Back to Journals » Journal of Inflammation Research » Volume 18
Effects of Advanced Glycation End Products (AGEs) on Human Lung Macrophages: Implications for Pulmonary Inflammation
Authors Palestra F ![]() , Memoli G, Cristinziano L, Ferrara AL, Carucci L
, Memoli G, Cristinziano L, Ferrara AL, Carucci L ![]() , La Rocca A, Illiano A
, La Rocca A, Illiano A ![]() , Modestino L, Poto R
, Modestino L, Poto R ![]() , Galdiero MR, Varricchi G, Spadaro G, Berni Canani R, Marone G, Mercadante E, Loffredo S
, Galdiero MR, Varricchi G, Spadaro G, Berni Canani R, Marone G, Mercadante E, Loffredo S
Received 22 April 2025
Accepted for publication 6 August 2025
Published 5 November 2025 Volume 2025:18 Pages 15463—15477
DOI https://doi.org/10.2147/JIR.S531689
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qing Lin
Advanced Glycation End Products in Human Lung Macrophages – Video S2 [531689]
Views: 81
Francesco Palestra,1,2 Gina Memoli,1,2 Leonardo Cristinziano,2,3 Anne Lise Ferrara,1,2 Laura Carucci,1,4 Antonello La Rocca,5 Amalia Illiano,5 Luca Modestino,1,2,6 Remo Poto,1,2 Maria Rosaria Galdiero,1– 3,6 Gilda Varricchi,1– 3 Giuseppe Spadaro,1– 3,6 Roberto Berni Canani,1,4 Gianni Marone,1– 3 Edoardo Mercadante,5 Stefania Loffredo1– 3
1Department of Translational Medical Sciences, University of Naples “Federico II”, Naples, Italy; 2WAO Center of Excellence, Naples, 80131, Italy; 3Center for Basic and Clinical Immunology Research (CISI), University of Naples “Federico II”, Naples, Italy; 4ImmunoNutritionLab at CEINGE-Advanced Biotechnologies, University of Naples “Federico II”, Naples, Italy; 5Thoracic Surgery Unit - Istituto Nazionale Tumori IRCCS Fondazione G. Pascale, Naples, Italy; 6Department of Internal Medicine and Clinical Immunology, University Hospital of Naples Federico II, Naples, 80131, Italy
Correspondence: Stefania Loffredo, Email [email protected]
Introduction: Advanced glycation end products are a diverse, complex group of chemicals that are mostly produced by the Maillard reaction. The Maillard reaction happens when reducing sugar interacts non-enzymatically with amino acids found in proteins, lipids, or DNA. AGEs can be produced not only endogenously but also exogenously, as a result of certain cooking conditions or food processing. AGEs can interact with their receptor RAGE (receptor for AGE), expressed in various tissues including lung, inducing inflammatory effects and oxidative stress conditions that could damage target organs. Recent studies demonstrated that AGEs could be implicated in pathophysiology of chronic obstructive pulmonary disease (COPD) and idiopathic pulmonary fibrosis (IPF).
Objective: Considering that human lung macrophages (HLMs) are the most abundant immune cells in the lung, the aim of this project was to investigate the effects of AGEs on HLM activation and RAGE expression in these cells.
Results: AGEs induce, in a concentration-dependent manner, the release of cytokines (IL-6, IL-1β, TNF-α) and chemokines (CXCL1, CXCL2, CXCL8) from HLMs but not their de novo synthesis, except for IL-6. AGEs affect cellular kinetic proprieties and phagocytosis. HLMs constitutively express both sRAGE (soluble form of the receptor) and flRAGE (membrane-bound form of the receptor) mRNAs, and AGEs interaction slightly upregulate sRAGE expression. RAGE protein is constitutively expressed by about 60% of HLMs. Pre-incubation of HLMs with anti-RAGE antibody inhibits the release of cytokines and chemokines induced by AGEs. Finally, HLMs store and spontaneously release AGEs.
Conclusion: AGE accumulation promotes a pro-inflammatory phenotype in HLMs, potentially contributing to lung injury and respiratory dysfunction in AGEs-related diseases through the release of pro-inflammatory mediators. These findings highlight HLMs as a potential target in AGE-mediated pulmonary inflammation and suggest novel therapeutic strategies targeting the AGE-HLM axis.
Keywords: cytokines, chemokines, COPD, RAGE, lung injury
Introduction
Advanced glycation end products (AGEs) are mainly formed through a process called the Maillard reaction, which occurs when reducing sugars react non-enzymatically with amino acids in proteins, lipids, or DNA.1,2 This reaction occurs in three phases: in the first phase, glucose attaches to a free amino acid (such as lysine and arginine) to form a Schiff base; the Schiff base undergoes a chemical rearrangement to yield Amadori products, also known as early glycation products; if there is an accumulation of Amadori products, they undergo further chemical rearrangements, leading to the formation of crosslinked proteins, which represent AGEs.2 In addition to the Maillard reaction, other pathways can also lead to the formation of AGEs, such as glucose oxidation, lipid peroxidation, and the polyol pathway.2 Another source of AGEs are reactive dicarbonyl compounds, such as methylglyoxal, which contribute to AGE formation through non-enzymatic glycation reactions.3
Endogenous AGEs physiologically increase with aging and their accumulation can be boosted by smoking status and exposure to air pollution.4,5 In addition to endogenous production, AGEs can also be obtained from external sources: certain cooking conditions or food processing (high temperatures for an extended period of time, low hydration, and high pH), air pollution can enhance AGEs formation and accumulation.2,6 AGEs can interact with their receptor RAGE (receptor for advanced glycation end products), inducing inflammatory effects and oxidative stress conditions that could damage target organs.7 RAGE exists in two forms: a membrane-bound form (flRAGE), and a soluble form (sRAGE).8 RAGE is expressed in various tissues including lung9,10 and its signalling has also been implicated in pulmonary inflammation triggered by infectious stimuli, such as LPS, and sepsis. In these models, both genetic deletion and pharmacological inhibition of RAGE significantly reduce lung tissue injury.11,12
Both the intake and endogenous production of AGEs have been associated with various diseases due to their role in promoting oxidative stress, inflammation, and tissue damage.2,9 Although the historical focus of AGEs toxicity has been on diabetes mellitus and cardiovascular diseases (such as atherosclerosis),13 recent studies demonstrated that AGEs could be implicated in pathophysiology of allergic14 and respiratory diseases.15,16 In particular, AGE levels have been observed to rise in COPD patients independently of factors such as smoking, diabetes mellitus, or cardiovascular diseases.16 Furthermore, AGEs accumulation seems to be associated with impaired respiratory function in healthy individuals (lower forced expiratory volume, forced vital capacity, and diffusing capacity for carbon monoxide values).15
Macrophages are the most representative immune cell type in lung tissue and play a crucial role in inflammatory and phagocytic processes by releasing cytokines (eg, IL-6, IL-1β, TNF-α, IL-12), chemokines (eg, CXCL8), and enzymes.17–20 Recent evidences demonstrate that AGEs activate murine macrophages inducing pro-inflammatory mediator release and enhance their polarization into M1 phenotype.21,22 At the same time, AGEs inhibit anti-inflammatory factor release and efferocytosis in murine macrophages.21 To date, there are no data on the effects of the AGEs on human lung macrophages (HLMs). Considering the crucial role of HLMs in lung homeostasis and dysfunction, and that AGEs accumulation could be associated with lung injury and respiratory disease development, the aim of this project was to investigate if AGEs could influence the activation of HLMs and the expression of their receptor, RAGE.
Materials and Methods
Reagents
All reagents were obtained from commercial suppliers. L-glutamine, fetal bovine serum (FBS), lipopolysaccharide (LPS) from Escherichia coli serotype 026:B6, Percoll®, piperazine-N, N-bis-2-ethanesulphonic acid (PIPES), phosphate-buffered saline (PBS), Triton X-100, and an antibiotic-antimycotic solution (10,000 IU/mL penicillin, 10 mg/mL streptomycin and 25 µg/mL amphotericin B) were purchased from Lonza. RPMI 1640 was acquired from Microgem, and dimethyl sulfoxide (DMSO) from Merck Millipore. Accutase was purchased from StemCell Technologies. Primers targeting sRAGE, flRAGE, CXCL1, CXCL2, CXCL8, IL-1β, IL-6, TNF-α, and GAPDH were designed with Beacon Designer 3.0 software and synthesized and purified by Life Technologies.
Isolation and Purification of Human Lung Macrophages (HLMs)
HLMs were obtained from resected lung tissue of 37 patients undergoing surgery. These patients had tested negative for hepatitis C virus, hepatitis B surface antigen, and HIV. The study protocol received ethical approval from the Comitato Etico Territoriale Campania 1 (Protocol 36/23 OSS), and all participants provided informed consent. The donor cohort had a mean age of 62 years (range: 40–83 years) and consisted of approximately 41% females and 59% males. The cohort included a mix of smokers and non-smokers (54% smokers, 46% non-smokers). Lung tissue underwent mechanical processing and was rinsed with PIPES buffer using a 120 µm pore size Nitex cloth to create a cell suspension. Macrophages were enriched to 75–85% purity via discontinuous Percoll® density gradient centrifugation. The enriched cells were cultured in RPMI 1640 supplemented with 5% FBS, 2 mM L-glutamine, 1% antibiotic-antimycotic solution, and 1% non-essential amino acids. The cells were then cultured at 37°C in 24-well plates. After 16 hours, non-adherent cells were removed by washing, resulting in an adherent population of >98% macrophages confirmed through flow cytometry (as previously described).18 This method also effectively excludes dendritic cell contamination, as confirmed by the absence of CD169-/CD123+ cells.18
AGE Stimulation and RAGE Inhibition in HLMs
AGEs (Abcam, Cambridge, UK) were reconstituted and used as recommended. AGEs were diluted in complete medium before each experiment. HLMs were cultured in 24-well plates (0.15–2 × 106 cells/well) in complete medium and then stimulated (1 to 16 hours) with increasing concentrations of AGEs (1–100 µg/mL) or with LPS (1 µg/mL). For RAGE inhibition experiments, HLMs were pre-incubated with an RAGE neutralizing antibody (Abcam, Cam, UK) at a concentration of 4 µg/mL prior to stimulation with AGEs, as described previously.14 At the end of incubations, supernatants were harvested, centrifuged (500 × g, 4°C, 10 min), and stored at −80°C for subsequent determination of mediator release. Lysis of the remaining cells in the plates was carried out using 0.1% Triton X-100 for total protein quantification by a Bradford assay (Bio-Rad Laboratories, Segrate MI, Italy).
Cell Viability
After treatments, cell viability was evaluated as mitochondrial activity, determined by the MTT (3-(4,5-dimethylthiazol-2-yl)2,5-diphenyl tetrazolium bromide) assay, as reported previously.23 HLMs were incubated with AGEs, 1% v/v Triton X-100 (positive control for cell death), or medium alone (negative control) for the specific times. At the end of the incubation, supernatants were removed, and cells were incubated with MTT solution (0.5 mg/mL) at 37°C for 1 hour. The cells were washed with PBS, 0.5 mL of DMSO was added to solubilize the formazan crystals, and absorbance was measured at 540 nm. Cell viability was calculated as a percentage of the negative control, which was set to 100%. Figure S1A represents the percentage of viable cells relative to the untreated control.
ELISA
Concentrations of CXCL1, CXCL2, CXCL8, IL-6, TNF-α, and IL-1β in HLM supernatants were quantified in duplicate using commercially available ELISA kits (R&D Systems, Minneapolis, MN, USA). To account for variations in cell number between wells and experiments, cytokine concentrations were normalized to total protein content, determined by Bradford assay, in the corresponding cell lysates (0.1% Triton X-100). Results are expressed as ng cytokine/mg total protein.
RT-PCR
Expression of sRAGE, flRAGE, CXCL1, CXCL2, CXCL8, IL-6, TNF-α, and IL-1β mRNA was investigated. HLMs were stimulated with AGEs (100 µg/mL) or LPS (1 µg/mL). Following stimulation, total RNA was extracted using TRIzol® reagent (Euroclone, Milan, Italy). RNA quality and integrity were assessed using a Nanodrop spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). Reverse transcription was performed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). Real-time RT-PCR was conducted with SYBR Green Supermix (Bio-Rad) on a CFX96 real-time detection system (Bio-Rad Laboratories, Segrate, MI, Italy). GAPDH served as the housekeeping gene for normalization using the 2−ΔCt formula. PCR efficiency and specificity were evaluated by analyzing amplification curves with serial dilutions of the template cDNA and their dissociation curves. Each cDNA sample was analyzed in triplicate, and the corresponding no-RT mRNA sample was included as a negative control. The data were analyzed with iCycleriQ analysis software (Bio-Rad Laboratories, Segrate, MI, Italy), and the changes in CXCL1, CXCL2, CXCL8, IL-1β, IL-6, TNF-α, sRAGE, and flRAGE mRNAs were expressed as 2−ΔCt.
Flow Cytometric Analysis
HLMs fresh or cultured 16 hours with or without AGEs (100 µg/mL) and detached using Accutase, were washed with PBS and then stained with fluorophore-conjugated antibodies: PE-conjugated anti-CD11c (Miltenyi Biotec, Bologna Italy) and Alexa Fluor 647 anti-RAGE antibody (EPR21171 clone, Abcam, Mi Italy) for 30 minutes at 4°C in the dark. After staining, cells were washed and then fixed with 4% paraformaldehyde (PFA) and analyzed using the MACS Quant Analyzer 10 (Miltenyi Biotec, Bergisch Gladbach, Germany) and the FlowJo software, v.10. Doublets and debris were identified based on forward- and side-scatter properties and excluded from the analysis. Data were expressed as percentage of RAGE positive fresh HLMs (Figure 6D) Complete example of the gating strategy is represented in Supplementary Figure 2.
Time-Lapse and High-Content Microscopy
To assess the impact of AGEs on macrophage motility, time-lapse and high-content microscopy experiments were conducted using the Operetta High-Content Imaging System (PerkinElmer, MA, USA), as previously described.24,25 HLMs were cultured in Falcon® 24-well Clear Flat Bottom plates. For time-lapse experiments, HLMs were stimulated with AGEs (100 µg/mL) and monitored for 12 hours. Within this time window, digital phase contrast images of 15 fields/well were captured every 10 min via a 10× objective, allowing for detailed tracking of individual cell movements. To quantify cell tracking features, bright-field snapshots were taken at 6 fields/well. PhenoLOGIC (PerkinElmer) software was employed to analyse the following proprieties: current displacement and Y (measure the change in position along the x- and y-axes, respectively), displacement X and Y mean per well (average the displacement across multiple cells in a well along the x- and y-axes, respectively), current square displacement (reflects the overall area covered by a cell over time), displacement mean per well (average the square displacement across multiple cells), and current speed (quantifies the rate of cell movement) and turning angle (measures the change in direction of cell movement). These parameters collectively provide a comprehensive assessment of macrophage motility, including speed, directionality, and exploratory behaviour, allowing for a detailed analysis of the effects of AGEs on these crucial aspects of macrophage function.
Phagocytosis Assay
HLMs (1 × 105 cells/well) were seeded in imaging-compatible plates and allowed to adhere for 1 day at 37°C. Cells were then stimulated with AGEs (100 µg/mL) for 24 hrs. Following stimulation, the medium was replaced with pHrodo™ Green E. coli BioParticles™ Conjugate suspension (Chow et al, 2004). Phagocytosis was monitored by measuring fluorescence intensity (excitation 509 nm, emission 533 nm) over 3 hours, with readings taken every hour, using an EnSpire Multimode Plate Reader (PerkinElmer). Results are reported as relative fluorescence units (RFU). Since pHrodo™ Green fluorescence increases in acidic environments, such as phagosomes, it enables quantification of internalized particles in the assay.
Determination of AGEs
Two sets of human lung macrophages were used for AGE quantification. The first set consisted of freshly isolated HLMs. Immediately following Percoll® gradient isolation and washing, 5 × 106 of HLMs were lysed in 500 µL of 0.1% Triton X-100. The second set of HLMs (1 × 106 cells/well) were cultured in 24-well plates containing complete medium. After 16 hours incubation (37°C), the supernatants were collected, and the adherent cells were lysed in 500 µL of 0.1% Triton X-100. AGE concentrations in both the supernatants and cell lysates were measured using a commercially available ELISA kit for AGEs (MyBioSource, SD, USA).
Statistical Analysis
Statistical analysis was performed using Prism 8 software. Data are presented as mean ± standard deviation of the indicated number of experiments. Statistical significance was assessed using Student’s t-test or one-way ANOVA, followed by Dunnett’s test for comparisons against a control group or Bonferroni’s test for multiple pairwise comparisons. A p-value ≤0.05 was considered statistically significant. Due to technical constraints in isolating resident lung cells, the number of donors per endpoint varies. To ensure adequate statistical power, a post-hoc power analysis was conducted for each endpoint based on observed paired differences (mean and SD of differences). The power for all endpoints was above 80%, indicating sufficient statistical robustness.
Results
Effects of AGEs on Cytokine and Chemokine Release from HLMs
In a series of six different experiments, we evaluated the effects of increasing concentrations of AGEs (1–100 µg/mL) on HLM activation. AGEs activated HLMs inducing chemokine release including CXCL1 (Figure 1A), CXCL2 (Figure 1B) and CXCL8 (Figure 1C). Moreover, AGEs affect also the cytokine production; in particular, they induce IL-1β (Figure 1D), IL-6 (Figure 1E) and TNF-α (Figure 1F) release. The effect of AGEs was predominantly significant at 30 and 100 µg/mL. In these experiments, we used LPS, the main component in the cell wall of Gram-negative bacteria, as a positive control.19,26 LPS was a potent stimulus for the release of all tested (Figure 1). The percentage of viable HLMs measured by MTT assay 16 hours after AGEs treatment did not differ from that of untreated cells (Supplementary Figure 1A).
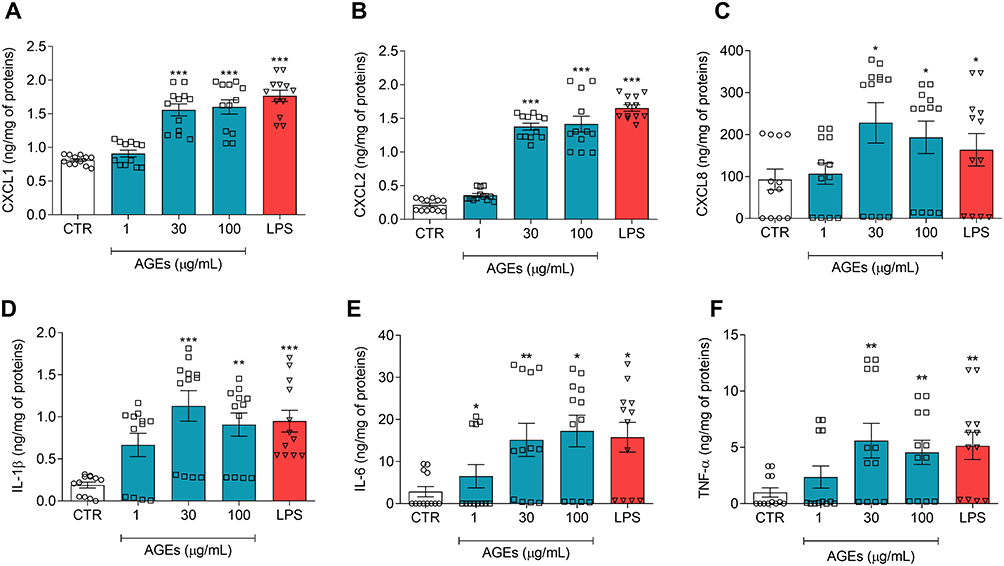 |
Figure 1 Effects of AGEs on chemokine and cytokine release from HLMs. HLMs (106 cells/well) were incubated (16 hours, 37°C, 5% CO2) with complete medium (CTR), or increasing concentrations of AGEs (1–100 μg/mL) or LPS (1 μg/mL). CXCL1 (A), CXCL2 (B), CXCL8 (C), IL-1β (D), IL-6 (E), and TNF-α (F) proteins in supernatants were evaluated by ELISA and expressed as ng or pg of mediators for mg of total proteins. Data are shown as individual values with mean ± SEM of 12 independent experiments obtained from different donors. * p< 0.05; ** p < 0.01; *** p < 0.001 vs CTR. |
To exclude the possibility that the activating effects of AGEs could be due to minor LPS contamination, HLMs were stimulated with AGEs or in the presence of polymyxin B (50 µg/mL), a potent inactivator of LPS.27 Supplementary Figure 1B shows that polymyxin B did not modify the spontaneous release of IL-6, a selected cytokine. Incubation of AGEs with polymyxin B did not modify the activating property of AGEs on the release of IL-6 from HLMs. As a control, polymyxin B significantly reduced the release of cytokines/chemokines induced by LPS (Supplementary Figure 1B).
Kinetics of Cytokine and Chemokine Release Induced by AGEs from HLMs
In the next group of experiments, we examined the kinetics of cytokine and chemokine release from HLMs stimulated with the highest tested concentration of AGEs (100 µg/mL). As shown in Figure 2, CXCL1, CXCL2, TNF-α, and IL-1β release peaked at 3 hours and then decreased by 16 hours. Conversely, CXCL8, IL-6, production markedly began after 1 h, and progressively increased after 16 hours of incubation. Only and IL-1β release was significant after 3 h of AGE stimulation (Figure 2). TNF-α and IL-1β were the only cytokines whose release was significant after just 1 hour of AGE stimulation.
 |
Figure 2 Kinetic of cytokine/chemokine release induced by AGEs from HLMs measured as fold increase vs control (CTR) respectively at all times. HLMs (106 cells/well) were stimulated (from 1 to 16 hours, 37°C, 5% CO2) with RPMI alone or with AGEs (100 µg/mL). CXCL1, CXCL2, CXCL8, IL-1β, IL-6, and TNF-α protein in supernatants were evaluated by ELISA. Data are the mean ± SEM of 7 independent experiment obtained from different donors. * p< 0.05; ** p < 0.01, and Δ p < 0.001 vs respective time CTR. |
The kinetic curve of IL-6 led us to hypothesize that the release of this cytokine was due both to a release of pre-formed mediator and to a new synthesis. Therefore, we also evaluated the effects of AGEs on CXCL1, CXCL2, CXCL8, IL-1β, IL-6, and TNF-α mRNA expression in HLMs by real-time quantitative PCR. The stimulation of HLMs with AGEs for 6 hours induced the mRNA expression for IL-6 (Figure 3E), but not for CXCL1 (Figure 3A), CXCL2 (Figure 3B), CXCL8 (Figure 3C), IL-1β (Figure 3D), and TNF-α (Figure 3F). These results indicate that AGEs induced preformed CXCL1, CXCL2, CXCL8, IL-1β, and TNF-α from HLMs but not their de novo synthesis, except for IL-6.
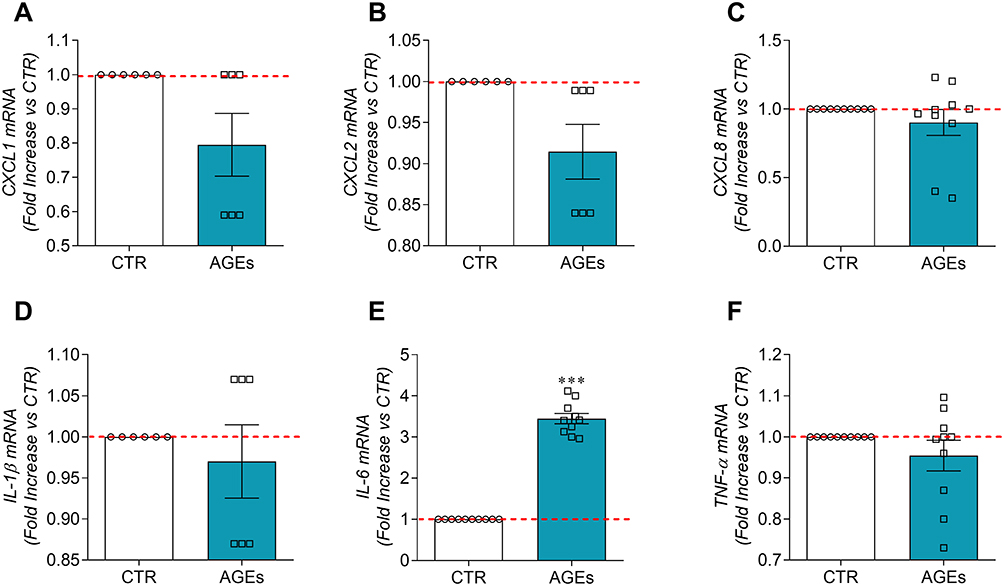 |
Figure 3 Effects of AGEs on mRNA expression for cytokines/chemokines in HLMs. Highly purified HLMs (6 × 106 cells/well) were incubated (6 hours, 37°C, 5% CO2) in the absence (CTR) or in the presence of AGEs (100 µg/mL). At the end of incubation, CXCL1 (A), CXCL2 (B), CXCL8 (C), IL-1β (D), IL-6 (E), and TNF-α (F) mRNAs were determined by quantitative RT-PCR. Data are shown as individual values with mean ± SEM of 10 independent experiments obtained from different donors. *** p< 0.001 vs CTR. |
Expression of RAGE Receptor on HLMs
Receptor for advanced glycation end products (RAGE) is a multi-ligand receptor that is able to bind a variety of ligands, including advanced glycation end products.28 RAGE is expressed on monocyte and macrophages,29 but its expression has not been demonstrated on HLMs. Therefore, we evaluated the constitutive mRNA expression of sRAGE and flRAGE in fresh HLMs by real-time quantitative PCR. HLMs constitutively expressed flRAGE and sRAGE mRNAs (Figure 4A). sRAGE was significantly more expressed than flRAGE (Figure 4A).
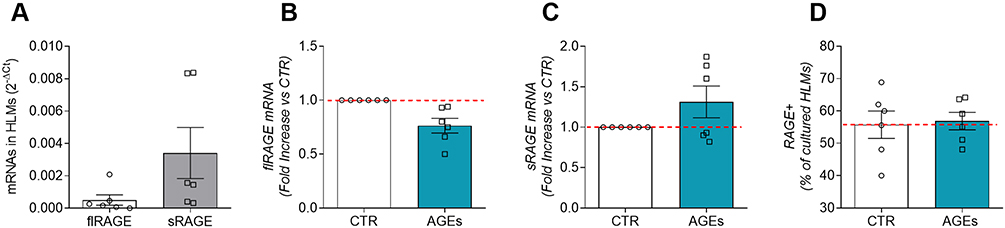 |
Figure 4 Expression of full length (flRAGE) and soluble form (sRAGE) of RAGE in HLMs. Fresh HLMs (6 × 106 cells) were lysed and RNA was extracted. mRNAs constitutive expression for flRAGE and sRAGE were determined by quantitative RT-PCR (A). HLMs (6 × 106 cells/well) were incubated (6 hours, 37°C) with complete media (CTR), or AGEs (100μg/mL). At the end of incubation, HLMs were lysed and RNA was extracted. mRNA expression for flRAGE (B) and sRAGE (C) was evaluated by quantitative RT-PCR. Unstimulated and AGE-stimulated (100 µg/mL) HLMs (D) were also stained with a specific anti-RAGE antibody (EPR21171, Abcam) and analyzed by flow cytometry. Data were expressed as percentage of RAGE positive fresh HLMs (D) and cultured HLMs. The red dotted line for each panel represents the control values. Data are shown as individual values with mean ± SEM of 6 independent experiments obtained from different donors. |
Then, we investigated the effect of AGEs (100 µg/mL) on flRAGE and sRAGE mRNA expression in HLMs. Figure 4 shows that AGEs did not affect mRNA expression for flRAGE (panel B), while slightly upregulate sRAGE mRNA (panel C).
Flow cytometry analysis was assessed to determine the percentage of RAGE-positive HLMs under both basal conditions and under stimulation with AGEs (100 µg/mL). Approximately 60% of fresh HLMs express RAGE (data not shown). Moreover, cultured (16 hours, 37°C, 5% CO2) HLMs were RAGE-positive under basal conditions (Figure 4D) and their stimulation with AGEs did not alter the percentage of RAGE-positive cells (Figure 4D).
Given the constitutive expression of RAGE on HLMs, we next sought to determine whether AGE-induced cytokine release was RAGE-dependent. To investigate this, HLMs were pre-incubated with an anti-RAGE antibody (4µg/mL) prior to stimulation with AGEs, as previously described.14 We observed that pre-incubation with anti-RAGE antibody significantly reduced the release of key pro-inflammatory cytokines from HLMs stimulated with AGEs (Figure 5). By contrast, pre-incubation with anti-RAGE antibody did not affect LPS-induced cytokine release. These results suggest that AGE-induced cytokine release in HLMs is, at least in part, mediated through RAGE signaling.
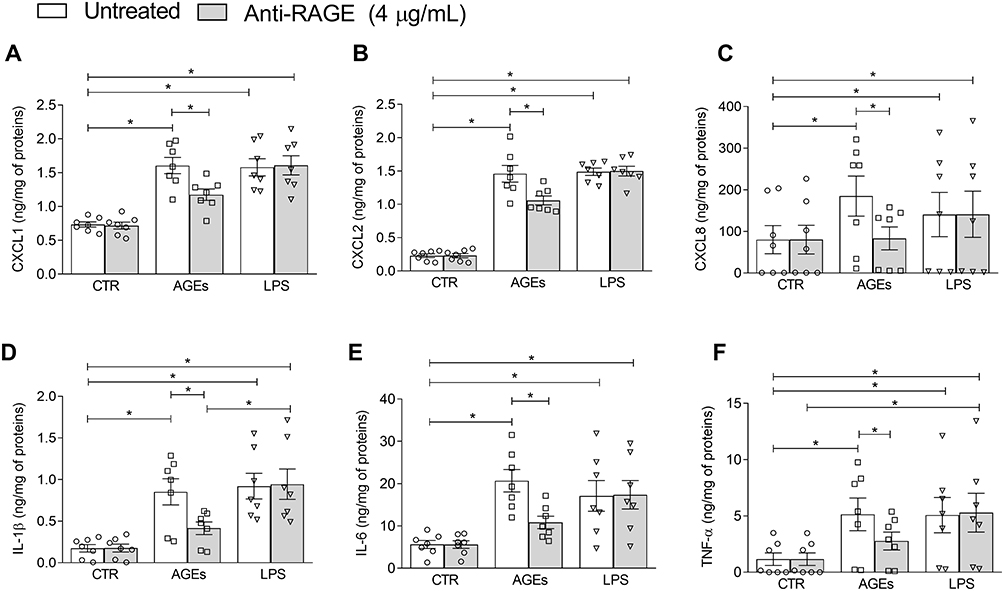 |
Figure 5 Effects of anti-RAGE antibody on AGEs-induced chemokine and cytokine release from HLMs. HLMs (106 cells/well) were pre-incubated (1 hour, 37°C, 5% CO2) with an anti-RAGE blocking antibody (4 μg/mL), followed by stimulation with AGEs (100 μg/mL) or LPS (1 μg/mL) for 16 hours. CXCL1 (A), CXCL2 (B), CXCL8 (C), IL-1β (D), IL-6 (E), and TNF-α (F) levels were measured in supernatants by ELISA and expressed as ng of mediator per mg of total protein. Data are shown as individual values with mean ± SEM of 7 independent experiments obtained from different donors. * p < 0.05 vs all other groups. |
Effects of AGEs on Kinetic Properties of HLMs
Different stimuli can change cell morphology as well as their movement.30 Macrophages exhibit migratory and adaptive behaviour within the tissue in response to various physiological and pathological conditions.31 In this context, we investigated HLM tracking induced by AGEs with an Operetta High-Content Imaging System (PerkinElmer). To this end, HLMs were incubated with AGEs (100 µg/mL) or complete medium alone for 12 hours at 37°C and 5% of CO2. AGEs significantly altered HLM displacement along the X-axis (Figure 6A and B), indicating a directional bias in their movement. This suggests that AGEs may influence the directional migration of HLMs, potentially disrupting their normal chemotactic responses or interactions with the extracellular matrix. No significant change in displacement was observed along the Y-axis (Figure 6C and D), suggesting that the effect of AGEs on HLM movement is directionally specific. Moreover, AGEs subtly modulated HLM motility by slightly altering turning angle (Figure 6E) and reduced area movement (Figure 6F) indicating a potential restriction in their overall surveillance capacity. This finding aligns with the observed directional bias in displacement and suggests that AGEs may limit the ability of HLMs to effectively patrol and respond to stimuli within the lung tissue. Finally, AGEs had no effect on HLM speed (Figure 6G and H). A representative visual aid of macrophage motility is provided in Supplementary Videos 1 (CTR) and 2 (AGEs-stimulated), which illustrates the differences in movement dynamics between conditions.
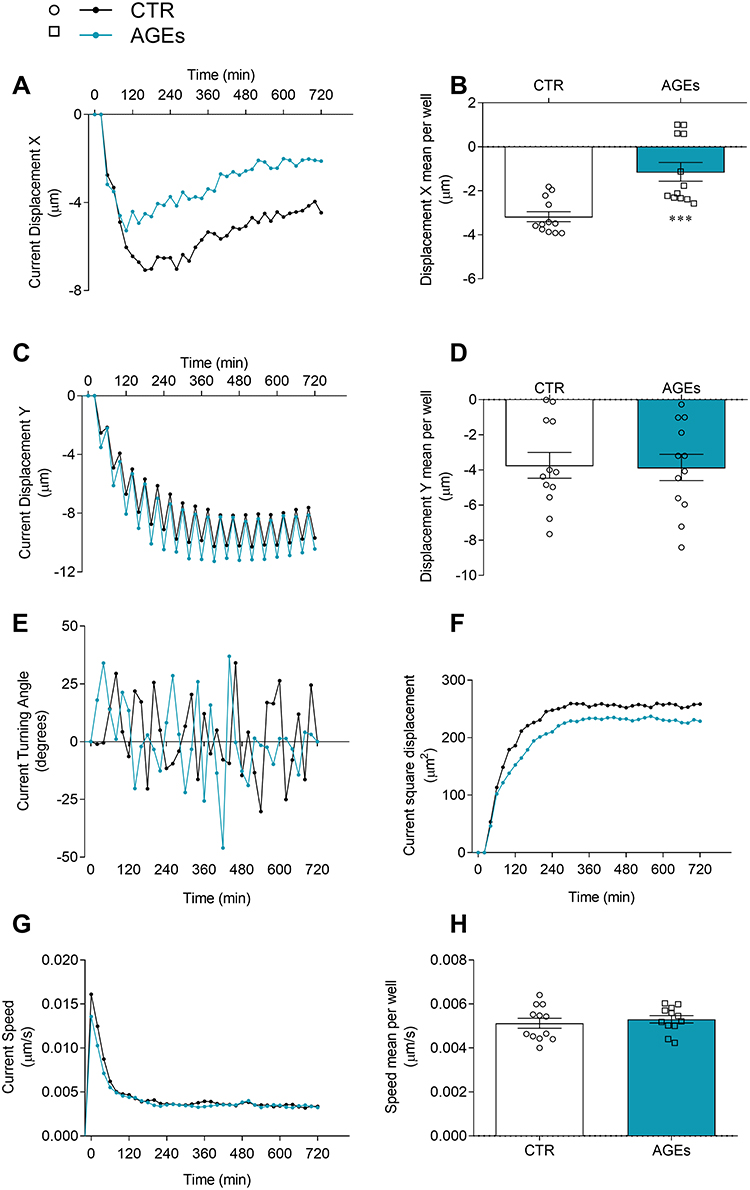 |
Figure 6 Effects of AGEs on kinetic properties of HLMs. HLMs (105 cells/well) were incubated (12 hours, 37°C, 5% CO2) with RPMI alone (CTR) or AGEs (100μg/mL). The incubation was carried out with time-lapse and high-content microscopy Operetta High-Content Imaging System (PerkinElmer) to investigate the tracking characteristics as current displacement X (A), displacement X mean per well (B), current displacement Y (C), displacement Y mean per well (D), current turning angle (E), current square displacement (F), current speed (G), and speed mean per well (H). Data are shown as individual values with mean ± SEM of 12 independent experiments obtained from different donors. *** p< 0.001 vs CTR. |
Effects of AGEs on Phagocytosis
Phagocytosis is a protective mechanism of innate immunity that plays a key role in the elimination of microbial agents and debris.32 In the next group of experiments, we evaluated whether AGEs could affect phagocytosis by HLMs. To this end, spectrophotometric reader was used to measure the uptake of pHrodo™ green labeled E. coli particles over time in the same well. Figure 7 shows that while AGEs did not significantly affect HLM phagocytic activity during the first 2 hours of incubation, they significantly inhibited phagocytosis after 3 hours.
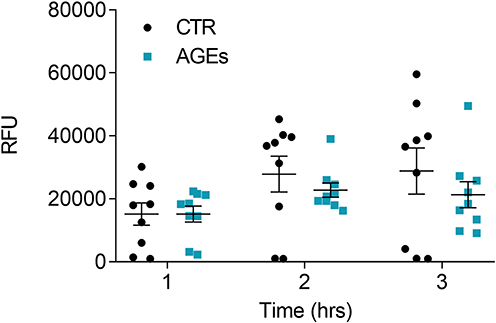 |
Figure 7 Effect of AGEs on HLM phagocytic activity. HLMs (105 cells/well) were incubated (16 hours, 37°C, 5% CO2) with RPMI alone (CTR), and with AGEs (100 µg/mL) one day before phagocytosis assay. The day after, the cell culture medium was removed and pHrodo™ Green E. coli BioParticles™ Conjugate suspension was added to the wells. The plate was placed in an EnSpire Multimode Plate Reader (PerkinElmer). The data were expressed as Relative Fluorescence Units (RFU) measured up to 3 hours with a 1 h span at an excitation wavelength of 509 nm, and emission at 533 nm. Data are shown as individual values with mean ± SEM of 9 independent experiments obtained from different donors. ** p< 0.01 vs CTR. |
Determination of AGEs
In a last group of experiments, we evaluated the presence of AGEs in fresh and cultured HLMs. The lysate of fresh HLMs contained AGEs (Figure 8). In addition, after 16 hours of incubation (at 37°C, 5% CO2) HLMs stored and spontaneously release the same concentration of AGEs (Figure 8).
 |
Figure 8 Expression of AGEs in HLMs. HLMs (5 x 106) were freshly lysed with 0.1% Triton X-100. HLMs were cultured (16 hours) and then lysed in the plates with 0.1% Triton X-100, while supernatants were collected. Concentration of AGEs in supernatants and in cellular lysates were measured using commercially available ELISA kit for AGEs. Data are shown as individual values with mean ± SEM of 6 independent experiments obtained from different donors. |
To assess the basal content of AGEs in HLMs, we quantified AGE levels in both freshly isolated and cultured HLM preparation. Freshly isolated HLMs, consisting of approximately 85% pulmonary macrophages18 directly lysed after percoll gradient, exhibited detectable levels of AGEs. Cultured HLMs, representing a more purified population (98% macrophages) after adherence,18 showed a lower AGE content in cell lysates compared to freshly isolated cells. It is possible that the non-macrophage cell types present in the fresh HLM preparation (approximately 15% of the total cells) contribute to the higher AGE levels observed in these samples.
Furthermore, we investigated the basal release of AGEs from cultured HLMs. After 16 hours of incubation (at 37°C, 5% CO2), the concentration of AGEs in the culture supernatant was measured and found to be comparable to that in the cultured HLM lysates. This finding suggests that HLMs may passively release or actively secrete AGEs into the extracellular environment, potentially contributing to the overall AGE burden in the lung microenvironment.
Discussion
In the present study, we investigated the effect of AGEs on the activation of HLMs isolated from non-tumour human lung tissue from a total of 30 patients who underwent lung surgery. We demonstrate that AGEs induce cytokines/chemokines release, but not production except for IL-6, from HLMs and they affect kinetic proprieties of HLMs. We also reveal that HLMs constitutively express both sRAGE and flRAGE mRNAs and RAGE protein. In addition, AGEs stimulation slightly upregulates sRAGE mRNA expression but do not affect RAGE protein. Finally, HLMs store and spontaneously release AGEs.
AGEs can be formed endogenously as part of normal metabolism, but their accumulation is accelerated in conditions characterized by smoking status, exposure to air pollution, oxidative stress, inflammation, and aging.2,4–6,33,34 Their harmful effects on various tissues, including the lung, are ascribed to their chemical, pro-oxidant, and inflammatory properties.15,16,33,35,36 In line with these papers, here we demonstrate that the predominant immune cells in the lung, macrophages, are activated by AGEs showing a proinflammatory profile. In fact, they release, upon AGEs stimulation, CXCL1, CXCL2 and CXCL8 that act as chemoattractants for immune cells, especially neutrophils, to the site of injury playing a crucial role in regulation of immune and inflammatory responses.37,38 Mouse macrophage cell lines release IL-6, TNF-α and IL-1β under stimulation with AGEs,39,40 here we show that also AGEs-activated HLMs release cytokines (IL-1β, IL-6, TNF-α) that are responsible for immune activation in response to cell damage.41 These mediators are markedly increased in induced sputum of COPD patients regulating inflammatory responses and immune cell recruitment in the lung.42–45 Recent studies demonstrated that AGEs could be implicated in pathophysiology of COPD and IPF.15,16 Therefore, we could hazard the hypothesis that, in some variants of COPD and IPF, the elevated levels of chemokines/cytokines could be due also to the effects of AGEs on macrophages. Here we show, by kinetic and mRNA experiments, that AGEs induce the release of pre-formed mediators stored in HLMs but not the production, except for IL-6 whom release is accentuated after 16 hours. These results reflect the potential ability of AGEs to induce chronic inflammatory disease after both short- and long-term exposure.
The biological impact of AGEs operates through two distinct mechanisms: one independent of the receptor and another that involves the receptor RAGE.2 AGEs interacting with the RAGE activates intracellular signal pathways leading the transcription factor NF-κB and in turn the production of pro-inflammatory cytokines.2 Previous studies have demonstrated that RAGE is constitutively highly expressed in the lung and play critical role of RAGE signalling in modulating the inflammatory and immune responses of alveolar macrophages.9,10,46 RAGE seems to be expressed in murine alveolar macrophages,47 but there was currently no evidence of RAGE expression in human models of pulmonary macrophages. We demonstrate that fresh and cultured HLMs constitutively express both RAGE mRNA and protein. More in detail, sRAGE mRNA seems to be more expressed in HLMs, compared with flRAGE mRNA. Although RAGE expression was increased under its ligands accumulation,10 stimulation of HLM with AGEs did not affect RAGE expression. However, clinical studies have shown that circulating levels of sRAGE are significantly reduced in patients with COPD and IPF.48,49 This systemic sRAGE deficiency could reduce the neutralization of AGEs, favouring their interaction with flRAGE on lung macrophages. We hypostasize that this deficiency in sRAGE could thus favour activation of lung macrophages, contributing to the persistent inflammation observed in these patients. Support for a RAGE-dependent mechanism in our system comes from experiments in which HLMs were pre-treated with a neutralizing anti-RAGE antibody: this intervention significantly reduced AGE-induced cytokine release, indicating that at least part of the pro-inflammatory response is RAGE-mediated.
Studying cell tracking by time-lapse and high-content microscopy, we showed that AGEs affect HLM kinetic proprieties. Specifically, AGEs induced significant changes in cell displacement along X-axis, reducing HLM movement earlier in the space. This finding raises important questions about the potential mechanisms by which AGEs influence macrophage motility and the implications for pulmonary health. The mechanism of macrophage movement in the space is still not fully understood. Murine alveolar macrophages have been shown to crawl along the alveolar space in response to inhaled pathogens and debris50 and adhesion molecules, such as integrins, seems to play a crucial role in mediating cell adhesion to the extracellular matrix and promoting cell movement.51 Reduced expression or dysfunction of these molecules can hinder macrophage migration.51 It would be interesting to evaluate whether AGEs could potentially impair integrin expression and/or function, but further investigations are needed. However, our suggests that AGEs may restrict the surveillance capacity of HLMs, potentially impairing their ability to effectively reach and clear pathogens and debris from the lung. This aligns with previous studies demonstrating that AGEs can compromise various protective mechanisms of macrophages.52,53
Macrophages are phagocytic cells that reside in the blood and tissues waiting to engulf foreign substances.54 The phagocytic ability of macrophages is impaired by several pathological conditions55 including chronic alcohol ingestion56 and HIV infection.57 AGEs impaired phagocytic function in murine macrophages,52,53 our results suggest that AGEs accumulation could potentially inhibit phagocytosis in HLMs.
It was initially hypothesized that AGEs were slightly engrossed by cells, and their impact on human health was largely ignored. Afterwards, it has been discovered the presence of a significant pool of AGEs in the cells58,59 and that the excessive AGEs accumulation in the cells may affect both long- and short-lived proteins and peptides.60 Our study reveals that HLMs also could contribute to the overall AGE pool, potentially through intracellular AGE formation and release. However, the primary focus of our investigation was the effect of exogenous AGEs on HLM activation.
Conclusions
In conclusion, since AGEs accumulation seems to be associated with impaired respiratory function,15 our preliminary findings provide novel insights into the cellular mechanisms underlying the impact of AGEs accumulation on respiratory system. Our findings demonstrate that AGEs induce HLM activation primarily through interaction with the RAGE, leading to increased production of pro-inflammatory cytokines and chemokines, such as CXCL8, IL-6, and TNF-α. This inflammatory response, triggered by AGE accumulation, may contribute to lung injury and impaired respiratory function. Notably, while we observed increased release of several pro-inflammatory mediators, IL-6 stood out as the only cytokine produced de novo, suggesting a potential role in chronic inflammation. Furthermore, we show that AGEs can inhibit phagocytosis by macrophages, a critical protective mechanism for clearing pathogens and cellular debris, which could indirectly affect the lung response to infection. While our experiments utilized purified HLMs, as confirmed by flow cytometry,18 it is important to acknowledge the existing heterogeneity within the HLM population,61,62 and future research should investigate the specific responses of these subpopulations to AGEs. Moreover, our study focused on tissue-resident human lung macrophages obtained from surgical samples, which limited the possibility of comparing macrophages from healthy individuals and patients with inflammatory lung conditions. Future studies could consider the use of macrophages derived from bronchoalveolar lavage or in vitro models, such as alveolar macrophage-like cells differentiated from peripheral blood mononuclear cells,63 although these approaches may not fully recapitulate the features of tissue-resident macrophages. Despite these limitations, our study provides valuable insights into the potential role of HLMs in driving AGE-related pulmonary inflammation and suggests that targeting the AGE-RAGE axis in HLMs may hold therapeutic promise.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author ([email protected]; [email protected]) upon reasonable request.
Ethics Approval
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of University of Naples Federico II (Prot. 7/19 and 301/12).
Patient Consent
Informed consent was obtained from all individual participants included in the study.
Acknowledgments
The authors thank scientists from CISI Laboratory not listed as authors for invaluable collaborations, the administrative staff (Roberto Bifulco, Anna Ferraro and Gjada Criscuolo), without whom we could not function as an integrated team. The authors apologize to colleagues whose papers could not be cited due to space and citation restrictions.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Supplementary Material
The online version of this article includes the following supplementary materials. Supplementary Figure 1: experiments showing cell viability assessed with MTT assay (a) and IL-6 release upon treatment with AGEs, LPS, and the effect of polymyxin B, to exclude LPS contamination (b). Supplementary Figure 2: flow cytometry gating strategy used to identify RAGE+ HLMs.
Funding
This work was supported in part by grants from the CISILab Project (University of Naples Federico II), PRIN 2022TC5JJW, investigator-initiated research grant IIT-ITA-002138, TIMING Project and Campania Bioscience (Regione Campania) and by PNRR-MAD-2022-12376769, PRIN-MUR P202282S5M, AIRC under MFAG 2020 (25123).
Disclosure
The authors declare no conflict of interest.
References
1. Prasad C, Davis KE, Imrhan V, et al. Advanced glycation end products and risks for chronic diseases: intervening through lifestyle modification. Am J Lifestyle Med. 2019;13(4):384–404. doi:10.1177/1559827617708991
2. Luevano-Contreras C, Chapman-Novakofski K. Dietary advanced glycation end products and aging. Nutrients. 2010;2(12):1247–1265. doi:10.3390/nu2121247
3. Antognelli C, Talesa VN. Glyoxalases in urological malignancies. Int J Mol Sci. 2018;19(2):415. doi:10.3390/ijms19020415
4. Prasad K, Dhar I, Caspar-Bell G. Role of advanced glycation end products and its receptors in the pathogenesis of cigarette smoke-induced cardiovascular disease. Int J Angiol. 2015;24(2):75–80. doi:10.1055/s-0034-1396413
5. Chen CY, Zhang JQ, Li L, et al. Advanced glycation end products in the skin: molecular mechanisms, methods of measurement, and inhibitory pathways. Front Med. 2022;9:837222. doi:10.3389/fmed.2022.837222
6. Mengstie MA, Chekol Abebe E, Behaile Teklemariam A, et al. Endogenous advanced glycation end products in the pathogenesis of chronic diabetic complications. Front Mol Biosci. 2022;9:1002710. doi:10.3389/fmolb.2022.1002710
7. Chaudhuri J, Bains Y, Guha S, et al. The role of advanced glycation end products in aging and metabolic diseases: bridging association and causality. Cell Metab. 2018;28(3):337–352. doi:10.1016/j.cmet.2018.08.014
8. Oczypok EA, Perkins TN, Oury TD. All the “RAGE” in lung disease: the receptor for advanced glycation endproducts (RAGE) is a major mediator of pulmonary inflammatory responses. Paediatr Respir Rev. 2017;23:40–49. doi:10.1016/j.prrv.2017.03.012
9. Byun K, Yoo Y, Son M, et al. Advanced glycation end-products produced systemically and by macrophages: a common contributor to inflammation and degenerative diseases. Pharmacol Ther. 2017;177:44–55. doi:10.1016/j.pharmthera.2017.02.030
10. Robinson AB, Johnson KD, Bennion BG, et al. RAGE signaling by alveolar macrophages influences tobacco smoke-induced inflammation. Am J Physiol Lung Cell Mol Physiol. 2012;302(11):L1192–9. doi:10.1152/ajplung.00099.2012
11. Johnson LL, Tekabe Y, Zelonina T, et al. Blocking RAGE expression after injury reduces inflammation in mouse model of acute lung injury. Respir Res. 2023;24(1):21. doi:10.1186/s12931-023-02324-6
12. Lutterloh EC, Opal SM, Pittman DD, et al. Inhibition of the RAGE products increases survival in experimental models of severe sepsis and systemic infection. Crit Care. 2007;11(6):R122. doi:10.1186/cc6184
13. Reddy VP, Aryal P, Darkwah EK. Advanced glycation end products in health and disease. Microorganisms. 2022;10(9):1848. doi:10.3390/microorganisms10091848
14. Paparo L, Coppola S, Nocerino R, et al. How dietary advanced glycation end products could facilitate the occurrence of food allergy. J Allergy Clin Immunol. 2024;153(3):742–758. doi:10.1016/j.jaci.2023.11.023
15. Zaigham S, Persson M, Jujic A, et al. Measures of lung function and their relationship with advanced glycation end-products. ERJ Open Res. 2020;6(2). doi:10.1183/23120541.00356-2019
16. Hoonhorst SJ, Lo Tam Loi AT, Hartman JE, et al. Advanced glycation end products in the skin are enhanced in COPD. Metabolism. 2014;63(9):1149–1156. doi:10.1016/j.metabol.2014.06.006
17. Palestra F, Poto R, Ciardi R, et al. SARS-CoV-2 spike protein activates human lung macrophages. Int J Mol Sci. 2023;24(3):3036. doi:10.3390/ijms24033036
18. Balestrieri B, Granata F, Loffredo S, et al. Phenotypic and functional heterogeneity of low-density and high-density human lung macrophages. Biomedicines. 2021;9(5):505. doi:10.3390/biomedicines9050505
19. Granata F, Frattini A, Loffredo S, et al. Production of vascular endothelial growth factors from human lung macrophages induced by group IIA and group X secreted phospholipases A2. J Immunol. 2010;184(9):5232–5241. doi:10.4049/jimmunol.0902501
20. Loffredo S, Staiano RI, Granata F, et al. Immune cells as a source and target of angiogenic and lymphangiogenic factors. Chem Immunol Allergy. 2014;99:15–36. doi:10.1159/000353316
21. Mao QY, He SY, Hu QY, et al. Advanced glycation end products (AGEs) inhibit macrophage efferocytosis of apoptotic beta cells through binding to the receptor for AGEs. J Immunol. 2022;208(5):1204–1213. doi:10.4049/jimmunol.2100695
22. Jin X, Yao T, Zhou Z, et al. Advanced glycation end products enhance macrophages polarization into m1 phenotype through activating RAGE/NF-kappaB pathway. Biomed Res Int. 2015;2015:732450. doi:10.1155/2015/732450
23. Scorziello A, Pellegrini C, Secondo A, et al. Neuronal NOS activation during oxygen and glucose deprivation triggers cerebellar granule cell death in the later reoxygenation phase. J Neurosci Res. 2004;76(6):812–821. doi:10.1002/jnr.20096
24. Borriello F, Iannone R, Di Somma S, et al. GM-CSF and IL-3 modulate human monocyte TNF-alpha production and renewal in in vitro models of trained immunity. Front Immunol. 2016;7:680. doi:10.3389/fimmu.2016.00680
25. Borriello F, Iannone R, Di Somma S, et al. Lipopolysaccharide-elicited TSLPR expression enriches a functionally discrete subset of human CD14(+) CD1c(+) monocytes. J Immunol. 2017;198(9):3426–3435. doi:10.4049/jimmunol.1601497
26. Staiano RI, Loffredo S, Borriello F, et al. Human lung-resident macrophages express CB1 and CB2 receptors whose activation inhibits the release of angiogenic and lymphangiogenic factors. J Leukoc Biol. 2016;99(4):531–540. doi:10.1189/jlb.3HI1214-584R
27. Kolomaznik M, Liskayova G, Kanjakova N, et al. The perturbation of pulmonary surfactant by bacterial lipopolysaccharide and its reversal by polymyxin B: function and structure. Int J Mol Sci. 2018;19(7):1964. doi:10.3390/ijms19071964
28. Lee EJ, Park JH. Receptor for advanced glycation endproducts (RAGE), its ligands, and soluble RAGE: potential biomarkers for diagnosis and therapeutic targets for human renal diseases. Genomics Inform. 2013;11(4):224–229. doi:10.5808/GI.2013.11.4.224
29. Schmidt AM. 22016 ATVB plenary lecture: receptor for advanced glycation endproducts and implications for the pathogenesis and treatment of cardiometabolic disorders: spotlight on the macrophage. Arterioscler Thromb Vasc Biol. 2017;37(4):613–621. doi:10.1161/ATVBAHA.117.307263
30. Caballero D, Voituriez R, Riveline D. Protrusion fluctuations direct cell motion. Biophys J. 2014;107(1):34–42. doi:10.1016/j.bpj.2014.05.002
31. Wiesner C, Le-Cabec V, El Azzouzi K, et al. Podosomes in space: macrophage migration and matrix degradation in 2D and 3D settings. Cell Adh Migr. 2014;8(3):179–191. doi:10.4161/cam.28116
32. Gordon S. Phagocytosis: an Immunobiologic Process. Immunity. 2016;44(3):463–475. doi:10.1016/j.immuni.2016.02.026
33. Khalid M, Petroianu G, Adem A. Advanced glycation end products and diabetes mellitus: mechanisms and perspectives. Biomolecules. 2022;12(4):542. doi:10.3390/biom12040542
34. Twarda-Clapa A, Olczak A, Bialkowska AM, et al. Advanced glycation end-products (AGEs): formation, chemistry, classification, receptors, and diseases related to AGEs. Cells. 2022;11(8):1312. doi:10.3390/cells11081312
35. Rungratanawanich W, Y Q, Wang X, et al. Advanced glycation end products (AGEs) and other adducts in aging-related diseases and alcohol-mediated tissue injury. Exp Mol Med. 2021;53(2):168–188. doi:10.1038/s12276-021-00561-7
36. Davis KE, Prasad C, Vijayagopal P, et al. Advanced glycation end products, inflammation, and chronic metabolic diseases: links in a chain? Crit Rev Food Sci Nutr. 2016;56(6):989–998. doi:10.1080/10408398.2012.744738
37. Metzemaekers M, Gouwy M, Proost P. Neutrophil chemoattractant receptors in health and disease: double-edged swords. Cell Mol Immunol. 2020;17(5):433–450. doi:10.1038/s41423-020-0412-0
38. Sawant KV, Sepuru KM, Penaranda B, et al. Chemokine Cxcl1-Cxcl2 heterodimer is a potent neutrophil chemoattractant. J Leukoc Biol. 2023;114(6):666–671. doi:10.1093/jleuko/qiad097
39. He S, Hu Q, Xu X, et al. Advanced glycation end products enhance M1 macrophage polarization by activating the MAPK pathway. Biochem Biophys Res Commun. 2020;525(2):334–340. doi:10.1016/j.bbrc.2020.02.053
40. Qiang X, Liang S, Lv Y, et al. Advanced glycation end products (AGEs) impair the intestinal epithelial barrier via STAT3 activation mediated by macrophages. Food Chem Toxicol. 2024;192:114966. doi:10.1016/j.fct.2024.114966
41. Marcella S, Apicella B, Secondo A, et al. Size-based effects of anthropogenic ultrafine particles on activation of human lung macrophages. Environ Int. 2022;166:107395. doi:10.1016/j.envint.2022.107395
42. Dawson RE, Jenkins BJ, Saad MI. IL-6 family cytokines in respiratory health and disease. Cytokine. 2021;143:155520. doi:10.1016/j.cyto.2021.155520
43. Henrot P, Prevel R, Berger P, et al. Chemokines in COPD: from Implication to Therapeutic Use. Int J Mol Sci. 2019;20(11):2785. doi:10.3390/ijms20112785
44. Barnes PJ. The cytokine network in chronic obstructive pulmonary disease. Am J Respir Cell Mol Biol. 2009;41(6):631–638. doi:10.1165/rcmb.2009-0220TR
45. Lazar M, Sandulescu M, Barbu EC, et al. The role of cytokines and molecular pathways in lung fibrosis following SARS-CoV-2 infection: a physiopathologic (Re)view. Biomedicines. 2024;12(3):639. doi:10.3390/biomedicines12030639
46. Sanders KA, Delker DA, Huecksteadt T, et al. RAGE is a critical mediator of pulmonary oxidative stress, alveolar macrophage activation and emphysema in response to cigarette smoke. Sci Rep. 2019;9(1):231. doi:10.1038/s41598-018-36163-z
47. Wolf L, Herr C, Niederstrasser J, et al. Receptor for advanced glycation endproducts (RAGE) maintains pulmonary structure and regulates the response to cigarette smoke. PLoS One. 2017;12(7):e0180092. doi:10.1371/journal.pone.0180092
48. Miniati M, Monti S, Basta G, et al. Soluble receptor for advanced glycation end products in COPD: relationship with emphysema and chronic cor pulmonale: a case-control study. Respir Res. 2011;12(1):37. doi:10.1186/1465-9921-12-37
49. Machahua C, Montes-Worboys A, Planas-Cerezales L, et al. Serum AGE/RAGEs as potential biomarker in idiopathic pulmonary fibrosis. Respir Res. 2018;19(1):215. doi:10.1186/s12931-018-0924-7
50. Neupane AS, Willson M, Chojnacki AK, et al. Patrolling alveolar macrophages conceal bacteria from the immune system to maintain homeostasis. Cell. 2020;183(1):110–125e11. doi:10.1016/j.cell.2020.08.020
51. Cui K, Ardell CL, Podolnikova NP, et al. Distinct migratory properties of M1, M2, and resident macrophages are regulated by alpha(D)beta(2) and alpha(M)beta(2) integrin-mediated adhesion. Front Immunol. 2018;9:2650. doi:10.3389/fimmu.2018.02650
52. Wang Q, Zhu G, Cao X, et al. Blocking AGE-RAGE signaling improved functional disorders of macrophages in diabetic wound. J Diabetes Res. 2017;2017:1428537. doi:10.1155/2017/1428537
53. Liu BF, Miyata S, Kojima H, et al. Low phagocytic activity of resident peritoneal macrophages in diabetic mice: relevance to the formation of advanced glycation end products. Diabetes. 1999;48(10):2074–2082. doi:10.2337/diabetes.48.10.2074
54. Santa K. Macrophages: phagocytosis, antigen presentation, and activation of immunity. Phagocytosis - Main Key of Immune System. 2023.
55. Hirayama D, Iida T, Nakase H. The phagocytic function of macrophage-enforcing innate immunity and tissue homeostasis. Int J Mol Sci. 2017;19(1):92. doi:10.3390/ijms19010092
56. Mehta AJ, Guidot DM. Alcohol and the Lung. Alcohol Res. 2017;38(2):243–254. doi:10.35946/arcr.v38.2.07
57. Presti RM, Flores SC, Palmer BE, et al. Mechanisms underlying HIV-associated noninfectious lung disease. Chest. 2017;152(5):1053–1060. doi:10.1016/j.chest.2017.04.154
58. Schalkwijk CG, Brouwers O, Stehouwer CD. Modulation of insulin action by advanced glycation endproducts: a new player in the field. Horm Metab Res. 2008;40(9):614–619. doi:10.1055/s-0028-1082085
59. Horace S, Isbell HLF. Mutarotation, hydrolysis, and rearrangement reactions of glycosylamines. J Org Chem. 1958;23(9).
60. Vistoli G, De Maddis D, Cipak A, et al. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): an overview of their mechanisms of formation. Free Radic Res. 2013;47 Suppl 1(sup1):3–27. doi:10.3109/10715762.2013.815348
61. Vieira Braga FA, Kar G, Berg M, et al. A cellular census of human lungs identifies novel cell states in health and in asthma. Nat Med. 2019;25(7):1153–1163. doi:10.1038/s41591-019-0468-5
62. Lavin Y, Kobayashi S, Leader A, et al. Innate immune landscape in early lung adenocarcinoma by paired single-cell analyses. Cell. 2017;169(4):750–765e17. doi:10.1016/j.cell.2017.04.014
63. Pahari S, Neehus AL, Trapnell BC, et al. Protocol to develop human alveolar macrophage-like cells from mononuclear cells or purified monocytes for use in respiratory biology research. STAR Protoc. 2024;5(2):103061. doi:10.1016/j.xpro.2024.103061
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Bacteriocin BacSp222 and Its Succinylated Forms Exhibit Proinflammatory Activities Toward Innate Immune Cells
Śmiałek J, Bzowska M, Hinz A, Mężyk-Kopeć R, Sołtys K, Mak P
Journal of Inflammation Research 2022, 15:4601-4621
Published Date: 12 August 2022
The Effects of Budesonide Inhalation Treatment on the Expression Levels of Serum IL-6, TGF-β1, and IgE and Pulmonary Function in Patients with Cough Variant Asthma and an Evaluation of Treatment Efficacy
Niu Y, Cao M, Li S, Mo J, Zhu Z, Wang H
International Journal of General Medicine 2024, 17:1253-1261
Published Date: 29 March 2024
Based on the Dual Pathway of Interaction-Mediated NF-κB in Cell Apoptosis and Immune Inflammation to Study the Effect of Danzhi Xiaoyao Powder on the Learning and Cognitive Ability of AD Model Rats
Wang HP, Li MC, Yang J, Zhou J, Meng ZP, Hu YY, Lyu YJ, Chen YQ, Han YM, Pei WL
Degenerative Neurological and Neuromuscular Disease 2025, 15:41-64
Published Date: 14 April 2025