Back to Journals » Drug Design, Development and Therapy » Volume 19
Rhubarb: Traditional Uses, Phytochemistry, Multiomics-Based Novel Pharmacological and Toxicological Mechanisms
Authors Miao H, Wang KE, Li P, Zhao YY ![]()
Received 4 August 2025
Accepted for publication 11 October 2025
Published 23 October 2025 Volume 2025:19 Pages 9457—9480
DOI https://doi.org/10.2147/DDDT.S557114
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Hua Miao,1 Kai-En Wang,1 Ping Li,2 Ying-Yong Zhao1
1School of Pharmaceutical Sciences, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China; 2Beijing Key Laboratory for Immune-Mediated Inflammatory Diseases, China-Japan Friendship Hospital, Beijing, People’s Republic of China
Correspondence: Ping Li, Beijing Key Laboratory for Immune-Mediated Inflammatory Diseases, China-Japan Friendship Hospital, Beijing, 100029, People’s Republic of China, Email [email protected] Ying-Yong Zhao, School of Pharmaceutical Sciences, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China, Email [email protected]
Abstract: Traditional Chinese medicines (TCMs) are practiced in clinical and healthcare settings and are widely employed as essential therapies for intervening diverse refractory illnesses. Rhubarb, which is prescribed as the root and rhizome of Rheum officinale Baill. Rheum tanguticum Maxim. ex Balf. and Rheum palmatum L. has been widely recognized as an important natural medicine for the management of cardiovascular diseases, cancer, and kidney disorders. The available information on traditional uses, phytochemistry, multiomics-based pharmacological and toxicological studies of rhubarb species and their components including anthraquinones, such as emodin, rhein, chrysophanol, aloe-emodin and physcion as well as their glycoside derivatives published from 2010 to 2025 was searched by several electronic database such as Web of Science, PubMed, Springer, ScienceDirect, Scopus, Google Scholar, CNKI, etc. Increasing evidence has indicated that rhubarb contain various bioactive compounds, such as anthraquinones, anthrones, stilbenes, and saccharides. Numerous studies have demonstrated that rhubarb exerts a broad spectrum of pharmacological effects, including anticardiovascular, anticancer, hepatoprotective, renoprotective, pulmoprotective, antidiabetic, anti-colitis, and antibacterial activities. In this review, traditional applications and major components of rhubarb are presented, and their diverse pharmacological and toxicological effects are discussed. Novel multiomics-based molecular mechanisms of the treatment of various diseases of rhubarb have been highlighted. Key challenges, such as rhubarb effect on cardiorenal syndrome and cardiovascular-kidney-metabolic syndrome, are also addressed. Current evidence suggests that rhubarb is a promising candidate drug for the prevention and treatment of various diseases. Mounting publications show that targeting multiomics-related targets are a promising therapeutic strategy in a myriad of refractory diseases. Rhubarb improves various diseases by reshaping microbial dysbiosis, restoring aberrant expression of ncRNAs and regulating maladaptive metabolite disorder that may provide new therapeutic targets for treatment of various diseases of rhubarb. This review is expected to development of rhubarb-derived anthraquinones as novel therapeutic agents for clinical applications in the future.
Keywords: rhubarb, gut microbiota, long non-coding RNAs, metabolome, anticardiovascular disease, anticancer, renoprotective activity, cardio-renoprotective effect, hepatotoxicity and nephrotoxicity
Introduction
Traditional Chinese medicines (TCMs) have accumulated extensive clinical experience and are recognized as essential therapeutic strategies for the prevention and treatment of various diseases and physiological conditions.1–3 Rhubarb (Rhei Radix et Rhizome) is a TCM primarily used as a strong purgative to remove pathogenic heat or toxins from the body.4,5 In the Chinese Pharmacopoeia, rhubarb is prescribed as the root and rhizome of Rheum officinale Baill. Rheum tanguticum Maxim. ex Balf., and Rheum palmatum L.6 Rhubarb has also been officially listed in the European Pharmacopoeia and Japanese Pharmacopoeia.
Several unofficial rhubarb species have been identified, although they display much weaker purgative effects than the official species, largely due to the absence of sennosides and their relatively low anthraquinone content. According to previous studies, 22 medicinal plants related to rhubarb have been used as folk medicines in Chinese Qinghai and Sichuan provinces as well as in the Xinjiang Uygur and Tibet Autonomous Regions.7 The major components of rhubarb include anthraquinones, anthrones, stilbenes, tannins, butyrophenones, and saccharides8,9 (Figure 1). More than 100 phenolic compounds have been identified, including sennosides, stilbenes, anthraquinones, glucose gallates, naphthalenes, and catechins.10 Rhubarb exhibits diverse biological effects, including anticancer, anti-inflammatory, antifibrotic, and antibacterial activities (Figure 2).
 |
Figure 1 Structure of the main rhubarb compounds. |
 |
Figure 2 Main components and pharmacological effects of rhubarb. Rhubarb contains various active components and has a wide range of pharmacological effects. |
In this review, the traditional uses and major components of rhubarb are presented, and their diverse pharmacological and toxicological effects are discussed. Novel multiomics-based molecular mechanisms underlying the therapeutic effects of rhubarb are also highlighted. Furthermore, key challenges, such as the role of rhubarb in cardiorenal syndrome, are addressed.
Traditional Uses
Rhubarb (Polygonaceae) was first documented in the classic herbal text Sheng Nong’s Herbal Classic.11 Traditionally, rhubarb has been described as effective for “eliminating extensive accumulation with catharsis”, “dampening heat and purging fire”, “cooling blood and detoxifying”, “removing blood stasis and obstructions”, and “disinhibiting dampness to ameliorate jaundice”.12 Rhubarb is considered as the first-line treatment for stagnation and constipation in clinical practice. Its primary cathartic components are sennosides and anthraquinone derivatives.13 Additionally, tannins are believed to cool the blood, flavonoids activate blood circulation, and free anthraquinones clear heat, remove dampness, and reduce jaundice.
Pharmacological differences between raw and steamed rhubarb have been reported. A previous study demonstrated that chrysophanol and emodin exhibited higher bioavailability in steamed rhubarb than in raw rhubarb.14 Traditionally, prepared rhubarb is obtained by steaming raw rhubarb with wine. This preparation process reduces the purgative effect while enhancing its efficacy in alleviating pathogenic heat and toxins, particularly in the treatment of noxious heat–blood stasis syndrome. Rhubarb can also be stir-fried to diminish its purgative effect, when such an action is considered a side effect.
Chemical Components
Anthraquinone
Among anthraquinones, the chemical structure of 9,10-anthraquinone was identified as the most abundant. The anthraquinone content in different rhubarb species ranges from 3% to 5%.15 These compounds can be classified into free and bound types (Figure 1).
Anthranone
Anthranones, including palmatal dianthranones A–C, rhubarb dianthranones A–C, and sennosides A–F, are considered the principal purgative components of rhubarb. As the reduced derivatives of anthraquinones, anthranones are the main active constituents of rhubarb-derived medicinal preparations. To date, 29 anthranones have been identified (Table 1 and Figure 1). These natural compounds are derived primarily from plants and microorganisms. Owing to their diverse and unique structural characteristics, anthranones have been shown to exert various biological activities, including immunoregulatory, antitumor, antibacterial, antioxidant, and anti-inflammatory effects.16 Sennosides have demonstrated therapeutic potential in the prevention and treatment of constipation, diabetes, obesity, fatty liver disease, and cancer.17 After administration, sennosides are metabolized to anthraquinones in vivo and exhibit strong laxative effects. In addition, earlier research identified anthranone esters (revandchinone-1 and revandchinone-2) and anthranone ethers (revandchinone-3 and revandchinone-4), which possess both antibacterial and antifungal activities.18
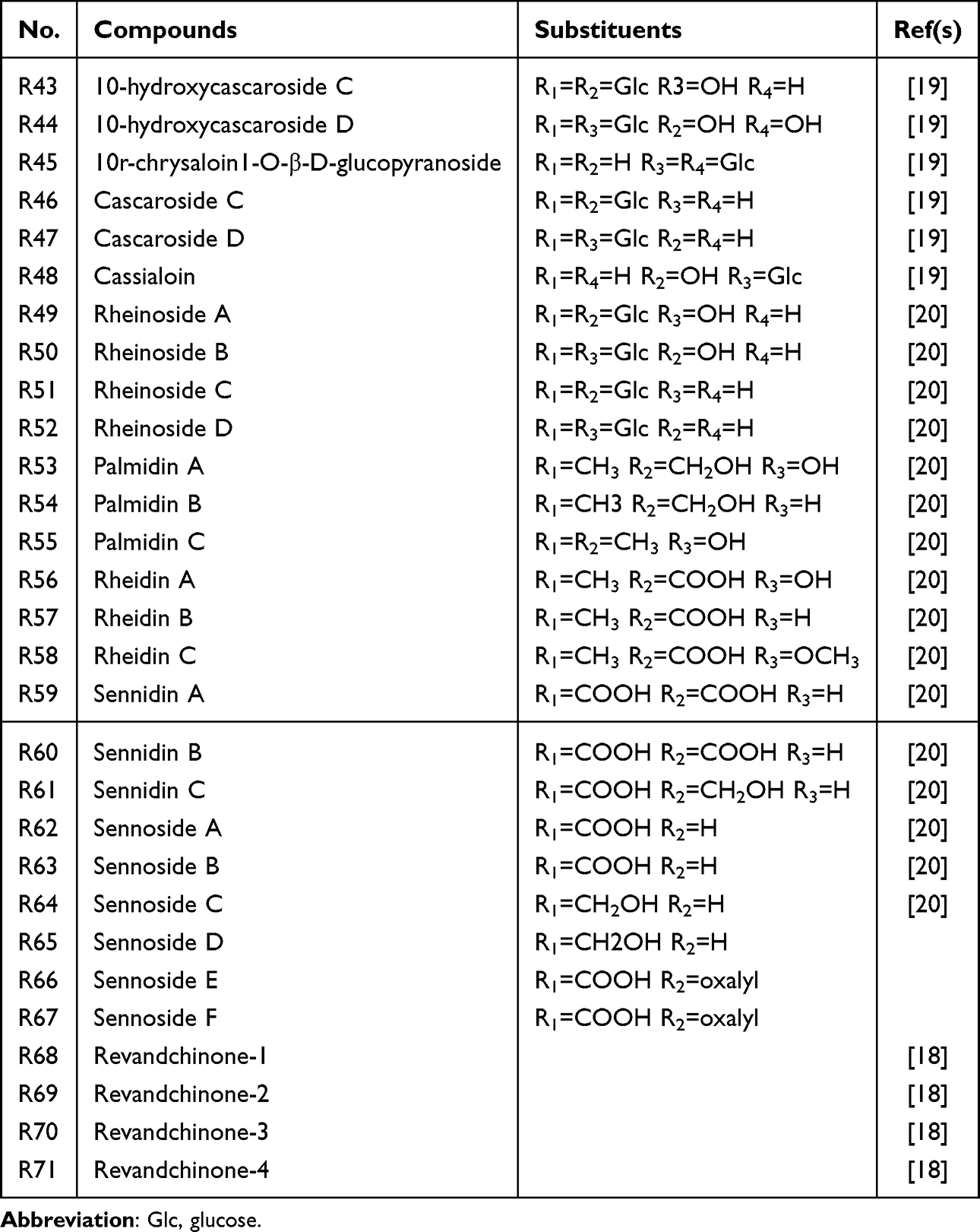 |
Table 1 Main Anthrones of Rhubarb |
Stilbene
Stilbenes, another key component of rhubarb, have been reported to possess strong free radical-scavenging and anti-aging properties.21 The primary active stilbenes were rhapontigenin and piceatannol (Table 2 and Figure 1). Among them, two stilbene glucoside gallates, rhaponticin 2″-O-gallate and rhaponticin 6″-O-gallate, have shown inhibitory effects on nitric oxide production by lipopolysaccharide-activated macrophages.22 Furthermore, rhapontigenin has been shown to scavenge reactive oxygen species (ROS), 1,1-diphenyl-2-picrylhydrazyl radicals, and hydrogen peroxide.23
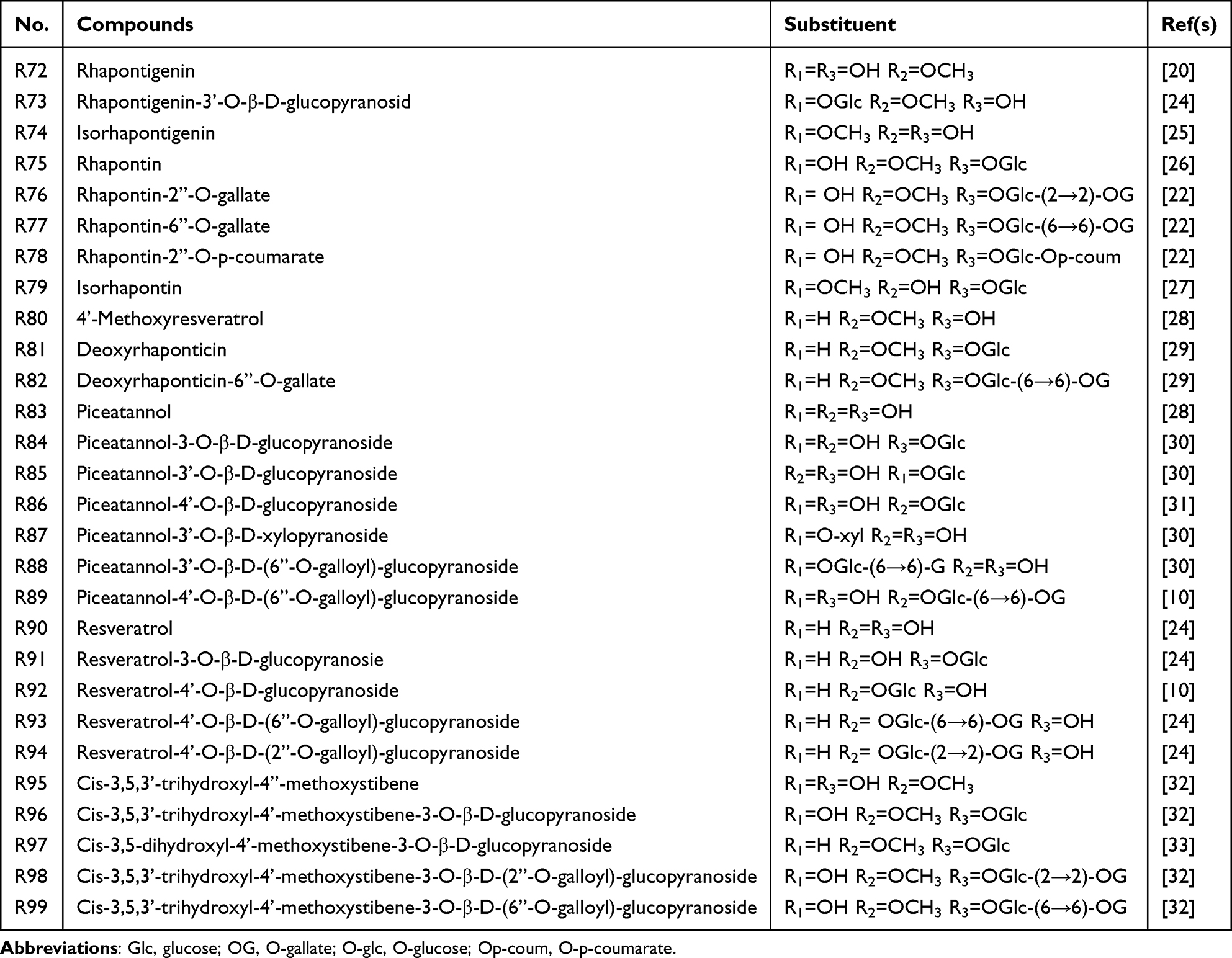 |
Table 2 Main Stilbenes of Rhubarb |
Tannin
Tannins are among the most important constituents of rhubarb. Two major types of tannins, hydrolyzable tannins and condensed tannins, have been identified. The principal monomers included gallic acid and D-catechins. Tannins are considered the bioactive components responsible for the styptic and constipating properties.34 Notably, the purgative activity of rhubarb increased significantly when tannin content was reduced. Forty tannins have been identified in rhubarb (Table 3 and Figure 1).35 Earlier studies identified several tannin compounds in rhubarb, including gallic acid, galloyl glucose, di-O-galloyl-glucose, glucopyranosyl-galloyl-glucose, coumaroyl-O-galloyl-glucose, catechin trimers, catechin-glucopyranoside, and catechin gallate.36
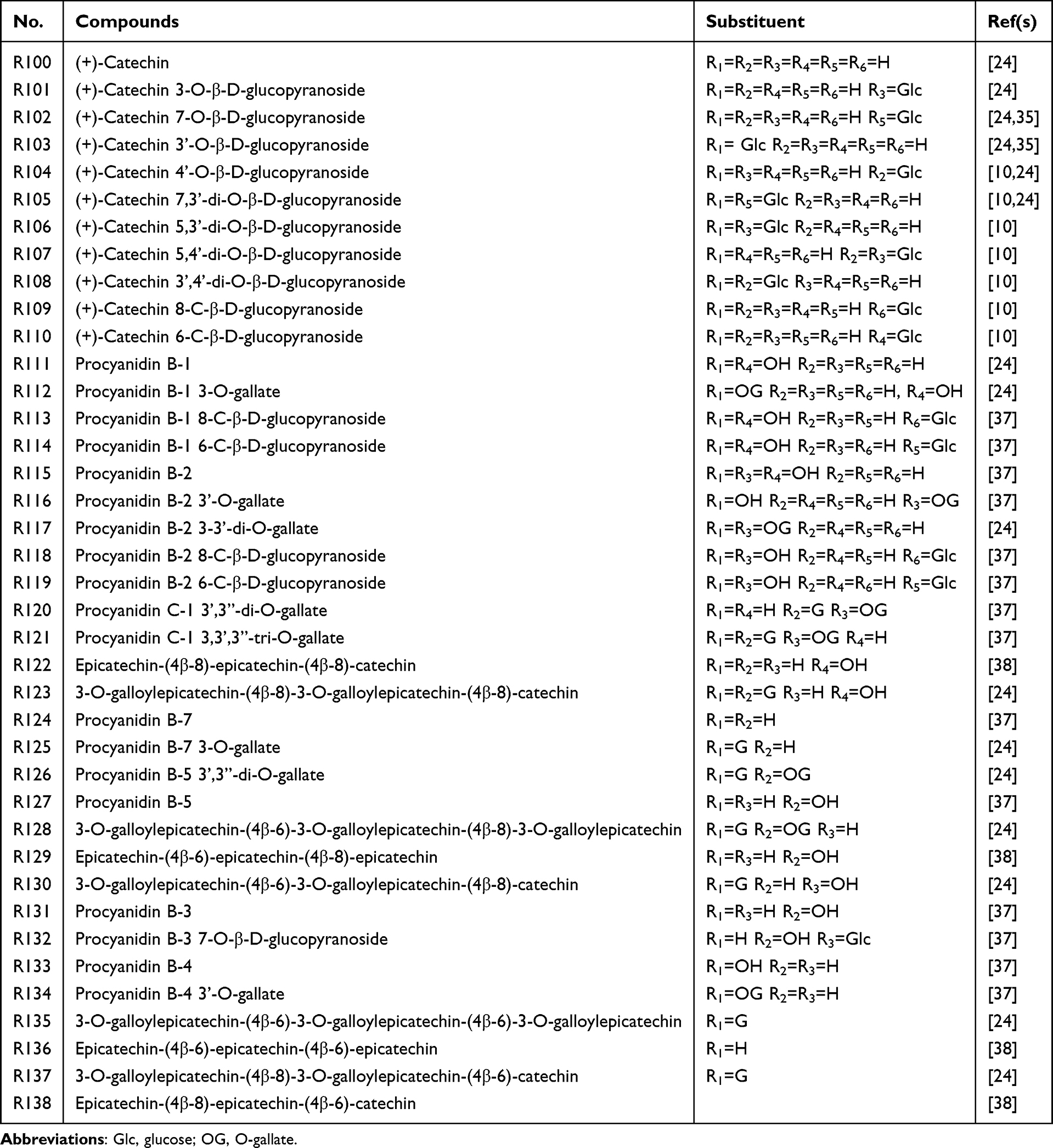 |
Table 3 Main Tannins of Rhubarb |
Butyrophenone and Chromone
Butyrophenones such as lotus palmatin, isobarmatin, and phenylbutadione glucoside have been reported to exhibit analgesic and anti-inflammatory activities. Eight phenylbutanones have been isolated to date (Table 4 and Figure 1).24 Chromones exhibit multiple pharmacological properties, including coronary vasodilation, cholesterol-lowering, antihypertensive, and antibacterial effects. A total of 26 chromone compounds have been identified in rhubarb,39 of which eight representative compounds are listed in this review (Table 4 and Figure 1).
 |
Table 4 Main Butyrophenones and Chromones of Rhubarb |
Other Isolated Components
Other phytochemicals identified in rhubarb include naphthalene glucosides, such as torachrysone 8-O-β-D-glucopyranoside.22 Additionally, flavonoids, such as kaempferol, quercetin, and apigenin-8-O-β-glucopyranoside have been reported. A recent study has identified 31 acyl glucosides, including 3-O-cinnamoyl-1-O-galloyl-β-D-glucopyranoside, 2-O-cinnamoyl-1-O-galloyl-β-D-glucopyranoside, 2-O-cinnamoyl-1,6-di-O-galloyl-β-D-glucopyranoside, and 6-O-cinnamoyl-1-O-galloyl-β-D-glucopyranoside.40 Previous reviews have shown that polysaccharides isolated from rhubarb exert immunomodulatory, lipid-lowering, antioxidant, and anticancer effects.41
Pharmacological Properties
Substantial evidence indicates that rhubarb and its constituents exhibit a wide spectrum of biological activities. These include anticardiovascular, anticancer, hepatoprotective, renoprotective, reproductive protective, pulmoprotective, cerebral protective, antidiabetic, anticolitis, and antibacterial effects. These therapeutic actions have been shown to be mediated through the regulation of various molecular mechanisms.
Several novel mechanisms, revealed through multiomics approaches, have been suggested. These include the modulation of microbial dysbiosis, dysregulation of noncoding RNAs (ncRNAs), such as long noncoding RNAs (lncRNAs) and microRNAs, as well as metabolic disturbances (Figures 3 and 4). Furthermore, rhubarb and its components have been found to activate or inhibit multiple signaling molecules and pathways, including phosphoinositide 3-kinases (PI3K), protein kinase B (Akt), mammalian target of rapamycin (mTOR), phosphatase and tensin homolog (PTEN), peroxisome proliferator-activated receptor γ (PPARγ), Janus kinase 2 (JAK2), signal transducer and activator of transcription 3 (STAT3), mitogen-activated protein kinase (MAPK), myeloid differentiation factor 88 (MyD88), toll-like receptor 4 (TLR4), transforming growth factor-β (TGF-β)/suppressor of Mothers against Decapentaplegic (Smad), inhibitor of kappa B (IκB)/nuclear factor kappa B (NF-κB) and Kelch-like ECH-associated protein 1 (Keap1)/nuclear factor erythroid 2-related factor 2 (Nrf2).
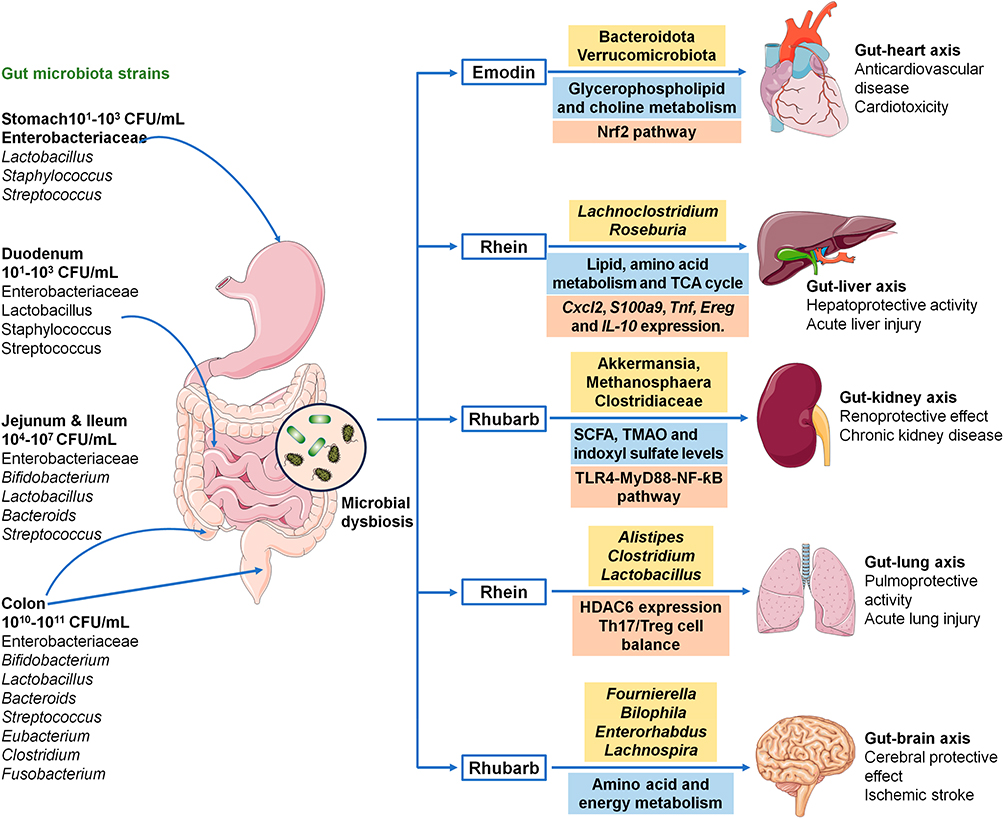 |
Figure 3 Rhubarb exerts its multi-organ protective effect by reshaping microbial dysbiosis and improving microbial-derived metabolite-mediated molecular mechanisms. The gut is the most abundant reservoir for human microbial ecosystems. Spatial composition and distribution of various types of bacteria in the human gastrointestinal tract. The healthy microbial ecosystem contributes to maintaining the normal physiological functions of the host. Dysbiosis of gut microbiota is involved in diverse diseases. Rhubarb exerts its anticardiovascular, hepatoprotective, renoprotective, pulmoprotective, and cerebral protective effects by regulating gut–heart axis, gut–liver axis, gut–kidney axis, gut–lung axis, and gut–brain axis by restoring metabolic disorders and underlying molecular mechanisms. |
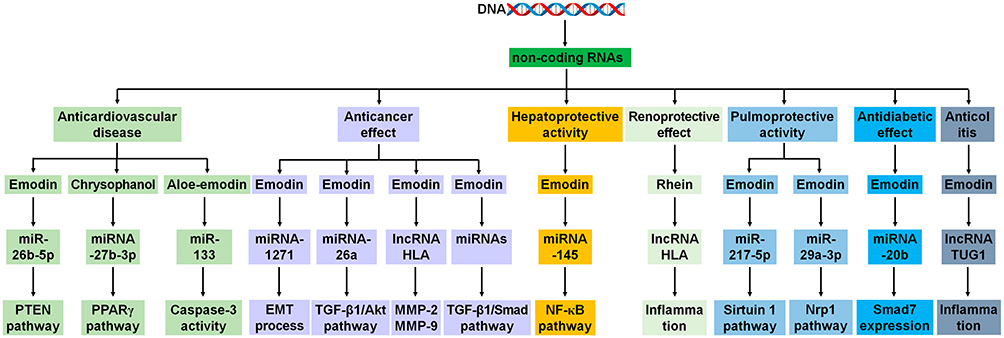 |
Figure 4 Rhubarb exerts multi-organ protective effects by regulating the underlying molecular mechanisms of non-coding RNAs. Aberrant ncRNAs, including miRNAs and lncRNAs, are associated with various diseases. Rhubarb exerts its anticardiovascular disease, anticancer, hepatoprotective, renoprotective, pulmoprotective, antidiabetic, and anti-colitis effects by improving the dysregulation of non-coding RNAs that mediate the underlying molecular mechanisms. |
Anticardiovascular Disease
Cardiovascular disease (CVD) remains a leading cause of morbidity and mortality worldwide, posing a significant burden on public health and healthcare systems. Recent reviews have suggested that rhubarb extracts possess cardioprotective properties and may serve as potential therapeutic options for CVD management.6,42,43 Among the chemical constituents of rhubarb, anthraquinones are considered the primary bioactive components, with emodin being the most extensively studied owing to its anticardiovascular effects. Several reviews have emphasized the beneficial roles of emodin, its underlying mechanisms, and therapeutic targets in the context of CVD.44–46 Recent studies have demonstrated that emodin improves cardiac function and alleviates chronic heart failure and myocardial fibrosis by inhibiting the PI3K/Akt/mTOR signaling axis and modulating the miR-26b-5p/PTEN pathway47,48 (Figure 4). Furthermore, cardiomyocyte pyroptosis induced by doxorubicin has been shown to be suppressed by emodin through direct binding to gasdermin D.49 Emodin has also been reported to inhibit cardiac hypertrophy by attenuating angiotensin-induced hypertension and modulating the gut microbiome.50 One study revealed that emodin restored the gut microbial balance in doxorubicin-induced mice by increasing Bacteroidetes and decreasing Verrucomicrobiota, which contributed to improved cardiac outcomes (Figure 3).51 Additionally, fecal microbiota transplantation (FMT) of emodin-treated mice led to changes in serum metabolite profiles and improved cardiac function in recipient mice, including a reduction in ferroptosis. However, these protective effects were not observed in Nfe2l2–/– mice, suggesting that Keap1/Nrf2 pathway may be essential for the cardioprotective activity51 (Figure 3).
Chrysophanol, the second most widely studied anthraquinone in rhubarb, has also shown significant cardioprotective potential.52 It has been reported to attenuate arrhythmia, cardiac injury, chronic heart failure, and cardiac fibrosis by inhibiting endoplasmic reticulum stress, pyroptosis, and inflammation.53–55 Additionally, chrysophanol has been shown to reduces sepsis-induced acute myocardial injury by modulating the miRNA-27b-3p/PPARγ axis56 (Figure 4), alleviates doxorubicin-induced cardiotoxicity by inhibiting cellular poly(adenosine diphosphate-ribosyl)ation57 and mitigates isoproterenol-induced cardiac hypertrophy by suppressing the JAK2/STAT3 pathway.58
Rhein attenuates myocardial injury by inhibiting mitochondrial fission, activating mitophagy, and suppressing apoptosis via the dynamin-related protein 1/PTEN-induced kinase 1/Parkin and p38 MAPK/heat shock protein 90/c-Jun/c-Fos signaling pathways.59,60 Rhein has also been reported to ameliorate diabetic cardiomyopathy by suppressing inflammatory injury, mitochondrial dynamics dysfunction, apoptosis, and cardiac hypertrophy.61,62 Recent studies have demonstrated that aloe-emodin mitigates radiation-induced heart disease by inhibiting P4HB lactylation and restoring kynurenine metabolism. It has also been shown to improve post-myocardial infarction cardiac remodeling by upregulating Smad7 and thereby blocking TGF-β/Smad signaling.63 In addition, aloe-emodin was found to protect against myocardial infarction and apoptosis by upregulating miR-133 expression64 (Figure 4). These findings support the potential use of rhubarb and its anthraquinone constituents as promising agents for the prevention and treatment of CVD.
Anticancer Effect
Cancer is a long-term disease involving aberrant cellular and immune functions. It is the second leading cause of death after CVD. Several studies have demonstrated that rhubarb extracts protect against lung, hepatocellular, colon, and breast cancers.6 An earlier clinical study showed that rhubarb extract ameliorated radiation-mediated lung toxicity and enhanced pulmonary function, probably by reducing serum levels of TGF-β1 and interleukin (IL)-6 in lung cancer patients.65 A recent analysis showed that rhubarb affects non-small cell lung cancer.66 Previous studies have reported that extracts of Rheum palmatum and rhubarb suppress hepatocellular carcinoma by inhibiting the expression of STAT3 and β-catenin.67–69 In addition, crude extracts of Rheum palmatum were found to inhibit migration and invasion by repressing matrix metalloproteinase-2/matrix metalloproteinase-9 via the MAPK and caspase signaling pathways in human colon cancer cells.70,71 Moreover, Rheum palmatum extracts exhibit antimetastatic effects in human breast cancer cells.72 Several recent reviews have demonstrated that rhubarb components, such as emodin, rhein, physcion, chrysophanol, aloe-emodin, and physcion 8-O-β-D-glucopyranoside, protect against various cancers, including liver, lung, and colon cancers.73–75 The ncRNAs that do not encode proteins have emerged as regulators of various cellular processes. A recent study revealed that emodin suppresses colorectal cancer cells by targeting the lncRNA human leukocyte antigen complex P576 (Figure 4). Moreover, emodin inhibited hepatocellular carcinoma growth by modulating macrophage polarization via the miRNA-26a/TGF-β1/Akt axis77 (Figure 4). Emodin suppressed the epithelial–mesenchymal transition and invasion of pancreatic cancer cells by upregulating miRNA-127178 (Figure 4). Furthermore, emodin blocked angiogenesis in pancreatic cancer by regulating the TGF-β1/Smad pathway and microRNAs79 (Figure 4). Therefore, published data suggests that rhubarb and its components have beneficial effects on cancer suppression.
Hepatoprotective Activity
Liver diseases, including acute liver failure and chronic liver disease, are major global health problems. Liver fibrosis is a major risk factor for the development of liver cancer. An earlier study reported that rhubarb ameliorated TGF-β1-induced expression of α-smooth muscle actin and collagen and attenuated hepatic stellate cell (HSC) activity, which was associated with the downregulation of Smad2/3 and c-Jun N-terminal kinase phosphorylation and a reduction in matrix metalloproteinase-2 activity.80 Metabolic disorders are a major global health concern. A recent study showed that rhubarb affects metabolic pathways such as fatty acid biosynthesis, bile acid biosynthesis, and the pentose phosphate pathway in high-fat diet-induced rats with nonalcoholic fatty liver disease.81 Gong et al demonstrated that rhubarb anthraquinone attenuated acute liver injury and reversed aberrant metabolite levels associated with the biosynthesis and metabolism of tryptophan, tyrosine, phenylalanine, nucleotide sugars, amino sugars, pyrimidines, and steroid hormone biosynthesis in carbon tetrachloride-induced rats.82
Chrysophanol is the most extensively studied component of rhubarb for the treatment of liver disease. Several studies have suggested that chrysophanol ameliorated liver injury by regulating oxidative stress, endoplasmic reticulum stress, ferroptosis, and apoptosis.83,84 In addition, chrysophanol-8-O-glucoside ameliorates acute liver injury and hepatic fibrosis by inhibiting HSC autophagy, liver-resident macrophage-mediated inflammation, and the STAT3 signaling pathway.85,86 Several recent reviews have reported that emodin and aloe-emodin ameliorate liver diseases.87,88 Emodin has been shown to promote HSC senescence, inhibit HSC activation, and attenuate liver fibrosis through nuclear receptor-mediated epigenetic regulation of glutaminase 1, as well as through the p38 MAPK and Smad signaling pathways.89,90
Lipopolysaccharide-induced liver inflammatory injury was mitigated by emodin via the upregulation of miRNA-14591 (Figure 4). Gong et al found that rhein reshaped microbial dysbiosis, reducing the relative abundance of Lachnoclostridium and Roseburia and reversing L-α-amino acids, ofloxacin-N-oxide, 1-hydroxy-1,3-diphenylpropan-2-one, and L-4-hydroxyglutamate semialdehyde to normal levels. Additionally, the expression of C-X-C motif chemokine ligand 2, S100 calcium binding protein A9, tumor necrosis factor, Epiregulin, and IL-10 was downregulated, whereas Mfsd2a and Bhlhe41 was upregulated in mice with acute liver injury92 (Figure 3). These findings demonstrate the therapeutic potential of rhubarb for the treatment of liver injury.
Renoprotective and Reproductive Protective Effects
Chronic kidney disease (CKD) is globally recognized as a public health problem, affecting 10–16% of the adult population.93,94 Renal fibrosis is considered the final manifestation of CKD,95–98 and is associated with activation of the renin-angiotensin system, oxidative stress, inflammation, aryl hydrocarbon receptor and Wnt1/β-catenin pathways, microbial dysbiosis, and metabolic disorders.99–104 Increasing evidence has shown that natural products are widely demonstrated as an important therapy for CKD and renal fibrosis through the regulation of various mechanisms, including the pathways mentioned above.105–111 An earlier study demonstrated that petroleum ether, ethyl acetate, and n-butanol extracts of rhubarb reduced the expression of alpha-smooth muscle actin, fibronectin, collagen I, collagen III, vimentin, and fibroblast-specific protein 1 in kidney tissues of adenine-induced CKD rats.112 This reduction was accompanied by downregulation of the protein expression of TGF-β1, TGF-β receptor I, TGF-β receptor II, Smad2, phosphorylated Smad2, Smad3, phosphorylated Smad3, and Smad4 as well as upregulation of Smad7 protein expression.112 Moreover, rhein has been shown to blunt uric acid nephropathy in rats by suppressing lncRNA antisense non-coding RNA in the INK4 locus-induced inflammation.113
Accumulating evidence has highlighted the roles of gut microbiota dysbiosis and dysregulated microbial-derived metabolites in CKD pathogenesis.114–118 Several studies have demonstrated that natural products and TCMs can improve kidney function and attenuate renal fibrosis by reshaping microbial dysbiosis.108 Ji et al demonstrated that rhubarb enema treatment reduced the relative abundance of Akkermansia, Methanosphaera, and Clostridiaceae in CKD rats, which was associated with inhibition of the TLR4–MyD88–NF-κB signaling pathway and reduction of systemic inflammation119 (Figure 3). The same research group further reported that rhubarb enema improved renal fibrosis by increasing serum levels of short-chain fatty acids (SCFAs), decreasing circulating trimethylamine N-oxide and indoxyl sulfate, and correcting microbial dysbiosis in 5/6 nephrectomized rats120–122 (Figure 3).
As a powerful analytical platform, metabolomics has recently been used extensively for novel biomarker discovery,101,123 disease diagnosis and prognosis,124–127 disease mechanism identification,128–131 pharmaceutical discovery and development,132,133 and drug efficacy and toxicity evaluation.134,135 An increasing number of studies have indicated that TCMs improve CKD and renal fibrosis by restoring metabolic disorders.102 A previous study showed that rhubarb extract restored the levels of 40 metabolites, including indoleacetaldehyde, docosahexaenoic acid, kynurenic acid, hypotaurine, and arachidonic acid, which are associated with tryptophan, taurine, hypotaurine, purine, pyrimidine, fatty acid, glycerophospholipid metabolism, and the tricarboxylic acid (TCA) cycle, in the kidney tissues of adenine-induced CKD rats.112 Another study showed that rhein restored the plasma concentrations of three aberrant biomarkers, arachidonic acid, indoxyl sulfate, and lysoPC(20:5), in rats with unilateral ureteral obstruction.136 A more recent study revealed that rhein ameliorated renal fibrosis by activating the fatty acid oxidation pathway and correcting aberrant lipid metabolism.137 Additionally, rhein attenuates renal fibrosis by enhancing Cpt1a-mediated fatty acid oxidation via the Sirtuin 1/STAT3/Twist1 pathway.138 In chronic glomerulonephritis, glomerular inflammation and injury are caused by immune-mediated mechanisms. Rhein was shown to restore normal levels of 16 metabolites involved in amino acid metabolism, arachidonic acid metabolism, and the TCA cycle, thereby inhibiting oxidative stress, inflammation, and immune dysregulation.139 These findings demonstrate that metabolomics provides profound insights into the therapeutic mechanisms of rhubarb.
Testicular torsion, a urological emergency in adolescents, is characterized by testicular ischemia due to an inadequate local blood supply, resulting in impaired testicular function.140 A previous study showed that pyroptosis and inflammation were activated following testicular ischemia–reperfusion injury, whereas emodin attenuated the injury by inhibiting the NOD-like receptor thermal protein domain-associated protein 3 (NLRP3) signaling pathway and pyroptosis.140 The ameliorative effect of emodin on pyroptosis has been attributed to suppression of NLRP3 inflammasome activation.140 The testes, which are the primary reproductive organs, are responsible for the production of androgens such as spermatozoa and testosterone. Wang et al reported that emodin improved sperm function and increased the levels of testosterone, follicle-stimulating hormone, and luteinizing hormone in cyclophosphamide-induced rats.141 Emodin also inhibits testicular oxidative stress and the expression of pro-inflammatory markers, including malondialdehyde (MDA), interleukin-6, and tumor necrosis factor-α (TNF-α). At the same time, it increased the levels of testicular antioxidant enzymes, such as catalase, glutathione peroxidase, glutathione, and superoxide dismutase (SOD).141 These findings indicate that emodin mitigates chemotherapy-induced testicular toxicity and provides protective effects in cyclophosphamide-induced rats by suppressing oxidative stress and inflammation.141 Moreover, a thermosensitive hydrogel loaded with emodin-loaded triple-targeted nanoparticles was found to improve chronic nonbacterial prostatitis by inhibiting inflammation and fibrosis.142 Therefore, emodin is a promising alternative treatment for reproductive diseases.
Pulmoprotective Activity
Acute lung injury (ALI) is a severe respiratory condition characterized by high morbidity and mortality rates.143 Hu et al reported that rhubarb abrogated hyperoxia-mediated lung lesions by suppressing inflammation in neonatal rats with bronchopulmonary dysplasia.144 Tang et al demonstrated that patients with ALI and lipopolysaccharide-induced ALI mice exhibited gut microbiota dysbiosis, which caused a Th17/Treg cell imbalance. In their study, rhubarb ameliorated ALI by increasing the relative abundance of Alistipes, Clostridium, and Lactobacillus, upregulating histone deacetylase 6 (HDAC6) expression, and restoring Th17/Treg cell balance145 (Figure 3). Rhubarb anthraquinones were found to mitigate lung injury by increasing alveolar epithelial tight junction protein expression via the Ras homolog family member A/Rho-associated coiled coil-containing protein kinase 1 pathway.146 Among the anthraquinone components, emodin has been identified as the most extensively studied compound with pulmoprotective effects.147 Emodin alleviates intestinal ischemia/reperfusion-induced lung injury by upregulating heme oxygenase-1 expression through the PI3K/Akt pathway.148 Additionally, ozone-mediated lung damage is attenuated by emodin via inhibition of the TLR4/MYD88/NF-κB pathway.149 Given the high mortality rate of ALI in severe acute pancreatitis (SAP), a growing body of research has demonstrated that emodin alleviates SAP-associated ALI by activation of the Keap1/Nrf2 pathway, inhibition of ferroptosis, and regulation of the miR-217-5p/Sirtuin 1 axis150–152 (Figure 4). Furthermore, emodin was found to modulate the miRNA expression profiles of exosomes in both the plasma and bronchoalveolar lavage fluid in rats with SAP-associated ALI. Among these, NOVEL-rno-miR-29a-3p in bronchoalveolar lavage fluid was identified to be specific to exosomes and played a key role in the therapeutic effects of emodin (Figure 4). Moreover, emodin was shown to ameliorate SAP-induced ALI by regulating lncRNA–mRNA interaction networks in rats153 (Figure 4). These findings suggest that rhubarb ameliorates acute lung injury by modulating the expression of non-coding RNAs and targeting inflammatory and oxidative stress pathways.
Cerebral Protective Effect
Recent studies have shown that rhubarb extract prevents ischemic stroke by regulating gut microbiota and metabolic pathways through the microbiota–gut–brain axis.154,155 Microbiome and metabolome analyses demonstrated that the aqueous extract of rhubarb reduced the abundance of Fournierella and Bilophila while increasing the abundance of Defluviitaleaceae, Christensenellaceae, Enterorhabdus, and Lachnospira. Altered levels of metabolites, including isoleucine, valine, methionine, 3-aminoisobutyric acid, trimethylamine N-oxide, betaine, choline, glucose, and lactate, were also observed, indicating their regulatory effects on amino acid and energy metabolism154 (Figure 3). Similar findings have indicated that treatment with raw rhubarb confers protection against ischemic stroke via the microbiota–gut–brain axis.155 Mao et al revealed that rhubarb mitigated cerebral ischemia–reperfusion injury by reducing harmful bacteria, correcting metabolic disturbances, restoring intestinal barrier function, attenuating blood–brain barrier dysfunction, and improving neurological outcomes.156
A recent review highlighted the cerebral protective effects of emodin and its molecular mechanisms in Alzheimer’s disease.157,158 Wang et al reported that emodin relieved neuropathic pain by inhibiting inflammation and increasing the abundance of beneficial bacteria and metabolites.159 Additionally, emodin was found to alleviate hypoxia-induced neuronal injury by increasing miRNA-25 expression in PC-12 cells160 (Figure 4). Based on a proteomic analysis identifying 1356 proteins, early studies indicated that rhubarb regulates the expression of neuron projection proteins involved in drug response and nervous system development. These proteins are associated with oxidative stress, calcium-binding protein regulation, vascularization, and energy metabolism.161 These findings suggest that the cerebral protective effects of rhubarb are mediated by the regulation of inflammation, microbial dysbiosis, and metabolic dysfunction.
Antidiabetic Effect
A previous study showed that rhubarb extract inhibits α-glucosidase activity in the small intestine of rats with type 1 diabetes.162 In another study, intestinal α-glucosidase activity was suppressed by rhubarb in rats with type 1 diabetes.163 Several recent reviews have emphasized that rhein and emodin exert antidiabetic effects by improving insulin resistance, oxidative stress, inflammation, microbial dysbiosis, dyslipidemia, mitochondrial dysfunction, and β-cell failure.164,165 An earlier study demonstrated that β-cells were protected by emodin from diabetogenic insults through inhibition of the IκB/NF-κB signaling pathway.166 In addition, multiple studies have shown that anthraquinones, such as emodin and aloe-emodin, mitigate diabetic cardiomyopathy by inhibiting the Akt/glycogen synthase kinase-3β and NLRP3 inflammasome signaling pathways.167,168 Moreover, glucose metabolism was improved by emodin by targeting miRNA-20b in insulin-resistant skeletal muscles169 (Figure 4). These findings demonstrated the antidiabetic properties of rhubarb and its constituents.
Anticolitis
Accumulating evidence has indicated that polysaccharides have multiple pharmacological effects. Rheum tanguticum polysaccharide (RTP) ameliorated colitis injury, reduced the levels of IL-1β, IL-6, and TNF-α, and downregulated MDA and myeloperoxidase levels in dextran sulfate sodium salt-induced colitis mice.170 These effects were accompanied by the regulation of Notch and NF-κB p65 mRNA expression as well as the restoration of gut microbiota dysregulation in colitis mice.170 Several earlier studies have demonstrated that RTP attenuates colitis in 2,4,6-trinitrobenzene sulfonic acid-induced rats by inhibiting the NF-κB signaling pathway.171 In addition, RTP was shown to improve intestinal bacterial balance, suppress TLR4 and NF-κB activation, and modulate Th1/Th2 cytokine production balance.172 Furthermore, targeting the mannose receptor by RTP and the subsequent reduction in the Th1-polarized immune response have been suggested as potential mechanisms underlying its effect in the treatment of colitis.173 Moreover, emodin suppressed acute pancreatitis by regulating lncRNA taurine upregulated 1 (TUG1) and exosomal lncRNA TUG1174 (Figure 4). These findings suggest that rhubarb components, particularly RTP, have promising therapeutic potential for the treatment of colitis.
Antibacterial Effect
Rhubarb has been shown to exhibit inhibitory effects against a variety of bacteria, including Escherichia coli, Staphylococcus aureus, Typhoid bacillus, Pseudomonas aeruginosa, and Dysentery bacillus.175 Mechanistically, the growth of S. aureus is inhibited by rhubarb through disruption of the bacterial cell wall structure and alteration of cell membrane permeability.4 Additionally, higher antibacterial activity against E. coli, S. aureus, and Klebsiella pneumoniae, as well as against fungi such as Candida albicans MTCC 277 and ATCC 90028, has been demonstrated by the chloroform subcomponents of rhubarb.176 Recent studies have highlighted the antibacterial properties of anthraquinones, particularly against Aeromonas hydrophila, suggesting potential applications of anthraquinones in the development of antibacterial agents. Previous findings have indicated that different substituents at the C3 and C6 positions of the anthraquinone benzene ring influence antibacterial activity. Specifically, the hydroxyl, hydroxymethyl, and carboxyl groups of hydroxyanthraquinones have been shown to enhance the antibacterial potency. Inhibitory effects against S. aureus have been reported for five types of free anthraquinones found in rhubarb, with rhein and emodin exhibiting stronger antibacterial activity than the other three hydroxyanthraquinones.177 Moreover, rhubarb extract has been shown to inhibit the growth of Helicobacter pylori. An in vivo study further confirmed its high efficiency in terms of dosage, tolerability, and eradication of active H. pylori infection.178 These findings indicated that rhubarb possesses broad-spectrum antibacterial activity.
Taken together, these findings suggest that Rheum and its components of anthraquinones such as emodin and chrysophanol display diverse pharmacological effects through anti-inflammatory and antifibrotic mechanisms, thereby suggesting their potential as multi-target therapeutics.
Rhubarb Toxicology
Although a wide range of pharmacological effects has been attributed to rhubarb, increasing evidence has demonstrated its potential toxicity, including hepatotoxicity, nephrotoxicity, cardiotoxicity, and neurotoxicity. A recent study reported that accumulated rhein is associated with the induction of apoptosis and autophagy.13 In earlier studies, emodin was shown to induce toxicity in mouse blastocysts and zebrafish embryos via apoptosis.179,180 Therefore, apoptosis has been implicated in rhubarb-induced toxicity.
Hepatotoxicity
Increasing studies have suggested that rhubarb hepatotoxicity was mediated by a variety of underlying molecular mechanisms, such as the aberrant expression of uridine 5′-diphospho-glucuronosyltransferase 2B7 (UGT2B7), multidrug resistance-associated protein 2 (MRP2) and cytochrome P450 family 2 subfamily C member 19 (CYP2C19) as well as oxidative stress, mitochondrial injury and metabolic disorders.181–185
Hepatotoxicity of Rhubarb Extract
In an earlier study, liver damage induced by rhubarb extract was primarily caused by increased liver fibrosis in normal rats, suggesting that the hepatotoxicity of rhubarb could be attributed to liver cell fibrosis.186 Metabolomic analyses showed that rhubarb-induced hepatotoxicity was associated with increased levels of lysoPE (18:2), cervonoyl ethanolamide, dynorphin B (10–13), and 3-hydroxyphenyl 2-hydroxybenzoate as well as decreased levels of dopamine, biopterin, choline, coenzyme Q9, and P1, P4-bis (5′-uridyl) tetraphosphate.187
Hepatotoxicity of Emodin Mediated by UGT2B7 and MRP2 Expression
Increasing publications have suggested that multiomics has been used to reveal emodin-mediated hepatotoxicity. In a recent genome-wide association study, four nucleotide polymorphisms (rs6093966, rs2868094, rs2071197, and rs6073433) located in the hepatocyte nuclear factor 4α gene were identified and found to be associated with emodin glucuronidation. Emodin has been emodin inhibited hepatocyte nuclear factor 4α expression, leading to downregulation of uridine UGT2B7 expression and subsequent hepatotoxicity.188 In an earlier study, higher hepatotoxicity was observed in female rats than in male rats.181 Metabolic and transcriptomic data from human liver and kidney tissues showed that UGT2B7 serves as the primary enzyme responsible for emodin glucuronidation.181 A genome-wide association study further identified rs11726899, located in UGT2B, as a variant that affects emodin metabolism. In HepG2 cells, the knockdown of UGT2B7 results in decreased emodin glucuronidation and increased cytotoxicity.181 In HepG2 cells treated with emodin, gene expression and protein levels of UGT2B7 were decreased, whereas those of MRP2 were increased.181 Long-term administration of emodin was shown to reduce intrinsic clearance in liver microsomes from both male and female rats, thereby contributing to emodin accumulation.181 However, higher self-induced MRP2 expression and lower hepatotoxicity have been observed in male rats than in females following emodin treatment.181 Moreover, emodin-induced hepatotoxicity was alleviated by probenecid, which suppressed the expression of uridine 5′-diphospho-glucuronosyltransferase and MRP2.182 Therefore, emodin hepatotoxicity has been shown to be mediated by the expression of UGT2B7 and MRP2.
Hepatotoxicity of Emodin Mediated by Mitochondrial Injury and Metabolic Disorders
Proteomic analyses have revealed that emodin-induced hepatotoxicity is associated with increased ROS and caspase-3 levels, decreased mitochondrial membrane potential, and impaired ATP synthesis, leading to mitochondrial damage and hepatocyte apoptosis.183 Metabolomic studies have further demonstrated that emodin-induced hepatotoxicity is related to disrupted hepatic antioxidant homeostasis, particularly involving glutathione and xanthine metabolism.184 Similarly, in human liver L-02 cells, emodin has been shown to exerts toxic effects by disturbing fatty acid and glutathione metabolism.189 Moreover, both in vitro and in vivo studies have shown that 3-methylcholanthrene enhances emodin-induced hepatotoxicity by activating the aryl hydrocarbon receptor and inducing cytochrome P450 family 1 subfamily A member 1 expression.185 These findings indicate that emodin hepatotoxicity is mediated by mitochondrial injury and metabolic disturbances.
Hepatotoxicity of Rhein Mediated by Oxidative Stress and CYP2C19 Expression
In a recent acute and subchronic toxicity study, no mortality was observed in immature mice, whereas mortality occurred in aged mice administered rhein.190 Biochemical parameters, including serum aspartate transaminase, alanine aminotransferase, IL-6, and TNF-α, along with pathological liver alterations, demonstrated rhein-induced liver injury. This hepatotoxicity is associated with increased ROS production, Nrf2, and MDA levels, along with decreased SOD levels, indicating that rhein-induced oxidative stress leads to mitochondrial dysfunction and apoptosis.190 However, MRP expression in the liver was not significantly altered.190 Rhein-induced hepatotoxicity involves altered levels of metabolites through the CYP2C19 expression.191 These findings suggest that rhein hepatotoxicity is mediated by oxidative stress and CYP2C19-related metabolic activation.
Hepatotoxicity of Aloe-Emodin Mediated by Oxidative Stress and MRP2 Expression
MRP2 is a key efflux transporter for substances involved in oxidative stress. In a recent study, aloe-emodin was shown to inhibit the transport activity of ATP-binding cassette subfamily C member 2 (ABCC2), downregulate ABCC2 expression, and disrupt intracellular redox balance, resulting in reduced intracellular glutathione levels, mitochondrial dysfunction, and apoptosis. Conversely, overexpression of ABCC2 attenuated aloe-emodin-induced oxidative stress and cell death, whereas ABCC2 knockdown enhanced these effects.192 Additionally, aloe-emodin was found to induce autophagy-mediated degradation of ABCC2, thereby exacerbating hepatotoxicity in mice.192 Another study demonstrated that aloe-emodin induced hepatic lesions in zebrafish, which was accompanied by upregulated expression of NF-κB and p53 at both the mRNA and protein levels, indicating that NF-κB and p53 signaling pathways contribute to aloe-emodin-induced hepatotoxicity.193
Nephrotoxicity of Rhubarb Mediated by Oxidative Stress
Chronic toxicity of rhubarb extract has been shown to primarily target the kidneys, with renal tubular epithelial cells identified as the most affected cell type in rats.194 In a subchronic toxicity study, increases in both spleen and kidney weights were observed along with renal swelling, dark discoloration, and hydronephrosis. Extensive pigment deposition has been detected in the renal tubular epithelial cells.195 Furthermore, total anthraquinones and emodin have both been shown to induce injury in HK-2 cells, with total anthraquinones causing more pronounced damage than emodin.196 The impact of emodin on glomerular endothelial cells was also assessed, which revealed that emodin compromised glomerular filtration barrier integrity and increased barrier permeability. These effects were associated with elevated levels of TNF-α, TGF-β1, IL-6, and monocyte chemoattractant protein-1 (MCP-1), suggesting that oxidative stress–mediated inflammation may underlie emodin-induced nephrotoxicity.197
Reproductive Toxicity of Emodin
Emodin impairs the function of human sperm. A study by Luo et al demonstrated that total motility, linear velocity, and progressive motility of human sperm were significantly decreased following emodin exposure. Additionally, the ability of sperm to penetrate viscous medium and undergo progesterone-induced capacitation and acrosome reaction was inhibited. These effects were associated with reduced intracellular Ca²⁺ levels and suppressed tyrosine phosphorylation, indicating that emodin impairs sperm function by inhibiting intracellular calcium signaling and phosphorylation processes.198 Furthermore, testicular toxicity was found to be mediated through insulin-like growth factor 1 receptor-dependent apoptosis. Additional effects involve alterations in casein kinase II, spermatogenesis, and sperm motility, which are modulated via four signaling pathways, including the tumor necrosis factor receptor 1 signaling pathway.199
Cardiotoxicity of Rhein Mediated by Fas Pathway
The cardiotoxic effects of rhein have been demonstrated both in vitro and in vivo. In H9c2 cells, reduced cell counts, cell atrophy, rounding, and detachment from the culture surface were observed following rhein treatment.200 Proliferation of H9c2 cells is inhibited through S phase arrest, accompanied by increased levels of ROS and lactate dehydrogenase (LDH), decreased mitochondrial membrane potential, and downregulated SOD expression.200 Rhein impaired cardiac function in vivo, as evidenced by reductions in the left ventricular ejection fraction and fractional shortening. Concurrently, elevated serum Ca²⁺, cardiac troponin T, creatine kinase, and LDH levels were observed. These changes were associated with the activation of the Fas-induced apoptotic pathway.200 These findings indicate that rhein-induced cardiotoxicity is mediated by Fas pathway–dependent apoptosis.
Neurotoxicity of Emodin Mediated by Autophagy
The role of autophagy in Alzheimer’s disease (AD) pathogenesis is being increasingly recognized. Emodin has been shown to reduce cell viability and LC3-I/LC3-II conversion ratio while decreasing LDH levels in amyloid-β protein precursor/presenilin 1 mice and PC12 cells.201 Both emodin and 3-methyladenine decreased the number of LC3-II–positive cells in the cortex, increased B-cell lymphoma 2 (Bcl-2) expression, and reduced the expression of Beclin-1 and human vacuolar protein sorting 34 (hVps34), particularly under amyloid-β 25–35 exposure, as confirmed by small interfering RNA silencing of Beclin-1 and Bcl-2.201 Additionally, it was found that the PI3K inhibitor LY294002 blocked LC3-I/LC3-II conversion and increased Bcl-2 expression while reducing hVps34 and Beclin-1 expression.201 These findings suggest that emodin-induced neurotoxicity is associated with the modulation of the PI3K/Beclin-1/Bcl-2 signaling pathway.
Pharmacokinetic Studies
Anthraquinones are the primary active components of rhubarb. These compounds are predominantly absorbed in the intestine and are largely distributed in tissues and organs with high blood flow.202 Among the free forms of anthraquinones, including rhein, aloe-emodin, and chrysophanol, the highest distribution was observed in the kidney, followed by the liver, with the lowest levels detected in the lungs.88 The metabolism of aloe-emodin, emodin, chrysophanol, and physcion has been reported to occur rapidly, with rhein identified as the final metabolic product. Glucuronidation has been shown to be the most efficient metabolic pathway for anthraquinones.203 However, most previous studies have concentrated on aglycones and their glycosides as naturally occurring in herbs, revealing that the oral bioavailability of anthraquinones is low due to the extensive conjugation of aglycones.
Emodin has been found to possess several limitations as a therapeutic agent, including poor water solubility, low oral bioavailability, hepatotoxicity, and nephrotoxicity. Rhein has been recognized as the major absorbable anthraquinone present in systemic circulation following oral administration. A previous study demonstrated a correlation between plasma rhein concentrations and three biomarkers directly associated with renal fibrosis.136 Furthermore, the pharmacokinetics of aloe-emodin, emodin, rhein, and chrysophanol were investigated in rats with CKD induced through 5/6 nephrectomy, adenine, and advanced oxidation protein products (AOPP).204 In these models, increased pharmacokinetic parameters for the four anthraquinones were observed, and these changes were positively correlated with AOPP levels in both 5/6 nephrectomized and adenine-treated rats.204 Elevated AOPP levels and the corresponding increases in pharmacokinetic parameters have also been reported in AOPP-induced rats.204
Collectively, the above-mentioned findings show that rhubarb has dual roles of multiple biological activities and toxic effects and further anthraquinones are not only the bioactive components of rhubarb, but also the toxicity components of rhubarb. These structural features underlie their role as the anti-inflammatory and antifibrotic effects of rhubarb.
Conclusion, Limitations and Future Directions
The unique multi-target efficacy of natural medicines in treating various diseases has been increasingly recognized worldwide over the past several decades. This summarizes traditional use, chemical diversity, broad pharmacological effects, mechanistic insights, toxicity and pharmacokinetics. Rhubarb is widely distributed across East Asia, Europe, and North America. However, its species have varied significantly owing to differences in origin, as revealed by chemical fingerprinting and metabolomic analyses. Traditionally, rhubarb is bitter and cold in nature, with the effects of removing stagnation, detoxifying, purging fire, and promoting blood circulation and menstruation, reducing swelling and relieving pain. Anthraquinones have been identified as the principal components of rhubarb. This review suggests that anthraquinones are not only the bioactive components that exert a broad spectrum of pharmacological effects but also the primary contributor to rhubarb-induced multi-organ toxicity. The anticardiovascular, anticancer, hepatoprotective and renoprotective effects are pharmacological activities of rhubarb and its anthraquinone components. The antiinflammatory and antifibrotic mechanisms are common molecular mechanisms by which rhubarb and its anthraquinone components exert their various pharmacological effects. Intriguingly, based on the studies of genomics, transcriptomics and metabolomics, emerging multiomics studies reveal that pharmacological effects including antiinflammatory and antifibrotic mechanisms of rhubarb and its anthraquinone components are closely associated with reshaping microbial dysbiosis, restoring aberrant expression of ncRNAs and regulating maladaptive metabolite disorder. The latest mounting publications show that targeting gut microbiota, ncRNAs and endogenous metabolites are a promising therapeutic strategy in a myriad of diseases. Therefore, the underlying molecular mechanism by which rhubarb and its anthraquinone components improve various diseases by reshaping microbial dysbiosis, restoring aberrant expression of ncRNAs and regulating maladaptive metabolite disorder may provide new therapeutic targets for treatment of various diseases of rhubarb. Similarly, the antiinflammatory mechanism is common toxicological mechanisms of rhubarb-induced multi-organ toxicity that are also demonstrated to be associated with the dysregulation of genomics, proteomics, and metabolomics. Thus, multiomics is also a powerful tool for uncovering rhubarb’s toxicological mechanisms. Although considerable progress has been made regarding the therapeutic use of rhubarb in various diseases and in elucidating novel multilayered mechanisms, several key issues regarding its application remain unresolved. Integrated with multiomics strategies will offer a new perspective for dissecting and improving the therapy of rhubarb in various diseases.205
According to previous reports, the clinical use of rhubarb has primarily relied on the experience of TCM practitioners and lacks standardized application guidelines. Furthermore, the species and origins of rhubarb have not been clearly specified in many studies. Essential patient characteristics, such as age, body weight, and race, have not been adequately addressed in rhubarb treatment protocols. Therefore, large-scale and scientifically rigorous clinical trials should be conducted to re-evaluate the clinical efficacy of rhubarb using advanced technologies, including metagenomics, proteomics, metabolomics, and phenomics.
Cardiorenal syndrome, a complex clinical condition characterized by the interplay between cardiac and renal dysfunction, is associated with high global morbidity and mortality.206 To date, only one study has reported that emodin attenuates mitochondrial injury in cardiorenal syndrome type 4 by activating PPARγ coactivator 1-alpha signaling.207 Additional experimental and clinical investigations are warranted to assess the pharmacological effects of rhubarb on cardiorenal syndrome. With the growing appreciation and understanding of pathophysiological interrelatedness of metabolic risk factors such as diabetes, obesity, CKD, and CVD, cardiovascular-kidney-metabolic (CKM) syndrome is a new construct recently defined by the American Heart Association.208 CKM syndrome reflects the interplay among metabolic risk factors, CKD, and CVD and has high morbidity and mortality. No report demonstrates that rhubarb is applied to treat CKM syndrome via multiomics-associated molecular mechanism. Therefore, the intervention study of rhubarb on CKM syndrome will provide a new avenue for the discovery of agents targeting CKM syndrome in the future.
Several critical concerns must be addressed based on prior studies. First, whether long-term rhubarb use contributes to carcinogenesis remains controversial. Second, sex-based differences may influence hepatotoxicity, nephrotoxicity, and reproductive toxicity of rhubarb. Therefore, the molecular mechanisms underlying rhubarb-induced toxicity remain unclear and require further investigation. Previous studies have suggested that rhubarb hepatotoxicity may be linked to emodin accumulation in the liver or interference with the oxidative phosphorylation pathway. A deeper understanding of rhubarb-related toxicity is essential for safe clinical application. The multiorgan toxicity of rhubarb must be comprehensively evaluated to ensure its safety and effectiveness in clinical practice.
Abbreviations
ABCC2, ATP-binding cassette subfamily C member 2; AD, Alzheimer’s disease; ALI, Acute lung injury; AOPP, advanced oxidation protein products; Bcl-2, B-cell lymphoma 2; CKD, chronic kidney disease; CKM, cardiovascular-kidney-metabolic; CVD, cardiovascular disease; CYP2C19, cytochrome P450 family 2 subfamily C member 19; FMT, fecal microbiota transplantation; HDAC6, histone deacetylase 6; HSC, hepatic stellate cell; hVps34, human vacuolar protein sorting 34; IL, interleukin; IκB, inhibitor of kappa B; JAK2, Janus kinase 2; Keap1, Kelch-like ECH-associated protein 1; MCP-1, monocyte chemoattractant protein-1; MRP2, multidrug resistance-associated protein 2; MyD88, myeloid differentiation factor 88; NF-κB, nuclear factor kappa B; NLRP3, NOD-like receptor thermal protein domain-associated protein 3; Nrf2, nuclear factor erythroid 2-related factor 2; PI3K, phosphoinositide 3-kinases; PPARγ, peroxisome proliferator-activated receptor γ; RTP, Rheum tanguticum polysaccharide; SAP, severe acute pancreatitis; SCFAs, short-chain fatty acids; Smad, suppressor of Mothers against Decapentaplegic; SOD, superoxide dismutase; TCA, tricarboxylic acid; TGF-β, transforming growth factor-β; TLR4, toll-like receptor 4; TNF-α, tumor necrosis factor-α; TUG1, taurine upregulated 1; UGT2B7, uridine 5′-diphospho-glucuronosyltransferase 2B7.
Funding
This study was supported by the Shaanxi Key Science and Technology Plan Project (No. 2023-ZDLSF-26) and National Natural Science Foundation of China (Nos. 82474062, 82274079 and 82274192).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Mullowney MW, Duncan KR, Elsayed SS, et al. Artificial intelligence for natural product drug discovery. Nat Rev Drug Discov. 2023;22:895–916. doi:10.1038/s41573-023-00774-7
2. Atanasov AG, Zotchev SB, Dirsch VM, Supuran CT. Natural products in drug discovery: advances and opportunities. Nat Rev Drug Discov. 2021;20:200–216. doi:10.1038/s41573-020-00114-z
3. Guo ZY, Wu X, Zhang SJ, et al. Poria cocos: traditional uses, triterpenoid components and their renoprotective pharmacology. Acta Pharmacol Sin. 2024;46:836–851. doi:10.1038/s41401-024-01404-7
4. Xiang H, Zuo J, Guo F, Dong D. What we already know about rhubarb: a comprehensive review. Chin Med. 2020;15:88. doi:10.1186/s13020-020-00370-6
5. Feng HY, Wang YQ, Yang J, Miao H, Zhao YY, Li X. Anthraquinones from Rheum officinale ameliorate renal fibrosis in acute kidney injury and chronic kidney disease. Drug Des Devel Ther. 2025;19:5739–5760. doi:10.2147/DDDT.S521265
6. Xu H, Wang W, Li X, et al. Botany, traditional use, phytochemistry, pharmacology and clinical applications of rhubarb (Rhei Radix et Rhizome): a systematic review. Am J Chin Med. 2024;52:1925–1967. doi:10.1142/S0192415X24500757
7. Kumar A, Jugran AK, Bargali SS, Bhatt ID. Ethno-medicinal uses, ecology, phytochemistry, biological activities, and conservation approaches for himalayan rhubarb species. Nat Prod Res. 2025;2025:1–14.
8. Wang Y, Yu F, Li A, et al. The progress and prospect of natural components in rhubarb (Rheum ribes L.) in the treatment of renal fibrosis. Front Pharmacol. 2022;13:919967. doi:10.3389/fphar.2022.919967
9. Gao HW, Liu J, Zhang M, et al. Anthraquinones from rhubarb as potential inhibitors of trimethylamine (TMA)-lyase for Alzheimer’s disease. Neurochem Res. 2022;16:295–301. doi:10.1134/S1819712422030035
10. Ye M, Han J, Chen H, Zheng J, Guo D. Analysis of phenolic compounds in rhubarbs using liquid chromatography coupled with electrospray ionization mass spectrometry. J Am Soc Mass Spectrom. 2007;18(1):82–91. doi:10.1016/j.jasms.2006.08.009
11. Liu Y, Li L, Xiao YQ, et al. Global metabolite profiling and diagnostic ion filtering strategy by LC-QTOF MS for rapid identification of raw and processed pieces of Rheum palmatum L. Food Chem. 2016;192:531–540. doi:10.1016/j.foodchem.2015.07.013
12. Chen JQ, Chen YY, Du X, et al. Fuzzy identification of bioactive components for different efficacies of rhubarb by the back propagation neural network association analysis of UPLC-Q-TOF/MSE and integrated effects. Chin Med. 2022;17:50. doi:10.1186/s13020-022-00612-9
13. Cheng Y, Zhang H, Qu L, et al. Identification of rhein as the metabolite responsible for toxicity of rhubarb anthraquinones. Food Chem. 2020;331:127363. doi:10.1016/j.foodchem.2020.127363
14. Zhang J, Fu Y, Li L, et al. Pharmacokinetic comparisons of major bioactive components after oral administration of raw and steamed rhubarb by UPLC-MS/MS. J Pharm Biomed Anal. 2019;171:43–51. doi:10.1016/j.jpba.2019.04.002
15. Wei SY, Yao WX, Ji WY, Wei JQ, Peng SQ. Qualitative and quantitative analysis of anthraquinones in rhubarbs by high performance liquid chromatography with diode array detector and mass spectrometry. Food Chem. 2013;141:1710–1715. doi:10.1016/j.foodchem.2013.04.074
16. Zofou D, Ntie-Kang F, Sippl W, Efange SM. Bioactive natural products derived from the Central African flora against neglected tropical diseases and HIV. Nat Prod Rep. 2013;30:1098–1120. doi:10.1039/c3np70030e
17. Le J, Ji H, Zhou X, et al. Pharmacology, toxicology, and metabolism of sennoside A, a medicinal plant-derived natural compound. Front Pharmacol. 2021;12:714586. doi:10.3389/fphar.2021.714586
18. Babu KS, Srinivas PV, Praveen B, Kishore KS, Murty US, Rao JM. Antimicrobial constituents from the rhizomes of Rheum emodi. Phytochemistry. 2003;62:203–207. doi:10.1016/S0031-9422(02)00571-X
19. Min BS, Lee JP, Na MK, et al. A new naphthopyrone from the root of Pleuropterus ciliinervis. Chem Pharm Bull. 2003;51:1322–1324. doi:10.1248/cpb.51.1322
20. Cao YJ, Pu ZJ, Tang YP, et al. Advances in bio-active constituents, pharmacology and clinical applications of rhubarb. Chin Med. 2017;12:36. doi:10.1186/s13020-017-0158-5
21. Matsuda H, Morikawa T, Toguchida I, Park JY, Harima S, Yoshikawa M. Antioxidant constituents from rhubarb: structural requirements of stilbenes for the activity and structures of two new anthraquinone glucosides. Bioorg Med Chem. 2001;9:41–50. doi:10.1016/S0968-0896(00)00215-7
22. Kageura T, Matsuda H, Morikawa T, et al. Inhibitors from rhubarb on lipopolysaccharide-induced nitric oxide production in macrophages: structural requirements of stilbenes for the activity. Bioorg Med Chem. 2001;9:1887–1893. doi:10.1016/S0968-0896(01)00093-1
23. Chen D, Liu JR, Cheng Y, et al. Metabolism of rhaponticin and activities of its metabolite, rhapontigenin: a review. Curr Med Chem. 2020;27:3168–3186. doi:10.2174/0929867326666190121143252
24. Zhang L, Liu H, Qin L, et al. Global chemical profiling based quality evaluation approach of rhubarb using ultra performance liquid chromatography with tandem quadrupole time-of-flight mass spectrometry. J Sep Sci. 2015;38:511–522. doi:10.1002/jssc.201400971
25. Chu XY, Yang SZ, Zhu MQ, et al. Isorhapontigenin improves diabetes in mice via regulating the activity and stability of PPARγ in adipocytes. J Agric Food Chem. 2020;68:3976–3985. doi:10.1021/acs.jafc.0c00515
26. Tao L, Cao J, Wei W, et al. Protective role of rhapontin in experimental pulmonary fibrosis in vitro and in vivo. Int Immunopharmacol. 2017;47:38–46. doi:10.1016/j.intimp.2017.03.020
27. Lee SW, Hwang BS, Kim MH, et al. Inhibition of LFA-1/ICAM-1-mediated cell adhesion by stilbene derivatives from Rheum undulatum. Arch Pharm Res. 2012;35:1763–1770. doi:10.1007/s12272-012-1008-8
28. Ha MT, Park DH, Shrestha S, et al. PTP1B inhibitory activity and molecular docking analysis of stilbene derivatives from the rhizomes of Rheum undulatum L. Fitoterapia. 2018;131:119–126. doi:10.1016/j.fitote.2018.10.020
29. Czop M, Bogucka-Kocka A, Kubrak T, et al. Imaging flow cytometric analysis of stilbene-dependent apoptosis in drug resistant human leukemic cell lines. Molecules. 2019;24:1896. doi:10.3390/molecules24101896
30. Setoguchi Y, Oritani Y, Ito R, et al. Absorption and metabolism of piceatannol in rats. J Agric Food Chem. 2014;62:2541–2548. doi:10.1021/jf404694y
31. Fei Y, Wang J, Peng B, et al. Phenolic constituents from Rheum nobile and their antioxidant activity. Nat Prod Res. 2017;31:2842–2849. doi:10.1080/14786419.2017.1303691
32. Vastano BC, Chen Y, Zhu N, Ho CT, Zhou Z, Rosen RT. Isolation and identification of stilbenes in two varieties of Polygonum cuspidatum. J Agric Food Chem. 2000;48:253–256. doi:10.1021/jf9909196
33. Zhao XH, Han F, Li YL, Yue HL. Preparative isolation and purification of three stilbene glycosides from the tibetan medicinal plant Rheum tanguticum maxim. Ex Balf. by high-speed counter-current chromatography. Phytochem Anal. 2013;24:171–175. doi:10.1002/pca.2397
34. Liu C, Zheng Y, Xu W, Wang H, Lin N. Rhubarb tannins extract inhibits the expression of aquaporins 2 and 3 in magnesium sulphate-induced diarrhoea model. Biomed Res Int. 2014;2014:619465. doi:10.1155/2014/619465
35. Lin CC, Wu CI, Lin TC, Sheu SJ. Determination of 19 rhubarb constituents by high-performance liquid chromatography-ultraviolet-mass spectrometry. J Sep Sci. 2006;29:2584–2593. doi:10.1002/jssc.200500307
36. Wang JB, Qin Y, Kong WJ. Identification of the antidiarrhoeal components in official rhubarb using liquid chromatography-tandem mass spectrometry. Food Chem. 2011;129:1737–1743. doi:10.1016/j.foodchem.2011.06.041
37. Ge YW, Zhu S, Kazuma K, Wei SL, Yoshimatsu K, Komatsu K. Molecular ion index assisted comprehensive profiling of B-type oligomeric proanthocyanidins in rhubarb by high performance liquid chromatography-tandem mass spectrometry. Anal Bioanal Chem. 2016;408:3555–3570. doi:10.1007/s00216-016-9433-z
38. Shoji T, Mutsuga M, Nakamura T, et al. Isolation and structural elucidation of some procyanidins from apple by low-temperature nuclear magnetic resonance. J Agric Food Chem. 2003;51(13):3806–3813. doi:10.1021/jf0300184
39. Wang Q, Lu Z, Zhang L, et al. Applying characteristic fragment filtering for rapid detection and identification of ingredients in rhubarb by HPLC coupled with linear ion trap-orbitrap mass spectrometry. J Sep Sci. 2017;40:2854–2862. doi:10.1002/jssc.201700203
40. Yao M, Li J, He M, et al. Investigation and identification of the multiple components of Rheum officinale Baill. using ultra-high-performance liquid chromatography coupled with quadrupole-time-of-flight tandem mass spectrometry and data mining strategy. J Sep Sci. 2021;44:681–690. doi:10.1002/jssc.202000735
41. Zong A, Cao H, Wang F. Anticancer polysaccharides from natural resources: a review of recent research. Carbohydr Polym. 2012;90:1395–1410. doi:10.1016/j.carbpol.2012.07.026
42. Wu L, Wang X, Jiang J, Chen Y, Peng B, Jin W. Mechanism of rhubarb in the treatment of hyperlipidemia: a recent review. Open Med (Wars). 2023;18:20230812. doi:10.1515/med-2023-0812
43. Gholami Z, Bahrami M, Kamalinejad M, Ahangar H, Reshadmanesh T, Gohari S. Efficacy of Rhubarb root extract on quality of life in patients with systolic heart failure: a randomized placebo controlled study. Med Sci. 2021;25:2096–2104.
44. Wang X, Yang S, Li Y, Jin X, Lu J, Wu M. Role of emodin in atherosclerosis and other cardiovascular diseases: pharmacological effects, mechanisms, and potential therapeutic target as a phytochemical. Biomed Pharmacother. 2023;161:114539. doi:10.1016/j.biopha.2023.114539
45. Guo Y, Zhang R, Li W. Emodin in cardiovascular disease: the role and therapeutic potential. Front Pharmacol. 2022;13:1070567. doi:10.3389/fphar.2022.1070567
46. Li Q, Gao J, Pang X, Chen A, Wang Y. Molecular mechanisms of action of emodin: as an anti-cardiovascular disease drug. Front Pharmacol. 2020;11:559607. doi:10.3389/fphar.2020.559607
47. Huang W, Zhou P, Zou X, Liu Y, Zhou L, Zhang Y. Emodin ameliorates myocardial fibrosis in mice by inactivating the ROS/PI3K/Akt/mTOR axis. Clin Exp Hypertens. 2024;46:2326022. doi:10.1080/10641963.2024.2326022
48. Gu Z, Zhang S, Zhao S, Cui Y, Sun L. Emodin improves the cardiac function in the rats with chronic heart failure through regulation of the miR-26b-5p/PTEN pathway. Arch Med Sci. 2024;20:655–663. doi:10.5114/aoms.2020.96345
49. Dai S, Chen Y, Fan X, et al. Emodin attenuates cardiomyocyte pyroptosis in doxorubicin-induced cardiotoxicity by directly binding to GSDMD. Phytomedicine. 2023;121:155105. doi:10.1016/j.phymed.2023.155105
50. Evans L, Price T, Hubert N, et al. Emodin inhibited pathological cardiac hypertrophy in response to angiotensin-induced hypertension and altered the gut microbiome. Biomolecules. 2023;13:1274. doi:10.3390/biom13091274
51. Hu S, Zhou J, Hao J, et al. Emodin ameliorates doxorubicin-induced cardiotoxicity by inhibiting ferroptosis through the remodeling of gut microbiota composition. Am J Physiol Cell Physiol. 2024;326:C161–c76. doi:10.1152/ajpcell.00477.2023
52. Su S, Wu J, Gao Y, et al. The pharmacological properties of chrysophanol, the recent advances. Biomed Pharmacother. 2020;125:110002. doi:10.1016/j.biopha.2020.110002
53. Liu C, Qiu S, Liu X, Huang R, Fang Z. Chrysophanol attenuates cardiac fibrosis and arrhythmia by suppressing the endoplasmic reticulum stress/pyroptosis axis and inflammation. Phytother Res. 2025;2025:1.
54. Zhu M, Tai S. Chrysophanol mitigates chronic heart failure in rats by modulating ROS-mediated parthanatos and pyroptosis. Int Heart J. 2025;66(1):126–136. doi:10.1536/ihj.24-387
55. Lian Y, Xia X, Zhao H, Zhu Y. The potential of chrysophanol in protecting against high fat-induced cardiac injury through Nrf2-regulated anti-inflammation, anti-oxidant and anti-fibrosis in Nrf2 knockout mice. Biomed Pharmacother. 2017;93:1175–1189. doi:10.1016/j.biopha.2017.05.148
56. Zhao H, Wang Y, Zhu X. Chrysophanol exerts a protective effect against sepsis-induced acute myocardial injury through modulating the microRNA-27b-3p/Peroxisomal proliferating-activated receptor gamma axis. Bioengineered. 2022;13:12673–12690. doi:10.1080/21655979.2022.2063560
57. Lu J, Li J, Hu Y, et al. Chrysophanol protects against doxorubicin-induced cardiotoxicity by suppressing cellular PARylation. Acta Pharm Sin B. 2019;9:782–793. doi:10.1016/j.apsb.2018.10.008
58. Yuan J, Hong H, Zhang Y, et al. Chrysophanol attenuated isoproterenol-induced cardiac hypertrophy by inhibiting Janus kinase 2/signal transducer and activator of transcription 3 signaling pathway. Cell Biol Int. 2019;43:695–705. doi:10.1002/cbin.11146
59. Chen Y, Tu Y, Cao J, Wang Y, Ren Y. Rhein alleviates doxorubicin-induced myocardial injury by inhibiting the p38 MAPK/HSP90/c-Jun/c-Fos pathway-mediated apoptosis. Cardiovasc Toxicol. 2024;24:1139–1150. doi:10.1007/s12012-024-09917-7
60. Li H, Jia Y, Yao D, Gao M, Wang L, Liu J. Rhein alleviates myocardial ischemic injury by inhibiting mitochondrial division, activating mitochondrial autophagy and suppressing myocardial cell apoptosis through the Drp1/Pink1/Parkin pathway. Mol Biol Rep. 2024;51:266. doi:10.1007/s11033-023-09154-1
61. Li H, Wang G, Tang Y, Wang L, Jiang Z, Liu J. Rhein alleviates diabetic cardiomyopathy by inhibiting mitochondrial dynamics disorder, apoptosis and hypertrophy in cardiomyocytes. Cell Signal. 2025;131:111734. doi:10.1016/j.cellsig.2025.111734
62. Zhao SY, Zhao HH, Wang BH, Shao C, Pan WJ, Li SM. Rhein alleviates advanced glycation end products (AGEs)-induced inflammatory injury of diabetic cardiomyopathy in vitro and in vivo models. J Nat Med. 2023;77:898–915. doi:10.1007/s11418-023-01741-7
63. Yu J, Zhao X, Yan X, et al. Aloe-emodin ameliorated MI-induced cardiac remodeling in mice via inhibiting TGF-β/SMAD signaling via up-regulating SMAD7. Phytomedicine. 2023;114:154793. doi:10.1016/j.phymed.2023.154793
64. Yu Y, Liu H, Yang D, et al. Aloe-emodin attenuates myocardial infarction and apoptosis via up-regulating miR-133 expression. Pharmacol Res. 2019;146:104315. doi:10.1016/j.phrs.2019.104315
65. Yu HM, Liu YF, Cheng YF, Hu LK, Hou M. Effects of rhubarb extract on radiation induced lung toxicity via decreasing transforming growth factor-β-1 and interleukin-6 in lung cancer patients treated with radiotherapy. Lung Cancer. 2008;59:219–226. doi:10.1016/j.lungcan.2007.08.007
66. Tan YR, Lu Y. Molecular mechanism of Rhubarb in the treatment of non-small cell lung cancer based on network pharmacology and molecular docking technology. Mol Divers. 2023;27:1437–1457. doi:10.1007/s11030-022-10501-w
67. Tan ZB, Fan HJ, Wu YT, et al. Rheum palmatum extract exerts anti-hepatocellular carcinoma effects by inhibiting signal transducer and activator of transcription 3 signaling. J Ethnopharmacol. 2019;232:62–72. doi:10.1016/j.jep.2018.12.019
68. El-Saied MA, Sobeh M, Abdo W, et al. Rheum palmatum root extract inhibits hepatocellular carcinoma in rats treated with diethylnitrosamine. J Pharm Pharmacol. 2018;70:821–829. doi:10.1111/jphp.12899
69. Tsai KH, Hsien HH, Chen LM, et al. Rhubarb inhibits hepatocellular carcinoma cell metastasis via GSK-3-β activation to enhance protein degradation and attenuate nuclear translocation of β-catenin. Food Chem. 2013;138:278–285. doi:10.1016/j.foodchem.2012.10.038
70. Ma YS, Hsiao YP, Lin JH, et al. Crude extract of Rheum palmatum L inhibits migration and invasion of LS1034 human colon cancer cells acts through the inhibition of matrix metalloproteinase-2/-9 by MAPK signaling. Environ Toxicol: Int J. 2015;30:852–863. doi:10.1002/tox.21962
71. Ma YS, Hsu SC, Weng SW, et al. Crude extract of Rheum palmatum L induced cell death in LS1034 human colon cancer cells acts through the caspase-dependent and -independent pathways. Environ Toxicol: Int J. 2014;29:969–980. doi:10.1002/tox.21827
72. Nho KJ, Chun JM, Lee AY, Kim HK. Anti-metastatic effects of Rheum Palmatum L. extract in human MDA-MB-231 breast cancer cells. Environ Toxicol Pharmacol. 2015;40:30–38. doi:10.1016/j.etap.2015.05.006
73. Yu L, Qin J, Zhang M, Gao Y, Zhao Y. Research progress on the anti-liver cancer mechanism and toxicity of rhubarb anthraquinone. Drug Des Devel Ther. 2024;18:6089–6113. doi:10.2147/DDDT.S489377
74. Sharifi-Rad J, Herrera-Bravo J, Kamiloglu S, et al. Recent advances in the therapeutic potential of emodin for human health. Biomed Pharmacother. 2022;154:113555. doi:10.1016/j.biopha.2022.113555
75. Adnan M, Rasul A, Hussain G, et al. Physcion and physcion 8-O-β-D-glucopyranoside: natural anthraquinones with potential anticancer activities. Curr Drug Targets. 2021;22:488–504. doi:10.2174/18735592MTEwDNjQiz
76. Yan CX, Sun L, Wang GB, et al. Anti-tumor efficacy of emodin on colorectal cancer cells via targeting lncRNA HLA complex P5. Chem Biol Drug Des. 2024;103:e14444.
77. Yin J, Zhao X, Chen X, Shen G. Emodin suppresses hepatocellular carcinoma growth by regulating macrophage polarization via microRNA-26a/transforming growth factor β1/protein kinase B. Bioengineered. 2022;13:9548–9563. doi:10.1080/21655979.2022.2061295
78. Li N, Wang C, Zhang P, You S. Emodin inhibits pancreatic cancer EMT and invasion by up‑regulating microRNA‑1271. Mol Med Rep. 2018;18:3366–3374. doi:10.3892/mmr.2018.9304
79. Lin SZ, Xu JB, Ji X, et al. Emodin inhibits angiogenesis in pancreatic cancer by regulating the transforming growth factor-β/drosophila mothers against decapentaplegic pathway and angiogenesis-associated microRNAs. Mol Med Rep. 2015;12:5865–5871. doi:10.3892/mmr.2015.4158
80. Lin YL, Wu CF, Huang YT. Effects of rhubarb on migration of rat hepatic stellate cells. J Gastroenterol Hepatol. 2009;24:453–461. doi:10.1111/j.1440-1746.2008.05573.x
81. Zhang F, Wu R, Liu Y, Dai S, Gong X, Li Y. Integration of pharmacodynamics and metabolomics to reveal rhubarb anthraquinone protection against nonalcoholic fatty liver disease rat model. J Pharm Pharmacol. 2024;76:381–390. doi:10.1093/jpp/rgae014
82. Gong X, Zhang F, Li Y, Peng C. Study on the mechanism of acute liver injury protection in Rhubarb anthraquinone by metabolomics based on UPLC-Q-TOF-MS. Front Pharmacol. 2023;14:1141147. doi:10.3389/fphar.2023.1141147
83. Kuo CY, Chiu V, Hsieh PC, et al. Chrysophanol attenuates hepatitis B virus X protein-induced hepatic stellate cell fibrosis by regulating endoplasmic reticulum stress and ferroptosis. J Pharmacol Sci. 2020;144:172–182. doi:10.1016/j.jphs.2020.07.014
84. Jiang WJ, Zhou R, Li PJ. Protective effect of chrysophanol on LPS/d-GalN-induced hepatic injury through the RIP140/NF-κB pathway. RSC Adv. 2016;6:38192–38200.
85. Park YJ, Lee KH, Jeon MS, et al. Hepatoprotective potency of chrysophanol 8-O-glucoside from Rheum palmatum L. against hepatic fibrosis via regulation of the STAT3 signaling pathway. Int J Mol Sci. 2020;21:9044. doi:10.3390/ijms21239044
86. Wang T, Lu Z, Qu XH, et al. Chrysophanol-8-O-glucoside protects mice against acute liver injury by inhibiting autophagy in hepatic stellate cells and inflammatory response in liver-resident macrophages. Front Pharmacol. 2022;13:951521. doi:10.3389/fphar.2022.951521
87. Hu N, Liu J, Xue X, Li Y. The effect of emodin on liver disease -- comprehensive advances in molecular mechanisms. Eur J Pharmacol. 2020;882:173269. doi:10.1016/j.ejphar.2020.173269
88. Dong X, Zeng Y, Liu Y, et al. Aloe-emodin: a review of its pharmacology, toxicity, and pharmacokinetics. Phytother Res. 2020;34:270–281. doi:10.1002/ptr.6532
89. Chen L, Liang B, Xia S, et al. Emodin promotes hepatic stellate cell senescence and alleviates liver fibrosis via a nuclear receptor (Nur77)-mediated epigenetic regulation of glutaminase 1. Br J Pharmacol. 2023;180:2577–2598. doi:10.1111/bph.16156
90. Wang X, Niu C, Zhang X, Dong M. Emodin suppresses activation of hepatic stellate cells through p38 mitogen-activated protein kinase and Smad signaling pathways in vitro. Phytother Res. 2018;32:2436–2446. doi:10.1002/ptr.6182
91. Xie R, Liu M, Li S. Emodin weakens liver inflammatory injury triggered by lipopolysaccharide through elevating microRNA-145 in vitro and in vivo. Artif Cells Nanomed Biotechnol. 2019;47:1877–1887. doi:10.1080/21691401.2019.1614015
92. Liu S, Yin R, Yang Z, Wei F, Hu J. The effects of rhein on D-GalN/LPS-induced acute liver injury in mice: results from gut microbiome-metabolomics and host transcriptome analysis. Front Immunol. 2022;13:971409. doi:10.3389/fimmu.2022.971409
93. Francis A, Harhay MN, Ong ACM, et al. Chronic kidney disease and the global public health agenda: an international consensus. Nat Rev Nephrol. 2024;20:473–485. doi:10.1038/s41581-024-00820-6
94. Khandpur S, Mishra P, Mishra S, Tiwari S. Challenges in predictive modelling of chronic kidney disease: a narrative review. World J Nephrol. 2024;13:97214. doi:10.5527/wjn.v13.i3.97214
95. Youhua L. Kidney fibrosis: fundamental questions, challenges, and perspectives. Integr Med Nephrol Androl. 2024;11:e24–00027.
96. Balakumar P. Unleashing the pathological role of epithelial-to-mesenchymal transition in diabetic nephropathy: the intricate connection with multifaceted mechanism. World J Nephrol. 2024;13:95410. doi:10.5527/wjn.v13.i2.95410
97. Jafry NH, Manan S, Rashid R, Mubarak M. Clinicopathological features and medium-term outcomes of histologic variants of primary focal segmental glomerulosclerosis in adults: a retrospective study. World J Nephrol. 2024;13:88028. doi:10.5527/wjn.v13.i1.88028
98. Ren LL, Miao H, Wang YN, Liu F, Li P, Zhao YY. TGF-β as a master regulator of aging-associated tissue fibrosis. Aging and Disease. 2023;14:1633–1650. doi:10.14336/AD.2023.0222
99. Miao H, Wang YN, Su W, et al. Sirtuin 6 protects against podocyte injury by blocking the renin-angiotensin system by inhibiting the Wnt1/β-catenin pathway. Acta Pharmacol Sin. 2023;45:137–149. doi:10.1038/s41401-023-01148-w
100. Wang YN, Miao H, Yu XY, et al. Oxidative stress and inflammation are mediated via aryl hydrocarbon receptor signalling in idiopathic membranous nephropathy. Free Radic Biol Med. 2023;207:89–106. doi:10.1016/j.freeradbiomed.2023.07.014
101. Chen D, Guo Y, Li P. New insights into a novel metabolic biomarker and therapeutic target for chronic kidney disease. Integr Med Nephrol Androl. 2024;11:e24–00019. doi:10.1097/IMNA-D-24-00019
102. Wang YN, Zhang ZH, Liu HJ, et al. Integrative phosphatidylcholine metabolism through phospholipase A2 in rats with chronic kidney disease. Acta Pharmacol Sin. 2023;44:393–405. doi:10.1038/s41401-022-00947-x
103. Vagopoulou A, Theofilis P, Karasavvidou D, et al. Pilot study on the effect of flavonoids on arterial stiffness and oxidative stress in chronic kidney disease. World J Nephrol. 2024;13:95262. doi:10.5527/wjn.v13.i3.95262
104. Raikou VD. Renoprotective strategies. World J Nephrol. 2024;13:89637. doi:10.5527/wjn.v13.i1.89637
105. Wang YN, Wu X, Shan QY, et al. Acteoside-containing caffeic acid is bioactive functional group of antifibrotic effect by suppressing inflammation via inhibiting AHR nuclear translocation in chronic kidney disease. Acta Pharmacol Sin. 2025. doi:10.1038/s41401-025-01598-4
106. Li XJ, Wang YN, Wang WF, et al. an aryl hydrocarbon receptor antagonist, ameliorates podocyte injury through inhibiting oxidative stress and inflammation. Front Pharmacol. 2024;15:1386604. doi:10.3389/fphar.2024.1386604
107. Wang YN, Li XJ, Wang WF, Zou L, Miao H, Zhao YY. Geniposidic acid attenuates chronic tubulointerstitial nephropathy through regulation of the NF‐ƙB/Nrf2 pathway via aryl hydrocarbon receptor signaling. Phytother Res. 2024;38:5441–5457. doi:10.1002/ptr.8324
108. Zhao H, Zhao T, Li P. Gut microbiota-derived metabolites: a new perspective of traditional Chinese medicine against diabetic kidney disease. Integr Med Nephrol Androl. 2024;11:e23–00024. doi:10.1097/IMNA-D-23-00024
109. Zhong X, Jia J, Tan R, Wang L. Hederagenin improves adriamycin-induced nephropathy by inhibiting the JAK/STAT signaling pathway. Integr Med Nephrol Androl. 2024;11:e22–00016. doi:10.1097/IMNA-D-22-00016
110. Mei H, Jing T, Liu H, et al. Ursolic acid alleviates mitotic catastrophe in podocyte by inhibiting autophagic P62 accumulation in diabetic nephropathy. Int J Biol Sci. 2024;20(9):3317–3333. doi:10.7150/ijbs.94096
111. Zou TF, Liu ZG, Cao PC, et al. Fisetin treatment alleviates kidney injury in mice with diabetes-exacerbated atherosclerosis through inhibiting CD36/fibrosis pathway. Acta Pharmacol Sin. 2023;44:2065–2074. doi:10.1038/s41401-023-01106-6
112. Zhang ZH, Li MH, Liu D, et al. Rhubarb protect against tubulointerstitial fibrosis by inhibiting TGF-β/Smad pathway and improving abnormal metabolome in chronic kidney disease. Front Pharmacol. 2018;9:1029. doi:10.3389/fphar.2018.01029
113. Hu J, Wang D, Wu H, et al. Long non-coding RNA ANRIL-mediated inflammation response is involved in protective effect of rhein in uric acid nephropathy rats. Cell Biosci. 2019;9(1):11. doi:10.1186/s13578-019-0273-3
114. Tao P, Huo J, Chen L. Bibliometric analysis of the relationship between gut microbiota and chronic kidney disease from 2001–2022. Integr Med Nephrol Androl. 2024;11:e00017.
115. Krukowski H, Valkenburg S, Madella AM, et al. Gut microbiome studies in CKD: opportunities, pitfalls and therapeutic potential. Nat Rev Nephrol. 2023;19:87–101. doi:10.1038/s41581-022-00647-z
116. Li XJ, Shan QY, Wu X, Miao H, Zhao YY. Gut microbiota regulates oxidative stress and inflammation: a double-edged sword in renal fibrosis. Cell Mol Life Sci. 2024;81:480. doi:10.1007/s00018-024-05532-5
117. Stepanova N. Probiotic interventions in peritoneal dialysis: a review of underlying mechanisms and therapeutic potentials. World J Nephrol. 2024;13:98719. doi:10.5527/wjn.v13.i4.98719
118. Miao H, Liu F, Wang YN, et al. Targeting Lactobacillus johnsonii to reverse chronic kidney disease. Signal Transduct Target Ther. 2024;9:195. doi:10.1038/s41392-024-01913-1
119. Ji C, Deng Y, Yang A, et al. Rhubarb enema improved colon mucosal barrier injury in 5/6 nephrectomy rats may associate with gut microbiota modification. Front Pharmacol. 2020;11:1092. doi:10.3389/fphar.2020.01092
120. Ji C, Lu F, Wu Y, et al. Rhubarb enema increasing short-chain fatty acids that improves the intestinal barrier disruption in CKD may be related to the regulation of gut dysbiosis. Biomed Res Int. 2022;2022:1896781. doi:10.1155/2022/1896781
121. Lu Z, Ji C, Luo X, et al. Nanoparticle-mediated delivery of emodin via colonic irrigation attenuates renal injury in 5/6 nephrectomized rats. Front Pharmacol. 2020;11:606227. doi:10.3389/fphar.2020.606227
122. Ji C, Li Y, Mo Y, et al. Rhubarb enema decreases circulating trimethylamine N-oxide level and improves renal fibrosis accompanied with gut microbiota change in chronic kidney disease rats. Front Pharmacol. 2021;12:780924. doi:10.3389/fphar.2021.780924
123. Liu HJ, Miao H, Yang JZ, Liu F, Cao G, Zhao YY. Deciphering the role of lipoproteins and lipid metabolic alterations in ageing and ageing-associated renal fibrosis. Ageing Res Rev. 2023;85:101861. doi:10.1016/j.arr.2023.101861
124. Gupta R, Kumari S, Senapati A, Ambasta RK, Kumar P. New era of artificial intelligence and machine learning-based detection, diagnosis, and therapeutics in Parkinson’s disease. Ageing Res Rev. 2023;90:102013. doi:10.1016/j.arr.2023.102013
125. Miao H, Cao G, Wu XQ, et al. Identification of endogenous 1-aminopyrene as a novel mediator of progressive chronic kidney disease via aryl hydrocarbon receptor activation. Br J Pharmacol. 2020;177:3415–3435. doi:10.1111/bph.15062
126. Pereira PR, Carrageta DF, Oliveira PF, Rodrigues A, Alves MG, Monteiro MP. Metabolomics as a tool for the early diagnosis and prognosis of diabetic kidney disease. Med Res Rev. 2022;42:1518–1544. doi:10.1002/med.21883
127. Corradi V, Caprara C, Barzon E, et al. A possible role of p-cresyl sulfate and indoxyl sulfate as biomarkers in the prediction of renal function according to the GFR (G) categories. Integr Med Nephrol Androl. 2024;11:e24–00002. doi:10.1097/IMNA-D-24-00002
128. Miao H, Wu XQ, Wang YN, et al. 1-Hydroxypyrene mediates renal fibrosis through aryl hydrocarbon receptor signalling pathway. Br J Pharmacol. 2022;179:103–124. doi:10.1111/bph.15705
129. Cao G, Miao H, Wang YN, et al. Intrarenal 1-methoxypyrene, an aryl hydrocarbon receptor agonist, mediates progressive tubulointerstitial fibrosis in mice. Acta Pharmacol Sin. 2022;43:2929–2945. doi:10.1038/s41401-022-00914-6
130. Li XJ, Fang C, Zhao RH, Zou L, Miao H, Zhao YY. Bile acid metabolism in health and ageing-related diseases. Biochem Pharmacol. 2024;225:116313. doi:10.1016/j.bcp.2024.116313
131. Li XJ, Suo P, Wang YN, et al. Arachidonic acid metabolism as a therapeutic target in AKI-to-CKD transition. Front Pharmacol. 2024;15:1365802. doi:10.3389/fphar.2024.1365802
132. Pang H, Hu Z. Metabolomics in drug research and development: the recent advances in technologies and applications. Acta Pharm Sin B. 2023;13:3238–3251. doi:10.1016/j.apsb.2023.05.021
133. Palermo A. Metabolomics- and systems-biology-guided discovery of metabolite lead compounds and druggable targets. Drug Discov Today. 2023;28:103460. doi:10.1016/j.drudis.2022.103460
134. Miao H, Wang YN, Yu XY, et al. Lactobacillus species ameliorate membranous nephropathy through inhibiting aryl hydrocarbon receptor pathway via tryptophan-produced indole metabolites. Br J Pharmacol. 2024;181:162–179. doi:10.1111/bph.16219
135. Miao H, Zhang YM, Yu XY, Zou L, Zhao YY. Membranous nephropathy: systems biology-based novel mechanism and traditional Chinese medicine therapy. Front Pharmacol. 2022;13:969930. doi:10.3389/fphar.2022.969930
136. Sun H, Luo G, Xiang Z, Cai X, Chen D. Pharmacokinetics and pharmacodynamics study of rhein treating renal fibrosis based on metabonomics approach. Phytomedicine. 2016;23:1661–1670. doi:10.1016/j.phymed.2016.10.002
137. Xiao Q, Yu X, Yu X, et al. An integrated network pharmacology and cell metabolomics approach to reveal the role of rhein, a novel PPARα agonist, against renal fibrosis by activating the PPARα-CPT1A axis. Phytomedicine. 2022;102:154147. doi:10.1016/j.phymed.2022.154147
138. Song X, Du Z, Yao Z, Tang X, Zhang M. Rhein improves renal fibrosis by restoring Cpt1a-mediated fatty acid oxidation through SirT1/STAT3/Twist1 pathway. Molecules. 2022;27:2344. doi:10.3390/molecules27072344
139. Yu W, Yang W, Zhao MY, Meng XL. Functional metabolomics analysis elucidating the metabolic biomarker and key pathway change associated with the chronic glomerulonephritis and revealing action mechanism of rhein. Front Pharmacol. 2020;11:554783. doi:10.3389/fphar.2020.554783
140. He KX, Ning JZ, Li W, Cheng F. Emodin alleviates testicular ischemia-reperfusion injury through the inhibition of NLRP3-mediated pyroptosis. Tissue Cell. 2023;82:102069. doi:10.1016/j.tice.2023.102069
141. Wang Y, Zou Z, Jaisi A, Olatunji OJ. Unravelling the protective effects of emodin against cyclophosphamide induced gonadotoxicity in male Wistar rats. Drug Des Devel Ther. 2021;15:4403–4411. doi:10.2147/DDDT.S333383
142. Ye Y, Zhong W, Luo R, et al. Thermosensitive hydrogel with emodin-loaded triple-targeted nanoparticles for a rectal drug delivery system in the treatment of chronic non-bacterial prostatitis. J Nanobiotechnology. 2024;22:33. doi:10.1186/s12951-023-02282-7
143. Lu X, Li G, Liu Y, et al. The role of fatty acid metabolism in acute lung injury: a special focus on immunometabolism. Cell Mol Life Sci. 2024;81:120. doi:10.1007/s00018-024-05131-4
144. Hu XC, Xu J, Liu N. Rhubarb alleviates hyperoxia induced lung injury in neonatal rats with bronchopulmonary dysplasia by inhibiting inflammation. Trop J Pharm Res. 2022;21:2619–2624. doi:10.4314/tjpr.v21i12.16
145. Tang T, Wang F, Liu J, Ye W, Zhao T, Li Z. Rhubarb alleviates acute lung injury by modulating gut microbiota dysbiosis in mice. Curr Microbiol. 2022;79:116. doi:10.1007/s00284-022-02811-x
146. Liu GY, Bai YL, Leng GX, et al. Rhubarb anthraquinones ameliorates inflammatory lung injury by enhancing alveolar epithelium tight junction proteins through RhoA/ROCK1 signalling. Pharmacogn Mag. 2023;19:359–370. doi:10.1177/09731296231158697
147. Liu L, Zhang Y, Tang XR, et al. Effect of emodin on acute lung injury: a meta-analysis of preclinical trials. BMC Pulm Med. 2024;24:596. doi:10.1186/s12890-024-03406-x
148. Chen M, Ji T, Liu YY, et al. Emodin alleviates intestinal ischemia/reperfusion-induced lung injury by upregulating HO-1 expression via PI3K/AkT pathway. Surgery. 2024;176:499–510. doi:10.1016/j.surg.2024.04.006
149. Zeng D, Xie W, Xiang Y, Tan ML, Qin XQ. Emodin attenuates ozone-induced lung injury via TLR4/MYD88/NF-κB signaling pathways. Pharm Chem J. 2023;57:957–964. doi:10.1007/s11094-023-02971-5
150. Chen Z, Dong X, Song Y, et al. Emodin: an alveolar macrophage protector in acute pancreatitis induced lung injury. Int J Med Sci. 2025;22:2075–2087. doi:10.7150/ijms.105965
151. Shen G, Wen H, Li H, et al. Emodin protects against severe acute pancreatitis-associated acute lung injury by activating Nrf2/HO-1/GPX4 signal and inhibiting ferroptosis in vivo and in vitro. BMC Gastroenterol. 2025;25:57. doi:10.1186/s12876-025-03660-1
152. Zhang Z, Luo Y, Zhuang X, Gao H, Yang Q, Chen H. Emodin alleviates lung injury via the miR-217-5p/Sirt1 axis in rats with severe acute pancreatitis. J Pharmacol Sci. 2024;156:188–197. doi:10.1016/j.jphs.2024.08.007
153. Xu C, Luo Y, Ntim M, et al. Effect of emodin on long non-coding RNA-mRNA networks in rats with severe acute pancreatitis-induced acute lung injury. J Cell Mol Med. 2021;25:1851–1866. doi:10.1111/jcmm.15525
154. Liu X, Wang Y, Tian Y, et al. The water extract of rhubarb prevents ischemic stroke by regulating gut bacteria and metabolic pathways. Metabolites. 2024;14:1.
155. Xian M, Ma Z, Zhan S, et al. Network analysis of microbiome and metabolome to explore the mechanism of raw rhubarb in the protection against ischemic stroke via microbiota-gut-brain axis. Fitoterapia. 2024;175:105969. doi:10.1016/j.fitote.2024.105969
156. Mao M, Cao X, Liang Y, et al. Neuroprotection of rhubarb extract against cerebral ischaemia-reperfusion injury via the gut-brain axis pathway. Phytomedicine. 2024;126:155254. doi:10.1016/j.phymed.2023.155254
157. Saha P, Ahmad F. Neuroprotective, anti-inflammatory and antifibrillogenic offerings by emodin against alzheimer’s dementia: a systematic review. ACS Omega. 2024;9:7296–7309. doi:10.1021/acsomega.3c07178
158. Mitra S, Anjum J, Muni M, et al. Exploring the journey of emodin as a potential neuroprotective agent: novel therapeutic insights with molecular mechanism of action. Biomed Pharmacother. 2022;149:112877. doi:10.1016/j.biopha.2022.112877
159. Wang C, Wu L, Zhou R, et al. Integration of microbiota and metabolomics reveals the analgesic mechanisms of emodin against neuropathic pain. Int Immunopharmacol. 2023;125:111170. doi:10.1016/j.intimp.2023.111170
160. Cao M, Fang Y, Jia W, Wang Y, Sun J, Tao D. Emodin relieves hypoxia-triggered injury via elevation of microRNA-25 in PC-12 cells. Artif Cells Nanomed Biotechnol. 2019;47:2678–2687. doi:10.1080/21691401.2019.1633339
161. Liu T, Zhou J, Cui H, et al. iTRAQ-based quantitative proteomics reveals the neuroprotection of rhubarb in experimental intracerebral hemorrhage. J Ethnopharmacol. 2019;232:244–254. doi:10.1016/j.jep.2018.11.032
162. Hou KJ, Chen C, Wang XH, et al. Rhubarb extract’s inhibiting effects on α-glucosidase’s activities in the small intestines of type 1 diabetic rats. West Indian Med J. 2016;65:2016.
163. Hou KJ, Lin CJ, Chen C, Wu BT, Wang XH, Zhu D. Inhibitory effect of rhubarb on intestinal α-glucosidase activity in type 1 diabetic rats. Trop J Pharm Res. 2017;16:1293–1297. doi:10.4314/tjpr.v16i6.12
164. Deng T, Du J, Yin Y, et al. Rhein for treating diabetes mellitus: a pharmacological and mechanistic overview. Front Pharmacol. 2022;13:1106260. doi:10.3389/fphar.2022.1106260
165. Martorell M, Castro N, Victoriano M, et al. An update of anthraquinone derivatives emodin, diacerein, and catenarin in diabetes. Evid Based Complement Alternat Med. 2021;2021:3313419. doi:10.1155/2021/3313419
166. Bae UJ, Song MY, Jang HY, et al. Emodin isolated from Rheum palmatum prevents cytokine-induced β-cell damage and the development of type 1 diabetes. JOURNAL of FUNCTIONAL FOODS. 2015;16:9–19. doi:10.1016/j.jff.2015.04.016
167. Wu Z, Chen Q, Ke D, Li G, Deng W. Emodin protects against diabetic cardiomyopathy by regulating the AKT/GSK-3β signaling pathway in the rat model. Molecules. 2014;19:14782–14793. doi:10.3390/molecules190914782
168. Hu Y, Zhang S, Lou H, et al. Aloe-emodin derivative, an anthraquinone compound, attenuates pyroptosis by targeting NLRP3 inflammasome in diabetic cardiomyopathy. Pharmaceuticals. 2023;16:1275. doi:10.3390/ph16091275
169. Xiao D, Hu Y, Fu Y, et al. Emodin improves glucose metabolism by targeting microRNA-20b in insulin-resistant skeletal muscle. Phytomedicine. 2019;59:152758. doi:10.1016/j.phymed.2018.11.018
170. Zhang Y, Liu Y, Luo J, et al. Rheum tanguticum polysaccharide alleviates DSS-induced ulcerative colitis and regulates intestinal microbiota in mice. Food Bioscience. 2023;53:102788. doi:10.1016/j.fbio.2023.102788
171. Liu L, Liu Z, Zhang T, et al. Combined therapy with Rheum tanguticum polysaccharide and low-dose 5-ASA ameliorates TNBS-induced colitis in rats by suppression of NF-κB. Planta Med. 2015;81:705–712. doi:10.1055/s-0035-1545945
172. Liu L, Yuan S, Long Y, et al. Immunomodulation of Rheum tanguticum polysaccharide (RTP) on the immunosuppressive effects of dexamethasone (DEX) on the treatment of colitis in rats induced by 2,4,6-trinitrobenzene sulfonic acid. Int Immunopharmacol. 2009;9(13–14):1568–1577. doi:10.1016/j.intimp.2009.09.013
173. Liu L, Guo Z, Lv Z, et al. The beneficial effect of Rheum tanguticum polysaccharide on protecting against diarrhea, colonic inflammation and ulceration in rats with TNBS-induced colitis: the role of macrophage mannose receptor in inflammation and immune response. Int Immunopharmacol. 2008;8:1481–1492. doi:10.1016/j.intimp.2008.04.013
174. Wen X, He B, Tang X, Wang B, Chen Z. Emodin inhibits the progression of acute pancreatitis via regulation of lncRNA TUG1 and exosomal lncRNA TUG1. Mol Med Rep. 2021;24:785. doi:10.3892/mmr.2021.12425
175. Wang J, Zhao H, Kong W, et al. Microcalorimetric assay on the antimicrobial property of five hydroxyanthraquinone derivatives in rhubarb (Rheum palmatum L.) to Bifidobacterium adolescentis. Phytomedicine. 2010;17(8–9):684–689. doi:10.1016/j.phymed.2009.10.009
176. Rolta R, Kumar V, Sourirajan A, Upadhyay NK, Dev K. Bioassay guided fractionation of rhizome extract of Rheum emodi wall as bio-availability enhancer of antibiotics against bacterial and fungal pathogens. J Ethnopharmacol. 2020;257:112867. doi:10.1016/j.jep.2020.112867
177. Wang YW, Gao WY, Xiao XH, Liu Y. Calorimetric investigation of the effect of hydroxyanthraquinones in Rheum officinale Baill on Staphylococcus aureus growth. Thermochim acta. 2005;429:167–170. doi:10.1016/j.tca.2005.03.008
178. Ibrahim M, Khan AA, Tiwari SK, Habeeb MA, Khaja MN, Habibullah CM. Antimicrobial activity of Sapindus mukorossi and Rheum emodi extracts against H pylori: in vitro and in vivo studies. World J Gastroenterol. 2006;12:7136–7142. doi:10.3748/wjg.v12.i44.7136
179. Chang MH, Huang FJ, Chan WH. Emodin induces embryonic toxicity in mouse blastocysts through apoptosis. Toxicology. 2012;299:25–32. doi:10.1016/j.tox.2012.05.006
180. He Q, Liu K, Wang S, Hou H, Yuan Y, Wang X. Toxicity induced by emodin on zebrafish embryos. Drug Chem Toxicol. 2012;35:149–154. doi:10.3109/01480545.2011.589447
181. Wu L, Han W, Chen Y, et al. Gender differences in the hepatotoxicity and toxicokinetics of emodin: the potential mechanisms mediated by UGT2B7 and MRP2. Mol Pharm. 2018;15:3931–3945. doi:10.1021/acs.molpharmaceut.8b00387
182. Wu L, Chen Y, Liu H, et al. Emodin-induced hepatotoxicity was exacerbated by probenecid through inhibiting UGTs and MRP2. Toxicol Appl Pharmacol. 2018;359:91–101. doi:10.1016/j.taap.2018.09.029
183. Tabassum R, Ramo JT, Ripatti P, et al. Genetic architecture of human plasma lipidome and its link to cardiovascular disease. Nat Commun. 2019;10:4329. doi:10.1038/s41467-019-11954-8
184. Ruan L, Jiang L, Zhao W, Meng H, Zheng Q, Wang J. Hepatotoxicity or hepatoprotection of emodin? Two sides of the same coin by 1H-NMR metabolomics profiling. Toxicol Appl Pharmacol. 2021;431:115734. doi:10.1016/j.taap.2021.115734
185. Wang M, Zhang Z, Ruan P, et al. Emodin-induced hepatotoxicity is enhanced by 3-methylcholanthrene through activating aryl hydrocarbon receptor and inducing CYP1A1 in vitro and in vivo. Chem Biol Interact. 2022;365:110089. doi:10.1016/j.cbi.2022.110089
186. Wang JB, Zhao HP, Zhao YL, et al. Hepatotoxicity or hepatoprotection? Pattern recognition for the paradoxical effect of the Chinese herb Rheum palmatum L. in treating rat liver injury. PLoS One. 2011;6:e24498.
187. Li S, Wang Y, Li C, et al. Study on hepatotoxicity of rhubarb based on metabolomics and network pharmacology. Drug Des Devel Ther. 2021;15:1883–1902. doi:10.2147/DDDT.S301417
188. Chen Y, Zhang T, Wu L, et al. Metabolism and toxicity of emodin: genome-wide association studies reveal hepatocyte nuclear factor 4α regulates UGT2B7 and emodin glucuronidation. Chem Res Toxicol. 2020;33:1798–1808. doi:10.1021/acs.chemrestox.0c00047
189. Liu XY, Liu YQ, Qu Y, Cheng MC, Xiao HB. Metabolomic profiling of emodin-induced cytotoxicity in human liver cells and mechanistic study. Toxicol Res. 2015;4:948–955. doi:10.1039/C4TX00246F
190. Yang D, Huang WY, Li YQ, et al. Acute and subchronic toxicity studies of rhein in immature and d-galactose-induced aged mice and its potential hepatotoxicity mechanisms. Drug Chem Toxicol. 2022;45:1119–1130. doi:10.1080/01480545.2020.1809670
191. He LN, Yang AH, Cui TY, et al. Reactive metabolite activation by CYP2C19-mediated rhein hepatotoxicity. Xenobiotica. 2015;45:361–372. doi:10.3109/00498254.2014.984794
192. Liu DM, Yang D, Zhou CY, et al. Aloe-emodin induces hepatotoxicity by the inhibition of multidrug resistance protein 2. Phytomedicine. 2020;68:153148. doi:10.1016/j.phymed.2019.153148
193. Quan Y, Gong L, He J, et al. Aloe emodin induces hepatotoxicity by activating NF-ƙB inflammatory pathway and P53 apoptosis pathway in zebrafish. Toxicol Lett. 2019;306:66–79. doi:10.1016/j.toxlet.2019.02.007
194. Yang W, Liu J, Zheng Y, et al. Study on chronic toxicity of rhubarb extract in Sprague-Dawley rats. Heliyon. 2022;8:e10907.
195. Liu C, Liu J, Zheng Y, et al. Subchronic oral toxicity study of rhubarb extract in Sprague-Dawley rats. Regul Toxicol Pharmacol. 2021;123:104921. doi:10.1016/j.yrtph.2021.104921
196. Cao C, Hui L, Li C, et al. In vitro study of the nephrotoxicity of total dahuang (Radix Et Rhizoma Rhei Palmati) anthraquinones and emodin in monolayer human proximal tubular epithelial cells cultured in a transwell chamber. J Tradit Chin Med. 2019;39:609–623.
197. Yang Z, Qin W, Chen D, et al. In vitro study of emodin-induced nephrotoxicity in human renal glomerular endothelial cells on a microfluidic chip. BIOCELL. 2022;47:125–131. doi:10.32604/biocell.2023.022937
198. Luo T, Li N, He YQ, et al. Emodin inhibits human sperm functions by reducing sperm [Ca2+]i and tyrosine phosphorylation. Reprod Toxicol. 2015;51:14–21. doi:10.1016/j.reprotox.2014.11.007
199. Oshida K, Hirakata M, Maeda A, Miyoshi T, Miyamoto Y. Toxicological effect of emodin in mouse testicular gene expression profile. J Appl Toxicol. 2011;31:790–800. doi:10.1002/jat.1637
200. Li GM, Chen JR, Zhang HQ, et al. Rhein activated Fas-induced apoptosis pathway causing cardiotoxicity in vitro and in vivo. Toxicol Lett. 2022;363:67–76. doi:10.1016/j.toxlet.2022.04.006
201. Sun YP, Liu JP. Blockade of emodin on amyloid-β 25-35-induced neurotoxicity in AβPP/PS1 mice and PC12 cells through activation of the class III phosphatidylinositol 3-kinase/Beclin-1/B-cell lymphoma 2 pathway. Planta Med. 2015;81:108–115. doi:10.1055/s-0034-1383410
202. Wang D, Wang XH, Yu X, et al. Pharmacokinetics of anthraquinones from medicinal plants. Front Pharmacol. 2021;12:638993. doi:10.3389/fphar.2021.638993
203. Lee JH, Kim JM, Kim C. Pharmacokinetic analysis of rhein in Rheum undulatum L. J Ethnopharmacol. 2003;84:5–9. doi:10.1016/S0378-8741(02)00222-2
204. Xun T, Wang X, Zhao J, et al. Advanced oxidation protein products regulate the pharmacokinetics of aloe-emodin, emodin, rhein, and chrysophanol in chronic kidney disease rats. Clin Complement Med Pharmacol. 2023;3:100087. doi:10.1016/j.ccmp.2023.100087
205. Miao H, Zhang SJ, Wu X, Li P, Zhao YY. Tryptophan metabolism as a target in gut microbiota, ageing and kidney disease. Int J Biol Sci. 2025;21:4374–4387. doi:10.7150/ijbs.115359
206. Zhao BR, Hu XR, Wang WD, Zhou Y. Cardiorenal syndrome: clinical diagnosis, molecular mechanisms and therapeutic strategies. Acta Pharmacol Sin. 2025;46:1539–1555. doi:10.1038/s41401-025-01476-z
207. Dong X, Wen R, Xiong Y, et al. Emodin alleviates CRS4-induced mitochondrial damage via activation of the PGC1α signaling. Phytother Res. 2024;38:1345–1357. doi:10.1002/ptr.8091
208. Ndumele CE, Rangaswami J, Chow SL, et al. Cardiovascular-kidney-metabolic health: a presidential advisory from the American heart association. Circulation. 2023;148:1606–1635. doi:10.1161/CIR.0000000000001184
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Integrated Small Intestine Microbiota and Serum Metabolomics Reveal the Potential Mechanisms of Wine Steaming in Alleviating Rhubarb-Induced Diarrhea

Bai YY, Tian R, Qian Y, Zhang Q, Zhao CB, Yan YG, Zhang L, Yue SJ, Tang YP
Journal of Inflammation Research 2024, 17:7851-7868
Published Date: 30 October 2024
Altered Gut Microbiota and Plasma Metabolome Profiles Characterize Depression Individuals with Ischemic Stroke: A Comparative Analysis
Cao N, Lv D, Liu Y, Zhang H, Zhang X
Neuropsychiatric Disease and Treatment 2025, 21:973-987
Published Date: 29 April 2025
Metabolomics and Gut Microbiota in Elderly Patients with Diabetic Peripheral Neuropathy and Sarcopenia
Wang X, Yu F, Cai Q, Li B, Li X, Fu C, Yu X, Yin W, Zeng S, Gao H, Cheng M
Diabetes, Metabolic Syndrome and Obesity 2025, 18:4335-4345
Published Date: 22 November 2025
Exploratory Pilot Multi-Omics Profiling of Gut Microbiota and Metabolic Features in Patients with Prolactinoma
Yang J, Nie D, Zhang Y, Li C
Cancer Management and Research 2026, 18:608026
Published Date: 16 June 2026