Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Orbital Fat is an Observation Model to Provide Insights into Adipocyte Hypertrophy and Hyperplasia During White Adipose Tissue Expansion
Authors Cheng Y, Zhang K, Liu J, Liu G ![]()
Received 19 February 2025
Accepted for publication 28 July 2025
Published 20 August 2025 Volume 2025:18 Pages 2977—2984
DOI https://doi.org/10.2147/DMSO.S521845
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Pablo Corral
Yu Cheng,1,* Kaili Zhang,1,* Jisong Liu,2 Guangpeng Liu1
1Department of Plastic and Reconstructive Surgery, Shanghai Tenth People’s Hospital, School of Medicine, Tongji University, Shanghai, People’s Republic of China; 2Department of Burn and Plastic Surgery, The Third People’s Hospital of Bengbu, Bengbu, Anhui, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Guangpeng Liu, Department of Plastic and Reconstructive Surgery, Shanghai Tenth People’s Hospital, School of Medicine, Tongji University, 301 Yanchang Road, Shanghai, 200072, People’s Republic of China, Email [email protected] Jisong Liu, Department of Burn and Plastic Surgery, The Third People’s Hospital of Bengbu, 38 Shengli Middle Road, Bengbu, Anhui, 233000, People’s Republic of China, Email [email protected]
Introduction: Obesity is a global health problem characterized by excessive white adipose tissue (WAT) distribution. Adipocyte hypertrophy (increased cell size) and hyperplasia (differentiation into new adipocytes from pre-adipocytes) are two ways for WAT to expand. The precedence of hypertrophy over hyperplasia leads to an enlarged adipocyte size, which is associated with multiple metabolic dysfunctions. Compared with abdominal subcutaneous fat (SF), orbital fat (OF) has smaller adipocytes with less inflammatory infiltration, better vascularization, and higher adipogenic and proliferative capacities, reflecting a healthy metabolic state. Polyunsaturated fatty acids (PUFAs) can stabilize energy homeostasis via G protein-coupled receptor 120 (GPR120) to alleviate insulin resistance and inflammation.
Methods: We used lipidomics analysis to reveal a greater accumulation of two PUFAs—arachidonic acid (AA) and docosapentaenoic acid (DPA) in OF than in SF and then hypothesized that AA/DPA is one factor regulating WAT morphological and biological heterogeneity.
Results: Mechanistically, the existing literature evidence suggests that AA/DPA signals may stimulate the co-activation and interaction of GPR120 and peroxisome proliferator-activated receptor γ (PPARγ), at least partially contributing to adipose metabolic health.
Conclusion: As the differential metabolites between OF and SF, AA and DPA, along with the relevant GPR120/PPARγ pathways, offer new therapeutic approaches for morbid obesity.
Keywords: adipose tissue, orbital fat, arachidonic acid, docosapentaenoic acid, GPR120, obesity
Graphical Abstract:

Introduction
Obesity is characterized by the expansion of white adipose tissue (WAT) and has become a challenging issue for public health.1 WAT expands through a combination of two ways: hypertrophy—enlargement in the size of mature adipocytes and hyperplasia—differentiation of pre-adipocytes to produce new small adipocytes.2 Increased adipocyte size is associated with numerous adverse metabolic consequences, such as inflammation, fibrosis, and insulin resistance.3,4 Conversely, the formation of new small adipocytes (adipogenesis) mitigates these metabolic declines and represents a healthy metabolic condition in WAT.5,6 Therefore, exploring factors that shift adipocyte development from hypertrophy to hyperplasia during WAT expansion, thereby storing surplus calories more safely, is crucial for promoting a “metabolically healthy obesity” (MHO) state.7
Orbital fat (OF) is a highly specialized WAT occupying the orbital cavities around the eyeballs. Healthy WAT is characterized by a high degree of vascularization, minimal hypoxia and fibrosis, and low-level inflammation.8 Our previous work, along with other scholars’, has identified that OF possesses smaller adipocytes and is likely to be a healthier adipose tissue than abdominal subcutaneous fat (SF).9–14 Consequently, OF may represent a natural MHO model to study ways to reverse “metabolically morbid obesity” (MMO) by prioritizing adipocyte hyperplasia over hypertrophy.
Studies have demonstrated the beneficial effects of polyunsaturated fatty acids (PUFAs) on obesity-associated metabolic disorders, including anti-inflammation, increasing insulin sensitivity, and healthy adipose tissue remodeling.15,16 As one of the lipid-sensitive G protein-coupled receptors highly expressed in adipose tissue, the G protein-coupled receptor 120 (GPR120), also known as the free fatty acid receptor 4 (FFAR4), plays an essential part in PUFA-regulated adipose metabolic homeostasis.17 In our comparisons of the lipid compositions between OF and SF, we found that the relative amounts of two specific PUFAs, arachidonic acid (AA, C20:4n-6) and docosapentaenoic acid (DPA, C22:5n-3), were unexpectedly higher in OF than in SF. Therefore, in this study, we hypothesized that, in addition to adipose cell-intrinsic heterogeneity (eg, distinct developmental origins and physiological properties), cell content, and cell subtype composition, the differential AA and DPA levels in adipose microenvironment may also influence the morphological differences between OF and SF and regulate adipose metabolism via GPR120.
Methods and Materials
Adipose tissue samples in both groups (n = 6) were, respectively, harvested from different overweight female patients (mean age, 39 years old (range 27–68); mean BMI, 28.3 kg/m2 (range 25–30)) undergoing lower eyelid blepharoplasty or abdominal liposuction in the plastic and reconstructive surgery department of Shanghai Tenth People’s Hospital. The OF samples were specifically obtained from the central orbital fat pads in the lower eyelid, and the SF samples were from the lower abdomen area. We confirmed that there were no significant inter-group differences in basic individual characteristics, including age, sex, ethnicity, BMI, percentage of fat, and waist-to-hip ratio, to ensure the comparability. None of the participants had any ophthalmologic diseases, obesity-related metabolic syndromes, or other severe systemic diseases, and none had a significant weight change within 3 months.
Lipid metabolomics analysis was performed after the digestion and extraction of adipose tissue samples, and the detailed procedures were shown in our previous work.11 Briefly, the total lipid extractions from OF and SF were centrifuged, and the supernatants were collected, dried, and reconstituted in a solvent mixture for the final sample preparation. A volume of 20 μL from each individual sample was pooled as a quality control (QC) sample, aiming to assess and ensure the analytical reproducibility and consistency. Subsequently, with specific setting parameters, an Ultimate 3000 UPLC coupled with a mass spectrometer Q-Exactive Plus (Thermo Fisher Scientific, USA) controlled by Xcalibur software was performed for LC-MS analysis. The raw data was further processed for peak extraction, peak alignment, lipid identification, and peak intensity quantification. Based on the processed metabolomics data, an online metabolomics data analysis tool—MetaboAnalyst (https://www.metaboanalyst.ca) was used to perform the classic univariate receiver operating characteristic (ROC) curve analyses. For statistical analysis, the Shapiro–Wilk test and Levene test were performed to assess the normality and homogeneity of variances, respectively. Comparisons between the two groups were conducted using the independent sample t-test or the Mann–Whitney U-test (as appropriate). The SPSS software (version 26.0; IBM Corp, USA) was used with a statistical significance set at p < 0.05.
Results and Evaluation of the Hypothesis
Importance of Studying the OF Microenvironment to Promote MHO
WAT depot-based morphological and biological differences have been widely proposed, but few have focused on the metabolic distinctions.18,19 Compared with SF, OF may be metabolically healthier, since OF has: (1) morphologically smaller adipocytes,9–11 which are associated with high insulin sensitivity and low-level inflammation;20 (2) more vascularization;10,13 (3) a more dynamic extracellular matrix (ECM) that better supports WAT expansion;10,13,21 (4) less macrophage infiltration (our unpublished data); (5) higher proliferative and adipogenic capacities in adipose-derived stem cells (our unpublished data and reference14).
The adipose microenvironment has been proven to cause regional WAT heterogeneity by both in vitro differentiated cells and in vivo adipose transplantation experiments.22,23 Therefore, we compared the lipid differences between OF and SF, hoping to find key metabolites in the OF microenvironment to improve metabolic disorders.
The Findings of AA/DPA Content Differences Between OF and SF
In this study, more accumulations of AA and DPA in OF than in SF were identified by lipidomics analysis (Figure 1a). ROC curves further demonstrated their discriminatory power between the OF and the SF microenvironments, with the mean area under the ROC curve (AUC) values of 0.833 (95% CI: 0.785–0.881) for AA and 0.750 (95% CI: 0.658–0.842) for DPA, respectively (Figure 1b). Both AA and DPA are common kinds of PUFAs with potential benefits to metabolic homeostasis.24–27 Despite AA’s dual pro- and anti-inflammatory properties in metabolism,26 comprehensive transcriptional profiling during adipogenesis from human mesenchymal stem cells (hMSCs) has already highlighted the critical role of AA in adipogenic differentiation and its potential in improving fat metabolism disorders.28 Therefore, we believe there is a link between differential AA/DPA levels and the morphological and biological distinctions between OF and SF, which is partly responsible for their eventual heterogeneity in metabolic health.
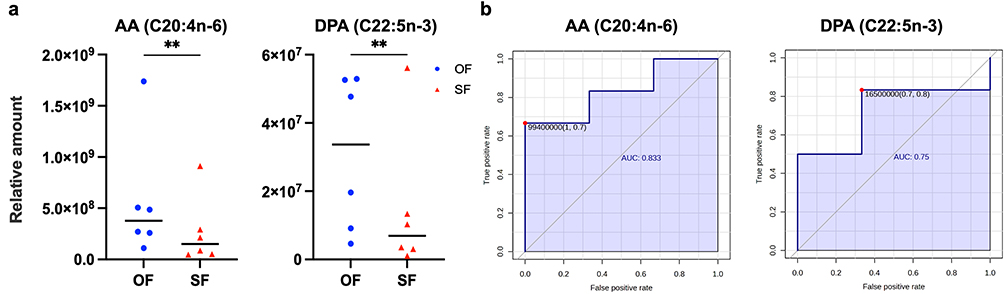 |
Figure 1 The findings of AA/DPA content differences between OF and SF. (a) Comparisons of the relative amounts of AA (C20:4n-6) and DPA (C22:5n-3) between OF and SF. Each spot represented a sample, and the median was shown as a short horizontal line in each group (n = 6, **p < 0.01). (b) Receiver operating characteristic (ROC) curves of AA and DPA. |
AA/DPA May Activate GPR120 to Promote MHO
Theoretically, AA and DPA can activate GPR120 to promote adipose health since these beneficial effects of GPR120 activation has been widely proven using other PUFAs.17,29 In fact, we have found the elevated GPR120 expressions in OF both at the gene level using quantitative real-time polymerase chain reaction (qRT-PCR) and at the protein level utilizing Western blot analysis (the full data is in preparation), which may be attributable to the greater accumulation of AA and DPA in this adipose region. These results, combined with our ongoing adipose GPR120 immunofluorescence staining, are currently being compiled as part of our one unpublished study.
AA/DPA May Regulate GPR120 and PPARγ Co-Activation and Interaction in Pre-Adipocytes
PUFA-mediated GPR120 activation promotes adipogenesis in 3T3-L1 pre-adipocytes.30 On the contrary, inhibiting GPR120 impairs adipogenesis significantly.31,32 PPARγ also can be directly stimulated by fatty acids in pre-adipocytes.33,34 Notably, the size of lipid droplets of PUFA-induced differentiated adipocytes is smaller than that of monounsaturated fatty acid (MUFA)-induced ones, indicating a positive effect of PUFA-mediated PPARγ activation on the adipocyte hypertrophy-hyperplasia balance.33 Metabolites derived from PUFAs also induce PPARγ-related adipogenesis.35,36 Moreover, PPARγ shows a similar pocket structure with GPR120,30 implying the possibility that it could be co-activated by AA/DPA and their metabolites.
The interplay between GPR120 and PPARγ after the co-activation magnifies their positive metabolic effects. GPR120 augments PPARγ-initiated adipogenesis by activating its upstream signal pathways37 and producing more PPARγ ligands.38 Conversely, GPR120 gene knockout inhibits PPARγ expressions.32 On the other hand, PPARγ agonist rosiglitazone increases the expression of GPR120 gene,39 which is subsequently confirmed to be a PPARγ target gene.38 The relevant pathways in pre-adipocytes were speculated to be activated by AA/DPA and were illustrated in Figure 2a.
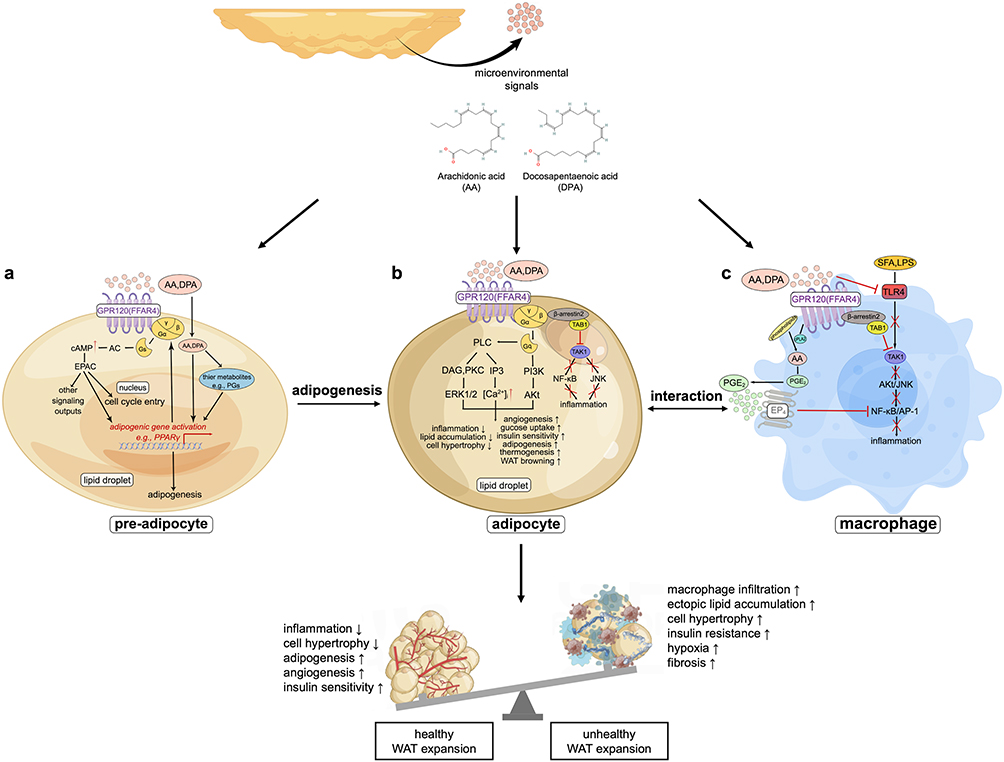 |
Figure 2 Hypothetical overview of the positive biological effects of AA/DPA on promoting metabolic health in WAT through GPR120 (FFAR4) and PPARγ co-activation and interaction. Specifically, as adipose tissue microenvironmental signals, AA/DPA could perhaps act on three kinds of cells: (a) In pre-adipocytes, AA/DPA may stimulate adipogenesis to increase hyperplasia over hypertrophy; (b) In mature adipocytes, AA/DPA may promote healthy cellular biological states; (c) In macrophages, AA/DPA may exert anti-inflammatory effects directly. Abbreviations: AA, arachidonic acid; DPA, docosapentaenoic acid; PGs, prostaglandins; GPR120 (FFAR4), G protein-coupled receptor 120 (free fatty acid receptor 4); AC, adenylate cyclase; cAMP, cyclic adenosine monophosphate; EPAC, exchange factor directly activated by cAMP; PPARγ, peroxisome proliferator-activated receptor γ; PI3K, phosphatidylinositol 3-kinase; Akt, serine/threonine-protein kinase; PLC, phospholipase C; IP3, inositol trisphosphate; DAG, diacylglycerol; PKC, protein kinase C; ERK, extracellular regulated kinase; TAK1, transforming growth factor-β activated kinase 1; TAB1, TAK1 binding protein 1; JNK, c-Jun N-terminal kinase; NF-κB, nuclear factor-kappa B; SFA, saturated fatty acid; LPS, lipopolysaccharide; TLR4, toll-like receptor 4; cPLA2, cytosolic phospholipase A2; PGE2, prostaglandin E2; EP4, prostaglandin receptor 4; AP-1, activator protein 1. |
AA/DPA May Regulate GPR120 Activation in Mature Adipocytes
GPR120 is also highly expressed on the plasma membrane of mature adipocytes and coupled with Gαq to exert various positive biological functions.17,31,40,41 For instance, n-3 PUFAs have anti-inflammatory and insulin-sensitizing effects on wild-type (WT) mice with a high-fat diet (HFD) through GPR120 activation; on the contrary, GPR120-deficient HFD-fed mice show obese phenotypes.17,31,42 PUFAs suppress inflammation through GPR120 by disrupting the bonds between transforming growth factor-β activated kinase 1 (TAK1) and TAK1 binding protein 1 (TAB1) in adipocytes.43 PUFAs also upregulate vascular endothelial growth factor A (VEGF-A) and WAT browning.44,45 Taken together, adipocyte GPR120 activation might be stimulated by AA/DPA to increase glucose uptake, insulin sensitivity, angiogenesis, thermogenesis, and browning in WAT, thereby inhibiting cell hypertrophy and inflammation (Figure 2b).
AA/DPA May Regulate GPR120 Activation in Macrophages
AA/DPA may also mediate GPR120 signals to exert anti-inflammatory effects directly in macrophages (Figure 2c). Through the GPR120 activation, PUFAs: (1) inhibit TAK1/TAB1 interaction to block the downstream IKKβ/NF-κB and JNK/AP pathways;17 (2) couple with β-arrestin2 to repress TLR4-induced inflammatory signaling;46 (3) activate cytosolic phospholipase A2 (cPLA2) to release prostaglandin E2 (PGE2), which subsequently stimulates prostaglandin E receptor 4 (EP4) to inhibit the NF-κB inflammatory pathway.47 Additionally, there is an interplay between GPR120-regulated macrophages and adipocytes: Inflammatory cytokines from macrophages inhibit GPR120 expression in adipocytes,48 and, in turn, metabolically disordered adipocytes induce pro-inflammatory macrophage phenotypes.49
Discussion
The WAT depot-based heterogeneity provides a comparison model for exploring ways to treat obesity-related metabolic dysfunctions. Numerous studies have compared SF and visceral fat (VF), showing that SF is metabolically healthier than VF, as accumulated VF is more closely associated with insulin resistance and lipotoxicity.50 However, anatomically different SF depots also respond to weight gain differently and are associated with distinct metabolic statuses.51,52 Abdominal SF mainly relies on fat cell hypertrophy to expand and demonstrates significant inflammatory insults, while lower-body (eg, gluteofemoral) SF seems to retain its biological functions and capacity to recruit new, small adipocytes and therefore is regarded as a metabolically protective WAT.51,52 Compared with extensive research on SF and VF, OF is often overlooked and has never been utilized as a model for novel obesity treatment targets. Our OF-centric study innovatively compared OF with SF and proposed OF possesses smaller adipocytes and a relatively healthier metabolic state. Further, we identified higher levels of AA and DPA in OF and hypothesized their important roles in regulating adipose health.
Almost all existing studies on the positive metabolic effects of the PUFA-mediated GPR120 activation focus on n-3 PUFAs.17,30 DPA, despite being an n-3 PUFA, has been reported to improve inflammation, arteriosclerosis, and other obesity-related metabolic dysfunctions, but no research has linked these effects to GPR120.25,27 As for AA, an n-6 PUFA, it plays a controversial and complicated role in obesity, as it is a precursor to multiple pro-inflammatory mediators, while its derivatives exert anti-inflammatory effects.24,26,53 However, as for now, no one has studied the impacts of AA/DPA-GPR120 signaling, not to mention in the context of adipose metabolism. We propose for the first time that the AA/DPA-mediated co-activation of GPR120 and PPARγ, as well as their interplay, at least partly contributes to OF’s metabolic profile, expanding the clinical applications of AA/DPA in metabolic diseases. Furthermore, based on our OF-SF comparison model to hypothesize the different GPR120 activation statuses caused by different adipose AA/DPA levels, we reinforce the feasibility and prospects of GPR120-related treatments in obesity (particularly through increasing GPR120 expression and its sensitivity). Nevertheless, the AA/DPA-GPR120/PPARγ axis we discuss here only offers a preliminary and partial explanation for the metabolic heterogeneity between OF and SF, as the effects of the axis itself need to be experimentally validated and numerous other undiscovered factors in OF also influence adipose health.54
To fully clarify our hypothesis that AA/DPA mediates WAT depot-specific distinctions in adipocyte hypertrophy–hyperplasia balance and metabolic health, we plan to: (1) Perform a target lipid metabolomics analysis to specify the differential AA/DPA metabolism patterns in OF and SF. (2) Study distinct transcriptomics profiles in OF and SF to explore the underlying mechanisms for the differential AA/DPA levels. (3) Observe whether AA/DPA eliminates the depot-based heterogeneity in vitro adipose stem and progenitor cells (ASPCs), like the tumor necrosis factor (TNF) stimulation reducing differences between visceral and subcutaneous ASPCs in a previous study.23 Concurrent with this, we will observe positive biological effects induced by AA/DPA in ASPCs, such as improved adipogenic and angiogenic capacities, insulin sensitivity, inflammatory level, etc. (4) Validate the anti-obesity effects of AA/DPA and the related GPR120/PPARγ signaling activation in an animal model. (5) Employ GPR120 antagonists and/or GPR120 gene-knockout experiments to prove that AA/DPA confers these biological benefits via the GPR120/PPARγ-related pathways. In addition, we could compare OF with other adipose sites to provide a more comprehensive view of adipose depot-specific heterogeneity in morphology and metabolic health.
Conclusion
This study introduces OF as a depot-specific adipose model with small adipocytes and a relatively healthy metabolic profile. This OF-based healthy adipose model offers a more valid and ideal way to study healthy WAT expansion by eliminating the intrinsic biological variables of individuals, such as genes, dietary structures, and exercise habits. Based on our OF observation model, we hypothesized that higher AA and DPA levels in adipose microenvironments may improve adipose inflammation and metabolism via GPR120/PPARγ. AA/DPA could perhaps serve as biomarkers for assessing adipose metabolic health, and new AA/DPA-related nutritional or pharmaceutical interventions may benefit clinical obesity. Altogether, AA/DPA and their targets—GPR120/PPARγ—inspire us with new therapeutic targets for improving metabolic profiles in obesity.
Data Sharing Statement
The data in this study are available upon request from the corresponding author.
Informed Consent and Institutional Review Board Statement
All participants provided written informed consent to participate in this study and to publish their case details. Experimental methods involving patients followed the tenets of the Declaration of Helsinki and were approved by the Medical Ethics Committee of Shanghai Tenth People’s Hospital (approval number: SHSY-LYZX-223).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agreed to be accountable for all aspects of the work.
Funding
This work was supported by grants from the National Natural Science Foundation of China (grant number: 31870974).
Disclosure
All authors declare that no known competing interests influence the work reported in this paper.
References
1. Goossens GH. The metabolic phenotype in obesity: fat mass, body fat distribution, and adipose tissue function. Obes Facts. 2017;10(3):207–215. doi:10.1159/000471488
2. Ghaben AL, Scherer PE. Adipogenesis and metabolic health. Nat Rev Mol Cell Biol. 2019;20(4):242–258. doi:10.1038/s41580-018-0093-z
3. Salans LB, Knittle JL, Hirsch J. The role of adipose cell size and adipose tissue insulin sensitivity in the carbohydrate intolerance of human obesity. J Clin Invest. 1968;47(1):153–165. doi:10.1172/JCI105705
4. Krotkiewski M, Björntorp P, Sjöström L, Smith U. Impact of obesity on metabolism in men and women. Importance of regional adipose tissue distribution. J Clin Invest. 1983;72(3):1150–1162. doi:10.1172/JCI111040
5. Yang J, Eliasson B, Smith U, Cushman SW, Sherman AS. The size of large adipose cells is a predictor of insulin resistance in first-degree relatives of type 2 diabetic patients. Obes Silver Spring Md. 2012;20(5):932–938. doi:10.1038/oby.2011.371
6. Lönn M, Mehlig K, Bengtsson C, Lissner L. Adipocyte size predicts incidence of type 2 diabetes in women. FASEB J off Publ Fed Am Soc Exp Biol. 2010;24(1):326–331. doi:10.1096/fj.09-133058
7. Denis GV, Obin MS. “Metabolically healthy obesity”: origins and implications. Mol Aspects Med. 2013;34(1):59–70. doi:10.1016/j.mam.2012.10.004
8. Scherer PE. The many secret lives of adipocytes: implications for diabetes. Diabetologia. 2019;62(2):223–232. doi:10.1007/s00125-018-4777-x
9. Bujalska IJ, Durrani OM, Abbott J, et al. Characterisation of 11β-hydroxysteroid dehydrogenase 1 in human orbital adipose tissue: a comparison with subcutaneous and omental fat. J Endocrinol. 2007;192(2):279–288. doi:10.1677/JOE-06-0042
10. Afanas’eva DS, Gushchina MB, Borzenok SA. Comparison of morphology of adipose body of the orbit and subcutaneous fat in humans. Bull Exp Biol Med. 2018;164(3):394–396. doi:10.1007/s10517-018-3997-x
11. Zhang K, Zhou Y, Xie Z, Liu G. Subcutaneous and orbital adipose tissues: intrinsic differences in carotenoid contents and lipidomic compositions. ACS Omega. 2023;8(31):28052–28059. doi:10.1021/acsomega.2c08013
12. Arner E, Westermark PO, Spalding KL, et al. Adipocyte turnover: relevance to human adipose tissue morphology. Diabetes. 2010;59(1):105–109. doi:10.2337/db09-0942
13. Ilankovan V, Soames JV. Morphometric analysis of orbital, buccal and subcutaneous fats: their potential in the treatment of enophthalmos. Br J Oral Maxillofac Surg. 1995;33(1):40–42. doi:10.1016/0266-4356(95)90085-3
14. Nepali S, Park M, Lew H, Kim O. Comparative analysis of human adipose-derived mesenchymal stem cells from orbital and abdominal fat. Stem Cells Int. 2018;2018:3932615. doi:10.1155/2018/3932615
15. Shahidi F, Ambigaipalan P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu Rev Food Sci Technol. 2018;9:345–381. doi:10.1146/annurev-food-111317-095850
16. Fisk HL, Childs CE, Miles EA, et al. Dysregulation of subcutaneous white adipose tissue inflammatory environment modelling in non-insulin resistant obesity and responses to omega-3 fatty acids – a double blind, randomised clinical trial. Front Immunol. 2022;13:922654. doi:10.3389/fimmu.2022.922654
17. Oh DY, Talukdar S, Bae EJ, et al. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell. 2010;142(5):687–698. doi:10.1016/j.cell.2010.07.041
18. Karastergiou K, Fried SK, Xie H, et al. Distinct developmental signatures of human abdominal and gluteal subcutaneous adipose tissue depots. J Clin Endocrinol Metab. 2013;98(1):362–371. doi:10.1210/jc.2012-2953
19. Macotela Y, Emanuelli B, Mori MA, et al. Intrinsic differences in adipocyte precursor cells from different white fat depots. Diabetes. 2012;61(7):1691–1699. doi:10.2337/db11-1753
20. Klöting N, Fasshauer M, Dietrich A, et al. Insulin-sensitive obesity. Am J Physiol Endocrinol Metab. 2010;299(3):E506–515. doi:10.1152/ajpendo.00586.2009
21. Soták M, Rajan MR, Clark M, et al. Healthy subcutaneous and omental adipose tissue is associated with high expression of extracellular matrix components. Int J Mol Sci. 2022;23(1):520. doi:10.3390/ijms23010520
22. Jeffery E, Wing A, Holtrup B, et al. The Adipose Tissue Microenvironment Regulates Depot-Specific Adipogenesis in Obesity. Cell Metab. 2016;24(1):142–150. doi:10.1016/j.cmet.2016.05.012
23. Mathur N, Severinsen MCK, Jensen ME, et al. Human visceral and subcutaneous adipose stem and progenitor cells retain depot-specific adipogenic properties during obesity. Front Cell Dev Biol. 2022;10:983899. doi:10.3389/fcell.2022.983899
24. Das UN. Syntaxin interacts with arachidonic acid to prevent diabetes mellitus. Lipids Health Dis. 2022;21(1):73. doi:10.1186/s12944-022-01681-3
25. Rissanen T, Voutilainen S, Nyyssönen K, Lakka TA, Salonen JT. Fish oil-derived fatty acids, docosahexaenoic acid and docosapentaenoic acid, and the risk of acute coronary events: the Kuopio ischaemic heart disease risk factor study. Circulation. 2000;102(22):2677–2679. doi:10.1161/01.cir.102.22.2677
26. Tortosa-Caparrós E, Navas-Carrillo D, Marín F, Orenes-Piñero E. Anti-inflammatory effects of omega 3 and omega 6 polyunsaturated fatty acids in cardiovascular disease and metabolic syndrome. Crit Rev Food Sci Nutr. 2017;57(16):3421–3429. doi:10.1080/10408398.2015.1126549
27. Hirako S, Hirabayashi T, Shibato J, et al. Docosapentaenoic acid-rich oil lowers plasma glucose and lipids in a mouse model of diabetes and mild obesity. Nutr Res. 2023;118:128–136. doi:10.1016/j.nutres.2023.08.004
28. Yi X, Liu J, Wu P, Gong Y, Xu X, Li W. The whole transcriptional profiling of cellular metabolism during adipogenesis from hMSCs. J Cell Physiol. 2020;235(1):349–363. doi:10.1002/jcp.28974
29. Mao C, Xiao P, Tao XN, et al. Unsaturated bond recognition leads to biased signal in a fatty acid receptor. Science. 2023;380(6640):eadd6220. doi:10.1126/science.add6220
30. Song T, Yang Y, Zhou Y, Wei H, Peng J. GPR120: a critical role in adipogenesis, inflammation, and energy metabolism in adipose tissue. Cell Mol Life Sci. 2017;74(15):2723–2733. doi:10.1007/s00018-017-2492-2
31. Ichimura A, Hirasawa A, Poulain-Godefroy O, et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nature. 2012;483(7389):350–354. doi:10.1038/nature10798
32. Gotoh C, Hong YH, Iga T, et al. The regulation of adipogenesis through GPR120. Biochem Biophys Res Commun. 2007;354(2):591–597. doi:10.1016/j.bbrc.2007.01.028
33. Madsen L, Petersen RK, Kristiansen K. Regulation of adipocyte differentiation and function by polyunsaturated fatty acids. Biochim Biophys Acta BBA - Mol Basis Dis. 2005;1740(2):266–286. doi:10.1016/j.bbadis.2005.03.001
34. Hihi AK, Michalik L, Wahli W. PPARs: transcriptional effectors of fatty acids and their derivatives. Cell Mol Life Sci CMLS. 2002;59(5):790–798. doi:10.1007/s00018-002-8467-x
35. Forman BM, Tontonoz P, Chen J, Brun RP, Spiegelman BM, Evans RM. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell. 1995;83(5):803–812. doi:10.1016/0092-8674(95)90193-0
36. Fujimori K. Prostaglandin D2 and F2α as regulators of adipogenesis and obesity. Biol Pharm Bull. 2022;45(8):985–991. doi:10.1248/bpb.b22-00210
37. Song T, Zhou Y, Peng J, et al. GPR120 promotes adipogenesis through intracellular calcium and extracellular signal-regulated kinase 1/2 signal pathway. Mol Cell Endocrinol. 2016;434:1–13. doi:10.1016/j.mce.2016.06.009
38. Paschoal VA, Walenta E, Talukdar S, et al. Positive reinforcing mechanisms between GPR120 and PPARγ modulate insulin sensitivity. Cell Metab. 2020;31(6):1173–1188.e5. doi:10.1016/j.cmet.2020.04.020
39. Muredda L, Kępczyńska MA, Zaibi MS, Alomar SY, Trayhurn P. IL-1β and TNFα inhibit GPR120 (FFAR4) and stimulate GPR84 (EX33) and GPR41 (FFAR3) fatty acid receptor expression in human adipocytes: implications for the anti-inflammatory action of n-3 fatty acids. Arch Physiol Biochem. 2018;124(2):97–108. doi:10.1080/13813455.2017.1364774
40. Bjursell M, Admyre T, Göransson M, et al. Improved glucose control and reduced body fat mass in free fatty acid receptor 2-deficient mice fed a high-fat diet. Am J Physiol Endocrinol Metab. 2011;300(1):E211–220. doi:10.1152/ajpendo.00229.2010
41. Kimura I, Ichimura A, Ohue-Kitano R, Igarashi M. Free fatty acid receptors in health and disease. Physiol Rev. 2020;100(1):171–210. doi:10.1152/physrev.00041.2018
42. Im DS. FFA4 (GPR120) as a fatty acid sensor involved in appetite control, insulin sensitivity and inflammation regulation. Mol Aspects Med. 2018;64:92–108. doi:10.1016/j.mam.2017.09.001
43. Yamada H, Umemoto T, Kakei M, et al. Eicosapentaenoic acid shows anti-inflammatory effect via GPR120 in 3T3-L1 adipocytes and attenuates adipose tissue inflammation in diet-induced obese mice. Nutr Metab. 2017;14:33. doi:10.1186/s12986-017-0188-0
44. Yang L, Lei XT, Huang Q, Wang T, Sun HB, Wang HY. A novel GPR120-selective agonist promotes insulin secretion and improves chronic inflammation. Life Sci. 2021;269:119029. doi:10.1016/j.lfs.2021.119029
45. Quesada-López T, Cereijo R, Turatsinze JV, et al. The lipid sensor GPR120 promotes brown fat activation and FGF21 release from adipocytes. Nat Commun. 2016;7:13479. doi:10.1038/ncomms13479
46. Li X, Yu Y, Funk CD. Cyclooxygenase‐2 induction in macrophages is modulated by docosahexaenoic acid via interactions with free fatty acid receptor 4 (FFA4). FASEB J. 2013;27(12):4987–4997. doi:10.1096/fj.13-235333
47. Liu Y, Chen LY, Sokolowska M, et al. The fish oil ingredient, docosahexaenoic acid, activates cytosolic phospholipase A2 via GPR120 receptor to produce prostaglandin E2 and plays an anti-inflammatory role in macrophages. Immunology. 2014;143(1):81–95. doi:10.1111/imm.12296
48. Trayhurn P, Denyer G. Mining microarray datasets in nutrition: expression of the GPR120 (n −3 fatty acid receptor/sensor) gene is down-regulated in human adipocytes by macrophage secretions. J Nutr Sci. 2012;1:e3. doi:10.1017/jns.2012.3
49. Rodriguez-Pacheco F, Gutierrez-Repiso C, Garcia-Serrano S, et al. The pro-/anti-inflammatory effects of different fatty acids on visceral adipocytes are partially mediated by GPR120. Eur J Nutr. 2017;56(4):1743–1752. doi:10.1007/s00394-016-1222-0
50. Ibrahim MM. Subcutaneous and visceral adipose tissue: structural and functional differences. Obes Rev off J Int Assoc Study Obes. 2010;11(1):11–18. doi:10.1111/j.1467-789X.2009.00623.x
51. Karpe F, Pinnick KE. Biology of upper-body and lower-body adipose tissue—link to whole-body phenotypes. Nat Rev Endocrinol. 2015;11(2):90–100. doi:10.1038/nrendo.2014.185
52. Piché ME, Poirier P, Lemieux I, Després JP. Overview of epidemiology and contribution of obesity and body fat distribution to cardiovascular disease: an update. Prog Cardiovasc Dis. 2018;61(2):103–113. doi:10.1016/j.pcad.2018.06.004
53. Zhao Y, Chen Y, Xiao Q, Li W. Arachidonic acid alleviates autoimmune diabetes in NOD mice. Int Immunopharmacol. 2024;127:111340. doi:10.1016/j.intimp.2023.111340
54. Chouchani ET, Kajimura S. Metabolic adaptation and maladaptation in adipose tissue. Nat Metab. 2019;1(2):189–200. doi:10.1038/s42255-018-0021-8
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Elevated CTSL Gene Expression Correlated with Proinflammatory Cytokines in Omental Adipose Tissue of Patients with Obesity
Zhou Q, Zhu Y, Li C, Li Z, Tang Z, Yuan B, Wang X, Zhang S, Wu X
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2277-2285
Published Date: 30 July 2022
Effects of NKT Cells on Metabolic Disorders Caused by High-Fat Diet Using CD1d-Knockout Mice

Ishikawa H, Nagashima R, Kuno Y, Sasaki H, Kohda C, Iyoda M
Diabetes, Metabolic Syndrome and Obesity 2023, 16:2855-2864
Published Date: 19 September 2023
Bariatric Surgery and Gut-Brain-Axis Driven Alterations in Cognition and Inflammation
Custers E, Franco A, Kiliaan AJ
Journal of Inflammation Research 2023, 16:5495-5514
Published Date: 22 November 2023
Body Weight-Related Differences in Adipokines and Inflammatory Markers Among Women with Systemic Lupus Erythematosus
Carvalho LM, Da Mota JC, Ribeiro AA, Souza LL, Sales RCDS, Gualano B, Martínez-Urbistondo M, Martínez JA, Borba EF, Nicoletti CF
Journal of Inflammation Research 2026, 19:579501
Published Date: 28 February 2026