstet Gynecol. 2020;32(3):205–212. doi:10.1097/GCO.0000000000000619
8. Dapas M, Dunaif A. Deconstructing a syndrome: genomic insights into PCOS causal mechanisms and classification. Endocr Rev. 2022;43(6):927–965. doi:10.1210/endrev/bnac001
9. Azziz R, Carmina E, Chen Z, et al. Polycystic ovary syndrome. Nat Rev Dis Primers. 2016;2(1):16057. doi:10.1038/nrdp.2016.57
10. Starace M, Orlando G, Alessandrini A, Piraccini BM. Female androgenetic alopecia: an update on diagnosis and management. Am J Clin Dermatol. 2020;21(1):69–84. doi:10.1007/s40257-019-00479-x
11. Goodman NF, Cobin RH, Futterweit W, et al. American association of clinical endocrinologists, American college of endocrinology, and androgen excess and pcos society disease state clinical review: guide to the best practices in the evaluation and treatment of polycystic ovary syndrome--part 1. Endocr Pract. 2015;21(11):1291–1300. doi:10.4158/EP15748.DSC
12. Azziz R. Polycystic Ovary Syndrome. Obstet Gynecol. 2018;132(2):321–336. doi:10.1097/AOG.0000000000002698
13. Throwba HP, Unnikrishnan L, Pangath M, et al. The epigenetic correlation among ovarian cancer, endometriosis and PCOS: a review. Crit Rev Oncol Hematol. 2022;180:103852. doi:10.1016/j.critrevonc.2022.103852
14. Rudnicka E, Kunicki M, Calik-Ksepka A, et al. Anti-mullerian hormone in pathogenesis, diagnostic and treatment of PCOS. Int J Mol Sci. 2021;22(22):12507. doi:10.3390/ijms222212507
15. Diamanti-Kandarakis E, Dunaif A. Insulin resistance and the polycystic ovary syndrome revisited: an update on mechanisms and implications. Endocr Rev. 2012;33(6):981–1030. doi:10.1210/er.2011-1034
16. Burghen GA, Givens JR, Kitabchi AE. Correlation of hyperandrogenism with hyperinsulinism in polycystic ovarian disease. J Clin Endocrinol Metab. 1980;50(1):113–116. doi:10.1210/jcem-50-1-113
17. Govind A, Obhrai MS, Clayton RN. Polycystic ovaries are inherited as an autosomal dominant trait: analysis of 29 polycystic ovary syndrome and 10 control families. J Clin Endocrinol Metab. 1999;84(1):38–43. doi:10.1210/jcem.84.1.5382
18. Franks S, Webber LJ, Goh M, et al. Ovarian morphology is a marker of heritable biochemical traits in sisters with polycystic ovaries. J Clin Endocrinol Metab. 2008;93(9):3396–3402. doi:10.1210/jc.2008-0369
19. Day F, Karaderi T, Jones MR, et al. Large-scale genome-wide meta-analysis of polycystic ovary syndrome suggests shared genetic architecture for different diagnosis criteria. PLoS Genet. 2018;14(12):e1007813. doi:10.1371/journal.pgen.1007813
20. Shi Y, Zhao H, Shi Y, et al. Genome-wide association study identifies eight new risk loci for polycystic ovary syndrome. Nat Genet. 2012;44(9):1020–1025. doi:10.1038/ng.2384
21. Chen ZJ, Zhao H, He L, et al. Genome-wide association study identifies susceptibility loci for polycystic ovary syndrome on chromosome 2p16.3, 2p21 and 9q33.3. Nat Genet. 2011;43(1):55–59. doi:10.1038/ng.732
22. Hayes MG, Urbanek M, Ehrmann DA, et al. Genome-wide association of polycystic ovary syndrome implicates alterations in gonadotropin secretion in European ancestry populations. Nat Commun. 2015;6(1):7502. doi:10.1038/ncomms8502
23. Day FR, Hinds DA, Tung JY, et al. Causal mechanisms and balancing selection inferred from genetic associations with polycystic ovary syndrome. Nat Commun. 2015;6(1):8464. doi:10.1038/ncomms9464
24. Zhang Y, Ho K, Keaton JM, et al. A genome-wide association study of polycystic ovary syndrome identified from electronic health records. Am J Obstet Gynecol. 2020;223(4):559e1–559e21. doi:10.1016/j.ajog.2020.04.004
25. Day F, Karaderi T, Jones MR, et al. Correction: large-scale genome-wide meta-analysis of polycystic ovary syndrome suggests shared genetic architecture for different diagnosis criteria. PLoS Genet. 2019;15(12):e1008517. doi:10.1371/journal.pgen.1008517
26. Brower MA, Hai Y, Jones MR, et al. Bidirectional Mendelian randomization to explore the causal relationships between body mass index and polycystic ovary syndrome. Hum Reprod. 2019;34(1):127–136. doi:10.1093/humrep/dey343
27. Zhao Y, Xu Y, Wang X, et al. Body mass index and polycystic ovary syndrome: a 2-sample bidirectional Mendelian randomization study. J Clin Endocrinol Metab. 2020;105(6):1778–1784. doi:10.1210/clinem/dgaa125
28. Ruth KS, Day FR, Tyrrell J, et al. Using human genetics to understand the disease impacts of testosterone in men and women. Nat Med. 2020;26(2):252–258. doi:10.1038/s41591-020-0751-5
29. Sun S, Jiao M, Han C, et al. Causal effects of genetically determined metabolites on risk of polycystic ovary syndrome: a Mendelian randomization study. Front Endocrinol. 2020;11:621. doi:10.3389/fendo.2020.00621
30. Zhu T, Cui J, Goodarzi MO. Polycystic ovary syndrome and risk of type 2 diabetes, coronary heart disease, and stroke. Diabetes. 2021;70(2):627–637. doi:10.2337/db20-0800
31. Pan JX, Tan YJ, Wang FF, et al. Aberrant expression and DNA methylation of lipid metabolism genes in PCOS: a new insight into its pathogenesis. Clin Clin Epigenet. 2018;10(1):6. doi:10.1186/s13148-018-0442-y
32. Sagvekar P, Kumar P, Mangoli V, Desai S, Mukherjee S. DNA methylome profiling of granulosa cells reveals altered methylation in genes regulating vital ovarian functions in polycystic ovary syndrome. Clin Clin Epigenet. 2019;11(1):61. doi:10.1186/s13148-019-0657-6
33. Kokosar M, Benrick A, Perfilyev A, et al. Epigenetic and transcriptional alterations in human adipose tissue of polycystic ovary syndrome. Sci Rep. 2016;6(1):22883. doi:10.1038/srep22883
34. Nilsson E, Benrick A, Kokosar M, et al. Transcriptional and epigenetic changes influencing skeletal muscle metabolism in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2018;103(12):4465–4477. doi:10.1210/jc.2018-00935
35. Lambertini L, Saul SR, Copperman AB, et al. Intrauterine reprogramming of the polycystic ovary syndrome: evidence from a pilot study of cord blood global methylation analysis. Front Endocrinol. 2017;8:352. doi:10.3389/fendo.2017.00352
36. Mimouni NEH, Paiva I, Barbotin AL, et al. Polycystic ovary syndrome is transmitted via a transgenerational epigenetic process. Cell Metab. 2021;33(3):513–530.e8. doi:10.1016/j.cmet.2021.01.004
37. Armanini D, Boscaro M, Bordin L, Sabbadin C. Controversies in the pathogenesis, diagnosis and treatment of PCOS: focus on insulin resistance, inflammation, and hyperandrogenism. Int J Mol Sci. 2022;23(8):4110. doi:10.3390/ijms23084110
38. Kim E, Seok HH, Lee SY, et al. Correlation between expression of glucose transporters in granulosa cells and oocyte quality in women with polycystic ovary syndrome. Endocrinol Metab. 2014;29(1):40–47. doi:10.3803/EnM.2014.29.1.40
39. Zhang CH, Liu XY, Wang J. Essential role of granulosa cell glucose and lipid metabolism on oocytes and the potential metabolic imbalance in polycystic ovary syndrome. Int J Mol Sci. 2023;24(22). doi:10.3390/ijms242216247
40. Chaudhry R, Varacallo MA. Biochemistry, glycolysis. StatPearls. 2025.
41. Fuller GG, Kim JK. Compartmentalization and metabolic regulation of glycolysis. J Cell Sci. 2021;134(20). doi:10.1242/jcs.258469
42. Mor I, Cheung EC, Vousden KH. Control of glycolysis through regulation of PFK1: old friends and recent additions. Cold Spring Harb Symp Quant Biol. 2011;76:211–216. doi:10.1101/sqb.2011.76.010868
43. Hallows WC, Yu W, Denu JM. Regulation of glycolytic enzyme phosphoglycerate mutase-1 by Sirt1 protein-mediated deacetylation. J Biol Chem. 2012;287(6):3850–3858. doi:10.1074/jbc.M111.317404
44. Bian X, Jiang H, Meng Y, Li YP, Fang J, Lu Z. Regulation of gene expression by glycolytic and gluconeogenic enzymes. Trends Cell Biol. 2022;32(9):786–799. doi:10.1016/j.tcb.2022.02.003
45. Xiong Y, Lei QY, Zhao S, Guan KL. Regulation of glycolysis and gluconeogenesis by acetylation of PKM and PEPCK. Cold Spring Harb Symp Quant Biol. 2011;76:285–289. doi:10.1101/sqb.2011.76.010942
46. Kierans SJ, Taylor CT. Glycolysis: a multifaceted metabolic pathway and signaling hub. J Biol Chem. 2024;300(11):107906. doi:10.1016/j.jbc.2024.107906
47. Bar-Even A, Flamholz A, Noor E, Milo R. Rethinking glycolysis: on the biochemical logic of metabolic pathways. Nat Chem Biol. 2012;8(6):509–517. doi:10.1038/nchembio.971
48. Juengel JL, McNatty KP. The role of proteins of the transforming growth factor-beta superfamily in the intraovarian regulation of follicular development. Hum Reprod Update. 2005;11(2):143–160. doi:10.1093/humupd/dmh061
49. de Castro FC, Cruz MH, Leal CL. Role of growth differentiation factor 9 and bone morphogenetic protein 15 in ovarian function and their importance in mammalian female fertility - A review. Asian-Australas J Anim Sci. 2016;29(8):1065–1074. doi:10.5713/ajas.15.0797
50. Persani L, Rossetti R, Di Pasquale E, Cacciatore C, Fabre S. The fundamental role of bone morphogenetic protein 15 in ovarian function and its involvement in female fertility disorders. Hum Reprod Update. 2014;20(6):869–883. doi:10.1093/humupd/dmu036
51. Hussein TS, Froiland DA, Amato F, Thompson JG, Gilchrist RB. Oocytes prevent cumulus cell apoptosis by maintaining a morphogenic paracrine gradient of bone morphogenetic proteins. J Cell Sci. 2005;118(Pt 22):5257–5268. doi:10.1242/jcs.02644
52. Chang HM, Cheng JC, Leung PC. Theca-derived BMP4 and BMP7 down-regulate connexin43 expression and decrease gap junction intercellular communication activity in immortalized human granulosa cells. J Clin Endocrinol Metab. 2013;98(3):E437–45. doi:10.1210/jc.2012-3851
53. Chang HM, Cheng JC, Taylor E, Leung PC. Oocyte-derived BMP15 but not GDF9 down-regulates connexin43 expression and decreases gap junction intercellular communication activity in immortalized human granulosa cells. Mol Hum Reprod. 2014;20(5):373–383. doi:10.1093/molehr/gau001
54. Havelock JC, Rainey WE, Carr BR. Ovarian granulosa cell lines. Mol Cell Endocrinol. 2004;228(1–2):67–78. doi:10.1016/j.mce.2004.04.018
55. Sugiura K, Pendola FL, Eppig JJ. Oocyte control of metabolic cooperativity between oocytes and companion granulosa cells: energy metabolism. Dev Biol. 2005;279(1):20–30. doi:10.1016/j.ydbio.2004.11.027
56. Thomas RE, Armstrong DT, Gilchrist RB. Bovine cumulus cell-oocyte gap junctional communication during in vitro maturation in response to manipulation of cell-specific cyclic adenosine 3’,5’-monophosophate levels. Biol Reprod. 2004;70(3):548–556. doi:10.1095/biolreprod.103.021204
57. Orisaka M, Jiang JY, Orisaka S, Kotsuji F, Tsang BK. Growth differentiation factor 9 promotes rat preantral follicle growth by up-regulating follicular androgen biosynthesis. Endocrinology. 2009;150(6):2740–2748. doi:10.1210/en.2008-1536
58. Honda A, Hirose M, Inoue K, et al. Large-scale production of growing oocytes in vitro from neonatal mouse ovaries. Int J Dev Biol. 2009;53(4):605–613. doi:10.1387/ijdb.082607ah
59. Artimani T, Karimi J, Mehdizadeh M, et al. Evaluation of pro-oxidant-antioxidant balance (PAB) and its association with inflammatory cytokines in polycystic ovary syndrome (PCOS). Gynecol Endocrinol. 2018;34(2):148–152. doi:10.1080/09513590.2017.1371691
60. Wang B, Hao M, Yang Q, Li J, Guo Y. Follicular fluid soluble receptor for advanced glycation endproducts (sRAGE): a potential protective role in polycystic ovary syndrome. J Assist Reprod Genet. 2016;33(7):959–965. doi:10.1007/s10815-016-0704-6
61. Zhang H, Wang X, Xu J, Zhu Y, Chen X, Hu Y. IL-18 and IL-18 binding protein concentration in ovarian follicular fluid of women with unexplained infertility to PCOS during in vitro fertilization. J Reprod Immunol. 2020;138:103083. doi:10.1016/j.jri.2020.103083
62. Rostamtabar M, Esmaeilzadeh S, Tourani M, et al. Pathophysiological roles of chronic low-grade inflammation mediators in polycystic ovary syndrome. J Cell Physiol. 2021;236(2):824–838. doi:10.1002/jcp.29912
63. Liu Y, Liu H, Li Z, et al. The release of peripheral immune inflammatory cytokines promote an inflammatory cascade in PCOS patients via altering the follicular microenvironment. Front Immunol. 2021;12:685724. doi:10.3389/fimmu.2021.685724
64. Castiglione Morelli MA, Iuliano A, Schettini SCA, et al. NMR metabolic profiling of follicular fluid for investigating the different causes of female infertility: a pilot study. Metabolomics. 2019;15(2):19. doi:10.1007/s11306-019-1481-x
65. Lord T, Aitken RJ. Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction. 2013;146(6):R217–27. doi:10.1530/REP-13-0111
66. Mu L, Ye Z, Hu J, et al. PPM1K-regulated impaired catabolism of branched-chain amino acids orchestrates polycystic ovary syndrome. EBioMedicine. 2023;89:104492. doi:10.1016/j.ebiom.2023.104492
67. Naigaonkar A, Dadachanji R, Hinduja I, Mukherjee S. Altered redox status may contribute to aberrant folliculogenesis and poor reproductive outcomes in women with polycystic ovary syndrome. J Assist Reprod Genet. 2021;38(10):2609–2623. doi:10.1007/s10815-021-02241-x
68. Seleem AK, El Refaeey AA, Shaalan D, Sherbiny Y, Badawy A. Superoxide dismutase in polycystic ovary syndrome patients undergoing intracytoplasmic sperm injection. J Assist Reprod Genet. 2014;31(4):499–504. doi:10.1007/s10815-014-0190-7
69. Bausenwein J, Serke H, Eberle K, et al. Elevated levels of oxidized low-density lipoprotein and of catalase activity in follicular fluid of obese women. Mol Hum Reprod. 2010;16(2):117–124. doi:10.1093/molehr/gap078
70. Wiener-Megnazi Z, Vardi L, Lissak A, et al. Oxidative stress indices in follicular fluid as measured by the thermochemiluminescence assay correlate with outcome parameters in in vitro fertilization. Fertil Steril. 2004;82 Suppl 3:1171–1176. doi:10.1016/j.fertnstert.2004.06.013
71. Nishihara T, Matsumoto K, Hosoi Y, Morimoto Y. Evaluation of antioxidant status and oxidative stress markers in follicular fluid for human in vitro fertilization outcome. Reprod Med Biol. 2018;17(4):481–486. doi:10.1002/rmb2.12229
72. Zhang X, Zhang W, Wang Z, et al. Enhanced glycolysis in granulosa cells promotes the activation of primordial follicles through mTOR signaling. Cell Death Dis. 2022;13(1):87. doi:10.1038/s41419-022-04541-1
73. Yang X, M A, Gegen T, Daoerji B, Zheng Y, Wang A. PHLPP1 inhibits the growth and aerobic glycolysis activity of human ovarian granular cells through inactivating AKT pathway. BMC Womens Health. 2024;24(1):25. doi:10.1186/s12905-023-02872-5
74. Zhang Q, Ren J, Wang F, et al. Mitochondrial and glucose metabolic dysfunctions in granulosa cells induce impaired oocytes of polycystic ovary syndrome through Sirtuin 3. Free Radic Biol Med. 2022;187:1–16. doi:10.1016/j.freeradbiomed.2022.05.010
75. Zhang Y, Liu L, Yin TL, Yang J, Xiong CL. Follicular metabolic changes and effects on oocyte quality in polycystic ovary syndrome patients. Oncotarget. 2017;8(46):80472–80480. doi:10.18632/oncotarget.19058
76. Chang AY, Lalia AZ, Jenkins GD, et al. Combining a nontargeted and targeted metabolomics approach to identify metabolic pathways significantly altered in polycystic ovary syndrome. Metabolism. 2017;71:52–63. doi:10.1016/j.metabol.2017.03.002
77. Zhao Y, Fu L, Li R, et al. Metabolic profiles characterizing different phenotypes of polycystic ovary syndrome: plasma metabolomics analysis. BMC Med. 2012;10(1):153. doi:10.1186/1741-7015-10-153
78. Liu K, Wei H, Nong W, et al. Nampt/SIRT2/LDHA pathway-mediated lactate production regulates follicular dysplasia in polycystic ovary syndrome. Free Radic Biol Med. 2024;225:776–793. doi:10.1016/j.freeradbiomed.2024.10.312
79. Hu R, Nong W, Huo P, et al. Dendrobium nobile-derived polysaccharides stimulate the glycolytic pathway by activating SIRT2 to regulate insulin resistance in polycystic ovary syndrome granulosa cells. Int J Biol Macromol. 2024;278(Pt 2):134780. doi:10.1016/j.ijbiomac.2024.134780
80. Liang A, Huang L, Liu H, et al. Resveratrol improves follicular development of PCOS rats by regulating the glycolytic pathway. Mol Nutr Food Res. 2021;65(24):e2100457. doi:10.1002/mnfr.202100457
81. Liang A, Zhang W, Wang Q, et al. Resveratrol regulates insulin resistance to improve the glycolytic pathway by activating SIRT2 in PCOS granulosa cells. Front Nutr. 2022;9:1019562. doi:10.3389/fnut.2022.1019562
82. Emidio GD, Placidi M, Rea F, et al. Methylglyoxal-dependent glycative stress and deregulation of SIRT1 functional network in the ovary of PCOS mice. Cells. 2020;9(1):209. doi:10.3390/cells9010209
83. Zhang S, Tu H, Yao J, et al. Combined use of Diane-35 and metformin improves the ovulation in the PCOS rat model possibly via regulating glycolysis pathway. Reprod Biol Endocrinol. 2020;18(1):58. doi:10.1186/s12958-020-00613-z
84. Min Z, Long X, Zhao H, et al. Protein lysine acetylation in ovarian granulosa cells affects metabolic homeostasis and clinical presentations of women with polycystic ovary syndrome. Front Cell Dev Biol. 2020;8:567028. doi:10.3389/fcell.2020.567028
85. Cao J, Huo P, Cui K, et al. Follicular fluid-derived exosomal miR-143-3p/miR-155-5p regulate follicular dysplasia by modulating glycolysis in granulosa cells in polycystic ovary syndrome. Cell Commun Signal. 2022;20(1):61. doi:10.1186/s12964-022-00876-6
86. Li Y, Tan Y. Bioinformatics analysis of ceRNA network related to polycystic ovarian syndrome. Comput Math Methods Med. 2021;2021:9988347. doi:10.1155/2021/9988347
87. Zhao R, Jiang Y, Zhao S, Zhao H. Multiomics analysis reveals molecular abnormalities in granulosa cells of women with polycystic ovary syndrome. Front Genet. 2021;12:648701. doi:10.3389/fgene.2021.648701
88. Cui X, Lei X, Huang T, et al. Follicular fluid-derived extracellular vesicles miR-34a-5p regulates granulosa cell glycolysis in polycystic ovary syndrome by targeting LDHA. J Ovarian Res. 2024;17(1):223. doi:10.1186/s13048-024-01542-w
89. Gu R, Dai F, Xiang C, et al. BMP4 participates in the pathogenesis of PCOS by regulating glucose metabolism and autophagy in granulosa cells under hyperandrogenic environment. J Steroid Biochem Mol Biol. 2023;235:106410. doi:10.1016/j.jsbmb.2023.106410
90. Jiao YY, Song N, Fang XY, et al. YTHDF2 regulates MSS51 expression contributing to mitochondria dysfunction of granulosa cells in polycystic ovarian syndrome patients. Mol Cell Endocrinol. 2024;592:112292. doi:10.1016/j.mce.2024.112292
91. Kitada M, Ogura Y, Koya D. The protective role of Sirt1 in vascular tissue: its relationship to vascular aging and atherosclerosis. Aging. 2016;8(10):2290–2307. doi:10.18632/aging.101068
92. Jalil AT, Zair MA, Hanthal ZR, et al. Role of the AMP-activated protein kinase in the pathogenesis of polycystic ovary syndrome. Indian J Clin Biochem. 2024;39(4):450–458. doi:10.1007/s12291-023-01139-y
93. Yaba A, Demir N. The mechanism of mTOR (mammalian target of rapamycin) in a mouse model of polycystic ovary syndrome (PCOS). J Ovarian Res. 2012;5(1):38. doi:10.1186/1757-2215-5-38
94. Song WJ, Shi X, Zhang J, Chen L, Fu SX, Ding YL. Akt-mTOR signaling mediates abnormalities in the proliferation and apoptosis of ovarian granulosa cells in patients with polycystic ovary syndrome. Gynecol Obstet Invest. 2018;83(2):124–132. doi:10.1159/000464351
95. Guo Z, Chen X, Feng P, Yu Q. Short-term rapamycin administration elevated testosterone levels and exacerbated reproductive disorder in dehydroepiandrosterone-induced polycystic ovary syndrome mice. J Ovarian Res. 2021;14(1):64. doi:10.1186/s13048-021-00813-0
96. Cai Z, He S, Li T, Zhao L, Zhang K. Plumbagin inhibits proliferation and promotes apoptosis of ovarian granulosa cells in polycystic ovary syndrome by inactivating PI3K/Akt/mTOR pathway. Anim Cells Syst. 2020;24(4):197–204. doi:10.1080/19768354.2020.1790416
97. Liu J, Wu DC, Qu LH, Liao HQ, Li MX. The role of mTOR in ovarian Neoplasms, polycystic ovary syndrome, and ovarian aging. Clin Anat. 2018;31(6):891–898. doi:10.1002/ca.23211
98. Fazleen NE, Whittaker M, Mamun A. Risk of metabolic syndrome in adolescents with polycystic ovarian syndrome: a systematic review and meta-analysis. Diabetes Metab Syndr. 2018;12(6):1083–1090. doi:10.1016/j.dsx.2018.03.014
99. Chantrapanichkul P, Indhavivadhana S, Wongwananuruk T, Techatraisak K, Dangrat C, Sa-Nga-Areekul N. Prevalence of type 2 diabetes mellitus compared between lean and overweight/obese patients with polycystic ovarian syndrome: a 5-year follow-up study. Arch Gynecol Obstet. 2020;301(3):809–816. doi:10.1007/s00404-019-05423-2
100. De Leo V, Musacchio MC, Morgante G, Piomboni P, Petraglia F. Metformin treatment is effective in obese teenage girls with PCOS. Hum Reprod. 2006;21(9):2252–2256. doi:10.1093/humrep/del185
101. Kurzthaler D, Hadziomerovic-Pekic D, Wildt L, Seeber BE. Metformin induces a prompt decrease in LH-stimulated testosterone response in women with PCOS independent of its insulin-sensitizing effects. Reprod Biol Endocrinol. 2014;12(1):98. doi:10.1186/1477-7827-12-98
102. Guan Y, Wang D, Bu H, Zhao T, Wang H. The effect of metformin on polycystic ovary syndrome in overweight women: a systematic review and meta-analysis of randomized controlled trials. Int J Endocrinol. 2020;2020:5150684. doi:10.1155/2020/5150684
103. Tao X, Cai L, Chen L, Ge S, Deng X. Effects of metformin and Exenatide on insulin resistance and AMPKalpha-SIRT1 molecular pathway in PCOS rats. J Ovarian Res. 2019;12(1):86. doi:10.1186/s13048-019-0555-8
104. Tao X, Chen L, Cai L, Ge S, Deng X. Regulatory effects of the AMPKalpha-SIRT1 molecular pathway on insulin resistance in PCOS mice: an in vitro and in vivo study. Biochem Biophys Res Commun. 2017;494(3–4):615–620. doi:10.1016/j.bbrc.2017.09.154
105. Reverchon M, Cornuau M, Cloix L, et al. Visfatin is expressed in human granulosa cells: regulation by metformin through AMPK/SIRT1 pathways and its role in steroidogenesis. Mol Hum Reprod. 2013;19(5):313–326. doi:10.1093/molehr/gat002
106. Chen Z, Wei H, Zhao X, et al. Metformin treatment alleviates polycystic ovary syndrome by decreasing the expression of MMP-2 and MMP-9 via H19/miR-29b-3p and AKT/mTOR/autophagy signaling pathways. J Cell Physiol. 2019;234(11):19964–19976. doi:10.1002/jcp.28594
107. Zhang Z, Huo P, Lei X, et al. Metformin activates SIRT2 to improve insulin resistance and promote granulosa cell glycolysis in a rat model of polycystic ovary syndrome. Reprod Biomed Online. 2024;50(5):104750. doi:10.1016/j.rbmo.2024.104750
108. Jin R, Chen A, Ye Y, et al. Effect of berberine combined with metformin on autophagy in polycystic ovary syndrome by regulating AMPK/AKT/mTOR pathway. Mol Reprod Dev. 2024;91(8):e23768. doi:10.1002/mrd.23768
109. Furat Rencber S, Kurnaz Ozbek S, Eraldemir C, et al. Effect of resveratrol and metformin on ovarian reserve and ultrastructure in PCOS: an experimental study. J Ovarian Res. 2018;11(1):55. doi:10.1186/s13048-018-0427-7
110. Huang L, Liang A, Li T, et al. Mogroside V improves follicular development and ovulation in young-adult PCOS rats induced by letrozole and high-fat diet through promoting glycolysis. Front Endocrinol. 2022;13:838204. doi:10.3389/fendo.2022.838204
111. Ren C, Zhang S, Ma J, et al. Nicotinamide mononucleotide alleviates bile acid metabolism and hormonal dysregulation in letrozole-induced PCOS mice. Biology. 2024;13(12):1028. doi:10.3390/biology13121028
112. Ochiai A, Kuroda K, Ozaki R, et al. Resveratrol inhibits decidualization by accelerating downregulation of the CRABP2-RAR pathway in differentiating human endometrial stromal cells. Cell Death Dis. 2019;10(4):276. doi:10.1038/s41419-019-1511-7
113. Lachmandas E, Eckold C, Bohme J, et al. Metformin alters human host responses to mycobacterium tuberculosis in healthy subjects. J Infect Dis. 2019;220(1):139–150. doi:10.1093/infdis/jiz064
114. Crowley MJ, Diamantidis CJ, McDuffie JR, et al. Clinical outcomes of metformin use in populations with chronic kidney disease, congestive heart failure, or chronic liver disease: a systematic review. Ann Intern Med. 2017;166(3):191–200. doi:10.7326/M16-1901
115. Mansour A, Samadi M, Sanginabadi M, et al. Effect of resveratrol on menstrual cyclicity, hyperandrogenism and metabolic profile in women with PCOS. Clin Nutr. 2021;40(6):4106–4112. doi:10.1016/j.clnu.2021.02.004
116. Hassan S, Shah M, Malik MO, Ehtesham E, Habib SH, Rauf B. Treatment with combined resveratrol and myoinositol ameliorates endocrine, metabolic alterations and perceived stress response in women with PCOS: a double-blind randomized clinical trial. Endocrine. 2023;79(1):208–220. doi:10.1007/s12020-022-03198-2
117. Ochiai A, Kuroda K, Ikemoto Y, et al. Influence of resveratrol supplementation on IVF-embryo transfer cycle outcomes. Reprod Biomed Online. 2019;39(2):205–210. doi:10.1016/j.rbmo.2019.03.205
118. Giuliani C, Iezzi M, Ciolli L, et al. Resveratrol has anti-thyroid effects both in vitro and in vivo. Food Chem Toxicol. 2017;107(Pt A):237–247. doi:10.1016/j.fct.2017.06.044
119. Yang W, Ma Y, Wu Y, Lei X, Zhang J, Li M. Study on the effects of Mogroside V in inhibiting NLRP3-mediated granulosa cell pyroptosis and insulin resistance to improve PCOS. J Ovarian Res. 2025;18(1):10. doi:10.1186/s13048-024-01563-5
120. Ahmed M, Riaz U, Lv H, et al. Nicotinamide mononucleotide restores NAD(+) levels to alleviate LPS-induced inflammation via the TLR4/NF-kappaB/MAPK signaling pathway in mice granulosa cells. Antioxidants. 2024;14(1). doi:10.3390/antiox14010039
121. Aflatounian A, Paris VR, Richani D, et al. Declining muscle NAD(+) in a hyperandrogenism PCOS mouse model: possible role in metabolic dysregulation. Mol Metab. 2022;65:101583. doi:10.1016/j.molmet.2022.101583
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Platelet-Activating Factor Promotes the Development of Non-Alcoholic Fatty Liver Disease
Yin H, Shi A, Wu J
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2003-2030
Published Date: 8 July 2022
Tumor Necrosis Factor Alpha Level in the Uterine Fluid of Patients with Polycystic Ovary Syndrome and Its Correlation with Clinical Parameters
Ha LX, Li WX, Du YD, Yuan YY, Qu XX
Journal of Inflammation Research 2022, 15:6015-6020
Published Date: 29 October 2022
Thyroid Hormones and Cardiometabolic Risk Factors in Saudi Women with Polycystic Ovary Syndrome: A Cross-Sectional Study
Saeed WM, Alsehli F
International Journal of Women's Health 2023, 15:1197-1203
Published Date: 26 July 2023
Correlation Between Hemoglobin Levels and Polycystic Ovary Syndrome Metabolic Disorder
Ha LX, Du YD, Qu XX, Wang JJ
Diabetes, Metabolic Syndrome and Obesity 2023, 16:3019-3027
Published Date: 28 September 2023
Correlation Between Serum Vitamin E and HOMA-IR in Patients with T2DM
Zhang J, Hou Y, Zhang Z, Shi Y, Wang Z, Song G
Diabetes, Metabolic Syndrome and Obesity 2024, 17:1833-1843
Published Date: 23 April 2024