Back to Journals » Drug Design, Development and Therapy » Volume 20
Obacunone Promotes Functional Recovery After Spinal Cord Injury by Attenuating Neuroinflammation by Targeting the TLR4/MyD88/p38 MAPK Pathway
Authors Kuang W, Zhang M, Zhang J, Huang H, Chen Q, Yu C, Peng B, Sun W, Lin J, Cheng J, Zhu L
Received 15 December 2025
Accepted for publication 25 February 2026
Published 13 March 2026 Volume 2026:20 577707
DOI https://doi.org/10.2147/DDDT.S577707
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leonidas D. Panos
Wenhao Kuang,1,* Mi Zhang,1,* Jiaqi Zhang,1,* Haoran Huang,1 Qifan Chen,2 Cheng Yu,3 Birong Peng,1 Wei Sun,1 Jiezhao Lin,4 Junjie Cheng,1 Lixin Zhu1
1Department of Spinal Surgery, Zhujiang Hospital, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China; 2Department of Spinal Surgery, The First Affiliated Hospital of Guangxi Medical University, Nanning, Guangxi, People’s Republic of China; 3Department of Orthopedics, The Third People’s Hospital of Chengdu, Chengdu, Sichuan, People’s Republic of China; 4Department of Spinal Surgery, Shantou Central Hospital, Shantou, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lixin Zhu, Department of Spinal Surgery, Zhujiang Hospital, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China, Email [email protected]
Purpose: Spinal cord injury (SCI) triggers a complex secondary injury process, among which inflammation and apoptosis are the key factors of nerve injury. Obacunone (Oba) is a natural limonoid that has demonstrated a variety of pharmacological effects, but its role in SCI remains unclear.
Methods: Network pharmacology and bioinformatics analysis were employed to predict the functions and mechanisms of Oba in SCI. Subsequently, in vivo validation was conducted in a mouse SCI model, with motor function recovery assessed using open field, footprint, and swimming tests. Spinal cord histomorphology was examined via hematoxylin and eosin (HE) staining and Nissl staining, while the anti‑inflammatory and anti‑apoptotic effects were evaluated by Western blot and immunofluorescence. To further elucidate the underlying mechanisms, a lipopolysaccharide (LPS)-induced inflammatory model in BV‑2 microglial cells was established to study the anti‑inflammatory mechanisms of Oba. Furthermore, a BV‑2/HT22 neuronal co‑culture system was constructed to investigate neuroprotective effects of Oba against apoptosis.
Results: In vivo, Oba treatment improved motor function, promoted neural repair, reduced inflammation and apoptosis. Correspondingly, Oba suppressed the expression of LPS-induced pro-inflammatory cytokines in BV-2 cells. In a microglia-neuron co-culture system, Oba protected HT22 neurons from microglia-mediated inflammatory apoptosis. Mechanistically, the anti‑inflammatory effects of Oba were mediated by inhibiting the activation of the TLR4/MyD88/p38 MAPK pathway.
Conclusion: This study identifies Oba as an effective compound that mitigates secondary injury by reducing inflammation and apoptosis and promotes nerve repair and functional recovery post-SCI, supporting its potential for further therapeutic development.
Keywords: obacunone, inflammation, apoptosis, TLR4/MyD88/p38 MAPK, spinal cord injury
Introduction
Spinal cord injury (SCI) is a severe disabling disorder of the central nervous system (CNS), often resulting in significant sensory, motor, and autonomic dysfunction. Most cases are caused by mechanical trauma.1 The pathological process of SCI is highly complex and consists of two main phases: the primary injury, which involves direct mechanical damage to spinal cord tissue, frequently leading to neuronal and axonal demyelination and necrosis; and the secondary injury, which encompasses a range of pathophysiological mechanisms such as inflammation, apoptosis, hemorrhage, ischemia, and edema.2 Secondary injury exacerbates damage to neural tissue and contributes to worse functional outcomes. Suppressing neuroinflammation is a key therapeutic strategy for improving prognosis.3 Microglia, as resident immune cells in the CNS, play a crucial role in maintaining neural homeostasis. Following SCI, activated microglia predominantly adopt a pro-inflammatory phenotype, which aggravates neuroinflammatory pathology and promotes neuronal apoptosis.4
Microglia express all natural immune signaling receptors, including cytokine receptors, complement receptors, chemokine receptors, purinergic receptors, pattern recognition receptors like TLR.5 Among TLRs, Toll-like receptor 4 (TLR4) plays a pivotal role in inflammation.6 Its activation initiates the MyD88-dependent pathway by recruiting the adaptor protein MyD88, subsequently triggering downstream effectors like MAPK and NF-κB pathways that propel inflammatory responses. Therefore, a promising therapeutic strategy for SCI involves targeting the TLR4/MyD88 axis to mitigate neuroinflammation and neuroapoptosis.7
Given the pathophysiological mechanisms of spinal cord injury, natural compounds derived from plants have emerged as a promising therapeutic strategy.8 Among these, monomers isolated from traditional Chinese medicine have recently garnered significant research attention, particularly for SCI treatment, owing to their documented low toxicity and reduced side effects.9 Obacunone (Oba), a limonoid compound abundant in various medicinal plants, exhibits a broad spectrum of pharmacological properties including anti-inflammatory, antioxidant, anti-cancer, and anti-fibrotic activities.10 Oba has been shown to alleviate inflammatory neuropathic pain through suppression of inflammatory responses and oxidative stress.11 Additionally, Oba protects HT22 cells from glutamate-induced toxicity by reducing reactive oxygen species (ROS) via upregulation of heme oxygenase-1 (HO-1) expression.12 Oba alleviates thalamic pain in mice through promoting LCN2-mediated astrocytic phagocytosis.13 Collectively, these findings underscore the multifaceted neuroprotective properties of Oba, highlighting its potential as a therapeutic agent for various neurological disorders. Nevertheless, the specific role of Oba in SCI remains largely unexplored. And its anti‑inflammatory effects and the related mechanisms in microglia are still unclear. These questions warrant further investigation.
This study comprehensively investigates the pharmacological role of the natural compound Oba in SCI through an integrated approach combining network pharmacology, bioinformatics, and experimental validation both in vivo and in vitro. This study suggest that Oba alleviates neuroinflammation by inhibiting the TLR4/MyD88/p38 MAPK signaling pathway, thereby reducing apoptosis and promoting functional recovery after SCI. These findings provide substantive evidence for the development of novel mechanism-based therapies for SCI.
Materials and Methods
Network Pharmacology
The CAS number and SMILES notation of Obacunone were retrieved from the PubChem database using “Obacunone” as the search term. These molecular descriptors were then submitted to the Swiss Target Prediction platform to forecast potential therapeutic targets. SCI-relevant targets were identified by systematically querying the GeneCards and OMIM databases using the key term “spinal cord injury”, after which the collected targets were integrated and duplicates were removed. Intersection analysis between the compound-target and disease-target datasets was conducted using R software (version 4.2.1) with the packages VennDiagram (v1.7.3) and ggplot2 (v3.4.4) to identify overlapping genes.
GO and KEGG Pathway Enrichment Analysis
The overlapping genes were subsequently annotated through functional enrichment analyses based on Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG), performed with the “clusterProfiler” package in R. The results were visualized using the “ggplot2” package.
Protein-Protein Interaction Network
A protein-protein interaction (PPI) network was constructed using the STRING database, incorporating genes with an interaction score threshold set above 0.4 to ensure high-confidence interactions among 96 overlapping genes.
Animals and SCI Model
Eight-week-old female C57BL/6J mice (body weight 22–25 g) were obtained from the Experimental Animal Center of Southern Medical University. The animals were maintained under standard housing conditions with ambient temperature of 22 ± 2 °C, relative humidity of 50–60%, and a 12 h/12 h light/dark cycle. Food and water were provided ad libitum. All experimental procedures were approved by the Ethics Committee of Zhujiang Hospital, Southern Medical University (Approval No. LAEC-2024-290). All animal procedures were conducted in compliance with the Guide for the Care and Use of Laboratory Animals.
A spinal cord compression injury model was established in this study, as previously described.14 Briefly, mice were anesthetized via intraperitoneal injection of 1.25% tribromoethanol (Sigma, USA) at a dose of 20 mL/kg. Following disinfection, the spinal cord was exposed at the T9 vertebral level and compressed using an arterial clip (30 g force, Kent Scientific Corporation, USA) for 10 seconds. Successful induction of injury was confirmed by immediate observations of spinal cord congestion, tail flick reflex, and hind limb rigidity. Postoperatively, Massage the bladder three times a day after the operation to assist urination.
Obacunone (≥98% purity, cat. no. X03975; ACMEC, Shanghai, China) was dissolved in dimethyl sulfoxide (DMSO) and then diluted with a vehicle consisting of 40% PEG300, 5% Tween 80, and 53% normal saline, resulting in a final DMSO concentration of 2%. The starting dose of 10 mg/kg and the intraperitoneal route of administration were chosen based on existing literature reports.11 The experimental timeline employed four separate cohorts of animals. The first cohort was randomly assigned to four groups (n=6 per group): Sham, SCI, Oba 10 (SCI+10 mg/kg Obacunone), and Oba 20 (SCI+20 mg/kg Obacunone). Treatment was administered once daily for 7 consecutive days, beginning immediately after spinal cord injury. To evaluate functional recovery, this cohort underwent behavioral testing on days 7, 14, 21, and 28 post‑SCI, and was euthanized for histopathological analysis on day 28. To assess acute inflammatory and apoptotic responses, spinal cord tissues were collected from two additional cohorts of animals at 3 days post-SCI. Tissues from Cohort 2 (groups: Sham, SCI, and SCI+Oba [20 mg/kg]; n=5 per group) were used for Western blot analysis of inflammatory and apoptotic markers. Tissues from Cohort 3 (groups: Sham, SCI, and SCI+Oba [20 mg/kg]; n=5 per group) were processed for immunofluorescence staining and for TUNEL staining. Tissues from Cohort 4 (groups: Sham, SCI, and SCI+Oba [20 mg/kg]; n=5 per group) were used for Western blot analysis of axonal regeneration marker on day 28. All euthanasia was conducted via CO2 inhalation.
Behavioral Assessments
All behavioral assessments were performed by two investigators who were blinded to the group assignments. These assessments were conducted on post-injury days 7, 14, 21, and 28.
The open-field test was employed to assess locomotor function, alongside the Basso Mouse Scale (BMS), a validated tool for evaluating motor recovery after SCI.15 At designated time points post-injury, mice were placed in an open field for a 1-minute observation period, during which BMS scores were assigned based on spontaneous movement.
Additionally, a swimming test was conducted to further evaluate motor performance. Mice were acclimated to swim in a glass tank filled with water and were rated according to an established swimming scale.16
Footprint analysis is used to detect changes in the stride length of mice. The forelimbs and hindlimbs of the mice were marked with red and blue non-toxic ink, respectively. Each mouse was then allowed to traverse a narrow runway (10 cm wide × 1 m long) lined with white paper, allowing for the subsequent analysis of footprint sequences.
Histological Staining
Following anesthesia, the mice underwent transcardial perfusion with normal saline followed by 4% paraformaldehyde. A 10-mm segment of the spinal cord centered on the lesion site was dissected and post-fixed in 4% paraformaldehyde for 24 hours. The tissue was then dehydrated through a graded series of sucrose solutions (10%, 20%, and 30%). Subsequently, frozen sections were prepared at a thickness of 10 μm and subjected to hematoxylin and eosin (HE) staining as well as Nissl staining.
The HE staining procedure began with a 3-minute incubation in hematoxylin, followed by a tap water rinse, brief differentiation in acid-alcohol for 3 seconds, and counterstaining with eosin for 30 seconds. After a final 3-minute rinse, the sections were dehydrated through a graded alcohol series, cleared in xylene, and mounted with neutral gum.
Nissl staining was performed using a commercial kit (G1430, Solarbio, China) according to the manufacturer’s instructions. The sections were immersed in the staining solution and incubated at 56 °C for 30 minutes. After being rinsed with distilled water to remove excess dye, they were differentiated in the Nissl differentiation solution for 10 seconds. Finally, the sections were dehydrated through a graded series of alcohols, cleared in xylene, and mounted with neutral resin. Finally, digital images of the stained sections were acquired using a digital slide scanner (3D HISTECH).
Immunofluorescent Staining
Tissue sections were fixed in 4% paraformaldehyde for 20 minutes at room temperature. After three washes with PBST, the sections were permeabilized with Triton X-100 (P0096, Beyotime, China) for 20 minutes. To minimize non-specific binding, the sections were blocked with QuickBlock™ blocking buffer (P0260, Beyotime, China) for 60 minutes at room temperature. Following a wash with PBST, the sections were incubated with primary antibodies overnight at 4°C. The next day, unbound primary antibodies were removed by washing with PBST, and the sections were then incubated with appropriate secondary antibody conjugates for 60 minutes at room temperature. Finally, the stained sections were coverslipped using an anti-fade mounting medium containing DAPI. The primary antibodies included rabbit anti-iNOS (13120, Cell signaling technology, 1: 200). The second antibodies included HyperFluor™ 488 Goat Anti- Rabbit IgG (H+L) Antibody (K1206, Apexbio, 1:500).
Western Blot
Tissue and cell samples from all experimental groups were lysed on ice using RIPA buffer (P0013B, Beyotime, China) supplemented with phosphatase and protease inhibitors (P003, P001, NCM Biotech, China) for 30 minutes. The lysates were centrifuged at 12,000 rpm for 15 minutes, and the resulting supernatants were mixed with 5× loading buffer. Protein denaturation was performed by heating at 100 °C for 10 minutes. Proteins were separated by SDS-PAGE and subsequently transferred to PVDF membranes. The membranes were blocked for 90 minutes at room temperature and then incubated with primary antibodies overnight at 4 °C. After washing with TBST, the membranes were incubated with HRP-conjugated secondary antibodies for 60 minutes at room temperature. Protein bands were visualized using an ECL detection system (P10100, NCM Biotech, China). The primary antibodies included rabbit anti-iNOS (13120, Cell signaling technology, 1: 1000), rabbit anti-TNFα (IPB9396, Baijia, 1:1000), rabbit anti-IL-1β (IPB0002, Baijia, 1: 1000), rabbit anti-GAP43 (16971-1-AP, Proteintech, 1:1000), mouse anti- α-tubulin (11224-1-AP, Proteintech, 1:1000), mouse anti-GAPDH (ab307799, Abcam, 1:5000), rabbit anti-Cleaved-caspase3 (AF7022, Affinity, 1: 1000), rabbit anti-Bax (505992-Ig, Proteintech, 1: 1000), rabbit anti-TLR4 (A17436, ABclonal, 1: 1000), rabbit anti-p38 MAPK (T55600, ABmart, 1: 1000), rabbit anti-Phospho-p38 MAPK (Thr180/Tyr182) (TA4001, ABmart, 1: 1000), rabbit anti-MyD88 (YM8747, immunoway, 1: 1000). The second antibodies included Goat Anti-Rabbit IgG (H+L) HRP (BS13278, Bioworld, 1: 5000) and Goat Anti-Mouse IgG (H+L) HRP (BS20242-Y, Bioworld, 1: 5000).
Tunel Staining
Spinal cord frozen sections were fixed with 4% paraformaldehyde for 20 minutes, followed by three washes in PBST. The sections were then permeabilized with Triton X-100 for 20 minutes at room temperature. The sections were then incubated with TUNEL staining solution (C1089, Beyotime, China) for 1 hour at 37 °C in the dark. After staining, the sections were coverslipped with an anti-fade mounting medium containing DAPI. Images were acquired using a Nikon confocal microscope.
Cell Culture
BV-2 microglial cells (CL-0493, Procell, Wuhan, China) and HT22 neuronal cells (CL-0697, Procell, Wuhan, China) were used in this study. Both cell lines were cultured in high-glucose Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum and maintained at 37 °C in a humidified atmosphere containing 5% CO2.
To establish an inflammatory model, BV-2 cells were pretreated with vehicle (DMSO) or 25 μM Oba or 50 μM Oba for 3 hours, followed by stimulation with lipopolysaccharide (LPS, 1 μg/mL) (Sigma-Aldrich, USA) for 24 hours.
To investigate the interaction between microglia and neurons, BV-2 and HT22 cells were co-cultured via a transwell system (Corning, USA). BV-2 cells were pretreated with LPS in the presence or absence of 50 μM Oba as mentioned previously. The pretreated BV-2 cells were then seeded into the upper chamber and co-cultured with HT22 cells seeded in the lower chamber for 24 hours. Subsequently, HT22 cells were harvested for further analysis.
To investigate the role of the p38 MAPK pathway in the anti-inflammatory effects of Oba, BV-2 cells were pre-treated for 3 hours with 50 μM Oba, 10 μM SB20358017 (a p38 MAPK inhibitor; HY-10256, MCE, China), or a combination of both, prior to stimulation with LPS for 24 hours.
CCK8 Assay
BV-2 cells were seeded in 96-well plates and allowed to adhere for 12 hours. Subsequently, the cells were treated with gradient concentrations (0, 3, 6, 12.5, 25 and 50μM) of Oba for 24 hours. Cell viability was then evaluated using the CCK-8 assay kit (K1018, Apexbio, USA) according to the manufacturer’s instructions. Absorbance was measured at 450 nm using a microplate reader.
Calcein AM /PI Cell Activity Assay
BV-2 cells were seeded in 24-well plates and treated with concentrations (0, 25 and 50μM) of Oba for 24 hours. Subsequently, an appropriate volume of Calcein AM/PI double staining solution (C2015S, Beyotime, China) was added to each well. The plates were incubated at 37 °C for 30 minutes in the dark. Post-incubation, cell viability was assessed using a fluorescence microscope: live cells (stained green with Calcein AM) and dead cells (stained red with PI) were visualized. All steps were carried out under light-protected conditions.
Real-Time Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
The total RNA was isolated from cells using the ALFA-SEQ RNA Kit (RZ319-02, Findrop, China). The cDNA was synthesized from 1 µg RNA using the Evo M-MLV RT Kit with gDNA removal (AG11705, Accurate Biotechnology, China). qPCR amplification was carried out with SYBR Green Premix Pro Taq HS qPCR Kit (AG11746, Accurate Biotechnology, China) on a real-time PCR system. The primer sequences used were: IL-1β: forward 5′–CTCGCAGCAGCACATCAACAAG–3′, reverse 5′–CCACGGGAAAGACACAGGTAGC–3′; TNFα: forward 5′-ATGGCCTCCCTCTCATCAGT–3′, reverse 5′–AAGGTACAACCCATCGGCTG–3′. β-actin: forward 5′–CATCCGTAAAGACCTCTATGCCAAC–3′, reverse 5′–ATGGAGCCACCGATCCACA–3′. The expression of target genes was normalized to β-actin and relative quantification was analyzed using the 2−ΔΔCt method.
Annexin V-FITC/PI Apoptosis Assay
Apoptosis of HT22 cells was assessed using an Annexin V-FITC/PI Apoptosis Detection Kit (CA1020, Solarbio, China) according to the manufacturer’s instructions. Briefly, after co-culture, HT22 cells were harvested, digested with trypsin, and resuspended in binding buffer. The cells were then stained with 5 µL of Annexin V-FITC for 10 minutes at room temperature in the dark, followed by staining with 5 µL of propidium iodide (PI) for 5 minutes under the same conditions. Apoptotic cells were subsequently analyzed using a flow cytometer (Beckman Coulter).
Molecular Docking
The 3D structure of Oba in SDF format was obtained from the PubChem database. The crystal structures of TLR4 (PDB ID: 2z62), MyD88 (PDB ID: 4dom), and p38 MAPK (PDB ID: 1a9u) were retrieved from the Protein Data Bank (PDB) and preprocessed using PyMOL. Molecular docking simulations of Oba with these target proteins were conducted using AutoDockTools-1.5.7. The results were visualized and analyzed with PyMOL and Discovery Studio 4.5 Client.
Cell Transfection
The pcDNA3.1-TLR4 plasmid and the negative control (empty) plasmid were designed and synthesized by Synbio Technologies, and were used to establish the Oe‑TLR4 and Oe‑NC groups, respectively. Transfection was performed using Lipofectamine 3000 reagent (Invitrogen, USA) according to the manufacturer’s instructions. Briefly, Lipofectamine 3000, P3000™ reagent, and the plasmid were separately diluted in Opti-MEM medium, combined, and incubated at room temperature for 20 minutes to form transfection complexes. The complex was then added to the cell culture medium. After 6 hours of transfection, the medium was replaced with fresh complete medium, and the cells were cultured for an additional 48 hours before subsequent experiments.
Statistical Analysis
All statistical analyses were performed using SPSS 24.0 and GraphPad Prism 9.4.1. The sample size (n) refers to the number of independent biological replicates. For in vivo studies, n is the number of individual mice. For in vitro experiments, n is the number of independent cultures, each derived from a separate experimental repeat. Data are presented as mean ± standard deviation (SD). The normality of data distribution was assessed using the Shapiro–Wilk test. Differences between two groups were assessed using Student’s t-test. The homogeneity of variances was verified using Levene’s, Brown-Forsythe or Bartlett’s test. For analyses where this assumption was violated, Welch’s ANOVA was employed, followed by Dunnett’s T3 post hoc test. The comparisons among multiple groups were analyzed by one-way or two-way ANOVAs followed by Tukey’s post hoc test. A p-value less than 0.05 was considered statistically significant.
Results
Network Pharmacology and Bioinformatics Analysis
Initially, the relevant functions and mechanisms of Oba in treating SCI were predicted using network pharmacology and bioinformatics. The chemical structure of Oba was shown in Figure 1A. A total of 107 potential therapeutic targets of Oba and 10,406 SCI-related targets were predicted. Venn diagram analysis revealed 96 overlapping genes between Oba and SCI (Figure 1B). Subsequent GO enrichment analysis suggested Oba’s potential involvement in SCI-related processes including regulation of inflammatory response, regulation of apoptotic signaling pathway, and the p38 MAPK cascade. KEGG pathway analysis indicated that Oba was related to Toll-like receptor signaling pathways, MAPK signaling pathways, apoptosis signaling pathways, and TNF signaling pathways in SCI (Figure 1C). A protein-protein interaction (PPI) network was constructed to visualize the functional associations among 96 overlapping genes (Figure S1). Together, these results suggest that Oba may alleviate SCI by exerting anti-inflammatory and anti-apoptotic effects, potentially through the modulation of the Toll-like receptor and p38 MAPK signaling pathways.
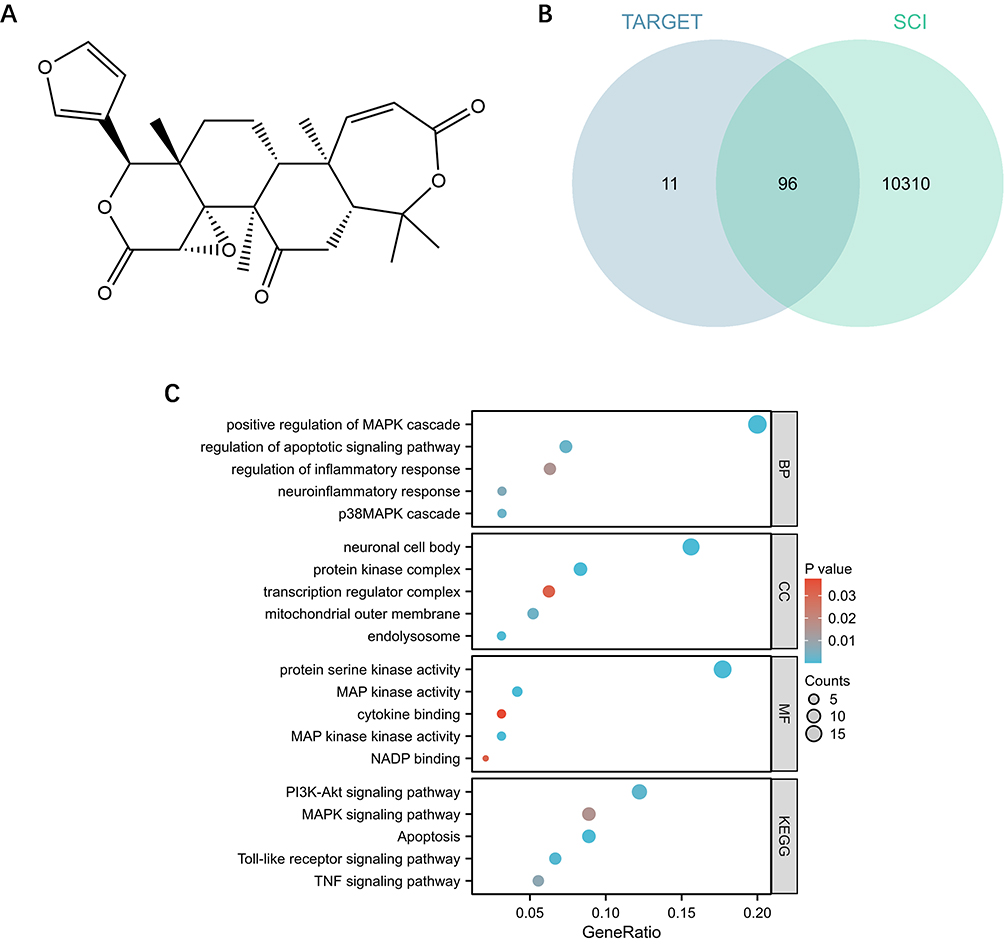 |
Figure 1 Network pharmacology and bioinformatics analysis. (A) Chemical structure of Oba. (B) Venn diagram illustrating 96 overlapping genes between Oba and SCI. (C) GO and KEGG enrichment analysis of 96 genes. |
Obacunone Facilitated Functional Recovery and Promoted Neural Regeneration Following SCI
To assess the therapeutic potential of Oba in SCI, an experimental timeline was designed (Figure 2A) and a mouse SCI model was established (Figure 2B). Oba was administered at doses of 10 mg/kg and 20 mg/kg. Following SCI, mice exhibited paralysis of both hindlimbs. Compared with the SCI group, administration of 20 mg/kg Oba improved hindlimb motor function on day 28. This improvement was evidenced by increased BMS scores (p=0.0133, two-way ANOVA with Tukey’s post hoc test) (Figure 2C), enhanced swimming performance (p=0.0220, two-way ANOVA with Tukey’s post hoc test) (Figure 2D and E), and greater stride length (p=0.0003, two-way ANOVA with Tukey’s post hoc test) (Figure 2F and G). In contrast, the 10 mg/kg dose did not elicit improvements in these behavioral tests compared to the SCI group (BMS scores: p=0.1468; swimming performance: p=0.0856; stride length: p=0.0707; two-way ANOVA with Tukey’s post hoc test). These findings suggest that Oba administered at 20 mg/kg promotes functional recovery after SCI. Since the comprehensive behavioral evaluation indicated that the 10 mg/kg dose did not provide a significant therapeutic benefit, it was excluded from further study. The 20 mg/kg dose, which demonstrated efficacy, was advanced for all subsequent investigations.
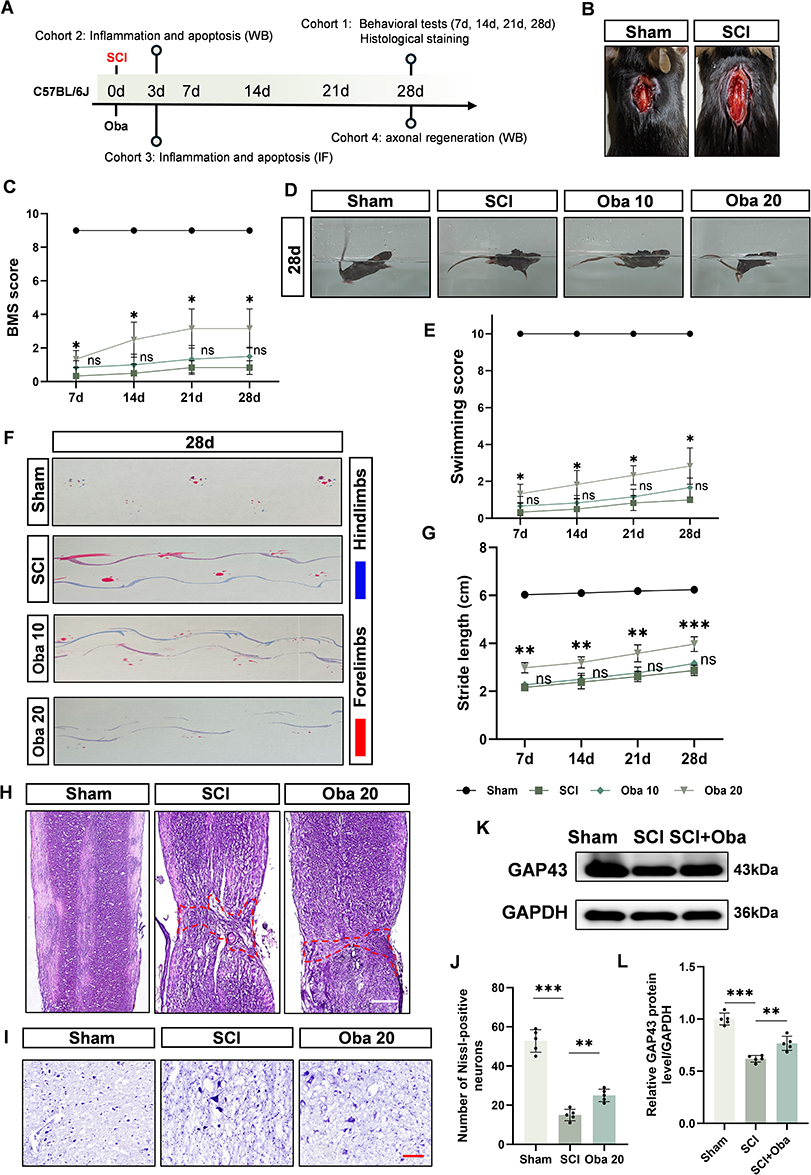 |
Figure 2 Oba Promotes Functional Recovery and Neural Regeneration after SCI. (A) Schematic of the experimental timeline. (B) Representative image of the spinal cord injury model. (C) Quantitative assessment of locomotor recovery evaluated by the BMS over time. (D) Representative images from the swimming test on day 28 post-injury. (E) Quantitative analysis of swimming performance scores. (F) Representative footprint patterns recorded on day 28. (G) Quantification of hindlimb stride length (cm). All behavioral assessments were performed in the following four experimental groups (n=6 per group), represented by distinct symbols: Sham (circle), SCI (square), Oba 10 (SCI + 10 mg/kg Oba; diamond), and Oba 20 (SCI + 20 mg/kg Oba; triangle). Statistical comparisons for all parameters were made against the SCI group. (H) Representative images of HE staining at 28 days post-SCI; scale bar = 500μm. The area within the red dotted line represents the extent of the injury. (I) Representative images of Nissl staining at 28 days post-SCI; scale bar = 100μm. (J) Quantitative analysis of Nissl-positive neurons (n = 5). (K) Representative Western blot image of GAP43 in spinal cord tissues from Sham, SCI, and SCI + Oba (20 mg/kg) groups at 28 days post-SCI. (L) Quantitative analysis of GAP43 protein level (n = 5). The values are presented as mean ± SD. *p<0.05, **p<0.01, ***p<0.001; NS, not significant. |
Next, we evaluated the repair effect of 20 mg/kg Oba on spinal cord tissue. HE staining showed that the tissue structure of the sham group was intact and well-organized. In contrast, the SCI group showed severe structural damage at the lesion site, with a wide range of injuries (red dotted line) and the appearance of vacuoles. Oba treatment reduced the extent of tissue damage and partially restored the integrity of the injured area (Figure 2H). Neuronal regeneration was further assessed by Nissl staining. A decrease in the number of Nissl-positive neurons was observed near the injury site in the SCI group, suggesting that substantial neuronal loss and impaired survival had occurred. Compared with the SCI group, Oba treatment increased the count of Nissl-positive neurons (p=0.0065, one-way ANOVA with Tukey’s post hoc test), suggesting enhanced neuronal survival (Figure 2I and J). To evaluate axonal regeneration after Oba treatment, we used Western blot to detect the regeneration marker GAP43 at 28 days after SCI. Western blot analysis confirmed that GAP43 protein expression was downregulated in the SCI group compared to the Sham group. Compared with the SCI group, Oba treatment increased GAP43 protein levels (p=0.0027, one-way ANOVA with Tukey’s post hoc test) (Figure 2K and L). These findings suggest that Oba promotes neuronal survival and axonal regeneration after SCI, contributing to the recovery of motor function.
Obacunone Mitigated Inflammation and Reduced Apoptosis Following SCI
To verify whether Oba exerts its effects in SCI through anti-inflammatory and anti-apoptotic pathways as predicted, we examined changes in the inflammatory markers (iNOS, IL-1β, TNFα) and apoptotic markers (Bax, Cleaved caspase-3) in spinal cord tissues at 3 days post-SCI. Western blot analysis showed that compared with the Sham group, the expression of iNOS, IL-1β and TNFα in the SCI group were increased. Compared to the SCI group, Oba treatment significantly reduced the expression of pro-inflammatory markers iNOS, IL-1β, and TNFα (p=0.0004, p=0.0010, and p=0.0055; one-way ANOVA with Tukey’s post hoc test) (Figure 3A and B). Immunofluorescence revealed increased infiltration of iNOS-positive cells in spinal cord tissues after SCI, and this effect was significantly attenuated by Oba administration (SCI+Oba group vs SCI group: p<0.0001, one-way ANOVA with Tukey’s post hoc test) (Figure 3C and D). Subsequently, we investigated the apoptosis after SCI. Western blot analysis indicated that the expression of pro-apoptotic proteins Bax and Cleaved caspase-3 was significantly upregulated in the SCI group compared to the Sham group. Compared to the SCI group, Oba treatment significantly suppressed the upregulation of both Bax (p=0.0008, one-way ANOVA with Tukey’s post hoc test) and Cleaved caspase-3 (p=0.0037, one-way ANOVA with Tukey’s post hoc test) (Figure 3E and F). TUNEL staining confirmed that Oba treatment significantly reduced the number of apoptotic cells in spinal cord tissues compared to the SCI group (p<0.0001, one-way ANOVA with Tukey’s post hoc test) (Figure 3G and H). These results indicate that Oba can mitigate secondary injuries, such as inflammation and apoptosis, consistent with the predictions from our network pharmacology and bioinformatics analyses.
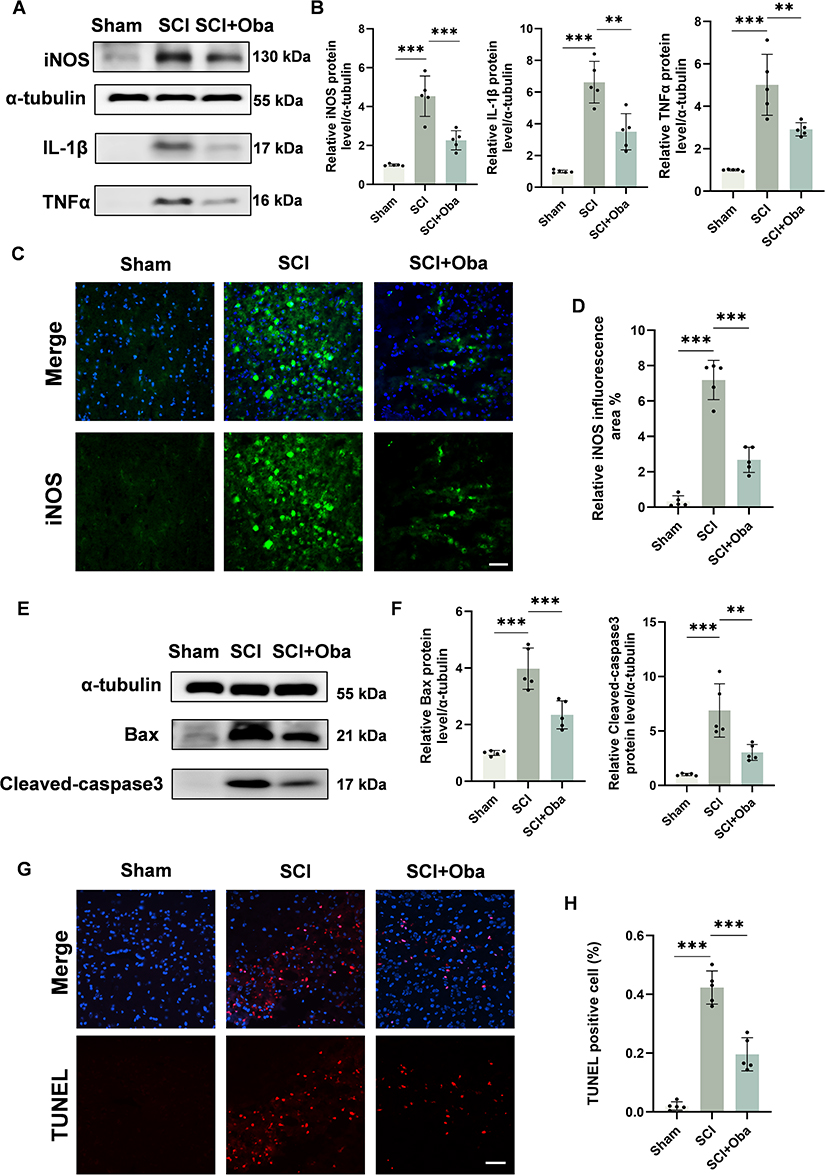 |
Figure 3 Oba attenuated neuroinflammation and reduced cellular apoptosis following SCI. All experiments were performed using tissues from the Sham, SCI, and SCI + Oba (20 mg/kg) groups at 3 days post-SCI. (A) Representative Western blot image of iNOS, IL-1β and TNFα at 3 days post-SCI. (B) Quantitative analysis of iNOS, IL-1β and TNFα protein levels (n = 5). (C) Representative immunofluorescence images of iNOS (green) and DAPI (blue) in spinal cord tissue at 3 days post-SCI; Scale bar = 50 μm. (D) Quantitative analysis of iNOS fluorescence areas (n = 5). (E) Representative Western blot image of Bax and Cleaved-caspase3 at 3 days post-SCI. (F) Quantitative analysis of Bax and Cleaved-caspase3 protein levels (n = 5). (G) Representative immunofluorescence images of TUNEL (red) and DAPI (blue) in spinal cord tissue at 3 days post-SCI; Scale bar = 50 μm. (H) Quantitative analysis of TUNEL positive cells (n = 5). The values are presented as mean ± SD. *p<0.05, **p<0.01, ***p<0.001; NS, not significant. |
Obacunone Inhibits Microglial Inflammation and Consequently Reduces the Apoptosis of HT22 Neurons
Based on the anti-inflammatory properties of Oba and the critical role of microglial inflammation in secondary injury, we further investigated the anti-inflammatory effects of Oba on microglia using an LPS-induced BV-2 microglial cell model. The safe concentration range of Oba was first determined using the CCK-8 assay and Calcein AM/PI assay. The CCK-8 assay demonstrated that various concentrations of Oba did not impair cell viability (25 μM vs 0 μM: p=0.9631; 50 μM vs 0 μM: p=0.6004; one-way ANOVA with Tukey’s post hoc test) (Figure 4A). This was further confirmed by Calcein AM/PI staining, which showed no significant cell death at the tested concentrations (25 μM vs 0 μM: p=0.6479; 50 μM vs 0 μM: p=0.8559; one-way ANOVA with Tukey’s post hoc test) (Figure 4B and C). The experimental results confirm that the selected doses of 25 μM and 50 μM are within the safe range. Therefore, we further utilized Oba at concentrations of 25 μM and 50 μM for subsequent anti-inflammatory studies. Western blot analysis revealed that LPS stimulation markedly upregulated the expression of iNOS, TNFα and IL-1β in BV-2 cells. Compared to the LPS group, treatment with 50 μM Oba significantly reduced the levels of these inflammatory markers (iNOS: p<0.0001; TNFα: p=0.0002; IL-1β: p=0.0001; one-way ANOVA with Tukey’s post hoc test), whereas 25 μM Oba failed to suppress IL-1β expression effectively (p=0.3675, one-way ANOVA with Tukey’s post hoc test) (Figure 4D and E). Compared to the LPS group, immunofluorescence results showed that Oba, particularly at 50 μM, significantly decreased iNOS fluorescence intensity (p<0.0001, one-way ANOVA with Tukey’s post hoc test) (Figure 4F and G). Moreover, qPCR analysis indicated that 50 μM Oba also downregulated the mRNA expression of TNFα (p=0.0002, one-way ANOVA with Tukey’s post hoc test) and IL-1β (p=0.0015, one-way ANOVA with Tukey’s post hoc test) compared to the LPS group (Figure 4H). These results suggest that Oba effectively attenuates inflammatory responses in BV-2 cells. Based on these findings, a concentration of 50 μM was selected for subsequent in vitro experiments.
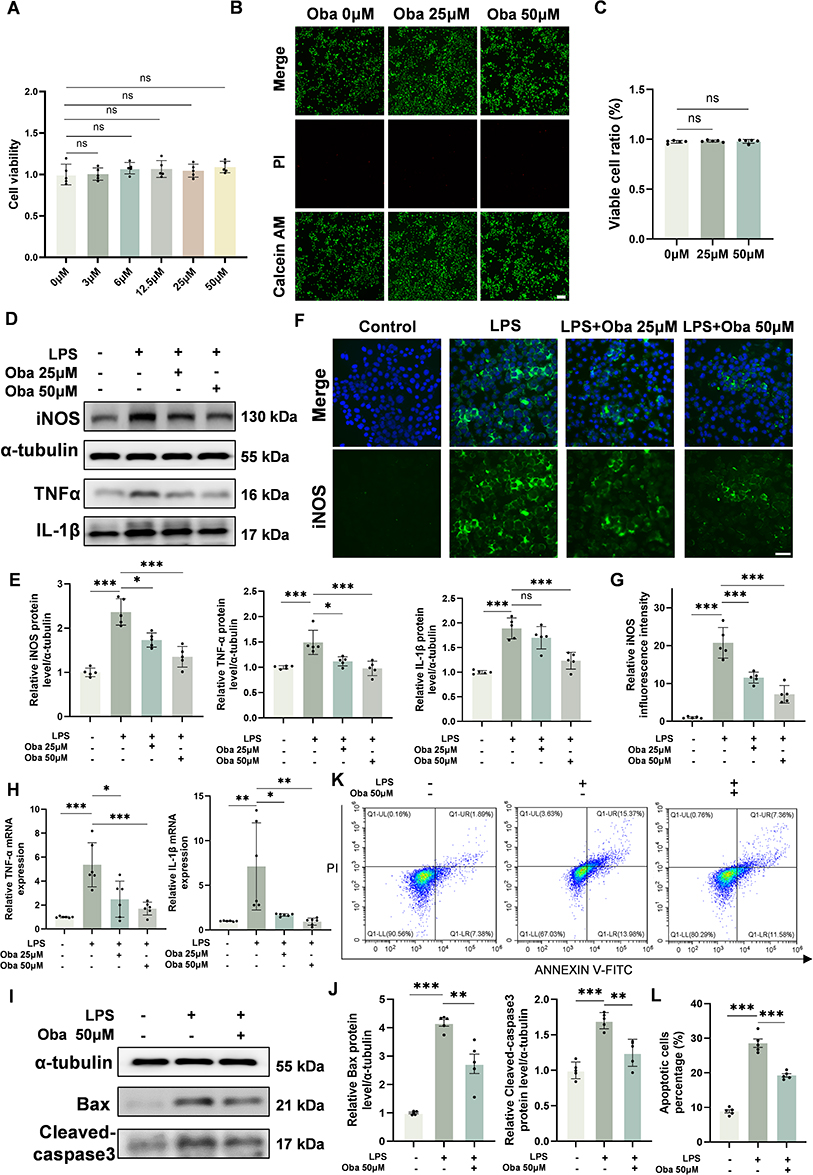 |
Figure 4 Oba suppresses microglial inflammation and blocks the subsequent apoptosis of HT22 neurons. (A) Viability of BV-2 cells treated with different concentrations of Oba, as assessed by CCK-8 assay (n = 5). (B) Representative Calcein AM (green)/PI (red) staining images of BV-2 cells following treatment with Oba at the indicated concentrations (0, 25, and 50 μM). Scale bar = 100 μm. (C) Quantitative analysis of the ratio of viable cells (n = 5). (D) Representative Western blot images of iNOS, TNFα, and IL-1β expression in BV-2 cells. (E) Quantitative analysis of iNOS, TNFα and IL-1β protein levels (n = 5). (F) Representative immunofluorescence images of iNOS (green) and DAPI (blue) in BV-2 cells treated as follows: vehicle (Control), LPS, LPS+Oba 25 μM, and LPS+Oba 50 μM. Scale bar = 50 μm. (G) Quantitative analysis of fluorescence intensity of iNOS (n = 5). (H) mRNA expression levels of TNFα and IL-1β in BV-2 cells measured by qPCR (n = 6). (I) Representative Western blot image of Bax and Cleaved-caspase3 protein level in HT22 cells. (J) Quantitative analysis of Bax and Cleaved-caspase3 protein levels (n = 5). (K) Representative flow cytometry plots of HT22 cells stained with Annexin V-FITC (x-axis) and PI (y-axis). (L) The percentage of apoptotic HT22 cells, as determined by flow cytometry. (n = 5). The plus and minus signs (+, -) denote the presence or absence of the following reagents in culture: LPS, 25 μM Oba, and 50 μM Oba. The values are presented as mean ± SD. *p<0.05, **p<0.01, ***p<0.001; NS, not significant. |
Subsequently, a co-culture system of BV-2 microglia and HT22 neuronal cells was employed to evaluate whether Oba could protect neurons by mitigating microglia-mediated inflammation. The Western blot analysis showed that the LPS group exhibited a significant increase in the expression of Bax and Cleaved caspase-3, indicating enhanced neuronal apoptosis. Compared to the LPS group, Oba treatment effectively attenuated this upregulation of pro-apoptotic proteins (Bax: p=0.0010; Cleaved caspase-3: p=0.0010; one-way ANOVA with Tukey’s post hoc test) (Figure 4I and J). Flow cytometry results further confirmed the anti-apoptotic effect of Oba, demonstrating a significant reduction in apoptosis rate relative to the LPS group (p<0.0001, one-way ANOVA with Tukey’s post hoc test) (Figure 4K and L). These findings indicate that Oba may protect neuronal cells from apoptosis by suppressing microglial inflammatory responses.
Obacunone Modulates the TLR4/MyD88/p38 MAPK Signaling Pathway in Both in vivo and in vitro
To validate whether Oba exerts its effects via the Toll-like receptor and p38 MAPK signaling pathways as hypothesized, we performed the following experiments. Molecular docking analysis revealed that Oba binds effectively within the active pockets of TLR4 (Figure 5A–D), MyD88 (Figure 5E–H), and p38 MAPK proteins (Figure 5I–L). The predominant binding pose for each target exhibited favorable binding energies: TLR4 (−8.1 kcal/mol), MyD88 (−7.6 kcal/mol), and p38 MAPK (−7.8 kcal/mol), indicating a stable interaction with Oba. Western blot results showed that Oba treatment significantly attenuated the LPS-induced upregulation of TLR4, MyD88, and p38 phosphorylation in BV-2 cells (TLR4: p=0.0020; MyD88: p=0.0007; p38 phosphorylation: p=0.0013; one-way ANOVA with Tukey’s post hoc test) (Figure 5M and N). Similarly, in vivo experiments showed that Oba suppressed the SCI-induced upregulation of TLR4, MyD88, and p38 phosphorylation (p=0.0006, p=0.0015, and p=0.0011; one-way ANOVA with Tukey’s test) (Figure 5O and P). These findings suggest that Oba may mitigate inflammation following SCI by downregulating TLR4 and MyD88 expression and suppressing p38 phosphorylation.
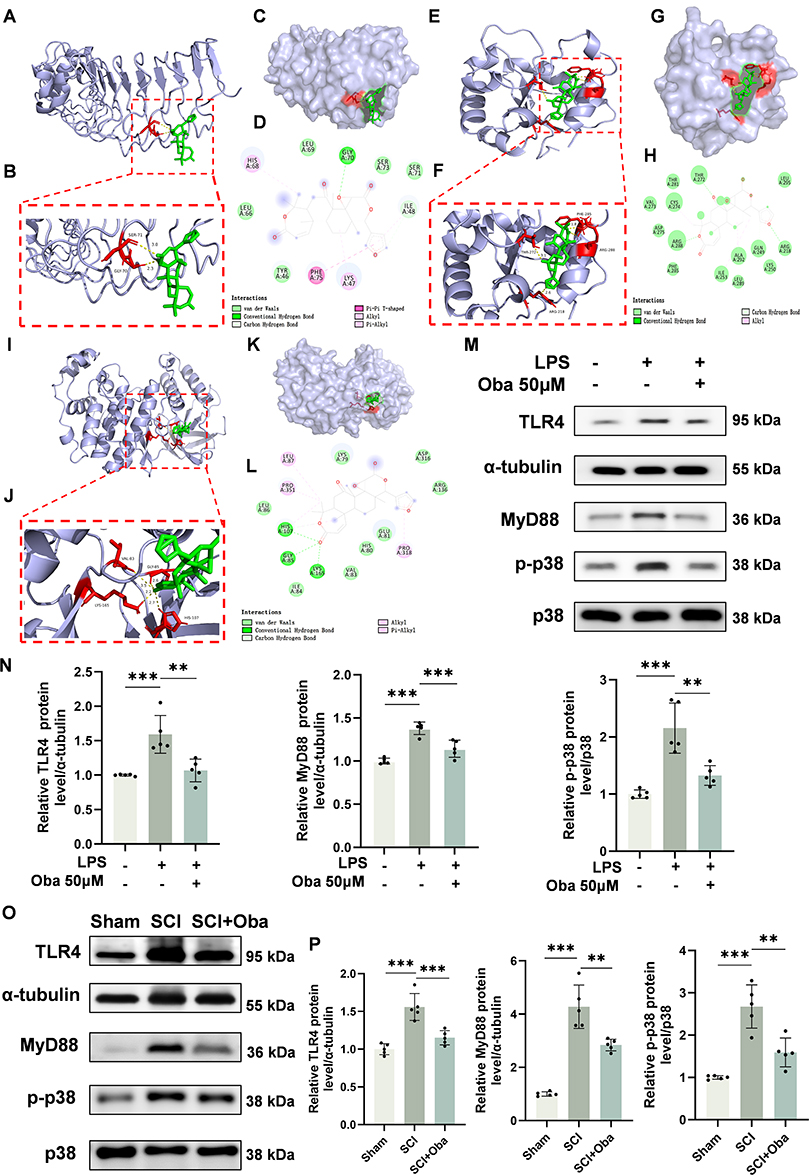 |
Figure 5 Oba inhibits the TLR4/MyD88/p38 MAPK signaling pathway in both in vivo and in vitro experiments. (A–D) Molecular docking analysis of Oba with TLR4. (E–H) Molecular docking analysis of Oba with MyD88. (I–L) Molecular docking analysis of Oba with p38. For each protein, the analysis is presented as: an overall docking pose with the binding region outlined by a red dashed box (A, E, I); a close-up view of the binding region (B, F, J); a surface representation (C, G, K); and a two-dimensional interaction diagram (D, H, L). (M) Representative Western blot image of TLR4, MyD88, p-p38 and p38 protein in BV-2 cells. The plus and minus signs (+, -) denote the presence or absence of the following reagents in culture: LPS, and 50 μM Oba. (N) Quantitative analysis of TLR4 and MyD88 protein levels and p38 phosphorylation in BV-2 cells (n = 5). (O) Western blot analysis of TLR4, MyD88, p-p38, and p38 in spinal cord tissues from Sham, SCI, and SCI + Oba (20 mg/kg) groups at 3 days post-SCI. (P) Quantitative analysis of TLR4 and MyD88 protein levels and p38 phosphorylation in spinal cord tissues (n = 5). The values are presented as mean ± SD. *p<0.05, **p<0.01, ***p<0.001; NS, not significant. |
Obacunone Modulates the TLR4/MyD88/p38 MAPK Signaling Pathway to Suppress Inflammatory Responses
To determine whether the anti-inflammatory effects of Oba are mediated through the TLR4/MyD88/p38 MAPK signaling pathway, we performed the following experiments. Western blot analysis revealed that transfection with pcDNA3.1-TLR4 significantly increased TLR4 protein expression in BV-2 cells compared to cells transfected with the empty vector (p<0.0001, unpaired t-test) (Figure 6A and B). Western blot analysis showed that LPS stimulation increased the protein levels of the inflammatory mediators iNOS, TNFα, and IL-1β, and Oba treatment inhibited the expression of these inflammatory factors. However, TLR4 overexpression markedly attenuated the protective effects of Oba (iNOS, TLR4, TNFα, and IL-1β: p=0.0426, p=0.0027, p=0.0018, and p=0.0002; one-way ANOVA with Tukey’s test) (Figure 6C–G). To further assess the involvement of p38 MAPK in Oba-mediated suppression of pro-inflammatory mediators, we pretreated BV-2 cells for 3 hours with Oba, SB203580, or their combination before LPS exposure. Oba treatment reduced the protein levels of iNOS, TNFα, IL-1β and inhibited p38 phosphorylation, consistent with the effect of the p38 inhibitor SB203580. Moreover, compared with SB203580 treatment alone, the combination of SB203580 and Oba further reduced the phosphorylation level of p38 (p=0.0002, one-way ANOVA with Tukey’s post hoc test). Similarly, relative to SB203580 monotherapy, co-treatment with SB203580 and Oba significantly decreased the expression of iNOS protein (p=0.0075, one-way ANOVA with Tukey’s post hoc test) (Figure 6H–L). Collectively, these results indicate that Oba attenuates inflammation through the TLR4/MyD88/p38 MAPK signaling cascade.
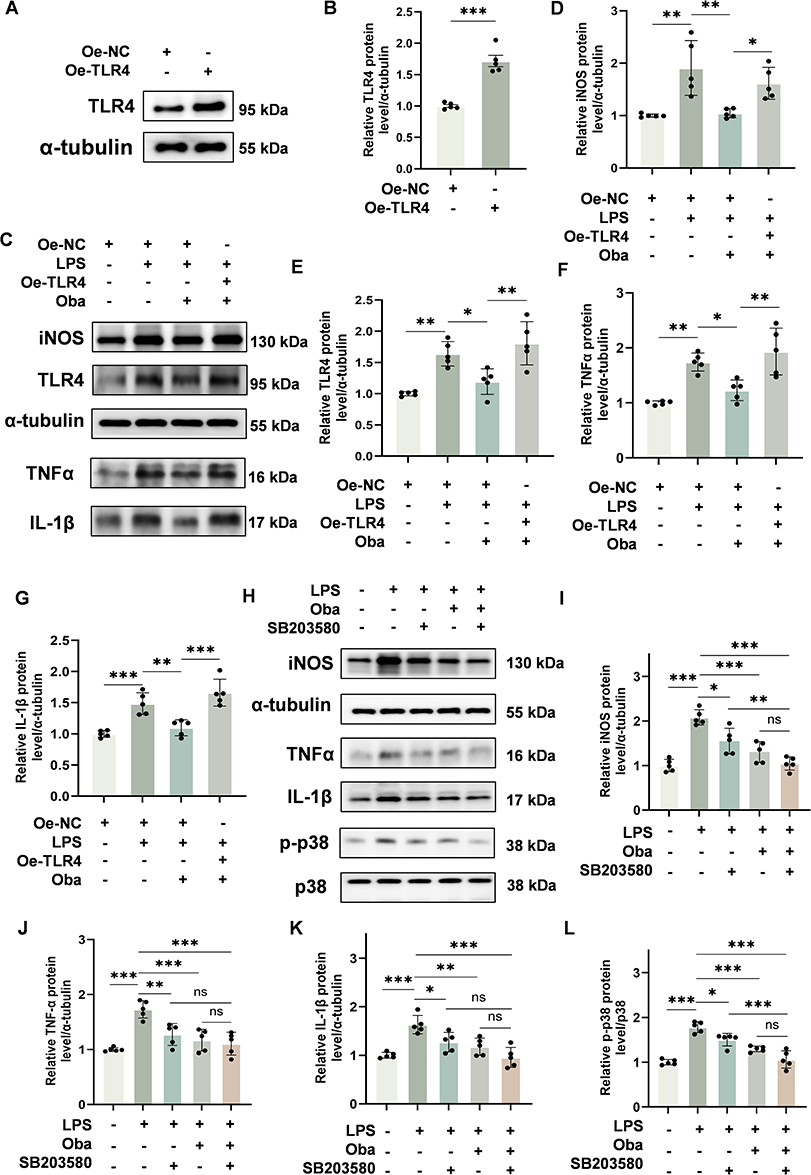 |
Figure 6 Oba attenuates inflammation by targeting the TLR4/MyD88/p38 MAPK pathway in BV-2 microglial cells. (A) Representative Western blot of TLR4 expression in BV-2 cells transfected with a TLR4 overexpression plasmid (pcDNA3.1-TLR4) or an empty vector. The plus and minus signs (+, -) denote the presence or absence of the following reagents in culture: empty plasmid (Oe-NC), and TLR4 overexpression plasmid (Oe-TLR4). (B) Quantitative analysis of TLR4 protein levels (n = 5). (C) Western blot images of iNOS, TLR4, TNFα and IL-1β protein expression in BV-2 cells. The plus and minus signs (+, -) denote the presence or absence of the following treatments in culture: Oe-NC, Oe-TLR4, LPS, and Oba. (D–G) Quantitative analysis of iNOS, TLR4, TNFα and IL-1β protein levels (n = 5). (H) Representative Western blot images of iNOS, TNFα, IL-1β, p-p38 and p38 in BV-2 cells. The plus and minus signs (+, -) denote the presence or absence of the following reagents in culture: LPS, Oba, and SB203580. (I–L) Quantitative analysis of iNOS, TNFα, IL-1β protein levels and p38 phosphorylation (n = 5). The values are presented as mean ± SD. *p<0.05, **p<0.01, ***p<0.001; NS, not significant. |
Discussion
The complex inflammatory response following secondary spinal cord injury is driven by multiple cell types—including activated microglia, astrocytes, and peripherally derived macrophages—as well as a wide array of inflammatory mediators such as TNFα, IL-1β, and IL-6. This response exacerbates tissue damage and promotes cellular death.18 Neuroinflammation is a critical factor influencing the extent of spinal cord damage. Modulating the inflammatory response represents a key therapeutic strategy to promote neural regeneration and ultimately improve functional recovery.19 Meanwhile, apoptosis, induced through multiple mechanisms such as inflammatory injury and cytokine-mediated pathways, also plays a critical role in the pathogenesis of SCI.20 The progressive nature of apoptosis contributes directly to the expansion of secondary damage, leading to substantial loss of neurons and oligodendrocytes, and ultimately resulting in long-term neurological impairment.2 Effective suppression of apoptosis has been demonstrated to play a crucial role in improving outcomes after SCI.21 The design of innovative treatment approaches for spinal cord injury—prioritizing precision, specificity, minimal invasiveness, and cost efficiency—has gained significant attention as a key direction in current research.22 This study investigated the therapeutic potential of Oba in SCI using an integrated approach combining network pharmacology, bioinformatics, as well as in vivo and in vitro experiments. Our results demonstrate that Oba promotes functional recovery and nerve repair following spinal cord injury, largely attributable to its neuroprotective effects in mitigating inflammation and apoptosis. Given its natural abundance, Oba represents a promising candidate for SCI treatment.
Toll-like receptors (TLRs) are a class of innate immune receptors that play a significant role in modulating glial and neuronal functions within the central nervous system under both physiological and pathophysiological conditions. They are critically involved in innate immune responses and neuroinflammatory processes.23 In the context of SCI, inhibition of TLR4 has been demonstrated to effectively attenuate post-injury inflammation and represents a promising therapeutic strategy. For instance, sarcoside has been shown to promote functional recovery after SCI by suppressing TLR4-mediated neuroinflammation and neuronal apoptosis.24 Similarly, fibroblast growth factor 10 (FGF10), secreted by neurons and microglia/macrophages, activates the FGFR2/PI3K/Akt pathway in neurons while inhibiting TLR4/NF-κB-dependent neuroinflammation in microglia/macrophages, thereby improving recovery outcomes.25 Additionally, baicalin has been found to alleviate neuroinflammation by targeting the TLR4/MD2 complex on microglia and modulating the PI3K/Akt/NF-κB axis.6 Furthermore, regulation of TLR4 signaling influences cytokine expression and extracellular matrix remodeling in the chronic phase of SCI, contributing to reduced secondary damage and enhanced functional recovery.26 Notably, Oba has been shown to alleviate inflammation and preserve the intestinal epithelial barrier through attenuation of the TLR4/NF-κB signaling cascade In ulcerative colitis.27 Building on this premise, we therefore performed KEGG pathway enrichment analysis. The results indicated that Oba may be involved in the regulation of the Toll-like receptor signaling pathway following SCI. We therefore hypothesized that Oba exerts anti-inflammatory effects via the TLR4 signaling pathway. This hypothesis was supported by molecular docking simulations, which revealed stable binding interactions between Oba and both TLR4 and MyD88, with binding free energies of –8.1 kcal/mol and –7.6 kcal/mol, respectively. Subsequent in vitro and in vivo experiments consistently demonstrated that Oba significantly downregulated the protein expression of TLR4 and MyD88. Concomitantly, Oba markedly suppressed the protein expression of key downstream pro-inflammatory mediators, including iNOS, TNFα, and IL-1β, at the protein level. Importantly, the protective effects of Oba were attenuated by TLR4 overexpression in BV-2 microglial cells. Collectively, these findings demonstrate that TLR4 and MyD88 mediate the anti-inflammatory effects of Oba in SCI.
The p38 MAPK signaling pathway plays a critical role in the regulation of inflammation.28 Its activation contributes to the release of key pro-inflammatory cytokines during the secondary injury phase following SCI.29 Previous studies have demonstrated that inhibition of p38 phosphorylation after SCI effectively suppresses neuroinflammation and promotes functional recovery. For example, oleanolic acid has been shown to attenuate inflammation and apoptosis by inhibiting the activation of p38 and JNK MAPK pathways, thereby improving outcomes in a mouse model of SCI.30 Similarly, salidroside reduces inflammatory responses and enhances motor function recovery in SCI rats through suppression of NF-κB, p38, and ERK signaling.31 Additionally, kaempferol pretreatment alleviates microglia-driven neuroinflammation following secondary SCI by inhibiting the p38 pathway and associated apoptosis.32 These prior findings collectively position p38 MAPK as a promising therapeutic target for SCI. Intriguingly, Oba has also been shown to modulate this pathway. In RAW264.7 cells, Oba has been reported to stabilize MKP-1 protein expression by targeting MIF, leading to dephosphorylation of p-p38 and subsequent reduction in pro-inflammatory mediator production.33 Furthermore, Oba mitigates osteoarthritis progression by binding to HDAC1 and inhibiting p38 MAPK signaling.34 Guided by this literature, we performed GO and KEGG enrichment analyses, which indicated that Oba may elicit therapeutic benefits in SCI through modulation of the p38 cascade (GO term) and the MAPK signaling pathway (KEGG pathway). Molecular docking simulations further supported this direction by suggesting a stable binding interaction between Oba and the p38 protein, with a binding free energy of –7.8 kcal/mol. Therefore, we hypothesized that Oba may confer neuroprotection in SCI via inhibiting the p38 MAPK pathway. Subsequent in vivo and in vitro experiments verified this hypothesis. In vivo, Oba suppressed the phosphorylation of p38. In LPS-stimulated BV-2 microglia, Oba markedly downregulated the expression of key pro-inflammatory mediators such as iNOS, TNFα, IL-1β, and concurrently inhibited p38 phosphorylation, which was mimicked by the p38 MAPK inhibitor SB203580. Moreover, in a BV-2/HT22 co-culture model, Oba treatment effectively decreased neuronal apoptosis. These results collectively suggest that the neuroprotective effect of Oba is mediated through the inhibition of the p38 MAPK pathway in microglia, leading to reduced neuroinflammation and ultimately less neuronal apoptotic damage.
In the osteoporosis model, studies have confirmed that Oba directly binds to and inhibits MIF, thereby suppressing its downstream ROS, NF-κB, and MAPK signaling pathways, ultimately impeding osteoclast differentiation and alleviating bone loss.35 In the spinal cord injury model, inhibition of HIF-1α has been shown to reduce MIF levels and promote motor function recovery.36 Based on these two mechanisms, we hypothesize that Oba may also exert neuroprotective effects and facilitate functional recovery in spinal cord injury by modulating MIF and its downstream pathways. This constitutes a promising and mechanistically interesting direction for further investigation. Modulating the NF-κB and MAPK signaling pathways, as well as reducing ROS levels, represent effective therapeutic strategies for spinal cord injury.31 In the ulcerative colitis model, Oba has been shown to effectively inhibit the TLR4/NF-κB signaling cascade.27 In a liver fibrosis model, Oba significantly reduced ROS levels by activating the Nrf2/GPx-4 pathway.37 Therefore, Oba may exert a synergistic effect in alleviating inflammation and oxidative damage after SCI through a dual mechanism of suppressing the NF-κB inflammatory pathway and activating the antioxidant pathway, representing a promising direction for future research. Neuropathic pain is the main complication of spinal cord diseases.38 The endogenous factor TAFA4 has been confirmed to play a key role in pain signal processing at the spinal cord level.39 Meanwhile, studies have shown that Oba can alleviate inflammatory neuropathic pain by suppressing inflammatory responses and oxidative stress.11 Investigating the potential relationship between Oba and the endogenous analgesic system TAFA4 is expected to further reveal the mechanism of Oba in alleviating pain after SCI, thereby more comprehensively highlighting its potential as a multidimensional therapeutic strategy.
According to the existing literature, a maximum dose of 100 mg/kg of Obacunone administered via intraperitoneal injection has been reported to be safe.40 In the present study, a lower dose of 20 mg/kg (i.p.) was employed, further supporting the reliability and safety profile of our dosing regimen. In vitro, assessments of cellular viability and cytotoxicity—conducted via CCK-8 assay and Calcein AM /PI Cell activity Assay—confirmed that 50 μM is a safe concentration of Oba treatment in BV-2 cells. Moreover, based on the dose of 20 mg/kg administered to mice in this study, the corresponding Human Equivalent Dose (HED) was calculated to be approximately 1.62 mg/kg using the well-established interspecies dose conversion methodology.41 It provides a valuable reference for the initial dose in future clinical trials involving patients with SCI. Taken together, the demonstrated efficacy and favorable safety profile of Oba underscore its translational potential for the treatment of SCI.
This study has several limitations. First, while we have identified an interaction between Oba and TLR4/MyD88/p38 signaling, whether additional mechanisms contribute to its therapeutic effects remains unclear. Second, as there is no universally accepted standard drug therapy for SCI, our experimental design did not include a positive control group, which limits a direct comparative assessment of Oba’s efficacy. Further work is needed to explore additional mechanisms of Oba and to evaluate its efficacy relative to existing candidate therapeutics.
Conclusion
In a mouse model of SCI, Oba exerted its therapeutic effects primarily by inhibiting the TLR4/MyD88/p38 MAPK pathway to attenuate neuroinflammation, thereby contributing to reduced neuronal apoptosis and enhanced neural repair, with a final improvement in motor function. These findings support the therapeutic feasibility of Oba for SCI and provide an experimental foundation for its further development.
Abbreviations
SCI, spinal cord injury; Oba, obacunone; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; PPI, protein-protein interaction; BMS, Basso Mouse Scale; HE, hematoxylin and eosin; CNS, central nervous system; TLR4, Toll-like receptor 4.
Ethics Approval and Informed Consent
All animal experiments were performed in accordance with Institutional Animal Care and Use Committee guidelines and approved protocols at the Southern Medical University (Ethics Number: LAEC-2024–290).
Acknowledgments
The author wishes to express sincere gratitude to all the teachers, classmates, and colleagues for their invaluable guidance and support throughout this research.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation of Guangdong Province (2024A1515013023) and the National Natural Science Foundation (81974329).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zhou M, Xu Z, Zhong H, Ning G, Feng S. Spinal cord injury and inflammatory mediators: role in “fire barrier” formation and potential for neural regeneration. Neural Regeneration Res. 2026;21(3):923–18. doi:10.4103/NRR.NRR-D-24-00792
2. Shi Z, Yuan S, Shi L, et al. Programmed cell death in spinal cord injury pathogenesis and therapy. Cell Proliferation. 2021;54(3):e12992. doi:10.1111/cpr.12992
3. Zhang Y, Deng Q, Hong H, Qian Z, Wan B, Xia M. Caffeic acid phenethyl ester inhibits neuro-inflammation and oxidative stress following spinal cord injury by mitigating mitochondrial dysfunction via the SIRT1/PGC1α/DRP1 signaling pathway. J Transl Med. 2024;22(1):304. doi:10.1186/s12967-024-05089-8
4. Xia M, Zhang Y, Wu H, et al. Forsythoside B attenuates neuro-inflammation and neuronal apoptosis by inhibition of NF-κB and p38-MAPK signaling pathways through activating Nrf2 post spinal cord injury. Int Immunopharmacol. 2022;111:109120. doi:10.1016/j.intimp.2022.109120
5. Jadhav VS, Khatri DK. Understanding the influence of TLR-mediated immune system on necroptosis-induced neurodegeneration in Parkinson’s disease. Ageing Res Rev. 2025;112:102872. doi:10.1016/j.arr.2025.102872
6. Lu Y, Zhou R, Zhu R, et al. Baicalin ameliorates neuroinflammation by targeting TLR4/MD2 complex on microglia via PI3K/AKT/NF-κB signaling pathway. Neuropharmacology. 2025;267:110296. doi:10.1016/j.neuropharm.2025.110296
7. Liu W, Ma Z, Kang J, et al. Grape seed proanthocyanidins exert a neuroprotective effect by regulating microglial M1/M2 polarisation in rats with spinal cord injury. Mediators of Inflammation. 2022;2022:1–23. doi:10.1155/2022/2579003
8. Chen W, Zhang L, Zhong G, et al. Regulation of microglia inflammation and oligodendrocyte demyelination by Engeletin via the TLR4/RRP9/NF-κB pathway after spinal cord injury. Pharmacol Res. 2024;209:107448. doi:10.1016/j.phrs.2024.107448
9. Zhang Y, Xiao S, Dan F, et al. Phillygenin inhibits neuroinflammation and promotes functional recovery after spinal cord injury via TLR4 inhibition of the NF-κB signaling pathway. J Orthopaedic Transl. 2024;48:133–145. doi:10.1016/j.jot.2024.07.013
10. Zhou Y, Gu J, Li J, et al. Obacunone, a promising phytochemical triterpenoid: research progress on its pharmacological activity and mechanism. Molecules. 2024;29(8):1791. doi:10.3390/molecules29081791
11. Nan F, Tian Q, Chen S. Obacunone alleviates inflammatory pain by promoting M2 microglial polarization and by activating Nrf2/HO-1 signaling pathway. DDDT. 2024;18:1265–1275. doi:10.2147/DDDT.S451281
12. Jeong GS, Byun E, Li B, Lee DS, An RB, Kim YC. Neuroprotective effects of constituents of the root bark of Dictamnus dasycarpus in mouse hippocampal cells. Arch Pharm Res. 2010;33(8):1269–1275. doi:10.1007/s12272-010-0818-9
13. Li YR, Xie LL, Cai HB, et al. Obacunone alleviates thalamic pain via promoting LCN2-mediated phagocytosis of astrocytes in mice. ACS Chem Neurosci. 2025;16(13):2546–2558. doi:10.1021/acschemneuro.5c00371
14. Xie C, Shen X, Xu X, et al. Astrocytic YAP promotes the formation of glia scars and neural regeneration after spinal cord injury. J Neurosci. 2020;40(13):2644–2662. doi:10.1523/JNEUROSCI.2229-19.2020
15. Basso DM, Fisher LC, Anderson AJ, Jakeman LB, Mctigue DM, Popovich PG. Basso mouse scale for locomotion detects differences in recovery after spinal cord injury in five common mouse strains. J Neurotrauma. 2006;23(5):635–659. doi:10.1089/neu.2006.23.635
16. Gu Y, Cheng X, Huang X, et al. Conditional ablation of reactive astrocytes to dissect their roles in spinal cord injury and repair. Brain Behav Immun. 2019;80:394–405. doi:10.1016/j.bbi.2019.04.016
17. Feng Y, Wei Z, Li R, Shi Q, Cai J. Effects and mechanism of berberine in ameliorating microglia-mediated hypothalamic inflammation by downregulating the MAPK and NF-κB pathway. J Ethnopharmacol. 2026;355:120695. doi:10.1016/j.jep.2025.120695
18. Hellenbrand DJ, Quinn CM, Piper ZJ, Morehouse CN, Fixel JA, Hanna AS. Inflammation after spinal cord injury: a review of the critical timeline of signaling cues and cellular infiltration. J Neuroinflammation. 2021;18(1):284. doi:10.1186/s12974-021-02337-2
19. Sterner RC, Sterner RM. Immune response following traumatic spinal cord injury: pathophysiology and therapies. Front Immunol. 2023;13:1084101. doi:10.3389/fimmu.2022.1084101
20. Lu J, Ashwell KWS, Waite P. Advances in secondary spinal cord injury: role of apoptosis. Spine. 2000;25(14):1859–1866. doi:10.1097/00007632-200007150-00022
21. Abbaszadeh F, Fakhri S, Khan H. Targeting apoptosis and autophagy following spinal cord injury: therapeutic approaches to polyphenols and candidate phytochemicals. Pharmacol Res. 2020;160:105069. doi:10.1016/j.phrs.2020.105069
22. Freyermuth-Trujillo X, Segura-Uribe JJ, Salgado-Ceballos H, Orozco-Barrios CE, Coyoy-Salgado A. Inflammation: a target for treatment in spinal cord injury. Cells. 2022;11(17):2692. doi:10.3390/cells11172692
23. Acioglu C, Heary RF, Elkabes S. Roles of neuronal toll-like receptors in neuropathic pain and central nervous system injuries and diseases. Brain Behav Immun. 2022;102:163–178. doi:10.1016/j.bbi.2022.02.016
24. Xiao S, Zhong N, Yang Q, et al. Aucubin promoted neuron functional recovery by suppressing inflammation and neuronal apoptosis in a spinal cord injury model. Int Immunopharmacol. 2022;111:109163. doi:10.1016/j.intimp.2022.109163
25. Chen J, Wang Z, Zheng Z, et al. Neuron and microglia/macrophage-derived FGF10 activate neuronal FGFR2/PI3K/Akt signaling and inhibit microglia/macrophages TLR4/NF-κB-dependent neuroinflammation to improve functional recovery after spinal cord injury. Cell Death Dis. 2017;8(10):e3090. doi:10.1038/cddis.2017.490
26. Ryan F, Francos-Quijorna I, Hernández-Mir G, et al. Tlr4 deletion modulates cytokine and extracellular matrix expression in chronic spinal cord injury, leading to improved secondary damage and functional recovery. J Neurosci. 2024;44(6):e0778232023. doi:10.1523/JNEUROSCI.0778-23.2023
27. Luo X, Yue B, Yu Z, et al. Obacunone protects against ulcerative colitis in mice by modulating gut microbiota, attenuating TLR4/NF-κB signaling cascades, and improving disrupted epithelial barriers. Front Microbiol. 2020;11:497. doi:10.3389/fmicb.2020.00497
28. Wang J, Liu Y, Guo Y, et al. Function and inhibition of P38 MAP kinase signaling: targeting multiple inflammation diseases. Biochem Pharmacol. 2024;220:115973. doi:10.1016/j.bcp.2023.115973
29. Kasuya Y, Umezawa H, Hatano M. Stress-activated protein kinases in spinal cord injury: focus on roles of p38. IJMS. 2018;19(3):867. doi:10.3390/ijms19030867
30. Wang JL, Ren CH, Feng J, Ou CH, Liu L. Oleanolic acid inhibits mouse spinal cord injury through suppressing inflammation and apoptosis via the blockage of p38 and JNK MAPKs. Biomed Pharmacother. 2020;123:109752. doi:10.1016/j.biopha.2019.109752
31. Su Y, Zong S, Wei C, et al. Salidroside promotes rat spinal cord injury recovery by inhibiting inflammatory cytokine expression and NF‐κB and MAPK signaling pathways. J Cell Physiol. 2019;234(8):14259–14269. doi:10.1002/jcp.28124
32. Liu Z, Yao X, Sun B, et al. Pretreatment with kaempferol attenuates microglia-mediate neuroinflammation by inhibiting MAPKs–NF–κB signaling pathway and pyroptosis after secondary spinal cord injury. Free Radic Biol Med. 2021;168:142–154. doi:10.1016/j.freeradbiomed.2021.03.037
33. Gao Y, Hou R, Liu F, et al. Obacunone causes sustained expression of MKP‐1 thus inactivating p38 MAPK to suppress pro‐inflammatory mediators through intracellular MIF. J Cell Biochemistry. 2018;119(1):837–849. doi:10.1002/jcb.26248
34. Gao Y, Wang K, Shi C, Gao Y, Kong DQ. Obacunone acts as a histone deacetylase 1 inhibitor to limit p38MAPK signaling and alleviate osteoarthritis progression. J Orthop Surg Res. 2025;20(1):441. doi:10.1186/s13018-025-05804-1
35. He J, Zheng L, Li X, et al. Obacunone targets macrophage migration inhibitory factor (MIF) to impede osteoclastogenesis and alleviate ovariectomy-induced bone loss. J Adv Res. 2023;53:235–248. doi:10.1016/j.jare.2023.01.003
36. Hao H, Hou Y, Li A, et al.
37. Bai Y, Wang W, Wang L, et al. Obacunone attenuates liver fibrosis with enhancing anti-oxidant effects of GPx-4 and inhibition of EMT. Molecules. 2021;26(2):318. doi:10.3390/molecules26020318
38. Kubota A, Nakajima H, Honjoh K, Watanabe S, Takahashi A, Matsumine A. Differential microglial dynamics and neuroinflammation underlying neuropathic pain in the central nervous system: comparative insights from spinal cord injury and compressive myelopathy models. Front Cell Neurosci. 2026;20:1769004. doi:10.3389/fncel.2026.1769004
39. Zhu S, Hu X, Bennett S, Mai Y, Xu J. Molecular structure, expression and role of TAFA4 and its receptor FPR1 in the spinal cord. Front Cell Dev Biol. 2022;10:911414. doi:10.3389/fcell.2022.911414
40. Qiu Z, He J, Shao G, et al. Obacunone retards renal cyst development in autosomal dominant polycystic kidney disease by activating NRF2. Antioxidants. 2021;11(1):38. doi:10.3390/antiox11010038
41. Nair A, Jacob S. A simple practice guide for dose conversion between animals and human. J Basic Clin Pharm. 2016;7(2):27. doi:10.4103/0976-0105.177703
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Vitamin D Ameliorates Apoptosis and Inflammation by Targeting the Mitochondrial and MEK1/2-ERK1/2 Pathways in Hyperoxia-Induced Bronchopulmonary Dysplasia
Hu J, Wu Z, Wang H, Geng H, Huo J, Zhu X, Zhu X
Journal of Inflammation Research 2022, 15:4891-4906
Published Date: 25 August 2022
Eriochloa villosa Alleviates Progression of Benign Prostatic Hyperplasia in vitro and in vivo
Baek EB, Hwang YH, Park S, Hong EJ, Won YS, Kwun HJ
Research and Reports in Urology 2022, 14:313-326
Published Date: 24 September 2022
Engeletin Alleviates the Inflammation and Apoptosis in Intervertebral Disc Degeneration via Inhibiting the NF-κB and MAPK Pathways
Li B, Yang X, Zhang P, Guo J, Rong K, Wang X, Cao X, Zhou T, Zhao J
Journal of Inflammation Research 2022, 15:5767-5783
Published Date: 10 October 2022
PANoptosis: A Cell Death Characterized by Pyroptosis, Apoptosis, and Necroptosis
Shi C, Cao P, Wang Y, Zhang Q, Zhang D, Wang Y, Wang L, Gong Z
Journal of Inflammation Research 2023, 16:1523-1532
Published Date: 12 April 2023
Wnt3a Inhibits Inflammatory Responses and Improves Survival in Immortalised Schwann Cells
Zheng J, Zhang Y, Nan G
Journal of Inflammation Research 2025, 18:17715-17730
Published Date: 18 December 2025