Back to Journals » Breast Cancer: Targets and Therapy » Volume 17
Differential TGF-β1/SMAD4 Signaling Regulates PMN-MDSC Differentiation and Reshapes the Immune Microenvironment in Breast Cancer
Authors Cao J, Qiao E, Shao H, Wang C, Xia M, Qin B
Received 9 October 2025
Accepted for publication 23 December 2025
Published 26 December 2025 Volume 2025:17 Pages 1369—1383
DOI https://doi.org/10.2147/BCTT.S566995
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Pranela Rameshwar
Jie Cao,1,2 Enqi Qiao,1,2 Hui Shao,2,3 Chunlei Wang,1,2 Manzhi Xia,1,2 Bo Qin4
1Department of General Surgery, Shaoxing Maternity and Child Health Care Hospital, Shaoxing, Zhejiang, People’s Republic of China; 2School of Medicine, Maternity and Child Health Care Affiliated Hospital, Shaoxing University, Shaoxing, Zhejiang, People’s Republic of China; 3Infection Control Office, Shaoxing Maternity and Child Health Care Hospital, Shaoxing, Zhejiang, People’s Republic of China; 4Department of Laboratory Medicine, Shaoxing Maternity and Child Health Care Hospital, Shaoxing, Zhejiang, People’s Republic of China
Correspondence: Bo Qin, Department of Laboratory Medicine, Shaoxing Maternity and Child Health Care Hospital, Shaoxing, Zhejiang, People’s Republic of China, Email [email protected]
Purpose: Polymorphonuclear myeloid-derived suppressor cells (PMN-MDSCs) exert strong immunosuppressive effects and are associated with poor prognosis in breast cancer. However, the mechanism underlying their differentiation remains unclear. Here, we aimed to elucidate how transforming growth factor-β 1 (TGF-β 1)/SMAD4 signaling regulates PMN-MDSC differentiation and modulates the breast cancer immune microenvironment.
Methods: We analyzed neutrophil subpopulation heterogeneity in breast cancer tissues using single-cell RNA sequencing combined with spatial transcriptomics (GSE176078 dataset). A 4T1 mouse breast cancer metastasis model was established, and tumor progression was tracked via in vivo fluorescence imaging. Colony formation and differentiation outcomes following Tgfb1 overexpression or knockdown were assessed in spleen-derived hematopoietic stem cells using flow cytometry. Gene expression profiles and signaling pathways were characterized using RNA sequencing and Western blotting.
Results: Neutrophil subpopulations in breast cancer tissues exhibited significant heterogeneity, particularly the N3 subset, which showed enhanced intracellular communication and regulatory potential. Tgfb1 was significantly upregulated along the neutrophil developmental trajectory in both spleen tissue and serum of breast cancer mice. In normal mice, the number of metastatic foci significantly correlated with neutrophil proportions. Tgfb1 overexpression promoted PMN-MDSC differentiation, colony formation, and SMAD4 upregulation in normal mice but exerted opposite effects in tumor-bearing mice.
Conclusion: Our findings implicate TGF-β 1/SMAD4 signaling in the differential regulation of PMN-MDSC differentiation in breast cancer models. The TGF-β 1 pathway represents a potential therapeutic target for modulating the immune microenvironment in breast cancer, although further validation in clinical settings is required to determine therapeutic efficacy.
Plain Language Summary: This study demonstrates that the transforming growth factor-β 1 (TGF-β 1)/SMAD4 signaling pathway differentially regulates the differentiation of polymorphonuclear myeloid-derived suppressor cells (PMN-MDSCs) in breast cancer. Specifically, TGF-β 1 promotes the development of immunosuppressive cells under healthy conditions, whereas it exerts opposing effects in tumor-bearing environments. These findings suggest that TGF-β 1 represents a promising therapeutic target for modulating the immune microenvironment in breast cancer and enhancing clinical outcomes for patients.
Keywords: immunotherapy, polymorphonuclear neutrophil, myeloid-derived suppressor cell, tumor microenvironment
Introduction
Breast cancer is the most common malignancy among women worldwide and presents a highly complex tumor microenvironment (TME), which plays a central role in facilitating tumor immune escape. The breast cancer TME contains diverse immune cell populations, among which myeloid-derived suppressor cells (MDSCs), a heterogeneous group of immature myeloid cells, exhibit potent immunosuppressive properties. MDSCs can inhibit T cell activation and proliferation, promote tumor immune escape, and are closely associated with tumor progression and metastasis.1
Polymorphonuclear MDSCs (PMN-MDSCs), a major MDSC subtype, share phenotypic similarities with neutrophils but possess markedly enhanced immunosuppressive activity. PMN-MDSCs suppress CD8+ T-cell activation and cytotoxicity by secreting immunosuppressive mediators, including arginase 1 and reactive oxygen species (ROS), thereby promoting tumor cell proliferation, invasion, and metastasis.2 Despite their importance, the developmental pathways and regulatory mechanisms underlying PMN-MDSC differentiation within the breast cancer microenvironment remain poorly understood.
Transforming growth factor-β1 (TGF-β1) is a multifunctional cytokine that regulates cell proliferation, differentiation, apoptosis, and immune responses. Within the TME, TGF-β1 presents dual functions: it acts as a tumor suppressor by inhibiting epithelial cell proliferation during early tumorigenesis but promotes metastasis in advanced tumors by inducing epithelial–mesenchymal transition, enhancing angiogenesis, and suppressing antitumor immunity.3 Significantly higher serum TGF-β1 levels have been reported in patients with breast cancer than in healthy controls, which positively correlates with tumor size, lymph node metastasis, and poor prognosis.4
TGF-β1 signals through the canonical SMAD-dependent pathway; the SMAD protein family functions as key intracellular effectors of canonical TGF-β1 signaling, which regulates hematopoietic cell fate decisions and myeloid differentiation.5 When the TGF-β1 ligand binds to its receptor complex (TβRII and TβRI), SMAD2 and SMAD3 are phosphorylated and form a heterotrimeric complex with the common mediator SMAD4. This SMAD2/3-SMAD4 complex translocates to the nucleus, where it regulates the transcription of myeloid lineage-specific genes and immunosuppressive effector molecules.6 However, the role of TGF-β1/SMAD4 signaling in regulating PMN-MDSC differentiation and modulating the breast cancer microenvironment remains largely undefined.
Recent mechanistic studies have revealed that TGF-β signaling exhibits context-dependent effects on myeloid cell differentiation, with low concentrations promoting myeloid-biased hematopoietic stem cell proliferation and granulocytic differentiation, whereas higher concentrations induce quiescence.7 Furthermore, TGF-β1 directly enhances MDSC expansion, particularly the monocytic MDSC subset, by upregulating arginase 1 and other immunosuppressive molecules through SMAD-dependent transcriptional activation.8
SMAD4 functions as a critical determinant of the MDSC phenotype and TME. Loss of SMAD4 in gastric cancer cells has been associated with increased accumulation of granulocytic MDSCs, driven by altered secretome profiles, highlighting SMAD4 as a key regulator linking tumor-derived signals to MDSC recruitment and differentiation.9 However, the precise mechanistic role of TGF-β1/SMAD4 signaling in directing PMN-MDSC differentiation from hematopoietic precursors in breast cancer is inadequately understood, and the paradoxical effects of TGF-β1 on myeloid differentiation under physiological versus pathological conditions have not been systematically investigated.Despite the importance of TGF-β1 in immune regulation and the role of PMN-MDSCs in breast cancer progression, the specific molecular mechanisms through which TGF-β1/SMAD4 signaling governs PMN-MDSC differentiation in tumor-bearing versus healthy states remain unclear. Specific questions that remain to be answered include whether and how the TGF-β1/SMAD4 pathway exerts differential regulatory effects on granulocytic differentiation and PMN-MDSC development under normal physiological conditions compared to the TME. Understanding these context-dependent mechanisms is essential for developing targeted immunotherapeutic strategies that can modulate MDSC-mediated immunosuppression without disrupting normal hematopoietic homeostasis. Therefore, this study aimed to elucidate the regulatory mechanisms through which the TGF-β1/SMAD4 pathway influences PMN-MDSC differentiation under both normal and tumor-bearing conditions and to determine how differential transduction of this signaling cascade reshapes the breast cancer immune landscape. By addressing this critical gap in the “tumor signaling–immune cell regulation–microenvironment remodeling” axis, our findings may inform new strategies for overcoming current limitations in breast cancer immunotherapy.
Materials and Methods
Single-Cell RNA Sequencing and Spatial Transcriptomics Analysis
The GSE176078 dataset was obtained from the Gene Expression Omnibus database (ncbi.nlm.nih.gov/geo/). It comprises 26 primary tumor samples representing 4 major clinical subtypes of breast cancer. Neutrophil subpopulation heterogeneity was examined using single-cell RNA sequencing (scRNA-seq) data. Uniform manifold approximation and projection (UMAP) dimension reduction was performed using Seurat (version 4.3.0) and the following parameters: n_neighbors = 15, min_dist = 0.5, and 2D projection. Differential gene expression analysis was conducted using DESeq2 (version 1.34.0), with significance defined as P < 0.01 and |log2 fold change| > 1.0. Cell–cell communication networks were quantified using CellChat (version 1.6.1) to identify ligand–receptor interactions and assess intercellular communication strength. Genomic stability across neutrophil subpopulations was evaluated using Inference of Copy Number Variations (InferCNV) with the N1 subpopulation as reference. Pseudo-temporal trajectory analysis reconstructed developmental pathways of neutrophil subpopulations in two-dimensional space, which were subsequently mapped onto tissue sections through spatial transcriptomics (stRNA-seq) to elucidate the spatial distribution patterns of differentially expressed genes.
Animals and Cell Lines
Animal procedures were conducted in accordance with the National Regulations for the Administration of Laboratory Animals in China (2017 Revision) and NIH Guide for the Care and Use of Laboratory Animals (8th edition, 2011). All animal experiments were approved by the Institutional Animal Care and Use Committee of Shaoxing Maternity and Child Health Care Hospital (approval no. 2025–075).
Forty-eight BALB/c female mice (18–22 g) were purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China) and housed under controlled conditions (20–26°C, 40–60% relative humidity, and a 12-h light–dark cycle) with ad libitum access to food and water and uniform supply of bedding materials by the facility to ensure experimental consistency. After one week of acclimatization, the mice were divided into two groups (n = 24 per group) based on their weight: normal and model.
The 4T1 mouse breast cancer cells (Yipu Biotechnology Co., Ltd., Wuhan, China) were confirmed to be mycoplasma-free and authenticated through short tandem repeat (STR) profiling before use. Cells were cultured in RPMI-1640 medium with 10% fetal bovine serum and antibiotics at 37°C, 5% CO2, and ≥95% humidity.
Breast Cancer Mouse Model and Observation of Metastasis
Luciferase-labeled 4T1 cells in the logarithmic growth phase were harvested, washed twice with phosphate buffer saline (PBS), and resuspended at 5×106 cells/mL in sterile PBS. Model group mice (n = 24) received orthotopic injections of 100 μL cell suspension (5 × 105 cells total) into the right fourth mammary fat pad under anesthesia with isoflurane (3% induction, 1.5% maintenance). Control mice (n = 24) were injected with an equal volume of PBS. Tumor dimensions were measured every 3 days using digital calipers, and the tumor volume was calculated using the equation: V = ½ × L × W2, where L is the longest diameter and W is the perpendicular diameter. Model establishment was confirmed when the tumor volume reached 500 mm3 (typically, 10–12 days after inoculation).
For chemotherapy intervention, a randomly selected subset of model mice (n = 6) received intraperitoneal doxorubicin (5 mg/kg; Yeasen, CAS#25316-40-9) once weekly for three consecutive weeks, beginning when tumors reached 200 mm3. Humane endpoints were established according to institutional guidelines, including a tumor burden exceeding 2000 mm3, body weight loss >20%, ulceration, or signs of distress.
Bioluminescence imaging was performed on days 7, 14, 21, and 28 after inoculation using a Xenogen IVIS 200 imaging system (PerkinElmer Inc., Waltham, MA, USA). Mice received intraperitoneal D-luciferin (150 mg/kg; YLKBIO, CAS#115144-35-9) and were imaged 10 min after injection under isoflurane anesthesia. The signal intensity was quantified using Living Image software (version 4.5.2), and the metastatic burden was assessed by counting bioluminescent foci in distant organs. At the end of the experiment (day 28), mice were euthanized via CO2 inhalation followed by cervical dislocation, and the tissues were immediately collected for analysis.
Preparation of Spleen Cell Suspension
Fresh spleens from normal and model groups were placed in small dishes containing PBS supplemented with 1% penicillin–streptomycin double antibiotic and washed several times. Connective tissues were removed, and single-cell suspensions were prepared by gently pressing the spleens through a 200 μm sieve. The cell suspension was centrifuged at 1000 rpm for 5 min, and the pellet was resuspended in PBS. Cell viability was assessed using trypan blue staining. After counting, the cell concentration was adjusted to 2×107 cells/mL and inoculated into culture flasks.
CCK-8 Assay for Evaluating Cell Viability Post TGF-β1 Administration
Splenic cells were seeded in 96-well plates at 2×103 cells per well. Cells were treated with recombinant mouse TGF-β1 (Abbkine Biotechnology Co., Ltd., Wuhan, China) at concentrations of 0, 1, 2, 4, or 8 ng/mL for 48 h. The medium was refreshed every 24 h. Cell viability was assessed using Cell Counting Kit-8 (CCK-8, Abbkine Biotechnology Co., Ltd., Wuhan, China) according to the manufacturer’s instructions. Briefly, 10 μL CCK-8 reagent was added to each well and incubated for 2 h at 37°C. Absorbance was measured at 450 nm using a microplate reader (Molecular Devices Inc., Shanghai, China), and experiments were repeated three times.
siTgfb-R1 Transfection Efficiency Using Quantitative Real-Time Polymerase Chain Reaction
Spleen cells from the normal and model groups were transfected with siTgfb-R1. After 48 h of culture, the cells were harvested, and total RNA was extracted using TRIzol reagent (Yisheng Biotechnology Co., Ltd., Shanghai, China). cDNA was synthesized using reverse transcription, and the expression level of TgfbR1 mRNA was quantified using quantitative real-time polymerase chain reaction (PCR). Each experimental group included three biological replicates.
Colony Formation Assay
The hematopoietic stem cell differentiation capacity was evaluated using methylcellulose-based colony-forming unit assays. Splenic cell suspensions were diluted to 1×106 cells/mL, and 400 μL of the suspension was mixed with 4 mL methylcellulose medium (STEMCELL Technologies, Vancouver, Canada, Cas#9004-67-5). Following thorough mixing and 10-min incubation at 4°C to remove air bubbles, 1.1 mL aliquots were dispensed into 35 mm culture dishes using blunt-end needles. Dishes were placed in a humidified chamber containing a 150 mm dish with 3 mL sterile water to prevent desiccation and incubated at 37°C with 5% CO2 for 10–14 days. Colony-forming unit granulocyte (CFU-G) colonies (clusters containing ≥50 cells with granulocytic morphology) were enumerated using an inverted microscope (Motic China Group Co., Ltd., Xiamen, China) at 40× magnification by two independent observers blinded to the experimental conditions. Each experimental group included three biological replicates.
Flow Cytometry for the Detection of Cell Differentiation
For PMN-MDSC quantification, splenic single-cell suspensions were first blocked with anti-mouse CD16/CD32 antibody (Abcam Biotechnology Co., Ltd., Cambridge, UK) for 10 min at 4°C to prevent non-specific binding. Subsequently, 100 μL of cell suspension was transferred to a 1.5 mL microcentrifuge tube, and CD11b and Ly6G antibodies (5 μL; Abcam Biotechnology Co., Ltd. Cambridge, UK) were added. After incubation at 4°C for 30 min in the dark, the cells were washed twice and then resuspended in 200 μL PBS. Data were collected on a flow cytometer (Agilent Technologies Inc., Santa Clara, CA, USA). At least 50,000 events were acquired per sample. The gating strategy was as follows: FSC-A/SSC-A for singlet discrimination, followed by FSC-H/FSC-W doublet exclusion, then CD11b+ population selection, and final identification of Ly6G+ cells within the CD11b+ gate. Data were analyzed by analysts blinded to the experimental groups using FlowJo software (version 10.8.1). Each experimental group included three biological replicates.
RNA-Seq Correlation Analysis
Comparative transcriptomic analysis was conducted on RNA-seq data to evaluate gene expression profiles between normal and model mouse samples. Pearson correlation coefficients were calculated to generate correlation matrices, facilitating the systematic identification of expression relationships across samples. Differentially expressed genes were subsequently identified using established significance thresholds and expression fold-change criteria. To elucidate the biological importance of transcriptomic alterations, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis was performed on the identified differentially expressed gene sets. The enrichment factor (Rich factor) was computed as the proportion of differentially expressed genes within each pathway relative to the total gene count in that pathway. Significance was determined using the hypergeometric test. False discovery rate correction was implemented to derive adjusted Q-values and ensure robust statistical inference.
Western Blot Analysis
Total cellular proteins were extracted using radioimmunoprecipitation assay lysis buffer, and protein concentrations were determined using the bicinchoninic acid assay. Equal amounts of protein lysates were separated using 8–10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto a polyvinylidene fluoride membrane. The membrane was then placed in an incubation box with a blocking solution containing 5% skim milk. Subsequently, the membrane was incubated overnight at 4°C with diluted primary antibodies under gentle agitation. The membrane was then washed with tris-buffered saline with Tween 20 (TBST) and incubated with a secondary antibody diluted in 5% skim milk-blocking solution. After an additional TBST wash, an enhanced chemiluminescence reagent was added to visualize the protein signals. The results were analyzed using ChemiCapture software (Beijing Saizhi Chemiluminescence Rapid Imaging Software, V3.5.1).
Statistical Analysis
Biostatistical analysis and plotting were conducted in R (version 4.5.0), and statistical analyses were performed using SPSS 25.0 software. Data meeting normality and homogeneity of variance were analyzed using one-way ANOVA for single-factor analysis of variance and Tukey’s test for multiple comparisons among groups. Data meeting normality but deviating from the assumptions of homogeneity of variance were analyzed using Dunnett’s T3 test or the independent sample t-test. Non-normally distributed data were analyzed using the Kruskal–Wallis H-test. A P-value of <0.05 was considered significant.
Results
Heterogeneity of Neutrophil Subpopulations Revealed by scRNA-Seq and stRNA-Seq
Using scRNA-seq, we identified the main immune cell types and their subpopulation distributions in breast cancer tissues. UMAP dimensionality reduction analysis revealed the spatial distribution of different immune cell populations (Figure 1A). In addition, neutrophil subpopulations were further subdivided, indicating their significant heterogeneity (Figure 1B). Mapping of these transcriptomic features onto tumor tissue sections using stRNA-seq localized the neutrophil subpopulations to specific regions of the TME (Figure 1C). Subsequently, differential gene expression analysis was used to identify significantly upregulated and downregulated genes among neutrophil subpopulations (Figure 1D), which were subjected to functional annotation and biological characteristic analysis. Analysis of the complexity of cell communication networks and the diversity of signaling pathways among neutrophil subpopulations revealed significantly higher communication intensity in the N3 subpopulation compared with other subpopulations (Figure 1E). N3 showed the strongest autocrine signal, with the most active intercellular communication observed between N3 and N5 (Figure 1F). The circular string plot showed that the N3 subpopulation exhibits a strong signal interaction (Figure 1G).
 |
Figure 1 Heterogeneity of neutrophil subpopulations in breast cancer tissues. (A) Identification of major immune cell infiltration types and their distribution characteristics in breast cancer tissues using single-cell RNA-sequencing (scRNA-seq). (B) In-depth analysis of neutrophil heterogeneity and subpopulation classification in breast cancer tissues using scRNA-seq. (C) Spatial distribution patterns of specific transcriptomic features of neutrophil subpopulations obtained by mapping them onto tissue sections using spatial transcriptomics. (D) Differentially expressed genes identified between neutrophil subpopulations. (E) Intensity of cell communication networks among neutrophil subpopulations. (F) Number of cell communication networks among neutrophil subpopulations. (G) Visualization model of the cell communication network among neutrophil subpopulations. |
TGF-β1 Expression Significantly Changes During Neutrophil Differentiation
The developmental trajectories of neutrophil subpopulations in two-dimensional space, as determined through pseudo-temporal analysis, are shown in Figure 2A. Their localization onto tumor tissue sections using stRNA-seq is shown in Figure 2B. Using pseudo-temporal analysis, we identified multiple genes with significant expression changes, among which the expression level of Tgfb1 showed a significant upward trend (Figure 2C). A three-dimensional spatial distribution map of Tgfb1 expression was constructed, revealing that areas of low expression were primarily located in the center of the tissue. In contrast, high-expression areas were concentrated at margins and local hotspots (Figure 2D). Analysis of genomic stability differences among neutrophil subpopulations using InferCNV, with the N1 subpopulation as a reference, revealed large-scale genomic structural variations in N2, N3, N4, and N5 subpopulations, indicating significant heterogeneity among neutrophil subpopulations derived from breast cancer tissues (Figure 2E).
 |
Figure 2 Developmental trajectory of neutrophil subpopulations and the expression level of Tgfb1 in breast cancer tissues. (A) Pseudo-temporal trajectory analysis of neutrophil subpopulations (N1, N2, N3, N4, N5) displayed in two-dimensional component space. The color gradient indicates pseudotime progression from early (dark blue) to late (light blue) developmental states, with the black trajectory line connecting sequential differentiation stages among the identified subpopulations. (B) Mapping the developmental trajectory onto tumor tissue sections using stRNA-seq. (C) Key genes with significantly altered expression levels during the pseudo-temporal process. (D) Marking of the Tgfb1 expression region in three-dimensional space. (E) InferCNV analysis of genomic structural variations among neutrophil subpopulations. |
Abnormal Activation of Spleen Neutrophil Differentiation in Breast Cancer Mice is Associated with Disease Progression
The evaluation of the tumor burden and effect of chemotherapeutic intervention on neutrophils revealed significantly higher proportions and absolute counts of neutrophils in the spleen in the model group than in the normal control group. In contrast, doxorubicin treatment reduced these elevated levels, although the levels were higher than those in the normal control group (Figure 3A and B). Pearson correlation analysis demonstrated a significant positive correlation between the neutrophil proportion and the number of metastatic foci in the model group (r = 0.602; Figure 3C). Additionally, tumor weight was significantly negatively correlated with the mean erythrocyte hemoglobin content (r = −0.348; Figure 3D). Flow cytometry analysis revealed a significant shift neutrophil distribution in the model group, characterized by enhanced forward and side-scatter signals, indicating altered cell size and granularity. In contrast, red blood cells and platelets maintained characteristic distributions in both model and normal groups, and the measured parameters remained consistent (Figure 3E).
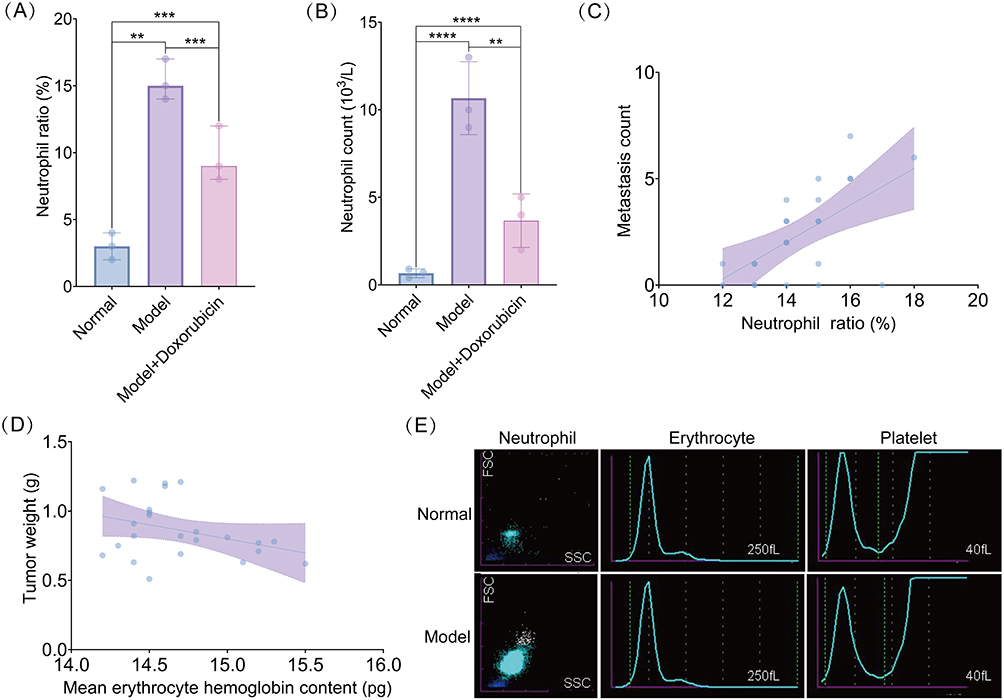 |
Figure 3 Abnormal and active differentiation of neutrophils in the spleen of breast cancer mice, which is associated with disease progression indicators. (A and B) Proportion of neutrophils (A) and count of neutrophils (B) in the control, model, and model + doxorubicin groups. (C and D) Correlation between the proportion of neutrophils and number of metastatic foci (C) and tumor mass and mean corpuscular hemoglobin content (D) in the breast cancer mouse model group. (E) Comparison of the distribution characteristics of neutrophils, red blood cells, and platelets between control and model groups. **P < 0.01, ***P < 0.001, ****P < 0.0001. |
Bidirectional Regulatory Effects of TGF-β1 on Granulocytic Differentiation of Hematopoietic Stem Cells in Normal and Breast Cancer Mice
Comparative analysis revealed significantly elevated Tgfb1 levels in both the splenic organ and serum of the model group compared with those in the normal group (Figure 4A). To determine the optimal concentration of TGF-β1 for in vitro administration, a CCK-8 assay was performed, which showed that the TGF-β1 concentration at 8 ng/mL significantly increased the activity of splenocytes in both normal and model mice compared with that at 0 ng/mL (Figure 4B). CFU-G assays were conducted to assess the granulocytic differentiation capacity. Breast cancer mice exhibited a significantly higher number of CFU-G colonies than normal mice. Furthermore, TGF-β1 administration decreased the CFU-G colony count in the spleens of normal mice but increased the numbers in model mice. Similar effects were observed when tumor serum was administered to normal and model mice (Figure 4C). Flow cytometry analysis showed that TGF-β1 treatment did not affect the proportion of PMN-MDSCs in normal mice (Figure 4D). In contrast, the proportion was increased in model mice (Figure 4E). Furthermore, TGF-β1 significantly decreased the number of CFU-G colonies in normal mice (Figure 4F), with contrasting effects observed in the model mice (Figure 4G).
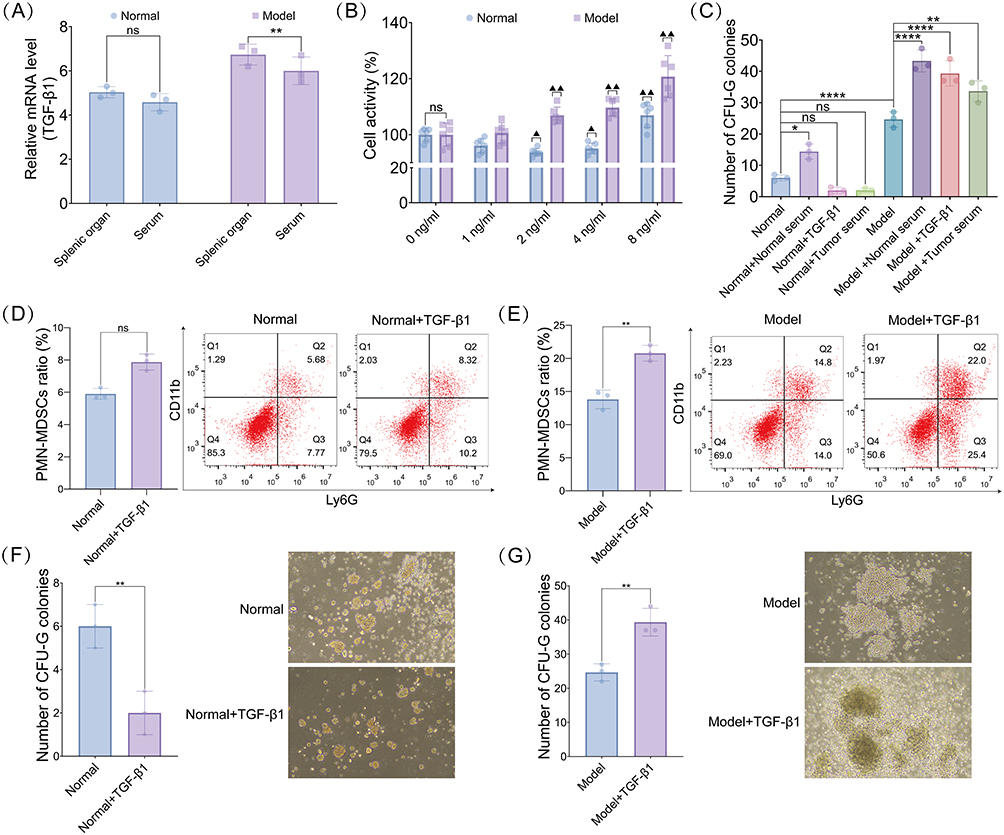 |
Figure 4 Bidirectional regulation of granulocytic differentiation of hematopoietic stem cells by TGF-β1 in normal and breast cancer mice. (A) Tgfb1 mRNA expression in the spleen tissues and serum of normal and model mice. (B) Optimal concentration of TGF-β1 for administration into normal and model mice spleen cells assessed using CCK-8 assay. ▲P < 0.05, ▲▲P < 0.01 vs 0 ng/mL group. (C) Changes in the number of CFU-G colonies formed after normal serum, exogenous TGF-β1, and tumor serum treatment in normal and breast cancer groups.(D and E) Flow cytometry detection of PMN-MDSC differentiation in normal (D) and breast cancer (E) mice. (F and G) Number of CFU-G colonies formed by hematopoietic stem cells in the spleen of normal (F) and breast cancer (G) mice. ns (not significant) P > 0.05, *P < 0.05, **P < 0.01, ****P < 0.0001. |
Tgfb1 Knockdown Inhibits Granulocytic Differentiation of Hematopoietic Stem Cells in Breast Cancer Mice
In normal mouse spleen cells, Tgfb1 knockdown resulted in a significant reduction in Tgfb1 expression (Figure 5A). Subsequent analysis showed that in normal mice, neither Tgfb1 overexpression nor knockdown significantly affected the proportion of PMN-MDSCs in the spleen (Figure 5B and C). However, Tgfb1 overexpression significantly decreased the number of CFU-G colonies in the spleen of normal mice, whereas Tgfb1 knockdown partially reversed this effect (Figure 5D and E). In breast cancer mice, Tgfb1 overexpression significantly increased the proportion of PMN-MDSCs (Figure 5F and G) and the number of CFU-G colonies (Figure 5H and I). In contrast, Tgfb1 knockdown effectively inhibited CFU-G colony formation and granulocytic differentiation ability (Figure 5F and I).
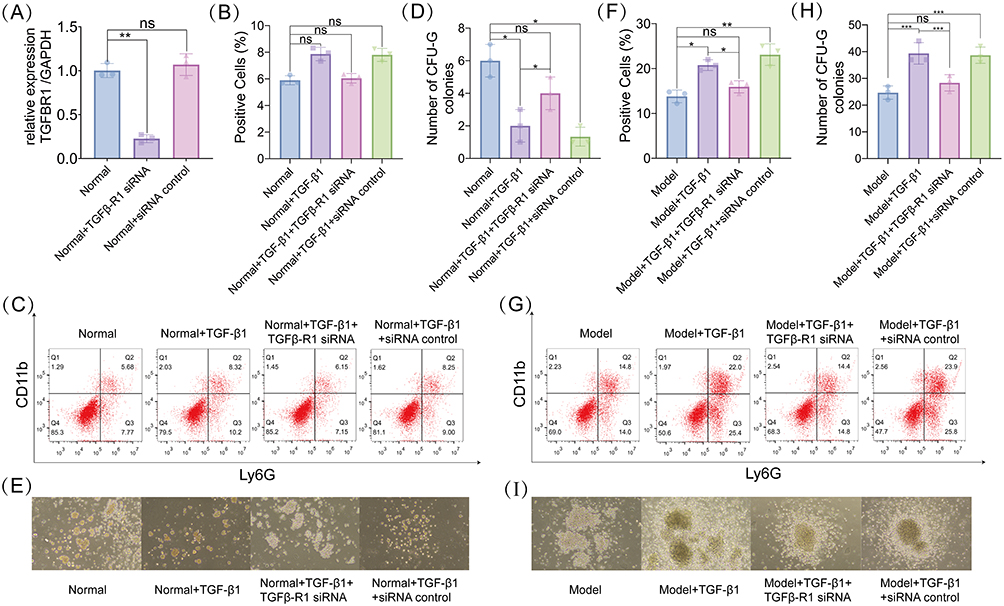 |
Figure 5 Tgfb1 knockdown inhibits granulocytic differentiation of hematopoietic stem cells in breast cancer mice. (A) Quantitative real-time (qPCR) detection of siTgfb-R1 transfection efficiency in splenocytes of mice. (B and C) Proportion of PMN-MDSCs (B) and flow cytometry detection of hematopoietic stem cell differentiation in normal mouse models (C). (D and E) Number of CFU-G colonies formed by hematopoietic stem cells (D) and typical CFU-G colonies of each experimental group (E) in the spleen of normal mouse models. (F and G) Proportion of PMN-MDSCs (F) and flow cytometry detection of hematopoietic stem cell differentiation (G) in the spleen of breast cancer mouse models. (H and I) Number of CFU-G colonies formed by hematopoietic stem cells (H) and typical CFU-G colonies of each experimental group (I) in the spleen of breast cancer mouse models. ns (not significant) P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001. |
Differential Transduction of TGF-β1/SMAD4 Signaling Affects PMN-MDSC Differentiation
RNA-seq with correlation analysis was performed to compare the gene expression levels between normal and model mouse samples. Hierarchical clustering based on log10(FPKM+1) values of differentially expressed genes revealed clear expression patterns, where FPKM represents fragments per kilobase of transcript per million mapped reads, a normalized measure of gene expression that accounts for both transcript length and sequencing depth (Figure 6A). In particular, Tgfb1 expression was significantly upregulated (Figure 6A). Using a threshold Q-value of ≤0.05, we identified the 16 most significantly enriched pathways (Figure 6B), with the SMAD pathway being the most significantly enriched. Western blot analysis further confirmed the regulatory effect of TGF-β1 on SMAD family proteins. In normal mice, Tgfb1 overexpression significantly increased the expression of SMAD2, SMAD3, and SMAD4 in the spleen, whereas Tgfb1 knockdown decreased the expression of the proteins (Figure 6C–F). In contrast, in model mice, Tgfb1 overexpression significantly upregulated SMAD2 and SMAD3 but decreased SMAD4 expression, which was reversed by Tgfb1 knockdown (Figure 6C–F).
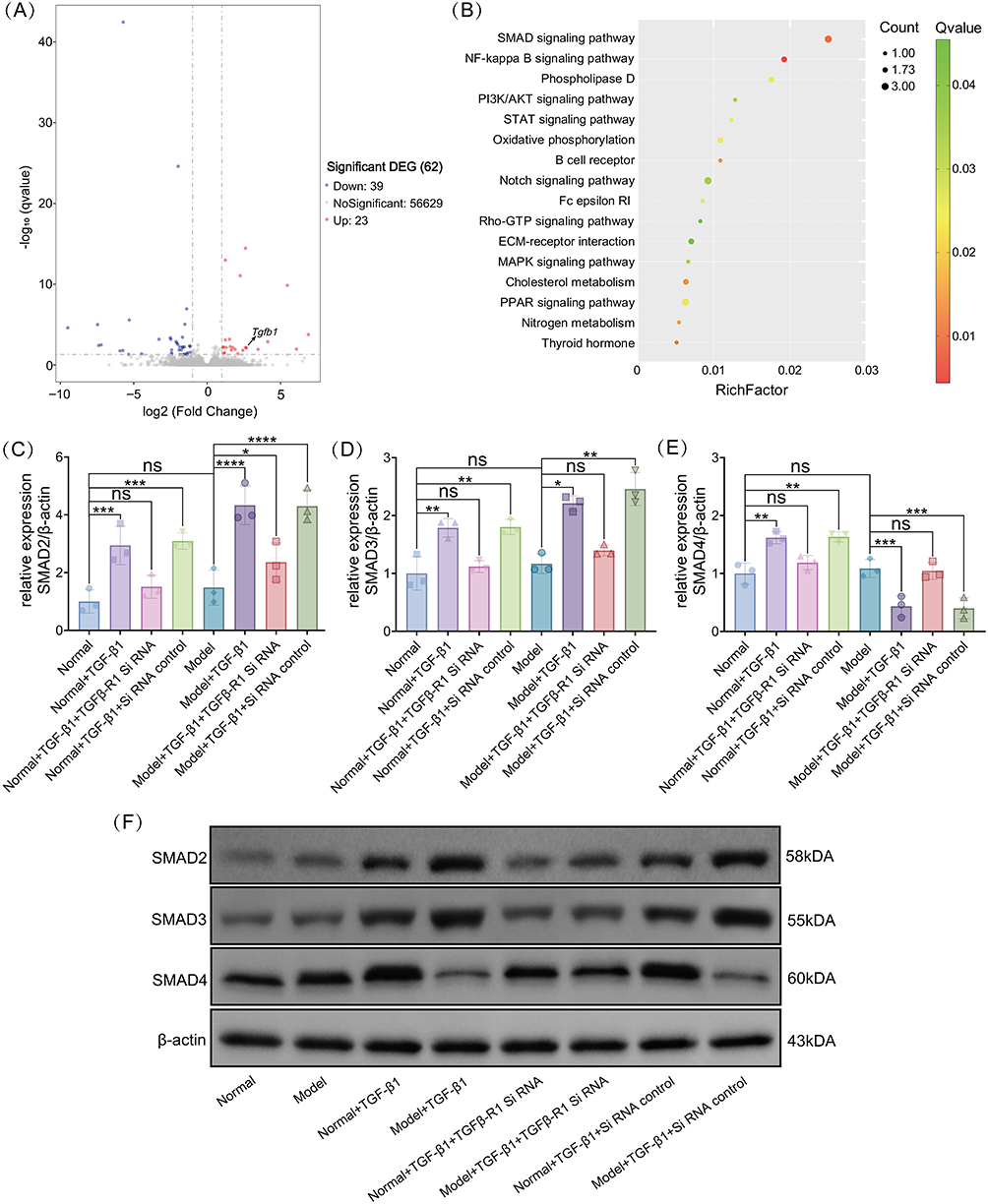 |
Figure 6 Differential transduction of transforming growth factor-β1 (TGF-β1)/SMAD4 signaling affects PMN-MDSCs differentiation. (A) Volcano plot of differentially expressed genes. Red dots represent upregulated genes, blue dots represent downregulated genes, the x-axis represents the fold change in gene expression between different samples, and the y-axis represents the statistical significance of the difference in gene expression. (B) Scatter plot of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment of differentially expressed genes. The y-axis shows the pathway name, and the x-axis represents the enrichment factor (Rich factor), that is, the ratio of the number of differentially expressed genes in a specific pathway to the total number of genes in that pathway. In the gene enrichment directed acyclic graph, the depth of color indicates the degree of enrichment, from deep red (most significant) to light green (least significant). (C–E) Expression levels of SMAD2 (C), SMAD3 (D), and SMAD4 (E) in the spleen of each experimental group in normal and breast cancer mice. (F) Representative Western blot image for SMAD2, SMAD3, and SMAD4 in the spleen of each experimental group in normal and breast cancer mice. ns (not significant) P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. |
Discussion
Neutrophils have a dual function in the TME. As immune effector cells, they can directly kill tumor cells by releasing cytotoxic molecules such as ROS and matrix metalloproteinase 9. Additionally, they induce programmed cell death in tumor cells by upregulating tumor necrosis factor-related apoptosis-inducing ligand and CD95/APO-1 ligand, thereby activating the extrinsic apoptotic signaling pathway.10 Notably, neutrophils in the TME can undergo phenotypic and functional reprogramming and transform into immunosuppressive subsets that secrete various immune suppressive and pro-angiogenic factors to create a tumor-supportive microenvironment, ultimately suppressing antitumor immune responses.11 In this study, we identified five neutrophil subsets (N1–N5), each exhibiting unique transcriptomic characteristics, genomic stability, and cell communication networks. Among these, the N3 subset exhibited significantly higher cell communication intensity and numbers than the other subsets. This finding is consistent with those of a previous study,12 which highlighted the functional heterogeneity of neutrophils in TME, particularly the key role of PMN-MDSCs under different pathophysiological conditions, as an important mechanism in tumor immune escape.
TGF-β1 is a multifunctional cytokine whose function depends on the cellular and environmental context. In normal tissues and early-stage tumors, TGF-β1 mainly exerts tumor-suppressive effects by inducing cell cycle arrest and promoting apoptosis. However, its role shifts during tumor progression, promoting the release of inflammatory mediators, activating macrophages and other inflammatory cells, and driving immune dysfunction, metabolic imbalance, and systemic damage.13 A previous study has shown that TGF-β1 expression is significantly higher in breast cancer tissues than in adjacent normal tissues.14 The study also showed a significant increase in the TGF-β1 concentration in the serum of patients with breast cancer compared with that in healthy individuals.14 This bidirectional regulatory mechanism of TGF-β1 may be related to its dual regulatory function in the hematopoietic system.15 Our findings indicate a significant increase in Tgfb1 expression in the spleen tissue and serum of breast cancer mice, consistent with its role in tumor progression. Furthermore, Tgfb1 overexpression significantly increased the proportion of PMN-MDSCs and the number of CFU-G colonies, confirming its role in promoting granulocytic differentiation in the TME. A previous study has shown that BMP4, a member of the TGF-β superfamily, can effectively block the metastasis of breast tumor cells by inhibiting the activity of MDSCs.16 This finding is consistent with the reduction in CFU-G colony formation and granulocytic differentiation induced by Tgfb1 knockdown observed in the present study. One of the key findings of this study is the observed correlation between the differential transduction of the TGF-β1/SMAD4 signaling pathway in normal and breast cancer mice and PMN-MDSC differentiation. Although these results demonstrate a strong correlation, it has been previously shown that the absence of SMAD4 enhances tumor invasion and metastasis by inhibiting TGF-β1 signaling. SMAD4 (V465M) mutation in patients with breast cancer is closely related to poor prognosis.17 Furthermore, endocrine therapy reduces SMAD4 expression in patients with breast cancer, potentially by promoting therapy resistance via integration of estrogen receptor and erythroblastic oncogene B signaling.18 These previous results provided a new molecular perspective to understand the mechanism underlying breast cancer treatment resistance.
Mechanistic studies demonstrated that the SMAD2/3-SMAD4 protein complex is a key effector molecule in TGF-β1 signal transduction and that disrupting this protein–protein interaction effectively inhibits the activation of the TGF-β1 signaling pathway.19 This highlights a potential therapeutic target for diseases related to abnormal TGF-β1 signaling. Recent evidence suggests that SMAD signaling complexity may involve context-dependent regulation through canonical and non-canonical pathways.20 Targeted therapies against the TGF-β1 signaling pathway have been assessed in clinical trials. For example, Formenti et al21 reported that the combination of TGF-β1 inhibitors and radiotherapy can enhance the antitumor immune response. Mariathasan et al22 showed that blocking TGF-β signaling enhances the efficacy of anti-PD-L1 by modulating T-cell infiltration. Together, these findings suggest that targeting the TGF-β1/SMAD4 signaling pathway is a promising approach for improving therapeutic outcomes in breast cancer, particularly by overcoming immune evasion and therapy resistance mechanisms.
This study has certain limitations. The mouse model system may not entirely reflect the complexity of breast cancer in humans. Although our results demonstrated strong correlations between TGF-β1/SMAD4 signaling and PMN-MDSC differentiation, the precise molecular mechanisms underlying these observations warrant further investigation. Interactions between the TGF-β1/SMAD4 signaling pathway and other cytokine networks remain unclear. Specific markers for neutrophil subpopulations and their validation in clinical samples require further research. Furthermore, the high plasticity of tumor-associated neutrophils presents both a challenge and an opportunity for research and therapeutic development. Future studies should focus on establishing humanized models to facilitate the clinical translation of these findings. Furthermore, a deeper understanding of the plasticity of PMN-MDSCs may pave the way for breakthroughs in precision immunotherapy in breast cancer.
Conclusions
This study uncovered heterogeneity among neutrophil subpopulations within the breast cancer TME and their regulatory mechanisms of differentiation using multi-omics analysis and functional experiments. Notably, our findings indicate that TGF-β1/SMAD4 signaling exerts bidirectional regulation on PMN-MDSC differentiation, promoting granulocytic differentiation and immunosuppressive phenotype under normal physiological conditions, whereas it exerts the opposite effects in the tumor-bearing state. This context-dependent, differential transduction mechanism of TGF-β1/SMAD4 signaling highlights its potential as a therapeutic target for modulating the tumor immune microenvironment. However, further studies, particularly validation in clinical samples and humanized model systems, are needed to fully elucidate the translational implications of these findings and to develop targeted immunotherapy strategies for breast cancer.
Abbreviations
CFU-G, colony-forming unit granulocyte; InferCNV, Inference of Copy Number Variations; KEGG, Kyoto Encyclopedia of Genes and Genomes; PBS, phosphate buffer saline; PMN-MDSC, polymorphonuclear myeloid-derived suppressor cell; ROS, reactive oxygen species; stRNA-seq, spatial transcriptomics; TBST, tris-buffered saline with Tween 20; TGF-β1, transforming growth factor-β1; TME, tumor microenvironment.
Data Sharing Statement
All data generated or analyzed during this study are included in this published article. Additional datasets supporting the conclusions are available from the corresponding author upon reasonable request.
Ethics Statement
This study was approved by the Ethics Committees of Shaoxing Maternity and Child Health Care Hospital, Maternity and Child Health Care Affiliated Hospital, Shaoxing University (No. 2025-075).
Acknowledgments
We would like to express our sincere gratitude to all clinical colleagues and research team members for their tireless efforts. In particular, we would like to express our sincere gratitude to Ms. Jiaqi Shen, the wife of Dr. Jie Cao, for her strong support of our work.
Author Contributions
All authors made a significant contribution to the work reported, whether in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Zhejiang Provincial Science and Technology Program for Traditional Chinese Medicine (2021ZQ014).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kreger J, Roussos Torres ET, MacLean AL. Myeloid-derived suppressor–cell dynamics control outcomes in the metastatic niche. Cancer Immunol Res. 2023;11(5):614–628. doi:10.1158/2326-6066.CIR-22-0617
2. Raskov H, Orhan A, Gaggar S, Gögenur I. Neutrophils and polymorphonuclear myeloid-derived suppressor cells: an emerging battleground in cancer therapy. Oncogenesis. 2022;11:22. doi:10.1038/s41389-022-00398-3
3. Batlle E, Massagué J. Transforming growth factor-β signaling in immunity and cancer. Immunity. 2019;50:924–940. doi:10.1016/j.immuni.2019.03.024
4. Mutiso JM, Gitonga CM, Gicheru MM. TGF-β levels significantly increases in patients with stage III and IV breast cancer and can be explored as a target for tumour diagnosis and staging. Scand J Immunol. 2023;
5. Wang L, Wang H, Zhu M, et al. Platelet-derived TGF-β1 induces functional reprogramming of myeloid-derived suppressor cells in immune thrombocytopenia. Blood. 2024;144:99–112. doi:10.1182/blood.2023022738
6. Giarratana AO, Prendergast CM, Salvatore MM, et al. TGF-β signaling: critical nexus of fibrogenesis and cancer. J Transl Med. 2024;22:594. doi:10.1186/s12967-024-05411-4
7. Blank U, Karlsson S. TGF-β signaling in the control of hematopoietic stem cells. Blood. 2015;125:3542–3550. doi:10.1182/blood-2014-12-618090
8. Lee CR, Lee W, Cho SK, Park SG. Characterization of multiple cytokine combinations and TGF-β on differentiation and functions of myeloid-derived suppressor cells. Int J Mol Sci. 2018;19:869. doi:10.3390/ijms19030869
9. Wang Q, Xiong F, Wu G, et al. SMAD proteins in TGF-β signalling pathway in cancer: regulatory mechanisms and clinical applications. Diagnostics. 2023;13:2769. doi:10.3390/diagnostics13172769
10. Yu X, Li C, Wang Z, et al. Neutrophils in cancer: dual roles through intercellular interactions. Oncogene. 2024;43(16):1163–1177. doi:10.1038/s41388-024-03004-5
11. Wen J, Liu D, Zhu H, Shu K. Microenvironmental regulation of tumor-associated neutrophils in malignant glioma: from mechanism to therapy. J Neuroinflammation. 2024;21(1):226. doi:10.1186/s12974-024-03222-4
12. Hedrick CC, Malanchi I. Neutrophils in cancer: heterogeneous and multifaceted. Nat Rev Immunol. 2022;22:173–187. doi:10.1038/s41577-021-00571-6
13. Li H, Hou M, Zhang P, et al. Wedelolactone suppresses breast cancer growth and metastasis via regulating TGF-β1/Smad signaling pathway. J Pharm Pharmacol. 2024;76:1038–1050. doi:10.1093/jpp/rgae065
14. Gao HC, Huang YZ, Liu YQ, et al. Role of TG2 and TGF‑β1 in the pathogenesis of human breast cancer. Oncol Lett. 2020;20:221. doi:10.3892/ol.2020.12057
15. Gautheron F, Georgievski A, Garrido C, Quéré R. Bone marrow-derived extracellular vesicles carry the TGF-β signal transducer Smad2 to preserve hematopoietic stem cells in mice. Cell Death Discov. 2023;9:117. doi:10.1038/s41420-023-01414-0
16. Cao Y, Slaney CY, Bidwell BN, et al. BMP4 inhibits breast cancer metastasis by blocking myeloid-derived suppressor cell activity. Cancer Res. 2014;74:5091–5102. doi:10.1158/0008-5472.CAN-13-3171
17. Balasundaram A, UK S, TK D, et al. The targeted next-generation sequence revealed SMAD4, AKT1, and TP53 mutations from circulating cell-free DNA of breast cancer and its effect on protein structure–A computational approach. J Biomol Struct Dyn. 2023;41:15584–15597. doi:10.1080/07391102.2023.2191122
18. Li K, Shu D, Li H. Abstract PO2-23-11: SMAD4 depletion contributes to endocrine therapy resistance by ERBB signaling in HR+HER2- breast cancer. Cancer Res. 2024;
19. Nong J, Shen S, Hong F, et al. Verteporfin inhibits TGF-β signaling by disrupting the Smad2/3–Smad4 interaction. Mol Biol Cell. 2024:
20. Massagué J, Sheppard D. TGF-β signaling in health and disease. Cell. 2023;186:4007–4037. doi:10.1016/j.cell.2023.07.036
21. Formenti SC, Hawtin RE, Dixit N, et al. Baseline T cell dysfunction by single cell network profiling in metastatic breast cancer patients. J Immunother Cancer. 2019;7:177. doi:10.1186/s40425-019-0633-x
22. Mariathasan S, Turley SJ, Nickles D, et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 2018;554:544–548. doi:10.1038/nature25501
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Overexpression of TREM1 is Associated with the Immune-Suppressive Microenvironment and Unfavorable Prognosis in Pan-Cancer
Zhou X, Lin K, Fu L, Liu F, Lin H, Chen Y, Zhuang B, Liang H, Deng Q, Wang Z, Chen W, Luo J, Cao J, Li P
Journal of Inflammation Research 2023, 16:1375-1391
Published Date: 27 March 2023
Exploring the Correlation Between GPR176, a Potential Target Gene of Gastric Cancer, and Immune Cell Infiltration
Gu X, Shen H, Xiang Z, Li X, Zhang Y, Zhang R, Su F, Wang Z
Pharmacogenomics and Personalized Medicine 2023, 16:519-535
Published Date: 1 June 2023
Integrating Bulk and Single-Cell RNA Sequencing Reveals Heterogeneity, Tumor Microenvironment, and Immunotherapeutic Efficacy Based on Sialylation-Related Genes in Bladder Cancer
Tan Z, Chen X, Zuo J, Fu S, Wang J, Wang H
Journal of Inflammation Research 2023, 16:3399-3417
Published Date: 14 August 2023
NOTCH1 Mutations Predict Superior Outcomes of Immune Checkpoint Blockade in Non-Small Cell Lung Cancer
Huang Q, Cao H, Yao Q, Zhou X, Li H, Bai Q, Hu H
ImmunoTargets and Therapy 2023, 12:165-173
Published Date: 5 December 2023
Gut Microbiota–Tumor Microenvironment Interactions: Mechanisms and Clinical Implications for Immune Checkpoint Inhibitor Efficacy in Cancer
Said SS, Ibrahim WN
Cancer Management and Research 2025, 17:171-192
Published Date: 25 January 2025