Back to Journals » International Journal of Nanomedicine » Volume 20
Understanding the Molecular Toxicity of Metal-Based Nanoparticles Through Nanotoxicomics
Authors González-Vega JG, Fabian-Ortiz E, Hernández-Pérez J, Hinojosa Alvarez S, Chavez Santoscoy RA ![]()
Received 12 July 2025
Accepted for publication 14 October 2025
Published 4 November 2025 Volume 2025:20 Pages 13293—13315
DOI https://doi.org/10.2147/IJN.S553276
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Krishna Nune
Jesús Gabriel González-Vega,* Emilio Fabian-Ortiz,* Jesús Hernández-Pérez, Silvia Hinojosa Alvarez, Rocio Alejandra Chavez Santoscoy
School of Engineering and Science, Tecnologico de Monterrey, Monterrey, NL, Mexico
*These authors contributed equally to this work
Correspondence: Rocio Alejandra Chavez Santoscoy, School of Engineering and Science, Tecnologico de Monterrey, Monterrey, NL, Mexico, Email [email protected]
Abstract: Metal and metal oxide nanoparticles (NPs) are prevalent in industrial, medical, and consumer applications, yet concerns persist regarding their potential impact on human health. Traditional endpoint-based toxicological studies often face challenges in explaining the complex molecular mechanisms underlying the interactions between the physicochemical properties of NPs (eg, chemical composition, size, coating) and biological factors (eg, cell type). To address this gap, we introduce nanotoxicomics, a stepwise framework that integrates nanotoxicology with advanced omics technologies—transcriptomics, proteomics, and metabolomics. This approach enables multilevel analysis of NP effects, supporting the extrapolation of in vitro findings to tissues, organs, or whole systems. Moreover, this framework provides insights into the molecular mechanisms underlying consumer-product interactions and occupational exposure to NPs by enabling early detection of transcriptomic biomarkers, guiding downstream proteomic validation, and supplying metabolomic functional readouts that can be translated into potential clinical outcomes and enhanced risk assessment strategies. This review compiles and critically discusses evidence on the interactions of metal and metal oxide NPs with human cells and occupationally exposed populations. Key mechanisms include differential expression of genes related to oxidative stress, inflammation, and DNA damage (eg, SOD1, IL-6, MAP3K14, TP53), validation of protein biomarkers such as heat shock proteins and metallothioneins, and metabolite shifts reflecting ATP depletion, mitochondrial dysfunction, and redox imbalance (eg, GSH, PGE2, ATP). Finally, we highlight the value of nanotoxicomics and bioinformatics for designing safer NPs that can function as nanosensors, real-time monitoring agents, and drug delivery systems in nanomedicine, particularly in oncology, neurology, immunology, and cardiology. Overall, this study provides valuable insights into the molecular basis of metal and metal oxide nanoparticles and lays the groundwork for future research in predictive nanotoxicology, biomonitoring, and omics-driven risk assessment.
Keywords: nanotoxicology, transcriptomics, proteomics, metabolomics, nanomaterials, occupational exposure, metal-based nanoparticles
Introduction
Nanotechnology involves the synthesis and manipulation of nanomaterials (NMs) with dimensions less than 100 nm and has found widespread applications across diverse industries, including biomedicine, electronics, food, and aerospace. Among these NMs, nanoparticles (NPs) stand out due to their modifiable physicochemical properties—such as size, shape, and surface charge—their ease of synthesis, and their ability to be functionalized with bioactive molecules, including drugs, nucleic acids, proteins, polymers, or lipids. These properties make NPs ideal for use with antimicrobial agents, cancer therapeutics, orthopedic implants, and drug delivery systems.1–3
Despite these advantages, NPs can also confer the potential for toxicity, including the induction of oxidative stress, inflammation, and DNA damage, which may affect the musculoskeletal, neurological, respiratory, cardiovascular, and reproductive systems.4–7 Such toxic effects have been observed in vitro, including reduced cell viability, increased apoptosis, and altered proliferation in various human and animal cell lines, as well as in vivo, where exposure can lead to organ-specific damage, immune system activation, and physiological alterations in animal models.7–10 Given that studies predict approximately eight million people will be exposed to engineered and naturally occurring metal-based nanoparticles by 2029, understanding their toxic effects is crucial for assessing health risks and informing safer design and regulatory strategies.11 This is particularly significant for metal-based nanoparticles, which are among the most widely used nanomaterials in consumer and biomedical applications, and whose unique redox activity can amplify toxic responses compared to other classes of nanomaterials.8
Traditional nanotoxicology approaches, employing both in vitro and in vivo models, are essential for elucidating the cytotoxicity and pharmacokinetic aspects of NPs in biological systems. In vitro models enable researchers to evaluate key factors such as cell viability, proliferation, and changes in cell morphology, which can aid in determining toxicity in a cost-effective and highly reproducible manner.12 In contrast, in vivo models provide insights into the systemic effects of these NMs, allowing for the analysis of specific responses in various organs through histopathological examination, as well as the assessment of immune responses; physiological and behavioral changes; and, importantly, pharmacokinetic parameters related to absorption, distribution, metabolism, and excretion. While these approaches effectively evaluate important aspects of NP bioactivity, they offer limited insights into the underlying mechanisms and biological responses associated with these NMs at the different molecular levels. Hence, to close these gaps in knowledge researchers are starting to adopt cutting-edge technologies such as omics sciences to incorporate them in traditional nanotoxicology assessment, giving birth to a new field that we propose as Nanotoxicomics.
Nanotoxicomics integrates nanotoxicology—the study of the toxicological effects and mechanisms of NPs on biological systems—with omics technologies—including transcriptomics, proteomics, and metabolomics. This framework provides insights into the global biological responses to NPs in various biological models, including cell lines, primary cultures and human biological samples. Unlike targeted analyses, these broader approaches have enhanced our understanding of NP-induced cytotoxicity, pharmacokinetics, and occupational exposure to the organ, tissue, and cellular levels.13 For instance, while conventional cytotoxicity assays failed to reveal any significant effects of TiO2 nanoparticles at sublethal concentrations, omics-based analyses identified alterations in DNA methylation patterns and lncRNA expression, highlighting molecular perturbations not detectable by traditional methods.14 However, the widespread adoption of these advanced techniques remains limited due to the high cost of equipment and data analysis, lack of familiarity with omics methods, and the complexity of data integration. Moreover, most nanotoxicomics studies still rely on individual omics approaches, which provide only a partial view of NP toxicity. Notably, multiomics studies—combining two or more omics techniques—are gaining acceptance, offering more robust datasets and a clearer understanding of NP toxicity mechanisms.15–19
This review aims to present the latest findings in nanotoxicomics by (1) exploring the primary nanotoxicological effects of NPs in biological systems and their potential risks to human health; (2) describing the known molecular processes involved in NP interactions at the levels of genes, proteins, and metabolites; (3) providing a comprehensive analysis of the signaling cascades triggered by NP-biological interactions; (4) exploring the correlations between the physicochemical properties of NPs and biological factors such as cell type and discussing the implications of their mechanisms of action at both the cellular and systemic levels; (5) proposing integrative strategies to move from single to multiomics approaches; and (6) providing insights into biomarker identification and strategies for safer usage and design of NPs, informed by a multiomics approach within the context of nanotoxicology.
Exploring the Primary Nanotoxicological Effects of NPs in Biological Systems and Their Potential Risks to Human Health
Exposure to NPs can occur primarily through inhalation, ingestion, ocular intake and/or dermal absorption.20 This can be attributed to occupational, environmental, and consumer-related sources, where nanoparticles interact with biological barriers biological barriers such as the skin, respiratory epithelium, ocular surface and gastrointestinal tract. Once internalized, NPs may accumulate in tissues and trigger diverse toxicological outcomes. Among the most documented examples we have, we have orthopedic toxicity, which can arise from titanium NP liberation derived from the corrosion of orthopedic and dental implants. In humans carrying titanium spinal implants, high levels of inflammation and fibrosis have been reported in surrounding tissues via activation of the SNAI2–SPP1–PI3K/AKT axis.14 Another relevant case is that of magnetic imaging agents such as iron oxide NPs, which at sizes of 2.3 and 4.2 nm have been shown to induce mortality in male ICR mice as a result of oxidative damage to several organs, particularly the heart.9
Beyond these clinical and experimental reports, occupational exposure represents one of the most concerning scenarios, as workers may experience sustained and repeated contact with NPs, significantly increasing the risk of long-term toxicological effects. In recent years, the focus of studies in this area has been directed towards healthcare workers, manufacturing industries, preprinting centers, and research and development laboratories.21,22 However, the estimation of occupational nanotoxicity has largely been restricted to a tiny family of biomarkers associated with free radical reactions including lipid, nucleic acid, and protein oxidation.23 Numerous techniques such as immunological assays, DNA amplification techniques, and statistical analysis have been employed to detect the presence of these biomarkers in urine, blood serum, and exhaled breath condensate samples after the exposure to NPs24 These occupational exposure biomarkers have been linked to the development of pathological conditions such as cancer, sarcoidosis and idiopathic pulmonary fibrosis, among many others.25 Despite this fact, the molecular interactions between nanoparticles and biological systems at the cellular and organ levels remain poorly understood (Figure 1).
 |
Figure 1 Impact of metal and metal oxide nanoparticles on human health. The figure illustrates how metallic and metal oxide NPs can affect different organs in the human body after inhalation, ingestion, ocular intake or dermal absorption. Each organ is labeled to highlight specific health risks associated with NP exposure. |
The presence of NPs in biological samples such as blood, urine, and feces demonstrates their ability to cross biological barriers, including the air‒blood, gut‒blood, and blood‒brain barriers.26 This capacity is largely due to their nanoscale size, which increases their surface area and facilitates biocorona formation—where cellular components and proteins adsorb to NP surfaces, altering their physicochemical properties and conferring a new biological identity. Once NPs trespass these barriers, they enter the bloodstream and are distributed to various organs and tissues.27 Multiple pharmacokinetic studies have revealed that metal and metal oxide NPs, such as gold, iron oxide, and titanium dioxide, can be bioaccumulated and metabolized by biological systems depending on both their physicochemical properties (eg, shape, size, and charge) and biological factors (eg, cell type, biocorona formation, and individual genetic traits).28 For example, Rosário et al (2022)29 demonstrated that silver nanoparticles (AgNPs) accumulate in different organs depending on their size. NPs between 5 and 10 nm in size bioaccumulate in a dose-dependent manner in organs such as the lungs, spleen, kidneys, heart, and brain after intratracheal exposure, whereas smaller AgNPs tend to accumulate in the kidneys, liver, and blood more easily. On the other hand, Brandão et al (2020)30 reported significant differences in the uptake profiles of 25 nm TiO2-NPs across various cell types, including adenocarcinoma alveolar epithelial cells (A549), hepatocellular carcinoma epithelial cells (HepG2), glioblastoma glial cells (A172), and neuroblastoma neuronal cells (SH-SY5Y), when exposed to concentrations ranging from 10 to 200 µg/mL. Similarly, a study by Mahmood et al (2022)31 reported that 33.24 nm copper oxide nanoparticles (CuO-NPs) caused a significant reduction in cell viability in an invasive breast carcinoma cell line (AMJ13) and a human breast adenocarcinoma cell line (MCF-7), whereas no significant inhibition was observed in a healthy human breast epithelial cell line (HBL-100) within the concentration range of 6.25–100 µg/mL. These findings highlight the complex, cell-specific nature of NP toxicity and emphasize the importance of considering both physicochemical and biological factors when assessing NP safety.
In addition to their systemic distribution, nanoparticles can undergo various metabolic transformations within biological systems. Metal and metal oxide NPs, for example, may partially dissolve to release ions, interact with enzymatic pathways, or be sequestered by organelles such as lysosomes, all of which influence their toxicity and clearance. Silver NPs, for instance, can be metabolized through two main pathways: either dissolving into toxic silver ions or transforming into less harmful silver sulfide, which reduces toxicity by decreasing ion bioavailability. AgNPs are excreted through saliva, sweat, feces, and urine, but their clearance can take from several days to months.19,32 The slow clearance and persistent excretion of NPs suggest that they can accumulate in the environment and be reabsorbed by humans and other organisms, creating a cycle of exposure.33 Chronic exposure to AgNPs may lead to bioaccumulation in humans, manifesting as silver granules in tissues and causing argyria, characterized by bluish‒gray skin discoloration. Despite these insights into AgNP fate and chronic accumulation, the molecular mechanisms underlying their interactions with cellular pathways, gene expression, and protein networks remain largely unexplored. This highlights the need for integrative omics approaches, including transcriptomic, proteomic, and metabolomic analyses, to fully characterize nanoparticle-induced perturbations and better predict potential toxicological outcomes.
While traditional cytotoxicity assays such as MTT (measuring mitochondrial activity), Comet (evaluating DNA strand breaks), and LDH (detecting membrane damage) remain important, they may fail to detect subtle or low-dose effects of nanoparticles.34 Integrating these assays with functional omics approaches can provide a more comprehensive understanding of NP interactions with biological systems, particularly for chronic or low-level exposures. Nevertheless, challenges related to variability, reproducibility, and protocol standardization persist. Careful evaluation of these strengths and limitations is essential before delving into the molecular processes involved in NP interactions at the levels of genes, proteins, and metabolites.
Describing the Molecular Processes Involved in NP Interactions at the Levels of Genes, Proteins, and Metabolites
Transcriptomic Data for Gene Expression Analysis
Transcriptomics focuses on the comprehensive study of RNA molecules to evaluate gene expression and has become an essential tool in elucidating cellular responses to diverse stimuli, including NPs. By quantifying messenger RNA (mRNA) levels, transcriptomics facilitates the investigation of dynamic changes in gene activity under specific experimental conditions being used in several studies for nanoparticle-cell interactions.35 It is well known that transcriptomics offers a direct measure of gene expression, allowing for the detection of changes at an earlier stage than proteomics or metabolomics. Unlike proteomics, which struggle with low-abundance proteins, or metabolomics, which is influenced by the complexity and variability of metabolic pathways, it enables high-throughput analysis with broad sensitivity. Additionally, it is cost-effective and supported by robust computational tools, making it an accessible method for large-scale studies. However, since transcriptomics does not directly measure the functional output of genes, such as protein activity or metabolic changes, its results may not always correlate with protein abundance or metabolic activity because of factors such as posttranslational modifications or protein degradation.
Transcriptomic techniques have evolved to become crucial for studying gene expression, each with its own strengths and limitations. Hybridization-based technologies, such as DNA microarrays, represent major advancements in high-throughput gene expression profiling by enabling the simultaneous analysis of thousands of genes. However, their reliance on predefined probes limits the discovery of novel transcripts and can introduce challenges, such as cross-hybridization.36 Advances in sequencing technologies, particularly RNA sequencing (RNA-Seq), have overcome these limitations, providing a broader dynamic range, high sensitivity for detecting rare transcripts, and the ability to identify novel transcripts.37 Despite its limitations, transcriptomics remains a powerful tool in nanotoxicomics, offering valuable insights into gene expression changes and cellular responses to nanoparticles, particularly when large-scale gene profiling and pathway analysis are needed.
Transcriptomic Insights Into the Mode of Action of Metal and Metal Oxide Nanoparticles in Human Cell Lines
Studies of metal and metal oxide nanoparticles have revealed their profound impact on gene expression across various cell lines. For example, 20-nm AgNPs at a concentration of 38.6 μg/mL can activate the Nrf2-mediated oxidative stress response pathway, the G2/M DNA damage checkpoint, and Toll-like receptor signaling in human adenocarcinoma alveolar epithelial cells (A549).16 On the other hand, 20 µg/mL of 18.18 nm titanium dioxide nanoparticles (TiO2-NPs) can upregulate angiogenesis, TGF-beta and chemokine and cytokine signaling pathways in human glioblastoma cells (T98G).4 These effects are not uniform but instead reflect the cell’s capacity to respond through diverse molecular mechanisms and pathways, which are shaped by the nature of the nanoparticle and the characteristics of the biological system under study. Similar to traditional cytotoxicity assays, the interplay between NP physicochemical properties—such as type, size, and coating—and biological factors such as cell type profoundly influences the diversity and extent of DEGs resulting from this NP‒cell interaction. For example, metal and metal oxide NPs may trigger DEGs related to oxidative stress, DNA repair mechanisms, metabolic reprogramming, or inflammation, depending on their physicochemical properties and biological factors (PPxBF).17,38–41 Nevertheless, unlike traditional cytotoxicity assays that provide a clear and well-defined biological endpoint, transcriptomic data from gene expression analysis offer a vast and complex dataset on the possible mechanisms leading to these biological endpoints. This further reinforces the need to explore complex NP‒cell interactions within an integrative framework such as nanotoxicomics.
In this context, omics studies have revealed that AgNPs elicit diverse molecular pathways depending on the cell model. Chronic exposure of normal human bronchial epithelial cells (Beas-2B) to 10-nm citrate-coated AgNPs (1 µg/mL for six weeks) induces the expression of DEGs associated with oxidative stress, extracellular matrix remodeling, and DNA damage, potentially contributing to carcinogenesis.5 In contrast, acute exposure of HepG2 cells to 10 µg/mL AgNPs results in DEGs associated with metabolic pathways such as glutathione biosynthesis, fatty acid metabolism, and glycolysis, highlighting the metabolic adaptability of hepatic cells under oxidative stress.42 Furthermore, transcriptomic analysis of T98G glioblastoma cells exposed to 40 µg/mL 16-nm AgNPs revealed DEGs related to neuroinflammation, suggesting that AgNPs may compromise neuronal cell survival through pathways involving the transcription factor JUN-B, the interleukins IL-6 and IL-8, and mitogen-activated protein kinases (MAPKs).43
Similarly, gold nanoparticles (AuNPs) have been shown to modulate gene expression linked not only to oxidative stress and inflammation but also to important pathological conditions. For example, in human colorectal adenocarcinoma (Caco-2) cells exposed to 14 nm citrate-coated AuNPs, heme oxygenase 1 (HMOX1), heat shock protein 90 kDa beta member 1 (HSP90B1), and carnitine palmitoyltransferase 1A (CPT1A), which are involved in important biological pathways such as fatty acid metabolism, protein processing, and insulin resistance, have been shown to be upregulated.44 Moreover, the authors reported the upregulation of other biologically important genes, such as hemopexin (HPX) and fatty acid synthase (FASN), suggesting a possible role of AuNPs in immunotoxicity and steatosis development. In a comparable manner, studies on MRC5 lung fibroblasts exposed to 20 nm AuNPs revealed DEGs associated with blood stress defense, hemostasis, coagulation, and sterol metabolism.45 Furthermore, the upregulation of microRNA-155 and the downregulation of PROS1 support a possible proinflammatory or procoagulant state, potentially leading to conditions such as autoimmune disorders, thrombosis, or carcinogenesis. In addition, in gastric adenocarcinoma (SNU5), AuNPs induce DEGs related to immune pathways, including Fcγ receptor-mediated phagocytosis and complement cascades, which are influenced by nanoparticle size, surface functionalization, and time of exposure.17
Copper oxide nanoparticles (CuO-NPs) are likely involved in inflammatory-related transcriptomic responses, with the upregulation of oxidative stress markers such as HMOX1, heat shock proteins (eg, HSPA1A), and metallothioneins (eg, MT1X) in both lung cancer and non-cancer cell lines. For example, exposure of the A549 lung cancer cell line to approximately 85.5 μg/mL of 46 nm uncoated CuO-NPs for 24 hours led to the overexpression of HMOX1, HSPA1A, and MT1X, whereas JUN expression decreased.37 Interestingly, the upregulation of HMOX1, HSPA1A, MT1X, and JUN was also observed in noncancer lung cells (BEAS-2B) exposed to 10 μg/mL of 55 nm CuO-NPs for 8 hours.40 HMOX1, which is involved in oxidative stress and inflammatory protection, appears to be a key gene involved in CuO-NP-induced effects in lung cells, as its overexpression has been observed under all experimental conditions, including in vivo in ICR female mouse models exposed to a concentration of 8 × 105 CuO NPs/cm3 with a size of 29.4 nm at 2 and 6 weeks and 3 months.46 Moreover, overexpression was associated primarily with immune responses at 2 weeks, cell cycle regulation at 6 weeks, and cell adhesion and migration at 3 months. These findings suggest that independent of PPxBF, CuO-NPs can induce broad oxidative stress and inflammation, along with possible DNA damage, which may be linked to systemic conditions such as cystic fibrosis due to impaired regulation of oxidative stress, as observed with disruptions in HMOX1.47
In contrast, cerium oxide nanoparticles (CeO2-NPs) have shown selective effects depending on the cell type. In lung cancer cells (A549), CeO2-NPs upregulated genes linked to oxidative stress and inflammation, whereas in noncancer bronchial epithelial cells, they mitigated oxidative damage, highlighting their potential as selective anticancer agents.16 Furthermore, these formulations seem to repress the ID1 pathway in A549 cells, which is associated mainly with uncontrolled cell growth and cell chemoresistance in various types of cancers, thus indicating the possible suppression of cancer cell proliferation by this type of metal oxide NPs.16,48,49 Conversely, non-transcriptomic validation studies on 9.52 nm CeO2-NPs revealed the mitigation of HMOX1 and SOD2 gene expression in Beas-2B cells pretreated with the oxidative stress-promoting agent KBrO3.50 These findings suggest the potential to counteract oxidative stress in noncancer cells. Remarkably, compared with the more robust RNA expression profiles obtained for other types of metal and metal oxide NPs in both healthy and tumoral cells, the effects of CeO2-NPs in these RNA-Seq studies revealed low to moderate effects on DEGs. As the sizes and concentrations of CeO2-NPs used in transcriptomic and non-transcriptomic validation studies are similar, a selective cytotoxic effect on lung cancer cells may be considered. In healthy models, CeO2 NPs help counteract oxidative stress; however, in tumor cells, they can exert a moderate cytotoxic effect by increasing susceptibility to ROS and suppressing cancer survival mechanisms.
TiO2-NPs are associated with inflammation, genotoxic stress, and DNA damage, with differential effects depending on the cell type. As illustration, bone marrow stromal cells exposed to TiO2-NPs exhibited increased expression of the reported DNA damage response genes RPA3 and CINP and the cell cycle arrest gene PRKACA on bone marrow stromal cells exposed to 100 μg/mL TiO2-NPs, regardless of NP size.51 Similarly, lower concentrations of 10 mg/mL of 25-nm TiO2-NPs downregulated the expression of cell cycle arrest genes, including CDK2, CCNA1, and CCNB1, in human keratinocyte cells. Notably, the upregulation of p53 and downregulation of p21, a key master regulator of the cell cycle, were observed in human keratinocytes but were not significantly deregulated in bone marrow stromal cells.15 In contrast, in cancer glioblastoma cells, 20 mg/mL 18.18 nm TiO2-NPs produced a different transcriptomic profile, as observed in the deregulation of inflammation, and cell‒cell interaction-related genes, such as c-Fos, interleukin 8, JUN-B, and Integrin beta 34. Overall, the DEGs associated with TiO2-NP exposure suggest that TiO2-NPs can promote cell death mechanisms in both cancer and noncancer cells by causing cell cycle arrest at the S‒G2/M phase, interfering with cell preparation for mitosis. At the systemic level, this could lead to loss of organ function, chronic inflammation, and even malignant transformation. As TiO2-NPs are among the most commercialized NMs, it is crucial to conduct further nanotoxicomics research to understand the potential risks of acute and chronic exposure to TiO2-NPs to human health.
The cellular effects of zinc oxide nanoparticles (ZnO-NPs) are complex and have become evident in studies such as that of Cao et al (2022),52 where 20 nm ZnO-NPs at 64 μg/mL affected multiple organelles in a 3D Caco-2 spheroid model. Like TiO2-NPs, ZnO-NPs altered immune response genes (JUN, MAP3K14) and oxidative stress mediators (Hsp40), alongside genes related to DNA damage and repair (GADD45G, PARP1, LMNB1). These changes suggest involvement in mitochondrial, endoplasmic reticulum, and lysosomal dysfunction, as evidenced by deregulated mitochondrial permeability; cytochrome C release; ER stress-induced transcription; and autophagy biological processes. The exposure of K562 leukemic cells to 40 μg/mL 32-nm ZnO-NPs triggered a stress response, with the upregulation of oxidative stress genes (eg, SOD3, Hsp70, and FOS) and the downregulation of survival genes (eg, VEGFA, IGF1, and BCL2A1), which activated the NF-κB and JAK-STAT pathways.35 Similarly, exposure of A549 cells to 141-nm and 142-nm ZnO-NPs at 15 µg/mL induced oxidative stress and cellular defense pathways, with upregulation of MT1H, MT1E, SLC30A1, HSPA1A, SOD1, and CAT, alongside the stress-related genes FOS and JUN. Activation of the B-cell receptor signaling pathway and HMGB1 suggests inflammation and immune responses, with the Nrf2 pathway being initially activated but repressed after 24 hours 12. The upregulation of oxidative stress genes and downregulation of survival genes, along with the activation of the NF-κB and JAK-STAT pathways, suggest that ZnO-NPs induce complex inflammatory and immune responses. The activation and subsequent repression of the Nrf2 pathway further highlight the dynamic nature of nanoparticle interactions, which traditional nanotoxicity methods may not capture. These transcriptomic data reveal the mechanisms by which ZnO-NPs induce oxidative stress, inflammation, and mitochondrial-dependent apoptosis in various cancer cell types, suggesting their potential as anticancer agents. However, the lack of recent studies on the cytotoxicity of ZnO-NPs in healthy human cells highlights the need for further research to assess their impact on normal cell functions and safety profiles.
Current transcriptomic studies on the toxicity of metal and metal oxide NPs highlight the differential expression of key genes in a PPxBF-dependent manner. Although gene expression varies depending on the type of NP and the cell type, a general mode of action can be identified. Metallic NPs typically interact with cellular receptors, promoting their internalization via a trojan horse mechanism. Once inside the cell, they are released from the endosome, where they dissolve and oxidize in a slightly acidic environment, generating metal ions that lead to the production of ROS.28 This oxidative stress activates transcription factors and epigenetic modulators (eg, acetylation, methylation, and phosphorylation), altering gene expression.53 This transcriptomic mode of action for each formulation is summarized in (Figure 2).
 |
Figure 2 Side to side transcriptomic mode of action of metal and metal oxide NPs on cancer and non-cancer human cells. (A) DNA damage and regulation: Metal and metal oxide nanoparticles can directly interact with DNA, causing damage such as strand breaks or base modifications, (B) Cell cycle disruption: Exposure to nanoparticles can perturb the normal progression of the cell cycle, leading to aberrant cell cycle regulation and cell cycle arrest., (C) Mitochondrial dysfunction: Metal and metal oxide nanoparticles can interfere with mitochondrial function, disrupting oxidative phosphorylation and ATP production, (D) Antioxidant defense: Exposure to nanoparticles can disrupt the cellular antioxidant defense mechanisms, leading to oxidative stress and damage to biomolecules such as DNA, proteins, and lipids, (E) Metabolism imbalance: Nanoparticle exposure can alter cellular metabolism, affecting pathways involved in energy production, nutrient utilization, and biosynthesis of amino acids, (F) Inflammation: Metal and metal oxide nanoparticles can induce inflammatory responses in cells and tissues, leading to the secretion of pro-inflammatory cytokines, chemokines, and other inflammatory mediators, (G) EMT: Exposure to nanoparticles can induce EMT, a process by which epithelial cells acquire mesenchymal characteristics, such as increased motility and invasiveness, (H) Cell survival: Exposure to NPs can promote cell survival mechanisms of healthy cells, in particular those related to cell growth and proliferation. |
These pathways diverge based on PPxBF, but similarities, such as oxidative stress and inflammation, are consistently observed. This suggests that while different NPs share similar biological endpoints, their mechanisms at the genetic level are not necessarily the same.
Although transcriptomic analysis provides an initial understanding of the NP-system interaction landscape and potential biomarkers, notably, a differentially expressed gene does not necessarily translate into a functional protein. It reflects only the amount of mRNA produced from a specific gene compared with control. Following transcription, mRNAs undergo maturation and may be translated into proteins by ribosomes. To address the challenges of understanding the complex relationship between gene expression and protein synthesis in NP-system interactions, additional nanotoxicological approaches, such as proteomics and metabolomics, are needed to validate the most promising biomarkers for future applications.
Proteomic Analysis
Proteomics involves the large-scale study of proteins within biological systems, focusing on the identification, characterization, and quantification of proteins in cells, tissues, and organisms. Unlike transcriptomics, which measures gene expression, proteomics provides a deeper understanding of biological interactions by revealing changes in protein expression, modifications, and interactions in response to stimuli such as NP exposure. Proteomics complements transcriptomics by validating biomarkers and offering a more comprehensive view of cellular metabolism and stress responses, which might occur after transcription processes, particularly in the context of NP exposure.16
Notably, proteomic techniques, such as phosphoproteomics and redox proteomics, enable the evaluation of posttranslational protein modifications, providing detailed insights into cellular functions. These techniques are particularly valuable for studying the effects of NPs, as stress responses often involve rapid protein modifications or changes in protein function and structure.54,55
Early proteomic studies utilized 2D gel electrophoresis to separate and quantify proteins based on spot intensity, followed by sequencing methods such as Edman degradation and mass spectrometry (MS).56 Advances in tandem MS and computational techniques, such as database search algorithms, have set the foundations for bottom-up proteomics to become a widely used approach. This method involves protein extraction, digestion, peptide separation via liquid chromatography, and identification via bioinformatics tools that match mass spectra to protein databases. Bottom-up proteomics is a highly sensitive approach capable of detecting peptides at low concentrations and is relatively accessible, making it an attractive technique for nanotoxicomics studies.
Recently, top-down proteomics has gained attention in the study of NP interactions in clinical research.57 Unlike bottom-up proteomics, which analyzes peptides, top-down proteomics studies intact proteins, providing a full view of protein isoforms, variants, and posttranslational modifications. This method involves extracting proteins from biological samples, separating them by size or charge (via liquid chromatography or electrophoresis), and analyzing their mass via electrospray ionization and fragmentation techniques such as collisional activation dissociation or high-energy collisional dissociation. While top-down proteomics offers a more detailed characterization of proteins, it faces challenges such as complex instrumentation, larger sample requirements, and difficulties in efficiently separating large proteins. Despite these limitations, this method can still be utilized in nanotoxicomics studies, particularly when a detailed analysis of posttranslational modifications or specific NP-protein binding sites is needed.
Proteomic Insights Into the Mode of Action of Metal and Metal Oxide Nanoparticles on Human Cell Lines
Proteomic research offers valuable insights into the complex interactions between NPs and cellular proteins. Some studies have shown that metal and metal oxide NPs can trigger unique molecular responses that may differ from those observed via transcriptomic analysis. For example, Gao et al (2022)58 reported that after exposure of human induced pluripotent stem cell (iPSC)-derived hepatocyte-like cells (HLCs) to 20 nm citrate-coated AgNPs (10 μg/mL) for 24 hours, 557 DEGs were identified via transcriptomics, whereas 665 differentially expressed proteins (DEPs) were identified via proteomics, with only 29 of them overlapping. This discrepancy may arise from differences between gene expression and protein translation, as not all mRNAs are translated into proteins. Additionally, proteins can undergo posttranslational modifications, such as phosphorylation or acetylation, which are not reflected in transcriptomic data.55 Proteomics may also detect a broader range of proteins, including those present at low levels or with post-translational modifications that impact their function, whereas transcriptomics captures only mRNA expression. Besides, Gene Ontology (GO) analysis revealed that both the DEGs and DEPs influenced similar biological functions, including inflammation, cell metabolism, and oxidative stress. Moreover, the authors found that metallothionein and heat shock proteins were among the most upregulated genes and proteins, which linked to the observed upregulation of the NRF2-mediated Oxidative Stress Response canonical pathway, suggesting a coordinated response to overcome cellular stress. However, a significant upregulation of SPINK1 and FAT10 cancer signaling pathways was also observed, indicating that cells are unable to cope with the damage induced by AgNPs, leading to chronic liver disease or hepatocellular carcinoma.58
Likewise, a differential expression of proteins related to cellular metabolism and oxidative stress response, such as MT1G, HMOX1, and PDHA2, was shown as a response to oxidative stress in hepatocellular carcinoma HepG2 cells exposed to 2.5 μg/mL of 20-nm and 60-nm citrate-coated AgNPs. This was confirmed by a size-dependent upregulation of metabolic pathways, where smaller NPs (20 nm) activated pyruvate metabolism and the tricarboxylic acid cycle (TCA), whereas larger NPs (60 nm) affected mainly endocytosis and TGF-beta receptor signaling. In contrast, normal liver cells (L02) respond differently, with a focus on promoting cell survival as demonstrated by the overexpression of proteins involved in macroautophagy, DNA repair, and regulation of biological pathways.59 These results suggest a cytoselective mechanism in liver cells, where cancer cells promote AgNPs internalization by an altered metabolic state while non-cancer cells can maintain cellular homeostasis by displaying several defense mechanisms such as DNA repair and blocking cellular internalization.
In lung models, AgNPs also exhibited cell type-dependent effects. Holmila et al (2021)54 reported that 25-nm PVP-coated AgNPs (20 μg/mL) activated oxidative stress responses in both AgNP-sensitive (Calu-1) and resistant (NCI-H358) lung cancer cells, involving proteins such as HMOX1, SIRT3, and heat shock proteins. These proteins were related to the overactivation of NRF2, sirtuin pathways, and mitochondrial dysfunction pathways, suggesting that respiratory exposure may contribute to the development of respiratory diseases such as lung cancer, asthma, and COPD.38,60
Proteomic studies on 20-nm AuNPs at concentrations of μg/mL also highlighted mitochondrial dysfunction as a key cytotoxic mechanism in THP-1 cells, as evidenced by the downregulation of proteins involved in oxidative phosphorylation and autophagy, such as CYT C, AIF, and mTOR, leading to apoptosis at low concentrations and necrosis at high doses.18 Conversely, on gastric cancer cells, AuNPs triggered selective antitumor mechanisms via phagosome- and FcγR-mediated pathways, with rod-shaped AuNPs exhibiting stronger apoptotic effects than spherical AuNPs.17
TiO2-NPs affect mitochondrial function and protein translation. For example, Montalvo-Quiros et al (2019)15 reported that 25-nm TiO2-NPs upregulated the mitochondrial antioxidant protein PRDX5 while inhibiting SOD2 and ATP5L in human keratinocyte cells, indicating mitochondrial disruption. TiO2-NPs also interfered with the cell cycle by downregulating proteins involved in cell division, leading to cell cycle arrest. These findings suggest potential disruption of the electron transport chain, leading to mitochondrial dysfunction and increased oxidative stress in a human keratinocyte cell line (HaCaT). In addition, the overexpression of the eukaryotic translation initiation factor EIF4E and the inhibition of the eukaryotic translation initiation factors EIF4G2 and EIF4H imply that TiO2-NPs can also play a pivotal role in translation. Notably, this formulation did not induce a significant amount of apoptosis, but significant cell cycle arrest at the S‒G2/M phase was mediated by the downregulation of the CDK2, CCNA1, and RanGAP1 proteins. This mechanism of impaired cell cycle progression induced by TiO2-NPs has also been identified by Mathilde Biola-Clier et al (2020),55 who proposed that the dysregulation of the phosphorylation of proteins related to disturbances in the actin cytoskeleton and microtubules, such as KIF4A, TLN2, and ZYX as the pivotal factor leading to this cellular effect in the alveolar lung cancer cell line A549. In addition, several proteins involved in RNA maturation, the transport of molecules and the inhibition of apoptosis, such as ADAR, SQSTM1, and TPX2, were also detected confirming the potential of TiO2-NPs to influence epigenetic machinery.
Research on CuO-NPs and CeO2-NPs in human cells is still limited, with only a handful of studies investigating their nanotoxicological effects in the past decade. Despite this scarcity, the findings from these studies are highly significant. CuO-NPs, especially larger particles, have detrimental effects linked to the deregulation of key cellular pathways, including mTOR signaling, leukocyte extravasation, and protein ubiquitination.61 These findings suggest that CuO-NPs can influence protein translation and potentially contribute to carcinogenesis through mechanisms such as IF2 signaling and remodeling of epithelial adherens junctions. In contrast, 4-nm CeO2-NPs demonstrated cytoprotective effects in HepG2 cells at the phosphoproteomic level, particularly by reducing phosphopeptides associated with oxidative stress, cell growth, angiogenesis, and carcinogenesis.62 Both CuO and CeO2 NPs appear to modulate the mTOR pathway, which plays a central role in regulating cellular functions such as growth, metabolism, survival, and autophagy. At the systemic level, alterations in mTOR signaling caused by both CuO-NPs and CeO2-NPs could exacerbate tumor progression. Therefore, further studies are essential to investigate the proteomic effects of these nanoparticles on both healthy and cancerous cells.
Proteomics is a powerful tool for revealing the functional pathways involved in NP-induced cytotoxicity. The key pathways influenced by metal and metal oxide NPs include NRF2-mediated oxidative stress, mitochondrial dysfunction, and metallothionein binding. On the other hand, metal oxide NPs significantly impact cellular metabolism by modulating the TCA cycle and mTOR signaling pathways, among other processes. However, proteomic studies on NPs remain limited due to their high cost and complexity. Further research is needed to fully understand the mechanisms of NP toxicity at the protein level and the influence of the physicochemical properties of NPs on the cellular responses of different cells and tissue types (Figure 3).
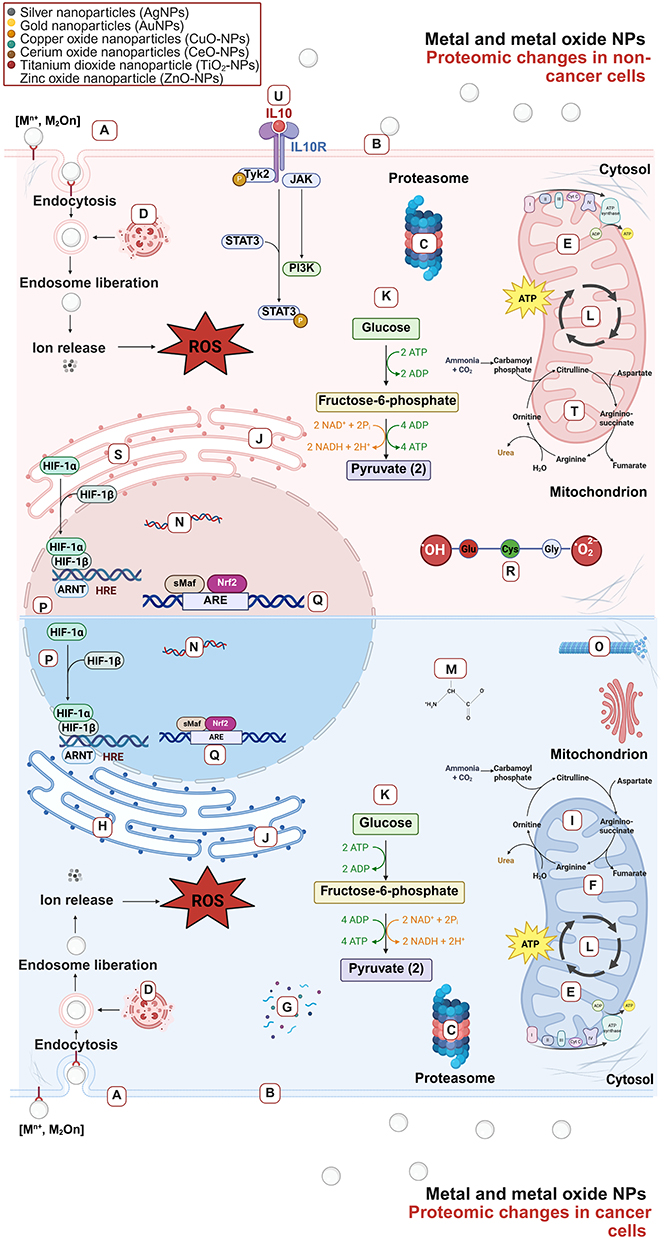 |
Figure 3 Side to side proteomics mode of action of metal and metal oxide NPs on cancer and non-cancer human cells. Metal and metal oxide NPs elicit wide effects on human proteomic mechanisms, impacting the expression and regulation of proteins of cancer and non-cancer cells. Besides, this proteins can be related to several important bio-logical mechanism such as: (A) Regulation of endocytosis, (B) mTOR signaling pathway, (C) regulation of proteolysis, (D) lysosome, (E) oxidative phosphorylation, (F) apoptosis, (G) metallothionein binds metal, (H) energy and lipid metabolism, (I) mitochondrial dysfunction, (J) gluconeogenesis, (K) glycolysis, (L) TCA cycle, (M) biosynthesis of amino acids, (N) DNA repair and transcription, (O) cytoskeleton, (P) HIF1α signaling, (Q) NRF2 signaling, (R) GSH detoxification, (S) COX2, (T) urea cycle, (U) IL10 signaling. |
Metabolomic Analysis
Metabolites are small organic molecules produced through metabolic processes in living organisms that play key roles in biological functions and cellular activities. These molecules include amino acids, lipids, sugars, nucleotides, and other small compounds. Metabolites can act as building blocks for larger molecules or as regulators of enzymes. Researchers utilize metabolomic approaches, either targeted or untargeted, to study these molecules. They often employ techniques such as nuclear magnetic resonance (NMR), which is nondestructive and provides structural information. These techniques are complemented by mass spectrometry (MS) approaches, which offer increased sensitivity and excel in detecting low-abundance metabolites.63
Untargeted metabolomics aims to quantify as many metabolites as possible without a predefined focus, allowing for the discovery of new biomarkers by comparing metabolite profiles before and after nanoparticle exposure. Targeted metabolomics, on the other hand, focuses on measuring specific metabolites, such as lipids, amino acids, or nucleotides, to test specific hypotheses or validate biomarkers. While untargeted metabolomics offers a broader view, it may have lower sensitivity and specificity than targeted approaches do, making data interpretation more challenging. The choice between these approaches depends on research goals: untargeted metabolomics is ideal for exploratory studies, whereas targeted metabolomics is better suited for hypothesis-driven investigations.64
Lipidomics, a targeted approach within metabolomics, plays a critical role in examining the effects of NPs on lipid profiles in human cells. Lipids are integral to cell membrane integrity, energy storage, and signaling processes. By analyzing changes in lipid expression across classes, such as fatty acids, glycerophospholipids, sphingolipids, and sterol lipids, lipidomics provide insights into the mechanisms of NP toxicity. For example, studies have shown that silver nanoparticles disrupt glycerophospholipid metabolism, contributing to oxidative stress and altered cellular function. These findings not only enhance our understanding of NP toxicity but also inform broader applications, such as nanoparticle-based drug delivery systems and therapeutic strategies.65
Metabolomic Insights Into the Mode of Action of Metal and Metal Oxide Nanoparticles in Human Cell Lines
Metabolomics provides a direct measure of small molecules, including metabolites such as amino acids, nucleotides, lipids, and small peptides, which reflect the biochemical state of a cell or organism after exposure to metal and metal oxide nanoparticles. It is considered a functional endpoint in nanotoxicomics because it captures downstream effects from transcriptomics and proteomics, linking molecular changes to observable phenotypes and potential clinical outcomes. Metabolomics can reveal adaptive metabolic changes in response to cellular stress, such as reductions in antioxidant levels like glutathione and disruptions in lipid metabolism. Additionally, it can detect metabolite signatures associated with oxidative DNA damage, which may ultimately contribute to cell death.16,66,67 For example, Swee Ling Lim et al (2019)6 demonstrated that 10 and 50 mg/mL ZnO-NPs significantly repressed the production of amino acids, such as glutamic acid and glycine, which are critical components of glutathione (GSH) in healthy bronchial cells, suggesting oxidative stress-related protein dysfunction. ZnO-NPs also disrupted the TCA cycle and reduced ATP production, impairing cellular energy metabolism. This accumulation of toxic molecules and proteins led to the overproduction of ROS and triggered apoptosis. In contrast, 2.5 and 5.0 mg/mL CuO-NPs (<50 nm) stimulated ceramide production in human colon carcinoma cells, promoting autophagy and oxidative stress.68
The effects of the physicochemical properties of NPs on glutathione and the TCA cycle have also been explored in other cell models and engineered nanoparticles. For example, in human dermal fibroblasts (HDFs), exposure to 39.4 μg/mL of 20 nm AuNPs and AgNPs elicit different metabolomic profiles after 4, 8, and 24 hours. Although both formulations involve the deregulation of several key metabolites and pathways, screening criteria have identified glutathione and uridine metabolites as the key agents affected by AuNPs. Moreover, for AgNPs, malic acid has the greatest effect on the metabolism of HDFs, particularly on the TCA cycle.41 Interestingly, other formulations of small 10-nm AgNPs and 4.6-nm ZnO-NPs have also been shown to deregulate the TCA cycle, as one of the principal cytotoxic mechanisms in lung and liver cancer cells, at concentrations of 128 µg/mL and 5 µg/mL, respectively.16,42 The TCA cycle, also known as the Krebs cycle, serves as a central hub for cell metabolism in the mitochondria of eukaryotic cells and can receive input from various substrates to promote the oxidation of acetyl-CoA to generate energy in the form of ATP. In this sense, the ability of nanoparticles to act as upstream regulators of the TCA cycle can affect the production of significant amounts of energetic biomolecules, such as ATP, ADP, and NAD+, which are common biological endpoints of nanotoxicity.42 Disruption of energy production and cellular homeostasis, particularly through the inhibition of TCA cycle intermediates such as citrate, glutamic acid, and aspartic acid, are common findings among NP studies.
It has been previously described that cells can switch to alternative energy pathways, such as glycolysis, when oxidative phosphorylation is impaired. Furthermore, numerous studies have reported overactivation of glycolysis, pyruvate, and sucrose metabolism after NP exposure.41,69,70 Additionally, key amino acids involved in glycolysis, such as alanine, glycine, and serine, have also been shown to be deregulated, indicating broader metabolic disruption.
On the other hand, mitochondrial dysfunction and energy imbalance are often linked to proinflammatory processes. In this context, studies on HDFs and prostate cells exposed to AuNPs revealed an accumulation of oxidative stress markers and eicosanoids, which are proinflammatory molecules. Eicosanoids, such as leukotrienes and thromboxanes, can modulate ROS production, activate transcription factors such as NF-κB, and induce cell death via pyroptosis.71 Similarly, small CeO2-NPs can trigger inflammation through the upregulation of eicosanoid levels.72
Lipidomics, a key tool in nanotoxicomics, can reveal how NPs affect the composition and function of the cellular membrane. Metal and metal oxide NPs can cause lipid peroxidation, especially in polyunsaturated fatty acids, leading to cell membrane damage and ferroptosis, a type of regulated cell death. For example, Adeyemi et al (2020)73 reported that high concentrations of AgNPs and Fe3O4 NPs induced lipid peroxidation, leading to necrosis in HepG2 cells. CuO-NPs in colorectal cancer cells have also been shown to trigger lipid alterations, promoting autophagy over ferroptosis.68
Metabolomics is a powerful approach for understanding the systemic effects of NPs. This review helps to validate toxicological mechanisms by revealing disruptions in energy pathways, such as the TCA cycle, glycolysis, and oxidative phosphorylation. These disturbances have been shown to lead to altered lipid and amino acid production, which are directly linked to inflammation and cell death mechanisms, including apoptosis, autophagy, pyroptosis, and ferroptosis. The integration of metabolomics with other omics approaches in the context of nanotoxicomics can provide a comprehensive understanding of the key molecules affected as a result of NP-cell interactions and their relationships with observed biological endpoints, as summarized in (Figure 4).
 |
Figure 4 Side-to-side metabolomics mode of action of metal and metal oxide NPs on cancer and non-cancer human cells. (A) Adaptive response to epigenetic changes and nucleotide deregulation, (B) Adaptive changes to cellular and intracellular NP-membrane interaction, (C) Adaptive changes to ROS-mediated mitochondrial dysfunction, (D) Adaptive responses to oxidative stress, (E) Adaptive changes to energy depletion, (F) Adaptive changes to proinflammatory cytokines. |
Proposing Integrative Strategies to Move From Single to Multiomics Approaches
The bioinformatics era has revolutionized how biological data from high-throughput technologies, such as those that can be applied to study nanoparticle toxicity, are analyzed and interpreted. By integrating data from transcriptomics, proteomics, metabolomics, and lipidomics, researchers can uncover deeper biological insights and systemic responses to NPs. This holistic approach enhances nanodrug development, biomarker discovery, occupational exposure risk assessment, and intelligent design of safer NPs.
Integrating multiple omics datasets via bioinformatic pipelines, such as differential expression and network analysis, may help to reveal the complex mechanisms underlying NP toxicity rather than focusing solely on isolated effects. For example, techniques such as weighted gene correlation network analysis (WGCNA) have proven to be effective in correlating experimental data with large databases, enabling comparisons with other NPs that share similar or different physicochemical properties via R-3.5.0.74 In this context, this multiomics study involving proteomics, phosphoproteomics, and metabolomics of seven different NMs revealed strong positive correlations between agglomerate size and pathways such as cytokine signaling, phagocytosis, the NRF2-mediated oxidative stress response, and mitochondrial dysfunction. Conversely, the results revealed a negative correlation with toxicological endpoints such as LDH release, driven by key molecules such as phosphatidylcholines, asparagine, and spermidine, which were highly consistent across the different omics datasets in mouse alveolar macrophages (NR8383). Therefore, this study provides an integral mechanism of NM-induced toxicity at different molecular levels as well as important biological markers within the context of nanotoxicomics. Without this integration, there is a significant risk of missing underlying toxic effects, potentially leading to false-positive or false-negative interpretations of NP toxicity.
Significance-based modules integrating the transcriptome and epigenome (SMITE) is another technique that has also been applied to integrate multilayered datasets from transcriptomics, microRNA, and methylome analyses. This approach was used to study focal adhesion and cell adhesion mediated by the PI3K–Akt signaling pathway in response to ten different carbon nanoparticles in the Beas-2B, A549, and THP-1 cell lines.75
To further enhance robustness and reduce biases in multiomics integration, combining network analysis with machine learning algorithms offers promising results. For example, Shin et al (2021)19 15 employed a k-NN algorithm alongside differential expression pathway analysis to integrate triple-omics data. These findings highlighted microglial activation and dysfunction, which are mediated by ROS and altered glucose uptake, in primary rat microglia and BV2 cells exposed to silica-coated cobalt ferrite magnetic nanoparticles. Similarly, Zhang et al (2022)76 used multilayered integration of transcriptome and proteome data to reveal mechanisms of cancer progression, including epithelial‒mesenchymal transition (EMT), hypoxia, and TNF-α signaling via NF-κB, in workers exposed to black carbon and diesel nanoparticles.
As research into NP safety advances, bioinformatics platforms such as MetaboAnalyst will likely become increasingly popular for integrating multiomics datasets, leading to more comprehensive insights into NP toxicity and safer nanoparticle design.77
Multiomics Insights Into Occupational and Consumer Related Exposure to Metal and Metal Oxide Nanoparticles
Occupational exposure to metal-based and metal oxide nanoparticles (OEtNPs) presents significant health risks associated with prolonged contact in industrial and laboratory environments. Conventional biomonitoring techniques and cytotoxicity assays often fail to capture subtle molecular perturbations induced by chronic exposure. Nanotoxicomics, integrating multiomics approaches, provides a comprehensive framework to detect these effects and identify biomarkers relevant for worker safety. By uncovering the mechanistic pathways affected by nanoparticles—such as oxidative stress, inflammation, DNA damage, mitochondrial dysfunction, and altered metabolic signaling—nanotoxicomics enables a direct connection between molecular-level perturbations observed in controlled in vitro and in vivo studies and potential health outcomes in occupational settings. This mechanistic integration is crucial for translating laboratory findings into actionable occupational risk assessment and biomonitoring strategies that can inform the establishment of occupational exposure limits (OELs).
Transcriptomics can serve as a first-line approach, providing early detection of gene-level perturbations in response to NP exposure. For instance, DEGs related to oxidative stress, inflammation, DNA damage, and protein folding can act as biomarkers in exposed workers, even before cytotoxicity occurs. Once these early markers are identified, they can guide the selection of targets for downstream proteomic and metabolomic analyses, which offer functional and biochemical validation, respectively.
In this regard, the expression of proteins involved in oxidative stress (eg, HMOX1, MT1G, SOD), mitochondrial dysfunction (eg, SIRT3, PRDX5), autophagy (eg, CYTC, AIF, mTOR), and cell cycle regulation (eg, CDK2, CCNA1, and RanGAP1), as well as proteins affecting translation and RNA maturation, such as EIF4E, ADAR, and SQSTM1, can provide more detailed insights into the molecular pathways and potential adverse effects resulting from acute and chronic exposure to metal and metal oxide nanoparticles.
Metabolomics provides a functional readout of these molecular perturbations by detecting small-molecule signatures associated with oxidative stress, mitochondrial dysfunction, and lipid peroxidation. Disruptions in glutathione metabolism, the tricarboxylic acid (TCA) cycle, and glycolysis have been consistently reported in dermal fibroblasts, bronchial cells, and hepatocytes exposed to metal and metal oxide nanoparticles, providing key indicators of OEtNP exposure that are particularly relevant in dermal, ocular, and respiratory occupational settings. These metabolic alterations can manifest as reductions in antioxidant defenses, accumulation of reactive oxygen species, and the release of proinflammatory eicosanoids—all of which may be translatable into biomarker discovery in occupational biofluids such as saliva, plasma, or exhaled breath condensate.
Despite its potential, multiomics studies in OEtNPs area remain scarce, emphasizing the urgent need for expanded research on the impact of NPs in occupational settings.78 For example, Araujo et al (2023)79 conducted a multiomics study on Brazilian workers in the welding and jewelry assembly industries. Using MetaboAnalyst 4.0 to integrate proteomic saliva and metabolomic plasma datasets, they reported that high levels of metals such as Ni, Cu, Zn, Sn, Sb, and Pb were linked to 11 key biological pathways. Most of these pathways are related to amino acid regulation, but they also identify pathways involved in disease progression, such as central carbon metabolism in cancer, ABC transporters, and arachidonic acid metabolism. Cancer cells are known to reprogram carbon metabolism to support rapid growth, relying on amino acids such as glycine, serine, and methionine for energy production and biosynthesis. Changes in carbon metabolism can also affect arachidonic acid availability, influence lipid biosynthesis, promote tumor growth and angiogenesis.80 Additionally, deregulation of ABC transporters has been linked to neurodegenerative diseases and drug resistance, including conditions such as Alzheimer’s disease and Parkinson’s disease.81
Another example of multiomics in occupational exposure involves metabolomic profiling and gene expression analysis in truck drivers exposed to diesel exhaust. Diesel nanoparticles, often composed of metals such as Fe, Ni, and Zn, were studied by Walker et al (2022)82 showing that while no significant metabolic changes were detected in the plasma samples, the multiomics analysis revealed pathways related to oxidative stress, endothelial dysfunction, and inflammation. These pathways have been implicated in cardiopulmonary diseases such as COPD, asthma, and coronary artery disease. This case highlights the value of nanotoxicomics in revealing molecular mechanisms of toxicity that may remain undetected via conventional non-omics approaches, providing critical information for developing occupational biomonitoring strategies and OELs.
A more recent example that exemplifies the integration of this nanotoxicomics framework in biomonitoring involves users of electronic nicotine delivery systems (ENDS). This study has demonstrated that puff volume-dependent exposure to metallic nanoparticles from ENDS aerosols can induce metabolomic and proteomic alterations in salivary samples enhancing the expression of inflammatory cytokines like TNFα, IL-1β, and IL-6 and oxidative makers such as MDA, SOD, 8-OHdG. Moreover, by performing a joint pathway analysis using MetaboAnalyst (version 5.0) for significant proteins and metabolites, the authors identified ABC transporters and tyrosine metabolism as among the most enriched pathways, with their significance increasing in a puff volume–dependent manner. In addition, the use of this integrative framework enabled the generation of a protein–metabolite–disease network, in which schizophrenia, Alzheimer’s disease, Canavan disease, and certain respiratory conditions (eg lung cancer, idiopathic pulmonary fibrosis, and COPD) were predicted as the most likely outcomes in participants, particularly those in high puff volume group.83
These findings emphasize the importance of adopting a stepwise nanotoxicomics framework, as it not only enables the identification of early molecular perturbations but also bridges mechanistic understanding with practical occupational health assessment. Continued integration of multiomics approaches into regulatory and monitoring strategies will be essential for establishing evidence-based OELs, improving worker safety, and promoting responsible use of metal-based nanoparticles.
Nanotoxicomics Into the Smart Design of NPs
Aside from unveiling the complex mechanisms underlying NP cytotoxicity in biological systems, nanotoxicomics can also provide valuable insights for the smart design of NPs. In this regard, transcriptomics, proteomics, and metabolomics can be employed to improve the precision of in vitro models for high-throughput screening of engineered NPs and nanotherapeutics. Moreover, nanotoxicomics serves as an excellent tool to quantitatively extrapolate in vitro results to the systemic level.84 To this end, several new methods, such as single-cell multi-omics, are gaining popularity in the research community. Among the advantages offered by single-cell multi-omics are detailed evaluations at high spatial resolution, enabling the investigation of multiple omics layers while maintaining spatial consistency. Additionally, new platforms, such as microfluidic-based technologies, have accelerated the development of smart NPs with specific physicochemical properties and high biocompatibility for biomedical applications.85
As the most relevant and recent examples, the development of safer metallic and metal oxide NPs has employed multi-omics strategies to guide smart design. As illustration, the CompSafeNano project, funded under the European Union’s Horizon 2020 program, which integrates nanoinformatics, computational modelling, and predictive toxicology to enable the design of safer NMs from the earliest stages of development. More precisely, this project has generated atomistic and quantum-mechanical descriptors for a wide array of NMs, including Ag, Au, TiO2, CuO, graphene oxide, and functionalized PLGA-Au NPs; evaluated protein–NP interactions across multiple protein datasets; and developed predictive models for toxicity based on multi-omics data.86 These approaches have informed safer-by-design (SbD) strategies by predicting biocorona formation, identifying molecular initiating events, and anticipating downstream adverse effects, thereby facilitating the production of NPs with reduced cytotoxicity and improved biocompatibility. Additionally, several partners have implemented advanced in vitro models, including 3D co-cultures and inhalation systems, to validate these predictive models experimentally, ensuring that the designed NMs retain desired functionalities while minimizing potential health risks.
Overall, integrating multi-omics data into NP design pipelines not only improves the mechanistic understanding of NP–cell interactions but also enables the SbD development of NPs. This nanotoxicomics approach not only strengthens preclinical evaluation but also provides a stepwise framework that allows the correlation of SbD development with regulatory standards, making it easier to translate findings into real-world applications (Figure 5).
 |
Figure 5 Integrated nanotoxicomics framework linking molecular mechanisms of metal-based nanoparticles to SbD synthesis and applications. Schematic representation of an integrated nanotoxicomics framework, showing how transcriptomic, proteomic, and metabolomic data can be integrated to reveal molecular mechanisms underlying the biological effects of metal-based nanoparticles. These insights support Safe-by-Design synthesis strategies and their translation into biomedical and industrial applications. |
Nanotoxicomics Into Personalized Nanomedicine
Nanotoxicomics is also opening new possibilities for personalized nanomedicine. This approach tailor’s nanotechnology-based treatments to individual patients by considering genetic, demographic, and environmental factors. Consequently, personalized medicine requires a holistic comparison of individual patient traits with larger, well-established clinical and nanotoxicomics databases. This need has marked the importance of bioinformatics tools and, more recently, artificial intelligence (AI) in the field.87 In this context, AI can utilize patient molecular profiling data and query nanotoxicomics databases to select the most appropriate treatment. Furthermore, machine learning (ML) can generate predictive models and monitor patient responses and progression under specific treatments.
Through the development of SbD NPs nanotoxicomics have enhanced personalized nanomedicine in multiple therapeutic areas. For example, in oncology, nanoparticles have shown to improve the isolation of circulating tumor DNA or exosomes in liquid biopsies for genomic and proteomic profiling, guiding targeted therapies and real-time monitoring of chemoresistance.88 On the other hand, nanosensors can display a key role in detecting small changes in protein aggregates or metabolic markers in cerebrospinal fluids, supporting early intervention of patients suffering from neurodegenerative diseases like Alzheimer.89 Furthermore, nanosensors and nanotoxicomics have allowed real-time monitoring of cardiac biomarkers, guiding personalized therapeutic adjustments and sustained release of cardioprotective medications to prevent Heart failure.90 These examples illustrate how integrating nanotoxicomics with patient-specific data can transform preventive, diagnostic, and therapeutic approaches.
Nevertheless, developing personalized nanomedicines using AI and nanotoxicomics presents several challenges, including the limited availability of high-quality datasets and the complexity and heterogeneity of biomedical data. Additionally, significant ethical and privacy concerns arise regarding clinical data management. Finally, integrating these cutting-edge methods into clinical practice may encounter resistance from the medical community due to the need to modify conventional workflows and comply with the rigorous validation procedures required for ML-based clinical tools.91 Overcoming these limitations is therefore essential for the effective implementation of nanotoxicomics in the SbD generation of NMs for AI-assisted personalized nanomedicine.
Conclusion
We introduce nanotoxicomics as an approach to increase our understanding of how NPs interact with biological systems at the molecular level. By integrating high-throughput technologies such as transcriptomics, proteomics, metabolomics, and lipidomics, this multiomics approach offers a comprehensive view of NP-induced toxicity. Each layer provides insights into how NPs affect gene expression, protein function, metabolic pathways, and lipid homeostasis, ultimately revealing how NPs cause cellular stress, dysfunction, and death.
Transcriptomic studies have shown that metal and metal oxide NPs trigger differential gene expression based on physicochemical properties such as size, coating, and shape, with changes linked to oxidative stress, inflammation, and DNA damage. On the other hand, proteomics highlights how NPs disrupt important biological processes by inducing the expression of heat shock proteins, metallothionein’s, and oxidative stress regulators. Finally, metabolomics identifies functional readouts that can translate into observable phenotypes, such as the inhibition of the TCA cycle and glycolysis, which are common in NP-induced cytotoxicity across several models. Integrating these omics layers provides an integral understanding of NP toxicity, with oxidative stress and mitochondrial dysfunction being key features across many types of NPs and omics layers. Integrative studies have confirmed that alterations in genes, proteins, amino acids, lipids, and energy metabolites contribute to the activation and regulation of complex pathogenic biological mechanisms such as EMT, mitochondrial depletion, musculoskeletal deterioration, neurodegeneration, and carcinogenesis at the systemic level.
Despite progress, the application of multiomics to occupational and real-world NP hazards remains limited. Studies on workers exposed to metal NPs have highlighted health risks, including oxidative stress, endothelial dysfunction, and diseases such as cancer, asthma, and cardiovascular conditions in workers and consumers exposed to metal-based nanoparticles. This highlights the need for more nanotoxicomics research on NP-induced health effects in occupational settings.
Future research in nanotoxicomics should aim to apply integrated multiomics approaches in practical real-world and clinically relevant settings. For instance, single-cell and spatial omics technologies could capture the heterogeneity of cellular responses to NPs, while longitudinal multiomics studies in exposed workers may provide dynamic insights into both acute and chronic health risks. The generation of SbD NPs will further support innovations in personalized nanomedicine, including nanosensors, imaging nanodevices, and drug delivery systems, which will be highly valuable in medical disciplines such as oncology, cardiology, neurology, and immunology. These advances will strengthen biocompatibility, enable the construction of predictive multiomics–disease networks, and guide the determination of safe dose–response parameters in nanomedical applications. Beyond biomedical applications, nanotoxicomics will be essential for regulatory frameworks, enabling the establishment of reliable OELs and global nanotoxicomics databases. Advances in machine learning and systems biology will accelerate the integration of complex datasets, potentially leading to the creation of digital replicas of biological systems and to real-time biomonitoring through bionanosensors and wearable devices. This integrated framework is already being employed in recent innovation projects such as CompSafeNano, which report valuable insights into the safer design of NP formulations through the generation of multiomics datasets combined with atomistic and quantum-mechanical descriptors. However, it is essential that we prioritize ethical considerations, such as data privacy and environmental sustainability, to ensure the responsible application of nanotoxicomics findings. Altogether, these directions suggest a future where nanotoxicomics not only help us understand the complex molecular mechanisms involved in NP-induced toxicity but also support safer NP design, personalized nanomedicine, and the sustainable development of nanotechnology.
Abbreviations
NPs, nanoparticles; NMs, nanomaterials; OEtNPs, occupational exposure to nanoparticles; ROS, reactive oxygen species; DEGs, differential expression of genes; AgNPs, silver nanoparticles; TiO2-NPs, titanium dioxide nanoparticles; NP, nanoparticle; PPxBF, physicochemical properties and biological factors; MAPKs, mitogen-activated protein kinases; AuNPs, gold nanoparticles; HMOX1, heme oxygenase 1; HSP90B1, heat shock protein 90 kDa beta member 1; CPT1A, carnitine palmitoyltransferase 1A; HPX, hemopexin; FASN, fatty acid synthase; CeO2-NPs, cerium oxide nanoparticles; ZnO-NPs, zinc oxide nanoparticles; MS, mass spectrometry; HLCs, hepatocyte-like cells; DEPs, differentially expressed proteins; GO, gene ontology; TCA, tricarboxylic acid cycle; HDFs, human dermal fibroblasts; WGCNA, weighted gene correlation network analysis; SMITE, significance-based modules integrating the transcriptome and epigenome; electronic nicotine delivery systems ENDS, SbD, safer by design; OELs, Occupational exposure limits.
Acknowledgments
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. They sincerely acknowledge BioRender for providing a high-quality platform for the creation of scientific illustrations.
Funding
This study was funded by the Challenge-Based Research Funding Program at Tecnológico de Monterrey (Grant No. E004 - EIC-TM – 10-372). Additionally, this work was supported by Azure sponsorship credits granted by Microsoft’s AI for Good Research Lab.
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Zhang L, Haddouti EM, Welle K, Burger C, Kabir K, Schildberg FA. Local cellular responses to metallic and ceramic nanoparticles from orthopedic joint arthroplasty implants. Int J Nanomed. 2020;15:6705–6720. doi:10.2147/IJN.S248848
2. Al-Khedhairy AA, Wahab R. Silver nanoparticles: an instantaneous solution for anticancer activity against human liver (HepG2) and breast (MCF-7) cancer cells. Metals. 2022;12(1):148. doi:10.3390/met12010148
3. Shineh G, Mobaraki M, Afzali E, Alakija F, Velisdeh ZJ, Mills DK. Antimicrobial metal and metal oxide nanoparticles in bone tissue repair. Biomed. Mater. Devices. 2024;2;(2):918–41. doi:10.1007/s44174-024-00159-3
4. Fuster E, Candela H, Estévez J, Vilanova E, Sogorb MA. Titanium dioxide, but not zinc oxide, nanoparticles cause severe transcriptomic alterations in t98g human glioblastoma cells. Int J Mol Sci. 2021;22(4):2084. doi:10.3390/ijms22042084
5. Gliga AR, Di Bucchianico S, Lindvall J, Fadeel B, Karlsson HL. RNA-sequencing reveals long-term effects of silver nanoparticles on human lung cells. Sci Rep. 2018;8(1):6668. doi:10.1038/s41598-018-25085-5
6. Lim SL, Ng CT, Zou L, et al. Targeted metabolomics reveals differential biological effects of nanoplastics and nanoZnO in human lung cells. Nanotoxicology. 2019;13(8):1117–1132. doi:10.1080/17435390.2019.1640913
7. Dianová L, Tirpák F, Halo M, et al. Effects of selected metal nanoparticles (Ag, ZnO, TiO2) on the structure and function of reproductive organs. Toxics. 2022;10(8):459. doi:10.3390/toxics10080459
8. Karlsson HL, Toprak M, Fadeel S. Toxicity of metal and metal oxide nanoparticles. Handbook on the Toxicology of Metals Elsevier. 2022:87–126. doi:10.1016/B978-0-12-823292-7.00002-4
9. Wu L, Wen W, Wang X, et al. Ultrasmall iron oxide nanoparticles cause significant toxicity by specifically inducing acute oxidative stress to multiple organs. Part Fibre Toxicol. 2022;19(1):24. doi:10.1186/s12989-022-00465-y
10. Horie M, Nishio K, Kato H, et al. The expression of inflammatory cytokine and heme oxygenase-1 Genes in THP-1 cells exposed to metal oxide nanoparticles. J Nano Res. 2015;30:116–127. doi:10.4028/www.scientific.net/JNanoR.30.116
11. Omari Shekaftik S, Mehrparvar N, Peivandi Z, Shahtaheri SJ. Risk assessment of occupational exposure to engineered and incidental nanomaterials: differences and challenges. J Nanopart Res. 2024;26(4):67. doi:10.1007/s11051-024-05980-x
12. Habanjar O, Diab-Assaf M, Caldefie-Chezet F, Delort L. 3D cell culture systems: tumor application, advantages, and disadvantages. Int J Mol Sci. 2021;22(22):12200. doi:10.3390/ijms222212200
13. Alijagic A, Scherbak N, Kotlyar O, et al. A novel nanosafety approach using cell painting, metabolomics, and lipidomics captures the cellular and molecular phenotypes induced by the unintentionally formed metal-based (nano)particles. Cells. 2023;12(2):281. doi:10.3390/cells12020281
14. Zhang G, Deng L, Jiang Z, et al. Titanium nanoparticles released from orthopedic implants induce muscle fibrosis via activation of SNAI2. J Nanobiotechnol. 2024;22(1):522. doi:10.1186/s12951-024-02762-4
15. Montalvo-Quiros S, Luque-Garcia JL. Combination of bioanalytical approaches and quantitative proteomics for the elucidation of the toxicity mechanisms associated to TiO2 nanoparticles exposure in human keratinocytes. Food Chem. Toxicol. 2019;127:197–205. doi:10.1016/j.fct.2019.03.036
16. Dekkers S, Williams TD, Zhang J, et al. Multi-omics approaches confirm metal ions mediate the main toxicological pathways of metal-bearing nanoparticles in lung epithelial A549 cells. Environ Sci Nano. 2018;5(6):1506–1517. doi:10.1039/C8EN00071A
17. Fan L, Wang W, Wang Z, Zhao M. Gold nanoparticles enhance antibody effect through direct cancer cell cytotoxicity by differential regulation of phagocytosis. Nat Commun. 2021;12(1):6371. doi:10.1038/s41467-021-26694-x
18. Gallud A, Klöditz K, Ytterberg J, et al. Cationic gold nanoparticles elicit mitochondrial dysfunction: a multi-omics study. Sci Rep. 2019;9(1):4366. doi:10.1038/s41598-019-40579-6
19. Shin TH, Manavalan B, Lee DY, et al. Silica-coated magnetic-nanoparticle-induced cytotoxicity is reduced in microglia by glutathione and citrate identified using integrated omics. Part Fibre Toxicol. 2021;18(1):42. doi:10.1186/s12989-021-00433-y
20. Saini PK, Kumar N, Keshu S. Health issues and risk assessment of nanomaterials. In: Handbook of Green and Sustainable Nanotechnology. Springer International Publishing; 2023:2553–2579. doi:10.1007/978-3-031-16101-8_30
21. Felici G, Lachowicz JI, Milia S, et al. A pilot study of occupational exposure to ultrafine particles during 3D printing in research laboratories. Front Public Health. 2023:11.doi 10.3389/fpubh.2023.1144475.
22. Mohajerani B, Smith K, Milas A, et al. Nanoparticles in construction materials and other applications, and implications of nanoparticle use. Materials. 2019;12(19):3052. doi:10.3390/ma12193052
23. Ghafari J, Moghadasi N, Shekaftik SO. Oxidative stress induced by occupational exposure to nanomaterials: a systematic review. Ind Health. 2020;58(6):492–502. doi:10.2486/indhealth.2020-0073
24. Lischkova L, Pelclova D, Hlusicka J, et al. Detection and identification of engineered nanoparticles in exhaled breath condensate, blood serum, and urine of occupationally exposed subjects. Monatshefte Für Chemie - Chemical Monthly. 2019;150(3):511–523. doi:10.1007/s00706-019-2379-z
25. Forest V, Pourchez J, Pélissier C, Audignon Durand S, Vergnon JM, Fontana L. Relationship between occupational exposure to airborne nanoparticles, nanoparticle lung burden and lung diseases. Toxics. 2021;9(9):204. doi:10.3390/toxics9090204
26. Jia J, Wang Z, Yue T, Su G, Teng C, Yan B. Crossing biological barriers by engineered nanoparticles. Chem Res Toxicol. 2020;33(5):1055–1060. doi:10.1021/acs.chemrestox.9b00483
27. Cox A, Andreozzi P, Dal Magro R, et al. Evolution of nanoparticle protein corona across the blood–brain barrier. ACS Nano. 2018;12(7):7292–7300. doi:10.1021/acsnano.8b03500
28. González-Vega JG, García-Ramos JC, Chavez-Santoscoy RA, Castillo-Quiñones JE, Arellano-Garcia ME, Toledano-Magaña Y. Lung models to evaluate silver nanoparticles’ toxicity and their impact on human health. Nanomaterials. 2022;12(13):2316. doi:10.3390/nano12132316
29. Rosário F, Creylman J, Verheyen G, et al. Impact of particle size on toxicity, tissue distribution and excretion kinetics of subchronic intratracheal instilled silver nanoparticles in mice. Toxics. 2022;10(5):260. doi:10.3390/toxics10050260
30. Brandão F, Fernández-Bertólez N, Rosário F, et al. Genotoxicity of TiO2 Nanoparticles in Four Different Human Cell Lines (A549, HEPG2, A172 and SH-SY5Y). Nanomaterials. 2020;10(3):412. doi:10.3390/nano10030412
31. Mahmood RI, Kadhim AA, Ibraheem S, et al. Biosynthesis of copper oxide nanoparticles mediated Annona muricata as cytotoxic and apoptosis inducer factor in breast cancer cell lines. Sci Rep. 2022;12(1):16165. doi:10.1038/s41598-022-20360-y
32. Andriamasinoro SN, Dieme D, Marie-Desvergne C, et al. Kinetic time courses of inhaled silver nanoparticles in rats. Arch Toxicol. 2022;96(2):487–498. doi:10.1007/s00204-021-03191-0
33. Pan B, Li S, Peng H, Ao C, Wei Z, Xing B. Advances in understanding the processes and cycling of nanoparticles in the terrestrial environment. Adv Agron. 2023:1–79. doi:10.1016/bs.agron.2023.06.001
34. Waldmann T, Grinberg M, König A, et al. Stem cell transcriptome responses and corresponding biomarkers that indicate the transition from adaptive responses to cytotoxicity. Chem Res Toxicol. 2017;30(4):905–922. doi:10.1021/acs.chemrestox.6b00259
35. Alsagaby SA, Vijayakumar R, Premanathan M, et al. Transcriptomics-based characterization of the toxicity of ZnO nanoparticles against chronic myeloid leukemia cells. Int J Nanomed. 2020;15:7901–7921. doi:10.2147/IJN.S261636
36. Mantione KJ, Kream RM, Kuzelova H, et al. Comparing bioinformatic gene expression profiling methods: microarray and RNA-Seq. Med Sci Monit Basic Res. 2014;20:138–141. doi:10.12659/MSMBR.892101
37. Hufnagel M, Schoch S, Wall J, Strauch BM, Hartwig A. Toxicity and gene expression profiling of copper- and titanium-based nanoparticles using air–liquid interface exposure. Chem Res Toxicol. 2020;33(5):1237–1249. doi:10.1021/acs.chemrestox.9b00489
38. Thiruvengadam R, Venkidasamy B, Samynathan R, Govindasamy R, Thiruvengadam M, Kim JH. Association of nanoparticles and Nrf2 with various oxidative stress-mediated diseases. Chem Biol Interact. 2023;380:110535. doi:10.1016/j.cbi.2023.110535
39. Biola-Clier M, Beal D, Caillat S, et al. Comparison of the DNA damage response in BEAS-2B and A549 cells exposed to titanium dioxide nanoparticles. Mutagenesis. 2017;32(1):161–172. doi:10.1093/mutage/gew055
40. Strauch BM, Hubele W, Hartwig A. Impact of endocytosis and lysosomal acidification on the toxicity of copper oxide nano- and microsized particles: uptake and gene expression related to oxidative stress and the DNA Damage response. Nanomaterials. 2020;10(4):679. doi:10.3390/nano10040679
41. Huang Y, Lü X, Chen R, Chen Y. Comparative study of the effects of gold and silver nanoparticles on the metabolism of human dermal fibroblasts. Regen Biomater. 2020;7(2):221–232. doi:10.1093/rb/rbz051
42. Aragoneses-Cazorla G, Buendia-Nacarino MP, Mena ML, Luque-Garcia JL. A multi-omics approach to evaluate the toxicity mechanisms associated with silver nanoparticles exposure. Nanomaterials. 2022;12(10):1762. doi:10.3390/nano12101762
43. Fuster E, Candela H, Estévez J, Arias AJ, Vilanova E, Sogorb MA. Effects of silver nanoparticles on T98G human glioblastoma cells. Toxicol Appl Pharmacol. 2020;404:115178. doi:10.1016/j.taap.2020.115178
44. Wusu AD, Sibuyi NRS, Moabelo KL, Goboza M, Madiehe A, Meyer M. Citrate-capped gold nanoparticles with a diameter of 14 nm alter the expression of genes associated with stress response, cytoprotection and lipid metabolism in CaCo-2 cells. Nanotechnology. 2022;33(10):105101. doi:10.1088/1361-6528/ac3c7c
45. Ng CT, Dheen ST, Yip WCG, Ong CN, Bay BH, Lanry Yung LY. The induction of epigenetic regulation of PROS1 gene in lung fibroblasts by gold nanoparticles and implications for potential lung injury. Biomaterials. 2011;32(30):7609–7615. doi:10.1016/j.biomaterials.2011.06.038
46. Rossner P, Vrbova K, Rossnerova A, et al. Gene expression and epigenetic changes in mice following inhalation of copper(ii) oxide nanoparticles. Nanomaterials. 2020;10(3):550. doi:10.3390/nano10030550
47. Di Pietro C, HH Ö, Murray TS, Bruscia EM. Targeting the heme oxygenase 1/carbon monoxide pathway to resolve lung hyper-inflammation and restore a regulated immune response in cystic fibrosis. Front Pharmacol. 2020;11. doi:10.3389/fphar.2020.01059
48. Meng J, Liu K, Shao Y, et al. ID1 confers cancer cell chemoresistance through STAT3/ATF6-mediated induction of autophagy. Cell Death Dis. 2020;11(2):137. doi:10.1038/s41419-020-2327-1
49. Tang Y, Zhang S, Li J, Wu C, Fan Q. ID1 marks the tumorigenesis of pancreatic ductal adenocarcinoma in mouse and human. Sci Rep. 2022;12(1):13555. doi:10.1038/s41598-022-17827-3
50. Rubio L, Annangi B, Vila L, Hernández A, Marcos R. Antioxidant and anti-genotoxic properties of cerium oxide nanoparticles in a pulmonary-like cell system. Arch Toxicol. 2016;90(2):269–278. doi:10.1007/s00204-015-1468-y
51. Xiang Y, Ran Q, Wu C, et al. Single-cell transcriptomics uncovers the impacts of titanium dioxide nanoparticles on human bone marrow stromal cells. Chem Eng J. 2022;440:135814. doi:10.1016/j.cej.2022.135814
52. Cao W, Gu M, Wang S, Huang C, Xie Y, Cao Y. Effects of epigallocatechin gallate on the stability, dissolution and toxicology of ZnO nanoparticles. Food Chem. 2022;371:131383. doi:10.1016/j.foodchem.2021.131383
53. Rossnerova A, Honkova K, Pelclova D, et al. DNA methylation profiles in a group of workers occupationally exposed to nanoparticles. Int J Mol Sci. 2020;21(7):2420. doi:10.3390/ijms21072420
54. Holmila R, Wu H, Lee J, Tsang AW, Singh R, Furdui CM. Integrated redox proteomic analysis highlights new mechanisms of sensitivity to silver nanoparticles. Mol Cell Proteomics. 2021;20:100073. doi:10.1016/j.mcpro.2021.100073
55. Biola-Clier M, Gaillard JC, Rabilloud T, Armengaud J, Carriere M. Titanium dioxide nanoparticles alter the cellular phosphoproteome in a549 cells. Nanomaterials. 2020;10(2):185. doi:10.3390/nano10020185
56. Dupree EJ, Jayathirtha M, Yorkey H, Mihasan M, Petre BA, Darie CC. A critical review of bottom-up proteomics: the good, the bad, and the future of this field. Proteomes. 2020;8(3):14. doi:10.3390/proteomes8030014
57. Melby JA, Roberts DS, Larson EJ, et al. novel strategies to address the challenges in top-down proteomics. J Am Soc Mass Spectrom. 2021;32(6):1278–1294. doi:10.1021/jasms.1c00099
58. Gao X, Li R, Yourick JJ, Sprando RL. Transcriptomic and proteomic responses of silver nanoparticles in hepatocyte-like cells derived from human induced pluripotent stem cells. Toxicol in vitro. 2022;79:105274. doi:10.1016/j.tiv.2021.105274
59. Wong TY, Yan N, Kwan KKL, et al. Comparative proteomic analysis reveals the different hepatotoxic mechanisms of human hepatocytes exposed to silver nanoparticles. J Hazard Mater. 2023;445:130599. doi:10.1016/j.jhazmat.2022.130599
60. Fredenburgh LE, Perrella MA, Mitsialis SA. The role of heme oxygenase-1 in pulmonary disease. Am J Respir Cell Mol Biol. 2007;36(2):158–165. doi:10.1165/rcmb.2006-0331TR
61. Edelmann MJ, Shack LA, Naske CD, Walters KB, Nanduri B. SILAC-based quantitative proteomic analysis of human lung cell response to copper oxide nanoparticles. PLoS One. 2014;9(12):e114390. doi:10.1371/journal.pone.0114390
62. Carvajal S, Perramón M, Casals G, et al. Cerium oxide nanoparticles protect against oxidant injury and interfere with oxidative mediated kinase signaling in human-derived hepatocytes. Int J Mol Sci. 2019;20(23):5959. doi:10.3390/ijms20235959
63. Lv M, Huang W, Chen Z, et al. Metabolomics techniques for nanotoxicity investigations. Bioanalysis. 2015;7(12):1527–1544. doi:10.4155/bio.15.83
64. Robertson DG, Watkins PB, Reily MD. Metabolomics in toxicology: preclinical and clinical applications. Toxicol Sci. 2011;120(Supplement 1):S146–S170. doi:10.1093/toxsci/kfq358
65. Essa N, O’Connell F, Prina-Mello A, O’Sullivan J, Marcone S. Gold nanoparticles and obese adipose tissue microenvironment in cancer treatment. Cancer Lett. 2022;525:1–8. doi:10.1016/j.canlet.2021.10.017
66. Kitchin KT, Grulke E, Robinette BL, Castellon BT. Metabolomic effects in HepG2 cells exposed to four TiO 2 and two CeO 2 nanomaterials. Environ Sci. 2014;1(5):466–477. doi:10.1039/C4EN00096J
67. Nirmala JG, Meher K, Lopus M. Proteomic and metabolomic profiling combined with in vitro studies reveal the antiproliferative mechanism of silver nanoparticles in MDA-MB-231 breast carcinoma cells. J Mater Chem B. 2022;10(13):2148–2159. doi:10.1039/D1TB02760C
68. Chavez Soria NG, Aga DS, Atilla-Gokcumen GE. Lipidomics reveals insights on the biological effects of copper oxide nanoparticles in a human colon carcinoma cell line. Mol Omics. 2019;15(1):30–38. doi:10.1039/C8MO00162F
69. Wu B, Wu J, Liu S, et al. Combined effects of graphene oxide and zinc oxide nanoparticle on human A549 cells: bioavailability, toxicity and mechanisms. Environ Sci Nano. 2019;6(2):635–645. doi:10.1039/C8EN00965A
70. Govindasamy C, Al-Numair KS, Prabhakaran VS, Ganesan R. Proton NMR spectroscopy-based metabolomics profiling of doxorubicin-silver nanoparticles for metabolites characterization, and chemical metabolism in breast cancer. Process Biochem. 2023;130:695–704. doi:10.1016/j.procbio.2023.05.020
71. Chen Q, Wang N, Zhu M, et al. TiO2 nanoparticles cause mitochondrial dysfunction, activate inflammatory responses, and attenuate phagocytosis in macrophages: a proteomic and metabolomic insight. Redox Biol. 2018;15:266–276. doi:10.1016/j.redox.2017.12.011
72. Cui L, Wang X, Zhao X, Sun B, Xia T, Hu S. CeO2 nanoparticles induce pulmonary fibrosis via activating S1P pathway as revealed by metabolomics. Nano Today. 2022;45:101559. doi:10.1016/j.nantod.2022.101559
73. Adeyemi JA, Sorgi CA, Machado ART, et al. Phospholipids modifications in human hepatoma cell lines (HepG2) exposed to silver and iron oxide nanoparticles. Arch Toxicol. 2020;94(8):2625–2636. doi:10.1007/s00204-020-02789-0
74. Bannuscher A, Karkossa I, Buhs S, et al. A multi-omics approach reveals mechanisms of nanomaterial toxicity and structure–activity relationships in alveolar macrophages. Nanotoxicology. 2020;14(2):181–195. doi:10.1080/17435390.2019.1684592
75. Scala G, Kinaret P, Marwah V, Sund J, Fortino V, Greco D. Multi-omics analysis of ten carbon nanomaterials effects highlights cell type specific patterns of molecular regulation and adaptation. NanoImpact. 2018;11:99–108. doi:10.1016/j.impact.2018.05.003
76. Zhang J, Li X, Cheng W, et al. Chronic carbon black nanoparticles exposure increases lung cancer risk by affecting the cell cycle via circulatory inflammation. Environ. Pollut. 2022;305:119293. doi:10.1016/j.envpol.2022.119293
77. Pang Z, Zhou G, Ewald J, et al. Using MetaboAnalyst 5.0 for LC–HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat Protoc. 2022;17(8):1735–1761. doi:10.1038/s41596-022-00710-w
78. Chen Z, Han S, Zhang J, et al. Exploring urine biomarkers of early health effects for occupational exposure to titanium dioxide nanoparticles using metabolomics. Nanoscale. 2021;13(7):4122–4132. doi:10.1039/D0NR08792K
79. Araujo ANM, Leroux IN, Furtado DZS, et al. Integration of proteomic and metabolomic analyses: new insights for mapping informal workers exposed to potentially toxic elements. Front Public Health. 2023:10.doi 10.3389/fpubh.2022.899638.
80. Locasale JW. Serine, glycine and one-carbon units: cancer metabolism in full circle. Nat Rev Cancer. 2013;13(8):572–583. doi:10.1038/nrc3557
81. Katzeff JS, Kim WS. ATP-binding cassette transporters and neurodegenerative diseases. Essays Biochem. 2021;65(7):1013–1024. doi:10.1042/EBC20210012
82. Walker DI, Hart JE, Patel CJ, et al. Integrated molecular response of exposure to traffic-related pollutants in the US trucking industry. Environ Int. 2022;158:106957. doi:10.1016/j.envint.2021.106957
83. He X, Meister M, Jeon J, et al. Multi-omics assessment of puff volume-mediated salivary biomarkers of metal exposure and oxidative injury associated with electronic nicotine delivery systems. Environ Health Perspect. 2025;133(1). doi:10.1289/EHP14321
84. Cattelani L, Del Giudice G, Serra A, et al. Quantitative in vitro to in vivo extrapolation for human toxicology and drug development.arXiv preprint arXiv:2401.03277. 2024
85. Mohammadi M, Ahmed Qadir S, Mahmood Faraj A, et al. Navigating the future: microfluidics charting new routes in drug delivery. Int J Pharm. 2024:124142.doi 10.1016/j.ijpharm.2024.124142.
86. Zouraris D, Mavrogiorgis A, Tsoumanis A, et al. CompSafeNano project: nanoInformatics approaches for safe-by-design nanomaterials. Comput Struct Biotechnol J. 2025;29:13–28. doi:10.1016/j.csbj.2024.12.024
87. Ahmad F, Mahmood A, Muhmood T. Machine learning-integrated omics for the risk and safety assessment of nanomaterials. Biomater Sci. 2021;9(5):1598–1608. doi:10.1039/D0BM01672A
88. Nikanjam M, Kato S, Kurzrock R. Liquid biopsy: current technology and clinical applications. J Hematol Oncol. 2022;15(1):131. doi:10.1186/s13045-022-01351-y
89. Coskun A, Ertaylan G, Pusparum M, et al. Advancing personalized medicine: integrating statistical algorithms with omics and nano-omics for enhanced diagnostic accuracy and treatment efficacy. Biochim. biophys. acta, Mol. basis dis. 2024;1870(7):167339. doi:10.1016/j.bbadis.2024.167339
90. V JR, Devasiya T, N VR, et al. Cardiovascular biomarkers in body fluids: progress and prospects in optical sensors. Biophys Rev. 2022;14(4):1023–1050. doi:10.1007/s12551-022-00990-2
91. Pammi M, Aghaeepour N, Neu J. Multiomics, artificial intelligence, and precision medicine in perinatology. Pediatr Res. 2023;93(2):308–315. doi:10.1038/s41390-022-02181-x
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Vitamin B6 Metabolic Pathway is Involved in the Pathogenesis of Liver Diseases via Multi-Omics Analysis
Mei M, Liu D, Tang X, You Y, Peng B, He X, Huang J
Journal of Hepatocellular Carcinoma 2022, 9:729-750
Published Date: 11 August 2022
In vitro Activity of Cefepime/Avibactam Against Carbapenem Resistant Klebsiella pneumoniae and Integrative Metabolomics-Proteomics Approach for Resistance Mechanism: A Single-Center Study
Wen L, Luo C, Chen X, Liu T, Li X, Wang M
Infection and Drug Resistance 2023, 16:6061-6077
Published Date: 11 September 2023
Protective Effect of Modified Suanmei-Tang on Metabolic-Associated Fatty Liver Disease: An Integrated Strategy of Network Pharmacology, Metabolomics, and Transcriptomics

Wang C, Zhao M, Yue Y, Hu C, Zhou C, Zhang Z, He Y, Luo Y, Shen T, Dang S, Yang Y, Zhang Y
Drug Design, Development and Therapy 2024, 18:5161-5182
Published Date: 13 November 2024
The Impact of Immune Cells, Metabolites, Inflammatory Factors, and Circulating Proteins on Atopic Dermatitis: Insights from a Mendelian Randomization Study
Zhou D, Gan G, Song S, Zi C, Bao Y, Hao W, Chen Q
Clinical, Cosmetic and Investigational Dermatology 2024, 17:2999-3011
Published Date: 21 December 2024
Distinct and Shared Molecular Mechanisms in Pediatric Antrochoanal Polyps and Chronic Rhinosinusitis with Nasal Polyps: A Proteomic and Metabolomic Integrative Analysis
Chen YC, Wang X, Pan YW, Teng YS, Pan HG
Journal of Inflammation Research 2025, 18:4435-4447
Published Date: 26 March 2025