Back to Journals » Journal of Pain Research » Volume 19
Exploring Causal Associations Between Plasma Metabolites and Sciatica: A Two-Sample Mendelian Randomization Analysis
Authors Yu S, Wei Q, Luo Y, Wu G
Received 14 October 2025
Accepted for publication 22 January 2026
Published 13 February 2026 Volume 2026:19 568746
DOI https://doi.org/10.2147/JPR.S568746
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Alaa Abd-Elsayed
Shaoyong Yu,1 Qiuyu Wei,2 Yi Luo,2 Gang Wu2
1Traumatic Joint First Ward, Liuzhou Traditional Chinese Medicine Hospital, Liuzhou, Guangxi, People’s Republic of China; 2Department of Neurosurgery, Liuzhou Traditional Chinese Medicine Hospital, Liuzhou, Guangxi, People’s Republic of China
Correspondence: Qiuyu Wei, Department of Neurosurgery, Liuzhou Traditional Chinese Medicine Hospital, Liuzhou, 545000, People’s Republic of China, Email [email protected] Gang Wu, Department of Neurosurgery, Liuzhou Traditional Chinese Medicine Hospital, Liuzhou, 545000, People’s Republic of China, Email [email protected]
Background: Sciatica is a prevalent and highly disabling radicular pain syndrome, often involving mixed nociceptive and neuropathic components, which severely impacts patients’ quality of life and work capacity, underscoring the need to identify biomarkers for early intervention. Although recent metabolomics studies have revealed significant associations between altered plasma metabolite levels and sciatica, the causal relationship with plasma metabolites remains unverified. This study aims to analyze the causal association between plasma metabolites and sciatica from a genetic perspective and provide relevant evidence.
Methods: Public datasets comprising 1,400 plasma metabolites (as exposures) and sciatica (as outcome) were collected. Mendelian randomization analyses were performed using Rstudio, incorporating inverse-variance weighted, MR-Egger, weighted median, simple mode, and weighted mode methods. Robustness and reliability were ensured through heterogeneity testing, horizontal pleiotropy assessment, and Steiger directionality tests.
Results: We identified four plasma metabolites with significant causal relationships to sciatica after false discovery rate correction (PFDR < 0.05). Two distinct forms of the bilirubin degradation product C17H20N2O5 were identified as risk factors (Form 1: OR = 1.040, 95% CI = 1.015– 1.066, P = 0.001, PFDR=0.003; Form 2: OR = 1.039, 95% CI = 1.014– 1.065, P = 0.002, PFDR = 0.004). In contrast, N-Acetyl-β-alanine (OR = 0.960, 95% CI = 0.931– 0.991, P = 0.011, PFDR = 0.014) and Glycosyl-N-stearoyl-sphingosine (d18:1/18:0) (OR = 0.953, 95% CI = 0.920– 0.989, P = 0.010, PFDR = 0.012) were protective factors. The analysis was based on 106 independent instrumental variables (single nucleotide polymorphisms).
Conclusion: This study provides genetic evidence supporting a causal role of specific plasma metabolites in sciatica risk, highlighting the involvement of bilirubin and amino acid/sphingolipid metabolism in its pathogenesis. These findings offer novel insights into the biology of sciatica and may inform future research into therapeutic strategies.
Keywords: plasma metabolites, sciatica, causal association, mendelian randomization, genetics, single nucleotide polymorphism, genome-wide association study
Introduction
Sciatica, a common form of radiculopathy, is characterized by radiating pain originating from pathology of the lumbosacral nerve roots (L4-S1).1 It represents a mixed pain syndrome encompassing both neuropathic (due to nerve root involvement) and nociceptive (inflammatory) components, with pain typically projecting along specific dermatomes.2 Its prevalence ranges from 2–5% in the general population but escalates to 43% among specific occupational groups, indicating substantial morbidity and disability risks.3 While lumbar disc herniation is the most common structural cause, accounting for a majority of cases, the resultant pain is mediated not only by mechanical compression but also by a cascade of inflammatory mediators (eg, cytokines, chemokines, prostaglandins) released from the herniated disc material and surrounding tissues.4,5 This biochemical dimension highlights the significant, yet underexplored, role of metabolic pathways in sciatica pathogenesis. Although acute sciatica generally has a favorable prognosis, approximately 45% of patients still experience persistent pain one year later, which severely impacts quality of life and imposes a significant socioeconomic burden.6 Therefore, elucidating the pathogenesis of sciatica and identifying early-stage biomarkers and therapeutic targets has become a crucial objective for achieving precision medicine.
The metabolome, comprising the repertoire of small-molecule products within a biological system, provides a dynamic readout of physiological state and disease processes.7 Consequently, alterations in plasma metabolite profiles are increasingly recognized for their role in the pathophysiology of neuropathic pain, including sciatica. Specifically in sciatica, a neuropathic pain condition often stemming from lumbar radiculopathy, alterations in circulating metabolites are postulated to mirror and contribute to the complex local pathophysiology. This extends beyond correlations with pain intensity to encompass several key mechanistic layers. Firstly, local nerve root compression or irritation induces oxidative stress, with markers like lipid peroxidation products (eg, MDA) and depleted antioxidants potentially serving as both drivers and indicators of neuronal and glial cell injury.8 Secondly, dysregulated lipid metabolism contributes directly to the generation of pro-inflammatory lipid mediators (eg, specific prostaglandins and leukotrienes), which exacerbate neuroinflammation at the nerve root level, sensitizing nociceptors.9 Critically, this neuroinflammation—mediated by activated glial cells (microglia and astrocytes) in the spinal dorsal horn and possibly at the dorsal root ganglion—is a pivotal mechanism underlying central sensitization and the transition from acute to chronic sciatica pain.10 Metabolic shifts can fuel this sustained glial activation. Finally, while the nerve root microenvironment and cerebrospinal fluid are partially shielded by barriers, plasma metabolite profiles may reflect these local changes due to several factors: compromised blood-nerve/blood-CSF barriers in pathological states, passive diffusion or active transport of signaling molecules into the circulation, and the systemic immune response elicited by the local injury, which releases mediators into the bloodstream.11–13 Thus, investigating plasma metabolites offers a viable, albeit indirect, window into the metabolic perturbations involved in oxidative damage, inflammatory lipid signaling, and neuroimmune interactions that characterize sciatica progression.14,15 Indeed, recent observational metabolomic studies have begun to identify specific plasma metabolite signatures associated with sciatica.16 However, existing evidence is predominantly derived from cross-sectional observational studies with limited sample sizes, making it difficult to eliminate interference from confounding factors and reverse causality. Consequently, while associations have been noted, a comprehensive understanding of the causal relationship between plasma metabolites and sciatica remains lacking.
To address this knowledge gap, Mendelian Randomization (MR) serves as a powerful causal inference method that utilizes genetic variants as instrumental variables, thereby substantially reducing confounding biases and reverse causality issues inherent in conventional observational studies.17 With the growing availability of genome-wide association study (GWAS) summary statistics for both plasma metabolites and sciatica, MR offers a unique opportunity to explore lifelong causal relationships between metabolic profiles and disease susceptibility. However, several methodological considerations specific to pain research must be acknowledged. First, genetic variants proxy for lifelong differences in exposure levels rather than short-term fluctuations, meaning MR is more suitable for identifying factors influencing long-term disease risk or development rather than acute pain episodes or immediate treatment targets. Second, while plasma metabolites may reflect systemic metabolic states, their association with localized nerve root inflammation—a central pathophysiological feature of sciatica—is likely indirect and mediated through complex neuroimmune and vascular interactions. Therefore, although MR can provide robust evidence regarding etiologic susceptibility, significant causal findings do not necessarily translate directly into actionable targets for acute therapeutic intervention. Despite these limitations, MR remains a valuable approach for generating novel hypotheses about the role of metabolic pathways in sciatica pathogenesis.
A recent MR study by Ren16 et al (2024) has investigated causal links between serum metabolites and a composite phenotype of sciatica and low back pain. Building upon and extending this approach, our study employs several key refinements: we utilize a substantially larger plasma metabolome dataset (encompassing >1,400 metabolites vs 486), apply stringent false discovery rate (FDR) correction to control for multiple testing, and aim for a more specific focus on sciatica (notwithstanding that the FinnGen phenotype M13, “sciatica accompanied by low back pain,” also incorporates a low back pain component, as detailed in the Limitations). These methodological distinctions are designed to enhance the specificity and statistical rigor of causal inference in this domain.
In this study, we performed a comprehensive two-sample MR analysis to investigate the lifelong causal effects of genetically predicted circulating metabolite levels on susceptibility to sciatica. Specifically, we aimed to identify which specific plasma metabolites exert a causal effect on sciatica risk. By identifying robust causal metabolites, we seek to pinpoint key metabolic pathways involved in the disease onset, thereby providing novel insights for understanding its pathogenesis and informing future strategies for early risk assessment.
Materials and Methods
Study Design
This study employed a two-sample MR approach to systematically evaluate potential causal associations between approximately 1,400 plasma metabolites and sciatica. Causal inference was performed using single nucleotide polymorphisms (SNPs) associated with metabolites as instrumental variables (IVs), with sciatica as the outcome variable. The MR analysis was based on the following three core assumptions: ① Relevance assumption: Instrumental variables exhibit significant associations with metabolite levels; ② Independence assumption: Instrumental variables are independent of potential confounding factors; (iii) Exclusivity assumption: The instrumental variable influences the outcome solely through the metabolite, with no alternative pathways.17 Only when this assumption holds do MR results possess causal interpretability. Therefore, this study combined multiple methods and sensitivity tests throughout the analysis to ensure the robustness and scientific validity of the conclusions. The research design flowchart is presented in Figure 1.
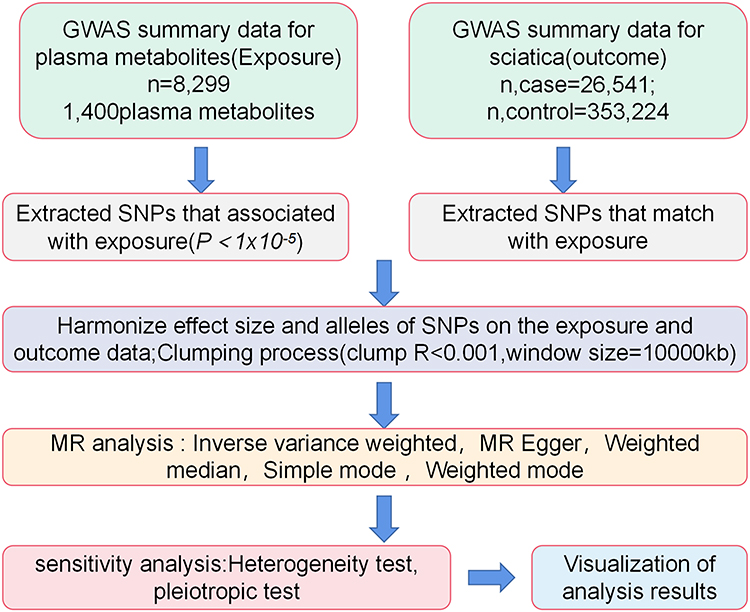 |
Figure 1 Research Design Flowchart. Abbreviations: GWAS, represents genome-wide association studies; MR, represents Mendelian randomization. |
Data Sources
The exposure data were obtained from a 2023 genome-wide association study on plasma metabolites published in Nature Genetics. This study included 8,299 individuals of European ancestry, identifying 1,091 independent metabolites and 309 metabolite ratios.18 All original summary statistics are publicly accessible through the GWAS Catalog website (https://www.ebi.ac.uk/gwas/downloads/summary-statistics), with accession numbers ranging from GCST90199621 to GCST90201020. The outcome data were sourced from the FinnGen research project Release 12 (https://storage.googleapis.com/finngen-public-data-r12/summary_stats/release/finngen_R12_M13_LOWBACKPAINORANDSCIATICA.gz), comprising 379,765 individuals of European ancestry.19 The specific phenotype used (FinnGen code: M13) is defined as “LOW BACK PAIN AND/OR SCIATICA”. Consequently, the case group (n = 26,541) comprises individuals diagnosed with “low back pain and/or sciatica”, while the control group consists of 353,224 individuals without this recorded diagnosis. Table 1 shows the comprehensive information of exposure data and outcome data. All data consisted of publicly available summary-level statistics, which had received ethics approval and informed consent in the original studies (Data summary is shown in Table 1). As this study did not involve individual-level raw data analysis, no additional ethics approval was required.
 |
Table 1 GWAS Data Summary |
Instrumental Variable Selection
To ensure the reliability and robustness of causal inference in this study, stringent quality control procedures were implemented during instrumental variable selection. First, the significance threshold for SNP screening was set at P < 1×10−5 to ensure significant and reliable associations between selected instrumental variables and plasma metabolites, while preventing insufficient instrumental variables due to overly stringent screening criteria.19 Regarding linkage disequilibrium (LD) processing, this study employed the “TwoSampleMR” package for clumping pruning, setting r2 < 0.001 with a 10,000 kb window. Referencing genotype data from the 1000 Genomes European population, we ensured independence among selected instrumental variables to prevent potential biases caused by linkage disequilibrium.20 Additionally, we utilized the F-statistic to assess instrumental variable strength, excluding weak instruments with F < 10 to prevent compromised analytical accuracy due to insufficient predictive power. The F-value was calculated using the formula: F = (N − K − 1) × R2 / [K × (1 − R2)], where N represents sample size, K denotes the number of SNPs, and R2 indicates the proportion of variance explained by SNPs in the exposure variable. Finally, to avoid pairing errors caused by inconsistent allele directions, we removed palindromic SNPs to ensure correct allele pairing for all instrumental variables, thereby further enhancing the analytical reliability.21
MR Analysis
All statistical analyses were performed using R software (version 4.4.1), primarily utilizing packages including TwoSampleMR and MendelianRandomization. To enhance the robustness of causal inference, this study comprehensively employed five MR methods: the inverse-variance weighted (IVW) method, MR-Egger regression, the weighted median method, the simple mode method, and the weighted mode method. Among these, IVW served as the primary analytical method, offering the highest statistical power and estimation accuracy when no significant horizontal pleiotropy existed among the instrumental variables.22,23 MR-Egger regression was employed to detect and adjust for horizontal pleiotropy, with an intercept term P-value > 0.05 indicating no significant bias affected the results;24 The weighted median method could yield consistent effect estimates even when ≤50% of instrumental variables were invalid;25 The mode-based method constructed causal models based on SNP clustering, providing supplementary validation for primary findings.26
To ensure result reliability, this study implemented a multi-method cross-validation strategy: causal associations between metabolites and sciatica were established when IVW, MR-Egger regression, and weighted median methods yielded consistent results with P < 0.05. Given the large number of exposure variables examined in this study, we further applied the False Discovery Rate (FDR) correction to control the false positive risk arising from multiple testing, setting the significance threshold at PFDR< 0.05. Additionally, the Steiger test was employed to verify the directionality of the causal relationship and exclude the possibility of reverse causality. Finally, the effect estimates are presented as odds ratios (ORs) with corresponding 95% confidence intervals (CIs), where an OR > 1 indicates that the metabolite increases the risk of sciatica, while an OR < 1 suggests a protective effect.
Sensitivity Analysis
Heterogeneity was assessed using Cochran’s Q test, where P > 0.05 indicated no significant heterogeneity.27 The MR-Egger intercept test was employed to detect pleiotropy, with P > 0.05 considered indicative of no significant bias.23 A leave-one-out analysis was conducted by sequentially removing individual SNPs; results were deemed robust if effect estimates exhibited no significant fluctuations.28
Results
MR Analysis Results
In this study, genetic data for plasma metabolites were stringently screened using thresholds of P < 1 × 10−5 and F > 10, ensuring selected SNPs showed significant associations with exposure variables. To ensure instrumental variable independence, we set r2 < 0.001 with a window size of 10,000 kb. Ultimately, 106 eligible SNPs were selected for subsequent causal relationship analysis. These SNPs satisfy the relevance, independence, and exclusion restriction assumptions in Mendelian randomization analysis, providing robust support for causal inference (Supplementary Tables 1 and 4).
Employing the FinnGen M13 phenotype, which captures a broad definition of sciatica alongside low back pain, as our outcome, we assessed the causal associations between plasma metabolites and sciatica risk. Among the 1,400 plasma metabolites analyzed, 4 metabolites demonstrated significant causal relationships with sciatica risk (P<0.05 and PFDR < 0.05). Specifically, elevated levels of Bilirubin degradation product, C17H20N2O5 (1) (OR=1.040, 95% CI=1.015–1.066, P=0.001, PFDR=0.003) and Bilirubin degradation product, C17H20N2O5 (2) (OR=1.039, 95% CI=1.014–1.065, P=0.002, PFDR=0.004) were associated with increased risk of sciatica. Conversely, N-Acetyl-β-alanine levels (OR=0.960, 95% CI=0.931–0.991, P=0.011, PFDR=0.014) and Glycosyl-N-stearoyl-sphingosine (d18:1/18:0) levels (OR=0.953, 95% CI=0.920–0.989, P=0.010, PFDR=0.012) demonstrated negative correlations with sciatica risk (as shown in Figures 2 and 3).
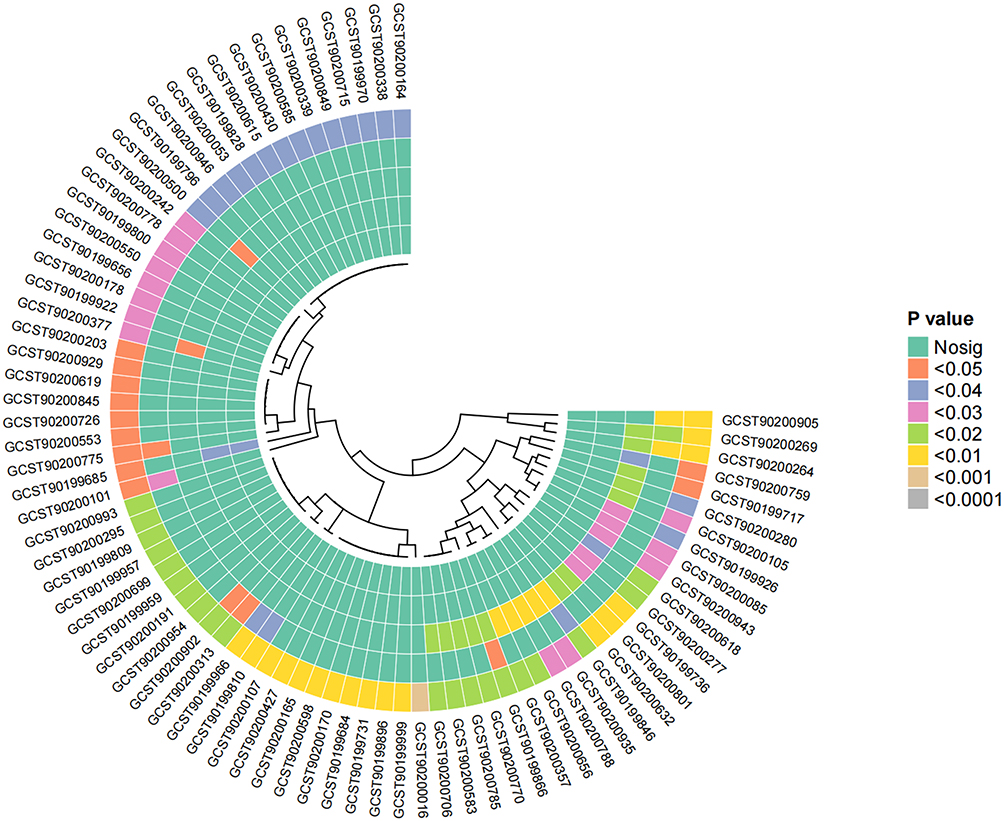 |
Figure 2 1,400 plasma metabolites and sciatica Mendelian randomization analysis results heat map. Notes: The heatmap displays the causal effect estimates from five MR methods, including IVW, MR-Egger, Weighted Median, Simple Mode, and Weighted Mode, for various genetic variants (SNPs). The P-values for each method are color-coded, with a color gradient indicating the statistical significance of the results. The following color scheme was used:No significance (Nosig): Represented in green, indicating P-values greater than 0.05.P < 0.05: Represented in yellow.P < 0.04: Represented in light orange.P < 0.03: Represented in dark orange.P < 0.02: Represented in light pink.P < 0.01: Represented in purple.P < 0.001: Represented in dark purple, highlighting highly significant results.Each row represents a genetic variant (SNP), while each column represents a different MR method. The clustering of SNPs based on their similarity in causal effect estimates across methods is visualized via hierarchical clustering, indicated by the dendrogram at the top of the heatmap. This clustering provides an overview of how each SNP’s estimated causal effect compares across different MR methods. |
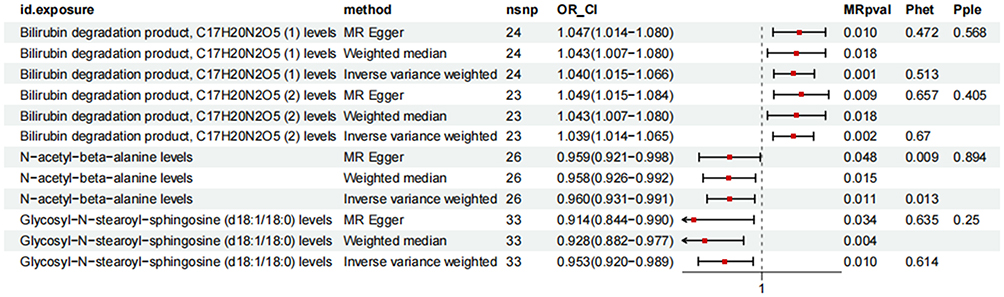 |
Figure 3 The overall forest map. Notes: This figure visually presents the overall causal relationship between exposures and outcomes, determining whether exposures exert significant causal effects on outcomes. If the confidence interval for the overall effect does not include the null line (represented as 1 in this figure), it indicates that the overall result is statistically significant. id.exposure denotes the exposure variable, method indicates the Mendelian randomization analysis approach, nsnp represents the number of instrumental variables, OR_CI indicates the 95% confidence interval, and MRpval denotes the p-value. |
Reliability Test
To verify the robustness of the model, multiple reliability tests were performed on the four metabolites exhibiting significant causal relationships with sciatica. First, Cochran’s Q test results showed that all Q values for the metabolites were greater than 0.05, indicating no significant heterogeneity among instrumental variables and demonstrating high consistency, as presented in Table 2 and Figure 4. Second, the intercept terms in MR-Egger regression were all close to zero with P-values exceeding 0.05, suggesting no significant horizontal pleiotropic bias was detected. This supports that the selected instrumental variables satisfy the independence and exclusion restriction assumptions. Further leave-one-out sensitivity analysis demonstrated that the causal effect estimates of each metabolite and their 95% confidence intervals remained stable after sequentially removing individual SNPs (Figure 5). The results were not dependent on any specific SNP, thereby enhancing the robustness of causal inference. Finally, Steiger directionality tests consistently showed Steiger_dir = TRUE, ruling out the possibility of reverse causality and further confirming the positive causal effect of metabolites on sciatica risk.
 |
Table 2 Heterogeneity Analysis and Horizontal Pleiotropy Analysis |
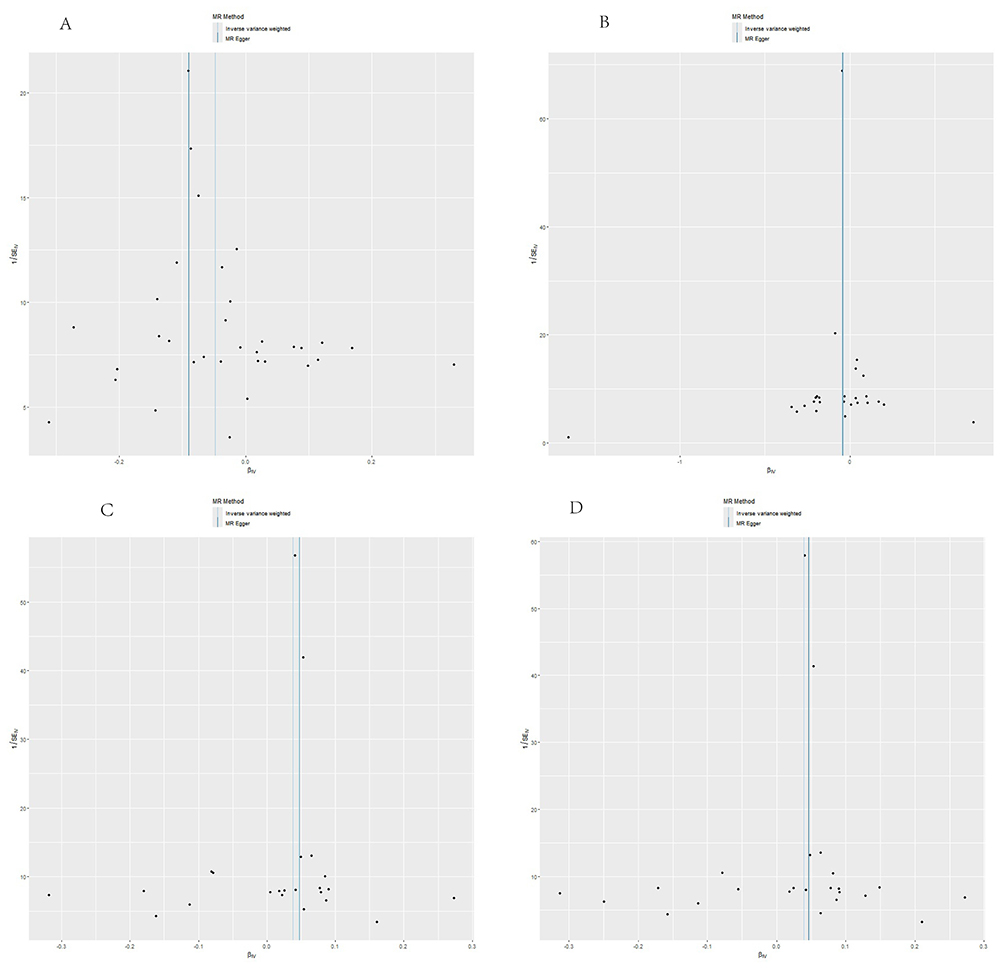 |
Figure 4 Funnel plot of heterogeneity analysis results. Notes: The funnel plot visualizes heterogeneity of single-nucleotide polymorphisms for four plasma metabolites causally associated with sciatica. This study used results from the inverse-variance weighted method as the primary reference standard. The distribution of single-nucleotide polymorphisms appears essentially symmetrical on both sides of the inverse-variance weighted reference line, indicating no significant bias. (A) shows the funnel plot for heterogeneity analysis between the level of bilirubin degradation product C17H20N2O5 (1) and Sciatica; (B) shows the funnel plot for heterogeneity analysis between the level of bilirubin degradation product C17H20N2O5 (2) and Sciatica; (C) shows the funnel plot for heterogeneity analysis between the level of N-Acetyl-β-alanine and Sciatica; (D) shows the funnel plot for heterogeneity analysis between Glycosyl-N-stearoyl-sphingosine and Sciatica. |
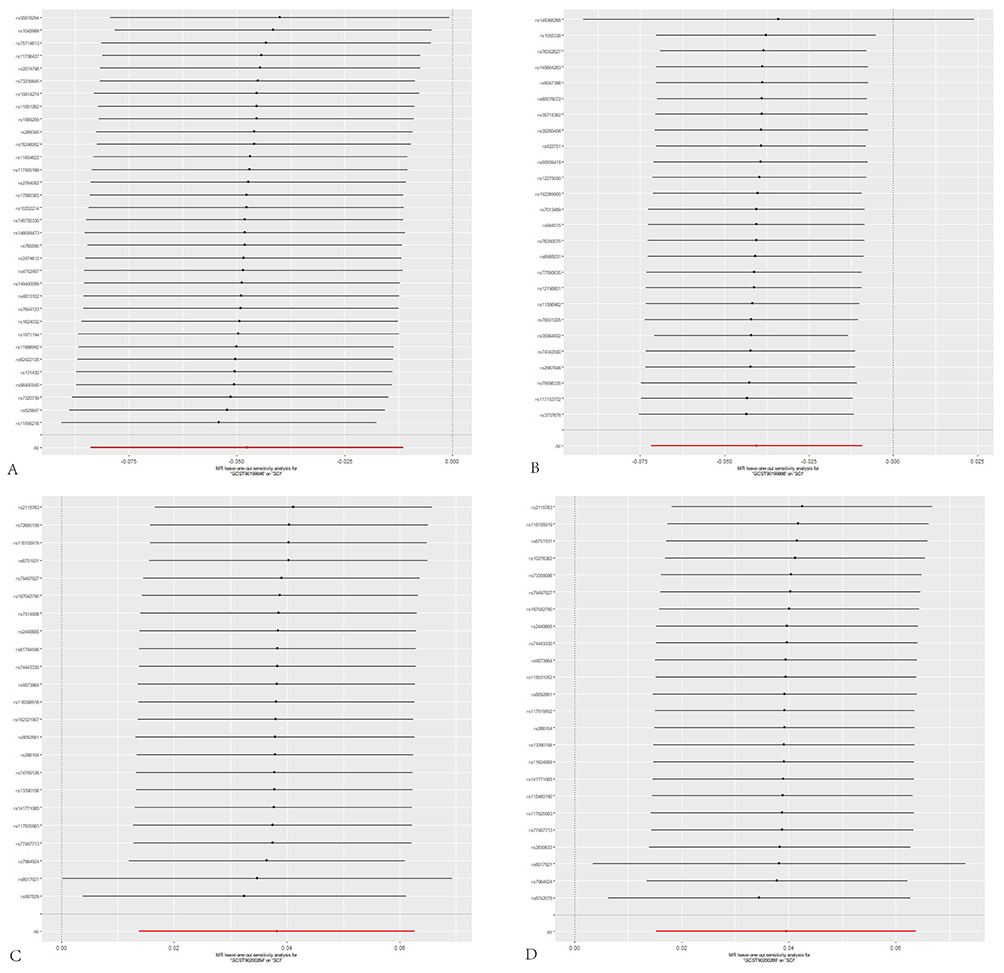 |
Figure 5 Leave-one-out test results forest plot. Notes: Primarily using the IVW method. The X-axis represents the IVW-specific values; black and red dots denote beta effect sizes, while line segments indicate the confidence intervals of the beta values. The Y-axis corresponds to each excluded rsid number and the IVW method without exclusions. (A) displays the leave-one-out sensitivity analysis for the association between bilirubin degradation product C17H20N2O5 (1) levels and sciatica; (B) shows the leave-one-out sensitivity analysis for bilirubin degradation product C17H20N2O5 (2) levels and sciatica; (C) presents the leave-one-out sensitivity analysis for N-Acetyl-β-alanine levels and sciatica; (D) represents the association between Glycosyl-N-stearoyl-sphingosine and Sciatica Leave-one-out sensitivity analysis plot. |
Discussion
This study systematically evaluated the causal relationships between 1,400 plasma metabolites and sciatica susceptibility using a two-sample MR approach. Our findings provide robust genetic evidence demonstrating that metabolic pathways play a significant role in driving sciatic nerve injury and associated pain. Specifically, bilirubin degradation product C17H20N2O5 (1) (OR=1.040, 95% CI=1.015–1.066, P=0.001, PFDR=0.003) and C17H20N2O5 (2) (OR=1.039, 95% CI=1.014–1.065, P=0.002, PFDR=0.004) were associated with increased risk of sciatica; Conversely, N-Acetyl-β-alanine (OR=0.960, 95% CI=0.931–0.991, P=0.011, PFDR=0.014) and Glycosyl-N-stearoyl-sphingosine (d18:1/18:0) (OR=0.953, 95% CI=0.920–0.989, P=0.010, PFDR=0.012) exhibited negative correlations. These four identified metabolites may serve as potential targets for future prevention or treatment of sciatica. More broadly, this study highlights the potential of genetic instrumental variable analysis to elucidate causal immune mechanisms underlying neuropathic pain.
Sciatica is a neuropathic pain caused by compression or irritation of the sciatic nerve. It typically presents with radiating pain, numbness, and motor dysfunction in the lower extremities. Its pathogenesis is complex and exhibits significant heterogeneity, involving multiple cellular processes including mitochondrial dysfunction, chemokine activation, and inflammatory factor release.29 In recent years, metabolomics has been progressively introduced into orthopedic disease research as a crucial approach for elucidating disease mechanisms and identifying potential biomarkers.30,31 A Mendelian randomization study has suggested causal associations between 28 serum metabolites and sciatica, indicating that some metabolites may play important roles in disease pathogenesis.16 However, their specific biological mechanisms require further elucidation. Moreover, studies have shown that modulating metabolite levels and the gut microbiota can alleviate pain associated with chronic nerve compression. The underlying mechanisms involve mitigating inflammatory responses, restoring microbial balance, and promoting nerve repair.32,33 Existing evidence indicates that gut-derived metabolites regulate neuropathic, inflammatory, and other chronic pain conditions.34,35 For instance, β-hydroxybutyrate, identified as a key plasma metabolite, correlates with reduced pain sensitivity and may alleviate inflammatory pain by enhancing expression.36 These studies provide a theoretical foundation for the marker potential and therapeutic value of metabolites in sciatica.
Our MR analysis identified bilirubin degradation products C17H20N2O5 (1) and (2) as potential risk factors for sciatica. This is paradoxical because bilirubin itself is a well-known endogenous molecule with antioxidant, anti-inflammatory, and neuroprotective effects.37 Under physiological conditions, bilirubin scavenges reactive oxygen species, inhibits pro-inflammatory cytokines, and promotes neuronal survival, helping maintain redox balance and neural homeostasis.38 Clinical observations further support its beneficial role, as higher serum bilirubin levels are inversely linked to the prevalence and severity of various inflammatory and metabolic diseases.39,40 The apparent discrepancy between the protective nature of bilirubin and the risk association of its downstream metabolites, C17H20N2O5 (1) and (2), necessitates a cautious and evidence-based interpretation.We propose several plausible explanations grounded in existing biological knowledge. First, the biological activity of a metabolic pathway is not solely defined by its initial substrate. Specific downstream degradation products can have distinct chemical properties and biological fates that differ from their parent compound.41 While bilirubin is protective, certain metabolites generated during its catabolism might participate in distinct and potentially harmful biological processes unrelated to its primary antioxidant function.42,43 Second, these metabolites may not be direct drivers of pathology but rather serve as biomarkers of an underlying pathological state. For instance, elevated levels of C17H20N2O5 could reflect a state of chronic, low-grade systemic inflammation or a broader metabolic dysfunction that also predisposes individuals to musculoskeletal and neuropathic pain conditions.44 This perspective is supported by a previous MR study linking the same metabolites to increased osteoporosis risk, suggesting a shared metabolic disturbance affecting both bone integrity and neural tissue, possibly through mechanisms like oxidative stress and neuro-immune-axis imbalance.45 Shared mechanisms such as oxidative stress and neuro-immune-axis imbalance could form a common pathophysiological ground. Third, the influence might be indirect. Bilirubin metabolite dysregulation could impact systems that secondarily affect sciatica risk, such as vascular health or the gut-brain axis, which is increasingly implicated in neuroinflammation and chronic pain.34,46,47 Critically, our MR estimates reflect lifelong genetic predisposition to higher circulating levels of these metabolites, not acute fluctuations. Therefore, the association likely points to a role in long-term susceptibility or in shaping a pro-inflammatory phenotype, rather than in triggering an acute pain episode. This aligns with the methodological principle that MR is better suited to identify factors influencing disease development rather than acute treatment targets. In summary, the causal link between bilirubin degradation products and sciatica risk revealed by our analysis highlights a complex and previously under-explored node in pain metabolism. It underscores that the functional output of a nominally protective pathway can be context-dependent and modified by downstream metabolism. Future research should prioritize elucidating the specific biological activities of C17H20N2O5 isomers, their roles in neuro-immune crosstalk (eg, through NF-κB or other inflammatory signaling pathways),38 and their interaction with systems like bone metabolism and the gut microbiome. Cellular and animal models of radiculopathy will be essential to distinguish whether these metabolites are active contributors to neuroinflammation, passive markers of systemic dysregulation, or involved in pre-conditioning the nervous system to injury.
Our analysis identified N-Acetyl-β-alanine as a metabolite inversely associated with sciatica risk. This compound is a downstream degradation product of carnosine (β-alanyl-L-histidine), a dipeptide that is highly concentrated in skeletal muscle. Carnosine functions as an intracellular pH buffer and antioxidant.48 In the context of chronic pain conditions like sciatica, reduced physical activity and possible muscle disuse could influence carnosine metabolism. The degradation of carnosine, potentially reflecting altered muscle physiology or a systemic acidic shift associated with chronic inflammation, releases β-alanine, which is subsequently acetylated to form N-Acetyl-β-alanine.49 Beyond its origin, N-Acetyl-β-alanine participates in central metabolic processes. It is synthesized through the acetylation of β-alanine, a reaction that consumes acetyl-CoA—a key molecule in energy and amide metabolism.50 Consequently, this links the metabolite to cellular energy homeostasis, amino acid detoxification, and neurotransmitter synthesis. Animal studies suggest a role in neuromodulation, with higher levels observed during active wakefulness compared to sleep, hinting at involvement in arousal and neurotransmitter dynamics.51 Furthermore, N-Acetyl-β-alanine may be metabolized to regenerate acetyl-CoA, feeding into the tricarboxylic acid (TCA) cycle to support mitochondrial energy production. Observations of its depletion in models of mitochondrial dysfunction, such as hepatic fibrosis, underscore its connection to cellular bioenergetics.52 Based on its metabolic origin and functions, we propose several mechanisms linking N-Acetyl-β-alanine to reduced sciatica risk. First, as a marker of carnosine turnover, its level may reflect muscle metabolic status or the body’s buffering capacity against inflammatory acidosis. Second, by contributing to acetyl-CoA pools and TCA cycle flux, it could support mitochondrial energy production in neurons or glial cells, helping to counter energy deficits linked to neuropathic pain. Third, it might influence neural excitability by modulating inhibitory neurotransmitter systems, such as GABAergic signaling;53 Finally, through its role in energy metabolism, it may indirectly reduce oxidative stress and pro-inflammatory pathways. In summary, the protective association of N-Acetyl-β-alanine with sciatica may arise from its dual significance: as a potential indicator of muscle-derived carnosine metabolism relevant to chronic pain pathophysiology, and as an active metabolite involved in maintaining neural energy homeostasis and modulating excitability. Future studies should directly measure carnosine and its degradation products in sciatica patients. Researchers should also use experimental models to determine whether N-Acetyl-β-alanine directly provides neuroprotection or mainly acts as a surrogate marker of related metabolic pathways.
Our MR analysis identified a protective association between higher levels of glycosyl-N-stearoyl-sphingosine (d18:1/18:0) and sciatica. Higher levels of this metabolite were associated with a reduced risk of developing the condition. As a glycosphingolipid, this metabolite is an essential component of cell membranes, particularly in neural tissues where sphingolipids contribute to myelin integrity, axonal function, and signal transduction.54,55 The potential mechanism underlying this association may involve the role of glycosphingolipids in peripheral nerve integrity and nociceptive signaling. Glycosphingolipids are critical for the formation and stability of myelin sheaths.56 Impaired myelination or demyelination can lead to increased neuronal excitability and aberrant pain signaling, which are hallmarks of neuropathic pain conditions such as sciatica.57 Moreover, sphingolipid metabolites, including ceramides and glycosphingolipids, participate in lipid raft-mediated signaling within nociceptive neurons and immune cells. These lipid microdomains facilitate the assembly and activation of receptors and signaling complexes involved in inflammatory pathways.58 For instance, glycosphingolipids can modulate NF-κB activation—a key pathway in neuroinflammation and radiculopathy—through their influence on membrane organization and cytokine receptor signaling.59,60 Additionally, sphingolipid signaling has been implicated in sensitization mechanisms within dorsal root ganglia and spinal cord circuits, potentially influencing pain chronicity.61 Alterations in sphingolipid metabolism may thus affect both structural integrity of nerves and functional pain pathways. While this study provides genetic evidence supporting a protective role for glycosyl-N-stearoyl-sphingosine in sciatica, the exact molecular mechanisms remain speculative. Future studies should directly examine its distribution in neural tissues, its role in myelin maintenance under compressive injury, and its interaction with inflammatory signaling in models of radicular pain. Furthermore, profiling sphingolipid species in patients with well-phenotyped sciatica could help validate its potential as a biomarker or therapeutic target.
Although the four identified metabolites come from different biochemical pathways, they may collectively affect sciatica risk through interconnected physiological systemsThe bilirubin degradation product C17H20N2O5 and N-Acetyl-β-alanine seem to have opposite roles in redox balance and energy homeostasis. While the former may indicate oxidative stress overflow, the latter supports mitochondrial function and metabolic resilience.62,63 Glycosyl-N-stearoyl-sphingosine, crucial for structural integrity, may be compromised by the same oxidative environments linked to bilirubin metabolite dysregulation.64,65 This suggests a model where sciatica susceptibility arises from an imbalance among neural energy needs, antioxidant capacity, and structural maintenance, especially during chronic inflammation or metabolic stress. Rather than acting independently, these metabolites likely form a network. Disturbances in one component, such as redox balance, may worsen vulnerabilities in others, including energy metabolism and myelin integrity. Together, these effects increase susceptibility to neuropathic pain.
This study still has certain limitations. First, the FinnGen M13 phenotype used as the outcome includes both sciatica and non-specific low back pain, which have different underlying pathophysiologies. This heterogeneity may dilute sciatica-specific metabolic signals. Therefore, the identified metabolite associations likely reflect broader mechanisms related to chronic spinal pain or generalized nociception, rather than specifically implicating neuroinflammatory pathways of radiculopathy. Second, since both exposure and outcome data were derived from European populations, the generalizability of these findings requires validation in other ethnic groups, particularly in East Asian cohorts. Third, although multiple sensitivity analyses and rigorous instrumental variable selection were employed to reduce bias, residual confounding or horizontal pleiotropy—especially within interconnected metabolic pathways—cannot be fully excluded. Although the sample size was relatively large, the number of significant associations was limited after strict FDR correction across approximately 1,400 metabolites. This indicates that the effects of individual metabolites on sciatica or broadly defined back pain may be modest. Therefore, larger GWAS samples with more precise phenotypic definitions—such as MRI-confirmed radicular pain—are needed to identify additional associations and enhance statistical power. Finally, this study used publicly available summary-level data. Although the observed causal links between metabolites and sciatica are statistically robust, they lack experimental validation of their biological roles. Future work should incorporate functional assays, animal models, and clinical biospecimens to verify the activity and mechanisms of these metabolites, especially since some have limited functional annotation in existing literature, which constrains interpretation of their physiological relevance. Notwithstanding these limitations, this study offers valuable insights relevant to Chinese populations. Against the backdrop of a growing burden of chronic and neuro‑musculoskeletal disorders such as sciatica in China’s primary care and rehabilitation systems, our findings on plasma metabolites may aid in early screening and tailored intervention.
Conclusion
In conclusion, this two-sample MR study identified four plasma metabolites—bilirubin degradation products C17H20N2O5 (1) and (2), N-Acetyl-β-alanine, and glycosyl-N-stearoyl-sphingosine (d18:1/18:0)—that exhibit significant causal associations with sciatica risk in European populations. The direction of these associations suggests distinct roles in disease etiology, with bilirubin derivatives potentially linked to risk-enhancing pathways, while N-Acetyl-β-alanine and glycosyl-N-stearoyl-sphingosine may be involved in protective mechanisms. These findings should be interpreted in context: the observed effect sizes are modest, and the identified metabolites are not intended to replace established clinical risk factors such as age, obesity, occupational load, or prior episode history, which remain paramount for individual risk assessment. Rather, this study provides genetic evidence that specific metabolic pathways may contribute to sciatica susceptibility independently of or in interaction with these traditional factors. It shifts the focus towards systemic metabolism as a component of sciatica pathophysiology. Future research should prioritize replicating these findings in independent and diverse cohorts, directly measuring these metabolites in patients, and elucidating their specific roles in neuroinflammatory and nociceptive processes through experimental models. Ultimately, this work highlights novel metabolic candidates for mechanistic investigation and lays a groundwork for future studies exploring their potential utility, in combination with clinical predictors, for understanding disease biology.
Data Sharing Statement
The original contributions presented in the study are included in the article/Supplementary materials, further inquiries can be directed to the corresponding author. The data that support the findings of this study are available from the corresponding author, Qiuyu Wei, upon reasonable request.
Ethics Statement
This study was a Mendelian randomization analysis based on publicly available summary-level data and did not involve any new human or animal experiments. All data used in this analysis were derived from publicly available Genome-Wide Association Study (GWAS) summary statistics. The original GWAS studies obtained ethical approval from their respective institutional review boards and informed consent from all participants. As this study involved secondary analysis of anonymized aggregated data, the Medical Ethics Committee of Liuzhou Hospital of Traditional Chinese Medicine (Liuzhou Zhuang Medical Hospital) reviewed and granted an exemption from formal ethics review for this study (Exemption Approval Date: January 3,2026).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from Zhongwei Traditional Chinese Medicine Development Research Center (No. ZW2023A003),Guangxi University of Traditional Chinese Medicine Joint Fund Project(No.2024LZ001).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Davis D, Taqi M, StatPearls VA. Treasure Island (FL); 2025.
2. Wei J, Chang S, Liu S, et al. Lysine-specific demethylase 1 in primary sensory neurons participates in chronic compression of dorsal root ganglion-induced neuropathic pain. Brain Res Bull. 2022;191:30–14. doi:10.1016/j.brainresbull.2022.10.010
3. Giménez-Campos MS, Pimenta-Fermisson-Ramos P, Díaz-Cambronero JI, Carbonell-Sanchís R, López-Briz E, Ruíz-García V. A systematic review and meta-analysis of the effectiveness and adverse events of gabapentin and pregabalin for sciatica pain. Aten Primaria. 2022;54(1):102144. doi:10.1016/j.aprim.2021.102144
4. Liu C, Ferreira GE, Abdel Shaheed C, et al. Surgical versus non-surgical treatment for sciatica: systematic review and meta-analysis of randomised controlled trials. BMJ. 2023;381:e070730. doi:10.1136/bmj-2022-070730
5. Gong G, Yan Z, Lai Q, et al. Inflammation preservation strategy: reconciling pain control and disc resorption in lumbar disc herniation. Front Immunol. 2025;16:1653681. doi:10.3389/fimmu.2025.1653681
6. Dove L, Jones G, Kelsey LA, Cairns MC, Schmid AB. How effective are physiotherapy interventions in treating people with sciatica? A systematic review and meta-analysis. Eur Spine J. 2023;32(2):517–533. doi:10.1007/s00586-022-07356-y
7. Pietzner M, Stewart ID, Raffler J, et al. Plasma metabolites to profile pathways in noncommunicable disease multimorbidity. Nat Med. 2021;27(3):471–479. doi:10.1038/s41591-021-01266-0
8. Wang W, Huang X, Zhang Y, et al. Transient compression injury triggers neuroinflammation in a new rat model of acute peripheral neuropathic pain. Pain Physician. 2024;27(1):E131–E145. doi:10.36076/ppj.2024.27.E131
9. Chen C. Inhibiting degradation of 2-arachidonoylglycerol as a therapeutic strategy for neurodegenerative diseases. Pharmacol Ther. 2023;244:108394. doi:10.1016/j.pharmthera.2023.108394
10. Fudong S, Zhiyi W, Jinyan Y, Yuzhang L, Zuoxu L, Shimin Z. Acupuncture mitigates sciatic neuropathic pain in lumbar disc herniation via inhibiting spinal CXCL12/CXCR4-driven glial activation and neuroinflammation. J Neuroimmunol. 2026;410:578797. doi:10.1016/j.jneuroim.2025.578797
11. Payam G, Yu G, Shruthi S, et al. 2-Deoxy-D-Glucose restores glial cell mitochondrial function and attenuates neuroinflammation. Sci Rep. 2025;15(1):38885. doi:10.1038/s41598-025-22677-w
12. Chang C, Wu C, Chern Y. Metabolic dysregulation in Huntington’s disease: neuronal and glial perspectives. Neurobiol Dis. 2024;201:106672. doi:10.1016/j.nbd.2024.106672
13. Bruno M, Bonomi CG, Ricci F, et al. Blood-brain barrier permeability is associated with different neuroinflammatory profiles in Alzheimer’s disease. Eur J Neurol. 2024;31(1):e16095. doi:10.1111/ene.16095
14. Yuan X, Yang P, Hu J, et al. Causal association between plasma metabolites and diverse autoimmune diseases: a two-sample bidirectional mendelian randomization study. Front Immunol. 2024;15:1437688. doi:10.3389/fimmu.2024.1437688
15. Di D, Tooki T, Zhou H, et al. Metal mixture and osteoporosis risk: insights from plasma metabolite profiling. Ecotoxicol Environ Saf. 2023;263:115256. doi:10.1016/j.ecoenv.2023.115256
16. Ren YM, Hou WY, Fan BY, et al. Causality of genetically determined serum metabolites on lower back pain or/and sciatica: a comprehensive Mendelian randomized study. Front Pain Res. 2024;5:1370704. doi:10.3389/fpain.2024.1370704
17. Skrivankova VW, Richmond RC, Woolf B, et al. Strengthening the reporting of observational studies in epidemiology using mendelian randomization: the STROBE-MR statement. JAMA. 2021;326(16):1614–1621. doi:10.1001/jama.2021.18236
18. Chen Y, Lu T, Pettersson-Kymmer U, et al. Genomic atlas of the plasma metabolome prioritizes metabolites implicated in human diseases. Nat Genet. 2023;55(1):44–53. doi:10.1038/s41588-022-01270-1
19. Zeng X, Zhong B. Investigating the causal relationship between human blood metabolites and pulmonary hypertension: a two-sample Mendelian randomization study. Front Cardiovasc Med. 2024;11:1304986. doi:10.3389/fcvm.2024.1304986
20. Machiela MJ, Chanock SJ. LDlink: a web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics. 2015;31(21):3555–3557. doi:10.1093/bioinformatics/btv402
21. Burgess S, Thompson SG. CRP CHD genetics collaboration. avoiding bias from weak instruments in mendelian randomization studies. Int J Epidemiol. 2011;40(3):755–764. doi:10.1093/ije/dyr036
22. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37(7):658–665. doi:10.1002/gepi.21758
23. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
24. Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44(2):512–525. doi:10.1093/ije/dyv080
25. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40(4):304–314. doi:10.1002/gepi.21965
26. Zhu Z, Zhang F, Hu H, et al. Integration of summary data from GWAS and eQTL studies predicts complex trait gene targets. Nat Genet. 2016;48(5):481–487. doi:10.1038/ng.3538
27. FD GM, Minelli C, Sheehan NA, Thompson JR. Detecting pleiotropy in Mendelian randomisation studies with summary data and a continuous outcome. Stat Med. 2015;34(21):2926–2940. doi:10.1002/sim.6522
28. Burgess S, Bowden J, Fall T, Ingelsson E, Thompson SG. Sensitivity analyses for robust causal inference from mendelian randomization analyses with multiple genetic variants. Epidemiology. 2017;28(1):30–42. doi:10.1097/EDE.0000000000000559
29. Nasir A, Afridi OK, Ullah S, Khan H, Bai Q. Mitigation of sciatica injury-induced neuropathic pain through active metabolites derived from medicinal plants. Pharmacol Res. 2024;200:107076. doi:10.1016/j.phrs.2024.107076
30. Gu Z, Wang Z, Zhang X, et al. Research advances in the study of traditional Chinese medicine formula granules on signaling pathway-mediated disease mechanisms. Front Pharmacol. 2025;16:1609211. doi:10.3389/fphar.2025.1609211
31. Song C, Liu F, Mei Y, et al. Integrated metagenomic and metabonomic mechanisms for the therapeutic effects of Duhuo Jisheng decoction on intervertebral disc degeneration. PLoS One. 2024;19(10):e0310014. doi:10.1371/journal.pone.0310014
32. Di Z, Zhenni C, Zifeng Z, et al. Danggui Sini Decoction normalizes the intestinal microbiota and serum metabolite levels to treat sciatica. Phytomedicine. 2024;132:155740. doi:10.1016/j.phymed.2024.155740
33. Zhang N, Guo P, Zhao Y, et al. Pharmacological mechanisms of puerarin in the treatment of Parkinson’s disease: an overview. Biomed Pharmacother. 2024;177:117101. doi:10.1016/j.biopha.2024.117101
34. Li JS, Su SL, Xu Z, et al. Potential roles of gut microbiota and microbial metabolites in chronic inflammatory pain and the mechanisms of therapy drugs. Ther Adv Chronic Dis. 2022;13:20406223221091177. doi:10.1177/20406223221091177
35. Magni G, Riboldi B, Ceruti S. Modulation of glial cell functions by the gut-brain axis: a role in neurodegenerative disorders and pain transmission. Cells. 2023;12(12):1612. doi:10.3390/cells12121612
36. Su M, Tang Y, Kong W, Zhang S, Zhu T. Genetically supported causality between gut microbiota, gut metabolites and low back pain: a two-sample Mendelian randomization study. Front Microbiol. 2023;14:1157451. doi:10.3389/fmicb.2023.1157451
37. Sheu A, White CP, Center JR. Bone metabolism in diabetes: a clinician’s guide to understanding the bone-glucose interplay. Diabetologia. 2024;67(8):1493–1506. doi:10.1007/s00125-024-06172-x
38. Zhang S, Chen T. To investigate the causal relationship between immune cell phenotype, blood metabolites, and myocarditis in a Mendelian randomization study. Medicine. 2025;104(23):e42713. doi:10.1097/MD.0000000000042713
39. Wu J, Su J, Wang Y, Chen J, Shang Y, Li J. Association between total bilirubin and bone mineral density level in adolescents. BMC Musculoskelet Disord. 2022;23(1):639. doi:10.1186/s12891-022-05592-3
40. Luo W, Li X, Zhou Y, Xu D, Qiao Y. Correlation between bone mineral density and type 2 diabetes mellitus in elderly men and postmenopausal women. Sci Rep. 2024;14(1):15078. doi:10.1038/s41598-024-65571-7
41. Voland RW, Wang H, Abruña HD, Lancaster KM. Nitrous oxide production via enzymatic nitroxyl from the nitrifying archaeon Nitrosopumilus maritimus. Proc Natl Acad Sci U S A. 2025;122(3):e2416971122. doi:10.1073/pnas.2416971122
42. Ryter SW, Tyrrell RM. The heme synthesis and degradation pathways: role in oxidant sensitivity. Heme oxygenase has both pro- and antioxidant properties. Free Radic Biol Med. 2000;28(2):289–309. doi:10.1016/S0891-5849(99)00223-3
43. Stevenson DK, Vreman HJ, Wong RJ, Contag CH. Carbon monoxide and bilirubin production in neonates. Semin Perinatol. 2001;25(2):85–93. doi:10.1053/sper.2001.23197
44. Aoi W, Tanimura Y. Roles of skeletal muscle-derived exosomes in organ metabolic and immunological communication. Front Endocrinol. 2021;12:697204. doi:10.3389/fendo.2021.697204
45. Gong L, Bai Z. Association of genetically predicted blood metabolites with osteopenia in individuals over 60 years of age: a Mendelian randomization study. Jt Dis Relat Surg. 2025;36(2):229–239. doi:10.52312/jdrs.2025.1991
46. Guo R, Chen L, Xing C, Liu T. Pain regulation by gut microbiota: molecular mechanisms and therapeutic potential. Br J Anaesth. 2019;123(5):637–654. doi:10.1016/j.bja.2019.07.026
47. Daulatzai MA. Chronic functional bowel syndrome enhances gut-brain axis dysfunction, neuroinflammation, cognitive impairment, and vulnerability to dementia. Neurochem Res. 2014;39(4):624–644. doi:10.1007/s11064-014-1266-6
48. Prakash MD, Fraser S, Boer JC, Plebanski M, de Courten B, Apostolopoulos V. Anti-cancer effects of carnosine-a dipeptide molecule. Molecules. 2021;26(6):1644. doi:10.3390/molecules26061644
49. Jukić I, Kolobarić N, Stupin A, et al. Carnosine, small but mighty-prospect of use as functional ingredient for functional food formulation. Antioxidants. 2021;10(7):1037. doi:10.3390/antiox10071037
50. Ji J, Zhu P, Blaženović I, et al. Explaining combinatorial effects of mycotoxins Deoxynivalenol and Zearalenone in mice with urinary metabolomic profiling. Sci Rep. 2018;8(1):3762. doi:10.1038/s41598-018-21555-y
51. Bourdon AK, Spano GM, Marshall W, et al. Metabolomic analysis of mouse prefrontal cortex reveals upregulated analytes during wakefulness compared to sleep. Sci Rep. 2018;8(1):11225. doi:10.1038/s41598-018-29511-6
52. Gao J, Qin XJ, Jiang H, et al. Detecting serum and urine metabolic profile changes of CCl(4)-liver fibrosis in rats at 12 weeks based on gas chromatography-mass spectrometry. Exp Ther Med. 2017;14(2):1496–1504. doi:10.3892/etm.2017.4668
53. Dong T, Wang X, Jia Z, Yang J, Liu Y. Assessing the associations of 1,400 blood metabolites with major depressive disorder: a Mendelian randomization study. Front Psychiatry. 2024;15:1391535. doi:10.3389/fpsyt.2024.1391535
54. Hanada K. Sphingolipids in infectious diseases. Jpn J Infect Dis. 2005;58(3):131–148. doi:10.7883/yoken.JJID.2005.131
55. Cantalupo A, Sasset L, Gargiulo A, et al. Endothelial sphingolipid de novo synthesis controls blood pressure by regulating signal transduction and no via ceramide. Hypertension. 2020;75(5):1279–1288. doi:10.1161/HYPERTENSIONAHA.119.14507
56. Belarbi K, Cuvelier E, Bonte MA, et al. Glycosphingolipids and neuroinflammation in Parkinson’s disease. Mol Neurodegener. 2020;15(1):59. doi:10.1186/s13024-020-00408-1
57. Yuchen W, Qiong S, Menachem H, et al. Demyelination-related pain: role of lysophosphatidic acid in satellite glial cell-neuron crosstalk. J Transl Med. 2025;23(1):1425. doi:10.1186/s12967-025-07568-y
58. Huang S, Abutaleb K, Mishra S. Glycosphingolipids in cardiovascular disease: insights from molecular mechanisms and heart failure models. Biomolecules. 2024;14(10):1265. doi:10.3390/biom14101265
59. Zhang T, de Waard AA, Wuhrer M, Spaapen RM. The role of glycosphingolipids in immune cell functions. Front Immunol. 2019;10:90. doi:10.3389/fimmu.2019.00090
60. Yokoyama N, Hanafusa K, Hotta T, Oshima E, Iwabuchi K, Nakayama H. Multiplicity of glycosphingolipid-enriched microdomain-driven immune signaling. Int J Mol Sci. 2021;22(17):9565. doi:10.3390/ijms22179565
61. Choi L, Vernon J, Kopach O, et al. The Fabry disease-associated lipid Lyso-Gb3 enhances voltage-gated calcium currents in sensory neurons and causes pain. Neurosci Lett. 2015;594:163–168. doi:10.1016/j.neulet.2015.01.084
62. Parfenova H, Leffler CW. Cerebroprotective functions of HO-2. Curr Pharm Des. 2008;14(5):443–453. doi:10.2174/138161208783597380
63. Ryter SW. Therapeutic potential of heme oxygenase-1 and carbon monoxide in acute organ injury, critical illness, and inflammatory disorders. Antioxidants. 2020;9(11):1153. doi:10.3390/antiox9111153
64. Liang W, Lu H, Wu B, et al. Bilirubin prevents hyperlipidaemic acute pancreatitis by inhibiting aryl hydrocarbon receptor-mediated acinar cell pyroptosis. Br J Pharmacol. 2025;182(23):5899–5912. doi:10.1111/bph.70168
65. Mohit MJ, Bajpai J, Verma S, Verma S. Disrupted bilirubin metabolism and gut microbiome dysregulation: a link to cardio-renal-hepatic-metabolic health in obstructive sleep apnea. Life Sci. 2025;379:123872. doi:10.1016/j.lfs.2025.123872
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Genetically Predicted Serum Albumin and Risk of Colorectal Cancer: A Bidirectional Mendelian Randomization Study
Lv L, Sun X, Liu B, Song J, Wu DJH, Gao Y, Li A, Hu X, Mao Y, Ye D
Clinical Epidemiology 2022, 14:771-778
Published Date: 21 June 2022
Associations Between Mean Platelet Volume and Risk of Deep Vein Thrombosis: A Mendelian Randomization Study and a Retrospective Study
Li J, Liang Y
International Journal of General Medicine 2023, 16:515-524
Published Date: 8 February 2023
Causal Relationship Between Endometriosis and Pelvic Inflammatory Diseases: Mendelian Randomization Study
Liu K, Liu X, Cao T, Cui X, Sun P, Zhang L, Wu X
International Journal of Women's Health 2024, 16:727-735
Published Date: 24 April 2024
Unveiling Novel Protein Biomarkers for Psoriasis Through Integrated Analysis of Human Plasma Proteomics and Mendelian Randomization
Mao R, Zhang T, Yang Z, Li J
Psoriasis: Targets and Therapy 2024, 14:179-193
Published Date: 7 December 2024
Assessing Causality Between Androgenetic Alopecia with Depression: A Bidirectional Mendelian Randomization Study
Li H, Cai H, Li P, Zeng Y, Zhang Y
Clinical, Cosmetic and Investigational Dermatology 2025, 18:445-451
Published Date: 22 February 2025