Back to Journals » Drug Design, Development and Therapy » Volume 19
Xenin-Derived Peptides: Multifaceted Regulators and Therapeutic Innovations in Metabolic Diseases
Authors Gong B, Liu X ![]() , Hu G, Li T, Chen F, Sun X, Sun L, Xu Y
, Hu G, Li T, Chen F, Sun X, Sun L, Xu Y
Received 3 September 2025
Accepted for publication 18 December 2025
Published 27 December 2025 Volume 2025:19 Pages 11799—11815
DOI https://doi.org/10.2147/DDDT.S565077
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Binbin Gong,1,2,* Xiyu Liu,1,2,* Guoqiang Hu,1 Tongtong Li,2 Fei Chen,2 Xueqing Sun,2 Lidan Sun,1,2 Yinghe Xu1
1Taizhou Hospital, Zhejiang University, Taizhou, 317000, People’s Republic of China; 2College of Medicine, Jiaxing University, Jiaxing, 314001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lidan Sun; Yinghe Xu, Taizhou Hospital, Zhejiang University, Taizhou, 317000, People’s Republic of China, Tel +86-18358328736, Fax +86-573-83643848, Email [email protected]; [email protected]
Abstract: Xenin, a 25-amino acid peptide hormone predominantly secreted by intestinal K cells, demonstrates evolutionary conservation with neuropeptides such as xenopsin and neurotensin. Functionally, it engages neurotensin receptor 1 (NTSR1) to regulate appetite via hypothalamic signaling pathways and modulates glucose homeostasis through synergistic interactions with incretin hormones. Preclinical studies highlight its dual role in suppressing appetite and enhancing pancreatic β-cell survival, while a single pilot human study suggests xenin-25 may delay gastric emptying and attenuate postprandial glucose excursions; however, these data await independent confirmation. Native xenin, however, is constrained by rapid proteolytic degradation and limited bioavailability. Advances in peptide engineering, including C-terminal truncation, site-directed amino acid substitution, and lipidation, have generated analogues that exhibit prolonged metabolic activity in rodent models, with plasma half-life extended from minutes to hours. In murine models of metabolic dysfunction, these derivatives enhance insulin secretion, improve glycaemic profiles and restore incretin responsiveness. Furthermore, multi-agonist peptides combining xenin with other gastrointestinal hormones show synergistic potential in preclinical studies, concurrently augmenting insulin secretion and reducing energy intake, though their clinical relevance remains to be validated in human trials. Despite promising preclinical outcomes, challenges persist in translating xenin-based therapies to clinical practice, including incomplete mechanistic insights into receptor cross-talk and species-specific variations in gastrointestinal responses. This review uniquely integrates the preclinical landscape of xenin biology, peptide-engineering principles, and emerging multi-agonist design, identifying knowledge gaps critical for future translation. We conclude that xenin-based therapeutics are a promising yet early-stage strategy whose efficacy and safety in human metabolic diseases remain to be established through rigorous pharmacokinetic profiling and phased clinical trials.
keywords: Xenin, type 2 diabetes, obesity, physiological function, structural modification
Introduction
Xenin is a 25-amino acid peptide hormone primarily secreted by enteroendocrine K cells in the gastrointestinal tract, particularly in the gastric and duodenal mucosa.1,2 Its secretion is closely associated with fasting and postprandial states, with plasma levels peaking during the migratory motor complex Phase III in humans.3 The species, in proteins of which the xenin motifs were identified with a highest degree of homology to human xenin. Motifs with 40% homology to human xenin are already present in prokaryotes. Homology reaches 84–96% in single-cell algae and plants, becoming complete since bony fishes (see Table 1).4
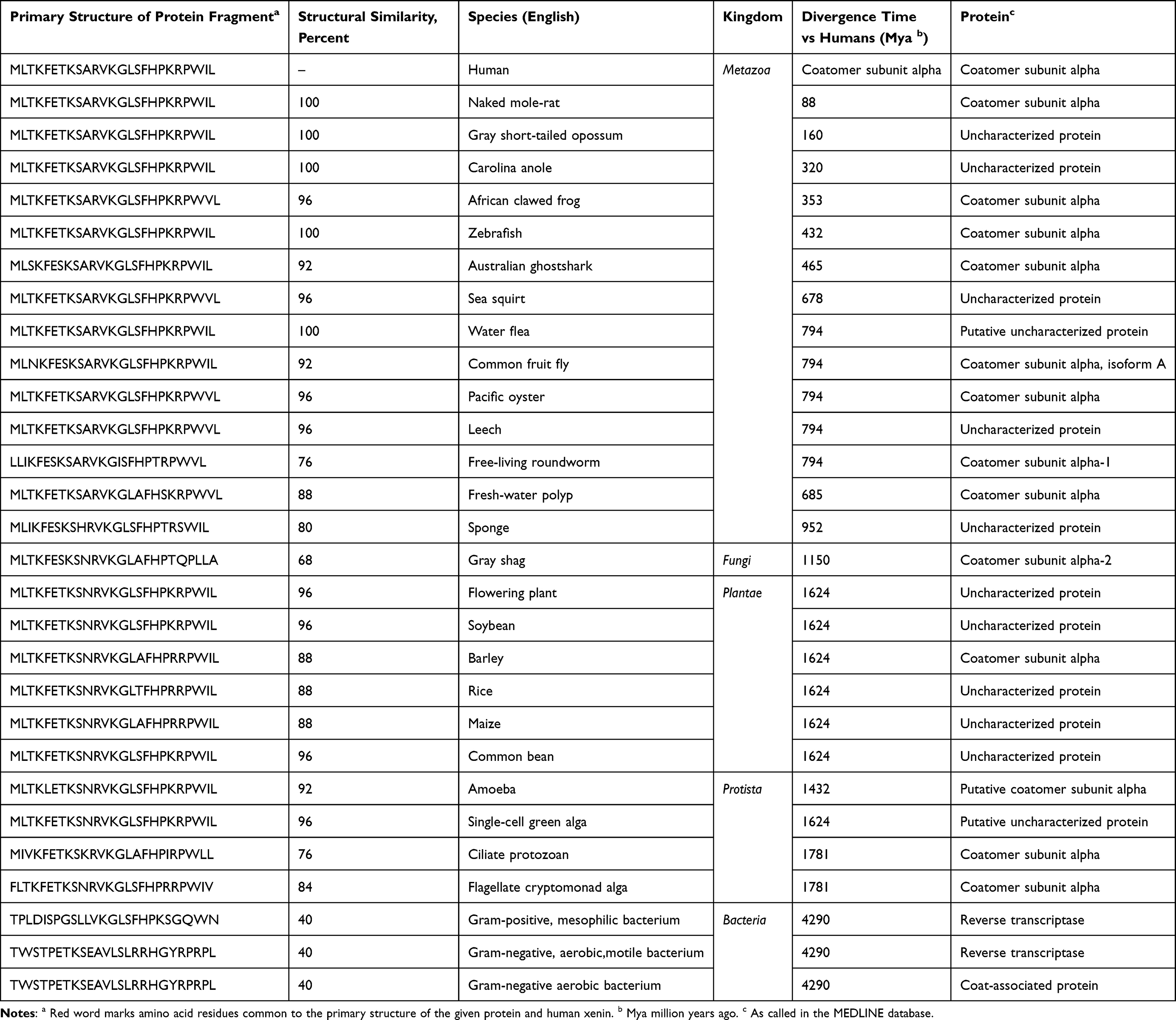 |
Table 1 Xenin Motifs in the Primary Structure of Proteins: From Bacteria to Human |
Xenin exhibits structural homology with neurotensin and amphibian xenopsin, sharing evolutionary conservation across vertebrates, and is derived from the proteolytic cleavage of its precursor, proxenin, which is homologous to cytoplasmic coatomer protein subunit α.4 Functionally, xenin modulates metabolic homeostasis through interactions with NTSR1, influencing appetite regulation, insulin secretion, and gastrointestinal motility (see Figure 1).5–7 In preclinical models, xenin enhances glucose-stimulated insulin release, suppresses glucagon secretion, and delays gastric emptying, positioning it as a potential therapeutic agent for obesity and type 2 diabetes mellitus (T2DM).8
 |
Figure 1 Production process and action sites of human xenin-25. |
 |
Figure 7 Sequences of Exendin-4/Gastrin/Xenin-8-Gln. |
However, the clinical translation of native xenin is hindered by rapid degradation via nonspecific plasma proteases and renal clearance, resulting in a short half-life of approximately 5–10 minutes.9 Unlike incretins such as Glucagon-Like Peptide-1 (GLP-1), xenin is not a substrate for dipeptidyl peptidase-IV (DPP-IV), but its instability necessitates structural modifications to improve pharmacokinetic profiles. Current strategies include amino acid substitutions (eg, xenin-25-Gln),10 lipidation (eg, xenin-25[Lys13PAL]),11 and hybrid peptide design (eg, GLP-1/xenin-8-Gln),12 which enhance metabolic stability and receptor specificity. These engineered analogues demonstrate prolonged activity in regulating glucose homeostasis and β-cell survival through PI3K/Akt signaling pathways in diabetic models.13,14 Unlike marketed GLP-1 receptor agonists (eg, liraglutide or semaglutide) or the dual GIP/GLP-1 co-agonist tirzepatide, xenin-based peptides have not yet progressed beyond pre-clinical studies; however, their unique ability to restore GIP sensitivity and activate NTSR1-dependent neuronal relays may offer mechanistic advantages that current incretin mimetics do not provide.
Recent advances also highlight the potential of multi-agonist peptides combining xenin with other gastrointestinal hormones, which synergistically target insulin sensitivity, appetite suppression, and lipid metabolism.15,16 Despite these innovations, challenges remain, including species-specific variations in gastrointestinal responses,17 incomplete understanding of receptor cross-talk mechanisms,18 and the need for rigorous clinical validation of long-term safety.19 Although scattered reports have highlighted xenin’s insulinotropic or anorexigenic effects, a systematic synthesis that integrates its evolutionary biology, structure–activity advances, and emerging multi-agonist engineering is still lacking. The present review fills this gap by providing the first comprehensive map of xenin-based peptide optimization and by explicitly delineating the pre-clinical–to–clinical translational hurdles that must be overcome before xenin hybrids can be positioned alongside approved incretin therapies. This review comprehensively examines xenin’s physiological roles, structural optimization strategies, and therapeutic potential, while proposing future directions to accelerate its development as a multi-target agent for metabolic disorders.
Physiological Function of Xenin
Xenin, an evolutionarily conserved gut hormone, was initially characterized for its regulatory role in modulating feeding behavior in piscine species.4,20 Xenin has been shown to exert significant effects on various physiological processes (see Figure 2), including insulin secretion, appetite control, and gastrointestinal motility.21 Its therapeutic potential in metabolic diseases such as diabetes and obesity has been extensively explored in both preclinical and clinical studies. This section provides an in-depth review of the diverse physiological functions of xenin, highlighting its mechanisms of action and potential therapeutic applications. We will discuss its roles in diabetes management, obesity treatment, appetite suppression, pancreatic function, polycystic ovary syndrome (PCOS), and other relevant physiological processes.22–25
 |
Figure 2 Physiological function of xenin-25. |
Diabetes
Xenin and its related peptides have garnered significant attention for their potential therapeutic applications in diabetes management. Accumulating evidence indicates that xenin modulates insulin secretion through multiple pathways, thereby exerting beneficial effects on glycemic control. For instance, Hasib et al (2018) demonstrated that xenin-25 significantly augments insulin secretion and improves glucose tolerance in high-fat diet-induced diabetic mice.10 In a 2023 study by Kucukbas GN et al, maternal serum levels of xenin-25 were assessed in women with gestational diabetes mellitus (GDM), revealing significant differences between normoglycemic and GDM pregnancies. These findings suggest that xenin-25 may play a role in the metabolic regulation of GDM.26 In a 2016 study by Karin Sterl et al, the metabolic effects of xenin-25 were investigated in healthy individuals and patients who underwent Roux-en-Y gastric bypass (RYGB). The findings revealed that xenin-25 induces diarrhea and suppresses GLP-1 release in healthy subjects, while its mechanisms of action and neural pathways differ markedly in RYGB patients.27
In type 2 diabetes, the impaired incretin effect, a key pathophysiological feature,28 mainly results from two factors: reduced secretion of glucagon-like peptide-1 (GLP-1) and defective insulinotropic action of glucose-dependent Insulinotropic Peptide (GIP), a sister incretin hormone to GLP-1.29 GLP-1 is predominantly produced by L cells in the distal bowel, and GIP is mainly produced by K cells in the proximal intestine. After eating, both peptides are immediately released into the blood and enhance glucose-stimulated insulin secretion (GSIS).30 Unlike GLP-1, which can stimulate insulin secretion in T2DM, persons with T2DM are thought to be resistant to the actions of GIP,31,32 so increasing GIP signaling has not been pursued as a therapeutic target for T2DM. Current GLP-1 mimetics only address the reduced GLP-1 secretion,33 with no impact on the compromised GIP bioactivity, which may explain the less-than-expected benefits in humans. However, xenin is known to enhance the insulinotropic action of GIP,13,34–36 offering a new possibility for improving the incretin effect in type 2 diabetes patients.
Additionally, hybrid peptides incorporating xenin, such as Exendin-4/xenin-8-Gln, have shown enhanced efficacy in diabetes treatment. These peptides not only potentiate insulin secretion but also restore sensitivity to GIP, thereby further optimizing glucose metabolism.37 And GIP plus xenin increases cholinergic input to islets equally well in humans with normal glucose tolerance (NGT), impaired glucose tolerance (IGT), and T2DM. Xenin potentiation of GIP is indirect and requires an intra-pancreatic neuronal relay. Antibody staining and single-nucleus RNA-seq show that NTSR1 is abundant on cholinergic neurons innervating mouse and human islets, but is absent from β-cells themselves.18,38 Xenin binding activates NTSR1→Gαq/11→PLCβ2→IP3-Ca2⁺ signalling in these neurons, evoking local acetylcholine (ACh) release. ACh then acts on β-cell M3 muscarinic receptors (CHRM3), amplifying cAMP generated by GIP-occupied GIPR and thereby enhancing glucose-dependent insulin exocytosis. Consistent with this circuit, blocking nicotinic or muscarinic transmission abolishes the xenin + GIP synergism in perfused mouse pancreas.35 Thus, xenin converts a paracrine GIP signal into a neuro-hormonal amplification loop, a mechanism not shared by GLP-1 or GLP-1/GIP co-agonists.39 The multifaceted mechanisms of action of xenin position it as a promising candidate for the development of novel antidiabetic therapies.
In addition, elevated serum xenin-25 has been reported in women with polycystic ovary syndrome, a finding that aligns with its association with insulin resistance in preclinical models;40 however, dedicated human intervention studies are absent.
Obesity
Xenin’s role in obesity management is equally noteworthy, given its capacity to modulate appetite and energy metabolism, which are pivotal in weight regulation. Research has shown that central administration of xenin significantly reduces food intake and body weight in obese mice, highlighting its potential as an anti-obesity agent.25 Furthermore, xenin has been shown to activate lipolysis in white adipose tissue, thereby reducing fat accumulation.41 A study by Onaga et al reported that xenin-25 evokes rumen motility in conscious sheep. While this highlights a conserved role in gut motility, the ruminant forestomach lacks an anatomical equivalent in monogastric mammals; thus, the finding serves primarily as proof-of-concept for xenin-induced smooth-muscle contraction and cannot be extrapolated to appetite regulation or energy intake in humans. Comparative studies using pig or dog models with a stomach anatomy closer to humans are still missing.42
In 2021, a study by Onaga et al first identified the complete sequence of ovine xenin and its mRNA expression profile, revealing that this sequence exhibits high homology (approximately 90%) with bovine xenin. Immunohistochemical analyses confirmed that xenin-positive cells are predominantly localized in the pyloric antrum region, while expression was also detected in the forestomach compartments (rumen, reticulum, and omasum), which are anatomically unique to ruminants. These findings provide critical anatomical insights for subsequent investigations into the mechanistic role of xenin in the gut-brain axis-mediated regulation of lipid metabolism in ruminants.43 This finding offers new insights into the role of xenin in the digestive system. These findings underscore the therapeutic potential of xenin in addressing obesity and related metabolic disorders.
The anorexigenic effects of xenin have been well-documented across various studies. Central administration of xenin has been shown to induce significant reductions in food intake through activation of specific neural pathways. For example, Kim et al (2014) demonstrated that xenin suppresses feeding via the hypothalamic IL-1β-IL-1RI signaling pathway.44 Regarding its appetite suppression effects, a 2023 study by Saito et al demonstrated that centrally administered xenin activates nesfatin-1 neurons in the rat hypothalamus, as evidenced by increased Fos expression, suggesting its involvement in feeding regulation through modulation of multiple neural circuits.45 In contrast, van de Sande-Lee et al (2013) found that while xenin reduces food intake via pathways independent of leptin and melanocortin systems, cerebrospinal fluid (CSF) xenin levels in obese individuals did not exhibit significant obesity-related changes during weight loss interventions.46 Additionally, xenin’s interaction with the brain regions responsible for appetite regulation suggests that it may offer a novel therapeutic approach for managing overeating and promoting satiety.7
Other Physiological Functions
Beyond its roles in diabetes, obesity, and appetite regulation, xenin exhibits a range of additional physiological functions. For example, xenin has been shown to modulate gastrointestinal motility, delay gastric emptying, and reduce postprandial glucose levels.8 In the context of metabolic responses in healthy individuals, xenin-25 exhibits significant physiological effects. A 2019 study by Kuwahara et al, utilizing Ussing chamber experiments, demonstrated that xenin-25 enhances anion secretion by activating non-cholinergic secretomotor neurons in the rat ileum.47 This finding suggests a potential critical role for xenin-25 in modulating intestinal function and electrolyte homeostasis. These effects may contribute to its overall metabolic regulatory actions.
Xenin exerts notable effects on pancreatic function, particularly in the context of insulin and glucagon secretion. Studies have shown that xenin enhances the secretion of pancreatic polypeptides through cholinergic signaling pathways.18 For instance, xenin-25 has been found to amplify the actions of GLP-1 on insulin and glucagon secretion, although this effect appears to be attenuated in individuals with type 2 diabetes.18 Furthermore, xenin’s influence on pancreatic exocrine function, such as the promotion of pancreatic fluid, protein, and bicarbonate output, has been demonstrated in sheep models.5 These findings collectively highlight xenin’s pivotal role in modulating pancreatic function and its potential applications in the treatment of pancreatic disorders.
Additionally, xenin’s influence on neurotensinergic pathways and its potential to enhance cognitive function in high-fat diet-induced neuroinflammation highlight its diverse therapeutic potential.48 A 2018 study by Khan et al demonstrated that xenin-25 potentiates GIP action via cholinergic neural signaling pathways, thereby stimulating insulin secretion in both murine and human pancreatic β-cells. This mechanism contrasts with the direct receptor-mediated effects of neurotensin (NT), suggesting that xenin’s regulatory influence relies on neuro-islet cross-communication.38 Moreover, in a 2014 study by Arslan et al, serum levels of xenin and ghrelin were measured in obese children, including those with comorbid non-alcoholic fatty liver disease (NAFLD) and insulin resistance. The study found significantly reduced serum xenin levels in these patients and demonstrated a positive correlation between xenin and ghrelin levels. These findings provided critical clinical evidence supporting the potential role of xenin in metabolic disorders such as obesity and NAFLD.49 Further exploration of these multifaceted functions of xenin is warranted to fully elucidate its potential applications in various clinical contexts.
Structural Modification of Xenin
The structural modification of xenin has emerged as a pivotal strategy to overcome its inherent pharmacokinetic limitations and amplify its metabolic regulatory effects. By engineering site-specific alterations, researchers have significantly enhanced proteolytic stability, receptor specificity, and therapeutic efficacy (see Figure 3). For instance, substitution of lysine and arginine residues with glutamine in xenin-25 (xenin-25-Gln) not only preserved insulinotropic activity but also reduced susceptibility to nonspecific protease degradation, achieving a 2.3-fold improvement in hypoglycemic potency.10 Hybrid peptide design, such as the GLP-1/xenin-8-Gln chimera, synergistically activates both GLP-1 and neurotensin receptors, restoring glucose-dependent insulin secretion and normalizing GIP responsiveness in diabetic models.37 Additionally, lipidation strategies (eg, xenin-25[Lys13PAL]) extend plasma half-life from minutes to hours through albumin binding, enabling dual peripheral and central metabolic modulation.9 These advancements underscore xenin’s versatility as a scaffold for next-generation therapies targeting β-cell dysfunction and insulin resistance.
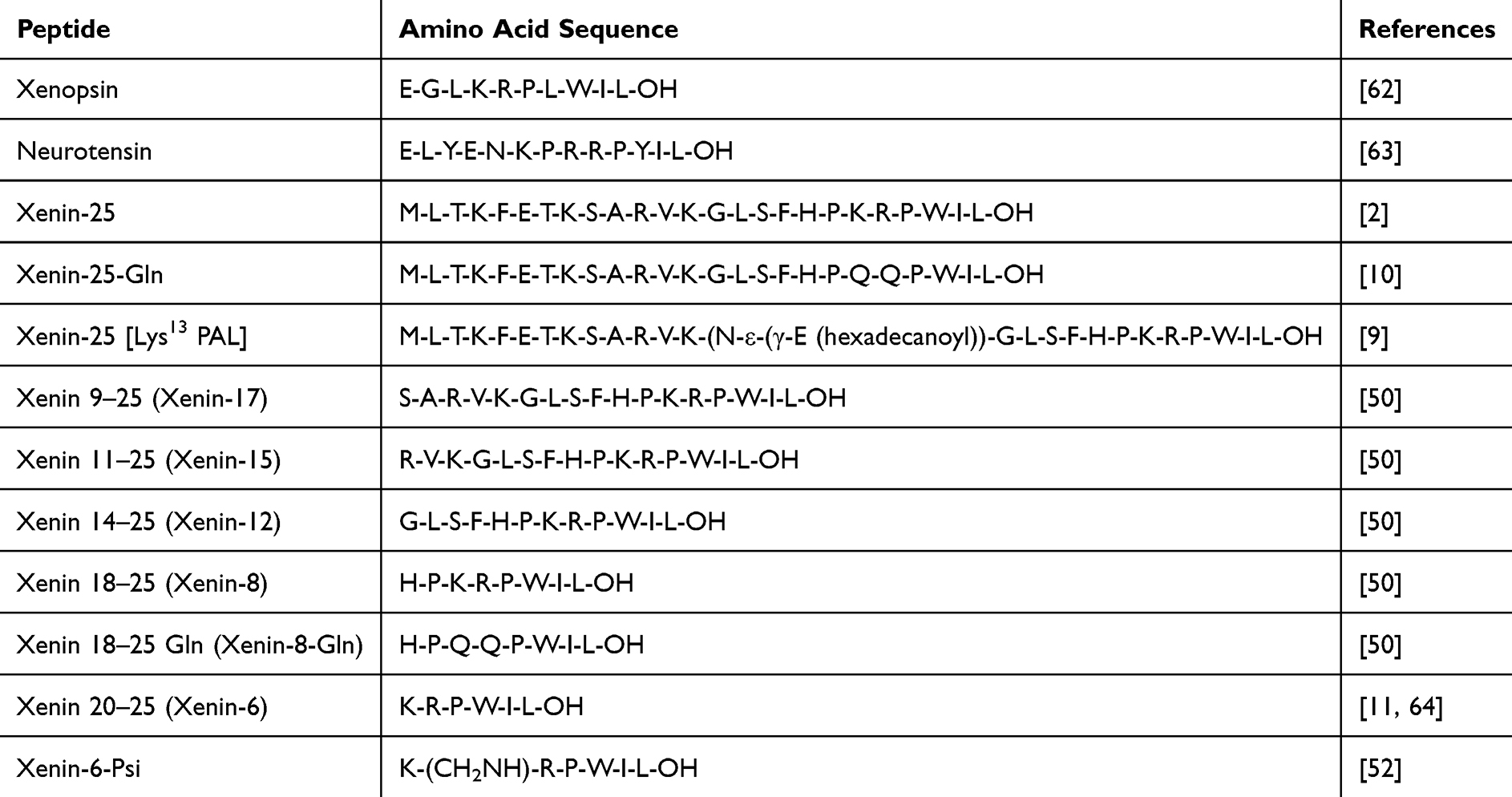 |
Table 2 Amino Acid Sequences of Xenin‐25 as Well as Its Related Stable Analogues and Naturally Occurring Fragment Peptides |
 |
Figure 3 Structural Modification of xenin-25. |
Fragmentation Modification
Proteolytic fragmentation of xenin has yielded bioactive peptide derivatives, notably xenin-8 (residues 18–25), which retain core insulinotropic activity while exhibiting enhanced metabolic stability compared to the full-length peptide (see Table 2). Studies demonstrate that xenin-8 stimulates glucose-dependent insulin secretion via partial agonism of NTSR1, with comparable efficacy to xenin-25 in pancreatic β-cell models.50,51 This C-terminal octapeptide (HPKRPVIL) is critical for receptor binding, as evidenced by structure-activity relationship analyses showing that truncation beyond residue 18 abolishes bioactivity.50 The fragmentation strategy not only delineates essential pharmacophores but also enables the development of target-selective therapeutics with reduced off-target effects.
In 2020, a study by Sarah L. Craig et al revealed that co-administration of Ψ-xenin-6 with sitagliptin enhanced anti-diabetic effects in high-fat high-fructose diet-fed (HFF) mice, although it failed to significantly improve glucose tolerance.52 In 2021, the Craig team further demonstrated that the MetAP2 inhibitor TNP-470 augmented the anti-diabetic efficacy of sitagliptin by enhancing insulin sensitivity and glucose tolerance.53
Despite these advantages, translational progress is hindered by insufficient pharmacokinetic characterization. Xenin-8 exhibits rapid plasma clearance (t1/2 <20 min) due to renal excretion and neutral endopeptidase (NEP)-mediated degradation, limiting its therapeutic utility.9 Current efforts focus on engineering stabilized analogs through lipid conjugation and D-amino acid substitutions to prolong systemic exposure.
Amino Acid Substitution
Amino acid substitution represents a cornerstone of rational peptide engineering, enabling precise modulation of xenin’s physicochemical properties and receptor interactions to optimize therapeutic potential. This technique involves the systematic replacement of specific residues to enhance proteolytic stability, receptor binding affinity, and metabolic activity while minimizing off-target effects.10 The complexity of this approach lies in preserving xenin’s core pharmacophore—particularly its C-terminal hexapeptide (residues 20–25), which is critical for NTSR1 activation—while introducing substitutions that mitigate enzymatic degradation and improve pharmacokinetics.10
Amino-acid substitution enhances proteolytic stability while preserving the NTSR1-pharmacophore (residues 20–25). Conservative Glu→Gln or Lys→Ala exchanges prolong t½ 2-3-fold in mouse plasma, whereas non-conservative hydrophobic insertions at position 4 reduce receptor affinity. Systematic scanning of non-natural residues is ongoing, but rodent-scale structure-immunogenicity data are still lacking.50 Structural super-position of the neurotensin–NTSR1 cryo-EM complex54 indicates that the xenin C-terminal hexapeptide (HPKRPW, residues 20–25) adopts an equivalent β-turn and inserts R23 and W26 into the same orthosteric pocket, triggering the canonical Gαq/11–PLCβ–IP3–Ca2⁺ cascade. Functional truncation studies show that deletion beyond residue 20 abolishes inositol phosphate production (>100-fold IC50 shift) in CHO-NTSR1 cells,38 confirming this motif as the minimal pharmacophore.
Despite these advances, key limitations persist. Non-conservative substitutions (eg, hydrophobic residues at position 4) may disrupt xenin’s α-helical structure, attenuating receptor binding.9 Furthermore, systemic studies on substitution-induced immunogenicity and tissue-specific biodistribution remain sparse. Emerging strategies, such as D-amino acid incorporation (eg, D-Ala4) and non-canonical residue substitutions (eg, β-homoarginine), show promise in evading enzymatic recognition while maintaining bioactivity.9 Combinatorial libraries and machine learning-guided design are now being leveraged to predict optimal substitution patterns, accelerating the development of xenin analogs with tailored pharmacokinetic-pharmacodynamic profiles.
Acylation
Acylation modifications, particularly palmitoylation of xenin, yield derivatives such as xenin-25 [Lys13PAL], which have been demonstrated to significantly enhance the peptide’s stability and prolong its duration of action.9 Specifically, xenin-25 [Lys13PAL] has exhibited robust hypoglycemic and insulinotropic effects in murine models of diet-induced diabetes.9 Additional acylated analogues include myristoyl (C14) and cholesteryl (C27) conjugates at Lys13 or Lys24, which extend mouse plasma t½ to 6.8 h and 11.3 h, respectively, and improve glucose-lowering by 1.7- and 2.1-fold vs native xenin-259,50 However, increased lipophilicity raised injection-site irritation and plasma protein binding (> 98%), issues that have not yet been solved.However, acylation may also induce alterations in the spatial configuration of xenin and its receptor binding dynamics. Therefore, further investigation is warranted to comprehensively elucidate the metabolic pathways and mechanisms of action of acylated xenin in vivo. Such insights will be crucial for optimizing its therapeutic efficacy and safety profile.
Fusion Peptide
The development of fusion peptides by hybridizing xenin with other bioactive peptides or proteins represents a promising strategy to amplify its therapeutic potential in metabolic diseases. This approach leverages the complementary actions of distinct peptides, yielding multifunctional molecules with superior efficacy compared to native xenin or its isolated analogs.
Xenin’s short half-life and limited receptor specificity have driven efforts to fuse it with other peptides. These fusion approaches aim to extend plasma stability, target additional pathways, and enhance tissue selectivity. Xenin fusion with hormones like GLP-1 or exendin-4 can prolong its stability in circulation. When combined with neurotensin, it may unlock new pathways for β-cell protection. Additionally, integration with GIP can improve tissue selectivity, helping to restore insulin sensitivity.
GLP-1/xenin Hybrids the combine incretin effects of GLP-1, which promotes glucose-dependent insulin secretion, with xenin’s β-cell proliferative properties. Preclinical studies have demonstrated improved glycemic control and β-cell mass preservation in diabetic models.37 Similarly, the Acetyleneurotensin /xenin-8-Gln Fusion (Ac-NT/XN-8-Gln) leverages neurotensin’s anti-apoptotic and anti-inflammatory effects alongside xenin’s metabolic actions. In high-fat-diet mice, Ac-NT/XN8--Gln significantly enhanced insulin secretion and reduced hyperglycemia, outperforming individual peptides.14 Additionally, the (DAla2)GIP/xenin-8-Gln fusion peptide, which combines a degradation-resistant GIP analog with xenin, has been shown to restore GIP receptor sensitivity and amplify β-cell function. This suggests potential utility in type 2 diabetes patients with GIP resistance.55
At present, the structural modification of xenin mainly includes the four methods described above (see Figure 4). These include fragmentation, amino acid substitution, acylation, and fusion peptides. Fragmentation produces bioactive derivatives like xenin-8, while substitution optimizes xenin’s properties. Acylation enhances stability, and fusion peptides combine xenin with other peptides to improve efficacy. However, challenges such as structural compatibility and pharmacokinetic studies persist.
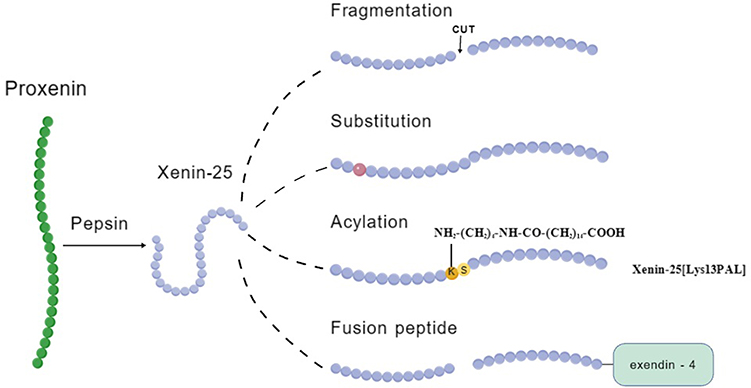 |
Figure 4 Structural modification on xenin. |
Fragmentation (xenin-8) raises t½ from 5 min to 18 min but loses the N-terminal domain required for β-cell proliferation signalling. Acylation gives the longest half-life (> 6 h) yet increases lipophilicity-related adverse events. Hybridisation combines two pharmacologies in one molecule but adds manufacturing complexity and potential immunogenicity. Thus, no single platform is superior; the choice depends on whether the intended indication demands rapid clearance (fragment), once-weekly injection (acylated), or multi-receptor synergy (hybrid).
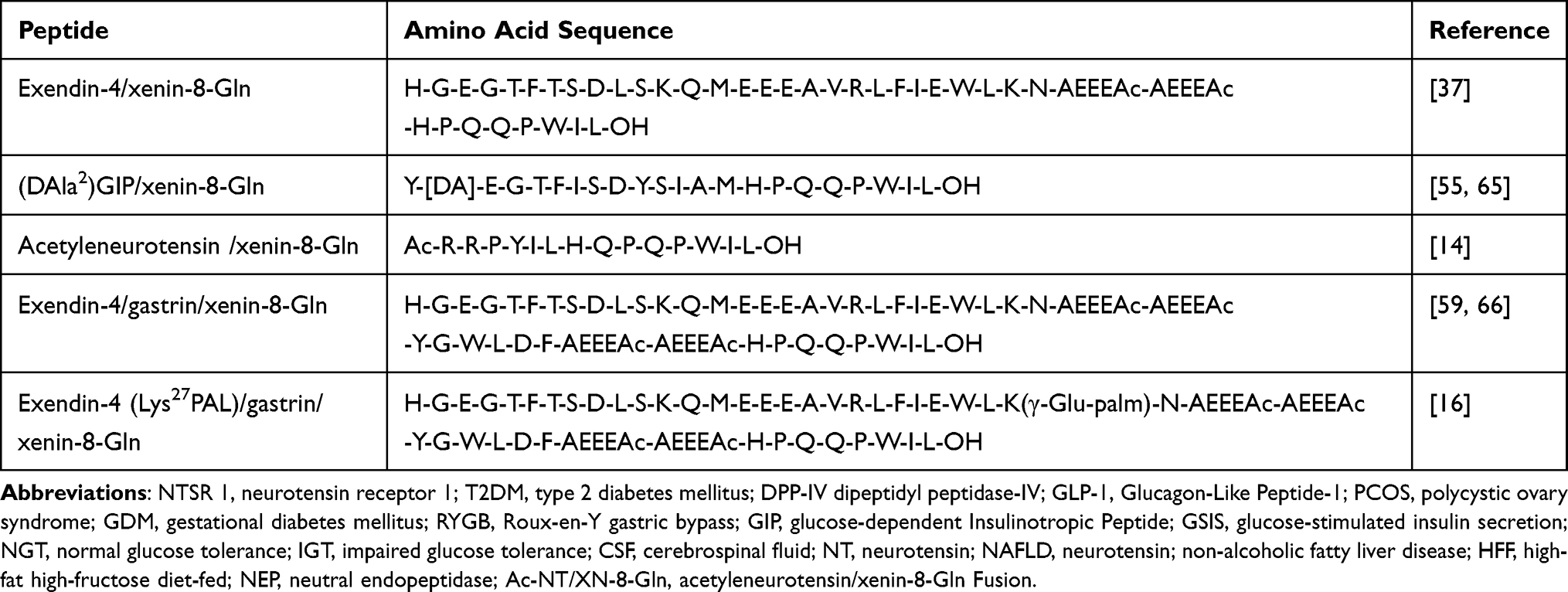 |
Table 3 Amino Acid Sequences of Xenin Incorporated Multi-Acting Hybrid Peptides |
Fusion of Xenin with Other Gastrointestinal Hormones
Current therapeutic strategies leveraging xenin demonstrate promising metabolic benefits, yet face limitations in durability and pathway selectivity. Building on the success of unimolecular multi-agonists like tirzepatide (GLP-1/GIP co-agonist), recent advances highlight xenin’s unique potential as a scaffold for hybrid peptides targeting complementary metabolic pathways (see Table 3). By integrating xenin with incretins (eg, GLP-1, GIP) or neuropeptides (eg, neurotensin) (see Figure 5), these co-agonists address multifactorial defects in diabetes pathogenesis—enhancing insulin secretion, suppressing glucagon, and modulating energy homeostasis through convergent receptor signaling (see Figure 6).13,14
 |
Figure 5 Sequences of Xenin, GLP-1, GIP, Neurotensin, and Gastrin. |
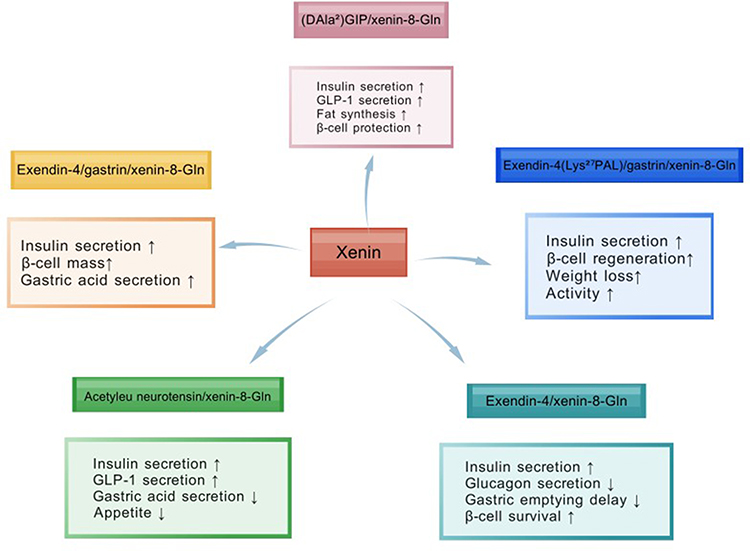 |
Figure 6 Synergistic effects of xenin with different receptor agonists. |
Glucagon-Like Peptide-1 (GLP-1)
GLP-1 is a crucial incretin hormone known to stimulate insulin secretion, suppress appetite, and decelerate gastric emptying, thereby demonstrating substantial efficacy in diabetes treatment.56 The amalgamation of xenin with GLP-1 is designed to combine the strengths of both, bolstering the regulation of blood glucose and body weight. For instance, exendin-4/xenin-8-Gln is a widely-studied fusion peptide of xenin and the GLP-1 analogue, exendin-4.12,37 Research has revealed that this fusion peptide exhibits pronounced antidiabetic effects in high-fat diet mice, markedly diminishing blood glucose levels, enhancing insulin sensitivity, and reestablishing the sensitivity of GIP.37 Its mechanism of action possibly operates through the concurrent activation of GLP-1 receptors and xenin-related signalling pathways, thereby promoting insulin secretion, amplifying insulin action, and modulating appetite and energy metabolism.12 However, there is still a need for further research regarding the differences in the efficacy of this fusion peptide across various diabetic subtypes, as well as potential safety concerns associated with its long-term use.
Glucose-Dependent Insulinotropic Peptide (GIP)
GIP is a crucial incretin hormone that stimulates insulin secretion postprandially.57 Investigating the fusion of xenin with GIP aims to optimize the regulatory mechanism of insulin secretion. A typical fusion peptide, (DAla2)GIP/xenin-8-Gln, has been shown to significantly enhance insulin secretion in vitro, effectively lower blood glucose levels in high-fat diet mice, improve glucose tolerance, and increase islet β-cell area.55 This enhancement may be attributed to the synergistic effect of the fusion peptide, combining GIP’s role in promoting insulin secretion and xenin’s function in regulating islet function to improve blood glucose metabolism. However, the current understanding of the pharmacokinetics and pharmacodynamics characteristics of this fusion peptide in humans remains limited, thus restricting its clinical application.
Neurotensin
Neurotensin is pivotal in modulating gastrointestinal motility, blood flow, and neuroregulation.58 The conjugation of xenin with neurotensin results in the Ac-NT/XN-8-Gln fusion peptide, demonstrating distinct metabolic regulatory capabilities.14 Investigations have revealed that this fusion peptide elicits a dose-dependent insulin-releasing response in BRIN-BD11 β cells and isolated mouse islets in vitro, and it can amplify the insulinotropic effect of GIP.14 In studies involving high-fat diet mice, administration of this fusion peptide over 32 days led to better blood glucose regulation and elevated circulating insulin levels. Notably, its combined application with exendin-4 resulted in heightened efficacy, manifested as reduced body fat, improved lipid profiles, and decreased glycosylated hemoglobin concentrations.14 Nevertheless, further studies are imperative to delineate the variances in the peptide’s effects under diverse physiological and pathological conditions and to ascertain potential long-term adverse reactions.
GLP-1/Gastrin/Xenin Receptor Tri-Agonist
Hasib et al ingeniously amalgamated exendin-4, Gastrin, and a modified version of xenin-8 to formulate the Exendin-4/gastrin/xenin-8-Gln hybrid peptide. Figure 7 displays the amino acid sequence of the novel hybrid peptide, and related parent peptides. The full sequence comprises the first N-terminal 28 amino acid residues of exendin-4, followed by gastrin-6, and then xenin-8-Gln, with each parent peptide coupled together by two 8-amino-3,6-dioxaoctanoic acid linker molecules (see Figure 7).59 This innovative approach seeks to harness the combined benefits of various gastrointestinal hormones, thereby achieving a multifaceted treatment strategy for diabetes.
The hybrid peptide is designed to simultaneously activate two distinct hormone signaling pathways by leveraging the dual agonist effects of GLP-1 and Gastrin. Through its activation of the GLP-1 receptor pathway, the peptide not only promotes insulin secretion but also aids in appetite suppression.56 Concurrently, the agonistic effect of Gastrin potentially influences insulin secretion and action indirectly by modulating the endocrine milieu of the gastrointestinal tract. Additionally, xenin-8 has been structurally modified to augment its stability and longevity. These modifications, which involve the introduction of specific chemical groups or alterations in the amino acid sequence, have enhanced xenin-8’s resistance to degradation in vivo.60 As a result, the peptide can sustainably regulate the function of pancreatic islet cells and synergistically interact with the effects of GLP-1 and Gastrin, ultimately leading to heightened insulin secretion and improved blood glucose management. Across pre-clinical models, GLP-1/xenin hybrids best suit short-term glycaemic rescue where β-cell mass preservation is critical; GIP/xenin co-agonists are preferred for GIP-resistant T2DM with concurrent obesity; neurotensin/xenin fusions display the strongest adipose-lipolytic profile and may serve as niche anti-obesity candidates.
Conclusion
Xenin has emerged as a promising therapeutic candidate in metabolic research due to its multifaceted roles in appetite suppression, glucose regulation, and β-cell protection, positioning it as a potential dual-target agent for T2DM and obesity. Native xenin, however, suffers from rapid proteolytic degradation and a short half-life (5–10 minutes), limiting its clinical application. Structural innovations—including C-terminal truncation (eg, xenin-8), amino acid substitutions (eg, Gln-modified analogues), lipidation (eg, xenin-25[Lys13PAL]), and fusion peptides—have markedly enhanced its metabolic stability, bioavailability, and receptor specificity. These modifications enable prolonged glycemic control, restored incretin sensitivity, and synergistic activation of pathways such as PI3K/Akt, underscoring xenin’s adaptability as a scaffold for next-generation therapies.
The development of multi-agonist peptides, such as GLP-1/xenin, GIP/xenin, and neurotensin/xenin hybrids, exemplifies a strategic shift toward targeting complementary pathways. These co-agonists amplify insulinotropic effects, suppress glucagon, and improve energy balance, outperforming single-target agents in preclinical models. For instance, GLP-1/xenin hybrids enhance β-cell mass and glucose tolerance, while neurotensin/xenin fusions demonstrate dose-dependent β-cell protection. Despite these advances, challenges persist, including species-specific gastrointestinal responses, incomplete mechanistic insights into receptor cross-talk, and the need for rigorous pharmacokinetic profiling and long-term safety evaluations.
Compared with approved multi-agonists (tirzepatide: GLP-1/GIP; survodutide: GLP-1/glucagon), xenin-containing hybrids remain exclusively in pre-clinical evaluation. While tirzepatide has demonstrated ≥2.0% HbA1c reduction and 15–20% weight loss in Phase III,61 xenin-8-Gln-based co-agonists have so far achieved ~1.2% HbA1c and 10% weight reduction in high-fat-fed mice,37 without long-term safety or dose-scaling data.13 Thus, xenin hybrids may offer mechanistic complementarity (NTSR1 engagement, GIP-sensitization), but their clinical positioning relative to validated GLP-1/GIP or GLP-1/GCG dual agonists remains speculative.
Looking ahead, xenin-based therapies hold potential to transcend traditional diabetes management. By integrating xenin into multi-agonist frameworks, researchers aim to replicate the metabolic benefits of bariatric surgery through pharmacological means. Additionally, optimizing oral delivery systems or exploring non-peptidyl xenin mimetics could address current limitations in administration and bioavailability. Future efforts must prioritize comprehensive clinical trials to validate efficacy in humans, alongside mechanistic studies to unravel receptor dynamics and tissue-specific actions. If successful, xenin-based multi-agonists have shown consistent efficacy in rodent models and offer mechanistic features (NTSR1 engagement, GIP re-sensitization) complementary to current GLP-1/GIP or GLP-1/GCG dual agonists. Nevertheless, their clinical impact remains speculative until safety, pharmacokinetics, and head-to-head trials in humans are completed.
Declaration of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this work, the author used ChatGPT and deep seek for language polishing. After using these services, the author reviewed and edited the content as needed and took full responsibility for the content of the published articles.
Acknowledgment
This work is supported by Zhejiang Province Medical and Health Science and Technology Plan Project (Grant No. 2025KY450), Zhejiang Province Postdoctoral Research Project (Grant No. ZJ2024136), China Postdoctoral Science Foundation Project (Grant No. 2025M772229), the Youth Science and Technology Talent Special Project of Jiaxing (Grant No. 2024AY40033), the Qin Shen Scholar Program of Jiaxing University.
Disclosure
The authors declare that there is no conflict of interest.
References
1. Hamscher G, Meyer HE, Feurle GE. Identification of proxenin as a precursor of the peptide xenin with sequence homology to yeast and mammalian coat protein alpha. Peptides. 1996;17(6):889–893. doi:10.1016/0196-9781(96)00150-7
2. Feurle GE, Hamscher G, Kusiek R, Meyer HE, Metzger JW. Identification of xenin, a xenopsin-related peptide, in the human gastric mucosa and its effect on exocrine pancreatic secretion. J Biol Chem. 1992;267(31):22305–22309. doi:10.1016/S0021-9258(18)41670-5
3. Schiavo-Cardozo D, Lima MMO, Pareja JC, Geloneze B. Appetite-regulating hormones from the upper gut: disrupted control of xenin and ghrelin in night workers. Clin Endocrinol. 2013;79(6):807–811. doi:10.1111/cen.12114
4. Maryanovich AT, Kormilets DY, Polyanovsky AD. Xenin: the oldest after insulin? Molecular Biology Reports. 2018;45(2):143–150. doi:10.1007/s11033-018-4147-2
5. Onaga T, Yasui Y, Hayashi H. Neurotensin and xenin stimulates pancreatic exocrine secretion through the peripheral cholinergic nerves in conscious sheep. Gen Comp Endocrinol. 2022;326:114073. doi:10.1016/j.ygcen.2022.114073
6. Kaji I, Akiba Y, Kato I, Maruta K, Kuwahara A, Kaunitz JD. Xenin Augments Duodenal Anion Secretion via Activation of Afferent Neural Pathways. J Pharmacol Exp Ther. 2017;361(1):151–161. doi:10.1124/jpet.116.238485
7. Kim ER, Lew PS, Spirkina A, Mizuno TM. Xenin-induced feeding suppression is not mediated through the activation of central extracellular signal-regulated kinase signaling in mice. Behav Brain Res. 2016;312:118–126. doi:10.1016/j.bbr.2016.06.026
8. Chowdhury S, Reeds DN, Crimmins DL, et al. Xenin-25 delays gastric emptying and reduces postprandial glucose levels in humans with and without type 2 diabetes. Am J Physiol Gastrointest Liver Physiol. 2014;306(4):G301–9. doi:10.1152/ajpgi.00383.2013
9. Gault VA, Martin CM, Flatt PR, Parthsarathy V, Irwin N. Xenin-25[Lys13PAL]: a novel long-acting acylated analogue of xenin-25 with promising antidiabetic potential. Acta Diabetologica. 2015;52(3):461–471. doi:10.1007/s00592-014-0681-0
10. Parthsarathy V, Irwin N, Hasib A, et al. A novel chemically modified analogue of xenin-25 exhibits improved glucose-lowering and insulin-releasing properties. Biochim Biophys Acta. 2016;1860(4):757–764. doi:10.1016/j.bbagen.2016.01.015
11. Craig SL, Gault VA, McClean S, Hamscher G, Irwin N. Effects of an enzymatically stable C-terminal hexapseudopeptide fragment peptide of xenin-25, ψ-xenin-6, on pancreatic islet function and metabolism. Mol Cell Endocrinol. 2019;496110523. doi:10.1016/j.mce.2019.110523
12. Hasib A, Khan D, Craig SL, Gault VA, Flatt PR, Irwin N. Antidiabetic effects and sustained metabolic benefits of sub-chronic co-administration of exendin-4/gastrin and xenin-8-Gln in high fat fed mice. Eur J Pharmacol. 2019;865:172733. doi:10.1016/j.ejphar.2019.172733
13. Tanday N, Moffett RC, Gault VA, Flatt PR, Irwin N. Enzymatically stable analogue of the gut-derived peptide xenin on beta-cell transdifferentiation in high fat fed and insulin-deficient Ins1(Cre/+);Rosa26-eYFP mice. Diabetes/Metab Res Rev. 2021;37(3):e3384. doi:10.1002/dmrr.3384
14. Perry RA, Craig SL, Gault VA, Flatt PR, Irwin N. A novel neurotensin/xenin fusion peptide enhances β-cell function and exhibits antidiabetic efficacy in high-fat fed mice. Biosci Rep. 2021;41(8). doi:10.1042/BSR20211275
15. Craig SL, Perry RA, Vyavahare SS, et al. A GIP/xenin hybrid in combination with exendin-4 improves metabolic status in db/db diabetic mice and promotes enduring antidiabetic benefits in high fat fed mice. Biochem Pharmacol. 2020;171:113723. doi:10.1016/j.bcp.2019.113723
16. Hasib A, Ng MT, Tanday N, et al. Exendin-4(Lys(27) PAL)/gastrin/xenin-8-Gln: a novel acylated GLP-1/gastrin/xenin hybrid peptide that improves metabolic status in obese-diabetic (ob/ob) mice. Diabetes Metab Res Rev. 2019;35(3):e3106. doi:10.1002/dmrr.3106
17. Kuwahara Y, Takahashi K, Akai M, et al. Minimum biological domain of xenin-25 required to induce anion secretion in the rat ileum. Peptides. 2022;147:170680. doi:10.1016/j.peptides.2021.170680
18. Wang S, Oestricker LZ, Wallendorf MJ, et al. Cholinergic signaling mediates the effects of xenin-25 on secretion of pancreatic polypeptide but not insulin or glucagon in humans with impaired glucose tolerance. PLoS One. 2018;13(2):e0192441. doi:10.1371/journal.pone.0192441
19. Sarnobat D, Moffett RC, Gault VA, et al. Effects of long-acting GIP, xenin and oxyntomodulin peptide analogues on alpha-cell transdifferentiation in insulin-deficient diabetic Glu(CreERT2);ROSA26-eYFP mice. Peptides. 2020;125:170205. doi:10.1016/j.peptides.2019.170205
20. Kerbel B, Badal K, Sundarrajan L, Blanco A, Unniappan S. Xenin is a novel anorexigen in goldfish (Carassius auratus). PLoS One. 2018;13(5):e0197817. doi:10.1371/journal.pone.0197817
21. Onaga T, Hayashi H, Yasui Y. Effects of xenin-25 on insulin and glucagon secretions in healthy conscious sheep. Domestic Animal Endocrinol. 2021;77:106635. doi:10.1016/j.domaniend.2021.106635
22. Craig SL, Irwin N, Gault VA. Xenin and Related Peptides: potential Therapeutic Role in Diabetes and Related Metabolic Disorders. Clin Med Insights Endocrinol Diabetes. 2021;14:11795514211043868. doi:10.1177/11795514211043868
23. English A, Craig SL, Flatt PR, Irwin N. Individual and combined effects of GIP and xenin on differentiation, glucose uptake and lipolysis in 3T3-L1 adipocytes. Biol Chem. 2020;401(11):1293–1303. doi:10.1515/hsz-2020-0195
24. Craig SL, Gault VA, Irwin N. Emerging therapeutic potential for xenin and related peptides in obesity and diabetes. Review. Diabet Metab Res Rev. 2018;34(6):e3006. doi:10.1002/dmrr.3006
25. Bhavya S, Lew PS, Mizuno TM. Central action of xenin affects the expression of lipid metabolism-related genes and proteins in mouse white adipose tissue. Neuropeptides. 2017;63:67–73. doi:10.1016/j.npep.2017.01.007
26. Kucukbas GN, Komuroglu AU, Dirik D, et al. Maternal serum xenin-25 levels in gestational diabetes mellitus. Eur Rev Med Pharmacol Sci. 2023;27(20):9902–9907. doi:10.26355/eurrev_202310_34168
27. Sterl K, Wang S, Oestricker L, et al. Metabolic responses to xenin-25 are altered in humans with Roux-en-Y gastric bypass surgery. Peptides. 2016;82:76–84. doi:10.1016/j.peptides.2016.06.001
28. Nauck MA, Meier JJ. The incretin effect in healthy individuals and those with type 2 diabetes: physiology, pathophysiology, and response to therapeutic interventions. Lancet Diabetes Endocrinol. 2016;4(6):525–536. doi:10.1016/s2213-8587(15)00482-9
29. Nauck MA, Heimesaat MM, Orskov C, Holst JJ, Ebert R, Creutzfeldt W. Preserved incretin activity of glucagon-like peptide 1 [7-36 amide] but not of synthetic human gastric inhibitory polypeptide in patients with type-2 diabetes mellitus. J Clin Invest. 1993;91(1):301–307. doi:10.1172/JCI116186
30. Fehmann HC, Göke R, Göke B. Cell and molecular biology of the incretin hormones glucagon-like peptide-I and glucose-dependent insulin releasing polypeptide. Endocrine Reviews. 1995;16(3):390–410. doi:10.1210/edrv-16-3-390
31. Vilsbøll T, Krarup T, Madsbad S, Holst JJ. Defective amplification of the late phase insulin response to glucose by GIP in obese Type II diabetic patients. Diabetologia. 2002;45(8):1111–1119. doi:10.1007/s00125-002-0878-6
32. Elahi D, McAloon-Dyke M, Fukagawa NK, et al. The insulinotropic actions of glucose-dependent insulinotropic polypeptide (GIP) and glucagon-like peptide-1 (7-37) in normal and diabetic subjects. Regul Pept. 1994;51(1):63–74. doi:10.1016/0167-0115(94)90136-8
33. Trujillo JM, Nuffer W. GLP-1 receptor agonists for type 2 diabetes mellitus: recent developments and emerging agents. Pharmacotherapy. 2014;34(11):1174–1186. doi:10.1002/phar.1507
34. Martin CMA, Gault VA, McClean S, Flatt PR, Irwin N. Degradation, insulin secretion, glucose-lowering and GIP additive actions of a palmitate-derivatised analogue of xenin-25. Biochem Pharmacol. 2012;84(3):312–319. doi:10.1016/j.bcp.2012.04.015
35. Wice BM, Wang S, Crimmins DL, et al. Xenin-25 potentiates glucose-dependent insulinotropic polypeptide action via a novel cholinergic relay mechanism. J Biol Chem. 2010;285(26):19842–19853. doi:10.1074/jbc.M110.129304
36. Taylor AI, Irwin N, McKillop AM, Patterson S, Flatt PR, Gault VA. Evaluation of the degradation and metabolic effects of the gut peptide xenin on insulin secretion, glycaemic control and satiety. J Endocrinol. 2010;207(1):87–93. doi:10.1677/joe-10-0085
37. Hasib A, Ng MT, Khan D, Gault VA, Flatt PR, Irwin N. A novel GLP-1/xenin hybrid peptide improves glucose homeostasis, circulating lipids and restores GIP sensitivity in high fat fed mice. Peptides. 2018;100:202–211. doi:10.1016/j.peptides.2017.10.015
38. Khan D, Vasu S, Moffett RC, Gault VA, Flatt PR, Irwin N. Locally produced xenin and the neurotensinergic system in pancreatic islet function and β-cell survival. Biol Chem. 2017;399(1):79–92. doi:10.1515/hsz-2017-0136
39. Chowdhury S, Wang S, Patterson BW, Reeds DN, Wice BM. The combination of GIP plus xenin-25 indirectly increases pancreatic polypeptide release in humans with and without type 2 diabetes mellitus. Regul Pept. 2013;187:42–50. doi:10.1016/j.regpep.2013.10.003
40. Guclu YA, Sahin E, Aksit M. The relationship between elevated serum xenin and insulin resistance in women with polycystic ovary syndrome: a case-control study. Gynecological Endocrinol. 2019;35(11):960–964. doi:10.1080/09513590.2019.1604663
41. Bhavya S, Lew PS, Mizuno TM. Stimulation of white adipose tissue lipolysis by xenin, a neurotensin-related peptide. Biochem Biophys Res Commun. 2018;498(4):842–848. doi:10.1016/j.bbrc.2018.03.067
42. Onaga T, Sakai A, Yasui Y. Intravenous administration of xenin-25 accelerates cyclic ruminal contractions in healthy conscious sheep. Neuropeptides. 2022;96:102293. doi:10.1016/j.npep.2022.102293
43. Onaga T, Sakai A, Kajita M, Fukuda H, Yasui Y, Hayashi H. Messenger RNA expression and localization of xenin in the gastrointestinal tract in sheep. Domestic Animal Endocrinol. 2021;74:106523. doi:10.1016/j.domaniend.2020.106523
44. Kim ER, Xu Y, Mizuno TM. Impaired suppression of feeding by the gut hormone xenin in type I interleukin-1 receptor-deficient mice. Behav Brain Res. 2014;261:60–64. doi:10.1016/j.bbr.2013.12.005
45. Saito S, Hashimoto H, Wakashin H, et al. Central administered xenin induced Fos expression in nesfatin-1 neurons in rats. Brain Res Bull. 2023;204:110788. doi:10.1016/j.brainresbull.2023.110788
46. van de Sande-Lee S, Cardoso AR, Garlipp CR, et al. Cerebrospinal fluid xenin levels during body mass reduction: no evidence for obesity-associated defective transport across the blood–brain barrier. Int J Obesity. 2013;37(3):416–419. doi:10.1038/ijo.2012.70
47. Kuwahara A, Kuwahara Y, Kato I, et al. Xenin-25 induces anion secretion by activating noncholinergic secretomotor neurons in the rat ileum. Am J Physiol. 2019;316(6):G785–G796. doi:10.1152/ajpgi.00333.2018
48. Denver P, Gault VA, McClean PL. Sustained high-fat diet modulates inflammation, insulin signalling and cognition in mice and a modified xenin peptide ameliorates neuropathology in a chronic high-fat model. Diabetes Obes Metab. 2018;20(5):1166–1175. doi:10.1111/dom.13210
49. Arslan N, Sayin O, Tokgoz Y. Evaluation of serum xenin and ghrelin levels and their relationship with nonalcoholic fatty liver disease and insulin resistance in obese adolescents. J Endocrinol Invest. 2014;37(11):1091–1097. doi:10.1007/s40618-014-0160-z
50. Martin CM, Parthsarathy V, Pathak V, Gault VA, Flatt PR, Irwin N. Characterisation of the biological activity of xenin-25 degradation fragment peptides. J Endocrinol. 2014;221(2):193–200. doi:10.1530/joe-13-0617
51. Gault VA, Martin CM, Parthsarathy V, Irwin N. A novel chemically modified analogue of xenin-25 exhibits improved glucose-lowering and insulin-releasing actions. Meeting Abstract. Diabetologia. 2014;57:S56–S57.
52. Craig SL, Gault VA, Hamscher G, Irwin N. Ψ-Xenin-6 enhances sitagliptin effectiveness, but does not improve glucose tolerance. J Endocrinol. 2020;245(2):219–230. doi:10.1530/joe-19-0557
53. Craig SL, Gault VA, Flatt PR, Irwin N. The methionine aminopeptidase 2 inhibitor, TNP-470, enhances the antidiabetic properties of sitagliptin in mice by upregulating xenin. Biochem Pharmacol. 2021;183:114355. doi:10.1016/j.bcp.2020.114355
54. Hagan T, Cortese M, Rouphael N, et al. Antibiotics-Driven Gut Microbiome Perturbation Alters Immunity to Vaccines in Humans. Cell. 2019;178(6):1313–1328e13. doi:10.1016/j.cell.2019.08.010
55. Hasib A, Ng MT, Gault VA, et al. An enzymatically stable GIP/xenin hybrid peptide restores GIP sensitivity, enhances beta cell function and improves glucose homeostasis in high-fat-fed mice. Diabetologia. 2017;60(3):541–552. doi:10.1007/s00125-016-4186-y
56. Drucker DJ. The biology of incretin hormones. Cell Metab. 2006;3(3):153–165. doi:10.1016/j.cmet.2006.01.004
57. Sarson DL, Wood SM, Holder D, Bloom SR. The effect of glucose-dependent insulinotropic polypeptide infused at physiological concentrations on the release of insulin in man. Diabetologia. 1982;22(1):33–36. doi:10.1007/bf00253866
58. Carraway R, Leeman SE. The isolation of a new hypotensive peptide, neurotensin, from bovine hypothalami. J Biol Chem. 1973;248(19):6854–6861. doi:10.1016/S0021-9258(19)43429-7
59. Hasib A, Ng MT, Khan D, Gault VA, Flatt PR, Irwin N. Characterisation and antidiabetic utility of a novel hybrid peptide, exendin-4/gastrin/xenin-8-Gln. Eur J Pharmacol. 2018;834:126–135. doi:10.1016/j.ejphar.2018.07.027
60. Martin CM, Parthsarathy V, Hasib A, et al. Biological Activity and Antidiabetic Potential of C-Terminal Octapeptide Fragments of the Gut-Derived Hormone Xenin. PLoS One. 2016;11(3):e0152818. doi:10.1074/jbc.M110.129304
61. Frías JP, Davies MJ, Rosenstock J, et al. Tirzepatide versus Semaglutide Once Weekly in Patients with Type 2 Diabetes. N Engl J Med. 2021;385(6):503–515. doi:10.1056/NEJMoa2107519
62. McConn BR, Park J, Gilbert ER, Cline MA. A novel role for xenopsin: stimulation of food intake. Behav Brain Res. 2015;292:500–507. doi:10.1016/j.bbr.2015.06.039
63. Mazella J, Béraud-Dufour S, Devader C, Massa F, Coppola T. Neurotensin and its receptors in the control of glucose homeostasis. Front Endocrinol. 2012;3:143. doi:10.3389/fendo.2012.00143
64. Feurle GE, Meyer HE, Hamscher G. Metabolism and potency of xenin and of its reduced hexapseudopeptide psi fragment in the dog. Life Sci. 2003;74(6):697–707. doi:10.1016/j.lfs.2003.05.009
65. Pathak NM, Pathak V, Gault VA, McClean S, Irwin N, Flatt PR. Novel dual incretin agonist peptide with antidiabetic and neuroprotective potential. Biochem Pharmacol. 2018;155:264–274. doi:10.1016/j.bcp.2018.07.021
66. Irwin N, Hasib A, Ng MT, Flatt PR, Gault VA. A novel exendin-4/gastrin/xenin-8-Gln fusion peptide, in combination with a stable GIP agonist, substantially improves metabolic control in high fat fed mice. Meeting Abstract Diabetol. 2018;61:S219–S219.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Evaluation of miRNA-143 and miRNA-145 Expression and Their Association with Vitamin-D Status Among Obese and Non-Obese Type-2 Diabetic Patients
Aladel A, Khatoon F, Khan MI, Alsheweir A, Almutairi MG, Almutairi SO, Almutairi FK, Osmonaliev K, Beg MMA
Journal of Multidisciplinary Healthcare 2022, 15:2979-2990
Published Date: 28 December 2022
Association of Serum Leptin and Adiponectin Concentrations with Type 2 Diabetes Biomarkers and Complications Among Saudi Women
Mohammed Saeed W, Nasser Binjawhar D
Diabetes, Metabolic Syndrome and Obesity 2023, 16:2129-2140
Published Date: 13 July 2023
DR10627, a Novel Dual Glucagon‑like Peptide‑1 and Gastric Inhibitory Polypeptide Receptor Agonist for the Treatment of Obesity and Type 2 Diabetes Mellitus
Shao Y, Chen Y, Zhu M, Liu Y, Fang C, Wang M, Sun P, Fu W, Huang J, Sheng S, Huang Y
Diabetes, Metabolic Syndrome and Obesity 2024, 17:1563-1573
Published Date: 6 April 2024
The Regulation of Metabolic Homeostasis by Incretins and the Metabolic Hormones Produced by Pancreatic Islets
Reed J, Bain SC, Kanamarlapudi V
Diabetes, Metabolic Syndrome and Obesity 2024, 17:2419-2456
Published Date: 13 June 2024
Relative Effectiveness and Safety of the GLP-1 (Glucagon-Like Peptide 1) Receptor Agonists, Semaglutide and Liraglutide in the Treatment of Obese Type 2 Diabetics: A Prospective Observational Cohort Study in Poland
Hoffmann K, Michalak M, Paczkowska A
Diabetes, Metabolic Syndrome and Obesity 2025, 18:2723-2738
Published Date: 7 August 2025