Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Metabolomics and Gut Microbiota in Elderly Patients with Diabetic Peripheral Neuropathy and Sarcopenia
Authors Wang X, Yu F, Cai Q, Li B, Li X, Fu C, Yu X, Yin W, Zeng S, Gao H, Cheng M
Received 30 April 2025
Accepted for publication 13 November 2025
Published 22 November 2025 Volume 2025:18 Pages 4335—4345
DOI https://doi.org/10.2147/DMSO.S537792
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Rebecca Baqiyyah Conway
Xue Wang,1– 3 Fei Yu,1– 3 Qian Cai,1– 3 Baoying Li,2,4 Xiaoli Li,5 Chunli Fu,1– 3 Xin Yu,1– 3 Wenbin Yin,1– 3 Shudong Zeng,1– 3 Haiqing Gao,1– 3 Mei Cheng1– 3
1Department of Geriatric Medicine & Laboratory of Gerontology and Anti-Aging Research, Qilu Hospital of Shandong University, Jinan, Shandong, People’s Republic of China; 2Key Laboratory of Cardiovascular Proteomics of Shandong Province, Qilu Hospital of Shandong University, Jinan, People’s Republic of China; 3Jinan Clinical Research Center for Geriatric Medicine (202132001), Jinan, Shandong, People’s Republic of China; 4Health Management Center (East Area), Qilu Hospital of Shandong University, Jinan, Shandong, People’s Republic of China; 5Department of Pharmacy, Qilu Hospital of Shandong University, Jinan, Shandong, People’s Republic of China
Correspondence: Mei Cheng, Department of Geriatric Medicine & Laboratory of Gerontology and Anti-Aging Research, Qilu Hospital of Shandong University, Jinan, Shandong, People’s Republic of China, Email [email protected]
Background: Diabetes peripheral neuropathy (DPN) is closely related to the occurrence and development of sarcopenia. However, the relevant early biological metabolites and their pathophysiological mechanism is unclear.
Aim: To explore the underlying mechanisms of Diabetic Peripheral Neuropathy with Sarcopenia (DPNS) by integrating metabolomics and 16S rRNA sequencing.
Methods: A total of 151 diabetic neuropathy patients were enrolled in the study. Untargeted metabolomics was performed using ultra-high-performance liquid. Chromatography-mass spectrometry, andgut microbiota was assessed through 16S rRNA sequencing. Differential metabolites and microbial taxa were identified, and theirassociations were explored using correlation analysis.
Results: A total of 376 differential metabolites were identified. Compared with the DPN group, the contents of glycerophosphocholine(GPC), taurine, and succinic acid in the DPNS group were significantly decreased, while that of 2-amino-1-methyl-6-phenylimidazo(4, 5-b)pyridine(PhIP), Sphingosine were increased. Gutmicrobiota analysis revealed reduced diversity in DPNS, with decreased beneficial genera (Faecalibacterium, Bacteroides) and increased pathogenic taxa (Streptococcus). Additionally, KEGG enrichment analysis showed that the mammalian target of necroptosis, sphingolipid metabolism, the mTOR signaling pathway, ABC transporters, and bile secretion pathways are closely related to DPNS.
Conclusion: We systematically explored the biomarkers and potential therapeutic targets in the patients with DPNS, which may provide new insights that may advance the treatment of sarcopenia.
Keywords: diabetes peripheral neuropathy, sarcopenia, gut microbiota, metabolome, biomarker
Introduction
Sarcopenia, a geriatric syndrome marked by progressive and generalized loss of skeletal muscle mass and function, arises from a complex, multifactorial pathogenesis that encompasses anabolic resistance, mitochondrial dysfunction, and chronic inflammation, yet its precise etiology remains incompletely delineated.1 Diabetic peripheral neuropathy (DPN), the most prevalent chronic complication of diabetes, affects up to 50% of diabetic patients.2 Intriguingly, our study revealed a strikingly high prevalence of Diabetic peripheral neuropathy with Sarcopenia (DPNS), reaching 30. 1%, which is significantly higher than the previously reported incidence of sarcopenia in diabetic patients (14. 8%).3 This coexistence of DPN and Sarcopenia has been shown to dramatically increase the risk of falls, fractures, all-cause mortality, and cardiovascular events in patients. Emerging evidence suggests a bidirectional relationship between these two conditions: DPN induces motor endplate reduction, axonal degeneration, and muscle denervation, thereby accelerating skeletal muscle atrophy and distal muscle weakness, which in turn exacerbates physical performance deterioration. Additionally, peripheral vascular disease and ischemia-reperfusion injury associated with DPN disrupt the skeletal muscle microenvironment, leading to an imbalance in muscle protein catabolism. Chronic inflammation and accumulation of advanced glycation end-products (AGEs) further inhibit anabolic pathways such as mTOR and activate the ubiquitin-proteasome system, resulting in decreased muscle protein synthesis and accelerated degradation.4–7 Despite these insights, the intricate interplay between DPN and Sarcopenia remains poorly understood and warrants further investigation. Given that the pathogenesis of DPNS may involve nutritional deficiencies, hormonal changes, insulin resistance, inflammation, neuro-muscular dysfunction, mitochondrial dysfunction, and genetic factors,8 elucidating the mechanisms underlying their interaction could provide a crucial theoretical foundation for developing targeted, combined intervention strategies.
The gut microbiota has emerged as a novel and promising research avenue, offering fresh insights into the pathogenesis of DPN and Sarcopenia. Accumulating evidence indicates that DPN patients exhibit significant alterations in their gut microbiota structure, characterized by a decline in beneficial bacteria that produce short-chain fatty acids (SCFAs) and a concurrent rise in harmful bacteria that generate endotoxins. This microbial dysbiosis impacts the pathological progression of DPN through multiple mechanisms, including inflammation and metabolic disturbances. Notably, interventions targeting the gut microbiota, such as fecal microbiota transplantation and probiotic supplementation, have shown efficacy in improving symptoms in DPN patients.9,10 In parallel, sarcopenia patients also display a marked reduction in gut microbial diversity.11 Emerging studies suggest that modulating the gut microbiota may represent a promising therapeutic approach for musculoskeletal diseases, including sarcopenia. Moreover, the application of metabolomics has further illuminated the pathophysiological processes underlying DPN and Sarcopenia. By elucidating their overall metabolic disturbances and underlying pathogenic mechanisms, metabolomics has the potential to identify novel biomarkers for clinical diagnosis and treatment. A key innovation in this field is the concept of the gut-muscle axis, which underscores the intricate bidirectional interactions between the gut and skeletal muscle. The gut microbiota produces a variety of metabolites that can influence muscle metabolism, growth, and repair through signaling pathways and by regulating inflammation. Conversely, muscle function can impact gut motility and barrier integrity. This dynamic interplay is crucial for nutrient absorption, energy homeostasis, and immune regulation. The gut–muscle axis concept offers a new lens through which to view the link between metabolic diseases and sarcopenia. Evidence shows that intestinal dysbiosis can intensify insulin resistance and muscle loss, a pattern especially evident in the comorbidity of metabolic syndrome and frailty syndrome. Older adults with diabetic peripheral neuropathy (DPN) frequently present with sarcopenia, yet diagnosis remains challenging: existing biomarkers lack sensitivity for early-stage changes, and the interplay between metabolic disturbances and microbiota-derived metabolites is still poorly understood. Although microbiota-targeted therapies have shown promise in metabolic syndrome, their efficacy in individuals with both DPN and sarcopenia has yet to be confirmed. This study aims to uncover the unique microbial–metabolic network disruptions in this population, providing a foundation for developing early diagnostic markers and personalized interventions. In addition, by unraveling the mechanisms of the gut-muscle axis, our study aims to provide a deeper understanding of the relationship between the gut microbiota and muscle health, thereby laying the groundwork for developing therapeutic strategies to improve muscle function and prevent related diseases.12,13
Given the complex interplay between DPN and Sarcopenia, as well as the emerging role of the gut microbiota and metabolic pathways in their pathogenesis, this study leverages metabolomics and gut microbiota analysis to identify serum metabolic biomarkers and characterize changes in microbial communities. Our goal is to elucidate the underlying mechanisms of DPNS and uncover potential therapeutic targets. By integrating these approaches, we aim to bridge the gap in understanding the multifaceted pathogenesis of DPNS and provide novel insights for targeted interventions.
Materials and Methods
Study Population
A total of 151 patients aged > 60 years were consecutively recruited from the Department of Geriatric Medicine, Qilu Hospital, Shandong University, Shandong Province, China, comprising diabetic neuropathy patients with sarcopenia (DPNS, n =35) and without sarcopenia (DPN, n = 116) were enrolled. From this patient population, we randomly selected 30 patients from each of the two groups. DPN was defined based on the standard criteria set by the 2022 Chinese Guidelines for the Prevention and Treatment of Type 2 Diabetes in the elderly in China.14 Sarcopenia was defined, based on the criteria recommended by the Asian Working Group for Sarcopenia (AWGS).15 Exclusion criteria included factors related to nerve damage, such as the use of neurotoxic drugs, vitamin B12 deficiency, renal insufficiency, cerebral infarction, cervical/lumbar spine disease, HIV infection, vasculitis, and hereditary neuropathies. Other exclusions were the use of antibiotics/probiotics within the past three months, the presence of other musculoskeletal diseases, significant limitations in limb mobility, severe organ dysfunction, hematologic disorders, cognitive/psychiatric impairments.
This study systematically evaluated the baseline characteristics and metabolic status of participants through a multidimensional data collection system. Standardized questionnaires were used to collect information on age, gender, and other demographic data. Height and weight were measured using calibrated instruments, and body mass index (BMI) was calculated. Blood pressure, waist circumference, hip circumference, and calf circumference were measured according to the World Health Organization (WHO) standards, and the waist-to-hip ratio was calculated. Fasting venous blood was collected from participants in the morning, and fasting blood glucose, lipid metabolism indicators, and liver function indicators were analyzed using an automated biochemical analyzer.
This study was approved by the Medical Ethics Committee of Qilu Hospital of Shandong University. All methods and analyses were conducted in accordance with the approved protocol and guidelines. Written informed consent was obtained from all study participants.
Skeletal Muscle Mass and Function Assessment
All participants underwent body composition analysis using the InBody 720 bioelectrical impedance analyzer. Body fat, muscle tissue, percentage of body fat (PBF), appendicular skeletal muscle mass (ASM), basal metabolic rate, visceral fat area (VFA), and bone mineral content were measured using bioelectrical impedance analysis (BIA) technology. The skeletal muscle mass index (ASMI) was calculated as ASM (in kilograms) divided by height squared (in meters squared) (ASMI = ASM/height²). The 6-meter walking speed test was conducted at least twice, and the results were recorded. Handgrip strength was measured using a Jamar dynamometer, with three repetitions performed to obtain the maximum value.
Metabolomics Analysis
Sample Processing and Data Analysis
All blood samples were centrifuged within 30 min of collection (3000 × g, 10 min, 4 °C). Serum aliquots were prepared within 1 h and transferred to −80 °C ultra-low-temperature freezers. After thawing, serum proteins were precipitated with methanol (100 µL serum + 300 µL methanol, incubated at −20 °C for 30 min, then centrifuged at 16000× g for 20 min). The supernatant was collected for ultra-high-performance liquid chromatography–mass spectrometry (UHPLC-MS) analysis.
Raw data were processed using MSDIAL software for peak extraction and metabolite identification (against HMDB, MassBank, and other databases). Differential metabolites were screened based on VIP > 1.0, fold change (FC) > 1.2 or < 0.8, and p < 0.05. KEGG enrichment analysis was performed using Fisher’s exact test, and significant pathways (P < 0.05) were identified after false discovery rate (FDR) correction for multiple comparisons.
Differential Metabolite Identification
All multivariate data analysis and modeling were conducted using R (version 4. 0. 3) and R packages. Data were centered by Pareto scaling. Principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA) were used to build models. All models were validated for overfitting using permutation tests. Differential metabolites were identified based on VIP values from OPLS-DA models and two-tailed Student’s t-tests on normalized raw data (p < 0.05). Metabolites with VIP > 1.0, FC > 1.2 or < 0.8, and p < 0.05 were considered statistically significant. Additionally, hierarchical clustering analysis of the identified differential metabolites was performed using R packages.
Gut Microbiota Analysis
Fecal samples (≥1 g) were collected from patients, snap-frozen in liquid nitrogen within 30 minutes of collection, and stored at −80 °C. PCR amplification was performed using Pfu high-fidelity DNA polymerase from Full Spectrum Gold. The amplified products were further purified and recovered using magnetic beads and quantified by fluorescence. Finally, sequencing libraries were prepared using the TruSeq Nano DNA LT Library Prep Kit from Illumina.
ELISA Validation
Serum concentrations of glutamate and succinate were measured using colorimetric assay kits. The optical density (OD) was determined using a microplate reader, and concentrations were calculated based on standard curves.
Statistical Methods
Statistical analyses were conducted using SPSS 26.0 software. The normality of the data was assessed using the Kolmogorov–Smirnov (K-S) test. Normally distributed continuous data were presented as mean ± standard deviation and analyzed using independent samples t-tests to compare differences between groups. Categorical data were expressed as proportions and analyzed using chi-square tests. A p-value of less than 0. 05 was considered statistically significant.
Results
Clinical Characteristics of Participants
The clinical characteristics of patients with diabetic peripheral neuropathy with sarcopenia are presented in Table 1. Compared with the DPN group, the DPNS group exhibited significant increases in age, duration of diabetes, pulse pressure, fasting blood glucose, HbA1c, waist circumference, hip circumference, and visceral fat area (P<0.01). In contrast, body weight, BMI, albumin, calf circumference, grip strength, basal metabolic rate, bone mineral content, and SMI were significantly decreased (P<0.01). Additionally, the levels of triglycerides, total cholesterol, low-density lipoprotein, and high-density lipoprotein did not show significant differences between the two groups.
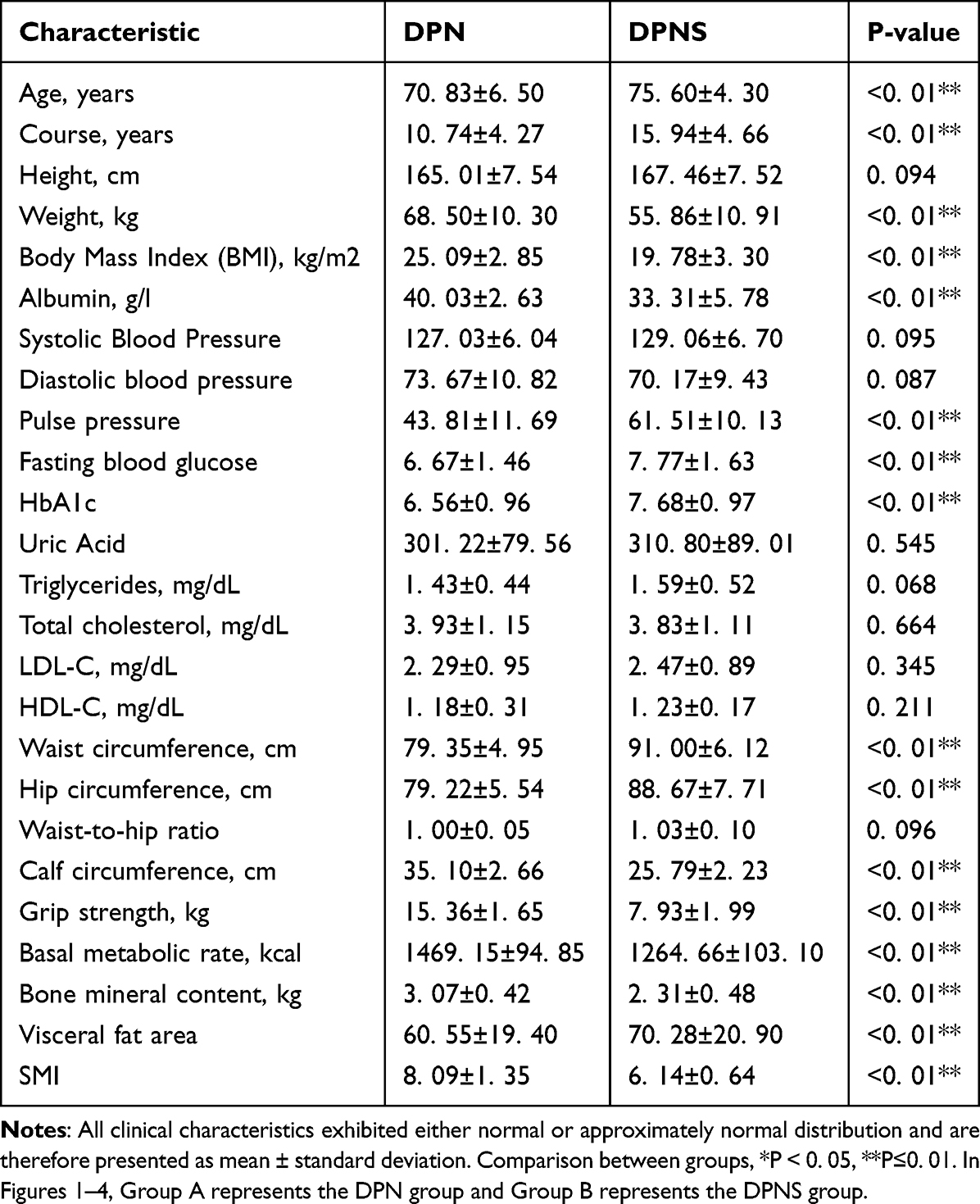 |
Table 1 Clinical Characteristics of Participants in the DPNS and DPN Control Groups |
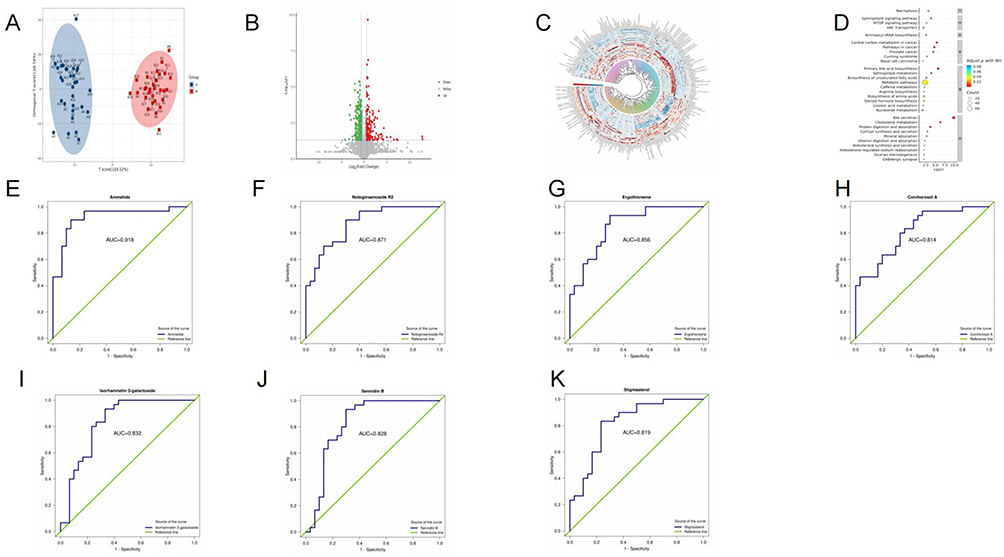 |
Figure 1 (A) OPLS-DA score plots. (B) Volcano plot. (C) Cluster heatmap. (D) KEGG signaling pathways of differentially regulated metabolites. (E–K) ROC analysis. |
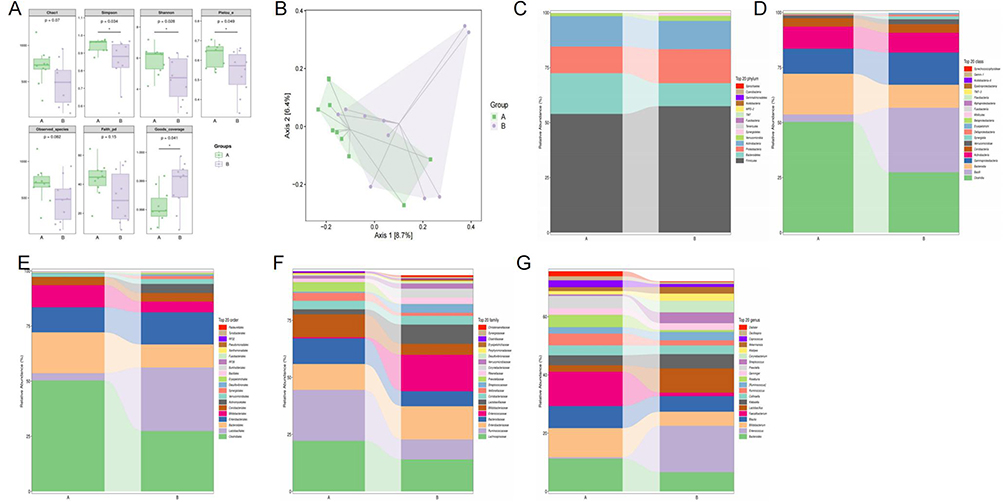 |
Figure 2 (A) Alpha diversity indexes (Chao 1, Simpson, Shannon, Pielou_e, Observed_species, Faith_pd, and Goods_coverage indexes). (B) Jaccard PCoA analysis of gut microbiota based on the ASV data. (C–G) Composition of gut microbiota at phylum, class, order, family, and genus levels. |
 |
Figure 3 (A) Cluster heatmap indicating significantly altered gut microbiota at genus levels. (B) LDA scores of gut microbiota. (C) Bar plot of differential taxonomic units between groups based on taxonomic hierarchy tree. (D and E) Manhattan plot of differential ASVs/OTUs based on metagenomeSeq analysis. |
 |
Figure 4 (A) Correlation coefficient matrix heatmap. (B) hierarchical clustering heatmap. (C) histogram. |
Detection of Dysregulated Metabolites by Untargeted Metabolomics Analysis
The OPLS-DA model was applied to identify differentially expressed metabolites between groups. The results demonstrated high predictive accuracy (R2Y and Q2 ≥ 0.5), confirming the model’s stability, reliability, and robust discriminative capacity (Figure 1A).
A total of 376 significantly differential metabolites were identified, primarily including lipids and lipid-like molecules, organic heterocyclic compounds, organic acids and derivatives, phenylpropanoids and polyketides, and benzene-related compounds. Volcano plot (Figure 1B) and cluster heatmap (Figure 1C) revealed a series of significantly altered metabolites. The abundance of these metabolites was represented by the log-transformed fold change between the two groups.
We employed KEGG enrichment analysis to identify the metabolic pathways associated with DPNS. The results showed that the mammalian target of necroptosis, sphingolipid metabolism, mTOR signaling pathway, ABC transporters, bile secretion, cholesterol metabolism, digestion and absorption of proteins and vitamins, synthesis and secretion of cortisol and aldosterone, ovarian steroidogenesis, and GABAergic synapses (Figure 1D).
ROC analysis was further conducted on the differential metabolites. Compared with the DPN group, the levels of Ammelide, Notoginsenoside R2, Corchorosol A, and Isorhamnetin 3-galactoside were increased in the DPNS group, while the levels of Ergothioneine and Stigmasterol were decreased. The AUC values of these significantly differential metabolites were all greater than 0.8 (Figure 1E–K).
Alteration of Gut Microbiota in DPNS
The α-diversity indexes have Chao 1, Simpson, Shannon, Pielou_e, Observed_species, Faith_pd, and Goods_coverage (Figure 2A). The richness and diversity of DPNS group were lower than DPN group. Moreover, the overall structure of gut microbiota obtained through Jaccard based PCoA indicates that separation was showed in the control group and the experimental group (Figure 2B). Based on univariate analysis, the composition of gut microbiota was analyzed at the phylum, class, order, family, and genus levels (Figure 2C–G).
Clustering heatmap-based species composition analysis revealed significant genus-level changes (Figure 3A). Genus levels plotted based on log-transformed fold change in abundance between the two groups (Figure 3B). The linear discriminant analysis effect size and cladogramd is playing differentially abundant taxonomic clade analysis were compared with between the two groups (Figure 3C). Dysfunction of gut microbiota was found in the DPNS, such as increase in [Eubacterium] and Streptococcus, and decrease in Faecalibacterium and Bacteroides. Furthermore, the Manhattan plot was utilized to visualize the results of the metagenomeSeq analysis, revealing upregulated features in the DPN group, including Bifidobacteriales, Bacteroidales and Clostridiales, while only Clostridiales were observed to be upregulated in the DPNS group (Figure 3D and E).
We further validated the possible interaction between the gut microbiome and serum metabolome by analyzing the correlation between the two datasets. Spearman correlation analysis between gut microbiota and metabolites revealed several significant associations in the two groups. Correlation coefficient matrix heatmap, hierarchical clustering heatmap, and histogram between control group and experimental group (Figure 4A–C). A total of 22 of 29 genera had notable relationships with circulating differential metabolites. Bacteroides, Faecalibacterium, Sutterella, Enterococcus, Oscillospira were all significantly correlated with at least 4 serum metabolites. For example, The increase in abundance of Faecalibacterium was mainly positively correlated with Tomatidine, Haematoporphyrin, and negatively correlated with 4-Benzyl-7-hydroxy-3-phenylcoumarin (P<0.05). The histogram of correlation coefficients indicates that the 25 pairs of data with positive correlation (r≥0.7) may represent that the abundance of certain metabolites or microbiota is positively correlated with the severity of DPNS. In contrast, the 14 pairs of data with negative correlation (r≤-0. 7) may suggest that certain factors have a protective effect against the disease or are inversely related to disease progression.
Discussion
Diabetic peripheral neuropathy with sarcopenia is increasingly prevalent among diabetic patients, posing significant health risks. The synergistic effects of nerve damage and muscle loss can further complicate disease management and contribute to higher healthcare costs. Given these challenges, identifying biomarkers for early detection and developing novel therapeutic strategies are urgently needed.
We conducted muscle function assessments and analyzed clinical parameters in the study cohort. The results revealed that DPNS exhibited advanced age, prolonged duration of type 2 diabetes mellitus (T2DM), and significantly reduced grip strength compared to DPN. In this study, we observed a significant decrease in basal metabolic rate and bone mineral content in patients from the DPNS group, along with a marked increase in visceral fat area compared to the DPN group. This phenomenon may be associated with the pathological mechanisms of sarcopenia: the reduction in muscle mass leads to a decrease in basal metabolic rate, thereby weakening the body’s energy expenditure capacity and promoting the abnormal accumulation of excess energy as fat in the visceral region. Hormonal abnormalities related to diabetes may further exacerbate insulin resistance, creating a vicious cycle. Notably, sarcopenia is often accompanied by a chronic inflammatory state. Elevated inflammatory cytokines not only disrupt adipocyte metabolic function but also promote their proliferation and differentiation, serving as a significant driving factor for the expansion of visceral fat. However, some studies have indicated that non-obese sarcopenia (e g, in patients with malnutrition or chronic wasting diseases) may exhibit reduced visceral fat, suggesting that sarcopenia caused by different etiologies may have distinct fat distribution patterns.16,17
Moreover, serum laboratory tests showed significant differences in albumin, HbA1c, blood glucose, and blood lipid levels between the two groups, which is consistent with previous studies in the literature.
In this study, we performed untargeted metabolomics analysis and found that the levels of metabolites such as glycerophosphocholine (GPC), taurine, succinic acid, and leucine were decreased in the serum of DPNS patients, while the level of 2-amino-1-methyl-6-phenylimidazo(4, 5-b)pyridine(PhIP) was increased. Through 16s rRNA analysis, the data showed that the abundance and diversity of the microbiota in the DPNSgroup were significantly reduced. Moreover, combined analysis revealed that the upregulated bacterial genera in the DPN group and the DPNS group were different.
Hyperosmotic stress, triggered by elevated extracellular osmolarity, induces intracellular dehydration. During ageing, this dehydration process progresses gradually alongside accumulating hyperosmotic stress, which not only correlates with inflammatory responses but also causes cellular volume loss and impairs both structural integrity and functional capacity of intracellular proteins. Several studies suggest that18 may have severe effects on muscle, leading to catabolism, anabolic resistance, and muscle wasting as well as impaired muscle contractile capacity.19,20 Glycerophosphocholine(GPC) is an abundant osmoprotective renal medullary organic osmolyte. Under normal physiological conditions, the kidneys adopt a series of adaptive measures to cope with high osmolarity, one of which is the accumulation of organic osmolytes such as GPC, betaine, inositol, sorbitol, and free amino acids.21,22 Among them, GPC is a major organic osmolyte. Therefore, if the level of GPC decreases and fails to maintain the normal osmotic balance in the body, it will affect the muscles, leading to muscle atrophy. Previous studies have identified through metabolomics that GPC is decreased in Duchenne muscular dystrophy(DMD) muscles.23 Thus, it is reasonable to assume that patients with sarcopenia also experience a decline in GPC levels, which is consistent with the results observed in the DPNS group. Therefore, GPC may represent a potential predictive biomarker for DPNS.
Additionally, compared with the DPN group, the level of Taurine was decreased in the DPNS group. Taurine is a sulfur-containing amino acid in the body and serves as an important bioactive substance with multiple functions, including antioxidant activity, osmoregulation, neuroprotection, stabilization of cell membranes, and involvement in ion transport.24 Muscle atrophy is often accompanied by an increase in protein degradation within muscle tissue, which is a complex pathological process. Taurine, as an effective bioactive substance, plays a role in inhibiting muscle atrophy through various mechanisms, such as reducing the activity of protein degradation pathways, suppressing the release of inflammatory factors, activating muscle satellite cells to participate in muscle repair and regeneration, enhancing muscle energy metabolism and calcium ion homeostasis, and inhibiting signaling pathways related to apoptosis. Thus, the significant reduction in taurine levels may provide a new direction for identifying predictive biomarkers of the disease.25,26
PhIP is a common heterocyclic amine compound that is significantly upregulated in patients with DPNS.27 Based on relevant literature, PhIP generates free radicals during its metabolism in the body, inducing oxidative stress and DNA damage. These effects can damage nerve cells, inhibit muscle protein synthesis, and accelerate muscle degradation, thereby exacerbating sarcopenia symptoms.28 Additionally, PhIP significantly alters the composition of the gut microbiota, decreasing the abundance of beneficial bacteria (such as Lactobacillus) while increasing the relative abundance of potentially harmful bacteria (such as certain Clostridium species). This microbial imbalance may impair gut barrier function and subsequently trigger systemic inflammatory responses. Inflammation is a common pathological feature of both Sarcopenia and DPN, and PhIP-induced inflammation may worsen the condition through multiple pathways. Furthermore, PhIP significantly inhibits the lipid metabolism functions of the gut microbiota, including glycerophospholipid and linoleic acid metabolism, further affecting the health of nerves and muscles.29 In summary, the upregulation of PhIP levels may exacerbate the condition of DPNS patients through multiple mechanisms, including inducing oxidative stress, altering gut microbiota composition, and disrupting lipid metabolism. These mechanisms interact with each other, further aggravating nerve damage and muscle atrophy in patients. Therefore, in the investigation and therapeutic development of DPNS, it is imperative to consider the potential impacts of PhIP exposure and elucidate its underlying mechanisms.
Recent investigations from both animal and human studies have illuminated gut microbiota can affect the host’s muscle mass and function. In this study, we found that the abundance and diversity of the microbiota in the DPNS group were significantly reduced. At the genus level, for example, the abundance of Faecalibacterium was decreased in DPNS patients. Numerous studies have underscored that low levels of Faecalibacterium are correlated with inflammatory conditions.30 Once the gut microbiota becomes imbalanced, it can lead to the entry of bacterial-derived inflammatory compounds into the circulation, thereby affecting muscle metabolism and mitochondrial function. We further utilized metagenomeSeq analysis and presented the differential ASVs/OTUs using a Manhattan plot, revealing significant downregulation of Bacteroides, Clostridium, Blautia, Bifidobacterium, and Faecalibacterium at the genus level.
The correlation analysis has revealed a significant association between gut microbiota (Faecalibacterium) and metabolites (Tomatidine), indicating that the gut−muscle axis is instrumental in the pathogenesis of DPNS. Faecalibacterium is a beneficial gut microbiota that produces butyrate, which can enhance intestinal immune regulation.30–33 Tomatidine, known for its anti-inflammatory and antioxidant properties, can inhibit systemic inflammation.34 Both may act synergistically through the “gut-muscle-nerve axis” to mitigate the inflammatory microenvironment in diabetic peripheral neuropathy with sarcopenia (DPNS). Tomatidine may also influence host metabolism (eg, lipid metabolism),35 indirectly modulating the composition of gut microbiota and promoting the proliferation of beneficial bacteria such as Faecalibacterium, thereby forming a positive feedback loop. In this study, we found that the decreased abundance of Faecalibacterium was positively correlated with the downregulation of Tomatidine metabolites, suggesting that both may influence glucose metabolism, inflammation, and muscle mass through a common pathway. Additionally, the histogram of correlation coefficients illustrates the distribution of correlation coefficients between metabolomics and gut microbiota. Although most variables do not exhibit significant correlations, a few strongly correlated variables may hold substantial significance for understanding disease mechanisms. We will focus on the 25 positively correlated and 14 negatively correlated variable pairs to further investigate their biological roles in the disease. These findings provide direction for subsequent in-depth research.
This study has several limitations. First, owing to time constraints, the sample size was relatively small; future work will expand the cohort to enhance the reliability of the findings. Second, as a cross-sectional investigation, we are currently unable to establish causal relationships or elucidate the specific molecular mechanisms linking serum metabolite alterations and gut-microbiota dysbiosis to diabetic peripheral neuropathy complicated by sarcopenia. Subsequent studies will employ animal experiments and/or longitudinal cohort designs to validate and further explore these underlying mechanisms.
Conclusion
This study systematically deciphered the unique “metabolome-microbiome” co-dysregulation pattern in DPNS by integrating metabolomics and gut microbiota multi-omics technologies. We identified several key metabolic biomarkers, including taurine, glycerophosphocholine, and succinate, as well as characteristic bacterial genera such as Bacteroides, Clostridium, Blautia, Bifidobacterium, and Faecalibacterium. Their dynamic changes not only serve as a non-invasive biomarker panel for early diagnosis of DPNS but also deeply participate in disease progression through the gut-muscle axis, particularly via the microbiota-host cometabolic network regulating metabolic homeostasis. Our findings confirm a significant association between metabolic pathway alterations and the abundance of specific gut microbiota, providing novel insights into the pathogenesis of DPNS and offering a new therapeutic direction for developing targeted microbiota reshaping strategies or precise metabolic pathway interventions.
Data Sharing Statement
The raw data supporting the conclusions of this article will be made available by the first author, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by the Medical Ethics Committee of the Qilu Hospital of Shandong University. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Tournadre A, Vial G, Capel F, et al. Sarcopenia. Joint Bone Spine. 2019;86(3):309–314. doi:10.1016/j.jbspin.2018.08.001
2. Feldman EL, Callaghan BC, Pop-Busui R, et al. Diabetic neuropathy. Nat Rev Dis Primers. 2019;5(1):42. doi:10.1038/s41572-019-0092-1
3. De Freitas MM, De Oliveira VLP, Grassi T, et al. Difference in sarcopenia prevalence and associated factors according to 2010 and 2018 European consensus (EWGSOP) in elderly patients with type 2 diabetes mellitus. Exp Gerontol. 2020;132:110835. doi:10.1016/j.exger.2020.110835
4. Fang Y, Yuan X, Zhang Q, et al. Causality between sarcopenia and diabetic neuropathy. Front Endocrinol. 2024;15:1428835. doi:10.3389/fendo.2024.1428835
5. Oh TJ, Kang S, Lee JE, et al. Association between deterioration in muscle strength and peripheral neuropathy in people with diabetes. J Diabetes Complications. 2019;33(8):598–601. doi:10.1016/j.jdiacomp.2019.04.007
6. Wannarong T, Sukpornchairak P, Naweera W, et al. Association between diabetic peripheral neuropathy and sarcopenia: a systematic review and meta-analysis. Geriatr Gerontol Int. 2022;22(9):785–789. doi:10.1111/ggi.14462
7. Sanz-Cánovas J, López-Sampalo A, Cobos-Palacios L, et al. Management of type 2 diabetes mellitus in elderly patients with frailty and/or sarcopenia. Int J Environ Res Public Health. 2022;19(14):8677. doi:10.3390/ijerph19148677
8. Papadopoulou SK. Sarcopenia: a contemporary health problem among older adult populations. Nutrients. 2020;12(5):1293. doi:10.3390/nu12051293
9. Zhang T, Cheng JK, Hu YM. Gut microbiota as a promising therapeutic target for age-related sarcopenia. Ageing Res Rev. 2022;81:101739. doi:10.1016/j.arr.2022.101739
10. Tanase DM, Gosav EM, Neculae E, et al. Role of gut microbiota on onset and progression of microvascular complications of type 2 diabetes (T2DM). Nutrients. 2020;12(12):3719. doi:10.3390/nu12123719
11. Ticinesi A, Nouvenne A, Cerundolo N, et al. Gut microbiota, muscle mass and function in aging: a focus on physical frailty and sarcopenia. Nutrients. 2019;11(7):1633. doi:10.3390/nu11071633
12. Mancin L, Wu GD, Paoli A. Gut microbiota-bile acid-skeletal muscle axis. Trends Microbiol. 2023;31(3):254–269. doi:10.1016/j.tim.2022.10.003
13. Chen LH, Chang SS, Chang HY, et al. Probiotic supplementation attenuates age-related sarcopenia via the gut-muscle axis in SAMP8 mice. J Cachexia Sarcopenia Muscle. 2022;13(1):515–531. doi:10.1002/jcsm.12849
14. Prevention D, Treatment of Clinical Guidelines Writing Group. Clinical guidelines for prevention and treatment of type 2 diabetes mellitus in the elderly in China (2022 edition)] [J. Zhonghua Nei Ke Za Zhi. 2022;61(1):12–50. doi:10.3760/cma.j.cn112138-20211027-00751
15. Chen LK, Woo J, Assantachai P, et al. Asian working group for sarcopenia: 2019 consensus update on sarcopenia diagnosis and treatment. J Am Med Dir Assoc. 2020;21(3):300–7.e2. doi:10.1016/j.jamda.2019.12.012
16. Sato M, Tamura Y, Murao Y, et al. Coexistence of high visceral fat area and sarcopenia is associated with atherosclerotic markers in old-old patients with diabetes: a cross-sectional study. J Diabetes Investig. 2024;15(10):1510–1518. doi:10.1111/jdi.14274
17. Xiao T, Zou S, Luo T, et al. Prevalence and risk factors of osteosarcopenia in elderly patients with uncontrolled type 2 diabetes. Endocrine. 2025;87(1):59–66. doi:10.1007/s12020-024-04001-0
18. Martínez-Navarro I, Montoya-Vieco A, Collado E, et al. Muscle cramping in the marathon: dehydration and electrolyte depletion vs. muscle damage. J Strength Cond Res. 2022;36(6):1629–1635. doi:10.1519/JSC.0000000000003713
19. Sumida TS. Hyperosmotic stress response regulates interstitial homeostasis and pathogenic inflammation. J Biochem. 2023;173(3):159–166. doi:10.1093/jb/mvad009
20. Stöhr D, Rehm M. Linking hyperosmotic stress and apoptotic sensitivity. Febs J. 2021;288(6):1800–1803. doi:10.1111/febs.15520
21. Baumgartner C, Wolf P, Beiglböck H, et al. Potential role of skeletal muscle glycerophosphocholine in response to altered fluid balance in humans: an in vivo nuclear magnetic resonance study. Am J Physiol Endocrinol Metab. 2023;324(4):E339–e46. doi:10.1152/ajpendo.00286.2022
22. Cikes D, Leutner M, Cronin SJ, et al. Gpcpd1-GPC metabolic pathway is dysfunctional in aging and its deficiency severely perturbs glucose metabolism. Nat Aging. 2024;4(1):80–94. doi:10.1038/s43587-023-00551-6
23. Dabaj I, Ferey J, Marguet F, et al. Muscle metabolic remodelling patterns in Duchenne muscular dystrophy revealed by ultra-high-resolution mass spectrometry imaging. Sci Rep. 2021;11(1):1906. doi:10.1038/s41598-021-81090-1
24. Seidel U, Huebbe P, Rimbach G. Taurine: a regulator of cellular redox homeostasis and skeletal muscle function. Mol Nutr Food Res. 2019;63(16):e1800569. doi:10.1002/mnfr.201800569
25. Barbiera A, Sorrentino S, Lepore E, et al. Taurine attenuates catabolic processes related to the onset of sarcopenia. Int J Mol Sci. 2020;21(22):8865. doi:10.3390/ijms21228865
26. Barbiera A, Sorrentino S, Fard D, et al. Taurine administration counteracts aging-associated impingement of skeletal muscle regeneration by reducing inflammation and oxidative stress. Antioxidants. 2022;11(5):1016. doi:10.3390/antiox11051016
27. Zhang P, Li T, Liu C, et al. Nano-sulforaphane attenuates PhIP-induced early abnormal embryonic neuro-development. Ann Anat. 2021;233:151617. doi:10.1016/j.aanat.2020.151617
28. Yang X, Peng H, Luo Z, et al. The dietary carcinogen PhIP activates p53-dependent DNA damage response in the colon of CYP1A-humanized mice. Biofactors. 2021;47(4):612–626. doi:10.1002/biof.1730
29. Zhao X, Liu H, Zhou X, et al. 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine induced colon injury by disrupting the intestinal bacterial composition and lipid metabolic pathways in rats. J Agric Food Chem. 2021;69(1):437–446. doi:10.1021/acs.jafc.0c06588
30. Leylabadlo HE, Ghotaslou R, Feizabadi MM, et al. The critical role of Faecalibacterium prausnitzii in human health: an overview. Microb Pathog. 2020;149:104344. doi:10.1016/j.micpath.2020.104344
31. Martín R, Rios-Covian D, Huillet E, et al. Faecalibacterium: a bacterial genus with promising human health applications. FEMS Microbiol Rev. 2023;47(4). doi:10.1093/femsre/fuad039
32. Wang X, Li Y, Wang X, et al. Faecalibacterium prausnitzii supplementation prevents intestinal barrier injury and gut microflora dysbiosis induced by sleep deprivation. Nutrients. 2024;16(8):1100.
33. Azuma N, Mawatari T, Saito Y, et al. Effect of continuous ingestion of bifidobacteria and dietary fiber on improvement in cognitive function: a randomized, double-blind, placebo-controlled trial. Nutrients. 2023;15(19):4175. doi:10.3390/nu15194175
34. Abdulaal WH, Omar UM, Zeyadi M, et al. Modulation of the crosstalk between Keap1/Nrf2/HO-1 and NF-κB signaling pathways by Tomatidine protects against inflammation/oxidative stress-driven fulminant hepatic failure in mice. Int Immunopharmacol. 2024;130:111732. doi:10.1016/j.intimp.2024.111732
35. Cai L, Hou B, Hu J. Tomatidine ameliorates high-fat-diet/streptozocin (HFD/STZ)-induced type 2 diabetes mellitus in mice. Arch Physiol Biochem. 2024;130(6):848–853. doi:10.1080/13813455.2023.2298404
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Association Between Sarcopenia and Diabetes: From Pathophysiology Mechanism to Therapeutic Strategy
Chen H, Huang X, Dong M, Wen S, Zhou L, Yuan X
Diabetes, Metabolic Syndrome and Obesity 2023, 16:1541-1554
Published Date: 30 May 2023
Altered Gut Microbiota and Plasma Metabolome Profiles Characterize Depression Individuals with Ischemic Stroke: A Comparative Analysis
Cao N, Lv D, Liu Y, Zhang H, Zhang X
Neuropsychiatric Disease and Treatment 2025, 21:973-987
Published Date: 29 April 2025
Rhubarb: Traditional Uses, Phytochemistry, Multiomics-Based Novel Pharmacological and Toxicological Mechanisms
Miao H, Wang KE, Li P, Zhao YY
Drug Design, Development and Therapy 2025, 19:9457-9480
Published Date: 23 October 2025
Exploratory Pilot Multi-Omics Profiling of Gut Microbiota and Metabolic Features in Patients with Prolactinoma
Yang J, Nie D, Zhang Y, Li C
Cancer Management and Research 2026, 18:608026
Published Date: 16 June 2026