Back to Journals » Neuropsychiatric Disease and Treatment » Volume 18
VRT-043198 Ameliorates Surgery-Induced Neurocognitive Disorders by Restoring the NGF and BNDF Expression in Aged Mice
Authors Tang Q, Guo Q, Li K, Fei F
Received 1 March 2022
Accepted for publication 6 May 2022
Published 16 May 2022 Volume 2022:18 Pages 1027—1037
DOI https://doi.org/10.2147/NDT.S364250
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yu-Ping Ning
Qi Tang, Qiang Guo, Ke Li, Fan Fei
Department of Anesthesiology, First People’s Hospital of Taicang, Soochow University, Suzhou, Jiangsu Province, People’s Republic of China
Correspondence: Qiang Guo; Ke Li, Tel +86 15862329291 ; +86 15962297450, Email [email protected]; [email protected]
Background: Perioperative neurocognitive disorders (PND) are common surgical complications in the elderly. Pyroptosis-associated inflammation has been suggested to participate in a series of neurocognitive diseases, including Alzheimer’s disease. Given that VRT-043198 can reportedly inhibit caspase-1-induced pyroptosis, this study sought to determine whether VRT-043198 reduced PND in a mouse model following abdominal exploratory laparotomy.
Methods: 20-month-old male C57/BL mice were used to establish an abdominal exploratory laparotomy (AEL) model of PND. VRT-043198 (1, 10 and 100 mg/kg) was administered intraperitoneally immediately after surgery. Thirty days post-surgery, the mice were evaluated in the Morris water maze test. Their number of neurons, neurotrophin nerve growth factor (NGF) levels and brain-derived neurotrophic factor (BDNF) were measured. In the hippocampus, A1-type astrocytes and M1-type microglia were assessed using an immunofluorescence assay and Western blot, respectively. Caspase-1 activity, IL-1β, IL-18, and PPAR-γ were also measured 24h after surgery.
Results: VRT-043198 administration increased the time to cross the platform and increased the ratio of distance and time in the targeted quadrant after surgery. Furthermore, it was found that VRT-043198 restored neuronal amount, increased NGF and BDNF and decreased the number of A1-type astrocytes and M1-type microglia. VRT-043198 also attenuated caspase-1 activity, downregulated IL-1β and IL-18, but increased PPAR-γ 24h post-surgery.
Conclusion: VRT-043198 improved PND in aged mice after abdominal exploratory laparotomy by restoring the NGF and BNDF expression. These results indicate that VRT-043198 may be a potential therapy for PND.
Keywords: cognitive dysfunction, elderly, caspase-1, pyrpotosis, inflammation
Introduction
Perioperative neurocognitive disorders (PND) are characterized by cognitive impairments such as learning and memory deficits in the elderly after surgery.1 The patient may experience difficulties performing his daily activities and an alteration of his communication abilities.2 An increase in morbidity rate, mortality rate, hospital length of stay and frequent admissions to residential care institutions is also observed.3 The actual mechanism of PND is unknown; however, some studies performed on animals have demonstrated that a series of events, including inflammation, oxidative stress, and neurodegeneration, play a key role in the evolution of PND due to perioperative stress.4,5
Neuroapoptosis has been acknowledged as a mechanism of PND after general anesthesia and surgery, which leads to the dysfunction of axonal regeneration, and synaptic plasticity.6,7 Several studies support that anesthesia and surgery cause an inflammatory response triggering PND.8 The innate immune response induced by a peripheral surgery causes interleukin (IL)-1β activation, subsequently accelerating the inflammatory process in the hippocampus region, which is related to memory and learning.9 IL-1β is a vital inducer and executor of pyroptosis. It is identified as a NOD-like receptor family pyrin domain-containing 3 inflammasome causing inflammation using caspase-1.10 In addition to an acute inflammatory response, pyroptosis is also involved in chronic inflammatory diseases and PND in the elderly.11,12 Recently, more studies suggested that pyroptosis plays a role in the pathogenesis of neurocognitive diseases, including Alzheimer’s and Parkinson’s disease.13,14 It was chiefly reported that silencing pyroptosis-associated inflammasome could attenuate surgery-induced cognitive impairments.15–17 Subsequently, the attenuated decline in learning and memory following the inhibition of caspase-1-associated pyroptotic inflammation after surgery was investigated.
VRT-043198, a selectively blood-brain barrier permeable caspase-1 inhibitor, is reported to inhibit lipopolysaccharide-induced cytokine secretion.18 VRT-043198 suppresses the release of proinflammatory cytokines, including IL-1β and IL-18 but has limited effect on other cytokines such as TNF-α and IL-1α, and can be still monitored in the plasma 24h post-oral administration.19 In addition, VRT-043198 provides little inhibition against cellular apoptosis because of its low affinity with caspase-3 and −6 to −9.18 According to recent evidence, caspase-3 and caspase-9 specifically block inflammatory responses,20 establishing apoptosis as an immunologically silent form of cell death. The latter specifies that the apoptotic caspase cascade attenuates the inflammatory response.21 Given the selective inhibition of caspase-1, VRT-043198 has no effect on apoptosis as it is an immunologically silent phenomenon. Based on these findings, data were analyzed to determine whether VRT-043198 administration reduced the process of PND.
In the present study, an abdominal exploratory laparotomy model was established, followed by an investigation to determine whether the administration of VRT-043198 mitigated PND in aged mice after surgical treatment. Moreover, the role of caspase-1-associated pyroptotic inflammation in the process of PND was explored.
Methods
All experiments involving animals were performed according to the National Institute of Health Guideline for the Care and Use of Laboratory Animals. The protocols involving animals were also approved by the Animal Review Board of First People’s Hospital of Taicang.
Experimental Animals
20-month-old adult male C57/BL mice were provided by Charles River Co., Ltd. (Beijing, China). All aged mice were allowed free access to food and water and kept in a 12-h alternating light and dark facility at 25°C±1°C (humidity 50–70%).
Grouping
Mice were assigned to one of the following five groups at random: (1) sham; (2) abdominal exploratory laparotomy (AEL); (3) AEL plus VRT-043198-low dose (1 mg/kg, 0.5 mL) (AEL+LV); (4) AEL plus VRT-043198-medium dose (10 mg/kg, 0.5 mL) (AEL+MV); (5) AEL plus VRT-043198-high dose (100 mg/kg, 0.5 mL) (AEL+HV) groups. VRT-043198 administration (Cat#HY-112226; MedChemExpress, NJ, USA) was performed immediately after surgery via intraperitoneal injection. VRT-043198 was dissolved in 10% dimethyl sulfoxide (DMSO) (Cat#HY-Y0320; MedChemExpress, NJ, USA), 20% SBE-β-CD (Cat#HY-17031; MedChemExpress, NJ, USA), and 70% sterile saline. The mice in the sham and AEL groups were only administered an equal amount of solvent without VRT-043198. Ten mice in each group were selected to participate in the Morris Water Maze test 30 days post-surgery (n =10). On day 34, the mice were terminated, immunofluorescence staining (n =5) and Western blot (n =5) were performed. Five mice in each group were sampled 24h after surgery to assess the inflammatory response (n =5), including caspase-1 activity, PPAR-γ, serum IL-1β and IL-18.
Abdominal Exploratory Laparotomy
Abdominal exploratory laparotomy was performed to stimulate PND in aged mice, as described by previous studies.22,23 Anesthetic induction started with 7–8% sevoflurane followed by 3–4% sevoflurane for maintenance via face mask. Mice were kept warm at 37°C–38°C using a warming pad. Abdominal exploratory laparotomy was performed as follows: (1) the abdomen was shaved and cleaned with 75% alcohol; (2) a 1-cm midline incision was given; (3) the liver, spleen, colon, and stomach were gently manipulated using a sterile probe (10 min); (4) the small intestine (approximately 5 cm) was exteriorized out of the peritoneal cavity; covered with saline gauze, and then gently kneaded with fingers (20 min); (5) the muscle and skin were sutured with 6-0 Prolene; (6) local anesthesia was given at the site of incision with 0.2% ropivacaine (100 μL) for analgesic purposes. In the sham group, a skin and muscle incision was sutured under anesthesia. Similarly, an incision block using 0.2% ropivacaine (100μL) was given.
Caspase-1 Activity, IL-1β and IL-18 ELISA Analysis
Twenty-four hours post-surgery, the mice were administered heparin saline via the aorta after deep anesthesia with sevoflurane. The hippocampal tissues were isolated using a stereoscopic microscope and homogenized. Total protein was then extracted and quantified by BCA assay. According to the manufacturer’s protocol, the level of caspase-1 activity (Cat#C1102, Beyotime) was analyzed. Lastly, the serum from the collected blood sample was used to evaluate the levels of IL-1β (Cat#PI301, Beyotime) and IL-18 (Cat#EK0411, Boster, Wuhan, China).
Morris Water Maze Test (MWM)
Thirty days post-surgery, the changes in memory and learning ability were assessed using the Morris Water Maze test. The mice were placed in a circular container (diameter, 2 m) filled with white water (25 ± 1 °C). At the training stage (day 1 to day 4), the mice searched for a fixed platform in quadrant I, 0.5 cm below the water from quadrants I–IV. If the mice were unable to find the platform within 90s, they were trapped on the platform for 30s. At the testing stage (day 5), the movements of the mice from quadrant III were video captured. The indicators, including escape latency, the crossing platform times, the ratio of distance and time in the targeted quadrant, and speed, were analyzed with software provided by XinRuan Co., Ltd. (Shanghai, China).
Immunofluorescence
At the end of the Morris water maze test (Thirty-four days post-surgery), mice were perfused with heparin saline and 4% paraformaldehyde via the aorta after deep anesthesia with sevoflurane. Their hippocampal tissue was dehydrated in a series of graded ethanol solutions, vitrified, and embedded in paraffin. Subsequently, 4-μm thick paraffin slices were cut and dewaxed. The slices were rehydrated with alcohol; incubated with 0.1% Triton X-100 (Cat#T8200, Solarbio, Bejing, China) for 30 min and sealed with 5% standard bovine serum (Cat#A8010, Solarbio) for 1 h at a temperature of 25°C. After rinsing with PBS, polyclonal goat anti-Iba1 (Cat#ab5076, 1:400; Abcam, Cambridge, UK), monoclonal mouse anti-GFAP (Cat#K200067M, 1:400; Solarbio), polyclonal rabbit anti-CD86 (Cat#K000343P, 1:400; Solarbio), polyclonal rabbit anti-C3 (Cat#K004838P, 1:400; Solarbio) and monoclonal mouse anti-NeuN (Cat#K009907M, 1:400; Solarbio) were used as a primary antibody. The slices were then infiltrated with the primary antibody at a temperature of 4°C overnight. Similarly, a secondary antibody was made consisting of Cy3-conjugated donkey anti-goat IgG (Cat#A0502, 1:1000; Beyotime), Cy3-conjugated goat anti-mouse IgG (Cat#A0521, 1:1000; Beyotime) and FITC goat anti-rabbit IgG (Cat#A0562, 1:1000; Beyotime). The latter was used to infiltrate the slices over 1 h after 3 consecutive washouts with PBS. The nuclei were labeled using 4′,6-diamidino-2-phenylindole (DAPI) (10μg/mL, Cat#C1002, Beyotime). A fluorescence microscope (DP70, Olympus, Japan) was used for imaging purposes. Six fields with a magnification of ×400 in 3 sections were randomly selected from each group. Subsequently, the intensity, and number of A1-type astrocytes and M1-type microglia were analyzed using Image-Pro Plus 6.0 (NIH, Bethesda, MD, USA).
Western Blot
Twenty-four hours and Thirty-four days post-surgery, protein samples were acquired as mentioned above (caspase-1 activity). Thirty microgrammes of hippocampal tissues were mixed with loading buffer (Cat#P0015, Beyotime), boiled, and the proteins were separated by SDS-PAGE (12%) and shifted to a PVDF membrane. 5% skimmed milk was then used to block the PVDF membrane for 2 hours at a temperature of 25 °C. After three washes (5minutes each) with TBS-T, the PVDF membrane was incubated with primary antibodies monoclonal rabbit anti-peroxisome proliferator-activated receptor-γ (PPAR-γ) (Cat#ab272718, 1:500; abcam), polyclonal rabbit anti-BDNF (Cat#K008206P, 1:400; Solarbio), and polyclonal rabbit anti-NGF (Cat#K101525P, 1: 500; Beyotime) at a temperature of 4°C overnight. On the following day, the PVDF membrane was incubated with a secondary antibody (HRP Conjugated AffiniPure Goat Anti-rabbit IgG, Cat# BA1055, 1:2000; Boster, Wuhan, China) at a temperature of 25°C over 1 hour. Following incubation with ECL chemiluminescence solution (Cat#AR1173, Boster) for 5 min at 25 °C, the intensity of the targeted bands was analyzed using Image Lab software (Version 6.0, Bio-Rad Laboratories, MD, USA). GAPDH (1:1000, Cat#K106389P, Solarbio) was used as an internal reference.
Statistical Analysis
In this study, all mice were randomly assigned to different groups, and none were excluded. According to previous reports, the incidence of PND in aged mice is about 40–50%,22,24 and the response rate of VRT-043198 against neurological dysfunction is about 50–60%.25,26 Sample sizes (n =5 each group) were predetermined using existing data to obtain an overall power of α =0.05 and β =0.8. All data were expressed as mean ± standard deviation (SD) and analyzed using GraphPad Prism 5 software (GraphPad Software, San Diego, CA, USA). The groups were compared using either one-way analysis of variance (ANOVA) followed by Bonferroni’s post hoc tests or two-way analysis of variance followed by Turkey’s post hoc (for escapes latency of Morris Water Maze test). P <0.05 was considered statistically significant.
Results
VRT-043198 Administration Reduces Cognitive Dysfunction Induced by Abdominal Exploratory Laparotomy
A remarkable decrease in escape latency was found (Two-way ANOVA; time: F3, 140 =591.6, P <0.0001; group: F4, 140 =37.18, P <0.0001; interaction: F12, 140 =6.043, P <0.0001; Figure 1A and B) in the five groups. There was a significant increase in escape latency at 3 days after training in the Morris Water Maze test in the AEL and AEL+LV group compared with the sham group (vs AEL, P <0.0001; vs AEL+LV, P <0.0001; Figure 1A and B). At 4 days after training, mice in the AEL (P <0.0001), AEL+LV (P <0.0001), AEL+MV (P =0.0386), and AEL+HV (P =0.0357) group exhibited higher escape latency than mice from the sham group. On the testing day, the mice exposed to abdominal exploratory laparotomy exhibited a decrease in the crossing platform times (P <0.0001; Figure 1C). Those mice also showed a reduced ratio of distance (P <0.0001; Figure 1D) and time (P <0.0001; Figure 1E) in the targeted quadrant relative to sham-treated mice. It was also found that medium- (vs AEL, P <0.0001) and high-dose (vs AEL, P =0.0008) of VRT-043198 administration both increased the crossing platform times after surgical exposure (Figure 1C). Moreover, mice in the AEL+MV and AEL+HV group both exhibited a higher ratio of distance (AEL+MV vs AEL, P =0.0019; AEL+HV vs AEL, P =0.0111) and time (AEL+MV vs AEL, P =0.0006; AEL+HV vs AEL, P =0.0033) in the targeted quadrant compared with the AEL group (Figure 1D and E). However, low-dose VRT-043198 administration failed to improve the crossing platform times the ratio of distance and time in the targeted quadrant. There was no significant difference in the speed among mice in the five groups (Figure 1F). Notably, there was no significant difference of the crossing platform times the ratio of distance and time in the targeted quadrant between mice exposed to medium- and high-dose VRT-043198 after surgery (Figure 1C–E). To sum up, high-dose VRT-043198 showed no advantage to reverse the abdominal exploratory laparotomy-induced cognitive dysfunction compared with medium-dose VRT-043198 that was further studied in the next step.
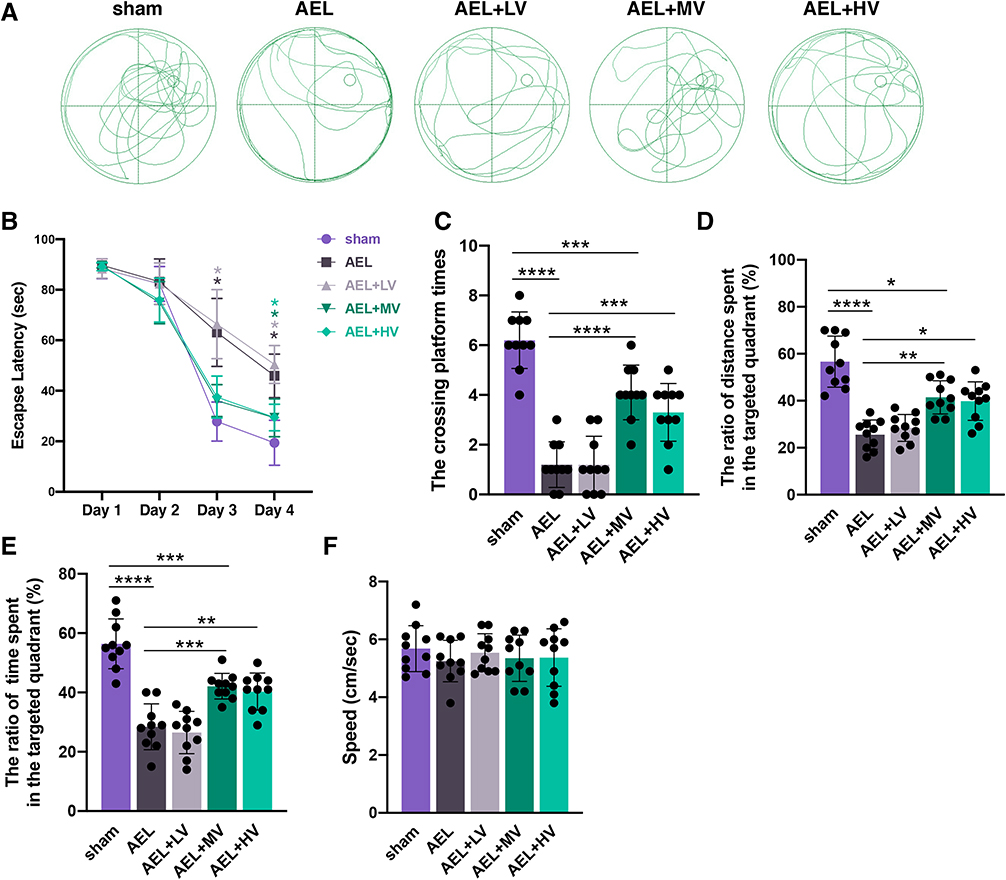 |
Figure 1 VRT-043198 administration mitigated abdominal exploratory laparotomy-induced memory and learning dysfunction (A) Trajectories generated from the Morris water maze test, 34 days post-surgery. AEL: mice were treated with abdominal exploratory laparotomy to stimulate perioperative neurocognitive disorders. Sham: mice underwent identical surgery except for abdominal exploratory laparotomy. LV, MV and HV: mice were administered with 1, 10 and 100 mg/kg VRT-043198, respectively, via intraperitoneal injection. (B) Escape latency from 30 to 33 days post-PND. (C–F) The platform crossing times, the ratio of the distance in the targeted quadrant, the time in the targeted quadrant, and the average swimming speed 34 days post-surgery. Data are presented as the mean ± SD (n =10). *P <0.05, **P <0.01, ***P <0.001, ****P <0.0001. |
VRT-043198 Administration Mitigates Pyroptosis-Associated Inflammation in the Early Stage After Surgery
The activity of caspase-1 in the hippocampus was also investigated in this study. The serum levels of IL-1β and IL-18 were evaluated to assess inflammatory response after surgery. In the present study, data showed that caspase-1 activity was significantly increased in mice that underwent surgery compared with mice in the sham group (vs sham, P <0.0001; Figure 2A). It was also found that the serum levels of IL-1β (vs sham, P <0.0001; Figure 2B) and IL-18 (vs sham, P <0.0001; Figure 2C) were both increased in the AEL group as compared to the sham group. However, medium-dose VRT-043198 administration in the AEL+MV group significantly attenuated caspase-1 activity (vs AEL, P =0.0009; Figure 2A), the levels of serum IL-1β (vs AEL, P =0.0005; Figure 2B) and IL-18 (vs AEL, P <0.0001; Figure 2C) compared to mice that underwent surgery. In addition, there was a significant difference in caspase-1 activity (vs sham, P <0.0001; Figure 2A), the levels of serum IL-1β (vs sham, P =0.0010; Figure 2B) and IL-18 (vs sham, P =0.0222; Figure 2C) between sham and AEL+MV groups. In addition, we found that the expression in PPAR-γ, which serves as a potential anti-inflammatory facor,27 was increased after surgical exposure (AEL vs sham, P <0.0001; Figure 2D and E), and further upregulated by VRT-043198 in the AEL+MV group compared with the AEL group (vs AEL, P <0.0001; Figure 2D and E).
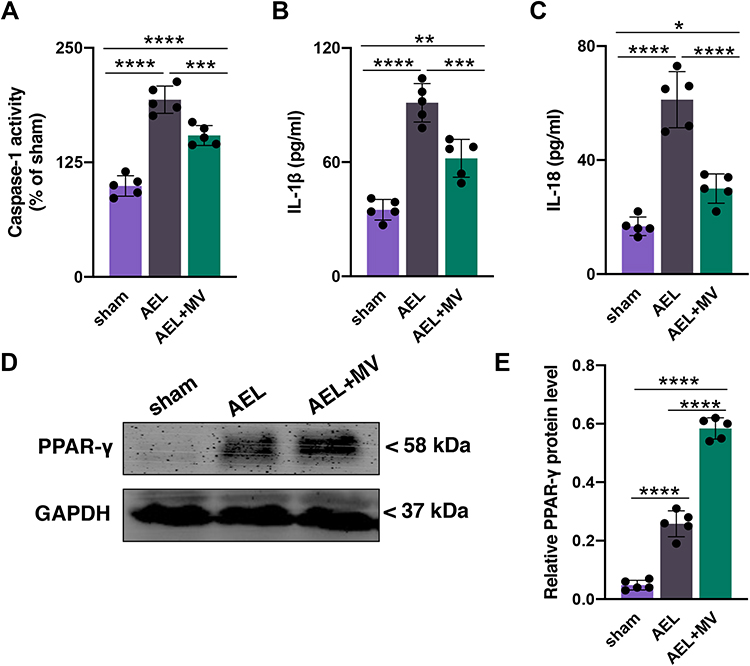 |
Figure 2 VRT-043198 administration mitigated abdominal exploratory laparotomy-induced pyroptosis-associated inflammation at the early stage. (A) Changes in hippocampal caspase-1 activity 24h post-surgery. (B–C) Changes in serum IL-1β and IL-18 24h post-surgery. (D) Representative blots of PPAR-γ 34 days post-surgery. (E) The ratio between the optical density value of PPAR-γ and GAPDH in the hippocampal tissue was evaluated by Western blot assay. Data are presented as the mean ± SD (n =5). Sham, AEL, and MV are described above. *P <0.05, **P <0.01, ***P <0.001, ****P <0.0001. |
VRT-043198 Administration Mitigates Neuronal Injury in the Late Stage After Surgery
Growing evidence demonstrated that the decline of memory and learning ability was associated with neuronal degeneration in the hippocampus.28,29 The neuronal amount indicated by NeuN-positive cells in the region of CA1 was significantly attenuated post- abdominal exploratory laparotomy in the AEL group compared with the sham group (vs sham, P =0.0001; Figure 3A and B). However, VRT-043198 administration after surgery induced a significant increase in neuronal amount (vs AEL, P =0.0241; Figure 3A and B). The neurotrophin nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF) in the brain are crucial for the survival, maintenance and regeneration of neurons.30 The study showed that the expression in NGF (vs sham, P <0.0001; Figure 3C and D) and BDNF (vs sham, P <0.0001; Figure 3C and E) were heavily reduced after surgical exposure. On the other hand, VRT-043198 increased the expression of NGF (vs AEL, P <0.0001; Figure 3C and D) and BDNF (vs AEL, P <0.0001; Figure 3C and E) in the AEL+MV group compared with the AEL group.
 |
Figure 3 VRT-043198 administration mitigated abdominal exploratory laparotomy-induced neuronal injury at the late stage. (A) Representative photomicrographs of NeuN staining. 400× magnification, scale bar =50 μm. (B) Changes in neuronal amount (NeuN+ cell count) in the CA1 region 34 days post-surgery. (C) Representative blots of NGF and BNDF 34 days post-surgery. (D–E) The ratio between the optical density value of NGF, BNDF and GAPDH in the hippocampal tissue was evaluated by Western blot assay. Data are presented as the mean ± SD (n =5). Sham, AEL, and MV are described above. *P <0.05, ***P <0.001, ****P <0.0001. |
VRT-043198 Administration Mitigates Inflammatory Response in the Late Stage After Surgery
Previous studies reported that chronic inflammatory response induced astrocytic and microglial activation in the hippocampus.31,32 In this study, it was found that abdominal exploratory laparotomy led to a significant increase in the intensity of GFAP (vs sham, P <0.0001; Figure 4A and B) and Iba1 (vs sham, P <0.0001; Figure 4D and E), while VRT-043198 administration partially reversed these changes (vs AEL, P <0.0001 for GFAP; vs AEL, P =0.0002 for Iba1; Figure 4A, B, D and E). In the present study, A1-type astrocytes indicated by C3 complement33 and M1-type microglia indicated by CD8634 were selected as biomarkers of inflammatory response in the late post-surgical stage. As seen in Figure 4, compared with sham-treated mice, a significant increase in mice exposed to abdominal exploratory laparotomy was observed in the A1-type astrocytes (vs sham, P <0.0001; Figure 4A and C) and M1-type microglia (vs sham, P <0.0001; Figure 4D and F) of the CA1 region. However, compared with mice in the AEL group, medium-dose VRT-043198 administration after surgery significantly reduced A1-type astrocytes (vs AEL, P =0.0002; Figure 4A and C) and M1-type microglia (vs AEL, P <0.0001; Figure 4D and F).
 |
Figure 4 VRT-043198 administration mitigated abdominal exploratory laparotomy-induced glial activation at the late stage. (A) Representative photomicrographs of GFAP and C3 complement plus DAPI staining (GFAP, red; C3 complement, green; DAPI, blue). 400× magnification, scale bar =50 μm. (B) Changes in GFAP intensity in the CA1 34 days post-surgery. (C) The percentage of GFAP combined with C3 complement-positive cells (A1-type). (D) Representative photomicrographs of Iba1 and CD86 complement plus DAPI staining (Iba1, red; CD86, green; DAPI, blue). 400× magnification, scale bar =50 μm. (E) Changes in Iba1 intensity in the CA1 region 34 days post-surgery. (F) The percentage of Iba1 combined with CD86-positive cells (M1-type). Data are presented as the mean ± SD (n =5). Sham, AEL, and MV are described above. **P <0.01, ***P <0.001, ****P <0.0001. |
Discussion
This study demonstrated that VRT-043198 administration alleviated abdominal exploratory laparotomy-induced cognitive dysfunction by inhibiting caspase-1 activity. Hence, significantly improving the crossing platform times and escape latency. VRT-043198 administration also increased the neuronal amount, NGF and BDNF expressions while decreasing A1-type astrocytes and M1-type microglia. Collectively, this study documented that VRT-043198 has a potential role against PND through restoring the NGF and BNDF expression.
Herein, the effect of VRT-043198 against PND in a model of abdominal exploratory laparotomy in aged mice was explored. Many studies have demonstrated that abdominal exploratory laparotomy could trigger the process of PND, inducing a significant long-term effect on memory and learning ability.35,36 Collected data revealed that abdominal exploratory laparotomy provoked a significant decline in cognition, evidenced by decreased platform crossing times. It is also associated with a reduced ratio of distance and time in the targeted quadrant, which was consistent with a previous study.22 Several studies suggested that PND following surgery was associated with neuronal injury and dysregulation of neurotrophic factors.37–39 Here, it was found that abdominal exploratory laparotomy reduced the neuronal amount and also attenuated the expressions of NGF and BDNF. Notably, the cerebral tissue was sampled under sevoflurane anesthesia, but sevoflurane anesthesia alone may have no impact on cognitive function as previous studies described.40,41 In addition, the control group eliminated the potential difference of pathology and molecules induced by sevoflurane anesthesia under sampling condition. These results suggested that cognitive dysfunction was successfully triggered in this PND model. Additionally, neuronal injury and the decline of neurotrophic factors in the hippocampus were related to PND.
It has been established that both acute and chronic inflammatory responses participate in the pathophysiology of perioperative neurocognitive disorders.42,43 In the present study, it was found that serum inflammatory factors, such as IL-1β and IL-18, were significantly increased at an early stage after surgery. In this context, it has been reported that proinflammatory cytokines, including IL-1β and IL-18, can lead to neuronal damage during ischemia or infection.9,44 Notable astrocytic and microglial activation in the CA1 was demonstrated in this study, which corroborates with a chronic inflammatory response in previous studies.45,46 In this regard, the blood-brain barrier dysfunction induced by inflammatory cytokines following surgical trauma reportedly facilitates the entry of IL-1β in the central nervous system, resulting in glial activation such as A1-type astrocytes and M1-type microglia in the CA1 region.47 IL-1β is also considered a vital contributor to the process of chronic neuroinflammation in the hippocampus.48 The chronic inflammatory response in the brain results in the decline of neurotrophic factors, subsequently leading to neuronal degeneration.49,50 These findings suggest that pyroptosis-associated inflammation participates in the process of PND.
VRT-043198 has been reported to be an anti-inflammatory agent that can inhibit caspase-1 activity. The latter is induced by inflammation.19 VRT-043198 was found to significantly attenuate caspase-1 activity, decrease the serum levels of IL-1β and IL-18, and increased PPAR-γ expression at the early post-surgical stage. Activated caspase-1 reportedly results in the activation of IL-1β and IL-18 and drives rapid pyroptosis.51 Downregulation of PPAR-γ mediated by activated caspase-1 is suggested to result in activation of transcription factors such as nuclear factor-κB (NF-κB), and liver-specific inhibitor of κB kinase β (Ikkb).52 There is overwhelming evidence that the attenuation of IL-1β and IL-18, and accumulation of PPAR-γ contribute chronic inflammatory response.52,53 The findings revealed that VRT-043198 administration reduces memory and learning dysfunction, as indicated by the Morris Water Maze tests. It also increases the neuronal amount and the levels of neurotrophic factors. In the late stage, VRT-043198 dampens the chronic inflammatory response of A1-type astrocytes and M1-type microglia. Studies have shown that chronic inflammatory response after early pyroptosis was involved in neuronal degeneration.54,55 In an apical periodontitis rat model, suppressing caspase-1 activity at the acute stage both reduces the early and chronic inflammatory responses.56 These results revealed that the VRT-043198-induced improvement in cognitive dysfunction might be mediated by suppressing pyroptosis-associated inflammation at both the early and late stages.
Several limitations of the current study should be documented. We only evaluated the expression of PPAR-γ under PND condition, but other anti-inflammatory factors including IL-10, and p38 mitogen-activated protein kinase (p38 MAPK), and transcription factors such as NF-κB, and Ikkb should further explored. In addition, only Morris Water Maze test was used to assess the learning and memory ability, and other types of behavioral tests for cognitive impairments should be performed in the further study.
In conclusion, VRT-043198 administration improves cognitive dysfunction in mice that underwent abdominal exploratory laparotomy. The mechanism involves the inhibition of pyroptosis-associated inflammation and the restoration of the NGF and BNDF expression. Although VRT-043198 demonstrates promising clinical prospects for PND treatment, further studies are required to have a better understanding of the underlying mechanisms.
Data Sharing Statement
All data generated and analysed during this study are included in this published article.
Funding
This study was supported by the Guiding Project of Suzhou City Science and Technology Bureau (SYSD2020027).
Disclosure
The authors declare that they have no competing interests.
References
1. Wiberg S, Holmgaard F, Zetterberg H, et al. Biomarkers of cerebral injury for prediction of postoperative cognitive dysfunction in patients undergoing cardiac surgery. J Cardiothorac Vasc Anesth. 2022;36(1):125–132. doi:10.1053/j.jvca.2021.05.016
2. Boone MD, Sites B, von Recklinghausen FM, Mueller A, Taenzer AH, Shaefi S. Economic burden of postoperative neurocognitive disorders among US medicare patients. JAMA Netw Open. 2020;3(7):e208931. doi:10.1001/jamanetworkopen.2020.8931
3. Rasmussen LS, Larsen K, Houx P, Skovgaard LT, Hanning CD, Moller JT. The assessment of postoperative cognitive function. Acta Anaesthesiol Scand. 2001;45(3):275–289. doi:10.1034/j.1399-6576.2001.045003275.x
4. Wang CM, Chen WC, Zhang Y, Lin S, He HF. Update on the mechanism and treatment of sevoflurane-induced postoperative cognitive dysfunction. Front Aging Neurosci. 2021;13:702231. doi:10.3389/fnagi.2021.702231
5. Lin X, Chen Y, Zhang P, Chen G, Zhou Y, Yu X. The potential mechanism of postoperative cognitive dysfunction in older people. Exp Gerontol. 2020;130:110791. doi:10.1016/j.exger.2019.110791
6. Han J, Pu CX, Xiao QX, et al. miRNA-124-3p targeting of LPIN1 attenuates inflammation and apoptosis in aged male rats cardiopulmonary bypass model of perioperative neurocognitive disorders. Exp Gerontol. 2021;155:111578. doi:10.1016/j.exger.2021.111578
7. Chen X, Gao F, Lin C, et al. mTOR-mediated autophagy in the hippocampus is involved in perioperative neurocognitive disorders in diabetic rats. CNS Neurosci Ther. 2022;28(4):540–553. doi:10.1111/cns.13762
8. Saxena S, Maze M. Impact on the brain of the inflammatory response to surgery. Presse Med. 2018;47(4):e73–e81. doi:10.1016/j.lpm.2018.03.011
9. Cibelli M, Fidalgo AR, Terrando N, et al. Role of interleukin-1β in postoperative cognitive dysfunction. Ann Neurol. 2010;68(3):360–368. doi:10.1002/ana.22082
10. Youm YH, Nguyen KY, Grant RW, et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat Med. 2015;21(3):263–269. doi:10.1038/nm.3804
11. Wu J, Lin S, Wan B, Velani B, Zhu Y. Pyroptosis in liver disease: new insights into disease mechanisms. Aging Dis. 2019;10(5):1094–1108. doi:10.14336/AD.2019.0116
12. Liu Z, Gan L, Xu Y, et al. Melatonin alleviates inflammasome-induced pyroptosis through inhibiting NF-κB/GSDMD signal in mice adipose tissue. J Pineal Res. 2017;63:e12414.
13. Moujalled D, Strasser A, Liddell JR. Molecular mechanisms of cell death in neurological diseases. Cell Death Differ. 2021;28(7):2029–2044. doi:10.1038/s41418-021-00814-y
14. de Brito Toscano EC, Rocha NP, Lopes BNA, Suemoto CK, Teixeira AL. Neuroinflammation in Alzheimer’s disease: focus on NLRP1 and NLRP3 inflammasomes. Curr Protein Pept Sci. 2021;22(8):584–598. doi:10.2174/1389203722666210916141436
15. Zuo Y, Yin L, Cheng X, et al. Elamipretide attenuates pyroptosis and perioperative neurocognitive disorders in aged mice. Front Cell Neurosci. 2020;14:251. doi:10.3389/fncel.2020.00251
16. Song P, Yi Z, Fu Y, et al. Reversing postcardiopulmonary bypass associated cognitive dysfunction using k-opioid receptor agonists to regulate microglial polarization via the NLRP3/Caspase-1 pathway. J Healthc Eng. 2021;2021:3048383. doi:10.1155/2021/3048383
17. Que YY, Zhu T, Zhang FX, Peng J. Neuroprotective effect of DUSP14 overexpression against isoflurane-induced inflammatory response, pyroptosis and cognitive impairment in aged rats through inhibiting the NLRP3 inflammasome. Eur Rev Med Pharmacol Sci. 2020;24(12):7101–7113. doi:10.26355/eurrev_202006_21704
18. Adriaenssens Y, Jiménez Fernández D, Vande Walle L, et al. Carboxylate isosteres for caspase inhibitors: the acylsulfonamide case revisited. Org Biomol Chem. 2017;15(35):7456–7473. doi:10.1039/C7OB01403A
19. Wannamaker W, Davies R, Namchuk M, et al. (S)-1-((S)-2-{[1-(4-amino-3-chloro-phenyl)-methanoyl]-amino}-3,3-dimethyl-butanoyl)-pyrrolidine-2-carboxylic acid ((2R,3S)-2-ethoxy-5-oxo-tetrahydro-furan-3-yl)-amide (VX-765), an orally available selective interleukin (IL)-converting enzyme/caspase-1 inhibitor, exhibits potent anti-inflammatory activities by inhibiting the release of IL-1beta and IL-18. J Pharmacol Exp Ther. 2007;321(2):509–516. doi:10.1124/jpet.106.111344
20. Taabazuing CY, Okondo MC, Bachovchin DA. Pyroptosis and apoptosis pathways engage in bidirectional crosstalk in monocytes and macrophages. Cell Chem Biol. 2017;24(4):507–514.e504. doi:10.1016/j.chembiol.2017.03.009
21. White MJ, McArthur K, Metcalf D, et al. Apoptotic caspases suppress mtDNA-induced STING-mediated type I IFN production. Cell. 2014;159(7):1549–1562. doi:10.1016/j.cell.2014.11.036
22. Xie X, Shen Z, Hu C, et al. Dexmedetomidine ameliorates postoperative cognitive dysfunction in aged mice. Neurochem Res. 2021;46(9):2415–2426. doi:10.1007/s11064-021-03386-y
23. Zhang Z, Li X, Li F, An L. Berberine alleviates postoperative cognitive dysfunction by suppressing neuroinflammation in aged mice. Int Immunopharmacol. 2016;38:426–433. doi:10.1016/j.intimp.2016.06.031
24. Wang J, Zhu S, Lu W, et al. Varenicline improved laparotomy-induced cognitive impairment by restoring mitophagy in aged mice. Eur J Pharmacol. 2022;916:174524. doi:10.1016/j.ejphar.2021.174524
25. Li J, Hao JH, Yao D, et al. Caspase-1 inhibition prevents neuronal death by targeting the canonical inflammasome pathway of pyroptosis in a murine model of cerebral ischemia. CNS Neurosci Ther. 2020;26(9):925–939. doi:10.1111/cns.13384
26. Liang Y, Song P, Chen W, et al. Inhibition of Caspase-1 ameliorates ischemia-associated blood-brain barrier dysfunction and integrity by suppressing pyroptosis activation. Front Cell Neurosci. 2020;14:540669. doi:10.3389/fncel.2020.540669
27. Monsalve FA, Pyarasani RD, Delgado-Lopez F, Moore-Carrasco R. Peroxisome proliferator-activated receptor targets for the treatment of metabolic diseases. Mediators Inflamm. 2013;2013:549627. doi:10.1155/2013/549627
28. Yassa MA, Stark CE. Pattern separation in the hippocampus. Trends Neurosci. 2011;34(10):515–525. doi:10.1016/j.tins.2011.06.006
29. Leblanc H, Ramirez S. Linking social cognition to learning and memory. J Neurosci. 2020;40(46):8782–8798. doi:10.1523/JNEUROSCI.1280-20.2020
30. Budni J, Bellettini-Santos T, Mina F, Garcez ML, Zugno AI. The involvement of BDNF, NGF and GDNF in aging and Alzheimer’s disease. Aging Dis. 2015;6(5):331–341. doi:10.14336/AD.2015.0825
31. Howe CL, LaFrance-Corey RG, Overlee BL, Johnson RK, Clarkson BDS, Goddery EN. Inflammatory monocytes and microglia play independent roles in inflammatory ictogenesis. J Neuroinflammation. 2022;19(1):22. doi:10.1186/s12974-022-02394-1
32. Nosi D, Lana D, Giovannini MG, Delfino G, Zecchi-Orlandini S. Neuroinflammation: integrated nervous tissue response through intercellular interactions at the ”Whole System” Scale. Cells. 2021;10(5):1195. doi:10.3390/cells10051195
33. Wang J, Hou Y, Zhang L, et al. Estrogen attenuates traumatic brain injury by inhibiting the activation of microglia and astrocyte-mediated neuroinflammatory responses. Mol Neurobiol. 2021;58(3):1052–1061. doi:10.1007/s12035-020-02171-2
34. Zhou T, Huang Z, Sun X, et al. Microglia polarization with M1/M2 phenotype changes in rd1 mouse model of retinal degeneration. Front Neuroanat. 2017;11:77. doi:10.3389/fnana.2017.00077
35. Muscat SM, Deems NP, D’Angelo H, et al. Postoperative cognitive dysfunction is made persistent with morphine treatment in aged rats. Neurobiol Aging. 2021;98:214–224. doi:10.1016/j.neurobiolaging.2020.11.008
36. Qiu LL, Ji MH, Zhang H, et al. NADPH oxidase 2-derived reactive oxygen species in the hippocampus might contribute to microglial activation in postoperative cognitive dysfunction in aged mice. Brain Behav Immun. 2016;51:109–118. doi:10.1016/j.bbi.2015.08.002
37. Subramaniyan S, Terrando N. Neuroinflammation and perioperative neurocognitive disorders. Anesth Analg. 2019;128(4):781–788. doi:10.1213/ANE.0000000000004053
38. Qiu LL, Pan W, Luo D, et al. Dysregulation of BDNF/TrkB signaling mediated by NMDAR/Ca(2+)/calpain might contribute to postoperative cognitive dysfunction in aging mice. J Neuroinflammation. 2020;17(1):23. doi:10.1186/s12974-019-1695-x
39. Xiong L, Duan L, Xu W, Wang Z. Nerve growth factor metabolic dysfunction contributes to sevoflurane-induced cholinergic degeneration and cognitive impairments. Brain Res. 2019;1707:107–116. doi:10.1016/j.brainres.2018.11.033
40. Alalawi R, Yasmeen N. Postoperative cognitive dysfunction in the elderly: a review comparing the effects of desflurane and sevflurane. J Perianesth Nurs. 2018;33(5):732–740. doi:10.1016/j.jopan.2017.04.009
41. Walters JL, Zhang X, Talpos JC, et al. Sevoflurane exposure has minimal effect on cognitive function and does not alter microglial activation in adult monkeys. Neurotoxicology. 2019;71:159–167. doi:10.1016/j.neuro.2018.12.008
42. Yu L, Wen G, Zhu S, Hu X, Huang C, Yang Y. Abnormal phosphorylation of tau protein and neuroinflammation induced by laparotomy in an animal model of postoperative delirium. Exp Brain Res. 2021;239(3):867–880. doi:10.1007/s00221-020-06007-2
43. Zhu Y, Wang Y, Yao R, et al. Enhanced neuroinflammation mediated by DNA methylation of the glucocorticoid receptor triggers cognitive dysfunction after sevoflurane anesthesia in adult rats subjected to maternal separation during the neonatal period. J Neuroinflammation. 2017;14(1):6. doi:10.1186/s12974-016-0782-5
44. Song L, Pei L, Yao S, Wu Y, Shang Y. NLRP3 inflammasome in neurological diseases, from functions to therapies. Front Cell Neurosci. 2017;11:63. doi:10.3389/fncel.2017.00063
45. Ragu-Varman D, Macedo-Mendoza M, Labrada-Moncada FE, et al. Anorexia increases microglial density and cytokine expression in the hippocampus of young female rats. Behav Brain Res. 2019;363:118–125. doi:10.1016/j.bbr.2019.01.042
46. Taipa R, Ferreira V, Brochado P, et al. Inflammatory pathology markers (activated microglia and reactive astrocytes) in early and late onset Alzheimer disease: a post mortem study. Neuropathol Appl Neurobiol. 2018;44:298–313. doi:10.1111/nan.12445
47. Skvarc DR, Berk M, Byrne LK, et al. Post-operative cognitive dysfunction: an exploration of the inflammatory hypothesis and novel therapies. Neurosci Biobehav Rev. 2018;84:116–133. doi:10.1016/j.neubiorev.2017.11.011
48. Jiang C, Zhang Y, Tang X, et al. IL-6 and IL-1β upregulation and tau protein phosphorylation in response to chronic alcohol exposure in the mouse hippocampus. Neuroreport. 2021;32(10):851–857. doi:10.1097/WNR.0000000000001661
49. DeLegge MH, Smoke A. Neurodegeneration and inflammation. Nutr Clin Pract. 2008;23(1):35–41. doi:10.1177/011542650802300135
50. Ertürk A, Mentz S, Stout EE, et al. Interfering with the chronic immune response rescues chronic degeneration after traumatic brain injury. J Neurosci. 2016;36(38):9962–9975. doi:10.1523/JNEUROSCI.1898-15.2016
51. Man SM, Karki R, Kanneganti TD. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol Rev. 2017;277(1):61–75. doi:10.1111/imr.12534
52. Meng QQ, Feng ZC, Zhang XL, et al. PPAR-γ activation exerts an anti-inflammatory effect by suppressing the NLRP3 inflammasome in spinal cord-derived neurons. Mediators Inflamm. 2019;2019:6386729. doi:10.1155/2019/6386729
53. Fortunato F, Berger I, Gross ML, Rieger P, Buechler MW, Werner J. Immune-compromised state in the rat pancreas after chronic alcohol exposure: the role of peroxisome proliferator-activated receptor gamma. J Pathol. 2007;213(4):441–452. doi:10.1002/path.2243
54. Cheng Q, Ding H, Fang J, et al. Aquaporin 9 represents a novel target of chronic liver injury that may antagonize its progression by reducing lipotoxicity. Oxid Med Cell Longev. 2021;2021:5653700. doi:10.1155/2021/5653700
55. Yan YQ, Fang Y, Zheng R, Pu JL, Zhang BR. NLRP3 inflammasomes in Parkinson’s disease and their regulation by Parkin. Neuroscience. 2020;446:323–334. doi:10.1016/j.neuroscience.2020.08.004
56. Cheng R, Feng Y, Zhang R, Liu W, Lei L, Hu T. The extent of pyroptosis varies in different stages of apical periodontitis. Biochim Biophys Acta Mol Basis Dis. 2018;1864(1):226–237. doi:10.1016/j.bbadis.2017.10.025
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Comparison of the Effects of Dexmedetomidine and Lidocaine on Stress Response and Postoperative Delirium of Older Patients Undergoing Thoracoscopic Surgery: A Randomized Controlled Trial
Lai Y, Chen Q, Xiang C, Li G, Wei K
Clinical Interventions in Aging 2023, 18:1275-1283
Published Date: 3 August 2023
Systemic Inflammation Response Index as a Predictor of Stroke Risk in Elderly Patients with Hypertension: A Cohort Study
Cai X, Song S, Hu J, Wang L, Shen D, Zhu Q, Yang W, Luo Q, Hong J, Li N
Journal of Inflammation Research 2023, 16:4821-4832
Published Date: 24 October 2023
The Mechanism of Pyroptosis and Its Application Prospect in Diabetic Wound Healing
Al Mamun A, Shao C, Geng P, Wang S, Xiao J
Journal of Inflammation Research 2024, 17:1481-1501
Published Date: 6 March 2024
A Mixture of Water-Soluble Polysaccharides Reduces Caspase-1 and IL-1β Inflammatory Responses by Cutibacterium acnes in vitro in Reconstructed Human Epidermis (RHE)
Gruber JV, Terpak N, Massard S, Chen X, Kim Y
Clinical, Cosmetic and Investigational Dermatology 2024, 17:2627-2633
Published Date: 21 November 2024
Clinical Relevance of Preoperative Lymphocyte-to-Monocyte Ratio in Predicting Postoperative Delirium Across Frailty Status in Older Surgical Patients
Zhang H, Han S, Li F, Hou D, Lv X, Lou J, Li H, Cao J, Mi W, Liu Y
Clinical Interventions in Aging 2026, 21:584851
Published Date: 7 May 2026