Back to Journals » Journal of Inflammation Research » Volume 18
Wnt3a Inhibits Inflammatory Responses and Improves Survival in Immortalised Schwann Cells
Authors Zheng J, Zhang Y, Nan G
Received 26 June 2025
Accepted for publication 24 November 2025
Published 18 December 2025 Volume 2025:18 Pages 17715—17730
DOI https://doi.org/10.2147/JIR.S549712
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adam Bachstetter
Jian Zheng,1– 3 Yanting Zhang,2,3 Guoxin Nan1,4
1Orthopedics Department, Dongguan Children’s Hospital Affiliated to Guangdong Medical University, Dongguan, People’s Republic of China; 2Chongqing Medical University, Chongqing, People’s Republic of China; 3Department of Pediatric Research Institute, Children’s Hospital of Chongqing Medical University, Chongqing, People’s Republic of China; 4Dongguan Eighth People’s Hospital, Dongguan, People’s Republic of China
Correspondence: Guoxin Nan, Orthopedics Department, Dongguan Children’s Hospital Affiliated to Guangdong Medical University, Dongguan, People’s Republic of China, Tel +86 13608397992, Email [email protected]
Background: Spinal cord injury (SCI) triggers a intense inflammatory response that hinders the success of cell transplantation therapies. Immortalised Schwann cells (iSCs) offer a renewable cell source, but their response to inflammation is poorly understood. Wnt3a regulates neural stem cells, but its role in modulating inflammatory responses in iSCs remains unclear.
Methods: Rat Schwann cells (SCs) were immortalised using SV40Tag. An inflammatory model was established by treating iSCs with LPS, in the presence or absence of Wnt3a protein. The inflammatory response, apoptosis, proliferation, and migration were assessed using quantitative PCR (qPCR), Western blotting, immunofluorescence, CCK-8 assay, TUNEL staining, flow cytometry, and scratch wound healing assay. An acute spinal cord injury model in rats was utilised for in vivo validation.
Results: This study shows that immortalized Schwann cells share some genotypic similarity with primary Schwann cells, but have a much faster proliferation rate than Schwann cells, which can be better used for neurological disease-related research. In addition, in the LPS-induced inflammatory environment, Wnt3a was able to inhibit the expression of IL-1β in immortalized Schwann cells, and enhance the expression of TGF-β by activating NF-κB. More importantly, Wnt3a inhibited the level of apoptosis in the inflammatory environment and promoted the proliferation and migration ability of cells, which also enhanced the function of immortalized Schwann cells.
Conclusion: Wnt3a modulates the inflammatory response in iSCs, primarily through NF-κB-mediated upregulation of TGF-β, and promotes iSC survival and function. The combination of iSCs and Wnt3a presents a promising strategy for improving cell transplantation therapy for SCI.
Keywords: spinal cord injury, immortalised schwann cells, inflammation, Wnt3a, NF-κB signaling pathway, apoptosis
Introduction
Spinal cord injury (SCI) is a severe traumatic disease of the central nervous system(CNS),1 and cell-based regenerative therapies have shown promise in the treatment of SCI, with exogenously transplanted cells improving the microenvironment, promoting vascular regeneration and playing a neuroprotective role.2,3 Schwann cells (SCs) are glial cells in the peripheral nervous system (PNS) that play a crucial role in peripheral nerve regeneration after traumatic injury and peripheral neuropathy.4,5 When transplanted, SCs act as a scaffold that guides regenerating axons to their intended target sites, thereby improving or restoring the conduction velocity of demyelinated axons.6,7
However, during the early stages of SCI, an increase in pro-inflammatory factors and chemokines leads to an intense inflammatory response and apoptosis at the site of injury.8,9 This inflammatory response and apoptosis reduce cell survival and proliferation, thus limiting the efficacy of detrimental to cell survival and proliferation, and the therapeutic means of cell transplantation may not be able to fully achieve the expected therapeutic effect.10,11 Therefore, modification of the exogenous cellular or CNS environment is necessary to overcome unfavourable environmental factors and reduce the level of apoptosis after cell transplantation. Primary SCs are difficult to obtain and have a limited capacity for expansion in culture. Immortalised Schwann cells (iSCs) provide a renewable and consistent cell source for research and potential therapy.
Cross-regulation between the NF-κB and Wnt/β-catenin signalling pathways plays an important role in the regulation of inflammation, immunity, development and oncogenesis.12,13 The NF-κB signalling pathway has a significant regulatory role in inflammatory responses, with NF-kB translocating to the nucleus to stimulate the transcription of cellular inflammatory factors.14,15 The Wnt/β-Catenin signalling pathway is a typical Wnt signalling pathway that plays an important role in neural development, axon guidance, neuropathic pain relief and neuronal survival.16,17 The Wnt/β-Catenin signalling pathway has either inhibitory or facilitatory effects on the activity of NF-κB in different cell types through different mechanisms.18 Wnt3a is a significant member of the Wnt family and is expressed during central nervous system development. Through the Wnt/β-Catenin signalling pathway, Wnt3a can regulate spinal neural stem cells differentiation and proliferation, inhibit neuronal death, and promote spinal axonal conduction and regeneration.19–21 However, it is unclear whether Wnt3a can regulate inflammatory responses through NF-κB.
This study first constructed rat immortalised Schwann cells (iSCs) to provide a scalable cell model for subsequent cell model construction. A cellular inflammation model was then created to investigate whether Wnt3a regulates the inflammatory response through NF-κB, as well as regulating apoptosis, proliferation and cellular function, providing potential therapeutic ideas for SCI.
Methods and Materials
Extraction, Culture and Immortalization of Primary Schwann Cells
Bilateral sciatic nerves were aseptically dissected from neonatal 24-hour-old rats following euthanasia via cervical dislocation and sterilization with alcohol. The nerves were minced and digested with 0.25% trypsin (BIOAGRIO, China). An equal volume of complete medium (DMEM (Gibco, USA) supplemented with 10% FBS (Gibco, USA) and 1% penicillin–streptomycin (Solarbio, China)) was added to terminate digestion, followed by filtration. The filtrate was centrifuged at 1000 rpm for 10 min, and the supernatant was discarded. The pellet was resuspended in complete medium and plated into culture dishes. Cells were maintained at 37 °C in a 5% CO2 incubator. To eliminate fibroblasts, 5 μg/mL cytarabine (MedChemExpress, USA) was added after 24 hours. Schwann cell purity was confirmed by S100β immunofluorescence. All cultures were routinely tested and verified to be free of mycoplasma contamination.
Primary Schwann cells were infected with a HEK293-packaged retrovirus containing a plasmid with the immortalisation gene SV40Tag, the hygromycin resistance gene and the green fluorescent protein gene GFP. After 24 hours of transfection, the medium was replaced with complete medium and the medium was changed every 3 days at half the amount. Upon reaching 80–90% confluence, the cells were passaged sequentially for 6–7 generations. Immortalized Schwann cells (iSCs) were then selected by culturing in medium supplemented with 0.1 g/L hygromycin for 3 days. All cell cultures were maintained at 37 °C in a 5% CO2 atmosphere.
Construction and Grouping of Cellular Inflammation Models
Cells were treated as follows: the LPS group was stimulated with 100 ng/mL LPS (Sigma, China) to induce inflammation; the Wnt3a group received 50 ng/mL recombinant Wnt3a protein(Med Chem Express, USA), which acts by binding to Frizzled and LRP5/6 co-receptors on the cell surface to activate downstream Wnt/β-catenin signaling; and the control group was maintained in complete medium. NF-κB expression was inhibited using BAY 11–7082 (MedChemExpress, USA).
Immunofluorescence
SCs were plated in confocal dishes at a density of 4×103 cells per well. When cells reached 60% confluence, the medium was removed. Cells were fixed with 4% paraformaldehyde for 15 min, permeabilized with 0.3% Triton X-100 for 15 min, and blocked with 5% fetal bovine serum for 30 min. Subsequently, the cells were incubated with primary antibodies at 4°C overnight, followed by incubation with secondary antibodies for 1.5 hours at room temperature protected from light. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Beyotime, China) for 10 min at room temperature. Images were acquired using a confocal microscope (Nikon C2, Japan). The same protocol was applied to detect Sox10 (1:2000, Proteintech, China) expression in both iSCs and Schwann cells.
Quantitative Real-Time PCR
Total RNA was extracted by The SteadyPure Quick RNA Extraction Kit (Accurate Biology, China) and reverse transcription was performed to synthesise cDNA. The Bio-Rad CFX96 instrument was used to perform RT-qPCR experiments. The reaction system was set to 95°C for 3 minutes, followed by 40 cycles of 95°Cfor 15seconds, 60°C for 30seconds, and 72°Cfor 30seconds. Gene expression was analysed using the 2−ΔΔCT method.22 The primers used are listed in Table 1.
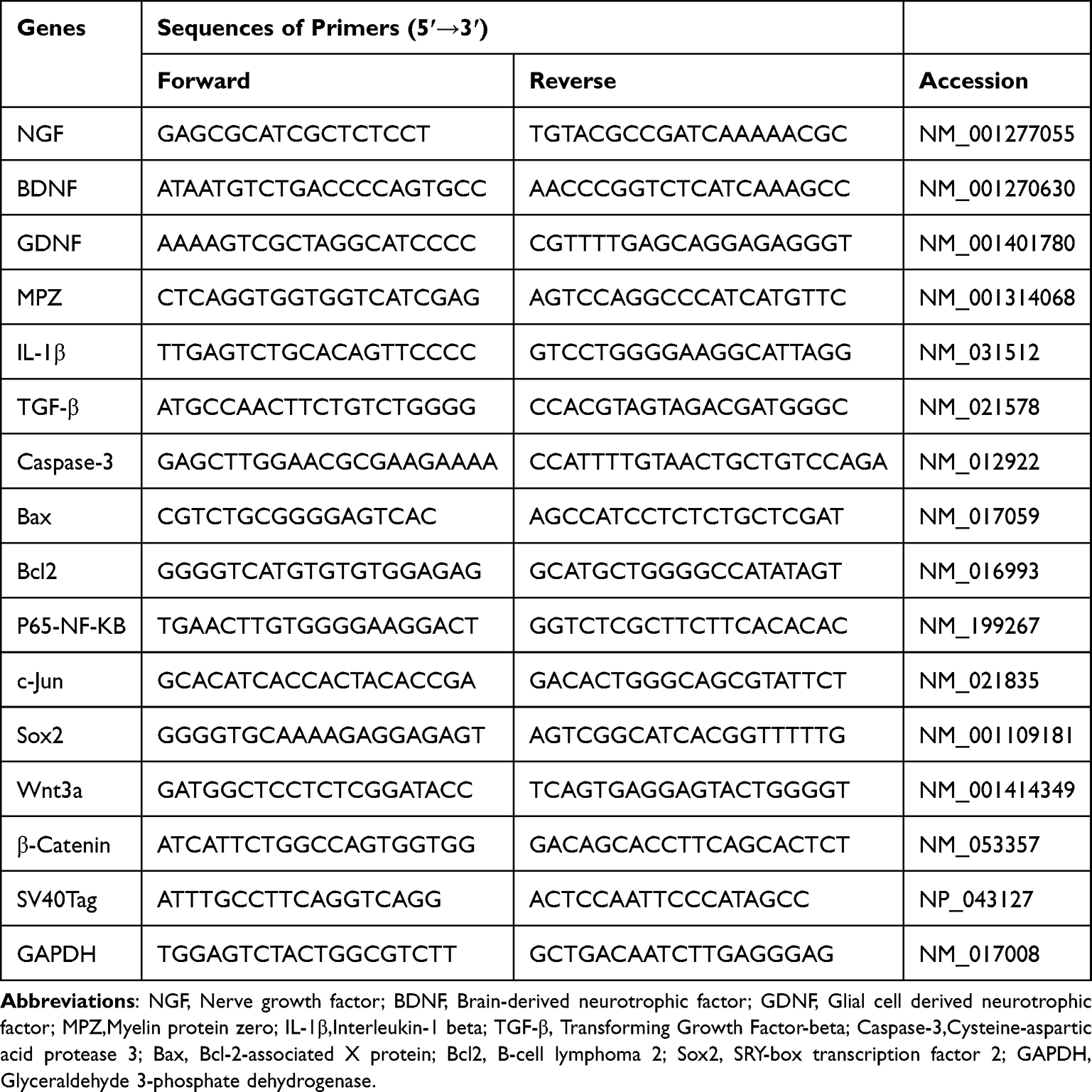 |
Table 1 Primer Sequences for RT-qPCR |
Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
The cDNAs of iSCs and SCs were amplified by RT-PCR, and the primer sequences are shown in Table 2. The reaction conditions were 98°C for 5 min, and the following 35 cycles consisted of 95°Cfor 30s, 55°C for 30s, and 72°C for 2 min. Afterwards, the mixtures were held at 72°C for 7 min, and finally stored at 4°C. The amplified products were analysed by agarose gel electrophoresis and ethidium bromide staining.
 |
Table 2 Primer Sequences for RT-PCR |
Western Blot
Total protein was extracted using RIPA lysis buffer (Beyotime, China) according to the manufacturer’s instructions. Protein concentration was determined with a BCA assay kit (Beyotime, China). Proteins were separated by 10% SDS-PAGE and transferred onto PVDF membranes (Millipore, IPVH00010). After blocking with 5% skim milk, the membranes were incubated with primary antibodies at 4°C overnight, followed by incubation with horseradish peroxidase-conjugated anti-rabbit or anti-mouse secondary antibodies for 2 hours at room temperature. Protein bands were visualized using an ECL system (BIO-RAD). The ratio of target protein to GAPDH was analyzed using Image Lab software (USA) to determine relative expression levels. Antibodies and their dilutions are listed in Table 3.
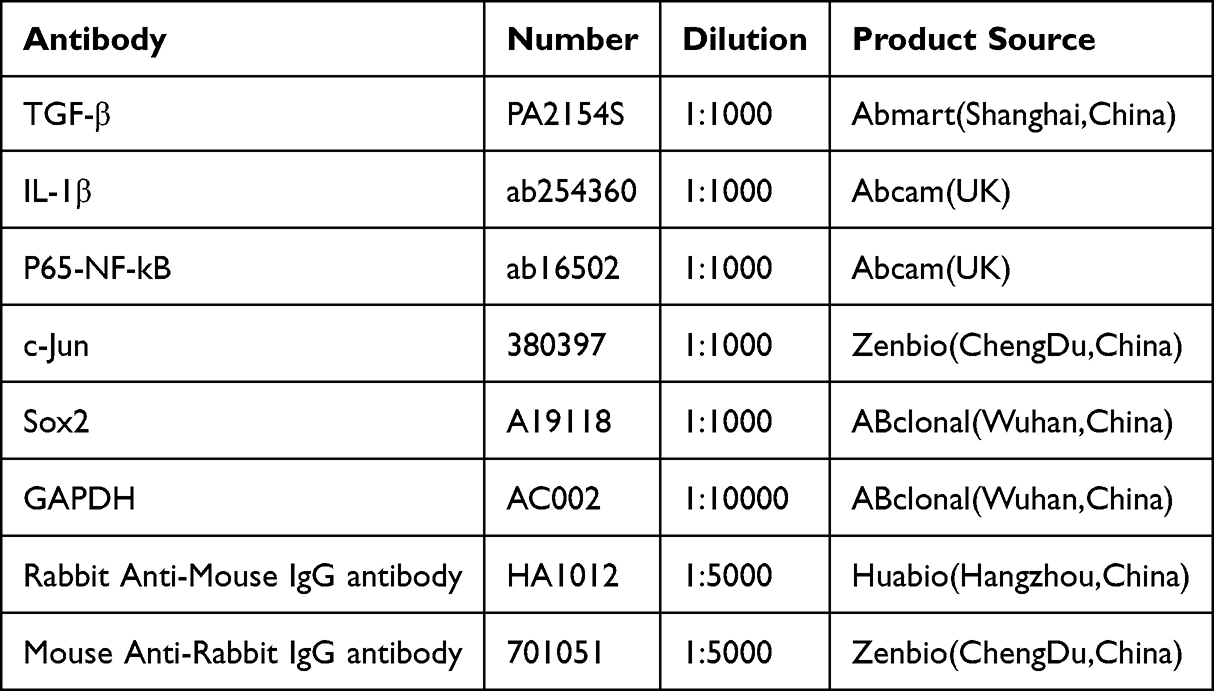 |
Table 3 Antibodies and Dilution Concentrations |
Cell Proliferation Assay
Cells were seeded in 96-well plates at a density of 2×103 cells per well and cultured for 24 hours. The medium was then replaced with 100 μL of serum-free medium. Subsequently, 10 μL of CCK-8 solution (Solarbio, China) was added to each well, followed by incubation at 37°C for 2 hours. The absorbance at 450 nm was measured using a microplate reader (BioTek, USA).
Scratch Test
The cells’ migratory capacity was assessed using a cell scratch test. After being separated with trypsin, the cells were placed in 6-well plates at a concentration of 1×105 cells per well. Once the cells reached 90% confluence, a 1 mL sterile pipette tip was used to create a uniform and flat scratch by scratching the six-well plate vertically. Wound healing was observed under the microscope at 0 and 24 hours, respectively, and photographs were taken to record the process. The wound closure rate was calculated using Image J software.
TUNEL Staining
The iSCs were inoculated into confocal dishes and cells were treated using the One-Step TUNEL Flow Cytometry Apoptosis Kit (Elabscience Biotechnology, China). After staining, the photographic conditions of the confocal microscope were adjusted according to the positive and negative control groups, and the treated groups were photographed under the same conditions.
Flow Cytometry
Cell apoptosis was assessed using an Annexin V-PE/7-AAD Apoptosis Detection Kit (MeilunBio, China) according to the manufacturer’s protocol. Briefly, cells were seeded in 6-well plates, subjected to the indicated treatments, harvested, washed with PBS, and resuspended in binding buffer at a density of 1×106 cells/mL. The cell suspension was stained with 5 μL Annexin V-PE and 5 μL 7-AAD, followed by incubation in the dark at room temperature for 15 min. Apoptotic cells were defined as Annexin V⁺/7-AAD− (early apoptotic) and Annexin V⁺/7-AAD⁺ (late apoptotic), and analyzed by flow cytometry within 1 hour.
Experimental Animals
The study used 20 female wild-type Sprague-Dawley rats weighing between 200–220 g, purchased from the Animal Experimentation Centre of Chongqing Medical University. The experimental protocols were approved by the China Association for Assessment and Accreditation of Laboratory Animal Care International, and all methods followed the ARRIVE guidelines. The rats were randomly divided into two groups: iSCs treatment (n = 10) and Wnt3a+iSCs treatment (n = 10). The animals were kept in a room with a temperature range of 22 to 26°C, a relative humidity of 40–60%, and a light/dark cycle of 12/12 hours.
Acute Spinal Cord Injury Model
A rat model of spinal cord injury (SCI) was established using a modified Allen’s method. Briefly, rats were anesthetized with sodium pentobarbital (10 g/L, 40 mg/kg; Sigma, USA). After hair removal and disinfection over the T9–T11 vertebrae, a dorsal midline incision was made, and the T10 lamina was removed to expose the spinal cord. A 10-g weight was dropped from a height of 3 cm through a guide tube to impact the spinal cord. Successful injury was confirmed by immediate tail flicking or curling. For treatment, the iSC group received an intraspinal injection of PBS containing 1×106 iSCs, while the Wnt3a+iSC group received PBS with 50 ng/mL Wnt3a and 1×106 iSCs. After hemorrhage control, muscle and skin layers were sutured sequentially. Rats were placed on a heating pad until fully awake and then returned to their cages with free access to food and water. Manual bladder expression was performed twice daily.
Frozen Slices
On day 2 after SCI, the experimental rats were anaesthetised. The chest cavity was opened to perfuse saline from the heart to remove blood from the tissue. Subsequently, spinal cord tissues were collected from the surgical area and fixed in paraformaldehyde solution for 24 h at 4°C. Each spinal cord tissue was dehydrated in 20% sucrose solution for 12 h, embedded in OCT medium, and stored at −80°C. Serial sagittal sections, 7 μm in thickness, were prepared using cryosectioning (Leica) and observed under a fluorescence microscope.
Statistical Analysis
All data were analysed using GraphPad Prism 9 software (San Diego, CA, USA) and results are expressed as mean ± standard deviation. Differences between groups were compared using two-way ANOVA or one-way ANOVA, followed by Tukey’s post-hoc test for multiple comparisons. P< 0.05 was considered statistically significant.
Results
Immortalisation of Rat Schwann Cells
After 7 days of culture, primary SCs were observed under a microscope. The cells gradually took on the form of small bipolar or tripolar elongated shapes arranged in a reticulated or swirling pattern (Figure 1A). Confocal microscopy revealed that the cells were small, shuttle-shaped, or bipolar. Upon staining with S100β antigen, the cytoplasm of the cells appeared green and the nucleus appeared blue (Figure 1B). It is known that the primary SCs can be successfully cultured in this experiment.
 |
Figure 1 Characterization of primary and immortalised Schwann cells. (A) Cell morphology of SCs, scale bar 100 μm; (B) Confocal image of S100β immunofluorescence staining of Schwann cells, microscope magnification 200x, scale bar 50 μm; (C) Cell morphology of iSCs(at passage 6–7), scale bar 100μm; (D) Agarose gel electrophoresisof SV40Tag PCR products; (E) Results of RT-qPCR assay of SV40Tag gene expression level (****P<0.0001). Data are presented as mean ± SD (n=3). |
Primary SCs at passage 2 (P2) were transfected with SV40T-Ag and cultured for 10 consecutive generations. Hygromycin was used to select for an immortalised Schwann cell line. iSCs exhibited a spindle-shaped morphology with slender processes, the majority being bipolar and a minority multipolar. (Figure 1C).
RNA was extracted from 12th generation iSCs and primary cells for agarose gel electrophoresis (Figure 1D) and RT-qPCR analysis (Figure 1E), respectively. The primary cells did not express SV40T-Ag, whereas the immortalised Schwann cell line stably expressed SV40T-Ag(P<0.01), indicating successful construction of the immortalised Schwann cell line.
Comparison of Primary and Immortalised Cells
The maintenance and regeneration of Schwann cells (SCs) are closely associated with Sox10 expression. In this study, immunofluorescence staining was performed to detect Sox10 expression in both primary SCs and immortalized SCs (iSCs). Following staining, the cytoplasm of both cell types exhibited red fluorescence, while the nuclei were counterstained blue (Figure 2A). Anti-Sox10 positivity was observed in both primary and immortalized cells.
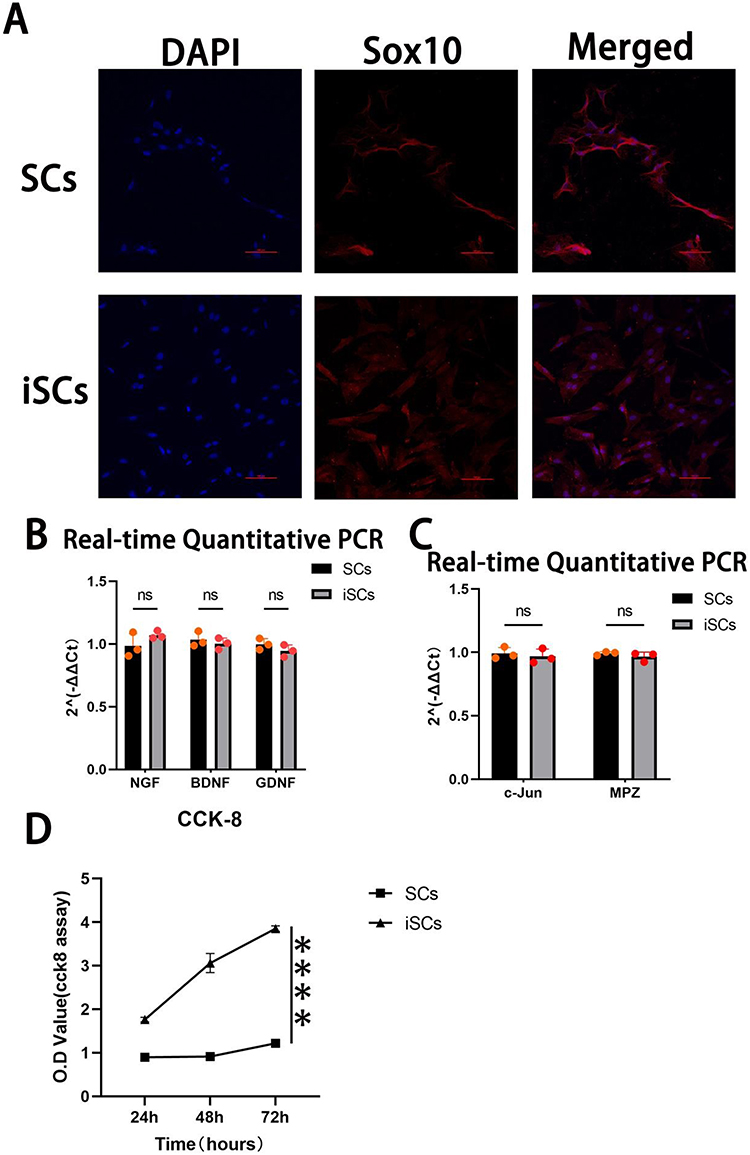 |
Figure 2 Comparison of primary SCs and iSCs. (A)Immunofluorescence staining of Sox10 in SCs and iSCs, microscope magnification 100x, scale bar 50μm; (B) and (C)gene expression levels of neurotrophic factors (B) and myelin-associated genes (C) in SCs and iSCs; (D)CCK-8 cell proliferation assay.(n=3) (`ns` for not significant, `****` for P<0.0001) Data are presented as mean ± SD (n=3). |
The expression levels of NGF, BDNF, and GDNF were assessed in primary SCs and iSCs using RT-qPCR. The results revealed no significant differences in the expression of these genes between the two cell types (Figure 2B). Additionally, the expression of c-Jun and MPZ, two key genes involved in myelin formation, was examined. Similarly, no significant differences were detected between iSCs and primary SCs (Figure 2C). These findings indicate that iSCs are genotypically comparable to primary SCs and suggest their suitability for neural-related research.
Furthermore, a CCK-8 assay was conducted to evaluate cell proliferation rates. The results demonstrated that the proliferation rate of iSCs was significantly higher than that of primary SCs (Figure 2D).
Wnt3a Inhibits LPS-Induced Inflammatory Response in iSCs
The iSCs were stimulated with induction medium containing 100 ng/mL LPS, and the expression of inflammatory factors was assessed via RT-qPCR at different time points. The results showed that, compared to the control group, the expression levels of IL-1β and TGF-β were altered to varying degrees after 24 h, 48 h, and 72 h of LPS treatment (Figure 3A and B). The highest expression of IL-1β was observed at 24 h, which was therefore selected as the optimal intervention time for subsequent experiments. Furthermore, LPS treatment led to a reduction in Wnt3a expression, accompanied by downregulation of β-catenin (Figure 3C and D).
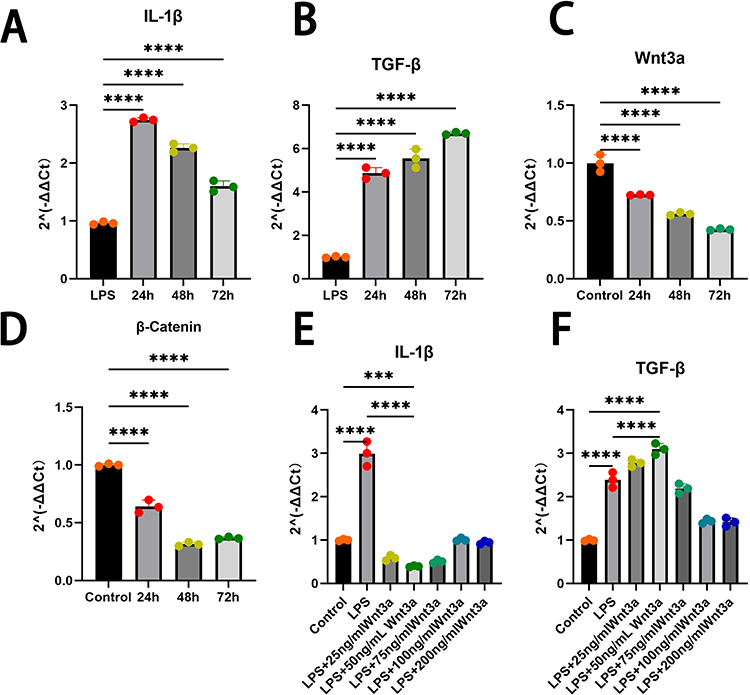 |
Figure 3 Wnt3a modulates LPS-induced inflammatory response in iSCs. (A) and (B):After LPS induction for different times, the expression levels of IL-1β and TGF-β were increased in the 24h, 48h and 72h groups compared to the Control group; (C) and (D) LPS inhibited the expression of Wnt3a and β-Catenin; (E) and (F) LPS-induced inflammatory response regulated by Wnt3a, which modulates IL-1β and TGF-β expression.(`ns` for not significant, `*` for P<0.05, `**` for P<0.01, `***` for P<0.001, `****` for P<0.0001) Data are presented as mean ± SD (n=3). |
In parallel, different concentrations of Wnt3a protein were applied for 24 h alongside LPS to evaluate its effect on the inflammatory response. qRT-PCR analysis revealed that Wnt3a treatment differentially inhibited IL-1β expression and enhanced TGF-β expression compared to the LPS-alone group (Figure 3E and F). Notably, treatment with 50 ng/mL Wnt3a recombinant protein significantly increased the anti-inflammatory factor TGF-β while markedly decreasing the pro-inflammatory factor IL-1β. Accordingly, 50 ng/mL was chosen as the effective concentration for further experiments.
Wnt3a Inhibits LPS-Induced Apoptosis and Promotes Cell Proliferation and Migration
TUNEL staining revealed a significantly higher proportion of apoptotic cells in the LPS group compared to the Control group. In contrast, the Wnt3a group (treated with 50 ng/mL Wnt3a) exhibited a marked reduction in TUNEL-positive cells relative to the LPS group, indicating that Wnt3a suppresses LPS-induced apoptosis (Figure 4A and B). Flow cytometry analysis further confirmed that Wnt3a significantly decreased the overall proportion of apoptotic cells, including a notable reduction in early apoptotic cells (Annexin V⁺/7-AAD−).
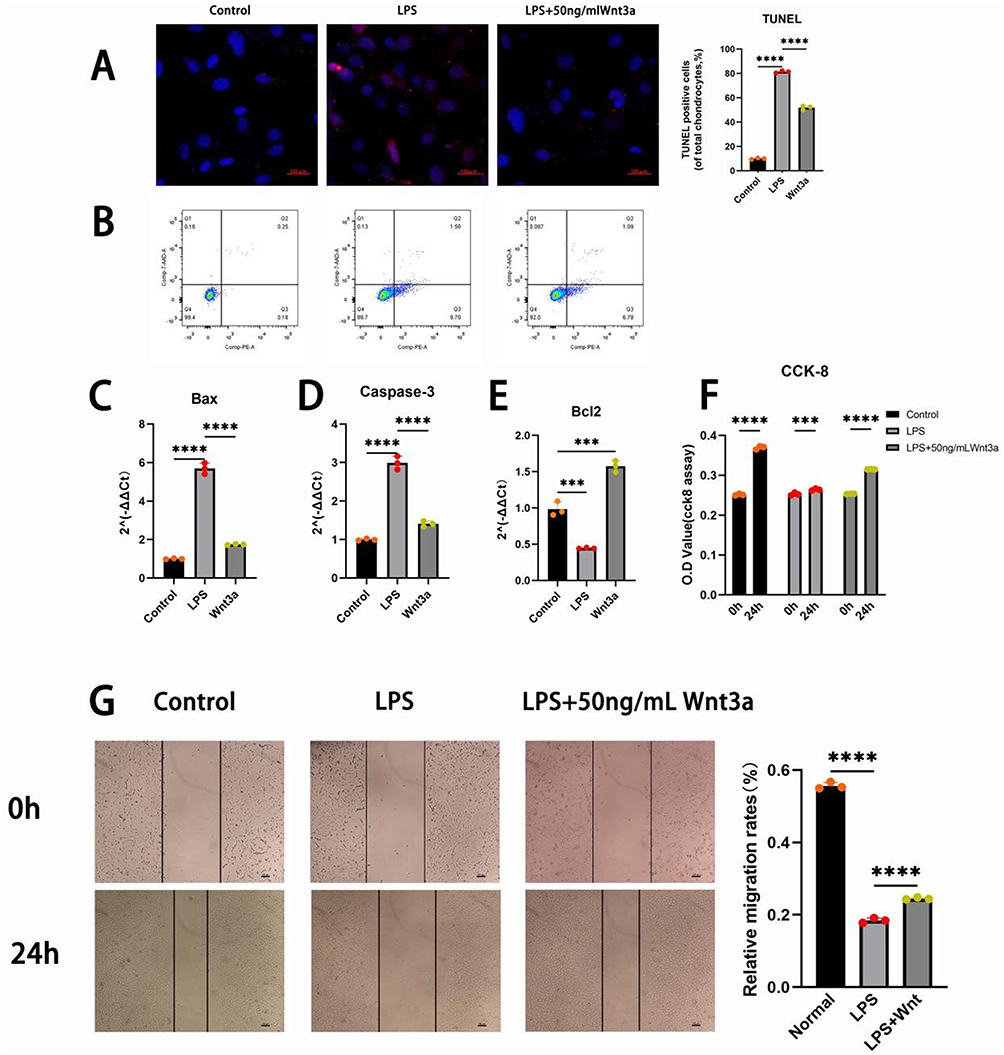 |
Figure 4 Wnt3a inhibits apoptosis and promotes proliferation and migration in iSCs under inflammatory conditions. (A) TUNEL assay, microscope magnification 100x, scale bar 150μm; (B) Apoptosis detection by flow cytometry; (C–E) Expression levels of apoptosis-related genes Bax, Caspase3, and Bcl2 measured by qPCR; (F) CCK-8 cell proliferation assay; (G) Wound healing assay and quantitative analysis of the results.(`*` for P<0.05, `**` for P<0.01, `***` for P<0.001, `****` for P<0.0001) Data are presented as mean ± SD (n=3). |
At the molecular level, LPS stimulation significantly upregulated the expression of Bax (Figure 4C) and activated Caspase-3 (Figure 4D), while downregulating the Bcl-2 (Figure 4E). Wnt3a treatment counteracted these effects, reducing Bax and cleaved Caspase-3 levels and increasing Bcl-2 expression (Figure 4C–E), suggesting that Wnt3a modulates apoptosis-related proteins to inhibit cell death.
CCK-8 assays indicated that LPS exposure significantly impaired iSC proliferation compared to the Control. However, supplementation with 50 ng/mL Wnt3a restored the proliferation rate, indicating that Wnt3a promotes iSC proliferation even under inflammatory conditions (Figure 4F).
Additionally, a scratch assay demonstrated that LPS inhibited cell migration compared to the Control, whereas Wnt3a significantly enhanced scratch closure (P < 0.01), confirming its promotive effect on cell migration (Figure 4G).
Wnt3a Enhances Neural Repair in iSCs Under Inflammatory Environment
The transcription factors c-Jun and Sox2 play crucial roles in the proliferation, differentiation, and myelin formation of SCs. In the LPS-induced inflammatory response (Figure 5), the expression of c-Jun and Sox2 was suppressed, which may have contributed to the slowdown of proliferation in iSCs. However, when treated with 50 ng/mL Wnt3a, the expression levels of c-Jun and Sox2 increased compared to the LPS group (Figure 5). This suggests that Wnt3a can enhance the neural repair function of iSCs.
 |
Figure 5 Wnt3a enhances expression of c-Jun and Sox2 in iSCs under inflammatory conditions. (A) Relative mRNA expression of c-Jun. (B) Relative mRNA expression of Sox2. (C) Protein expression changes of c-Jun and Sox2 (`**` for P<0.01, `***` for P<0.001, `****` for P<0.0001) Data are presented as mean ± SD (n=3). |
Wnt3a Regulates LPS-Induced Inflammatory Responses in iSCs Through the NF-kB Signalling Pathway
The specific inhibition of the NF-κB pathway by Bay 11–7082 has been extensively studied. Various concentrations of Bay 11–7082 were used to intervene with iSCs cells for 1 hour before inducing the inflammatory response. The expression of NF-kB was inhibited by Bay 11–7082 (Figure 6A). The strongest inhibitory effect on NF-kB was observed with 2 μM of Bay 11–7082, which was chosen as the experimental concentration for all subsequent experiments.
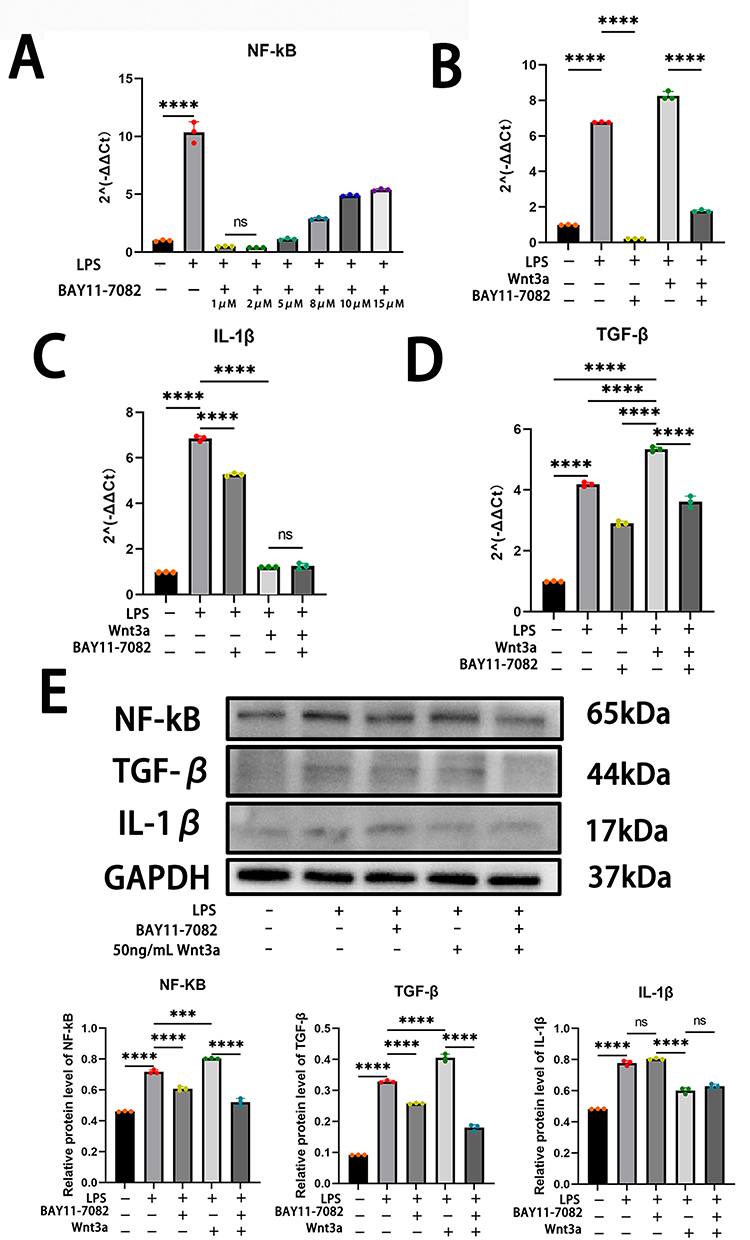 |
Figure 6 Wnt3a regulates inflammatory response via NF-κB signaling pathway. (A):BAY11-7082 produces significant inhibition of NF-κB after two hours of treatment(Ordinary one-way ANOVA, F(7,8),P<0.0001); (B–E) Wnt3a enhances the NF-κB signalling pathway to promote TGF-β expression. Additionally, the inhibition of IL-1β by Wnt3a does not involve the NF-κB signalling pathway.(`ns` for not significant, `*` for P<0.05, `**` for P<0.01, `***` for P<0.001, `****` for P<0.0001) Data are presented as mean ± SD (n=3). |
To investigate the potential molecular mechanism of Wnt3a on LPS-induced inflammatory response, the study evaluated the expression level of NF-kB in iSCs. The mRNA expression level of NF-kB was detected by using RT-qPCR, and the results showed that the expression of NF-kB increased significantly after 24 hours of LPS stimulation (Figure 6B). Then, Western Blot experiments at the protein level observed that the NF-kB expression level was also increased in the LPS group compared to the control group (Figure 6E). Furthermore, following the intervention with Wnt3a, it was observed that the activation level of NF-kB was once again enhanced (Figure 6B–E). This suggests that Wnt3a may regulate the inflammatory response by promoting the activation of NF-kB.
This was followed by exploring how Wnt3a inhibits the inflammatory response by modulating NF-kB. RT-qPCR was used to examine IL-1β and TGF-β mRNA levels. The results indicate that the expression level of IL-β decreased with the use of Bay11-7082 or Wnt3a alone. However, Wnt3a caused a more significant decrease in the expression level of IL-1β (Figure 6C). When both BAY11-7082 and Wnt3a were used, the expression level of IL-1β decreased compared to the LPS group. However, there was no significant change in IL-1β expression compared to when Wnt3a was used alone (Figure 6C). This suggests that the inhibition of IL-1β expression by Wnt3a was not due to the modulation of the NF-kB signalling pathway. After inhibiting NF-κB with Bay11-7082, the expression level of TGF-β decreased. However, when Wnt3a was introduced, the expression level of TGF-β increased (Figure 6D). When BAY11-7082 and Wnt3a were used together, the expression level of TGF-β decreased compared to Wnt3a alone (Figure 6D). This suggests that Wnt3a enhances the expression of TGF-β by increasing the activation of NF-kB. The validation was repeated using Western Blot, which yielded the same conclusion as RT-qPCR (Figure 6E).
7, Wnt3a increases survival of iSCs in injured spinal cord
The study investigated the effect of Wnt3a on reducing cell death in transplanted cells in animals. The study constructed spinal cord injury model using SD rats (Figure 7A). The iSCs solution was injected at the injury site with or without Wnt3a. After 24 hours, spinal cord frozen sections were viewed under a fluorescence microscope. Both the iSCs treatment and Wnt3a+iSCs treatment groups showed green fluorescence indicating the presence of iSCs cells. The number of cells in the Wnt3a+iSCs treatment group was higher than that in the iSCs treatment group (Figure 7B). Additionally, there was migration of iSCs cells in the damaged spinal cord, and some transplanted cells were observed outside the injury site (Figure 7B). These findings suggest that transplanted iSCs can survive in the injured spinal cord, and that the intervention of Wnt3a enhances their survival.
 |
Figure 7 Wnt3a enhances survival of transplanted iSCs in a rat model of spinal cord injury. Survival of transplanted cells in the rat spinal cord 24 hours after spinal cord injury. (A) Construction of SCI model and cell transplantation: (i):Exposure of spinal cord on T9-T10 after resection of the laminae; (ii) Cell transplantation is performed on the spinal cord after an injury. (B) Fluorescence microscopic observation of frozen sections of spinal cord tissue:(i) iSCs treatment Group; (ii) Wnt3a+iSCs treatment Group; microscope magnification 100x, scale bar 100μm. |
Discussion
Regenerative therapies with cell transplantation have been promising in the treatment of SCI.23 As a type of glial cells, SCs drive axon regeneration and myelin formation in the PNS and perform similar functions when transplanted into the spinal cord.24,25 SCs are involved in degrading the damaged myelin at an early stage after SCI,26 and SCs-secreted growth factors, extracellular components, and adhesion molecules are able to promote damaged axon regeneration.27,28 More importantly, demyelinated axons in the CNS can also be remyelinated by transplantation of SCs to improve conduction.29,30 However, obtaining primary SCs is a challenging task. These cells can be obtained from the sciatic nerve of neonatal rats, and their purity can be improved by removing fibroblasts with cytarabine.31 However, the obtained SCs cannot be passaged multiple times, and it is time-consuming to perform primary cell extraction and culture multiple times. In this study, immortalized Schwann cells were created through transfection with SV40Tag. The results showed significant genetic similarity between iSCs and SCs. SCs secrete various trophic factors, such as BDNF, NGF, and GDNF,32–34 which are crucial for maintaining neuronal function and repair. Additionally, the transcription factor c-Jun and Myelin Protein Zero (MPZ) expressed by SCs play vital roles in myelin formation.35,36 The study found no significant difference in gene expression levels between iSCs and SCs. After immortalisation, SCs did not lose their original functions, and iSCs have the same potential neurorestorative ability as SCs. More importantly, the ability of iSCs to undergo multiple passages and rapid proliferation facilitated the study. Secondly, while SV40Tag expression confirms immortalization, further validation through karyotyping or telomerase activity assays would strengthen the conclusion of stable immortalization.
Following spinal cord injury (SCI), imbalances in the microenvironment lead to low cell survival rates in the transplanted cells within the injured spinal cord region.37,38 While the acute phase of SCI is considered the optimal period for cell transplantation,39 this period is also associated with the most severe imbalanced microenvironment due to the inflammatory response and oxidative stress.40,41 Therefore, in addition to modifying the transplanted cells, how to improve the inflammatory response after spinal cord injury is a critical issue that needs to be urgently addressed.
Both the Wnt/β-catenin and NF-κB signalling pathways play a crucial role in mammalian development.42,43 While the Wnt/β-catenin signalling pathway is essential for development and tissue regeneration, NF-κB is primarily involved in regulating inflammatory responses.18 Research has demonstrated that β-catenin decreases the enzymatic activity of cAMP-responsive element-binding protein (CREB)-binding protein (CBP), which specifically inhibits the expression of pro-inflammatory subpopulations of NF-κB target genes.44 Then Wnt3a, as an activator of the Wnt/β-catenin signalling pathway,45,46 has potential research value for the regulation of inflammatory responses. The study demonstrated that Wnt3a enhances NF-κB activation, promotes the expression of the anti-inflammatory factor TGF-β, and inhibits the expression of IL-1β. Inhibition of the NF-κB signalling pathway by BAY11-7082 did not affect the expression level of IL-1β. Instead, the decrease in NF-κB activation led to a reduction in TGF-β expression. The findings suggest that Wnt3a did not regulate IL-1β through the NF-κB signalling pathway. The findings suggest that Wnt3a did not regulate IL-1β through the NF-κB signalling pathway. The association between elevated TGF-β expression levels and NF-κB signalling pathway activation was observed. Further exploration is needed to determine if the decrease in IL-1β is due to the high expression of TGF-β.
Apoptosis is closely related to tissue destruction and cellular stress. It is a strategy used by the body to control excessive inflammatory responses.47,48 Excessive apoptosis can cause nerve damage and further induce sensory and motor impairments.49 Research has shown that Wnt-3a can improve functional recovery by activating autophagy and inhibiting the mTOR signalling pathway after SCI.21,50 In this study, Wnt3a reduced apoptosis in the inflammatory response by regulating the expression of pro-apoptotic protein Bax and anti-apoptotic protein Bcl2. This suggests that Wnt3a may be a potential regulator of the inflammatory response. Based on this, a spinal cord injury model in SD rats was constructed in the study and validated in vivo in animals by injecting cytosol with iSCs with or without 50 ng/mL Wnt3a. The addition of Wnt3a significantly improved the survival of transplanted cells compared to the control group without Wnt3a. This finding provides crucial evidence for the potential use of cell transplantation as a therapeutic intervention during the acute phase of spinal cord injury.
Furthermore, in an inflammatory environment, the expression levels of c-Jun and Sox2 in iSCs decrease, which is unfavourable for promoting myelin tissue regeneration. However, the intervention of Wnt3a promotes the expression of both genes, which has greater significance for the application of Wnt3a to promote myelin repair in the injured spinal cord.
It is important to note that this study has some limitations. Firstly, only iSCs were selected for certain studies, while other cell types exist in the nervous system. Secondly, the mechanism of Wnt3a inhibiting the inflammatory response was not extensively explored in the study. The study has some limitations that need to be addressed. Cell transplantation experiments will be conducted in the future to investigate the role of Wnt3a and iSCs combination therapy in nerve repair for spinal cord injuries. This will provide additional treatment strategies for SCI.
In conclusion, this study aimed to facilitate the development of experiments. To achieve this, iSCs were constructed to avoid the inconvenience caused by multiple extractions of primary cells. An immortalised cell line with high similarity to primary cells was obtained. Secondly, our cellular experiments revealed that Wnt3a can regulate inflammatory responses by modulating the NF-kB signalling pathway. Additionally, it can inhibit apoptosis, promote cell proliferation, and improve the function of iSCs in inflammatory environments. Finally, we demonstrated that Wnt3a can enhance the neural repair function of iSCs.
Conclusion
Our study demonstrated that Wnt3a modulates inflammatory responses, reduces apoptosis, promotes cell proliferation and migration, and enhances cell function. The results suggest the potential use of iSCs in treating spinal cord injuries, and highlight the regulatory role of Wnt3a in modulating inflammatory responses and promoting neural recovery. Therefore, this study proposes a new cellular regimen for a cell transplantation therapeutic approach for spinal cord injury and provides further evidence for the use of Wnt3a in SCI treatment.
Data Sharing Statement
The data used in the study analyses can be made available by the corresponding author on reasonable request.
Ethics Statement
All animal procedures were conducted in accordance with the Basel Declaration and approved by the Ethics Committee of Chongqing Medical University (approval no. CHCMU-IACUC20230117002, approval date: 17 January 2023).
Acknowledgments
We are grateful to all study participants for their contributions.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors have no potential or existing conflicts of interest to declare.
References
1. Liu X, Jiang X, Yu Q, et al. Sodium alginate and naloxone loaded macrophage-derived nanovesicles for the treatment of spinal cord injury. Asian J Pharm Sci. 2022;17(1):87–101. doi:10.1016/j.ajps.2021.11.001
2. Kumamaru H, Lu P, Rosenzweig ES, Kadoya K, Tuszynski MH. Regenerating corticospinal axons innervate phenotypically appropriate neurons within neural stem cell grafts. Cell Rep. 2019;26(9):2329–2339e4. doi:10.1016/j.celrep.2019.01.099
3. Rodrigues P, Cassanego GB, Peres DS, et al. Alpha-lipoic acid reduces nociception by reducing oxidative stress and neuroinflammation in a model of complex regional pain syndrome type I in mice. Behav Brain Res. 2024;459:114790. doi:10.1016/j.bbr.2023.114790
4. Jessen KR, Arthur-Farraj P. Repair schwann cell update: adaptive reprogramming, EMT, and stemness in regenerating nerves. Glia. 2019;67(3):421–437. doi:10.1002/glia.23532
5. Horner SJ, Couturier N, Hafner M, Rudolf R. Schwann cells in neuromuscular in vitro models. Biol Chem. 2024;405(1):25–30. doi:10.1515/hsz-2023-0172
6. Fu H, Hu D, Chen J, et al. Repair of the injured spinal cord by schwann cell transplantation. Front Neurosci. 2022;16:800513. doi:10.3389/fnins.2022.800513
7. Chen CZ, Neumann B, Forster S, Franklin RJM. Schwann cell remyelination of the central nervous system: why does it happen and what are the benefits? Open Biol. 2021;11(1):200352. doi:10.1098/rsob.200352
8. Sunshine MD, Bindi VE, Nguyen BL, et al. Oxygen therapy attenuates neuroinflammation after spinal cord injury. J Neuroinflammation. 2023;20(1):303. doi:10.1186/s12974-023-02985-6
9. Li X, Fu J, Guan M, Shi H, Pan W, Lou X. Biochanin A attenuates spinal cord injury in rats during early stages by inhibiting oxidative stress and inflammasome activation. Neural Regen Res. 2024;19(9):2050–2056. doi:10.4103/1673-5374.390953
10. Hellenbrand DJ, Quinn CM, Piper ZJ, Morehouse CN, Fixel JA, Hanna AS. Inflammation after spinal cord injury: a review of the critical timeline of signaling cues and cellular infiltration. J Neuroinflammation. 2021;18(1):284. doi:10.1186/s12974-021-02337-2
11. He W, Li ZQ, Gu HY, et al. Targeted therapy of spinal cord injury: inhibition of apoptosis is a promising therapeutic strategy. Mol Neurobiol. 2023.
12. Guan X, He Y, Wei Z, et al. Crosstalk between Wnt/beta-catenin signaling and NF-kappaB signaling contributes to apical periodontitis. Int Immunopharmacol. 2021;98:107843. doi:10.1016/j.intimp.2021.107843
13. Li Y, Wang X, Lu J. Interleukin-35 promote osteogenesis and inhibit adipogenesis: role of Wnt/beta-Catenin and PPARgamma signaling pathways. Inflammation. 2023;46(2):522–533. doi:10.1007/s10753-022-01749-3
14. Sharma VK, Prateeksha P, Singh SP, Rao CV, Singh BN. Nyctanthes arbor-tristis bioactive extract ameliorates LPS-induced inflammation through the inhibition of NF-kappaB signalling pathway. J Ethnopharmacol. 2024;320:117382. doi:10.1016/j.jep.2023.117382
15. Chen Y, Lin W, Zhong L, et al. Bicyclol attenuates obesity-induced cardiomyopathy via inhibiting NF-kappaB and MAPK signaling pathways. Cardiovasc Drugs Ther. 2023;37(6):1131–1141. doi:10.1007/s10557-022-07356-6
16. Liu J, Xiao Q, Xiao J, et al. Wnt/beta-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Signal Transduct Target Ther. 2022;7(1):3. doi:10.1038/s41392-021-00762-6
17. Wu M, Dong H, Xu C, et al. The Wnt-dependent and Wnt-independent functions of BCL9 in development, tumorigenesis, and immunity: implications in therapeutic opportunities. Genes Dis. 2024;11(2):701–710. doi:10.1016/j.gendis.2023.03.012
18. Ma B, Hottiger MO. Crosstalk between Wnt/beta-Catenin and NF-kappaB signaling pathway during inflammation. Front Immunol. 2016;7:378. doi:10.3389/fimmu.2016.00378
19. Shen Z, Zhou Z, Gao S, et al. Melatonin inhibits neural cell apoptosis and promotes locomotor recovery via activation of the Wnt/beta-Catenin signaling pathway after spinal cord injury. Neurochem Res. 2017;42(8):2336–2343. doi:10.1007/s11064-017-2251-7
20. Lu P, Han D, Zhu K, Jin M, Mei X, Lu H. Effects of Sirtuin 1 on microglia in spinal cord injury: involvement of Wnt/beta-catenin signaling pathway. Neuroreport. 2019;30(13):867–874. doi:10.1097/WNR.0000000000001293
21. Gao K, Niu J, Dang X. Wnt-3a improves functional recovery through autophagy activation via inhibiting the mTOR signaling pathway after spinal cord injury. Neurosci Lett. 2020;737:135305. doi:10.1016/j.neulet.2020.135305
22. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
23. Xu B, Yin M, Yang Y, et al. Transplantation of neural stem progenitor cells from different sources for severe spinal cord injury repair in rat. Bioact Mater. 2023;23:300–313. doi:10.1016/j.bioactmat.2022.11.008
24. Monje PV, Deng L, Xu XM. Human schwann cell transplantation for spinal cord injury: prospects and challenges in translational medicine. Front Cell Neurosci. 2021;15:690894. doi:10.3389/fncel.2021.690894
25. Li J, Shangguan Z, Ye X, Wang Z, Liu W, Chen G. Modified FGF hydrogel for effective axon formation by enhanced regeneration of myelin sheath of schwann cells using rat model. Int J Nanomed. 2023;18:7225–7236. doi:10.2147/IJN.S417723
26. Arthur-Farraj PJ, Latouche M, Wilton DK, et al. C-Jun reprograms Schwann cells of injured nerves to generate a repair cell essential for regeneration. Neuron. 2012;75(4):633–647. doi:10.1016/j.neuron.2012.06.021
27. Bunge MB, Wood PM. Realizing the maximum potential of Schwann cells to promote recovery from spinal cord injury. Handb Clin Neurol. 2012;109:523–540.
28. Pan D, Zhu S, Zhang W, et al. Autophagy induced by Schwann cell-derived exosomes promotes recovery after spinal cord injury in rats. Biotechnol Lett. 2022;44(1):129–142. doi:10.1007/s10529-021-03198-8
29. Deng LX, Walker C, Xu XM. Schwann cell transplantation and descending propriospinal regeneration after spinal cord injury. Brain Res. 2015;1619:104–114. doi:10.1016/j.brainres.2014.09.038
30. Ghosh M, Pearse DD. Schwann cell-derived exosomal vesicles: a promising therapy for the injured spinal cord. Int J Mol Sci. 2023;24(24):17317. doi:10.3390/ijms242417317
31. Weinstein DE, Wu R. Isolation and purification of primary Schwann cells. Curr Protoc Neurosci Chapter. 2001;3.
32. Kim HS, Kim JY, Song CL, Jeong JE, Cho YS. Directly induced human Schwann cell precursors as a valuable source of Schwann cells. Stem Cell Res Ther. 2020;11(1):257. doi:10.1186/s13287-020-01772-x
33. Godinho MJ, Staal JL, Krishnan VS, et al. Regeneration of adult rat sensory and motor neuron axons through chimeric peroneal nerve grafts containing donor Schwann cells engineered to express different neurotrophic factors. Exp Neurol. 2020;330:113355. doi:10.1016/j.expneurol.2020.113355
34. Li X, Zhang T, Li C, et al. Electrical stimulation accelerates Wallerian degeneration and promotes nerve regeneration after sciatic nerve injury. Glia. 2023;71(3):758–774. doi:10.1002/glia.24309
35. Sohn EJ, Nam YK, Park HT. Involvement of the miR-363-5p/P2RX4 axis in regulating schwann cell phenotype after nerve injury. Int J Mol Sci. 2021;22(21):11601. doi:10.3390/ijms222111601
36. Gong X, Gui Z, Ye X, Li X. Jatrorrhizine ameliorates Schwann cell myelination via inhibiting HDAC3 ability to recruit Atxn2l for regulating the NRG1-ErbB2-PI3K-AKT pathway in diabetic peripheral neuropathy mice. Phytother Res. 2023;37(2):645–657. doi:10.1002/ptr.7641
37. Wang W, Huang X, Lin W, et al. Hypoxic preconditioned bone mesenchymal stem cells ameliorate spinal cord injury in rats via improved survival and migration. Int J Mol Med. 2018;42(5):2538–2550. doi:10.3892/ijmm.2018.3810
38. David BT, Curtin JJ, Brown JL, et al. Temporary induction of hypoxic adaptations by preconditioning fails to enhance Schwann cell transplant survival after spinal cord injury. Glia. 2023;71(3):648–666. doi:10.1002/glia.24302
39. Yan K, Zhang R, Sun C, et al. Bone marrow-derived mesenchymal stem cells maintain the resting phenotype of microglia and inhibit microglial activation. PLoS One. 2013;8(12):e84116. doi:10.1371/journal.pone.0084116
40. Luo S, Wu J, Qiu Y, et al. Hydrogen promotes the effectiveness of bone mesenchymal stem cell transplantation in rats with spinal cord injury. Stem Cells Int. 2023;2023:8227382. doi:10.1155/2023/8227382
41. Kaur J, Mojumdar A. A mechanistic overview of spinal cord injury, oxidative DNA damage repair and neuroprotective therapies. Int J Neurosci. 2023;133(3):307–321. doi:10.1080/00207454.2021.1912040
42. Song Y, Wu Z, Zhao P. The protective effects of activating Sirt1/NF-kappaB pathway for neurological disorders. Rev Neurosci. 2022;33(4):427–438. doi:10.1515/revneuro-2021-0118
43. Nejak-Bowen K, Monga SP. Wnt-beta-catenin in hepatobiliary homeostasis, injury, and repair. Hepatology. 2023;78(6):1907–1921. doi:10.1097/HEP.0000000000000495
44. Ma B, Fey M, Hottiger MO. WNT/beta-catenin signaling inhibits CBP-mediated RelA acetylation and expression of proinflammatory NF-kappaB target genes. J Cell Sci. 2015;128(14):2430–2436. doi:10.1242/jcs.168542
45. González-Fernández C, Fernández-Martos CM, Shields SD, Arenas E, Javier Rodríguez F. Wnts are expressed in the spinal cord of adult mice and are differentially induced after injury. J Neurotrauma. 2014;31(6):565–581. doi:10.1089/neu.2013.3067
46. Zhang S, Li J, Lea R, Vleminckx K, Amaya E. Fezf2 promotes neuronal differentiation through localised activation of Wnt/beta-catenin signalling during forebrain development. Development. 2014;141(24):4794–4805. doi:10.1242/dev.115691
47. Gao L, Dai C, Feng Z, Zhang L, Zhang Z. MiR-137 inhibited inflammatory response and apoptosis after spinal cord injury via targeting of MK2. J Cell Biochem. 2018;119(4):3280–3292. doi:10.1002/jcb.26489
48. Nano M, Montell DJ. Apoptotic signaling: beyond cell death. Semin Cell Dev Biol. 2024;156:22–34. doi:10.1016/j.semcdb.2023.11.002
49. Zhong ZX, Feng SS, Chen SZ, Chen ZM, Chen XW. Inhibition of MSK1 promotes inflammation and apoptosis and inhibits functional recovery after spinal cord injury. J Mol Neurosci. 2019;68(2):191–203. doi:10.1007/s12031-019-01298-9
50. Zhou Y, Xu J, Luo H, Meng X, Chen M, Zhu D. Wnt signaling pathway in cancer immunotherapy. Cancer Lett. 2022;525:84–96. doi:10.1016/j.canlet.2021.10.034
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Vitamin D Ameliorates Apoptosis and Inflammation by Targeting the Mitochondrial and MEK1/2-ERK1/2 Pathways in Hyperoxia-Induced Bronchopulmonary Dysplasia
Hu J, Wu Z, Wang H, Geng H, Huo J, Zhu X, Zhu X
Journal of Inflammation Research 2022, 15:4891-4906
Published Date: 25 August 2022
Eriochloa villosa Alleviates Progression of Benign Prostatic Hyperplasia in vitro and in vivo
Baek EB, Hwang YH, Park S, Hong EJ, Won YS, Kwun HJ
Research and Reports in Urology 2022, 14:313-326
Published Date: 24 September 2022
Engeletin Alleviates the Inflammation and Apoptosis in Intervertebral Disc Degeneration via Inhibiting the NF-κB and MAPK Pathways
Li B, Yang X, Zhang P, Guo J, Rong K, Wang X, Cao X, Zhou T, Zhao J
Journal of Inflammation Research 2022, 15:5767-5783
Published Date: 10 October 2022
PANoptosis: A Cell Death Characterized by Pyroptosis, Apoptosis, and Necroptosis
Shi C, Cao P, Wang Y, Zhang Q, Zhang D, Wang Y, Wang L, Gong Z
Journal of Inflammation Research 2023, 16:1523-1532
Published Date: 12 April 2023
Obacunone Promotes Functional Recovery After Spinal Cord Injury by Attenuating Neuroinflammation by Targeting the TLR4/MyD88/p38 MAPK Pathway
Kuang W, Zhang M, Zhang J, Huang H, Chen Q, Yu C, Peng B, Sun W, Lin J, Cheng J, Zhu L
Drug Design, Development and Therapy 2026, 20:577707
Published Date: 13 March 2026