Back to Journals » International Journal of Nanomedicine » Volume 21
The Role of Bacterial Outer Membrane Vesicles in Tumor Development: Facts, Potential Applications and Future Perspectives
Authors Gu Z ![]() , Zhang M, Shen F, Chen J, Gu Y, Gong Y, Xue Q, Zhou H
, Zhang M, Shen F, Chen J, Gu Y, Gong Y, Xue Q, Zhou H ![]() , Liu Y, Shao S
, Liu Y, Shao S ![]() , Zhang Y, Bai J
, Zhang Y, Bai J ![]() , Hua K
, Hua K ![]() , Zhou G, Ding J
, Zhou G, Ding J ![]()
Received 18 November 2025
Accepted for publication 16 February 2026
Published 11 March 2026 Volume 2026:21 582600
DOI https://doi.org/10.2147/IJN.S582600
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. RDK Misra
Zheng Gu,1,* Menglei Zhang,1,* Fang Shen,1,* Junhao Chen,1,* Yuanyuan Gu,1 Yingxin Gong,1 Qingxuan Xue,1 Hang Zhou,1 Yueheng Liu,1 Siqi Shao,1 Yue Zhang,1 Jiming Bai,1 Keqin Hua,1 Guannan Zhou,1,2,* Jingxin Ding1,*
1Department of Gynecology, Obstetrics & Gynecology Hospital of Fudan University, Shanghai Key Laboratory of Reproduction and Development, Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Shanghai, People’s Republic of China; 2Department of Medicine Huddinge, Karolinska Institutet, Center for Hematology and Regenerative Medicine, Stockholm, Sweden
*These authors contributed equally to this work
Correspondence: Guannan Zhou; Jingxin Ding, Email [email protected]; [email protected]
Abstract: Bacterial outer membrane vesicles (OMVs), nanosized lipid bilayer particles released by both Gram-negative and Gram-positive bacteria, are emerging as crucial mediators of host–microbe interactions in cancer biology. This review synthesizes current evidence on how OMVs modulate tumor initiation, progression, and therapeutic responses through multifaceted mechanisms aligned with the 14 hallmarks of cancer. Studies demonstrate that OMVs can either promote or inhibit neoplastic processes depending on their bacterial origin and cargo composition. Tumor-promoting OMVs enhance proliferative signaling, drive epithelial–mesenchymal transition, facilitate metastatic dissemination via barrier disruption and angiogenesis, and weaken antitumor immunity. Conversely, other OMVs exert antineoplastic effects by triggering intrinsic apoptosis, cell-cycle arrest, immunogenic cell death, and remodeling of the tumor immune microenvironment. Pre-clinical studies further highlight the translational potential of engineered OMVs as precision nano-vaccines, immunotherapeutic agents, and adjuvants that synergize with immune checkpoint blockade, chemotherapy, or photothermal therapy while minimizing systemic toxicity. Future directions could focus on mapping of OMV cargo–pathway–phenotype networks by multidisciplinary methods, programmable vesicle design using synthetic biology, and real-time microbiome-OMV monitoring in early-phase clinical trials to enable individualized onco-therapeutics. Collectively, OMVs represent a versatile platform to bidirectionally regulate oncogenesis and therapeutic responses. Exploiting their molecular plasticity through rational engineering and precision medicine frameworks would bring transformative potential for cancer prevention, diagnosis, and treatment.
Keywords: bacterial outer membrane vesicles, tumor microenvironment, carcinogenesis, metastasis, immunotherapy, nanomedicine
Introduction
Bacteria release membrane vesicles (MVs) of 20–400nm in diameter, with specific cargos and various functions.1 MVs were initially found to be produced by blebbing from the outer membrane of Gram-negative bacteria and are therefore termed as outer membrane vesicles (OMVs). However, more recent studies have indicated that Gram-positive species and bacteria that do not Gram stain can also release OMVs.2,3 OMVs consist of a single membrane bilayer, and encapsulate periplasmatic and cytosolic proteins, DNA and RNA, and virulence factors. Therefore, OMVs are considered as a specialized bacterial secretion pathway.4,5 Owning to their abundant cargos and membrane components, OMVs can affect diverse biological processes, including DNA transfer, cell-to-cell communication, the delivery of virulence factors and toxins to host cells, inflammatory and immunomodulatory effects, and so on.1,5,6
In addition to canonical OMVs, the MVs repertoire is now recognized to be markedly heterogeneous. Outer-inner membrane vesicles (OIMVs) have two membrane bilayers derived from both outer and inner membranes.7 DNA is specifically packed into these types of MVs.7,8 Biogenesis is thought to proceed via explosive cell lysis, a process that also generates explosive outer-membrane vesicles (EOMVs) that randomly contain cytoplasmic components.9 Cytoplasmic membrane vesicles (CMVs) are MVs released by Gram-positive bacteria, derived from “bubbling cell death” triggered by endolysin or other peptidoglycan-damaging enzymes or treatments.10 Tube-shaped membranous structures (TSMSs) are tube-like protrusions of the cytoplasmic membrane of Gram-positive bacteria or of the outer membrane of Gram-negative bacteria, acting as intercellular bridges to trade cytoplasmic molecules between biofilm cells.11 For more detailed classification and routes of formation of MVs, please refer to Toyofuku et al.12 For brevity, all bacteria-derived membrane vesicles discussed below are indiscriminately referred to as OMVs (Figure 1).
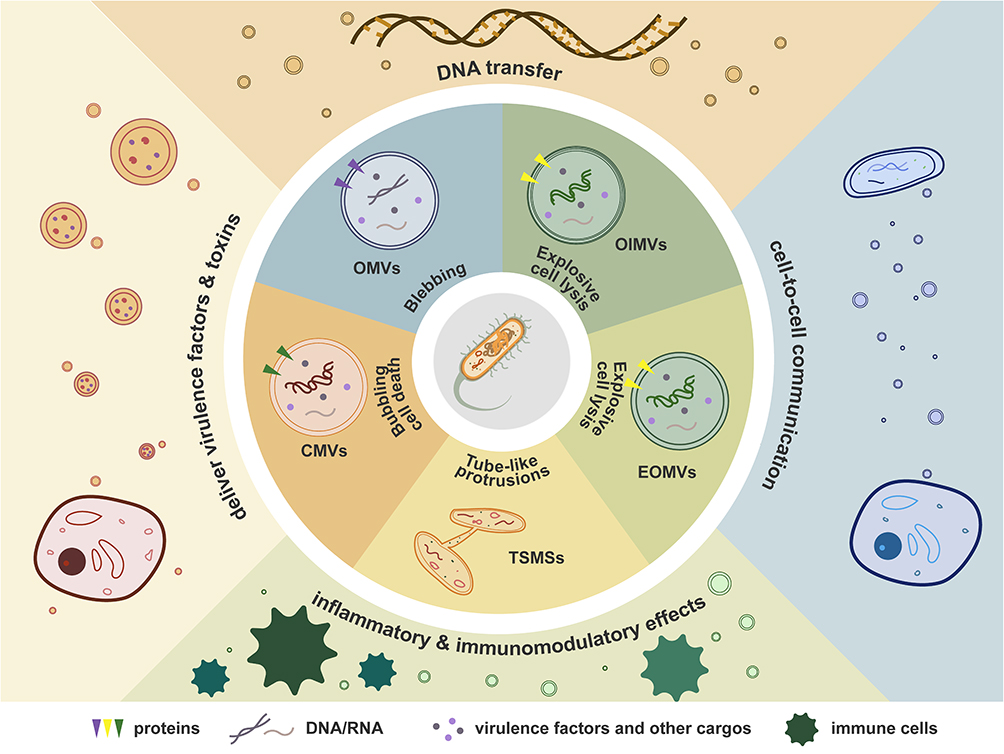 |
Figure 1 The biogenesis and functions of OMVs. Purple, green, and yellow arrowheads: trans-member proteins. Curve lines: DNA/RNA. Dots: virulence factors and other cargos. Abbreviations: OMVs, Outer membrane vesicles; OIMVs, Outer-inner membrane vesicles; EOMVs, Explosive outer-membrane vesicles, CMVs, Cytoplasmic membrane vesicles); TSMSs, Tube-shaped membranous structures. Notes: Triangles represent proteins; curved lines indicate DNA/RNA; dots denote virulence factors and other cargos. |
As a subpopulation of extracellular vesicle (EVs), OMVs share the characteristics and functions of EVs. The term “extracellular vesicles” was first suggested in 2011 as mobile membrane-limited vesicles in extracellular environment, including exosomes, activation- or apoptosis-induced microparticles and apoptotic bodies.13 EVs can be released by all cellular organisms, representing that vesiculation is a fundamental process of living matter.14
In recent years, the microbiome has emerged as a crucial role in carcinogenesis and prognosis, and can synergize cancer diagnosis and treatment.15 Few microbes are direct carcinogens, but many of them could affect tumor development, including biogenesis, growth, invasion and metastasis16–19 Dysbiosis or imbalance in the microbiome has also been mechanistically linked to local and systemic disorders, influencing different organs, immune dysregulation and oncogenesis.20 As the dominant secretory conduit of bacteria, OMVs are considered crucial in interaction between pathogens and host cells, both nearby and distant.21,22 Compared with eukaryotic extracellular vesicles, OMVs exhibit smaller size, higher intrinsic immunogenicity, and superior amenability to genetic engineering, while mass-production can be achieved cost-effectively through high-density fermentation.1,12,23 Besides, OMVs could target immune cells more and accumulate within inflammatory lesions, rendering them particularly suited for cargo delivery and immunomodulation.1,12 Due to their specific biostructure and activities, naturally secreted or bioengineered OMVs have been propelled to the forefront of next-generation vaccinology and targeted drug-delivery platforms.24 However, the mechanisms through which OMVs affect various biological processes in cancer development have been largely neglected. This review aims to synthesize current evidence delineating the multifaceted roles of OMVs in the initiation, progression and metastatic outgrowth of diverse malignancies, and critically appraise their translational potential for cancer prevention, early detection and precision therapeutics.
Human cells make their way from normalcy to malignant tumors by acquiring various functional capabilities. The most widely endorsed classification is the hallmarks of cancer put forward by Hanahan in 2000, 2011, and 2022, including 14 distinct characteristics, which are crucial for their ability to form malignant tumors. The 14 hallmarks include core hallmark capabilities that define the neoplastic phenotype at the single-cell level, and enabling characteristics that supply the cellular and microenvironmental substrate for acquisition of these functional traits.25–27 OMVs involve the development of cancer bidirectionally by interfering with the acquisition of these capabilities and the effect of these functions through various mechanisms. In this review, we categorize the above mechanisms based on the 14 hallmarks, aiming at providing innovative perspectives for future research, pharmaceutical development, and therapeutic strategies.
The Impacts of OMVs on Tumor Development
The association between OMVs and human pathobiology has become an area of intensive investigation in recent years. Many OMVs are demonstrated significant in both localized and systemic diseases.28 For example, Porphyromonas gingivalis OMVs has been found to be closely related to the development of cardiovascular disease, Alzheimer’s disease, rheumatoid arthritis, diabetes mellitus, non-alcoholic hepatitis, and cancer,29 among which cancer might be the one that gets the most attention. This part will therefore focus on the relationship between OMVs and cancer development.
Human oncogenesis is a multistep process driven by both well characterized and emerging influencing factors. The main capabilities necessary to tumor cells have been categorized by Hanahan into 14 hallmarks as is mentioned above. Based on Hanahan’s theory, mechanisms through which OMVs promote or inhibit tumor development are summarized as regulating tumor cell proliferation, affecting metabolic reprogramming, mediating invasion and metastasis, and modulating immune microenvironment. All these data establish OMVs as a crucial regulator in connecting microbiome and tumor biology (Figure 2).
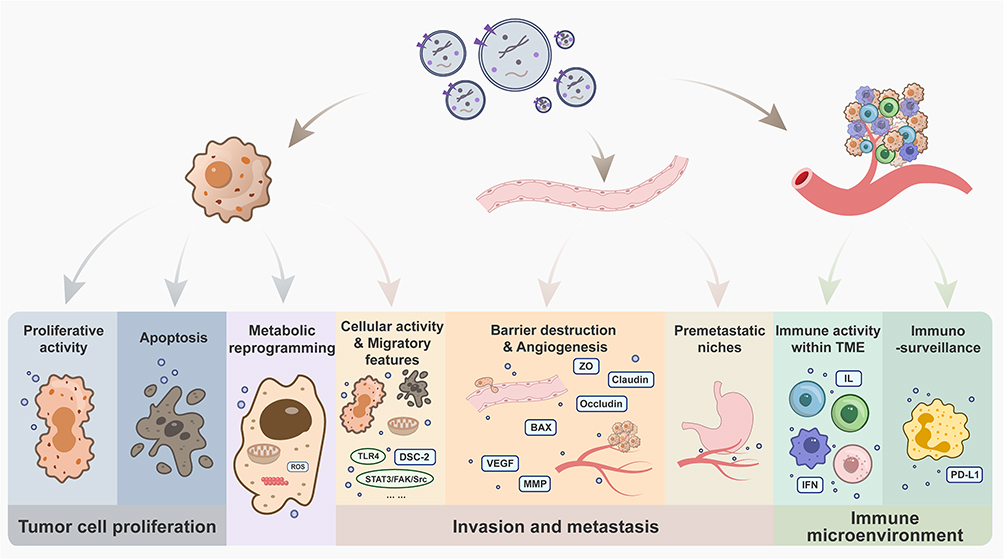 |
Figure 2 The impact of OMVs on tumor development. |
OMVs and Tumor Cell Proliferation
OMVs can regulate tumor cell proliferation by affecting their proliferative potential, apoptosis process, and metabolic progresses (Table 1).
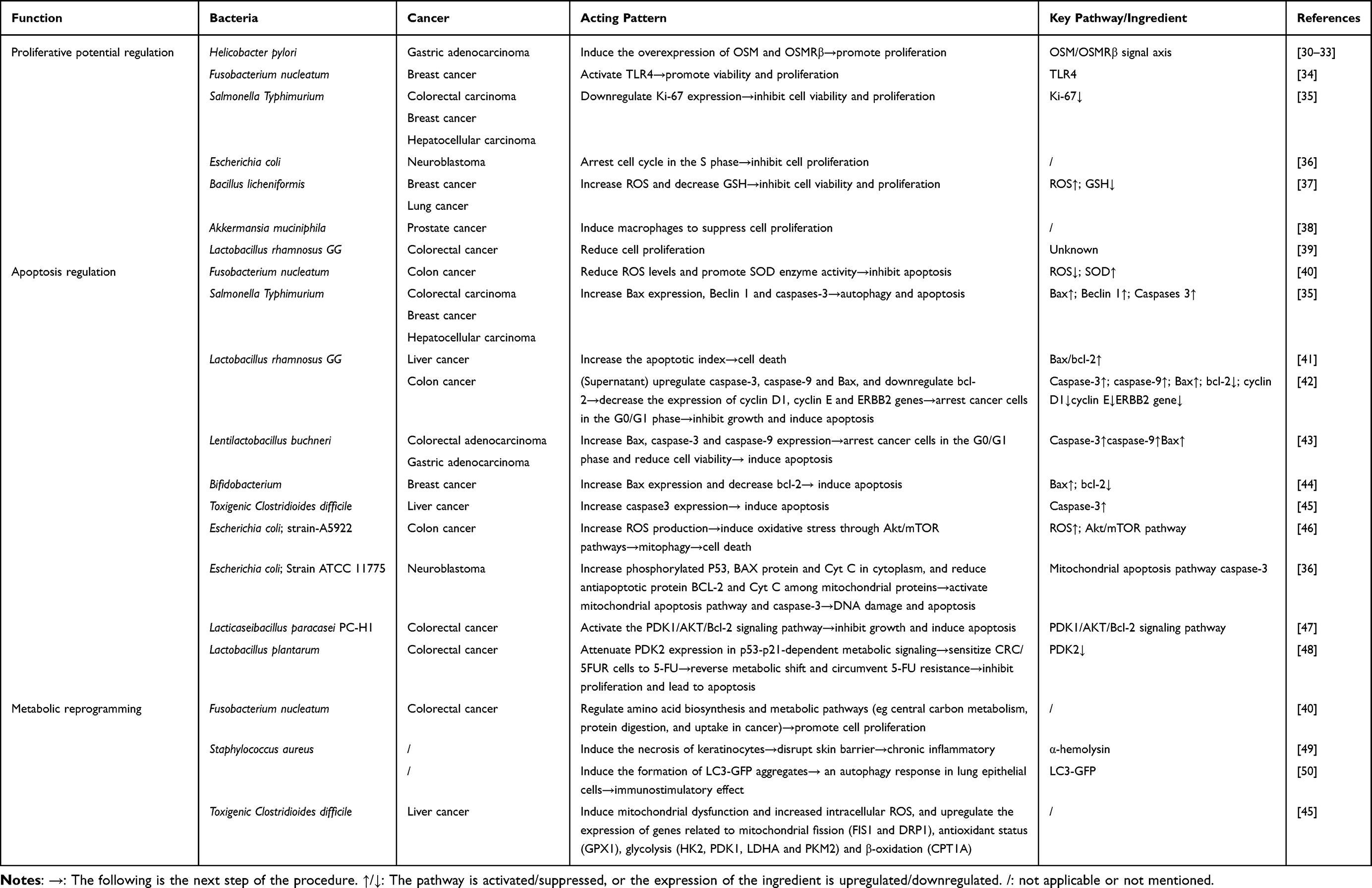 |
Table 1 The Role of OMVs in the Regulation of Tumor Proliferation |
OMVs Mediated Proliferative Ability
OMVs modulate neoplastic proliferation capacity at multiple molecular levels. Helicobacter pylori OMVs (H.p OMVs), a well-characterized cofactor in digestive system oncogenesis, has been reported to regulate the oncogenes or tumor suppressors genes in host cells in many studies. H.p OMVs induce the secretion of Oncostatin-M (OSM) and overexpression of its type II receptor OSMRβ in human gastric adenocarcinoma cell line (AGS), while similar overexpression is detectable in many other cancer types, such as pancreatic cancer, breast cancer and colon cancer.30,31 Elevated OSM is found in precancerous lesions and early gastric cancer biopsies, and OSMRβ is demonstrated to promote gastric cancer growth and metastasis upon treatment with OSM.32,33 The OSM/OSMRβ signal axis involves in many physical and pathological processes, including enhancement of cancer cell proliferation, metastasis, regulation of inflammation. Collectively, the H.p OMVs- OSM/OSMRβ axis functions as a context-dependent factor that converts bacterial signals into the full cascade of human tumor initiation and progression across anatomical sites.30–33 In breast cancer, OMVs derived from Fusobacterium nucleatum are also demonstrated to promote viability and proliferation through the activation of TLR4 pathway in vitro and in vivo.34
Conversely, some studies indicate that certain OMVs inhibit tumor cell proliferation. Salmonella Typhimurium OMVs inhibit tumor growth and cell proliferation by downregulating Ki-67 expression.35 Escherichia coli (E. coli) OMVs taken into cytoplasm and nucleus directly arrest cell cycle, blocking cells in the S phase.36 Bacillus licheniformis OMVs increase reactive oxygen species (ROS) and decrease glutathione (GSH) to inhibit cell viability and proliferation.37 In prostate cancer, Akkermansia muciniphila OMVs treated macrophages suppress the proliferation and invasion of tumor cells.38 Lactobacillus rhamnosus GG OMVs are able to reduce the cell proliferation of HT29 (colorectal adenocarcinoma cell line) and SW480 (colorectal cancer cell line) by mechanism that still needs exploration.39
OMVs Mediated Apoptosis
Apoptosis is a form of programmed cell death, essential for tissue homeostasis. It involves the activation, expression and regulation of a series of genes, such as the bcl-2 family and caspase family. By affecting the progress of apoptosis, OMVs can improve or reduce cell activity. OMVs from Fusobacterium nucleatum reduce ROS levels and oxidative stress response in colon cancer cells (CCC) to inhibit apoptosis.40 Meanwhile, these vesicles promote the activity of superoxide dismutase (SOD) enzyme, which enhance the ability to fight free radicals and protect from oxidative damage, thus enhancing the activity of CCC.40
Some OMVs induce cellular apoptosis, playing anti-tumor effects in vitro and in vivo. Salmonella Typhimurium OMVs elevate the expression of pro-apoptotic Bax gene, Beclin 1 (a marker for autophagy), and caspase-3 (a marker for cellular apoptosis), mediating non-inflammatory self-destruction of cancer cells.35 OMVs released by Lactobacillus rhamnosus GG increase the apoptotic index (bax/bcl-2 expression ratio) in liver cancer, and lead to cancer cell death.41 Meanwhile, the supernatant of Lactobacillus rhamnosus GG is confirmed to upregulate pro-apoptotic genes including caspase-3, caspase-9 and Bax, and downregulate bcl-2, resulting in a decrease in expression levels of cyclin D1, cyclin E and ERBB2 genes, and then arrest cancer cells in the G0/G1 phase of the cell cycle, thus inhibiting the growth of HT-29 cancer cells and induce apoptosis.42 Lentilactobacillus buchneri OMVs are also demonstrated apoptotic activity by increasing the expression of Bax, caspase-3 and caspase-9 genes, arresting AGS and HT‑29 cells in the G0/G1 phase and reducing cell viability.43 Likewise, the Bifidobacterium OMVs increase Bax expression and decrease bcl-2, inducing cellular apoptosis in triple-negative breast cancer,44 and Toxigenic Clostridioides difficile OMVs upregulate caspase3 gene expression to increase apoptosis in liver cancer cells.45 OMVs from E. coli strain-A5922 increase ROS production and induce oxidative stress via the activation of Akt/mTOR pathways, subsequently inducing mitophagy and culminating in cell death in CCC.46 OMVs from another E. coli strain, ATCC 11775, upregulate the expression of phosphorylated P53, BAX protein and Cyt C in cytoplasm, and downregulate the expression of antiapoptotic protein BCL-2 and Cyt C among mitochondrial proteins. Thus, the mitochondrial apoptosis pathway and cleaved caspase-3 are activated, resulting in DNA damage and apoptosis in neuroblastoma.36 Lacticaseibacillus paracasei PC-H1 OMVs activate the PDK1/AKT/Bcl-2 signaling pathway to inhibit the growth of colorectal cancer (CRC) cells and induce apoptosis.47 OMVs derived from Lactobacillus plantarum sensitize CRC/5FUR cells (5-fluorouracil-resistant CRC cells) to 5-FU by attenuating PDK2 expression in p53-p21-dependent metabolic signaling, to reverse the metabolic shift (such as the increased glycolysis) and circumvent 5-FU resistance, inhibiting cancer cell proliferation and leading to apoptosis.48
OMVs Mediated Metabolic Reprogramming
Effects on metabolic progresses can lead to diverse outcomes of tumor cell proliferation. OMVs from Fusobacterium nucleatum regulate amino acid biosynthesis and metabolic pathways in CRC cells, such as central carbon metabolism, protein digestion, and uptake in cancer, thus promoting cell proliferation.40 OMV-associated α-hemolysin of Staphylococcus aureus (S. aureus) induce necrosis of keratinocytes, disrupting skin barrier and resulting in chronic inflammatory.49 S. aureus OMVs can also induce the formation of LC3-GFP aggregates and an autophagy response in lung epithelial cells, leading to the immunostimulatory effect in host cells.50 Toxigenic Clostridioides difficile OMVs are demonstrated to induce mitochondrial dysfunction and increased intracellular ROS, and meanwhile upregulate the expression of genes related to mitochondrial fission (FIS1 and DRP1), antioxidant status (GPX1), glycolysis (HK2, PDK1, LDHA and PKM2) and β-oxidation (CPT1A) in liver cancer cells, with impacts on proliferation under exploration.45
OMVs Mediated Invasion and Metastasis
Invasive and metastatic competence is considered one of the hallmarks of cancer, denoting the biologic capacity of malignant cells to detach from the primary neoplasm to a new organ and establish secondary tumors. The complex process of invasion and metastasis has been summarized in a series of steps, often termed as invasion-metastasis cascade. The complete course includes the local invasion, detachment of tumor cells, intravasation into nearby blood and lymphatic vessels, transit through the lymphatic and hematogenous systems, attachment to endothelial cells that line such vessels, extravasation from the lumina to the parenchyma of tissue or organs, the formation of small nodules, and finally the growth from micro-metastatic lesions into tumors, which is termed as colonization.26,51 Accumulating evidence indicates that OMVs modulate the cascade in diverse aspects (Table 2).
 |
Table 2 The Role of OMVs in the Mediation of Tumor Invasion and Metastasis |
Effects on Tumor Cells
The effects of OMVs on tumor cells mainly include altered cellular activity and acquisition of migratory features. Among the OMVs mentioned above, some have been demonstrated to attenuate invasive and migrative capacity by inhibiting tumor cell proliferation and thus affecting cellular activity. Salmonella Typhimurium OMVs downregulate Ki-67 expression and increase Bax expression, Beclin 1 and caspases-3, inhibiting tumor cell proliferation. The inhibition also impact the invasive and migratory viability of tumor cells, playing significant anti-tumor effects.35 Similarly, Lentilactobacillus buchneri OMVs decrease migration viability by upregulating apoptotic activity.43 In addition, macrophages treated by Akkermansia muciniphila OMVs suppress the proliferation and invasion of prostate cancer cells, of which concrete procedures will be summarized in the following parts.38
OMVs exert bidirectional control over the acquisition of metastatic competence, either licensing or constraining the molecular and cellular programs that drive tumor cell dissemination. Fusobacterium nucleatum OMVs enhance the migration and invasion of breast cancer cells via TLR4,34 and have similar effects on colorectal cancer cells by increasing mitochondrial fusion.40,52 It can also promote cell migration and invasion of oral cancer by activating intracellular autophagy pathways and inducing the EMT (epithelial–mesenchymal transition) phenotype in cancer cells, downregulating E-cadherin and upregulating Vimentin and N-cadherin.53 Helicobacter pylori OMVs induce OSM secretion and OSMRβ overexpression, activate the STAT3/FAK/Src pathway, and finally prompt the migration and invasion potential of AGS.30–33 Porphyromonas gingivalis OMVs contain abundant sRNA23392, which is found to target and degrade Desmocollin-2 (DSC-2), a type of desmosomes, acting as a cadherin-type transmembrane adhesion molecule. With the decreased expression of DSC-2, the adhesion ability of oral squamous cell carcinoma cells is reduced, leading to the enhancement of migration and invasion ability.54
Effects on Vascular Systems
Hematogenous and lymphatic dissemination share a mechanistic prerequisite: the destruction of endothelial junctions that permits tumor cells to traverse the vessel wall in a process collectively termed trans-endothelial migration (TEM).64 Meanwhile, individualities exist in both metastases. For example, cytokines such as vascular endothelial growth factor C/D (VEGFC/D) and insulin-like growth factor 1 (IGF-1) secreted by cancer cells can induce lymphatic vessel formation and drive lymphatic metastasis,65–67 while neutrophils, monocytes-derived VEGF and metastasis-associated inflammatory macrophages can contribute to extravasation in hematogenous metastasis.60,64,68 As most studies explore the two systems with a same model rather than focus on their distinctions, we frame the impact of OMVs on vascular systems according to their underlying biologic processes—barrier destruction and angiogenesis—rather than by the anatomic distinction between blood and lymphatic vessels.
The integrity of endothelial barrier is guaranteed by tight junction proteins, which mainly consist of the claudin family, occludin family, and Zona Occludens (ZO) family. Fusobacterium nucleatum OMVs downregulate the expression of miR-574-5p to activate mir-574-5p/CARD3-dependent autophagy, reducing ZO-1 and claudin-1 and thus leading to epithelial barrier dysfunction in intestinal epithelial cells (IECs).55 The enteropathogenic E. coli (EPEC) OMVs downregulate ZO-1, ZO-2, occludin, claudin-14 expression and alter the subcellular localization of ZO-1, occludin and F-actin cytoskeleton. However, E.coli Nissle 1917 (EcN) are found to positively modulate the intestinal epithelial barrier by upregulating ZO-1, claudin-14 and downregulating claudin-2, which can also counteract EPEC-altered transepithelial resistance and paracellular permeability, by retaining ZO-1 and occludin at tight junctions in the cell boundaries and ameliorating F-actin disorganization by factors free-released or associated with OMVs.56,57 Likewise, Limosilactobacillus reuteri OMVs protect the integrity of epithelial barrier from the detrimental effect of enterotoxigenic E. coli (ETEC).58 Apart from the above, Limosilactobacillus johnsoni and Limosilactobacillus mucosae OMVs improve intestinal barrier function by increasing ZO-1 and occludin mRNA expression and reducing IL-1β, caspase-3, and bax gene expression in intestinal epithelial cells.59 Akkermansia muciniphila OMVs enter intestinal epithelial cells to stimulate the expression of ZO-1 and occludin, enhancing tight junction function and decreasing the gut permeability, thus improving the intestinal barrier integrity55,60 (Figure 3).
 |
Figure 3 The influence of OMVs on tight junction of endothelial barrier. Upward arrows: upregulate the expression of the molecules. Downward arrows: upregulate the expression of the molecules. |
Some OMVs downregulate angiogenesis to inhibit migration and invasion. Salmonella Typhimurium derived OMVs play the role of anti-angiogenesis by inhibiting the expression of VEGF gene.35 OMVs of E. coli (strain ATCC 11775) decrease the expression of MMP-2, MMP-9, VEGF, CD24 to inhibit migration and invasion of neuroblastoma cells.36
OMVs Mediated Formation of Premetastatic Niches in Distant Organs
Some OMVs counteract with distant organs and thus affect cancer development. CagA is a bacterial oncoprotein,61 and its delivery to distant organs and tissues mediated by Helicobacter pylori OMVs may be involved in the development of extragastric neoplasia.62 OMV-CagA is found to localize in proximity to ZO-1 in junctional area, and induce strong histone H1 affinity to ATP in differentiated and polarized epithelial monolayers, which may have possible effects on gene transcription, leading to different results of cancer development.63
Modulation of the Immune Microenvironment
The effects of OMVs on immune microenvironment are listed in Table 3.
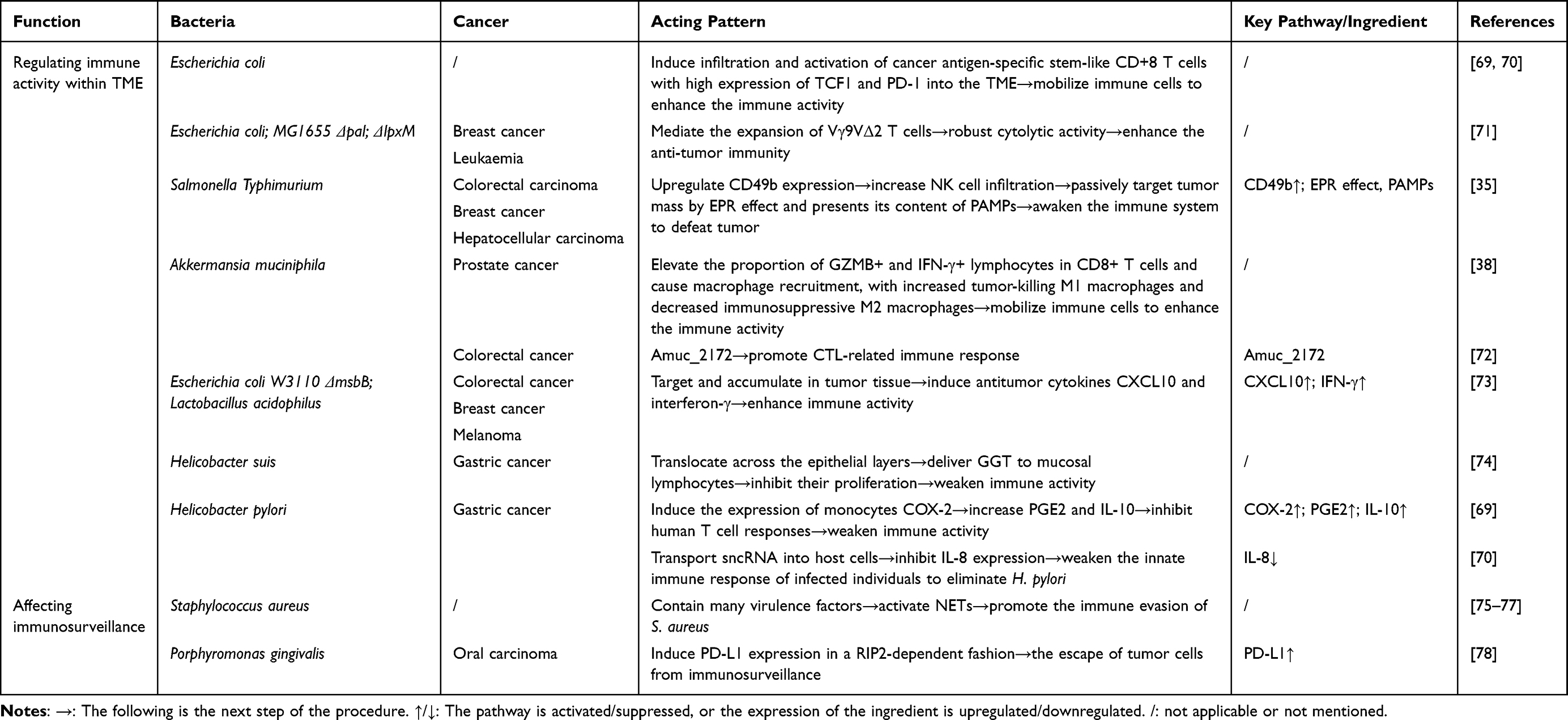 |
Table 3 The Role of OMVs in the Modulation of Immune Microenvironment |
OMVs Mediated Regulating Immune Activity within the Tumor Microenvironment (TME)
Tumor microenvironment (TME) is a highly integrated and dynamically evolving ecosystem composed of neoplastic cells, heterogeneous immune infiltrates and extracellular-matrix constituents. This multicellular niche governs every stage of tumorigenesis, from initial transformation to metastatic outgrowth.79 OMVs either amplify or attenuate the immune activity in TME, engaged in the modulation of immune microenvironment, thereby exerting control over cancer progression.
Some OMVs enhance the immune activity by mobilizing immune cells. E. coli OMVs are demonstrated to induce infiltration and activation of cancer antigen-specific stem-like CD+8 T cells with high expression of TCF1 and PD-1 into the TME.69,70 E. coli MG1655 Δpal ΔlpxM OMVs show anti-tumor immunity by mediating the expansion of Vγ9VΔ2 T cells, which then develop robust cytolytic activity against breast cancer and leukaemia cells.71 Salmonella Typhimurium derived OMVs are successfully exploited to passively target tumor mass by EPR (enhanced permeability and retention) effect and presents its content of PAMPs (pathogen-associated molecular patterns), and thus awaken patients’ immune system to defeat tumor. The anti-tumor effect is mediated by upregulating CD49b expression and then increasing NK cells infiltration.35 Akkermansia muciniphila OMVs play antitumor immunity against prostate cancer by elevating the proportion of granzyme B-positive (GZMB+) and interferon γ-positive (IFN-γ+) lymphocytes in CD8+ T cells and causing macrophage recruitment, with increased tumor-killing M1 macrophages and decreased immunosuppressive M2 macrophages.38 Also, Akkermansia muciniphila OMVs are found to promote CTL-related immune response against CRC with Amuc_2172 as active principle.72 In the meanwhile, some OMVs trigger anti-tumor responses by specifically targeting and accumulating in tumor tissue, thus inducing antitumor cytokines CXCL10 and interferon-γ.73
By avoiding immune destruction, some OMVs and their components weaken the immune activity within TME. OMVs from Helicobacter suis translocate across the epithelial layers and deliver γ-glutamyl transpeptidase (GGT) to mucosal lymphocytes, inhibiting their proliferation.74 OMVs from Helicobacter pylori induce the expression of monocytes COX-2 to increase PGE2 and IL-10, inhibiting T cell responses.80 Meanwhile, Helicobacter pylori OMVs are found to transport sncRNA into host cells to inhibit IL-8 expression, weakening the innate immune response of infected individuals to eliminate Helicobacter pylori, which may worsen chronic gastritis and gastric cancer.81
Affecting Immunosurveillance
The theory of immune surveillance proposes that cells and tissues are under the monitor of immune system, which is responsible for recognizing and eliminating the vast majority of incipient cancer cells and thus nascent tumors.26 Certain OMVs drive neoplastic progression by subverting immunosurveillance. OMVs derived from S. aureus are enriched in virulence factors which trigger a specialized mode of neutrophil death named as NETs (Neutrophil Extracellular Traps), to foster bacterial immune evasion of S. aureus from neutrophils, and establish a chronically inflamed microenvironment, thereby creating a pro-carcinogenic niche that facilitates tumor progression.75–77 Porphyromonas gingivalis OMVs induce programmed death ligand 1 (PD-L1) expression on human oral carcinoma cells in a receptor-interacting serine/threonine-protein kinase 2 (RIP2)-dependent fashion, which can then induce the dysfunction and apoptosis of T cells through the interaction between PD-L1 and programmed death 1 (PD1) on the surface of T cells, thus elevating the escape of tumor cells from immunosurveillance.78,82
Clinical Application and Potential
Owing to their distinctive structural characteristics, OMVs exhibit substantial potential for clinical translation. Natural or bioengineered OMVs serve as effective carriers to transport bacteria-, virus- and human-derived proteins, functioning as mucosal biologics, vaccine and drug delivery platform. In this section, the application of OMVs will be reviewed under three main categories: vaccines, immunotherapeutic agents and adjunctive therapy, and some other therapeutic options will also be briefly mentioned (Table 4 and Figure 4).
 |
Table 4 OMVs with Clinical Application Potential |
 |
Figure 4 The clinical application and potential of OMVs. Purple, green, and yellow arrowheads: trans-member proteins. Curve lines: DNA/RNA. Dots: virulence factors and other cargos. |
Meanwhile, it is necessary to ensure the quality and purity of the extracted OMVs before application to avoid risk bacterial infection and intoxication. While eukaryotic extracellular vesicles isolation is standardized in Minimal Information for Studies of Extracellular Vesicles (MISEV) updated in 2023 by the International Society for Extracellular Vesicles (ISEV),104 OMVs purification lacks consensus and the extraction methods should be carefully chosen according to specific bacterial species and culture conditions. Classical workflows include ultracentrifugation, ultrafiltration and characterization with nanoparticle tracking analysis, and newer techniques for isolation and characterization include acoustic trapping and resistive pulse sensing, respectively.105 Selecting appropriate centrifugation and filtration strategies will help researchers with purification, and in order to confirm the absence of viable bacteria, parallel validation by microbial planting is necessary.23
OMVs-Based Vaccines
Many studies have explored the clinical potential of OMVs, one of which OMVs-based vaccines constitute a major application. The first generation of FDA-approved OMVs-based vaccine is meningococcal group B vaccine in children, which is under the trade name Bexsero®, preventing infectious disease.83,84 Numerous additional OMVs are still being studied about the mechanisms by which they can function as a part of vaccines. Helicobacter pylori OMVs (ExHp-CD) are found to induce TH2-biased immune responses to resist H. pylori infection and induce huge production of IL-10 and IL-4. Thus, ExHp-CD can be considered as antigen-presenting vesicles and acellular vaccine. ExHp-CD and its associated cargos can also serve as effective immunogenic candidates.85,86 OMVs of some types of S. aureus can protect mice against S. aureus infection.75,87,88 Engineered S. aureus OMVs with non-toxic HlaH35L and the LukE monomer packaged are demonstrated to protect immunized mice against lethal sepsis.89 Some scientists engineer the Gram-negative bacterium Bacteroides thetaiotaomicron (Bt) to incorporate bacteria-, virus- and human-derived proteins into its OMVs to deliver the proteins to the respiratory and gastrointestinal tract. Bt OMVs with Typhimurium-derived vaccine antigens and influenza A virus (IAV)-derived vaccine antigens in or on the outer membrane can elicit antigen-specific immune and antibody responses in mucosal tissues and systemically, thus protecting against infection. Furthermore, Bt OMVs can stably express the human therapeutic protein, keratinocyte growth factor-2 (KGF-2), which reduce disease severity and promote intestinal epithelial repair and recovery when delivered orally in colitis models.90 Trained immunity-related vaccines (TIrV) is an emerging therapeutic tumor vaccine form, which can potentially foster antitumor activity by training innate immune cells and their derived hematopoietic progenitor cells without specific antigens. OMV nanohybrids (OMV-SIRPα@CaP/GM-CSF), an OMV-based TIrV, have been developed to target tumor-associated macrophages to enhance TIrV-generated antitumor activity, which shows possibilities of broad-spectrum vaccines and development of the next-generations of tumor vaccines.92
Immunotherapeutic Agents
OMVs can also serve as potent immunotherapeutic agents that elicit durable antitumor immunity for the treatment of various cancers. Kim et al have found gram-negative bacterial OMVs deprived of bacterial endotoxin lipopolysaccharide (LPS) can fully eradicate established tumors without notable adverse effects and induce long-term antitumor immune responses. When administered systematically, these OMVs specifically target and accumulate in tumor tissue and induce the production of antitumor cytokines CXCL10 and IFN-γ, leading to this IFN-γ-dependent antitumor effect.73 Park’s team have produced a kind of artificial OMV named as Synthetic Bacterial Vesicles (SyBV), which avoid activating the innate immune system and thus do not induce a severe toxic response. SyBV is found effective in activation of mouse bone marrow-derived dendritic cells. And co-immunization with SyBV and melanoma extracellular vesicles can elicit tumor regression in melanoma-bearing mice through Th-1 type T cell immunity and balanced antibody production.91 Feng et al have developed an OMV-based, controllable two-way adaptor, PEG/Se@OMV-CD47nb through genetic engineering and surface modification. OMV-CD47nb can simultaneously bind to both TAMs (tumor-associated macrophages) and tumor cells to induce TAM sensitization and CD47 blockade that synergistically promote robust phagocytosis of tumor cells by TAMs, and this multiple mechanism-based activation of macrophage phagocytosis will result in TME remodeling and stimulation of T cell-mediated immunity meanwhile. In addition, the surface PEG/Se layer modification endows the formulation with radiation-triggered controlled release of OMV-CD47nb, mitigating the side effects and enabling the precise release of OMV-CD47nb at the tumor site, thereby increasing the safety window of intravenous injection of the formulation.93 Su et al have designed a delivery platform developed by fusing a genetically engineered transferrin receptor-mediated lysosome-targeting chimera (TfR-LYTAC) to the surface of OMVs, which can be efficiently internalized via TfR-mediate endocytosis and targets PD-L1 for lysosomal degradation in cultured cells. The engineered OMV-LYTAC combines PD-1/PD-L1 pathway inhibition by LYTAC with immune activation by OMVs, significantly reducing tumor growth in vivo.94 Sun conjugated two types of nanobodies targeting CD47/SIRPα and PD-1/PD-L1 pathways onto the surfaces of the OMVs simultaneously and constructed an OMV-based nanosized immune cell engager (OMV-NICE) with dual-targeting abilities. OMV-NICE enhance the recruitment of macrophages and T cells to the tumor region and show superior antitumor activity compared to those of unconjugated nanobodies or OMVs alone.95 OMV/SaFeFA, derived from E. coli, is anchored with ferrous ions vis electrostatic interactions and loaded with the STING agonist-4, followed by tumor-targeting DSPE-PEG-FA decoration, aiming to activate ferroptosis and stimulate the immune responses. OMV/SaFeFA shows tumor-targeting capabilities, eliciting robust antitumor responses while minimizing side effects.96 Chen et al have developed melanin-loaded OMVs from EcN by genetic engineering and then coated them with calcium phosphate (CaP) to reduce systemic toxicity in tumor therapy, and named the nanoparticles as OMVMel@CaP. Under the co-stimulation of OMVMel as immunoadjuvants and the damage-associated molecular patterns (DAMPs) released by the photothermal effect, the efficiency of tumor photothermal/immunotherapy can be largely boosted through triggering antitumor immune responses.97
Adjunctive Therapy
Beyond their intrinsic activity, certain OMVs function as adjuvants that augment established therapies and enhance therapeutic efficacy. Acting as monotherapy or an adjuvant to the standard chemotherapy, Salmonella Typhimurium OMVs both have antitumor effects and can help avoid chemotherapy-related complications or side effects and reduce the rate of drug resistance.35 Bacillus licheniformis OMVs can be combined with doxorubicin to enhance mitochondrial dysfunction, DNA damage, and apoptosis when treating breast and lung cancer.37 E. coli OMVs have synergistic effects with anti-PD-1 combination therapy, inhibiting tumor growth substantially.69,70 The immunotherapeutic effect of the SyBV mentioned above can be synergistically enhanced by anti-PD-1 inhibitor, and SyBV shows significantly greater adjuvant activity than other classical adjuvants.91 Akkermansia muciniphila OMVs restore disturbed balance of the gut microbiota by selectively promoting the proliferation of beneficial bacteria through membrane fusion, and thus overcome primary resistance to PD-1 blockade in tumor immunotherapy.98 S. aureus OMVs increase p−ERK and p−AKT levels, while tamoxifen inhibits breast cancer cell growth by modulating PI3K/AKT, ERK, and IGF receptors, thereby enhancing tamoxifen efficacy when treating breast cancer.99 OMVs derived from transgenic E. coli, modified with αvβ3 integrin targeting ligand and indocyanine green (named as I-P-OMVs), are developed to induce the transdermal photo-TRAIL-programmed treatment in skin melanoma.100
Additionally, researchers have proposed loading OMVs with chemotherapeutics to integrate the advantages of nanotherapy, immunotherapy, and chemotherapy into a single platform. Especially, surface modification with cancer-targeting ligands can minimize off-tumor toxicity and multidrug resistance.35 Doxorubicin-loaded OMVs (DOX-OMV) are prepared and demonstrated to function as both biologic nanocarriers for chemotherapeutic agents and to recruit TME macrophages to synergize with DOX, inhibiting lung cancer progression.101 Nanoparticles, IR780@PLGA@HM, are constructed by fusing breast cancer cell membranes and OMVs to form a hybrid membrane (HM) and then encapsulating IR780-loaded PLGA, with tumor targeting, immune regulating, and Sonodynamic abilities to treat breast cancer bone metastasis.102
Other Therapeutic Options
Besides therapies mentioned above, multiple therapeutic methods with OMVs participated are emerging. For example, Tang et al develop the enzyme-powered bacterial OMV nanorobots by surface bioengineering the robotic body with cell-penetrating peptide for tumor targeting and penetration, which is further enhanced by active propulsion of nanorobots. These nanorobots can also effectively safeguard the loaded gene silencing tool, siRNA, from enzymatic degradation. Systematic in vitro and in vivo studies in rodent models have demonstrated that these OMV nanorobots can substantially enhance siRNA delivery and immune stimulation.103
Notably, OMVs are endowed with pathogen-associated molecular pattern (PAMP)-rich membranes and biogenesis-dependent cargoes encompassing proteins and nucleic acids.23 Consequently, every contemplated clinical application must rigorously address formulation safety, including scrutiny of endotoxin burden, immunogenic potential, and inter-batch variability. For example, in order to manage the immunogenicity of OMVs, calcium phosphate (CaP) shells can be employed to cover the surface of OMVs to alleviate the side effects of intravenous administration.106 Moreover, in the PEG/Se@OMV-CD47nb mentioned above, the surface PEG/Se layer endows the nanoparticles with radiation-triggered controlled release of OMV-CD47nb, also mitigating the side effects observed from intravenous injection of naked OMVs.93
Future Perspectives
Over the past few years, studies have established that OMVs can bidirectionally modulate tumor proliferation, metastasis and the immune microenvironment. Nevertheless, mechanistic dissection largely remains descriptive and confined to cataloguing discrete signaling pathways. By integrating published OMV cargo profiles with tumor-phenotype datasets and applying deep-learning algorithms, future work could construct a tripartite knowledge graph of “OMV cargo - host pathway - tumor outcome”. Mining this graph for recurrent patterns will provide perspectives for clarifying how cargo heterogeneity among OMVs of distinct bacterial origins dictates their pro- or anti-tumor selectivity, and how the same OMV elicits divergent phenotypes when exposed to different cancer type or microenvironment. Such a framework will yield molecular signatures for distinguishing “risk-associated” from “therapeutic” bacterial sources. Meanwhile, through interdisciplinary integration, the knowledge graph can be coupled with an artificial intelligence-driven predictive model. By means of transfer learning, this model could be extrapolated to novel bacterial strains or uncharacterized cargo, enabling in-silico screening and risk/efficacy prediction of OMV functionalities, and remarkably compressing the period from discovery to preclinical validation.
In addition to natural OMVs, engineered OMVs are a research focus in recent years. Diverse technologies have emerged to modify OMVs, including displaying tumor antigens on the vesicle surface for vaccination purposes by genetically editing the parental bacterium, presenting immunomodulatory ligands for onco-immune reprogramming, encapsulating therapeutic payloads for targeted delivery, or by coated and hybrid membrane technologies, constructing hybrid nanoplatforms that exert antitumor effects.107,108 Looking forward, researchers can exploit CRISPR-Cas systems and synthetic biology to install controllable switches at OMV biogenesis checkpoints, thereby enabling precision cargo loading and programmed release. Integration with existing immunotherapeutic formulations—for example, the PEG/Se@OMV-CD47nb construct mentioned above—could establish sequential regimens in which immune activation precedes drug release, thereby enhancing therapeutic efficacy and elevating OMVs from passive vectors to intelligent therapeutic systems.
Moreover, the clinical application of OMVs could be extended to encompass early-stage prevention, diagnostic assistance, post-metastatic maintenance, and so on. In imminent Phase I trials, the patients’ metagenomic profiling of microbiota and parallel characterization of circulating OMVs could be performed in real time to establish a “bacterial source–OMV–efficacy” monitoring framework. This will explore the potential of precision-medicine paradigms such as “same disease, different treatment” and “different diseases, same treatment,” thereby maximizing the clinical value of each OMV formulation. For patients with established metastases, it should be investigated whether OMVs engineered to carry specific cargos can re-program dormant micro-metastatic niches, thereby prolonging progression-free survival.
However, OMV-based therapeutics still face many challenges in its route to clinical deployment, including the absence of a standardized isolation protocol and quality control specification, side effects of intravenous administration, and the potential intoxication. Besides, most pre-clinical studies relied on model antigens or murine tumor antigens, and thus the adaptability of the delivery systems to human tumor antigens needs further study. The OMV-based strategies delineated above largely remain confined to pre-clinical investigation, with efficacy documented solely in animal models. It is anticipated that an expanding repertoire of OMV-based formulations will successfully advance through clinical translation.
Meanwhile, it remains to be systematically determined whether—and to what extent—OMVs release is modulated by extrinsic cues such as microbiome composition, dietary constituents, or antibiotic exposure, and how these variables subsequently shape cancer susceptibility or therapeutic outcomes. Elucidating and ultimately taking advantage of these modifiable determinants could markedly enhance the efficiency of OMV-based interventions in clinical appliances.
Conclusions
In conclusion, OMVs can bidirectionally modulate tumor development by influencing the cancer cells through their diverse molecular cargos in various pathways. Promising results from studies show that natural or engineered OMVs could serve as effective vaccines, immune-therapeutics and adjuvants and can also synergize with chemotherapy or checkpoint blockade to overcome resistance. To advance these findings, future efforts could combine them with different intelligent models through multidisciplinary tools, the isolation and purification process should be further standardized, and more factors of OMVs release should be studied, guiding the clinical transformation of OMVs from basic research to precision cancer prevention and treatment.
Abbreviations
EVs, Extracellular vesicles; MVs, Membrane vesicles; OMVs, Outer membrane vesicles; OIMVs, Outer-inner membrane vesicles; EOMVs, Explosive outer-membrane vesicles; CMVs, Cytoplasmic membrane vesicles); TSMSs, Tube-shaped membranous structures; h.p OMVs, Helicobacter pylori OMVs; OSM, Oncostatin-M; OSMRβ, Oncostatin-M type II receptor; AGS, Human gastric adenocarcinoma cell line; ROS, Reactive oxygen species; GSH, Glutathione; CCC, Colon cancer cells; SOD, Superoxide dismutase; CRC, Colorectal cancer cell; CRC/5FUR cells, 5-fluorouracil-resistant CRC cells; EMT, Epithelial–mesenchymal transition; DSC-2, Desmocollin-2; TEM, Trans-endothelial migration; VEGFC/D, Vascular endothelial growth factor C/D; IGF-1, Insulin-like growth factor 1; ZO, Zona Occludens; IECs, intestinal epithelial cells; EPEC, enteropathogenic E. coli; EcN, E. coli Nissle 1917; ETEC, enterotoxigenic E. coli; TME, Tumor microenvironment; EPR effect, enhanced permeability and retention effect; PAMPs, Pathogen-associated molecular patterns; GZMB+, Granzyme B-positive; IFN-γ+, Interferon γ-positive; GGT, γ-glutamyl transpeptidase; PD-L1, Programmed death ligand 1; RIP2, Receptor-interacting serine/threonine-protein kinase 2; IAV, Influenza A virus; KGF-2, Keratinocyte growth factor-2; TIrV, Trained immunity-related vaccines; SyBV, Synthetic Bacterial Vesicles; TAMs, Tumor-associated macrophages; TfR-LYTAC, transferrin receptor-mediated lysosome-targeting chimera; OMV-NICE, OMV-based nanosized immune cell engager; CaP, Calcium phosphate; DAMPs, damage-associated molecular patterns; HM, Hybrid membrane.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors declare that no financial support was received for the research, authorship, and/or publication of this article.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Schwechheimer C, Kuehn MJ. Outer-membrane vesicles from Gram-negative bacteria: biogenesis and functions. Nat Rev Microbiol. 2015;13(10):605–21. doi:10.1038/nrmicro3525
2. Remis JP, Wei D, Gorur A, et al. Bacterial social networks: structure and composition of Myxococcus xanthus outer membrane vesicle chains. Environ Microbiol. 2014;16(2):598–610. doi:10.1111/1462-2920.12187
3. McCaig WD, Koller A, Thanassi DG. Production of outer membrane vesicles and outer membrane tubes by Francisella novicida. J Bacteriol. 2013;195(6):1120–1132. doi:10.1128/JB.02007-12
4. Guerrero-Mandujano A, Hernández-Cortez C, Ibarra JA, Castro-Escarpulli G. The outer membrane vesicles: secretion system type zero. Traffic. 2017;18(7):425–432. doi:10.1111/tra.12488
5. Toyofuku M, Schild S, Kaparakis-Liaskos M, Eberl L. Composition and functions of bacterial membrane vesicles. Nat Rev Microbiol. 2023;21(7):415–430. doi:10.1038/s41579-023-00875-5
6. Kulp A, Kuehn MJ. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol. 2010;64(1):163–184. doi:10.1146/annurev.micro.091208.073413
7. Pérez-Cruz C, Carrión O, Delgado L, Martinez G, López-Iglesias C, Mercade E. New type of outer membrane vesicle produced by the Gram-negative bacterium Shewanella vesiculosa M7T: implications for DNA content. Appl Environ Microbiol. 2013;79(6):1874–1881. doi:10.1128/AEM.03657-12
8. Pérez-Cruz C, Delgado L, López-Iglesias C, Mercade E. Outer-inner membrane vesicles naturally secreted by gram-negative pathogenic bacteria. PLoS One. 2015;10(1):e0116896. doi:10.1371/journal.pone.0116896
9. Turnbull L, Toyofuku M, Hynen AL, et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat Commun. 2016;7(1):11220. doi:10.1038/ncomms11220
10. Toyofuku M, Cárcamo-Oyarce G, Yamamoto T, et al. Prophage-triggered membrane vesicle formation through peptidoglycan damage in Bacillus subtilis. Nat Commun. 2017;8(1):481.
11. Baidya AK, Bhattacharya S, Dubey GP, Mamou G, Ben-Yehuda S. Bacterial nanotubes: a conduit for intercellular molecular trade. Curr Opin Microbiol. 2018;42:1–6. doi:10.1016/j.mib.2017.08.006
12. Toyofuku M, Nomura N, Eberl L. Types and origins of bacterial membrane vesicles. Nat Rev Microbiol. 2019;17(1):13–24. doi:10.1038/s41579-018-0112-2
13. György B, Szabó TG, Pásztói M, et al. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell Mol Life Sci. 2011;68(16):2667–2688. doi:10.1007/s00018-011-0689-3
14. Gill S, Catchpole R, Forterre P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol Rev. 2019;43(3):273–303. doi:10.1093/femsre/fuy042
15. Sepich-Poore GD, Zitvogel L, Straussman R, Hasty J, Wargo JA, Knight R. The microbiome and human cancer. Science. 2021;371(6536):eabc4552. doi:10.1126/science.abc4552
16. Jin C, Lagoudas GK, Zhao C, et al. Commensal Microbiota Promote Lung Cancer Development via γδ T Cells. Cell. 2019;176(5):998–1013.e16. doi:10.1016/j.cell.2018.12.040
17. Matson V, Fessler J, Bao R, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. 2018;359(6371):104–108. doi:10.1126/science.aao3290
18. Tsay JCJ, Wu BG, Sulaiman I, et al. Lower Airway Dysbiosis Affects Lung Cancer Progression. Cancer Discov. 2021;11(2):293–307. doi:10.1158/2159-8290.CD-20-0263
19. Kadosh E, Snir-Alkalay I, Venkatachalam A, et al. The gut microbiome switches mutant p53 from tumour-suppressive to oncogenic. Nature. 2020;586(7827):133–138. doi:10.1038/s41586-020-2541-0
20. Basoya R, Singh B, Basi A, Aggarwal S. Chapter Three - Role of microbiome in cancer progression. In: Singh M. editor. International Review of Cell and Molecular Biology. Academic Press. 2025;Vol. 394:79–106. Available from: https://www.sciencedirect.com/science/article/pii/S1937644824001722.
21. Kuehn MJ, Kesty NC. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev. 2005;19(22):2645–2655. doi:10.1101/gad.1299905
22. Koeppen K, Hampton TH, Jarek M, et al. A Novel Mechanism of Host-Pathogen Interaction through sRNA in Bacterial Outer Membrane Vesicles. PLoS Pathog. 2016;12(6):e1005672. doi:10.1371/journal.ppat.1005672
23. Choi D, Lee EY. Standardizing bacterial extracellular vesicle purification: a call for consensus. J Microbiol Biotechnol. 2025;35:e2506011. doi:10.4014/jmb.2506.06011
24. Sartorio MG, Pardue EJ, Feldman MF, Haurat MF. Bacterial Outer Membrane Vesicles: from Discovery to Applications. Annu Rev Microbiol. 2021;75(1):609–630. doi:10.1146/annurev-micro-052821-031444
25. Hanahan D, Weinberg RA. The Hallmarks of Cancer. Cell. 2000;100(1):57–70. doi:10.1016/S0092-8674(00)81683-9
26. Hanahan D, Weinberg RA. Hallmarks of Cancer: the Next Generation. Cell. 2011;144(5):646–674. doi:10.1016/j.cell.2011.02.013
27. Hanahan D. Hallmarks of Cancer: new Dimensions. Cancer Discov. 2022;12(1):31–46. doi:10.1158/2159-8290.CD-21-1059
28. Abolhasani FS, Vaghefinanekaran N, Yarahmadi A, et al. Outer membrane vesicles in gram-negative bacteria and its correlation with pathogenesis. Front Immunol. 2025;16:1541636. doi:10.3389/fimmu.2025.1541636
29. Wu Z, Long W, Yin Y, et al. Outer membrane vesicles of Porphyromonas gingivalis: recent advances in pathogenicity and associated mechanisms. Front Microbiol. 2025;16:1555868. doi:10.3389/fmicb.2025.1555868
30. Masjedi A, Hajizadeh F, Beigi Dargani F, et al. Oncostatin M: a mysterious cytokine in cancers. Int Immunopharmacol. 2021;90:107158. doi:10.1016/j.intimp.2020.107158
31. Zoaiter M, Nasser R, Hage-Sleiman R, Abdel-Sater F, Badran B, Zeaiter Z. Helicobacter pylori outer membrane vesicles induce expression and secretion of oncostatin M in AGS gastric cancer cells. Braz J Microbiol. 2021;52(3):1057–1066. doi:10.1007/s42770-021-00490-7
32. Shi J, Xu X, Du J, Cui H, Luo Q. Expression of Oncostatin M in Early Gastric Cancer and Precancerous Lesions. Gastroenterol Res Pract. 2019;2019:3616140. doi:10.1155/2019/3616140
33. Yu Z, Li Z, Wang C, et al. Oncostatin M receptor, positively regulated by SP1, promotes gastric cancer growth and metastasis upon treatment with Oncostatin M. Gastric Cancer. 2019;22(5):955–966. doi:10.1007/s10120-019-00934-y
34. Li G, Sun Y, Huang Y, et al. Fusobacterium nucleatum-derived small extracellular vesicles facilitate tumor growth and metastasis via TLR4 in breast cancer. BMC Cancer. 2023;23(1):473. doi:10.1186/s12885-023-10844-z
35. Aly RGO, El-Enbaawy MIH, Abd El-Rahman SS, Ata NS. Antineoplastic activity of Salmonella Typhimurium outer membrane nanovesicles. Exp Cell Res. 2021;399(1):112423. doi:10.1016/j.yexcr.2020.112423
36. Jin L, Zhang Z, Tan X, et al. Antitumor effect of Escherichia coli-derived outer membrane vesicles on neuroblastoma in vitro and in vivo. Acta Biochim Biophys Sin. 2022;54(9):1301–1313. doi:10.3724/abbs.2022127
37. Gurunathan S, Ajmani A, Kim JH. Extracellular nanovesicles produced by Bacillus licheniformis: a potential anticancer agent for breast and lung cancer. Microb Pathog. 2023;185:106396. doi:10.1016/j.micpath.2023.106396
38. Luo Z-W, Xia K, Liu Y-W, et al. Extracellular Vesicles from Akkermansia muciniphila Elicit Antitumor Immunity Against Prostate Cancer via Modulation of CD8+ T Cells and Macrophages. Int J Nanomed. 2021;16:2949–2963. doi:10.2147/IJN.S304515
39. Keyhani G, Mahmoodzadeh Hosseini H, Salimi A. Effect of extracellular vesicles of Lactobacillus rhamnosus GG on the expression of CEA gene and protein released by colorectal cancer cells. Iran J Microbiol. 2022;14(1):90–96. doi:10.18502/ijm.v14i1.8809
40. Wu X, Xu J, Yang X, Wang D, Xu X. Integrating Transcriptomics and Metabolomics to Explore the Novel Pathway of Fusobacterium nucleatum Invading Colon Cancer Cells. Pathogens. 2023;12(2):201. doi:10.3390/pathogens12020201
41. Behzadi E, Mahmoodzadeh Hosseini H, Imani Fooladi AA. The inhibitory impacts of Lactobacillus rhamnosus GG-derived extracellular vesicles on the growth of hepatic cancer cells. Microb Pathog. 2017;110:1–6. doi:10.1016/j.micpath.2017.06.016
42. Dehghani N, Tafvizi F, Jafari P. Cell cycle arrest and anti-cancer potential of probiotic Lactobacillus rhamnosus against HT-29 cancer cells. Bioimpacts. 2021;11(4):245–252. doi:10.34172/bi.2021.32
43. Abedi A, Tafvizi F, Jafari P, Akbari N. The inhibition effects of Lentilactobacillus buchneri-derived membrane vesicles on AGS and HT-29 cancer cells by inducing cell apoptosis. Sci Rep. 2024;14(1):3100. doi:10.1038/s41598-024-53773-y
44. Jiang Y, Wang L, Yang B, et al. Bifidobacterium-derived membrane vesicles inhibit triple-negative breast cancer growth by inducing tumor cell apoptosis. Mol Biol Rep. 2023;50(9):7547–7556. doi:10.1007/s11033-023-08702-z
45. Caballano-Infantes E, Ho-Plágaro A, López-Gómez C, et al. Membrane Vesicles of Toxigenic Clostridioides difficile Affect the Metabolism of Liver HepG2 Cells. Antioxidants. 2023;12(4):818. doi:10.3390/antiox12040818
46. Marzoog TR, Jabir MS, Ibraheem S, et al. Bacterial extracellular vesicles induced oxidative stress and mitophagy through mTOR pathways in colon cancer cells, HT-29: implications for bioactivity. Biochim Biophys Acta Mol Cell Res. 2023;1870(6):119486. doi:10.1016/j.bbamcr.2023.119486
47. Shi Y, Meng L, Zhang C, Zhang F, Fang Y. Extracellular vesicles of Lacticaseibacillus paracasei PC-H1 induce colorectal cancer cells apoptosis via PDK1/AKT/Bcl-2 signaling pathway. Microbiol Res. 2021;255:126921. doi:10.1016/j.micres.2021.126921
48. An J, Ha EM. Extracellular vesicles derived from Lactobacillus plantarum restore chemosensitivity through the PDK2-mediated glucose metabolic pathway in 5-FU-resistant colorectal cancer cells. J Microbiol. 2022;60(7):735–745. doi:10.1007/s12275-022-2201-1
49. Hong SW, Choi EB, Min TK, et al. An important role of α-hemolysin in extracellular vesicles on the development of atopic dermatitis induced by Staphylococcus aureus. PLoS One. 2014;9(7):e100499. doi:10.1371/journal.pone.0100499
50. Bitto NJ, Cheng L, Johnston EL, et al. Staphylococcus aureus membrane vesicles contain immunostimulatory DNA, RNA and peptidoglycan that activate innate immune receptors and induce autophagy. J Extracell Vesicles. 2021;10(6):e12080. doi:10.1002/jev2.12080
51. Fidler IJ. The pathogenesis of cancer metastasis: the “seed and soil” hypothesis revisited. Nat Rev Cancer. 2003;3(6):453–458. doi:10.1038/nrc1098
52. Lin LT, Shi YC, Choong CY, Tai CJ. The Fruits of Paris polyphylla Inhibit Colorectal Cancer Cell Migration Induced by Fusobacterium nucleatum-Derived Extracellular Vesicles. Molecules. 2021;26(13):4081. doi:10.3390/molecules26134081
53. Chen G, Gao C, Jiang S, et al. Fusobacterium nucleatum outer membrane vesicles activate autophagy to promote oral cancer metastasis. J Adv Res. 2024;56:167–179. doi:10.1016/j.jare.2023.04.002
54. Liu D, Liu S, Liu J, Miao L, Zhang S, Pan Y. sRNA23392 packaged by Porphyromonas gingivalis outer membrane vesicles promotes oral squamous cell carcinomas migration and invasion by targeting desmocollin-2. Mol Oral Microbiol. 2021;36(3):182–191. doi:10.1111/omi.12334
55. Wei S, Zhang J, Wu X, et al. Fusobacterium nucleatum Extracellular Vesicles Promote Experimental Colitis by Modulating Autophagy via the miR-574-5p/CARD3 Axis. Inflamm Bowel Dis. 2023;29(1):9–26. doi:10.1093/ibd/izac177
56. Alvarez CS, Giménez R, Cañas MA, et al. Extracellular vesicles and soluble factors secreted by Escherichia coli Nissle 1917 and ECOR63 protect against enteropathogenic E. coli-induced intestinal epithelial barrier dysfunction. BMC Microbiol. 2019;19(1):166. doi:10.1186/s12866-019-1534-3
57. Alvarez CS, Badia J, Bosch M, Giménez R, Baldomà L. Outer Membrane Vesicles and Soluble Factors Released by Probiotic Escherichia coli Nissle 1917 and Commensal ECOR63 Enhance Barrier Function by Regulating Expression of Tight Junction Proteins in Intestinal Epithelial Cells. Front Microbiol. 2016;7:1981. doi:10.3389/fmicb.2016.01981
58. Pang Y, Ermann Lundberg L, Mata Forsberg M, et al. Extracellular membrane vesicles from Limosilactobacillus reuteri strengthen the intestinal epithelial integrity, modulate cytokine responses and antagonize activation of TRPV1. Front Microbiol. 2022;13:1032202. doi:10.3389/fmicb.2022.1032202
59. Li J, Feng S, Pi Y, et al. Limosilactobacillus johnsoni and Limosilactobacillus mucosae and Their Extracellular Vesicles Alleviate Gut Inflammatory Injury by Mediating Macrophage Polarization in a Lipopolysaccharide-Challenged Piglet Model. J Nutr. 2023;153(8):2497–2511. doi:10.1016/j.tjnut.2023.06.009
60. Qian B, Deng Y, Im JH, et al. A distinct macrophage population mediates metastatic breast cancer cell extravasation, establishment and growth. PLoS One. 2009;4(8):e6562. doi:10.1371/journal.pone.0006562
61. Hatakeyama M. Helicobacter pylori CagA and gastric cancer: a paradigm for hit-and-run carcinogenesis. Cell Host Microbe. 2014;15(3):306–316. doi:10.1016/j.chom.2014.02.008
62. Shimoda A, Ueda K, Nishiumi S, et al. Exosomes as nanocarriers for systemic delivery of the Helicobacter pylori virulence factor CagA. Sci Rep. 2016;6(1):18346. doi:10.1038/srep18346
63. Turkina MV, Olofsson A, Magnusson KE, Arnqvist A, Vikström E. Helicobacter pylori vesicles carrying CagA localize in the vicinity of cell-cell contacts and induce histone H1 binding to ATP in epithelial cells. FEMS Microbiol Lett. 2015;362(11):fnv076. doi:10.1093/femsle/fnv076
64. Reymond N, D’água BB, Ridley AJ. Crossing the endothelial barrier during metastasis. Nat Rev Cancer. 2013;13(12):858–870. doi:10.1038/nrc3628
65. Stacker SA, Achen MG, Jussila L, Baldwin ME, Alitalo K. Lymphangiogenesis and cancer metastasis. Nat Rev Cancer. 2002;2(8):573–583. doi:10.1038/nrc863
66. Stacker SA, Williams SP, Karnezis T, Shayan R, Fox SB, Achen MG. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat Rev Cancer. 2014;14(3):159–172. doi:10.1038/nrc3677
67. Björndahl M, Cao R, Nissen LJ, et al. Insulin-like growth factors 1 and 2 induce lymphangiogenesis in vivo. Proc Natl Acad Sci U S A. 2005;102(43):15593–15598. doi:10.1073/pnas.0507865102
68. Qian BZ, Li J, Zhang H, et al. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature. 2011;475(7355):222–225. doi:10.1038/nature10138
69. Pan J, Li X, Shao B, et al. Self-Blockade of PD-L1 with Bacteria-Derived Outer-Membrane Vesicle for Enhanced Cancer Immunotherapy. Adv Mater. 2022;34(7):e2106307. doi:10.1002/adma.202106307
70. Won S, Lee C, Bae S, et al. Mass-produced gram-negative bacterial outer membrane vesicles activate cancer antigen-specific stem-like CD8+ T cells which enables an effective combination immunotherapy with anti-PD-1. J Extracell Vesicles. 2023;12(8):e12357. doi:10.1002/jev2.12357
71. Firth J, Sun J, George V, Huang JD, Bajaj-Elliott M, Gustafsson K. Bacterial outer-membrane vesicles promote Vγ9Vδ2 T cell oncolytic activity. Front Immunol. 2023;14:1198996. doi:10.3389/fimmu.2023.1198996
72. Jiang Y, Xu Y, Zheng C, et al. Acetyltransferase from Akkermansia muciniphila blunts colorectal tumourigenesis by reprogramming tumour microenvironment. Gut. 2023;72(7):1308–1318. doi:10.1136/gutjnl-2022-327853
73. Kim OY, Park HT, Dinh NTH, et al. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat Commun. 2017;8(1):626. doi:10.1038/s41467-017-00729-8
74. Z G, D R, P F, et al. Effects of Helicobacter suis γ-glutamyl transpeptidase on lymphocytes: modulation by glutamine and glutathione supplementation and outer membrane vesicles as a putative delivery route of the enzyme. PLoS One. 2013;8(10):2.
75. Askarian F, Lapek JD, Dongre M, et al. Staphylococcus aureus Membrane-Derived Vesicles Promote Bacterial Virulence and Confer Protective Immunity in Murine Infection Models. Front Microbiol. 2018;9:262. doi:10.3389/fmicb.2018.00262
76. Reyes-Robles T, Torres VJ. Staphylococcus aureus Pore-Forming Toxins. Curr Top Microbiol Immunol. 2017;409:121–144. doi:10.1007/82_2016_16
77. Brinkmann V, Reichard U, Goosmann C, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532–1535. doi:10.1126/science.1092385
78. Groeger S, Denter F, Lochnit G, Schmitz ML, Meyle J. Porphyromonas gingivalis Cell Wall Components Induce Programmed Death Ligand 1 (PD-L1) Expression on Human Oral Carcinoma Cells by a Receptor-Interacting Protein Kinase 2 (RIP2)-Dependent Mechanism. Infect Immun. 2020;88(5):e00051–20. doi:10.1128/IAI.00051-20
79. Liao T, Chen X, Qiu F, et al. Correction: regulation of cancer‑associated fibroblasts for enhanced cancer immunotherapy using advanced functional nanomedicines: an updated review. J Nanobiotechnol. 2025;23(1):269. doi:10.1186/s12951-025-03344-8
80. Hock BD, McKenzie JL, Keenan JI. Helicobacter pylori outer membrane vesicles inhibit human T cell responses via induction of monocyte COX-2 expression. Pathog Dis. 2017;75(4). doi:10.1093/femspd/ftx034
81. Zhang H, Zhang Y, Song Z, et al. sncRNAs packaged by Helicobacter pylori outer membrane vesicles attenuate IL-8 secretion in human cells. Int J Med Microbiol. 2020;310(1):151356. doi:10.1016/j.ijmm.2019.151356
82. Keir ME, Butte MJ, Freeman GJ, Sharpe AH. PD-1 and its ligands in tolerance and immunity. Ann Rev Immunol. 2008;26(1):677–704. doi:10.1146/annurev.immunol.26.021607.090331
83. Zhang Y, Fang Z, Li R, Huang X, Liu Q. Design of Outer Membrane Vesicles as Cancer Vaccines: a New Toolkit for Cancer Therapy. Cancers. 2019;11(9):1314. doi:10.3390/cancers11091314
84. Perrett KP, McVernon J, Richmond PC, et al. Immune responses to a recombinant, four-component, meningococcal serogroup B vaccine (4CMenB) in adolescents: a Phase III, randomized, multicentre, lot-to-lot consistency study. Vaccine. 2015;33(39):5217–5224. doi:10.1016/j.vaccine.2015.06.103
85. Ahmed AAQ, Qi F, Zheng R, et al. The impact of ExHp-CD (outer membrane vesicles) released from Helicobacter pylori SS1 on macrophage RAW 264.7 cells and their immunogenic potential. Life Sci. 2021;279:119644. doi:10.1016/j.lfs.2021.119644
86. Liu Q, Li X, Zhang Y, et al. Orally-administered outer-membrane vesicles from Helicobacter pylori reduce H. pylori infection via Th2-biased immune responses in mice. Pathog Dis. 2019;77(5):ftz050. doi:10.1093/femspd/ftz050
87. Wang X, Lee JC. Staphylococcus aureus membrane vesicles: an evolving story. Trends Microbiol. 2024;32(11):1096–1105. doi:10.1016/j.tim.2024.04.003
88. Choi SJ, Kim MH, Jeon J, et al. Active Immunization with Extracellular Vesicles Derived from Staphylococcus aureus Effectively Protects against Staphylococcal Lung Infections, Mainly via Th1 Cell-Mediated Immunity. PLoS One. 2015;10(9):e0136021. doi:10.1371/journal.pone.0136021
89. Wang X, Thompson CD, Weidenmaier C, Lee JC. Release of Staphylococcus aureus extracellular vesicles and their application as a vaccine platform. Nat Commun. 2018;9(1):1379. doi:10.1038/s41467-018-03847-z
90. Carvalho AL, Fonseca S, Miquel-Clopés A, et al. Bioengineering commensal bacteria-derived outer membrane vesicles for delivery of biologics to the gastrointestinal and respiratory tract. J Extracell Vesicles. 2019;8(1):1632100. doi:10.1080/20013078.2019.1632100
91. Park KS, Svennerholm K, Crescitelli R, Lässer C, Gribonika I, Lötvall J. Synthetic bacterial vesicles combined with tumour extracellular vesicles as cancer immunotherapy. J Extracell Vesicles. 2021;10(9):e12120. doi:10.1002/jev2.12120
92. Liang J, Zhu F, Cheng K, et al. Outer Membrane Vesicle-Based Nanohybrids Target Tumor-Associated Macrophages to Enhance Trained Immunity-Related Vaccine-Generated Antitumor Activity. Adv Mater. 2023;35(46):e2306158. doi:10.1002/adma.202306158
93. Feng Q, Ma X, Cheng K, et al. Engineered Bacterial Outer Membrane Vesicles as Controllable Two-Way Adaptors to Activate Macrophage Phagocytosis for Improved Tumor Immunotherapy. Adv Mater. 2022;34(40):e2206200. doi:10.1002/adma.202206200
94. Su LY, Tian Y, Zheng Q, et al. Anti-tumor immunotherapy using engineered bacterial outer membrane vesicles fused to lysosome-targeting chimeras mediated by transferrin receptor. Cell Chem Biol. 2024;31(6):1219–1230.e5. doi:10.1016/j.chembiol.2024.01.002
95. Sun J, Tan L, Ye BC, Bi X. Engineered Outer Membrane Vesicles as Nanosized Immune Cell Engagers for Enhanced Solid Tumor Immunotherapy. ACS Nano. 2024;18(44):30332–30344. doi:10.1021/acsnano.4c07364
96. Sun Y, Ma YY, Shangguan S, et al. Metal ions-anchored bacterial outer membrane vesicles for enhanced ferroptosis induction and immune stimulation in targeted antitumor therapy. J Nanobiotechnology. 2024;22(1):474. doi:10.1186/s12951-024-02747-3
97. Chen X, Li P, Luo B, et al. Surface Mineralization of Engineered Bacterial Outer Membrane Vesicles to Enhance Tumor Photothermal/Immunotherapy. ACS Nano. 2024;18(2):1357–1370. doi:10.1021/acsnano.3c05714
98. Wang X, Lin S, Wang L, et al. Versatility of bacterial outer membrane vesicles in regulating intestinal homeostasis. Sci Adv. 2023;9(11):eade5079. doi:10.1126/sciadv.ade5079
99. An J, Kwon H, Lim W, Moon BI. Staphylococcus aureus-Derived Extracellular Vesicles Enhance the Efficacy of Endocrine Therapy in Breast Cancer Cells. J Clin Med. 2022;11(7):2030. doi:10.3390/jcm11072030
100. Peng LH, Wang MZ, Chu Y, et al. Engineering bacterial outer membrane vesicles as transdermal nanoplatforms for photo-TRAIL-programmed therapy against melanoma. Sci Adv. 2020;6(27):eaba2735. doi:10.1126/sciadv.aba2735
101. Kuerban K, Gao X, Zhang H, et al. Doxorubicin-loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-small-cell lung cancer. Acta Pharm Sin B. 2020;10(8):1534–1548. doi:10.1016/j.apsb.2020.02.002
102. Wang J, Liang S, Chen S, et al. Bacterial outer membrane vesicle-cancer cell hybrid membrane-coated nanoparticles for sonodynamic therapy in the treatment of breast cancer bone metastasis. J Nanobiotechnology. 2024;22(1):328. doi:10.1186/s12951-024-02619-w
103. Tang S, Tang D, Zhou H, et al. Bacterial outer membrane vesicle nanorobot. Proc Natl Acad Sci U S A. 2024;121(30):e2403460121. doi:10.1073/pnas.2403460121
104. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
105. Devati MS, Jnana A, Kidd SP, et al. Decoding bacterial extracellular vesicles: a review on isolation and characterization techniques. Arch Microbiol. 2025;208(1):63. doi:10.1007/s00203-025-04628-1
106. Qing S, Lyu C, Zhu L, et al. Biomineralized bacterial outer membrane vesicles potentiate safe and efficient tumor microenvironment reprogramming for anticancer therapy. Adv Mater. 2020;32(47):e2002085. doi:10.1002/adma.202002085
107. Liu J, Wang T, Zhou Y, et al. Bacterial outer membrane vesicles in tumor prevention and treatment: advancements in research and application. J Mater Chem B. 2025;2025:1.
108. Aytar Çelik P, Erdogan-Gover K, Barut D, et al. Bacterial Membrane Vesicles as Smart Drug Delivery and Carrier Systems: a New Nanosystems Tool for Current Anticancer and Antimicrobial Therapy. Pharmaceutics. 2023;15(4):1052. doi:10.3390/pharmaceutics15041052
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Overexpression of TREM1 is Associated with the Immune-Suppressive Microenvironment and Unfavorable Prognosis in Pan-Cancer
Zhou X, Lin K, Fu L, Liu F, Lin H, Chen Y, Zhuang B, Liang H, Deng Q, Wang Z, Chen W, Luo J, Cao J, Li P
Journal of Inflammation Research 2023, 16:1375-1391
Published Date: 27 March 2023
Exploring the Correlation Between GPR176, a Potential Target Gene of Gastric Cancer, and Immune Cell Infiltration
Gu X, Shen H, Xiang Z, Li X, Zhang Y, Zhang R, Su F, Wang Z
Pharmacogenomics and Personalized Medicine 2023, 16:519-535
Published Date: 1 June 2023
Integrating Bulk and Single-Cell RNA Sequencing Reveals Heterogeneity, Tumor Microenvironment, and Immunotherapeutic Efficacy Based on Sialylation-Related Genes in Bladder Cancer
Tan Z, Chen X, Zuo J, Fu S, Wang J, Wang H
Journal of Inflammation Research 2023, 16:3399-3417
Published Date: 14 August 2023
NOTCH1 Mutations Predict Superior Outcomes of Immune Checkpoint Blockade in Non-Small Cell Lung Cancer
Huang Q, Cao H, Yao Q, Zhou X, Li H, Bai Q, Hu H
ImmunoTargets and Therapy 2023, 12:165-173
Published Date: 5 December 2023
Nanodynamic Therapy in Colorectal Cancer: Engineering Precision Immunotherapy and Multimodal Synergy
Wang F, Zhang Z, Chen J, Qian L, Chen B, Zhai L, Zhao J, Chen T, Zhou J, Gou K, Zhao Z, Zhu X, Xiao Z
International Journal of Nanomedicine 2026, 21:598076
Published Date: 23 March 2026