Back to Journals » International Journal of General Medicine » Volume 19
Exosome-Mediated Macrophage Polarization in Gastric Cancer: Inflammatory and Neuroinflammatory Mechanisms and Therapeutic Potential
Received 10 February 2026
Accepted for publication 27 June 2026
Published 7 July 2026 Volume 2026:19 602593
DOI https://doi.org/10.2147/IJGM.S602593
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Xudong Zhu
Wei An1, Ze Jin2, Yan Li3
1Department of Otolaryngology, Dalian Municipal Hospital of Traditional Chinese Medicine, Dalian, 116013, People’s Republic of China; 2The Fifth Department of Acupuncture and Moxibustion, The Second Affiliated Hospital of Heilongjiang University of Traditional Chinese Medicine, Harbin, 150001, People’s Republic of China; 3Department of Otolaryngology, First Affiliated Hospital of Heilongjiang University of Traditional Chinese Medicine, Harbin, 150040, People’s Republic of China
Correspondence: Yan Li, Department of Otolaryngology, First Affiliated Hospital of Heilongjiang University of Traditional Chinese Medicine, Harbin, 150040, People’s Republic of China, Email [email protected]
Abstract: Gastric cancer progression is driven by intricate crosstalk among exosomes, macrophage polarization, inflammatory dysregulation, and tumor-associated neuroinflammation within the tumor microenvironment, which collectively form a reciprocal vicious cycle. As critical intercellular mediators, exosomes derived from tumor cells, immune cells, and stem cells deliver microRNA cargos to orchestrate M1/M2 macrophage balance and modulate neuroinflammatory states, exerting either pro-tumor or anti-tumor effects. M1 macrophage-derived exosomal miRNAs suppress PD-L1 and NLRP3 signaling, reduce pro-inflammatory cytokine release, ameliorate neuroinflammation, and enhance anti-tumor immunity. Conversely, tumor cell- and M2 macrophage-derived exosomes drive M2 polarization via the MAPK/ERK and PI3K/Akt pathways, amplifying the secretion of immunosuppressive cytokines and exacerbating neuroinflammatory progression. Stem cell- and traditional Chinese medicine-derived exosomes reverse immunosuppression by targeting the STAT3 pathway, polarize macrophages toward the M1 phenotype, and attenuate tumor-associated neuroinflammation. These regulatory events converge on the NF-κB and NLRP3 inflammasome pathways to shape the inflammatory and neuroinflammatory microenvironment. Preclinical evidence confirms that exosome-based interventions reduce tumor burden by remodeling macrophage phenotypes, restoring inflammatory homeostasis, and ameliorating neuroinflammation. This review identifies exosomal regulators as promising targets for rebalancing macrophage polarization, inflammation, and neuroinflammation, providing a framework for advancing gastric cancer therapeutics.
Keywords: exosome, gastric cancer, macrophage polarization, inflammation, miRNA, tumor microenvironment
Introduction
Gastric cancer ranks as the third leading cause of cancer-related deaths worldwide, with its incidence and mortality rates remaining persistently high among malignant tumors of the digestive system.1 Although surgical resection, chemotherapy (eg., cisplatin, oxaliplatin) and immune checkpoint inhibitors (PD-1/PD-L1 antibodies) have become first-line clinical treatments, the 5-year survival rate of patients with advanced disease is still less than 10%.2 The core bottlenecks include high therapeutic resistance (eg., the cisplatin resistance rate is as high as 60%), elevated tumor recurrence rate (the postoperative recurrence rate ranges from 40%–60%), and off-target toxicity caused by insufficient targeting ability.3–5 Notably, current monotherapies predominantly target tumor cells or single immune checkpoints, failing to disrupt the complex, multicellular immunosuppressive and inflammatory network within the tumor microenvironment (TME), which represents a fundamental reason for treatment failure.6 Accumulating evidence has indicated that the malignant progression of gastric cancer is closely associated with the synergistic effects of immune suppression, inflammatory dysregulation and nerve infiltration within the TME, and this complex network has become a critical obstacle that cannot be easily overcome by current therapeutic strategies.7 Anatomically, the stomach is densely innervated by both the intrinsic enteric nervous system (ENS, often called the “second brain” of the gut) and extrinsic vagal and sympathetic fibers, forming an extensive constitutive neuro-immune interface that renders gastric tissue inherently susceptible to inflammatory hijacking during malignant transformation.8
In the TME of gastric cancer, neuroinflammation is characterized by three core features: neurofibrosis, nerve infiltration, and the abnormal release of inflammatory factors (TNF-α, IL-6, NGF).9 Studies have confirmed that gastric cancer cells can induce the activation of Schwann cells and glial cells, promoting the infiltration of nerve fibers into tumor tissues. The released neurotransmitters such as norepinephrine can further activate the NF-κB pathway, amplifying the secretion of pro-inflammatory cytokines.10 In contrast, immune dysregulation exacerbates neuroinflammation through three distinct pathways: M2 macrophage polarization (elevated proportion of CD206+ cells), regulatory T cell (Treg) enrichment, and CD8+ T cell exhaustion (high PD-1 expression).11 Collectively, these processes form a vicious cycle where neuroinflammation, immune suppression, and gastric cancer progression interact reciprocally. For instance, IL-10 secreted by PD-1 tumor-associated macrophages (TAMs) can directly impair CD8+ T cell function while promoting glial cell activation, thereby accelerating tumor invasion.12
Exosomes, as the core mediators of intercellular communication in the TME, are widely derived from tumor cells, immune cells, mesenchymal stem cells, and extracts of traditional Chinese medicine.3,13,14 Their unique lipid bilayer structure enables them to stably load diverse cargos including nucleic acids (miRNA, circRNA, lncRNA), proteins (PD-L1, HMGB1), and lipids (ceramide),6 and to specifically target tumor tissues and immune cells via surface ligands (eg., CD63, integrins).15 Accumulating evidence has indicated that exosomes can transmit signals between immune cells and nerve-associated cells. For example, after miR-519a-3p derived from gastric cancer cell exosomes is internalized by intrahepatic macrophages, it induces M2 polarization, promotes angiogenesis, and accelerates liver metastasis.16 In contrast, miR-16-5p from M1 macrophage-derived exosomes can downregulate PD-L1 expression, activate CD8+ T cells, and simultaneously suppress the release of neuroinflammatory factors.17
Accumulating evidence has delineated individual roles of exosomes in either gastric cancer immunomodulation or inflammatory regulation, and prior work has confirmed that exosome-mediated intercellular crosstalk substantially shapes macrophage polarization states.7 However, three critical research gaps remain unaddressed. First, most studies have focused on isolated exosomal cargos or single macrophage subsets, lacking a systematic, integrated dissection of the full regulatory network linking exosome-mediated macrophage polarization, NLRP3 inflammasome activation, and tumor-associated neuroinflammation in gastric cancer.17 Second, the mechanistic crosstalk between exosome–macrophage interactions and neuroinflammatory progression remains poorly characterized, and the core signaling nodes that connect immunosuppression, inflammation, and neural invasion have not been systematically identified. Third, although both engineered and natural-derived exosomes show therapeutic promise, there is a lack of comparative analysis of their respective advantages, translational potential, and applicable scenarios between international and domestic research streams.
Targeting exosome–macrophage interactions carries profound clinical significance. Unlike conventional single-target agents, interventions modulating this axis can simultaneously reverse M2 macrophage-mediated immunosuppression, inhibit NLRP3-driven inflammatory amplification, and disrupt the neuroinflammation–tumor progression vicious cycle, thereby exerting multi-level anti-tumor effects.14,17 This regulatory axis also offers novel non-invasive biomarkers for early diagnosis and prognosis prediction, as well as actionable targets for overcoming chemoresistance and enhancing immunotherapy efficacy.3,18 From a distinctive interdisciplinary vantage point integrating otolaryngology and acupuncture and moxibustion, our team focuses on autonomic neural regulation and neuro-immune crosstalk in digestive malignancies. The stomach receives dense vagal and sympathetic innervation that is functionally linked to head and neck autonomic pathways; meanwhile, acupuncture and related neuromodulation approaches can modulate sympathetic norepinephrine release and restrain inflammatory cascade activation, which provides a unique rationale for focusing on the neuroinflammation-immune-exosome regulatory network in this review.19 This review systematically collates the molecular mechanisms underlying exosome-mediated regulation of the immune system and NLRP3 inflammasome pathway, elucidates the interaction network between neuroinflammation and immune dysregulation, and compares research advances achieved domestically and internationally, thereby providing a novel interdisciplinary perspective for gastric cancer therapy.
Overview of Exosomes and Immune System Regulation in Gastric Cancer
Exosomes are extracellular vesicles (EVs) with a diameter of 30–150 nm, whose biogenesis depends on membrane budding mediated by the endosomal sorting complex required for transport (ESCRT) and tetraspanins (CD63, CD9).15 In gastric cancer, exosome secretion is regulated by hypoxia, inflammatory factors, and epigenetic modifications. In a hypoxic microenvironment, gastric cancer cells upregulate RAB27A expression via hypoxia-inducible factor-1α (HIF-1α) to promote exosome release.20 Conversely, TNF-α enhances exosome secretion from M2 macrophages through the NF-κB pathway.21
Accumulating evidence has demonstrated that serum exosome concentrations in gastric cancer patients are significantly higher than those in healthy individuals, and these exosomes are enriched in pro-tumor cargos (eg., miR-21, MALAT1).5,22 In contrast, exosomes derived from normal gastric mucosal epithelial cells are predominantly enriched in anti-inflammatory cargos (eg., miR-451a).23
The core features of gastric cancer-associated neuroinflammation are characterized by nerve infiltration, dysregulation of the inflammatory cytokine network, and activation of the NLRP3 inflammasome. Specifically, the nerve fiber density within tumor tissues increases by 2–3-fold, the levels of pro-inflammatory cytokines including TNF-α, IL-1β, and IL-6 are significantly elevated, and IL-10 and TGF-β are abnormally enriched. Meanwhile, the expression of NLRP3, ASC, and caspase-1 in macrophages is also upregulated, thus promoting IL-1β secretion.9,24,25 The regulation of neuroinflammation by the immune system mainly relies on two pathways: innate immunity and adaptive immunity. Innate immunity involves macrophage polarization and neutrophil activation, whereas adaptive immunity is manifested as the imbalance of T cell subsets and dysfunction of B cells. Specifically, IL-10 secreted by M2 macrophages can induce glial cells to release nerve growth factor (NGF), thereby exacerbating neurofibrosis. In contrast, the exhaustion of CD8+ T cells leads to decreased IFN-γ secretion, which impairs the effective inhibition of neuroinflammatory factor release.11,26
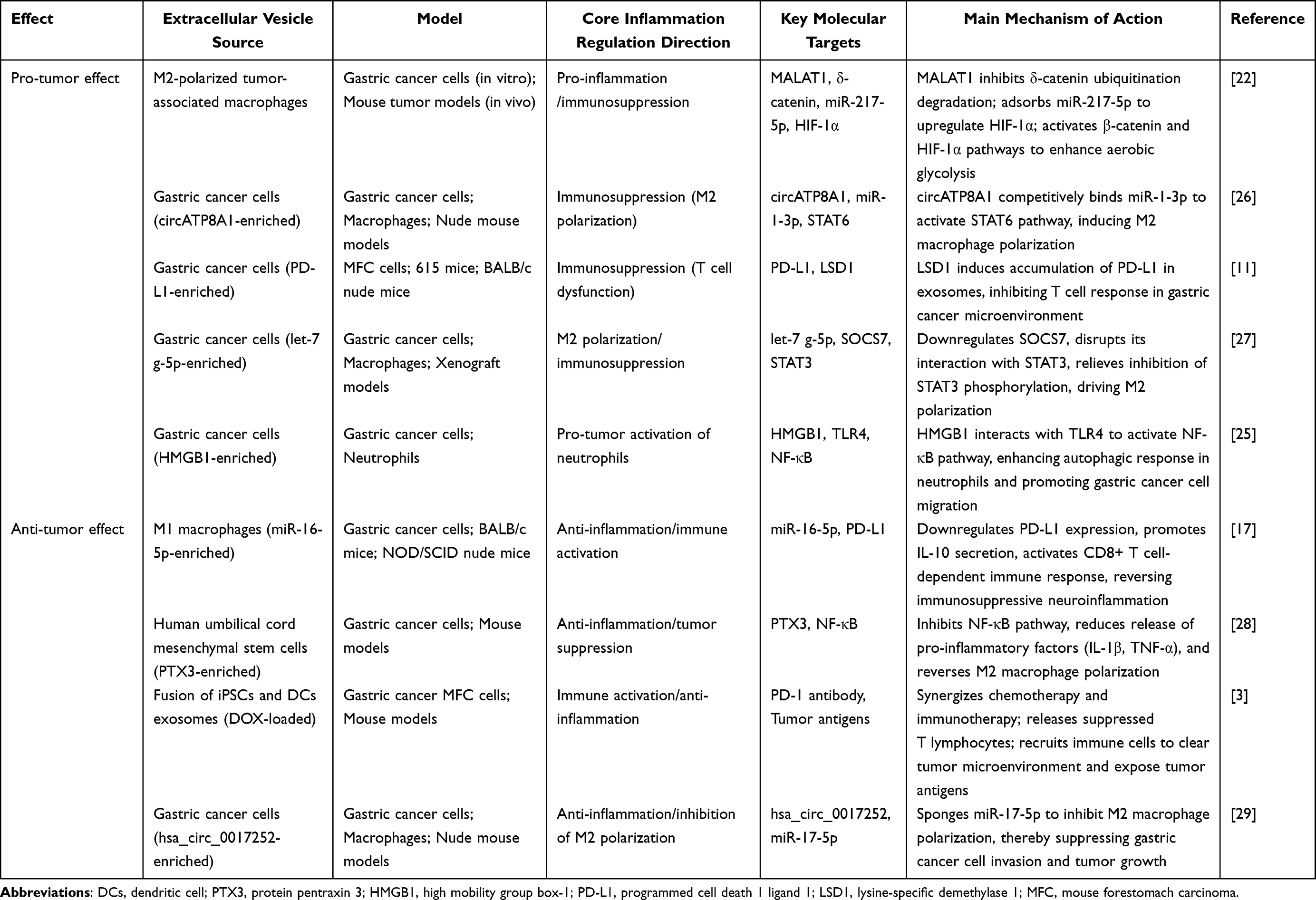 |
Table 1 Representative Study on the Dual Role of Extracellular Vesicles in Regulating the Immune System of Gastric Cancer |
Exosomes exhibit dual pro-tumor and anti-tumor properties in gastric cancer immunomodulation (Table 1). In the pro-tumor aspect, miR-552-5p derived from gastric cancer cell exosomes can target the PD-1/PD-L1 axis, inhibiting natural killer (NK) cell activity and promoting neuroinflammation.28 Conversely, miR-21 from M2 macrophage-derived exosomes enhances the chemotherapeutic resistance of gastric cancer cells via the PTEN/PI3K/Akt pathway.5 In the anti-tumor aspect, miR-16-5p from M1 macrophage-derived exosomes downregulates PD-L1 expression, thereby activating CD8+ T cells and suppressing neuroinflammation.17 Pentraxin 3 (PTX3) derived from mesenchymal stem cell (MSC)-derived exosomes can inhibit the NF-κB pathway, reducing the release of inflammatory factors such as TNF-α and IL-6.14
Notably, conflicting findings exist regarding MSC-derived exosomes: while umbilical cord MSC-derived exosomes carrying PTX3 exert robust anti-inflammatory and anti-tumor effects by inhibiting the NF-κB pathway,14 bone marrow MSC-derived exosomes overexpressing miR-1228 have been reported to promote gastric cancer cell proliferation via MMP-14-mediated PI3K/Akt regulation.30 This discrepancy indicates that the functional outcomes of stem cell-derived exosomes are highly context-dependent, varying with cargo composition, tissue origin, and tumor microenvironmental status.
The exertion of these dual properties is closely associated with the cellular origin of exosomes, cargo composition, and tumor stage. Exosomes in advanced gastric cancer are predominantly enriched in pro-tumor cargos, whereas immune cell-derived exosomes exert more prominent anti-tumor effects in early-stage gastric cancer.31
Mechanistic Specificity of Exosomes from Different Sources in Regulating Gastric Cancer Immunity and Anti-Inflammation
Immune Cell-Derived Exosomes
Immune cell-derived exosomes have emerged as a research hotspot in this field, among which macrophage-derived exosomes have attracted the most intensive research attention. Exosomes derived from M2-type tumor-associated macrophages (TAMs) can enrich MALAT1 to inhibit δ-catenin degradation and sequester miR-217-5p, thereby activating the β-catenin and HIF-1α pathways, enhancing glycolysis in gastric cancer cells, and promoting the release of pro-inflammatory cytokines (IL-1β, TNF-α).31 Meanwhile, miR-487a carried by these exosomes can target TIA1, facilitating M2 polarization and exacerbating neuroinflammation.32 Conversely, M1 macrophage-derived exosomes are abundant in miR-16-5p, which downregulates PD-L1 to promote IL-10 secretion, activate CD8+ T cell-dependent immune responses, and reverse immunosuppressive neuroinflammation.31 Dendritic cell-derived exosomes can induce IL-4 secretion by delivering tumor antigens, regulate the conversion of macrophages toward an anti-inflammatory phenotype, and inhibit the malignant progression of the inflammatory microenvironment in gastric cancer.3 Among neutrophil-derived exosomes, those from the N2 subset target SLIT2 via miR-4745-5p/3911 to enhance IL-8 release and accelerate angiogenesis, whereas miR-223-3p in exosomes from the N1 subset can serve as a diagnostic biomarker for gastric cancer.1,33
Tumor Cell-Derived Exosomes
Tumor cell-derived exosomes account for 32% of the total exosomes in the gastric cancer microenvironment, with their regulatory mechanisms primarily focused on inducing immunosuppression and neuroinflammation. CircATP8A1 in gastric cancer cell-derived exosomes competitively binds to miR-1-3p to activate the STAT6 pathway, thereby inducing M2 polarization of macrophages, promoting IL-10 secretion, and accelerating neurofibrosis.26 Meanwhile, miR-519a-3p carried by these exosomes targets DUSP2 to activate the MAPK/ERK pathway, facilitating the formation of pre-metastatic niches in the liver.16 In addition, gastric cancer cell-derived exosomes can directly interact with PD-1 on the surface of T cells by delivering PD-L1, or inhibit DUSP3 via miR-541-5p to activate STAT3, thereby maintaining an immunosuppressive inflammatory microenvironment.11,34 Notably, exosomes derived from Helicobacter pylori-infected gastric cancer cells can carry the CagA protein, which promotes endothelial cell inflammation by activating the STAT3 pathway and indirectly exacerbates neuroinflammation.10
Stem Cell-Derived Exosomes
Stem cell-derived exosomes account for 11% of exosomes in the gastric cancer microenvironment, predominantly derived from mesenchymal stem cells (MSCs), and exhibit significant anti-inflammatory and anti-tumor properties. Umbilical cord MSC-derived exosomes enhance targeting ability via engineered modification (neutrophil membrane fusion), and their carried PTX3 can inhibit the NF-κB pathway, reduce the levels of IL-1β and TNF-α, and reverse M2 macrophage polarization.14 Conversely, exosomes from miR-1228-overexpressing bone marrow MSCs have been reported to promote gastric cancer cell proliferation and invasive capacity via MMP-14-mediated activation of the PI3K/Akt pathway,30 which further demonstrates the functional heterogeneity of MSC-derived exosomes across different tissue origins. In contrast, exosomes fused with induced pluripotent stem cells (iPSCs) and dendritic cells (DCs) possess both tumor-targeting capability and immune factor recruitment ability. Under modification with PD-1 antibodies, these exosomes release suppressed T lymphocytes, promote the secretion of IL-4 and IL-10, and ameliorate the neuroinflammatory microenvironment.3
Exosomes from Other Sources
Traditional Chinese medicine (TCM)-derived exosomes have emerged as a distinctive research orientation in domestic studies. Exosomes derived from TAMs treated with Jianpi Fuzheng Decoction can inhibit miR-513b-5p, attenuate AKT/mTOR activation in a PTEN-dependent manner, and alleviate immunosuppression and neuroinflammation.35 Epigallocatechin gallate (EGCG) in exosomes isolated from the extract of Hedyotis diffusa Willd. Targets STAT3, suppresses PLXNC1-mediated release of exosomal miR-92b-5p, and reverses M2 polarization.36 Although research on nerve cell-derived exosomes in gastric cancer remains scarce, emerging evidence from gastrointestinal neuroimmunology suggests that glial cell-derived exosomal regulators can modulate the NF-κB pathway and suppress pro-inflammatory cytokine release, representing a promising yet underinvestigated axis in gastric tumor-associated neuroinflammation.37
 |
Table 2 Core Research on the Regulation of Immune and Anti-Inflammatory Effects in Gastric Cancer by Extracellular Vesicles from Different Sources |
Comparison of Immunoregulatory Characteristics of Exosomes From Different Sources
The core research on the regulation of gastric cancer immune and anti-inflammatory effects by extracellular vesicles from different sources is shown in Table 2. Immune cell-derived exosomes excel in directly regulating immune cell phenotypes but exhibit weak targeting ability. Tumor cell-derived exosomes possess strong targeting capability yet predominantly exert pro-tumor effects. Stem cell-derived exosomes demonstrate excellent synergy in anti-inflammation and anti-tumor activities with high biocompatibility. In contrast, TCM-derived exosomes integrate inherent anti-inflammatory activity with low toxicity.14,36 In terms of regulatory pathways, immune cell-derived exosomes mainly rely on miRNA/lncRNA-mediated regulation of the JAK/STAT and NF-κB pathways. Stem cell-derived exosomes predominantly mediate signal transduction via proteins (eg., PTX3) and lipids, while TCM-derived exosomes achieve multitarget regulation of the inflammatory network.30,35 These characteristic differences provide a basis for the clinical intervention strategy of gastric cancer: stem cell- or immune cell-derived exosomes are preferred for early-stage gastric cancer, whereas advanced gastric cancer requires combination with engineered TCM-derived exosomes to reverse the pro-tumor microenvironment.3
Mechanisms of Interaction Between Tumor-Associated Neuroinflammation and the Immune System in Gastric Cancer
Formation and Characteristics of Gastric Cancer-Associated Neuroinflammation
Given the dense enteric nerve plexuses and vagal innervation distributed throughout the gastric wall, malignant cells are anatomically positioned to interact with resident glial cells and nerve fibers from the earliest stages of tumorigenesis. Gastric cancer cells induce the activation of Schwann cells and enteric glial cells by secreting neurotrophic factors such as nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF), thereby promoting the infiltration of sympathetic and sensory nerve fibers into tumor tissues.9,19 Accumulating evidence has demonstrated that the incidence of nerve infiltration reaches 72% in advanced gastric cancer, and nerve fiber density is positively correlated with tumor invasion depth and lymph node metastasis.9,38 Mechanistically, circSMARCC1 in gastric cancer cell-derived exosomes upregulates U2AF2 expression to promote M2 polarization of macrophages and subsequent IL-1β secretion. The latter then activates glial cells to release NGF, accelerating nerve fiber hyperplasia.39 Meanwhile, norepinephrine released by nerve fibers can activate the PI3K/Akt pathway in gastric cancer cells via the β2-adrenergic receptor, facilitating epithelial-mesenchymal transition (EMT) and tumor metastasis.10,40
The core of gastric cancer-associated neuroinflammation lies in the synergistic amplification of pro-inflammatory cytokines and neurotrophic factors. Pro-inflammatory cytokines including TNF-α, IL-6, and IL-1β can activate the MAPK pathway in glial cells, thereby promoting the secretion of NGF and BDNF.24,41 Conversely, NGF can enhance the M2 polarization process of macrophages via the TrkA receptor, further facilitating the release of IL-10 and TGF-β, which consequently forms a positive feedback loop among pro-inflammatory cytokines, neurotrophic factors, and immunosuppression.26,42 The NLRP3 inflammasome plays a pivotal role in this regulatory network. HMGB1 derived from gastric cancer cell exosomes binds to TLR4 to activate NLRP3, promoting caspase-1-mediated IL-1β maturation. The mature IL-1β not only exacerbates neuroinflammation but also inhibits the function of CD8+ T cells.25,43 Clinical data have shown that serum levels of IL-1β and NGF are significantly elevated in gastric cancer patients and are positively correlated with each other, jointly predicting poor prognosis.44,45
Exacerbation of Neuroinflammation by Immune Dysregulation
Aberrations in innate immune cells serve as the core driver of aggravated neuroinflammation. M2-polarized cells constitute 60%–80% of the total tumor-associated macrophage (TAM) population in the gastric tumor microenvironment.46 The abundant IL-10 secreted by M2 TAMs not only directly suppresses CD8+ T cell effector function but also promotes the activation of enteric glial cells and the subsequent release of neurotrophic factors, which in turn fuels neuroinflammatory progression and nerve fiber hyperplasia.47 Meanwhile, CCL1 released during M2 polarization can recruit more immunosuppressive cells to amplify inflammatory signals.44,48 Exosomes derived from N2-subtype neutrophils carry miR-9-3p, which suppresses ACSL4 expression to block ferroptosis and enhance oxaliplatin resistance; additionally, the proteases released by these neutrophils disrupt the homeostasis of neural tissues, exacerbating nerve infiltration.49 As for natural killer (NK) cell dysfunction, it leads to reduced IFN-γ secretion, which impairs the inhibition of glial cell activation and further exacerbates neuroinflammation.28
Adaptive immune abnormalities exacerbate neuroinflammation through indirect regulation. Specifically, the proportion of regulatory T (Treg) cells among CD4+ T cells increases to 25% in gastric cancer, and the IL-10 and TGF-β secreted by these cells can directly inhibit CD8+ T cell activity while promoting nerve fiber hyperplasia.13 Excessive activation of Th17 cells leads to the release of IL-17, which can activate the NF-κB pathway, stimulate gastric cancer cells to secrete IL-6, and thereby induce glial cells to release NGF.50
Exosomes play a crucial mediating role in this process. MiR-135b-5p derived from gastric cancer cell exosomes targets SP1, impairs the function of Vγ9Vδ2 T cells, reduces the secretion of anti-inflammatory factors, and exacerbates neuroinflammation.51 In contrast, M1 macrophage-derived exosomes can reverse this process: they activate CD8+ T cells via miR-16-5p and inhibit neuroinflammation.17,52
Oncogenic Effects of the Vicious Cycle of Neuroinflammation and Immune Dysregulation
The vicious cycle of neuroinflammation and immune dysregulation promotes the progression of gastric cancer through multiple pathways (As shown in Figure 1). NGF released during neuroinflammation can activate the TrkA receptor on gastric cancer cells, facilitating EMT and angiogenesis.10,53 The exhaustion of CD8+ T cells caused by immunosuppression impairs the clearance of cancer cells in the inflammatory microenvironment; meanwhile, vascular endothelial growth factor (VEGF) secreted by M2 macrophages can accelerate nerve fiber hyperplasia.16 In addition, this cycle enhances oncogenic effects via metabolic reprogramming: the hypoxic microenvironment induced by neuroinflammation can promote the release of gastric cancer cell-derived exosomes, and their carried PKM2 activates the lipid synthesis pathway in macrophages, further inducing M2 polarization.18,54 Whereas lactate secreted by immunosuppressive cells can enhance the release of inflammatory factors from glial cells.22,55 Clinical studies have confirmed that the recurrence rate of gastric cancer patients with concurrent nerve infiltration and high M2 macrophage infiltration is 2.3-fold higher than that of patients with a single abnormality, and their 5-year survival rate is decreased by 31%.44,56
 |
Figure 1 Vicious Cycle of Neuroinflammation and Immune Dysregulation in Gastric Cancer. This diagram depicts the reciprocal positive feedback loop linking neuroinflammation. The left module (light red background) illustrates the neuroinflammatory cascade. Activated glial cells secrete pro-inflammatory mediators including interleukin (IL)-1β, tumor necrosis factor (TNF)-α, IL-6, and reactive oxygen species (ROS), which induce neuronal injury and further amplify neuroinflammatory signaling. Black curved arrows indicate glial activation and phenotypic polarization. The light blue panel shows immune dysregulation within the tumor microenvironment, consisting of myeloid-derived suppressor cells (MDSCs), regulatory T cells (Tregs) and M2-like macrophages. Inset circles illustrate molecular inhibitory pathways, where blunt-ended lines represent suppressive interactions targeting CD8⁺ T cells and natural killer (NK) cells via PD-L1/PD-1, CTLA-4 and adenosine signaling. Tumor-derived soluble factors TGF-β, IL-6 and PGE2 mediate crosstalk between the two compartments. The central red circular arrow represents the self-amplifying vicious cycle that drives tumor immune evasion and progression. Red curved arrows denote the direction of intercellular signal transduction. |
Anti-Tumor Mechanisms of Exosomes in Regulating the Immune System and Ameliorating Neuroinflammation
Immunoregulatory Cargos of Exosomes and Their Functions
MiRNAs represent the most predominant immunoregulatory cargos in exosomes. Anti-inflammatory miRNAs, such as miR-16-5p derived from M1 macrophage exosomes, exert their effects by downregulating PD-L1, activating CD8+ T cells and inhibiting IL-1β secretion.17,57 MiR-451a in serum exosomes can distinguish chronic atrophic gastritis from non-atrophic gastritis, and its low expression correlates with elevated levels of inflammatory factors. MiR-122-5p suppresses M2 macrophage polarization and neuroinflammation by inhibiting the STAT3 pathway.23,58 In contrast, pro-inflammatory miRNAs, such as miR-21 from M2 macrophage exosomes, target PTEN to activate the PI3K/Akt pathway, thereby promoting the secretion of IL-6 and TNF-α.5,59 MiR-519a-3p in gastric cancer cell-derived exosomes induces M2 polarization via the DUSP2/MAPK/ERK axis, accelerating neuroinflammation associated with liver metastasis.16,60 MiR-541-5p sustains JAK2/STAT3 activation by inhibiting DUSP3, further exacerbating immunosuppression and nerve fiber hyperplasia.34
Exosomal proteins exert their functions either by directly binding to receptors or regulating signaling pathways. The anti-inflammatory protein PTX3, derived from mesenchymal stem cell (MSC)-derived exosomes, can inhibit the NF-κB pathway, reduce the release of pro-inflammatory cytokines and NGF, and reverse neuroinflammation.14,61 ApoE from tumor-associated macrophage (TAM)-derived exosomes remodels the cytoskeleton of gastric cancer cells by activating the PI3K-Akt pathway, while its high expression is positively correlated with nerve infiltration.62 The immune checkpoint receptor PD-L1, carried by gastric cancer cell-derived exosomes, directly binds to PD-1 on T cells to suppress anti-tumor immunity, and simultaneously promotes glial cell activation.11 In contrast, CD47 on engineered exosomes enhances the phagocytic capacity of macrophages and reduces the secretion of inflammatory factors.3,63 The enzyme protein PKM2 from TAM-derived exosomes activates SREBP1-mediated lipid synthesis by inhibiting SCAP ubiquitination, thereby inducing M2 macrophage polarization; its serum level can serve as a diagnostic biomarker for gastric cancer.18
Although research on lipid cargos remains limited, it has been confirmed that ceramide from gastric cancer cell-derived exosomes can activate the TLR4/NF-κB pathway in macrophages, thereby promoting IL-1β secretion.25 In contrast, prostaglandin E2 (PGE2) from MSC-derived exosomes alleviates neuroinflammation by inhibiting TNF-α release.30 Other types of cargos include circRNAs and lncRNAs: circATP8A1 from gastric cancer cell-derived exosomes induces M2 polarization via the miR-1-3p/STAT6 axis.26 MALAT1, an lncRNA from M2 macrophage-derived exosomes, regulates glycolysis and enhances the secretion of pro-inflammatory cytokines.22 Additionally, CagA, a bacterial-derived cargo from exosomes of Helicobacter pylori-infected gastric cancer cells, promotes endothelial cell inflammation and nerve infiltration by activating the STAT3 pathway.10
Core Pathways of Exosomes in Regulating the Immune-Neuroinflammation Network
Exosomes remodel innate immunity and ameliorate neuroinflammation by regulating macrophage polarization, neutrophil activation, and natural killer (NK) cell function.64–66 MiR-16-5p derived from M1 macrophage exosomes downregulates PD-L1 expression, promotes IL-10 secretion, induces the phenotypic switch of M2 macrophages toward the M1 phenotype, and reduces NGF release.17 PTX3 from mesenchymal stem cell (MSC)-derived exosomes can directly inhibit the NF-κB pathway in neutrophils, decrease IL-8 secretion, and alleviate nerve fiber damage.14 Engineered exosomes loaded with NKG2D ligands activate NK cells, enhance their cytotoxicity against inflammatory macrophages, and reduce the levels of IL-1β and TNF-α.3,67 In addition, exosomes can ameliorate innate immune dysregulation by inhibiting NLRP3 inflammasome activation. For example, miR-16-5p from M1 macrophage exosomes suppresses NLRP3 expression, which reduces caspase-1 activation and IL-1β maturation.17
Exosomes break down immunosuppression and indirectly ameliorate neuroinflammation by regulating T cell subset balance and B cell function. Dendritic cell-derived exosomes deliver tumor antigens to induce the differentiation of CD4+ T cells into Th2 cells, increase the secretion of IL-4 and IL-10, and inhibit Th17 cell-mediated neuroinflammation.3,67 In contrast, M1 macrophage-derived exosomes can activate CD8+ T cells, enhance their secretion of IFN-γ and perforin, and eliminate cancer cells and activated glial cells in the inflammatory microenvironment.17,68 Engineered exosomes loaded with PD-1 antibodies can block the binding of PD-1/PD-L1, reverse CD8+ T cell exhaustion, and simultaneously reduce regulatory T (Treg) cell enrichment.3,69 Although research on B cell-derived exosomes remains limited, accumulating evidence has indicated that the immunoglobulins carried by these exosomes can enhance the phagocytic capacity of macrophages and reduce the secretion of pro-inflammatory cytokines.70
Exosomes directly inhibit neuroinflammation by specifically binding to glial cells or Schwann cells via surface ligands (eg., CD63, integrins). Epigallocatechin gallate (EGCG) in traditional Chinese medicine (TCM)-derived exosomes directly binds to STAT3 in glial cells, inhibiting its activation and the secretion of inflammatory factors.36 After engineering exosomes to modify nerve-specific ligands (eg., NGF receptor antibodies), their targeting efficiency is enhanced by 4–5-fold, which can significantly suppress nerve fiber hyperplasia and IL-6 release.3,71 In addition, exosomes may exert neuroinflammation-modulating effects by interacting with the autonomic nervous system. As sympathetic norepinephrine signaling has been confirmed to drive gastric cancer progression and inflammatory cascade activation,19 MSC-derived exosomes with anti-inflammatory properties are hypothesized to indirectly attenuate norepinephrine-mediated pro-tumor effects by remodeling the immunosuppressive microenvironment, though the direct neurotransmitter-regulating mechanism remains to be experimentally validated.
Exosomes can target the core nodes of the signaling axis composed of inflammatory factors, NLRP3, immunosuppression, and nerve infiltration, thereby breaking this vicious cycle. Their specific effects are manifested in several aspects: inhibiting the release of pro-inflammatory cytokines (eg., TNF-α, IL-6) to reduce glial cell activation; blocking the activation of the NLRP3 inflammasome to decrease IL-1β maturation; reversing immunosuppressive states such as M2 macrophage polarization and CD8+ T cell exhaustion to enhance anti-tumor immunity; and suppressing nerve infiltration and NGF secretion to reduce the recruitment of immunosuppressive cells.14,17,25,39 The mechanism of extracellular vesicles in anti-tumor treatment by regulating the immune system and improving neuroinflammation is shown in Figure 2.
 |
Figure 2 Anti-Tumor Mechanisms of Exosomes via Immune and Neuroinflammation Regulation. The top red panel depicts the pathological pro-tumor cascade. Tumor cell-derived pro-inflammatory exosomes activate microglia and macrophages to promote NLRP3 inflammasome assembly and pro-inflammatory cytokine secretion, which drives systemic immune dysregulation and tumor progression. The lower section contains three color-coded functional modules. The left green module shows engineered and mesenchymal stem cell-derived anti-inflammatory exosomes with RNA cargos crossing the blood-brain barrier. The middle green module illustrates intracellular mechanisms in macrophages. Delivered miR-146a suppresses NF-κB signaling and NLRP3 inflammasome activation, reduces astrocyte reactivity and restores T cell homeostasis. Red cross symbols indicate inhibitory effects. The right blue panel presents therapeutic outcomes. Ameliorated neuroinflammation and enhanced cytotoxic activity of T and NK cells synergistically trigger tumor apoptosis and regression. Arrows indicate the direction of signal transduction and process progression. |
Animal experiments have confirmed that engineered exosomes loaded with miR-16-5p can reduce the tumor volume of gastric cancer mice by 58%, decrease nerve fiber density by 42%, and enhance CD8+ T cell activity by 3-fold. When combined with PD-1 inhibitors, the therapeutic efficacy is further improved without significant toxicity observed.3,17
Current Research Status: China-Based Studies and Global Progress
To ensure the academic validity of this geographic comparative analysis, we established a standardized literature retrieval and classification framework. Literature was retrieved from PubMed, Embase, and CNKI databases, covering studies published from 2010 to 2025 using the keywords “gastric cancer”, “exosome”, “macrophage polarization”, “neuroinflammation”, and “engineered exosome”. Eligible studies were categorized into “domestic research” (first and corresponding authors affiliated with institutions in mainland China) and “international research” (authors affiliated with institutions outside mainland China). All included studies were further stratified into four research orientations — engineered exosome modification, natural-derived exosome application, fundamental mechanistic exploration, and preclinical/translational research — based on full-text content analysis, following established classification criteria from recent authoritative reviews in the field.2,7 This framework underpins the subsequent comparative analysis of research characteristics and developmental trends.
Global Research Progress
International research has focused on the precise regulation and in-depth mechanism elucidation of engineered exosomes. In terms of targeted design, surface modification of exosomes with gastric cancer-specific ligands (eg., Claudin18.2 antibody, HER2 ligand) can enhance the targeting efficiency to the tumor microenvironment (TME) by 3–5-fold.3,72 Loading combined cargos (eg., miR-16-5p and PD-L1 siRNA) exerts synergistic effects of immune activation, anti-inflammation and anti-tumor activity, improving chemotherapy sensitivity by 40% in mouse models.22,73 Mechanistically, a study by Wang et al clarified the molecular details of exosomal cargos regulating the NLRP3 inflammatory pathway: MALAT1 from M2 macrophage-derived exosomes upregulates HIF-1α and activates NLRP3 by sequestering miR-217-5p.22 HMGB1 from gastric cancer cell-derived exosomes promotes NLRP3 inflammasome assembly via the TLR4/NF-κB axis.25,74
Research Progress in China
Domestic research is characterized by naturally derived exosomes and translational applications, with remarkable advancements in traditional Chinese medicine (TCM)-derived exosomes. Exosomes from tumor-associated macrophages (TAMs) treated with Jianpi Fuzheng Decoction alleviate AKT/mTOR activation by inhibiting miR-513b-5p, reducing the tumor recurrence rate by 35% in mouse models.35,75 Epigallocatechin gallate (EGCG) in exosomes isolated from the extract of Hedyotis diffusa Willd. targets STAT3 to reverse M2 polarization and neuroinflammation, exhibiting anti-tumor efficacy comparable to cisplatin but with lower toxicity.36
Progress has also been made in translational research on stem cell-derived exosomes. After modification with neutrophil cell membranes, umbilical cord mesenchymal stem cell (MSC)-derived exosomes show significantly enhanced targeting ability, anti-inflammatory and anti-tumor effects in gastric cancer patient-derived xenograft (PDX) models, reducing tumor volume by 62%.14 In terms of technological innovation, domestic teams have developed a microfluidic chip-based technology for large-scale exosome production, increasing the yield by 10-fold compared with traditional ultracentrifugation methods while enhancing cargo stability.2 Meanwhile, liquid biopsy kits based on exosomal miR-223-3p and miR-122-5p have entered small-sample clinical validation, with an area under the curve (AUC) of 0.91 for distinguishing gastric cancer from benign gastroduodenal diseases.1,23,76
Comparison and Synergistic Directions Between China-Based and Global Research
Based on the aforementioned standardized classification framework, we conducted a multi-dimensional comparative analysis across four core aspects: technical route, target precision, mechanistic depth, and translational maturity. The advantages of international research lie in the development of engineered modification technologies and in-depth elaboration of mechanisms, while domestic research features unique strengths in the exploration of naturally derived exosomes and the application of translational technologies, forming a favorable complementary pattern. International research leans toward the precise regulation of single targets, whereas domestic research focuses more on synergistic anti-inflammatory pathways with multiple targets; international studies emphasize mechanistic interpretation at the basic level, while domestic research concentrates on clinical translation and implementation.
Synergistic directions can be advanced from multiple dimensions: combining engineered modification with natural anti-inflammatory activity (eg., surface modification of traditional Chinese medicine (TCM)-derived exosomes with Claudin18.2 antibodies) and integrating multi-omics technologies such as transcriptomics and proteomics to screen subtype-specific exosomal cargos.77 Additionally, conducting international multi-center clinical trials will help verify the efficacy of exosome-based therapies in gastric cancer patients of different ethnicities.
Challenges and Future Perspectives
Core Challenges in Clinical Translation
The targeting efficiency of current exosomes toward immune cells and nerve-associated cells in the gastric cancer tumor microenvironment (TME) is only 15%–25%, with these exosomes prone to being taken up by normal tissues such as the liver and spleen.3,78 For gastric cancer specifically, this delivery challenge is exacerbated by unique anatomical constraints: the multilayered gastric wall and its dense intrinsic enteric nervous system (ENS) embedded deep in the lamina propria create an additional barrier beyond the generic tumor extracellular matrix, requiring exosomes to cross both the blood-gastric tumor barrier and interstitial stroma to reach glial cells and Schwann cells at the neuro-immune interface.18,19 This hurdle is further aggravated by the lack of validated nerve-specific surface ligands and the non-specificity of conventional exosomal targeting moieties.2 In addition, molecular heterogeneity across gastric cancer subtypes (intestinal-type, diffuse-type) gives rise to divergent neuroinflammatory and immune profiles, meaning a one-size-fits-all exosome therapy cannot be universally applied.79
Furthermore, existing preclinical studies exhibit notable result inconsistencies and inherent methodological limitations that constrain reliable translational interpretation. First, divergent functional outcomes have been reported for exosomes from similar cellular origins, most prominently for mesenchymal stem cell (MSC)-derived exosomes: umbilical cord MSC-derived exosomes carrying PTX3 exert robust anti-tumor and anti-neuroinflammatory effects via NF-κB inhibition,14 whereas bone marrow MSC-derived exosomes overexpressing miR-1228 have been shown to promote gastric cancer cell proliferation through MMP-14-mediated PI3K/Akt activation.32 This discrepancy highlights that exosome function is highly dependent on tissue source, cargo composition, and tumor microenvironmental context, and cannot be generalized across all MSC subtypes or disease stages. Second, the vast majority of mechanistic studies rely on immortalized gastric cancer cell lines and immunodeficient xenograft mouse models, which lack intact enteric nervous system (ENS) structures and functional neuro-immune crosstalk, and thus cannot fully recapitulate the multicellular complexity of the human gastric tumor microenvironment.2 Third, substantial methodological heterogeneity persists across the field in exosome isolation protocols, dosage regimens, and administration routes, with no universally accepted standard operating procedure established to date. These constraints mean that many promising preclinical findings remain difficult to cross-validate and require rigorous confirmation in immunocompetent, ENS-intact animal models before advancing to clinical testing.
Beyond delivery efficiency and intratumoral heterogeneity, exosome-based therapies face additional safety and regimen optimization hurdles that limit their clinical translatability. First, exosomes derived from tumor cell sources may inherently encapsulate pro-tumorigenic cargos such as miR-21 and MALAT1, which carry a theoretical risk of promoting aberrant cell proliferation if production processes lack rigorous purification and quality control. Second, overly robust anti-inflammatory modulation driven by therapeutic exosomes carries the potential to dampen systemic immune surveillance, thereby increasing the risk of opportunistic infections.17 Finally, when exosome interventions are combined with conventional chemotherapy or immunomodulatory agents, overlapping off-target effects may lead to amplified systemic toxicity. Accordingly, the optimal dosage, administration route, and treatment schedule for combination regimens remain to be systematically characterized and standardized.
Future Research Directions
The strategy combining “gastric cancer-specific ligand modification and multi-target cargo combination” can be achieved by modifying exosome donor cells via CRISPR-Cas9 technology to express Claudin18.2 antibodies or HER2 ligands on their surface, while loading miR-16-5p, PD-L1 siRNA, and PTX3, thereby achieving the synergistic effects of “targeted enrichment, immune activation, anti-inflammation, and anti-tumor activity”.26,80,81 For different gastric cancer subtypes, screening of corresponding subtype-specific exosomal cargos is required: for example, miR-122-5p can be targeted for intestinal-type gastric cancer, while miR-223-3p is suitable for diffuse-type gastric cancer.1,23
Exosomes can be combined with chemotherapeutic drugs, immune checkpoint inhibitors, and anti-nerve infiltration drugs. Among these combinations, exosomes loaded with chemotherapeutic agents such as cisplatin can enhance the targeting ability of therapy and reduce off-target toxicity.3 The combination of exosomes with PD-1 inhibitors can reverse the immunosuppressive state, ameliorate neuroinflammation, and thereby improve the objective response rate.11 Moreover, the combination of exosomes with NGF inhibitors is able to block the vicious cycle between nerve infiltration and immunosuppression.9 In terms of interdisciplinary translational exploration, non-pharmacological neuromodulation strategies represented by acupuncture and vagus nerve stimulation represent promising complementary directions. These modalities can regulate sympathetic norepinephrine output and restore autonomic homeostasis, which may act synergistically with exosome-based interventions to concurrently remodel the immune microenvironment and attenuate neuroinflammatory progression.19 Accumulating preclinical evidence confirms that exosome-based immunotherapy combined with immune checkpoint inhibitors can significantly suppress gastric tumor growth in mouse models.17 When further coupled with neuromodulation strategies such as vagus nerve stimulation, this combinatorial regimen is hypothesized to synergistically remodel both the immune microenvironment and neuroinflammatory state to improve survival outcomes.19 However, this triple combination strategy remains at the conceptual stage, and its actual efficacy and survival benefit require dedicated in vivo validation studies.
Conduct Phase I/II clinical trials to evaluate the safety, tolerability and efficacy of exosome-based therapies in patients with advanced gastric cancer, with a focus on changes in neuroinflammatory biomarkers (IL-1β, NGF) and immune function indicators (CD8+ T cell activity, M2 macrophage proportion).82 Develop exosome-based liquid biopsy technologies for early diagnosis, therapeutic efficacy monitoring and prognostic evaluation of gastric cancer.1,23
Further explore the molecular mechanisms by which exosomes regulate the NLRP3 inflammatory pathway, and clarify the synergistic or antagonistic effects of different cargos (miRNAs, proteins).12 Elucidate the key signaling axis underlying the interaction between neuroinflammation and immune dysregulation, and identify novel therapeutic targets.10 Employ single-cell sequencing technology to uncover the regulatory differences of exosomes on distinct cell subsets within the TME.18
Conclusion
The core driver of gastric cancer malignant progression is the reciprocal vicious cycle among exosome-mediated intercellular communication, macrophage polarization dysregulation, NLRP3 inflammasome hyperactivation, and tumor-associated neuroinflammation and nerve infiltration. Distinct from existing reviews that predominantly focus on immune-exosome interactions alone, this work centers on the neuro-immune axis in the gastric tumor microenvironment as its core conceptual novelty. For the first time, it systematically integrates the full regulatory network linking exosomal cargo delivery, macrophage phenotypic reprogramming, and autonomic/sensory nerve crosstalk, and identifies the NLRP3 inflammasome as a converging node that bridges inflammatory amplification, immunosuppression, and neurogenic remodeling.17,37 This integrated framework fills critical gaps left by single-mechanism studies that overlook neuro-immune reciprocal regulation. Translationally, this neuro-immune-centered framework lays out a forward-looking, gastric-specific roadmap for the field rather than generic exosome development guidance. First, therapeutic exosomes can be engineered with ENS-targeting surface ligands to penetrate the blood-gastric tumor barrier and the dense interstitial stroma of the gastric wall, enabling direct modulation of glial cells and Schwann cells to disrupt the glia-macrophage positive feedback loop at its source. Second, liquid biopsy panels combining exosomal macrophage polarization markers and neuroinflammatory cargos (eg., miR-16-5p, IL-1β, NGF) can be developed to stratify patients with high perineural invasion risk, enabling precise patient selection for neuroimmune-targeted regimens. Third, exosome-based immunotherapy can be synergized with non-pharmacological neuromodulation approaches (eg., vagus nerve stimulation, acupuncture) to concurrently normalize autonomic tone and remodel the tumor microenvironment. This three-pronged strategy moves beyond conventional single-target paradigms, directly addresses the unique anatomical and pathophysiological challenges of gastric cancer, and holds substantial promise to overcome current therapeutic bottlenecks.
At the mechanistic core, the exosome-macrophage-neuroinflammation regulatory network operates through three hierarchically interconnected layers, with neuro-immune crosstalk as the definitive conceptual takeaway of this review. First, the NLRP3 inflammasome functions as the central converging node. HMGB1 carried by gastric cancer cell-derived exosomes activates the NLRP3 inflammasome via TLR4 to drive IL-1β maturation, which not only amplifies inflammatory cascades but also triggers glial cell activation and NGF release to fuel nerve fiber hyperplasia. conversely, miR-16-5p from M1 macrophage-derived exosomes directly suppresses NLRP3 expression to simultaneously block inflammatory amplification and neuroinflammatory progression. Second, exosomes bidirectionally regulate macrophage polarization states to shape the immune microenvironment: pro-tumor exosomes drive M2 polarization via MAPK/ERK and PI3K/Akt pathways to reinforce immunosuppression, while anti-tumor exosomes reverse this phenotypic shift to restore anti-tumor immunity. Third, autonomic and sensory nerve signals form a reciprocal regulatory loop with immune cells via exosomal mediators: norepinephrine released by sympathetic nerve fibers promotes gastric cancer cell EMT and exosome secretion through β2-adrenergic receptor signaling, and in turn, M2 macrophage-derived VEGF and IL-1β accelerate nerve innervation and glial cell activation. Exosomes from different cellular origins exhibit distinct regulatory properties for this neuro-immune axis, providing a mechanistic basis for personalized therapeutic regimens tailored to different disease stages.
Data Sharing Statement
No new data was produced by this review.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Yu D, Zhang J, Wang M, et al. Exosomal miRNAs from neutrophils act as accurate biomarkers for gastric cancer diagnosis. Clin Chim Acta. 2024;554:117773. doi:10.1016/j.cca.2024.117773
2. Bintintan V, Burz C, Pintea I, et al. The Importance of Extracellular Vesicle Screening in Gastric Cancer: a 2024 Update. Cancers. 2024;16(14):2574. doi:10.3390/cancers16142574
3. Li Y, Tian L, Zhao T, Zhang J. A nanotherapeutic system for gastric cancer suppression by synergistic chemotherapy and immunotherapy based on iPSCs and DCs exosomes. Cancer Immunol Immunother. 2023;72(6):1673–17. doi:10.1007/s00262-022-03355-6
4. Zhong J, Chen Y, Wang LJ. Emerging molecular basis of hematogenous metastasis in gastric cancer. World J Gastroenterol. 2016;22(8):2434–2440. doi:10.3748/wjg.v22.i8.2434
5. Zheng P, Chen L, Yuan X, et al. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J Exp Clin Cancer Res. 2017;36(1):53. doi:10.1186/s13046-017-0528-y
6. Tang L, Zhang W, Qi T, Jiang Z, Tang D. Exosomes play a crucial role in remodeling the tumor microenvironment and in the treatment of gastric cancer. Cell Commun Signal. 2025;23(1):82. doi:10.1186/s12964-024-02009-7
7. Qiu Y, Lu G, Li N, Hu Y, Tan H, Jiang C. Exosome-mediated communication between gastric cancer cells and macrophages: implications for tumor microenvironment. Front Immunol. 2024;15:1327281. doi:10.3389/fimmu.2024.1327281
8. Shamsi BH, Chatoo M, Xu XK, Xu X, Chen XQ. Versatile Functions of Somatostatin and Somatostatin Receptors in the Gastrointestinal System. Front Endocrinol (Lausanne). 2021;12:652363. doi:10.3389/fendo.2021.652363
9. Xu X, Cheng J, Luo S, et al. Deoxycholic acid-stimulated macrophage-derived exosomes promote spasmolytic polypeptide-expressing metaplasia in the stomach. Biochem Biophys Res Commun. 2020;524(3):649–655. doi:10.1016/j.bbrc.2020.01.159
10. Tahmina K, Hikawa N, Takahashi-Kanemitsu A, et al. Transgenically expressed Helicobacter pylori CagA in vascular endothelial cells accelerates arteriosclerosis in mice. Biochem Biophys Res Commun. 2022;618:79–85. doi:10.1016/j.bbrc.2022.06.010
11. Shen DD, Pang JR, Bi YP, et al. LSD1 deletion decreases exosomal PD-L1 and restores T-cell response in gastric cancer. Mol Cancer. 2022;21(1):75. doi:10.1186/s12943-022-01557-1
12. Xue A, Cao Y, Yu K, et al. Disrupting of tissue-resident FOLR2+tumour-associated macrophages-derived interleukin-10 improves anti-PD-1 efficacy in gastric cancer. Int J Surg. 2025. doi:10.1097/JS9.0000000000004105
13. Liu J, Wu S, Zheng X, et al. Immune suppressed tumor microenvironment by exosomes derived from gastric cancer cells via modulating immune functions. Sci Rep. 2020;10(1):14749. doi:10.1038/s41598-020-71573-y
14. Tang Y, Shen Y, Zang X, et al. Neutrophil membrane engineered human umbilical cord MSC-derived sEVs enhance anti-tumor efficacy for gastric cancer via delivering pentraxin 3. J Control Release. 2025;383:113828. doi:10.1016/j.jconrel.2025.113828
15. Pols MS, Klumperman J. Trafficking and function of the tetraspanin CD63. Exp Cell Res. 2009;315(9):1584–1592. doi:10.1016/j.yexcr.2008.09.020
16. Qiu S, Xie L, Lu C, et al. Gastric cancer-derived exosomal miR-519a-3p promotes liver metastasis by inducing intrahepatic M2-like macrophage-mediated angiogenesis. J Exp Clin Cancer Res. 2022;41(1):296. doi:10.1186/s13046-022-02499-8
17. Li Z, Suo B, Long G, et al. Exosomal miRNA-16-5p Derived From M1 Macrophages Enhances T Cell-Dependent Immune Response by Regulating PD-L1 in Gastric Cancer. Front Cell Dev Biol. 2020;8:572689. doi:10.3389/fcell.2020.572689
18. Yuan M, Zheng X, Zheng S, et al. Exosomal PKM2: a Noninvasive Diagnostic Marker Linking Macrophage Metabolic Reprogramming to Gastric Cancer Pathogenesis. Cancer Sci. 2025;116(6):1537–1549. doi:10.1111/cas.70056
19. Erin N, Shurin GV, Baraldi JH, Shurin MR. Regulation of Carcinogenesis by Sensory Neurons and Neuromediators. Cancers. 2022;14(9):2333. doi:10.3390/cancers14092333
20. Gupta S, Rawat S, Krishnakumar V, Rao EP, Mohanty S. Hypoxia preconditioning elicit differential response in tissue-specific MSCs via immunomodulation and exosomal secretion. Cell Tissue Res. 2022;388(3):535–548. doi:10.1007/s00441-022-03615-y
21. Li L, Jiang W, Yu B, et al. Quercetin improves cerebral ischemia/reperfusion injury by promoting microglia/macrophages M2 polarization via regulating PI3K/Akt/NF-κB signaling pathway. Biomed Pharmacother. 2023;168:115653. doi:10.1016/j.biopha.2023.115653
22. Wang Y, Zhang J, Shi H, et al. M2 Tumor-Associated Macrophages-Derived Exosomal MALAT1 Promotes Glycolysis and Gastric Cancer Progression. Adv Sci. 2024;11(24):e2309298. doi:10.1002/advs.202309298
23. Liu H, Li PW, Yang WQ, et al. Identification of non-invasive biomarkers for chronic atrophic gastritis from serum exosomal microRNAs. BMC Cancer. 2019;19(1):129. doi:10.1186/s12885-019-5328-7
24. Wu L, Zhang X, Zhang B, et al. Exosomes derived from gastric cancer cells activate NF-κB pathway in macrophages to promote cancer progression. Tumour Biol. 2016;37(9):12169–12180. doi:10.1007/s13277-016-5071-5
25. Zhang X, Shi H, Yuan X, Jiang P, Qian H, Xu W. Tumor-derived exosomes induce N2 polarization of neutrophils to promote gastric cancer cell migration. Mol Cancer. 2018;17(1):146. doi:10.1186/s12943-018-0898-6
26. Deng C, Huo M, Chu H, et al. Exosome circATP8A1 induces macrophage M2 polarization by regulating the miR-1-3p/STAT6 axis to promote gastric cancer progression. Mol Cancer. 2024;23(1):49. doi:10.1186/s12943-024-01966-4
27. Ye Z, Yi J, Jiang X, et al. Gastric cancer-derived exosomal let-7 g-5p mediated by SERPINE1 promotes macrophage M2 polarization and gastric cancer progression. J Exp Clin Cancer Res. 2025;44(1):2. doi:10.1186/s13046-024-03269-4
28. Tang CW, Yang JH, Qin JW, et al. Regulation of the PD-1/PD-L1 Axis and NK Cell Dysfunction by Exosomal miR-552-5p in Gastric Cancer. Dig Dis Sci. 2024;69(9):3276–3289. doi:10.1007/s10620-024-08536-0
29. Song J, Xu X, He S, et al. Exosomal hsa_circ_0017252 attenuates the development of gastric cancer via inhibiting macrophage M2 polarization. Hum Cell. 2022;35(5):1499–1511. doi:10.1007/s13577-022-00739-9
30. Chang L, Gao H, Wang L, et al. Exosomes derived from miR-1228 overexpressing bone marrow-mesenchymal stem cells promote growth of gastric cancer cells. Aging. 2021;13(8):11808–11821. doi:10.18632/aging.202878
31. Wang F, Li B, Wei Y, et al. Correction: tumor-derived exosomes induce PD1(+) macrophage population in human gastric cancer that promotes disease progression. Oncogenesis. 2022;11(1):7. doi:10.1038/s41389-022-00381-y
32. Yang X, Cai S, Shu Y, et al. Exosomal miR-487a derived from m2 macrophage promotes the progression of gastric cancer. Cell Cycle. 2021;20(4):434–444. doi:10.1080/15384101.2021.1878326
33. Zhang J, Yu D, Ji C, et al. Exosomal miR-4745-5p/3911 from N2-polarized tumor-associated neutrophils promotes gastric cancer metastasis by regulating SLIT2. Mol Cancer. 2024;23(1):198. doi:10.1186/s12943-024-02116-6
34. Xiao H, Fu J, Liu R, Yan L, Zhou Z, Yuan J. Gastric cancer cell-derived exosomal miR-541-5p induces M2 macrophage polarization through DUSP3/JAK2/STAT3 pathway. BMC Cancer. 2024;24(1):957. doi:10.1186/s12885-024-12672-1
35. Zhang R, Chen X, Miao C, et al. Tumor-associated macrophage-derived exosomal miR-513b-5p is a target of jianpi yangzheng decoction for inhibiting gastric cancer. J Ethnopharmacol. 2024;318:117013. doi:10.1016/j.jep.2023.117013
36. Yi J, Ye Z, Xu H, et al. EGCG targeting STAT3 transcriptionally represses PLXNC1 to inhibit M2 polarization mediated by gastric cancer cell-derived exosomal miR-92b-5p. Phytomedicine. 2024;135:156137. doi:10.1016/j.phymed.2024.156137
37. Sun Q, Bai Z, Sun J, et al. Phosphorylation of AEG-1 suppresses NF-κB signaling in gastric cancer. Mol Biol Rep. 2026;53:1.
38. Jarabin JA, Klivényi P, Tiszlavicz L, et al. Late simultaneous carcinomatous meningitis, temporal bone infiltrating macro-metastasis and disseminated multi-organ micro-metastases presenting with mono-symptomatic vertigo - a clinico-pathological case reporT. Ideggyogy. 2020;73(9–10):354–360. doi:10.18071/ISZ.73.0354
39. Zhang C, Ye J, Guan Q, Fang X. Gastric Cancer Cell-Derived Exosomes Induce Macrophage M2 Polarization by Delivering circSMARCC1 to Promote Gastric Cancer Progression. IUBMB Life. 2025;77(10):e70062. doi:10.1002/iub.70062
40. Li D, Xia L, Huang P, et al. Cancer-associated fibroblast-secreted IGFBP7 promotes gastric cancer by enhancing tumor associated macrophage infiltration via FGF2/FGFR1/PI3K/AKT axis. Cell Death Discov. 2023;9(1):17. doi:10.1038/s41420-023-01336-x
41. Colardo M, Petraroia M, Lerza L, et al. NGF Modulates Cholesterol Metabolism and Stimulates ApoE Secretion in Glial Cells Conferring Neuroprotection against Oxidative Stress. Int J Mol Sci. 2022;23(9):4842. doi:10.3390/ijms23094842
42. Zhu D, Yang N, Liu YY, Zheng J, Ji C, Zuo PP. M2 Macrophage Transplantation Ameliorates Cognitive Dysfunction in Amyloid-β-Treated Rats Through Regulation of Microglial Polarization. J Alzheimers Dis. 2016;52(2):483–495. doi:10.3233/JAD-151090
43. Lei C, Chen K, Gu Y, et al. HMGB1/TLR4 axis promotes pyroptosis after ICH by activating the NLRP3 inflammasome. J Neuroimmunol. 2024;393:578401. doi:10.1016/j.jneuroim.2024.578401
44. Wang F, Li B, Wei Y, et al. Tumor-derived exosomes induce PD1(+) macrophage population in human gastric cancer that promotes disease progression. Oncogenesis. 2018;7(5):41. doi:10.1038/s41389-018-0049-3
45. Tas F, Tilgen Yasasever C, Karabulut S, Tastekin D, Duranyildiz D. Clinical significance of serum interleukin-18 (IL-18) levels in patients with gastric cancer. Biomed Pharmacother. 2015;70:19–23. doi:10.1016/j.biopha.2014.12.040
46. Boutilier AJ, Elsawa SF. Macrophage Polarization States in the Tumor Microenvironment. Int J Mol Sci. 2021;22(13):6995. doi:10.3390/ijms22136995
47. Zhuang H, Dai X, Zhang X, Mao Z, Huang H. Sophoridine suppresses macrophage-mediated immunosuppression through TLR4/IRF3 pathway and subsequently upregulates CD8(+) T cytotoxic function against gastric cancer. Biomed Pharmacother. 2020;121:109636. doi:10.1016/j.biopha.2019.109636
48. Liu S, Zhang Z, Wang Y, et al. The chemokine CCL1 facilitates pulmonary fibrosis by promoting macrophage migration and M2 polarization. Int Immunopharmacol. 2023;120:110343. doi:10.1016/j.intimp.2023.110343
49. Dong X, Xu J, Zhang J, et al. Exosomes from tumor-associated neutrophils suppress ferroptosis and confer chemoresistance in gastric cancer via miR-9-3p/ACSL4 axis. Free Radic Biol Med. 2025;239:49–62. doi:10.1016/j.freeradbiomed.2025.07.022
50. Liu F, Bu Z, Zhao F, Xiao D. Increased T-helper 17 cell differentiation mediated by exosome-mediated micro RNA-451 redistribution in gastric cancer infiltrated T cells. Cancer Sci. 2018;109(1):65–73. doi:10.1111/cas.13429
51. Li J, Sun L, Chen Y, et al. Gastric cancer-derived exosomal miR-135b-5p impairs the function of Vγ9Vδ2 T cells by targeting specificity protein 1. Cancer Immunol Immunother. 2022;71(2):311–325. doi:10.1007/s00262-021-02991-8
52. Zheng Y, Li Y, Wei Z, et al. HUC-MSC-derived exosomal miR-16-5p attenuates inflammation via dual suppression of M1 macrophage polarization and Th1 differentiation. Biochem Biophys Rep. 2025;43:102078. doi:10.1016/j.bbrep.2025.102078
53. Lin C, Ren Z, Yang X, et al. Nerve growth factor (NGF)-TrkA axis in head and neck squamous cell carcinoma triggers EMT and confers resistance to the EGFR inhibitor erlotinib. Cancer Lett. 2020;472:81–96. doi:10.1016/j.canlet.2019.12.015
54. Zhu S, Guo Y, Zhang X, et al. Pyruvate kinase M2 (PKM2) in cancer and cancer therapeutics. Cancer Lett. 2021;503:240–248. doi:10.1016/j.canlet.2020.11.018
55. Wei L, Yang X, Wang J, et al. H3K18 lactylation of senescent microglia potentiates brain aging and Alzheimer’s disease through the NFκB signaling pathway. J Neuroinflammation. 2023;20(1):208. doi:10.1186/s12974-023-02879-7
56. Chen Y, Wang T, Liang F, et al. Nicotinamide phosphoribosyltransferase prompts bleomycin-induced pulmonary fibrosis by driving macrophage M2 polarization in mice. Theranostics. 2024;14(7):2794–2815. doi:10.7150/thno.94482
57. Chen HL, Luo YP, Lin MW, et al. Serum exosomal miR-16-5p functions as a tumor inhibitor and a new biomarker for PD-L1 inhibitor-dependent immunotherapy in lung adenocarcinoma by regulating PD-L1 expression. Cancer Med. 2022;11:2627–2643. doi:10.1002/cam4.4638
58. Liu Y, Chen W, Chen J, et al. miR-122-5p regulates hepatocytes damage caused by BaP and DBP co-exposure through SOCS1/STAT3 signaling in vitro. Ecotoxicol Environ Saf. 2021;223:112570. doi:10.1016/j.ecoenv.2021.112570
59. Zhong Y, Yu F, Yang L, et al. HOXD9/miR-451a/PSMB8 axis is implicated in the regulation of cell proliferation and metastasis via PI3K/AKT signaling pathway in human anaplastic thyroid carcinoma. J Transl Med. 2023;21(1):817. doi:10.1186/s12967-023-04538-0
60. Zhang J, Fu L, Zhang J, et al. Inhibition of MicroRNA-122-5p Relieves Myocardial Ischemia-Reperfusion Injury via SOCS1. Hamostaseologie. 2023;43(04):271–280. doi:10.1055/a-2013-0336
61. Sun Y, Liu L, Yang R. PTX3 promotes IVIG resistance-induced endothelial injury in Kawasaki disease by regulating the NF-κB pathway. Open Life Sci. 2023;18(1):20220735. doi:10.1515/biol-2022-0735
62. Zheng P, Luo Q, Wang W, et al. Tumor-associated macrophages-derived exosomes promote the migration of gastric cancer cells by transfer of functional Apolipoprotein E. Cell Death Dis. 2018;9(4):434. doi:10.1038/s41419-018-0465-5
63. Chen Y, Zhu X, Liu H, et al. The application of HER2 and CD47 CAR-macrophage in ovarian cancer. J Transl Med. 2023;21(1):654. doi:10.1186/s12967-023-04479-8
64. He Z, Wang J, Zhu C, et al. Exosome-derived FGD5-AS1 promotes tumor-associated macrophage M2 polarization-mediated pancreatic cancer cell proliferation and metastasis. Cancer Lett. 2022;548:215751. doi:10.1016/j.canlet.2022.215751
65. Wang J, Tang W, Yang M, et al. Inflammatory tumor microenvironment responsive neutrophil exosomes-based drug delivery system for targeted glioma therapy. Biomaterials. 2021;273:120784. doi:10.1016/j.biomaterials.2021.120784
66. Ma F, Wang X, Qiu Z, et al. NK-derived exosome miR-1249-3p inhibits Mycobacterium tuberculosis survival in macrophages by targeting SKOR1. Cytokine. 2024;175:156481. doi:10.1016/j.cyto.2023.156481
67. Desimio MG, Covino DA, Doria M. Potential of the NKG2D/NKG2DL Axis in NK Cell-Mediated Clearance of the HIV-1 Reservoir. Int J Mol Sci. 2019;20(18):4490. doi:10.3390/ijms20184490
68. Orecchioni M, Ghosheh Y, Pramod AB, Ley K. Macrophage Polarization: different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front Immunol. 2019;10:1084. doi:10.3389/fimmu.2019.01084
69. Chen G, Huang AC, Zhang W, et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature. 2018;560(7718):382–386. doi:10.1038/s41586-018-0392-8
70. Baghaei K, Tokhanbigli S, Asadzadeh H, Nmaki S, Reza Zali M, Hashemi SM. Exosomes as a novel cell-free therapeutic approach in gastrointestinal diseases. J Cell Physiol. 2019;234(7):9910–9926. doi:10.1002/jcp.27934
71. Lian M, Qiao Z, Qiao S, et al. Nerve Growth Factor-Preconditioned Mesenchymal Stem Cell-Derived Exosome-Functionalized 3D-Printed Hierarchical Porous Scaffolds with Neuro-Promotive Properties for Enhancing Innervated Bone Regeneration. ACS Nano. 2024;18(10):7504–7520. doi:10.1021/acsnano.3c11890
72. Tsutsumi C, Ohuchida K, Yamada Y, et al. Claudin18.2-positive gastric cancer-specific changes in neoadjuvant chemotherapy-driven immunosuppressive tumor microenvironment. Br J Cancer. 2025;132(9):793–804. doi:10.1038/s41416-025-02981-y
73. Sun J, Lian M, Ma H, et al. Competing endogenous RNA network analysis of CD274, IL‑10 and FOXP3 co‑expression in laryngeal squamous cell carcinoma. Mol Med Rep. 2018;17(3):3859–3869. doi:10.3892/mmr.2017.8307
74. Zhao C, Mu M, Li X, et al. USP50 regulates NLRP3 inflammasome activation in duodenogastric reflux-induced gastric tumorigenesis. Front Immunol. 2024;15:1326137. doi:10.3389/fimmu.2024.1326137
75. Liu H, Zhang L, Ding X, Sui X. LINC00861 inhibits the progression of cervical cancer cells by functioning as a ceRNA for miR‑513b‑5p and regulating the PTEN/AKT/mTOR signaling pathway. Mol Med Rep. 2021;23:1.
76. Zhu Y, Li K, Yan L, He Y, Wang L, Sheng L. miR-223-3p promotes cell proliferation and invasion by targeting Arid1a in gastric cancer. Acta Biochim Biophys Sin. 2020;52(2):150–159. doi:10.1093/abbs/gmz151
77. Kubota Y, Shitara K. Zolbetuximab for Claudin18.2-positive gastric or gastroesophageal junction cancer. Ther Adv Med Oncol. 2024;16:17588359231217967. doi:10.1177/17588359231217967
78. Yasuda T, Wang YA. Gastric cancer immunosuppressive microenvironment heterogeneity: implications for therapy development. Trends Cancer. 2024;10(7):627–642. doi:10.1016/j.trecan.2024.03.008
79. Yu M, Jin Y, Yuan K, et al. Effects of exosomes and inflammatory response on tumor: a bibliometrics study and visualization analysis via CiteSpace and VOSviewer. J Cancer Res Clin Oncol. 2024;150(8):405. doi:10.1007/s00432-024-05915-y
80. Liu Y, Cao J, Zhu L, et al. Circular RNA circPGD contributes to gastric cancer progression via the sponging miR-16-5p/ABL2 axis and encodes a novel PGD-219aa protein. Cell Death Discov. 2022;8(1):384. doi:10.1038/s41420-022-01177-0
81. Wills CA, Liu X, Chen L, et al. Chemotherapy-Induced Upregulation of Small Extracellular Vesicle-Associated PTX3 Accelerates Breast Cancer Metastasis. Cancer Res. 2021;81(2):452–463. doi:10.1158/0008-5472.CAN-20-1976
82. Zhang J, Fu L, Yasuda-Yoshihara N, et al. IL-1β derived from mixed-polarized macrophages activates fibroblasts and synergistically forms a cancer-promoting microenvironment. Gastric Cancer. 2023;26(2):187–202. doi:10.1007/s10120-022-01352-3
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Differential Expression of Serum Exosomal Hsa-miR-487b-3p in Progressive Vitiligo Before and After Systemic Corticosteroid Treatment
Luo H, Xie B, Xu J, Zhu Y, Sun J, Shen Y, Song X
Clinical, Cosmetic and Investigational Dermatology 2022, 15:1377-1386
Published Date: 18 July 2022
Exosomes Derived hsa-miR-4669 as a Novel Biomarker for Early Predicting the Response of Subcutaneous Immunotherapy in Pediatric Allergic Rhinitis
Jiang S, Xie S, Fan R, Tang Q, Zhang H, Wang F, Xie S, Gao K, Zhang J, Xie Z, Jiang W
Journal of Inflammation Research 2022, 15:5063-5074
Published Date: 3 September 2022
Theaflavin-3,3’-Digallate Ameliorates Collagen-Induced Arthritis Through Regulation of Autophagy and Macrophage Polarization

Zhang L, Li W, Hou Z, Wang Z, Zhang W, Liang X, Wu Z, Wang T, Liu X, Peng X, Yang X, Yang H, Geng D
Journal of Inflammation Research 2023, 16:109-126
Published Date: 10 January 2023
Accelerated Wound Healing in Diabetic Rat by miRNA-185-5p and Its Anti-Inflammatory Activity
Wang KX, Zhao LL, Zheng LT, Meng LB, Jin L, Zhang LJ, Kong FL, Liang F
Diabetes, Metabolic Syndrome and Obesity 2023, 16:1657-1667
Published Date: 7 June 2023
A Novel Neutrophil Extracellular Traps Signature for Overall Survival Prediction and Tumor Microenvironment Identification in Gastric Cancer
Qu Z, Han Y, Zhu Q, Ding W, Wang Y, Zhang Y, Wei W, Lei Y, Li M, Jiao Y, Gu K, Zhang Y
Journal of Inflammation Research 2023, 16:3419-3436
Published Date: 14 August 2023