Back to Journals » Therapeutics and Clinical Risk Management » Volume 21
Modulating Metabolic Health and Physiological Functions: Advances in Dietary Interventions Targeting Gut Microbiota
Authors Rabbani SA ![]() , El-Tanani M
, El-Tanani M ![]() , Janić M
, Janić M ![]() , Janež A, Tanani YE, Hajeer E, Matalka MI, Rizzo M
, Janež A, Tanani YE, Hajeer E, Matalka MI, Rizzo M ![]() , Kumar R
, Kumar R
Received 15 May 2025
Accepted for publication 24 November 2025
Published 13 December 2025 Volume 2025:21 Pages 1701—1733
DOI https://doi.org/10.2147/TCRM.S540144
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Garry Walsh
Syed Arman Rabbani,1 Mohamed El-Tanani,1 Miodrag Janić,2– 4 Andrej Janež,2,3 Yahia El Tanani,5 Eman Hajeer,6 Mohammad I Matalka,7 Manfredi Rizzo,1,4 Rakesh Kumar8,9
1RAK College of Pharmacy, RAK Medical and Health Sciences University, Ras Al Khaimah, United Arab Emirates; 2Department of Endocrinology, Diabetes and Metabolic Diseases, University Medical Centre Ljubljana, Ljubljana, Slovenia; 3Faculty of Medicine, University of Ljubljana, Ljubljana, Slovenia; 4PROMISE Department of Health Promotion Sciences Maternal and Infantile Care, Internal Medicine and Medical Specialties, School of Medicine, University of Palermo, Palermo, Italy; 5Royal Cornwall Hospital, NHS Trust, Treliske, Truro, UK; 6Faculty of Biology, Medicine and Health, University of Manchester, Manchester, UK; 7Department of Pathology and Microbiology, Faculty of Medicine, Jordan University of Science and Technology, Irbid, Jordan; 8Department of Pharmacy, Jagannath University, Bahadurgarh, Haryana, 124507, India; 9Amity Institute of Pharmacy, Amity University, Gurugram, Haryana, 122413, India
Correspondence: Syed Arman Rabbani; Miodrag Janić, Email [email protected]; [email protected]
Abstract: The human gut possesses a highly complex and metabolically functional microbial community. This microbial ecosystem, often termed a “super-organism”, plays a critical function in regulating the host’s metabolic processes, including gut motility, energy absorption, appetite, glucose and lipid metabolism, as well as hepatic fat storage. These metabolic functions of the gut microbiota (GM) play a central role in maintaining host homeostasis and overall metabolic health. This review synthesizes findings from recent clinical and preclinical studies, focusing on the interactions between gut microbiota, metabolic functions, and dietary interventions, to provide an evidence-based overview of current knowledge and future perspectives. Evidence was compiled through a narrative review of studies indexed in PubMed, Scopus, Web of Science, and Google Scholar using prespecified keywords related to gut microbiota, metabolic syndrome, diet, and dysbiosis.Recent advancements in nutritional science and microbiology have highlighted the substantial relation between the GM and multiple pathological conditions, including metabolic syndrome (MetS). A plethora of studies predict that disruptions in the GM, known as dysbiosis, may influence the progression of diabetes, obesity, and cardiovascular diseases (CVDs). Notably, elucidating the contributions of the GM in the pathogenesis of MetS could offer promising avenues for therapeutic interventions. Herein, we review the physiological and metabolic functions of the GM and its connection to MetS pathogenesis, while also highlighting the potential molecular mechanisms underlying these observed associations. Furthermore, we discuss the influence of different dietary approaches on MetS and the impact of nutritional therapeutic strategies to support the development of beneficial gut bacteria and alleviate dysbiosis. By integrating insights from both clinical and preclinical research, this study provides a comprehensive overview of how GM modulation can support metabolic health. The possibility of tailoring nutritional interventions based on individual microbiota profiles represents a promising frontier for personalized and effective approaches to improve metabolic health.
Keywords: gut microbiota, metabolic syndrome, obesity, diabetes, cardiovascular diseases, dietary patterns
Introduction
Metabolic syndrome (MetS) is listed as a prime contributor to the ongoing global cardiovascular health crisis, increasing the risk of premature mortality, significantly.1,2 MetS includes a group of interconnected cardio-metabolic disorders, including diabetes, obesity, dyslipidemia, hypertension, insulin resistance and chronic inflammation.3,4 A diagnosis of MetS is confirmed when at least three of these conditions are met, highlighting the urgent need for early identification and intervention. The onset of MetS is primarily influenced by an interplay of genetic predisposition and environmental factors, including a lack of physical activity and poor-dietary lifestyle, which contribute to excessive adiposity and, subsequently, to other metabolic disturbances.5,6 The global prevalence of MetS varies widely, ranging from less than 10% to as high as 84%, based on the geographic location, population characteristics, and the specific diagnostic criteria used.7,8 Despite this variability, the growing economic and societal burden of MetS underscores the importance of scientific research aimed to unravel the complex mechanisms underlying its pathogenesis.
Recent evidence suggests that the gut microbiota (GM) plays a significant role as a pathogenic factor influencing the human’s metabolic balance and contributing to metabolic disorders.9 Physiologically, GM exerts an important role in shaping the host’s immune system, regulating gut endocrine activity, facilitating digestion, modulating neurological signaling, altering drug metabolism and efficacy, detoxifying harmful substances, and synthesizing various bioactive compounds that affect host physiology.10,11 The alteration in the GM has been reported to be regulated by numerous factors, including an impaired gut barrier, the use of antibiotics, alteration in metabolism of bile acids, and the pleiotropic physiological functions of metabolites generated by microbes, all contributing to the risk of MetS.12 Notably, dietary factors are widely recognized as key contributors to metabolic dysfunction and are also considered the primary determinants of the GM composition. The gut microbiome exhibits remarkable sensitivity to both the quantity and composition of food, functioning as an effective dietary sensor for the host.13 The concept of a gut-centered mechanism in MetS was first proposed in 2007, based on a series of pre-clinical and clinical studies. These studies documented that chronic high-fat diet (HFD) consumption induces defects in the intestinal barrier, which allow the migration of intestinal contents—such as food antigens, bacteria and bacterial by-products—into the systemic circulation, with bacterial lipopolysaccharide (LPS) being a key component.12,14 The subsequent low-grade inflammation, which persists over time, has been termed “metabolic endotoxemia” due to its detrimental effects on normal glucose metabolism.14 This was the first evidence linking bacterial LPS to Toll-like receptors (TLRs) activation, triggering an immune response (innate) that compromises insulin sensitivity. Consequently, understanding factors that influence gut permeability has become a central focus in microbiome research. In addition, established areas of research in metabolic abnormalities including the role of microbial metabolites as bioactive molecules and presence of chronic inflammation have been shown to influence, and be influenced by, the gut microbiome. Insights from these research domains may contribute to the identification of new phenotypic characteristics of MetS, particularly those involving gut-related defects.
This review highlights the recent pre-clinical and clinical advancements in understanding the pathophysiological mechanisms of GM in metabolic disorders, including diabetes, obesity and CVDs. It also discusses the influence of different dietary approaches on the composition of the GM and the risk of MetS. Finally, the potential applications of GM modulation, through the use of dietary (probiotics, prebiotics), as well as therapeutic approaches in management of MetS are also discussed.
Gut Microbiota: Physiological and Pathological Importance
The concept of “microbiota” originated in the early 1900s, when it was discovered that a diverse array of microorganisms, including viruses, yeast and bacteria, inhabits different regions of the human body, such as the skin, gut, oral cavity and lungs.15 Importantly, GM is regarded as the most crucial microbial community for maintaining overall health.16 It exerts local metabolic effects in the gastrointestinal tract, influencing processes such as digestion, energy extraction, nutrient absorption, and the elimination of waste products from ingested food. Additionally, the microbiota modulates various functions, including gut motility and permeability, and the production of gastrointestinal hormones. However, the impact of the GM extends beyond the intestine; the microbes produce a range of small compounds that can influence the physiology of the host both within the gut and beyond, in other organs.17,18 The human GM is primarily composed of six phyla: Bacteroidetes, Proteobacteria, Actinobacteria, Firmicutes, Verrucomicrobia and Fusobacteria, with Bacteroidetes and Firmicutes being the predominant groups.19 The most extensively studied fungi within the GM include Saccharomyces, Cladosporium, Candida, and Malassezia.20 In addition to fungi and bacteria, the GM also harbors archaea, viruses and phages, with Methanobrevibacter smithii being the most commonly identified archaeon.
Physiological Role of Gut Microbiota
The equilibrium of the GM is crucial for maintaining human health and influencing the development of disease. The gastrointestinal tract, in particular, harbors a dense microbial community, with approximately 100 trillion microorganisms.21 A plethora of studies have investigated the significant interactions between GM and essential biological processes in humans. Under healthy conditions, the GM remains stable, resilient, and sustains a mutually beneficial relationship with the host. A healthy GM typically exhibits rich microbial gene content, high taxonomic diversity, and a stable core microbial community.22 However, it is important to recognize that the microbial composition is distinct for each human and can fluctuate within the same person over time. Gut bacteria perform several key roles, including food fermentation, pathogen protection, and immune response modulation.23 Interestingly, the fermentation of dietary plant fibers by gut microorganisms, results in the generation of short-chain fatty acids (SCFAs) such as propionate, acetate, and butyrate. These SCFAs are key metabolites in modulating the normal functioning of colonic cells and contribute to the maintenance of appetite, inflammatory processes and, insulin sensitivity. Additionally, the GM is implicated in the generation of secondary bile acids, degradation of xenobiotics, protein catabolism, and the synthesis of water-soluble vitamins. Beyond these functions, the GM is increasingly recognized for its essential role in the initiation and modulation of both adaptive and innate immunity, as well as gut-centric lymphoid tissue.24
In humans, the gut microbial composition is governed by factors such as age and environmental variables, including medication use.11 Moreover, there is variation in the microbial populations across different areas of the gastrointestinal tract. For instance, the small intestine harbors Proteobacteria, including Enterobacteriaceae, whereas the colon is predominantly populated by Bacteroidetes, such as Prevotellaceae, Bacteroidaceae, and Rikenellaceae.25 These differences are primarily attributable to the distinct environmental conditions in each region. The small intestine has a rapid transit time and high bile concentration, while the colon features slower transit, a more neutral pH, and a larger microbial population, with a higher prevalence of anaerobic bacteria.26 In addition to spatial variation, the composition of the GM also changes with age. Microbial diversity typically increases from childhood to adulthood, reaching its peak during early adulthood, but tends to decline in older age, particularly after the age of 70.27 Prior to reaching a relatively mature GM composition, the microbial diversity of children’s gut is predominantly characterized by species such as Veillonella, Bacteroides, Akkermansia muciniphila, Clostridium botulinum and Clostridium coccoides.28 By approximately age 3, the GM in children begins to resemble that of adults, containing three primary phyla—Bacteroidetes Actinobacteria and Firmicutes.29 As individuals age, changes in diet and immune function may influence the GM composition. Specifically, elderly individuals often exhibit a reduction in Bifidobacterium and an elevation in Proteobacteria and Clostridium.30 The decline in Bifidobacterium, an important anaerobic bacterium, is associated with a compromised immune response and heightened inflammatory status, reflecting its role in immune modulation.
Owing to the essential role of the GM in human health, including its active involvement in various physiological and pathological processes, research on the GM has expanded beyond microbial compositional analysis and the study of microbial associations. Increasing focus is now directed towards understanding the causality of microbial functions, particularly with the advent of advanced technologies such as high-throughput sequencing and interaction modeling and simulation of microbial components.31 Nonetheless, additional research is required to fully elucidate the roles of the human microbiota, which is essential for advancing microbiome-based theranostics and personalized medicine.
Pathophysiological Role of Gut Microbiome
Over the past decade, there has been an increasing interest in understanding the influence of GM in the pathogenesis of various metabolic ailments, including diabetes, obesity, cancer and CVDs32,33 (Figure 1). With the advent of bioinformatics and advanced sequencing technologies, much of the research on the microbiota has focused on investigating the link between alteration in microbiota composition and different disease states. The pathogens associated with these diseases, as well as the relevant signaling pathways, are examined in detail in the respective sections.
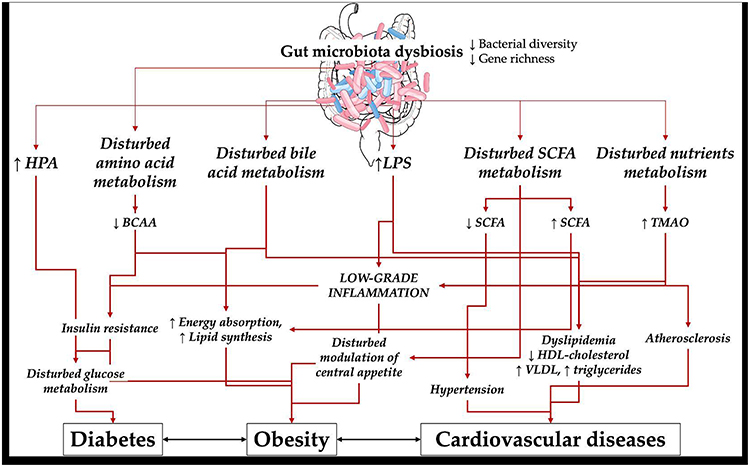 |
Figure 1 Complex pathogenetic associations between gut microbiota dysbiosis and diabetes, obesity and cardiovascular diseases. Image provided by Servier Medical Art (https://smart.servier.com), licensed under CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/). Part of this isIllustration from NIAID NIH BioArt Source (bioart.niaid.nih.gov/bioart/41). Abbreviations: HPA, hypothalamic-pituitary-adrenal axis; LPS, lipopolysaccharide; SCFA, short chain fatty acids; BCAA, branched chain amino acids; TMAO, Trimethylamine N-oxide; HDL, high density lipoprotein; VLDL, very low density lipoprotein. Notes: ↑ denotes increase, ↓ denotes decrease. |
Gut Microbiota and Diabetes Mellitus
Diabetes mellitus (DM) represents a paramount global health issue in the 21st century, imposing significant economic and social burdens.34 The World Health Organization (WHO) estimates that over 10% of the global population is either affected by DM or at elevated risk of having it.35 DM is a progressive metabolic disorder identified by dysfunction of the β-cells of the pancreas and peripheral tissue insulin resistance, which limit glucose metabolism and cause persistent low-grade inflammation.36,37 Type 1 DM (T1DM), Type 2 DM (T2DM), and gestational DM (GDM) are the three prevalent forms of the disease. T1DM is triggered by an autoimmune response that targets and destroys pancreatic β-cells, whereas the body’s incapacity to properly generate or utilize insulin is the hallmark of T2DM. GDM is a prevalent pregnancy complication associated with an increased risk of metabolic diseases for both the mother and the fetus.38 The onset of DM is strongly influenced by both environmental and genetic factors. Environmental factors, including excessive caloric intake, exposure to air pollution, nutrient composition and physical stagnancy, are key contributors to the rising prevalence of the disease.39 DM is a leading risk factor for blindness (diabetic retinopathy), renal dysfunction and cardiovascular complications. The interaction between GM and DM has been extensively studied, and there is a clear connection between the onset of the disease and gut microbial dysbiosis.40,41
Gut Dysbiosis and Type 2 Diabetes
Larsen et al in 2010, documented the first study to establish a strong link between T2DM and GM.42 Using a PCR approach and gene amplicon (16S rRNA) sequencing, the study examined the GM of a small cohort of T2DM patients and found reduced abundance of taxa from the class Clostridia and phylum Firmicutes, compared to controls. Additionally, the Bacteroidetes to Firmicutes ratio was found to correlate positively with altered glucose tolerance, indicating that the GM may be a useful biomarker for predicting the onset of DM. In a seminal study, Qin et al43 conducted the first metagenome-wide association study (MGWAS) in T2DM and revealed that T2DM patients display mild GM dysbiosis. This dysbiosis was primarily characterized by a decrease in butyrate-producing bacteria, such as Roseburia and Faecalibacterium prausnitzii, alongside an overrepresentation of opportunistic pathogens. The study also discovered that the T2DM cohort had an elevation of microbial activities linked to oxidative stress tolerance and sulfate reduction.
Following the initial MGWAS study in T2DM, a larger study conducted in Europe further investigated the intestinal microbiota in postmenopausal female T2DM patients using shotgun sequencing. In both cohort studies, Lactobacillus species and Clostridium clostridioforme were found to be enriched, while Roseburia_272, a key butyrate-producing bacterium, was notably depleted. Additionally, Allin et al44 demonstrated that pre-diabetic subjects exhibit an altered GM, featured by a decreased prevalence of Clostridium genus and Akkermansia muciniphila (mucin-degrading bacterium). The presence of A. muciniphila has been demonstrated to enhance glucose tolerance, reduce insulin resistance, and decrease adipose tissue inflammation.45 These results underscore the variability in gut microbiome composition and highlight the importance of regular microbiota analysis for guiding management strategies. The molecular mechanisms by which the GM contributes to T2DM may involve gut permeability, regulation of glucose metabolism and inflammation. T2DM is generally linked with elevated concentrations of pro-inflammatory molecules, with LPS being a well-established promoter of mild inflammation. A number of investigations have reported an elevated concentration of LPS in peripheral circulation of T2DM patients.46 Through its interaction with Toll-like receptor 4 (TLR4), LPS can activate macrophages, which in turn can activate the NF-κB signaling cascade. This cascade triggers the release of inflammatory cytokines, which ultimately impairs insulin secretion.47
Furthermore, GM plays a role in metabolizing bile acids into secondary bile acids, which can interact with farnesoid X receptor (FXR), triggering the production of fibroblast growth factor (FGF19/15), which enhances glucose tolerance and insulin sensitivity48. Consequently, GM dysbiosis may disrupt bile acid metabolism, potentially contributing to impaired glucose metabolism. Short-chain fatty acids (SCFAs) are another significant class of metabolites produced by the GM. Research indicates that SCFAs are essential for controlling glucose metabolism and enhancing insulin sensitivity through various signaling pathways. For example, the binding of SCFAs with free fatty acid receptors (FFAR2 or FFAR3) present on intestinal L cells, stimulate the secretion of peptide YY (PYY) and glucagon-like peptide-1 (GLP-1), both of which suppress glucagon release and stimulate insulin secretion.49 SCFAs also act as important anti-inflammatory agents, limiting autoimmune responses by elevating regulatory T cell production.50 Therefore, T2DM may occur as a result of a decreased population of microorganisms that produce SCFAs. Additionally, butyrates also preserve the intestinal barrier’s integrity, which may be weakened in T2DM because of low-grade inflammation.50
Long-term follow-up clinical studies have revealed a series of changes that occur during the development of T2DM, including alterations in the catabolism of branched-chain amino acids (BCAAs) and the composition of the bile acid pool.51 A recent study analyzed data on GM, insulin resistance and the pre-meal blood metabolome of 277 Danish subjects without DM, to examine how the GM may influence insulin resistance-related metabolic profiles. The blood metabolome of subjects with insulin resistance revealed an elevated capacity for BCAAs biosynthesis, which was associated with the presence of Bacteroides vulgatus and Prevotella copri in the microbiome of gut. Although these bacterial species lacked genes encoding bacterial transporters for BCAAs, they demonstrated an increased potential for production of these amino acids.52 Study results indicated that GM dysbiosis may influence the blood metabolome, potentially modulating systemic immunity and contributing to the development of insulin resistance.
One fast expanding field of DM research is altered intestinal permeability in T2DM patients, which leads to the subsequent transfer of microbes to various body regions.53,54 The gut’s normal microbiota typically helps regulate the nervous system’s stress response, whereas dysbiosis can lead to an exaggerated hypothalamic–pituitary–adrenal (HPA) axis response to stress.55 This heightened HPA activation results in increased cortisol production, which contributes to dysfunction of the gut barrier by promoting the extracellular matrix breakdown of the human gut. Chronic elevation of glucocorticoid levels can pose significant health risks, including the development of DM.
Critical Evaluation of Current Evidences
Collectively, studies investigating the gut microbiota in T2DM reveal a recurring theme of functional rather than purely compositional alterations. While early reports suggested shifts in the Firmicutes-to-Bacteroidetes ratio, subsequent metagenomic studies have not consistently reproduced these findings, highlighting the limited utility of phylum-level markers. More robust evidence points to depletion of SCFA-producing taxa (eg, Roseburia, Faecalibacterium prausnitzii) and enrichment of opportunistic pathogens, which together may exacerbate systemic inflammation and impair insulin sensitivity. However, many studies differ in methodology (16S sequencing vs shotgun metagenomics), population characteristics (European vs Asian cohorts), and disease stage (prediabetes vs overt T2DM), making direct comparison challenging. These inconsistencies suggest that T2DM-associated dysbiosis may not follow a universal taxonomic pattern but instead reflects context-dependent disruptions in microbial metabolic pathways, particularly those linked to bile acid metabolism, SCFA production, and LPS-driven inflammation. Future work should prioritize longitudinal and interventional studies to disentangle causal relationships from correlative associations.
Gut Dysbiosis and Type 1 Diabetes
Unlike T2DM, T1DM is characterized by the autoimmune disruption of insulin-secreting β cells of pancreas. While the underlying mechanisms of T1DM differ from those of T2DM, emerging evidence suggests that alterations in the GM are also present in T1DM.56–58 Interestingly, GM dysbiosis is predicted to develop early in life and exacerbate gut inflammation, potentially affecting immune system function before T1DM manifests clinically. Various studies have reported that in individuals developing T1DM, a pro-inflammatory gut environment is consistently linked with an elevated relative occurrence of Bacteroidetes and a decrease in Firmicutes, regardless of geographic location59 (Table 1). A longitudinal clinical study by Vatanen et al,58 examined the fecal samples of children (aged between three months to five years), and reported that children who were later diagnosed with T1DM had elevated levels of Alistipes shahii, Bifidobacterium pseudocatenulatum, and Roseburia hominis. In contrast, children who did not develop T1DM had higher levels of Streptococcus thermophilus and Lactococcus lactis, two species commonly present in dairy products. Furthermore, genes linked to the fermentation and production of SCFAs were more prevalent in the microbiomes of healthy children, indicating a possible protective function of SCFAs in the early development of T1DM.58 Given that T1DM typically begins in the early phase of life, the GM may be critical in preventing the onset and development of the disease by promoting the establishment of a healthy microbiota from birth.
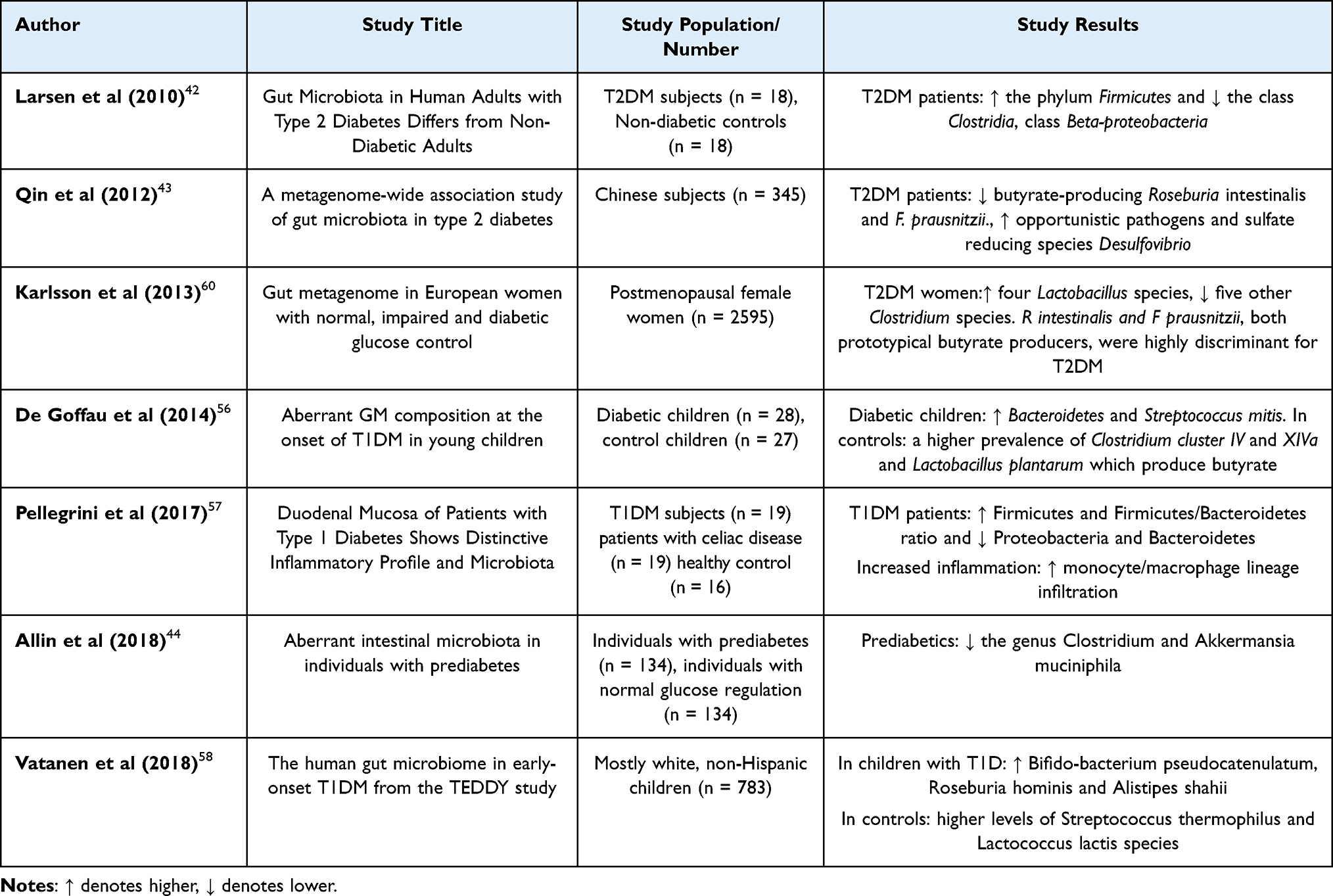 |
Table 1 Summary of Clinical Studies on Gut Microbiota in DM |
Scientific evidence proposed that the GM may contribute to the development of T1DM primarily through the modulation of immune responses. The pathophysiology of T1DM may be influenced by the disruption of the GM, given its role in the onset of chronic inflammation.
In support of this, Higuchi et al reported that blood IL-6 levels were significantly elevated in T1DM patients in comparison to normal controls, and this increase was associated with the presence of Ruminococcus and Ruminococcaceae.61 Leiva-Gea et al also documented that individuals with T1DM had higher levels of pro-inflammatory cytokines (IL-1β, IL-6 and TNF-α) alongside reduced levels of anti-inflammatory cytokines (IL-13 and IL-10). These alterations were linked with the prevalence of various bacterial species.62 Additionally, T1DM was shown to have elevated LPS, which is known to cause the release of proinflammatory cytokines and disrupt pancreatic β-cell function.63 Notably, clinical data have demonstrated that T1DM subjects exhibit significantly higher levels of ligands for TLR2 and TLR4, suggesting an upregulated activity of TLR2 and TLR4.64 As previously discussed, TLRs are crucial in both innate and adaptive immunity, serving to shield the body against infectious pathogens. Gülden et al showed that TLR4 promotes the progression of DM, indicating that TLR4 plays a role in the development of insulitis.65 The TLR4/MyD88 signaling pathway is involved in NF-κB activation and the regulation of pro-inflammatory cytokines, including TNF-α and IL-6.66 Wang et al created MyD88-deficient Non-Obese Diabetic (NOD) mice and observed that these mice did not develop T1DM. These results predict that the GM may promote the progression of T1DM through the TLR4/MyD88 signaling pathway.67
Overall, the findings from the studies discussed indicate that individuals with T1DM exhibit signs of gut dysbiosis. However, further research to fully elucidate the pathophysiological mechanism of GM dysbiosis and T1DM is needed.
Critical Evaluation of Current Evidences
Evidence for gut microbiota involvement in T1DM is compelling but remains fragmented. Across cohorts, children who develop T1DM consistently display reduced microbial diversity and an inflammatory signature characterized by increased Bacteroidetes and decreased Firmicutes, although the specific taxa reported vary widely between studies. Longitudinal data suggest that these microbial changes precede clinical onset, implying a role in disease initiation rather than progression. Mechanistic studies highlight the contribution of microbial modulation of TLR2/TLR4 pathways and SCFA production to shaping immune tolerance; however, most data are correlative and derived from small pediatric cohorts or mouse models, limiting generalizability. Importantly, whether dysbiosis drives autoimmunity or arises secondarily from genetic and environmental predispositions remains unresolved. Future investigations should focus on harmonizing sequencing approaches and integrating host genetics, diet, and immune profiling to clarify causality and therapeutic potential.
Gestational Diabetes Mellitus
In GDM, scientific studies have revealed GM as a key contributing factor in mediating inflammation, and insulin resistance during pregnancy. Women with GDM frequently experience metabolic disturbances, including reduced secretion of insulin and increased insulin resistance.68 The GM composition undergoes significant alterations during pregnancy, which may contribute to the onset of GDM. For instance, positive correlations have been observed between Collinsella and insulin levels, Coprococcus and gastrointestinal polypeptide, and Lachnospiraceae and Ruminococcaceae with adipokines69. Additionally, Koren et al reported that the GM undergoes significant changes from the first to the third trimester, characterized by elevated microbial diversity and reduced richness.70 Women with GDM had an elevated ratio of Firmicutes to Bacteroidetes, a key factor associated with the promotion of obesity and exacerbation of inflammation.71 Compared to healthy controls, the number of bacteria that produce SCFAs was significantly reduced in GDM pregnancies, indicating that alterations in the microbiota may contribute to elevated blood glucose levels.72 Additionally, studies have demonstrated that the GM composition of children born to women with GDM is different to that of children born to non-GDM women. According to Ponzo et al, proinflammatory microorganisms were more prevalent in GDM neonates than in healthy controls.73 This discovery was corroborated by additional research, which showed that GDM newborns had lower α-diversity than the control group, and that the mother’s GDM status affected several lactic acid bacteria.74 These findings suggest that the GM contributes to the progression of GDM and may also impact the microbiota composition in GDM offspring.
Critical Evaluation of Current Evidences
In summary, gut microbiota alterations appear to mirror the metabolic and inflammatory state of pregnancy. Multiple studies converge on a reduction of SCFA-producing bacteria and increased prevalence of proinflammatory taxa, with downstream effects on glucose tolerance and adipokine regulation. Notably, maternal dysbiosis is transferred to neonates, potentially predisposing offspring to future metabolic risk, underscoring the transgenerational impact of GDM. However, available studies are largely cross-sectional, with small sample sizes and heterogeneous diagnostic criteria, making it difficult to establish temporality or causation. Additionally, pregnancy itself induces profound physiological shifts in the microbiome, complicating differentiation between normal gestational adaptation and pathological dysbiosis. Thus, while the link between altered microbiota and impaired glucose metabolism in GDM is plausible, definitive mechanistic evidence is lacking, and future work should include well-controlled longitudinal studies tracking mother–infant dyads.
Gut Microbiota and Obesity
Obesity is a multifactorial metabolic disease, triggered by a range of environmental and genetic factors. According to the WHO reports, a body mass index (BMI) of 30 or higher is considered obese. Global assessments indicate that around one-third of the human population is classified as overweight, with approximately 10% categorized as obese.75 It is projected that by end of 2030, the global population of obese individuals will cross 1.12 billion.76 The health risks associated with obesity have raised significant concern, positioning it as a major global health issue.77 Obesity is not only characterized by alterations in physical appearance, but is also linked to disturbances in glucose metabolism, oxidative stress, systemic inflammation, and an elevated risk of various diseases, particularly CVDs, cancer and diabetes.78,79 Recent research has increasingly indicated that GM imbalance may contribute to the development of obesity.80,81 The hypothesis correlating GM dysbiosis, as a predominant environmental risk factor in obesity has prompted investigations into the microbiomes of obese subjects. Generally, obese patients exhibit reduced bacterial diversity and gene richness.82,83 Recent studies conducted on European subjects have shown that individuals with less diverse microbiomes tend to have higher levels of obesity, dyslipidemia, increased inflammatory response, and more prominent insulin resistance.84 Similarly, overweight and obese individuals with low microbial diversity experience an elevated microbial richness when subjected to an energy-restricted diet.85
Early research studies initially suggested that the ratio of Firmicutes-to-Bacteroidetes is elevated in obese subjects, which gradually reduces following weight loss through calorie-restricted diets or bariatric surgery.86 However, subsequent studies largely failed to replicate these findings.87–89 It is crucial to acknowledge that these studies often lacked sufficient statistical power to detect significant differences. A meta-analysis that identified no significant differences in the quantity of Firmicutes and Bacteroidetes, nor in their ratio, provided the strongest evidence against a major association between the ratio and obesity.90 Overall, research in this field has shed light on the association between microbiota and obesity, even though the ratio of these two phyla is not a valid indicator of obesity. Notably, these findings challenge the idea of a single, specific taxonomic signature for obesity, suggesting that future research should focus on identifying specific taxonomic markers that could be used to categorize patients into distinct subgroups. A 2025 comprehensive systematic review on microbiome-targeted interventions for obesity confirmed modest but significant reductions in body weight, adiposity, and inflammatory markers with probiotics, synbiotics, and FMT, though effects varied by strain and study design.91
Recent studies have linked obesity to specific bacterial families and genera, such as the Christensenellaceae (family) and the Methanobacteriales, Bifidobacterium, Lactobacillus, and Akkermansia (genera). Notably, the Christensenellaceae family was reported to be linked with weight reduction, with its relative prevalence inversely correlated with the host’s BMI.92 Akkermansia muciniphila has been identified as an important bacterium for weight reduction, with supplementation of this species improving metabolic parameters in obese individuals.93 Bifidobacterium and Lactobacillus are traditional probiotics, which exert a significant effect in regulating the human GM balance. A review by Crovesy et al94 reported that Lactobacillus has a species-specific effect on reducing body weight in overweight people. Specifically, an abundance of Lactobacillus paracasei was correlated negatively with obesity, whereas abundances of Lactobacillus gasseri and Lactobacillus reuteri were positively correlated with obesity. Evidence from pre-clinical studies suggests that Bifidobacterium may have a protective role against obesity, showing strain-dependent effects in diet-induced obesity models.95 Moreover, a reduction in the abundance of Bifidobacterium has been associated with obesity.96 These findings highlight the species-specific nature of obesity-related microorganisms, suggesting that microbes belonging to a specific genus may have opposing effects, potentially due to the complex metabolic mechanisms underlying obesity. More research is needed to elucidate the effect of other human gut microbial species in obesity.
Mechanistic Association of Gut Microbial Dysbiosis and Obesity
The altered GM is believed to contribute to the development of obesity by various mechanisms, such as the disturbance of energy balance, modulation of lipid production and storage, alteration of central appetite and feeding behaviors, and the promotion of chronic low-grade systemic inflammation.97
Absorption of Energy and Lipid Synthesis
GM metabolizes dietary macronutrients to generate bioactive metabolites, such as SCFAs, bile acids and amino acids (and their derivatives). In obese subjects, dysbiosis of the GM disrupts metabolite profile, which may contribute to enhanced lipid synthesis and energy absorption.98 Subsequently, the GM of obese mice exhibited an increased capacity to utilize non-digestible carbohydrates (NDC), for the synthesis of SCFAs and other monosaccharides which are utilized for energy production and fat storage, compared to the normal-weight mice. Elevated amounts of α-amylases and amylomaltases in the GM of obese mice were the main cause of these processes.99 Notably, SCFAs are essential as substrates for lipogenesis and energy production, meeting 60–70% of colonic epithelial cells’ caloric requirements and accounting for 5–15% of the body’s overall caloric intake.100,101
Despite their involvement in regulating the body’s energy metabolism, their effects on energy absorption and expenditure are complex and still subject to debate. SCFAs prevent intestinal epithelial cells from releasing fasting-induced adipocyte factors, which encourages the buildup of triglycerides into adipocytes that are guided by lipoprotein lipase and contribute to obesity.102 Conversely, SCFAs may promote fatty acid oxidation and thermogenesis through AMP-activated protein kinase (AMPK) activation, which would increase the expression of mitochondrial uncoupling protein 1 and PPAR-γ coactivator 1α, and provide protection against the development of obesity.103 Blaak et al reviewed a number of pre-clinical studies which showed that dietary supplementation with either single or mixed SCFAs significantly reduced high fat diet induced weight gain.104 However, it is crucial to mention that these conclusions are based on experiments on limited clinical research, particularly animal models, and small sample sizes.
In clinical settings, Schwiertz et al observed that obese individuals exhibited a distinct microbial and metabolite status, which resulted in an increased concentration of SCFAs with respect to normal subjects.86 Supporting this, a similar clinical analysis found that increased excretion of SCFAs was associated with gut permeability, gut dysbiosis, excess fat accumulation, and elevated CVD risk factors.105 However, well-controlled, long-term clinical trials examining SCFAs in human subjects are lacking. Future studies should focus on precise SCFAs metabolic kinetics and dosage, taking into account variations due to diverse metabolic conditions of humans.
The GM influences the metabolic status of amino acids as well as their byproducts, contributing to excessive lipid synthesis in the host. Liu et al identified a novel correlation between circulating amino acids, changes in the GM and obesity. Study results reported that the prevalence of glutamate-fermenting bacterium-Bacteroides thetaiotaomicron, was significantly reduced in overweight subjects and showed an inverse relationship with serum glutamate levels. Changes in body circumference, increased fat production, and significant weight gain were all linked to elevated serum glutamate levels.106 Pre-clinical (obese mice) and clinical (obese humans) studies revealed that a decrease in indole derivatives produced by the GM metabolizing tryptophan, results in an elevation of miR-181 family expression. This over-expression is linked to inflammation in adipose tissue and elevated lipid synthesis, both of which contribute to obesity development.107 While certain GM are known to generate various metabolites, the specific microbial contributors in the human gut are still not known. This necessitates further research to determine their exact functions within host pathways, and to understand the underlying mechanisms in various intestinal and other tissue cells.
Interestingly, the reduction of Lactobacillus and Bacteroides in over-weight subjects leads to a reduction in the levels of bile acids, which are crucial for managing fat storage and lipid synthesis.108 In the liver, bile acids stimulate the Farnesoid X Receptor (FXR), which decreases the liver receptor homolog 1 expression through a mechanism involving a small molecule heterodimer partner. This suppression limits the transactivation of sterol regulatory element-binding protein 1c (SREBP1c), a key regulator of genes involved in adipogenesis, thereby reducing synthesis of fatty-acids in human liver cells, and slowing the progression of obesity.109 Overall, these results emphasize the significant link between the GM, microbial metabolites, and energy storage by adipose tissue. A deeper understanding of these mechanisms could provide new insights and potential therapeutic targets for obesity treatment.
Chronic Systemic Inflammation and Reduced Insulin Function
Low-grade, chronic systemic inflammation is often considered as a major characteristic of metabolic disorders, primarily driven by increased serum levels of LPS.110 Obese individuals with GM dysbiosis typically exhibit an increased prevalence of LPS-releasing bacteria (Veillonella) and a reduction in beneficial bacteria (Akkermansia muciniphila), that maintain the gastrointestinal mucosal barrier integrity.45,111 These microbial imbalances lead to increased intestinal LPS concentrations, greater permeability of the intestinal barrier, and increased passage of harmful by-products from the gut into the circulation. In this context, excessive fat intake from a high-fat diet stimulates chylomicron generation in the gut after meals, which further promotes the LPS release into the bloodstream via the lymphatic system.112 Consequently, disruptions in the GM and remodeling of intestinal permeability may act as key contributors of inflammation in obesity, with unhealthy dietary factors potentially exacerbating this inflammatory response.
In healthy humans, an increase in fasting plasma glucose levels triggers insulin secretion, facilitating the uptake of glucose by peripheral tissues, which then leads to oxidative respiration and glycolysis. Insulin function is hampered by insulin resistance, which is characterized by a reduction or partial impairment of insulin sensitivity in peripheral organs. This leads to aberrant fat deposition in the liver and adipose tissue, as well as fasting hyperglycemia.113,114 The upregulation of inflammatory mediators in adipose tissue observed during obesity, accompanied by alterations in glucose tolerance and insulin sensitivity, was pivotal in establishing the connection between metabolic dysfunction and chronic inflammation.115 Cytokines (TNF-α and IL-1β), found in the adipose tissue of both humans and rodents, affect insulin sensitivity by regulating the gene expression of insulin receptor-1 (IRS-1), glucose transporter 4 (GLUT4), and PPAR-α, leading to insulin signaling desensitization.116 Subsequently, TNF-α can stimulate key inflammatory signaling components, including c-Jun N-terminal kinases and kappa B kinase inhibitor, which decreases the tyrosine phosphorylation of downstream insulin receptor substrates, attenuating insulin signaling.117 Changes in the GM notably affect intestinal metabolism of toxic by-products, promoting mild inflammation. Insulin resistance and obesity-related processes can be triggered by a variety of inflammatory mediators that affect many elements of the insulin signaling cascade, including transporters, insulin receptors, and downstream substrates.
Modulation of Central Appetite
GM exerts an important role in modulating feeding behavior and central appetite, by influencing gut metabolites, intestinal hormones, and neurotransmitters.
During feeding a variety of stimuli, including changes in intestinal osmotic pressure, the availability of nutrients, mechanical stimulation, and other elements, cause neuroendocrine processes to be activated. In response to these cues, enteroendocrine cells (EECs), which are widely dispersed throughout the intestinal epithelium, quickly release hormones that are essential for controlling metabolism. Key hormones secreted by EECs include pancreatic polypeptide (PP), GLP-1 and PYY, each exhibiting unique but overlapping physiological roles. PYY primarily acts to inhibit upper gastrointestinal motility, reduce electrolyte secretion and mucosal fluid, and slow colonic transit, collectively promoting satiety.118 Similarly, PP contributes to appetite regulation by delaying gastric emptying, enhancing satiety, and facilitating energy expenditure, highlighting its potential role in weight reduction mechanisms.119 GLP-1 plays a critical role in metabolic regulation by reducing glucagon secretion, delaying gastric emptying, stimulating insulin production, and decreasing food intake.120 Emerging research highlights the potent anorexigenic properties of GLP-1 and related hormones, which modulate appetite and feeding behavior. These effects are mediated through their entry into the circulation or interaction with the enteric nervous system, where they bind to specific receptors on vagal afferents, the hypothalamus, enteric neurons, as well as the brainstem.121 Notably, studies have revealed that GLP-1 and PYY levels are predominantly reduced in individuals with obesity compared to those with normal weight, and their blood levels increase progressively after bariatric surgery.122 An investigation by Schéle et al demonstrated that GM can suppress brain-derived neurotrophic factor and the anti-obesity neuropeptide GLP-1 precursor proglucagon (Gcg) expression. This suppression may lead to increased food acquisition, potentially aggravating obesity progression.123 Consequently, strategies aimed at modulating endogenous satiety hormones, or delivering exogenous hormone analogs to stimulate their specific receptors and enhance satiety signaling present a promising avenue for reducing food intake and managing obesity.
Research has shown that metabolites derived from microbiota, including bile acids, SCFAs, and indoles, play an important role in regulating the secretion of gut hormones.124,125 SCFAs interact with EECs by targeting G-protein-coupled receptors (GPR41 and GPR43), stimulating the secretion of GLP- 1 and PYY. This binding also influences the production of neuropeptides that regulate postprandial satiety.126 Notably, butyrate, a type of SCFAs, can penetrate the blood-brain barrier and attach to hypothalamic receptors, activating the vagus nerve, and subsequently modulating feeding behavior and appetite.127 Furthermore, lactate, produced by beneficial microbes such as Lactobacillus and Bifidobacterium, provides neuronal cells with energy and prolongs postprandial satiety.128
Taken together, findings link obesity to reduced microbial diversity and specific taxa such as Christensenellaceae and Akkermansia, yet no universal “obese microbiome” signature has emerged. Inconsistencies in the Firmicutes/Bacteroidetes ratio highlight the limitations of broad taxonomic markers. Instead, functional traits—such as altered SCFA metabolism, amino acid biosynthesis, and bile acid signaling—appear more relevant. This suggests that obesity is associated with diverse microbial imbalances rather than a single compositional pattern.
Critical Evaluation of Current Evidence
The evidence linking gut microbial dysbiosis with obesity highlights several mechanistic pathways, including enhanced energy harvesting, altered lipid synthesis, systemic inflammation, and disrupted amino acid metabolism. However, most mechanistic insights stem from animal models with limited translational value to humans, often constrained by small sample sizes and short follow-up periods. While SCFAs are presented as both obesogenic and anti-obesogenic, the duality of their function underscores unresolved contradictions that require clarification through controlled human studies. Similarly, inconsistencies in the Firmicutes-to-Bacteroidetes ratio across populations question its reliability as a universal biomarker of obesity. Findings on species such as Akkermansia muciniphila, Bifidobacterium, and Lactobacillus are promising but strain-specific, making generalization difficult. Moreover, metabolic effects attributed to GM—such as amino acid-driven lipid synthesis or bile acid–mediated regulation—remain correlative, with causality not firmly established. The section would benefit from integrating meta-analyses that emphasize inter-individual variability, host genetics, and dietary context as key modulators of microbiome-obesity interactions. Taken together, while GM dysbiosis is clearly associated with obesity, current data remain fragmented, requiring harmonized, longitudinal, and multi-omics approaches to move beyond descriptive correlations toward actionable biomarkers and therapies.
Gut Microbiota Dysbiosis and Cardiovascular Diseases
CVDs, encompassing conditions such as coronary heart disease, peripheral arterial disease and cerebrovascular disease, are the main causes of morbidity and mortality globally. Common risk factors include obesity, atherosclerosis, DM, hypertension, and dyslipidemia. Emerging evidence predicts the GM is essential for controlling cardiovascular health, and its dysregulation may be a factor in the onset and advancement of CVDs.129
Hypertension
Hypertension poses a serious threat to public health globally, and is the primary risk factor for CVDs, imposing a substantial economic burden on society. A compelling body of evidence has linked GM dysbiosis with hypertension. Li et al in 2017130 conducted an analysis of GM composition in healthy individuals, pre-hypertensives, and subjects with essential hypertension using metabolomic and metagenomic approaches. The study revealed that hypertensive patients exhibited reduced GM diversity, richness, and gene abundance compared to healthy controls. Furthermore, the GM composition in pre-hypertensive individuals closely resembled that observed in hypertensive individuals.
In a cross-sectional analysis, Sun et al131 explored the relationship between blood pressure and GM diversity in the Coronary Artery Risk Development in Young Adults (CARDIA) study cohort. Study results demonstrated a negative association between gut microbial diversity and hypertension, marking the first population-based cohort study to investigate this relationship. Subsequently, Liu et al132 utilized reverse transcription-quantitative polymerase chain reaction (RT-qPCR) to quantify gut microbial populations. Study results revealed a reduction in Bifidobacterium and Bacteroides thetaiotaomicron abundance, alongside an increase in Eubacterium rectale, in hypertensive patients. Palmu et al132 assessed the correlation between hypertension and GM in a cohort of 6,953 Finnish participants. Their findings revealed significant alterations in gut microbial composition among hypertensive individuals, particularly within the Firmicutes phylum. Similarly, Yan et al133 reported an increased prevalence of certain opportunistic pathogens, such as Streptococcus, Parabacteroides, and Klebsiella, in hypertensive patients. In contrast, the control group exhibited higher levels of SCFAs-producing microbes, including Bacillus freundii and Roseburia. A metagenomic analysis revealed significant alterations in the composition and functionality of GM in hypertensive subjects, including a marked reduction in fecal butyrate levels compared to healthy controls134. Additionally, specific gut bacterial populations were linked to variability in blood pressure and severity of hypertension. The relative abundance of Lactobacillus and Alistipes finegoldii were positively associated with lower blood pressure alteration, whereas Prevotella and Clostridium were correlated with higher variability in blood pressure.135
Significant variations in the relative abundance of particular taxa and the composition of GM were found in another clinical investigation between hypertensive patients with varying cardiovascular risk stratifications. Notably, probiotic abundance markedly decreased as cardiovascular risk increased, indicating that GM may contribute to the etiology and development of hypertension.136
Dyslipidemia
Owing to its regulatory effect on host lipid metabolism, GM has been closely linked to hyperlipidemia and associated diseases. GM functions as a critical metabolic “organ,” playing an essential role in host metabolism, homeostasis, and health maintenance, and is intricately linked to the onset and progression of dyslipidemia.137,138 Alterations in the gut environment can suppress the growth of beneficial bacteria such as Lactobacillus, Bifidobacterium, and butyrate-producing species while promoting the proliferation and dysregulation of Enterobacteria, leading to dyslipidemia.139 Conversely, dyslipidemia exacerbates gut microbiota dysbiosis. During digestion, various nutrients including dietary fiber, carnitine, polyunsaturated fatty acids (PUFAs) and bile acids, are metabolized by microbial enzymes into metabolites such as conjugated linoleic acid (CLA), SCFAs, secondary bile acids and trimethylamine N-oxide (TMAO).137 These microbial metabolites exert systemic effects on the host. For instance, TMAO reduces high-density lipoprotein (HDL) levels in the liver, thereby disrupting lipid metabolic balance.140 CLA and SCFAs bind to PPARs to lower elevated triglycerides (TG), HDL-c, and very-low-density lipoproteins (VLDL).141 Additionally, secondary bile acids engage with FXR receptors, which are linked to increased HDL levels, enhanced lipolysis, and reduced VLDL concentrations.142 Notably, individuals with hyperlipidemia exhibit reduced SCFA concentrations, elevated levels of LPS and TMAO, and alterations in the bile acid pool.116
Studies have reported that hyperlipidemic subjects exhibit significant disruptions in the structure and function of their GM. Adolescents and children with hyperlipidemia display reduced fecal levels of SCFAs, which are associated with SCFA-producing bacteria, including members of the families Ruminococcaceae, and Lachnospiraceae and genera Roseburia, Bacteroides, Akkermansia, and Faecalibacterium.143 A randomized controlled clinical study demonstrated that fecal samples from individuals with MetS, characterized by hyperlipidemia, contained lower abundances of probiotic species such as Lactobacillus, Bifidobacterium, Roseburia and Faecalibacterium prausnitzii, along with increased levels of LPS-producing bacteria like Enterobacter cloacae and Escherichia coli, compared to healthy controls.144 At present, there is no clear consensus on the precise compositional imbalances in gut microbiota linked to hyperlipidemia, potentially owing to the functional redundancy of GM.
Atherosclerosis
According to recent research, atherosclerotic plaques include bacterial DNA, and the same bacterial taxa have been found in both the plaques and the guts of the same subjects.145 These findings indicate that the bacterial communities in these locations could serve as a bacterial source in the plaques, potentially influencing plaque stability and the progression of CVDs. Metagenomic analysis of fecal microbiota demonstrated that microbial abundance differs between subjects with unstable and stable plaques. Specifically, unstable plaques were characterized with lower fecal prevalence of the genus Roseburia, an elevated theoretical ability of the microbiome to release pro-inflammatory peptidoglycans, and a reduced capacity to produce anti-inflammatory carotenoids.146 Thus, the gut microbiome in CVD patients may contribute to inflammation by generating more pro-inflammatory molecules. A recent systematic review and meta-analysis (Martins et al, 2024) synthesizing 67 studies with nearly 12,000 participants confirmed consistent microbial differences between CVD patients and healthy controls, particularly involving TMAO, Streptococcus, and SCFA-producing taxa. This reinforces the role of dysbiosis in CVD, though population and methodological variability remain significant.147
Mechanistic Insights About Gut Microbiota Dysbiosis and CVDs
GM regulates the metabolism of phosphatidylcholine, choline, and carnitine, ultimately producing TMAO, which has been implicated in regulating cholesterol homeostasis and bile acid levels, and is also linked with the early stages of atherosclerosis and an increased risk of CVDs. Independent cohorts have demonstrated a link between elevated TMAO levels and adverse clinical outcomes.148 In a clinical trial involving over 4,000 individuals undergoing elective coronary angiography, higher TMAO levels were linked with higher incidence of major adverse cardiovascular events (MACE), such as stroke, myocardial infarction, and death, during a 3-year follow-up period.149 Notably, patients with highest quartile of circulating levels of TMAO had a 2.5-fold higher risk of experiencing a MACE compared to patients in the lowest quartile. Since these early studies, additional research has consistently shown associations between elevated levels of TMAO and risk of CVD.150–152 TMAO, in circulation, was correlated with the incidence of plaque rupture, coronary plaques and chronic risks of CVDs in subjects with acute coronary syndrome.153
Mechanistically, TMAO can stimulate the expression of inflammatory cytokines (TNF-α, IL-6. IL-8), by activating NF-κB and MAPK signaling cascades in smooth muscle and endothelial cells. NF-κB is widely recognized as a critical mediator in the stimulation, differentiation, and effector functions of inflammatory immune cells. Consequently, NF-κB dysregulation may be a major factor in the pathogenesis of atherosclerosis by facilitating the recruitment of monocytes.154 Moreover, LPS has been reported to promote vascular inflammation and endothelial dysfunction by inducing vascular oxidative burden.155 A retrospective study performed by Yoshida et al revealed that CVD patients display elevated levels of fecal LPS, with respect to healthy subjects.156
The GM plays a central role in regulating dyslipidemia primarily through the production of cholesterol oxidase, which inhibits cholesterol synthesis while promoting its degradation and transformation. Additionally, GM and its metabolites, (SCFAs and bile acids), influence chronic inflammation, energy harvesting efficiency, and immune system activation, by altering intestinal barrier permeability. These mechanisms can disrupt reverse cholesterol transport and contribute to metabolic alterations, including dyslipidemia.157 Fu et al identified Bacteroides and Firmicutes as the predominant phyla affecting blood lipid levels, significantly influencing serum HDL-C and TG levels.158
Overall, evidence implicates gut dysbiosis in CVD through pathways involving TMAO, LPS, and reduced SCFA production. However, findings vary widely across cohorts, with functional redundancy and host–microbe interactions complicating causal inference. While some taxa show consistent associations with hypertension, dyslipidemia, and atherosclerosis, mechanistic validation in humans is limited. Future studies should integrate microbiome data with metabolic and immunological profiling to better define causality and therapeutic targets.
Critical Evaluation of Current Evidences
The growing body of evidence linking gut microbiota (GM) dysbiosis with cardiovascular diseases (CVDs) provides strong biological plausibility, especially through metabolites such as trimethylamine-N-oxide (TMAO), short-chain fatty acids (SCFAs), and secondary bile acids. Elevated TMAO levels have been associated with major adverse cardiovascular events across diverse cohorts, yet much of this evidence remains observational. The inability to establish clear causality raises questions as to whether TMAO is an active mediator of disease progression or a surrogate biomarker influenced by host diet and comorbidities.
Studies on hypertension and dyslipidemia consistently demonstrate associations with reduced microbial diversity and altered SCFA-producing taxa, but most are limited by cross-sectional design, modest sample sizes, and confounding factors such as medication use, dietary habits, and underlying metabolic disorders. This undermines the reproducibility of microbial signatures across populations, making it difficult to propose universal microbiome-based biomarkers of CVD.
Animal models have advanced mechanistic understanding by implicating inflammatory pathways such as NF-κB and MAPK in microbiota-driven vascular dysfunction. However, interspecies differences in microbiota composition and metabolite metabolism restrict translational relevance to human disease. Furthermore, functional redundancy within microbial communities means that shifts in taxa composition may not directly translate into consistent metabolic or clinical outcomes.
Taken together, while current findings underscore the potential contribution of GM dysbiosis to CVD pathophysiology, the field remains dominated by associative rather than causal evidence. Future studies should emphasize longitudinal cohort designs, standardized sequencing methodologies, and multi-omics integration to disentangle confounding factors, identify reproducible microbial markers, and advance microbiota-targeted therapeutic strategies for cardiovascular health.
Gut Microbiota: As a Metabolic Syndrome Biomarker or Diagnostic Predictor
A gut metagenomic linkage group associated with MetS risk has been identified, and taxonomic analysis revealed certain microbial risk markers characterized by moderate dysbiosis, reduced abundance of butyrate producers, increased prevalence of pathogenic bacteria, and an improvement of microbial functions associated with sulfate reduction resistance and oxidative stress.43 Emerging evidence strongly suggests that inter-individual deviations in GM not only predict disease progression but also influence responses to dietary interventions. Precision nutrition research has consistently highlighted the importance of GM composition in personalizing dietary recommendations. Variability in postprandial blood glucose responses to identical meals has been linked to alterations in GM composition. The PREVIEW trial, involving pre-diabetic overweight or obese subjects, consuming on low-energy diet (LED) for an 8-week period demonstrated that baseline GM features could predict reductions in body fat during the LED phase.159
Similarly, a cohort study involving 800 participants found significant inter-individual differences in post-prandial glucose responses, which could be linked to specific clinical and microbiome characteristics; this was confirmed in a follow-up study of 100 individuals.160 Furthermore, the PREDICT trial showed that GM composition accounted for 6.4% of post-meal glycemia, 7.5% of post-meal TG variability, and 5.8% of C-peptide levels (postprandial), independent of other individual characteristics, in over 1,000 twins and unrelated individuals, with findings validated in a US cohort of 100 participants.161
Certain microbial species, such as Blastocystis and Prevotella copri, have been identified as markers of advantageous postprandial glucose metabolism. Additionally, the overall composition of GM has been shown to predict a wide range of cardiometabolic blood indicators, such as fasting and postprandial lipemic, glycemic, and inflammatory indices.162 Notably, in obese individuals undergoing caloric restriction, improved insulin sensitivity has been observed in subjects having higher prevalence of Akkermansia muciniphila.163 Similarly, Parabacteroides distasonis has been linked with enhanced sensitivity to insulin in obese individuals.164
Crosstalk Between Dietary Interventions and Metabolic Syndrome
The prevalence and prevention of MetS is significantly influenced by modifiable lifestyle factors, especially eating habits, according to recent studies.165–167 The positive impacts of several dietary approaches on inflammatory markers linked to MetS were investigated by Steckhan et al.168 Similarly, Godos et al performed a meta-analysis demonstrating the preventative potential of healthy dietary lifestyle to decreased MetS prevalence169 (Table 2).Herein, we present the possible advantages of different dietary strategies for managing MetS and its related comorbidities, as well as their efficacy as preventative and therapeutic measures (Figure 2).
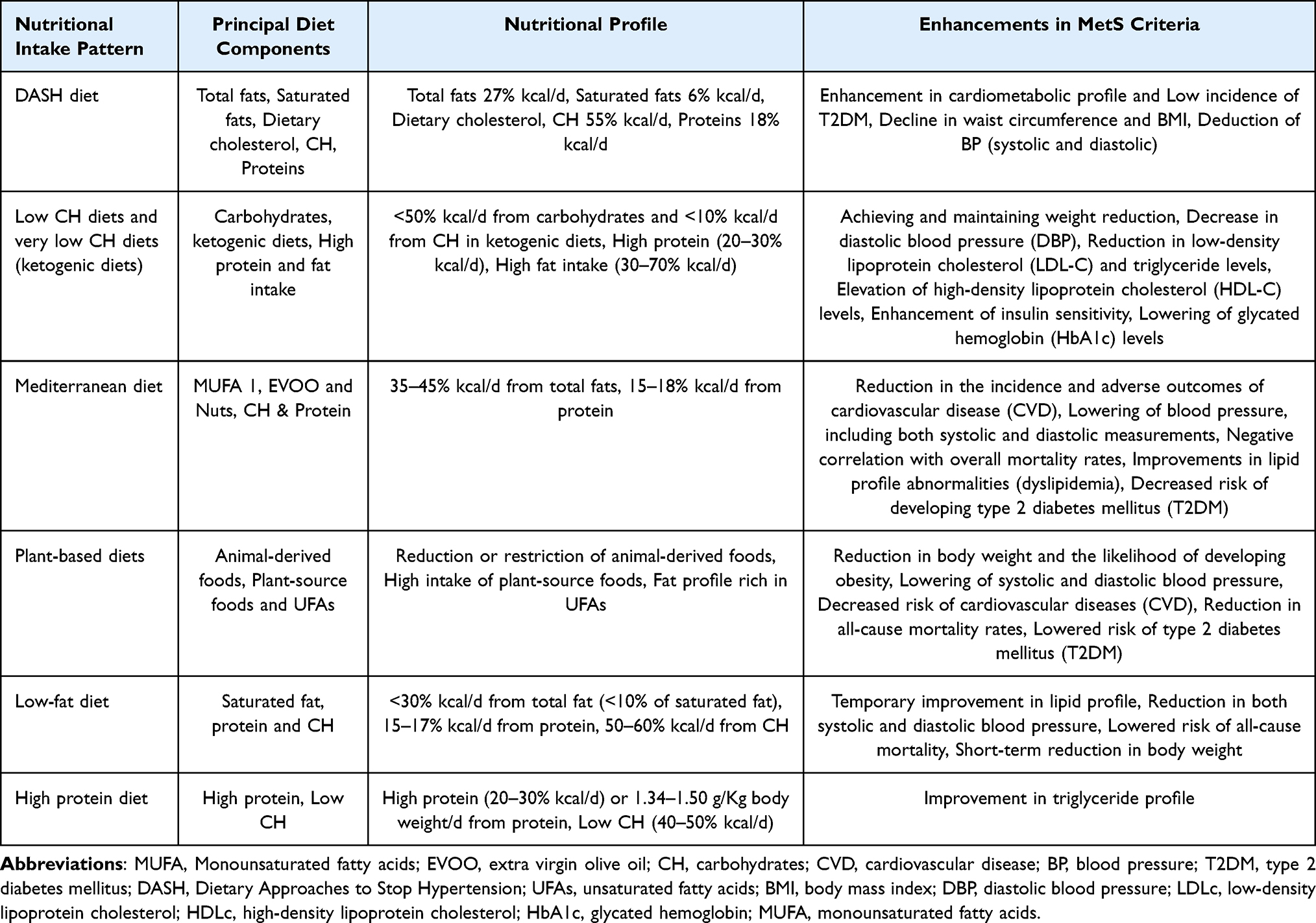 |
Table 2 Dietary Patterns and Their Influence on MetS |
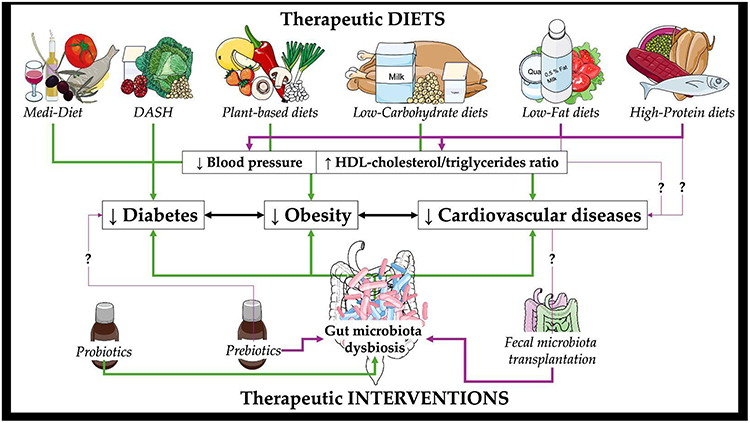 |
Figure 2 Therapeutic diets and therapeutic interventions on microbiota dysbiosis and their effects on diabetes, obesity and cardiovascular diseases. Image provided by Servier Medical Art (https://smart.servier.com), licensed under CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/). Part of this illustration is from NIAID NIH BioArt Source (bioart.niaid.nih.gov/bioart/41). Abbreviations: Medi-Diet, Mediterranean diet; DASH, Dietary Interventions to Stop Hypertension; HDL, high density lipoprotein. Notes: ↑ denotes increase, ↓ denotes decrease. |
Mediterranean Diet
The Mediterranean diet (Medi-Diet) encompasses the dietary patterns, cultural practices, and culinary traditions of populations residing in the Mediterranean Basin.170 Extensive scientific research has demonstrated its significant health benefits, including its role in the prevention of conditions such as T2DM, CVDs, and MetS.171,172 Beyond its health advantages, recent studies have also highlighted the Medi-Diet’s positive impact on cultural and sustainability preservation.173 Labelled by UNESCO as an Intangible Cultural Heritage of Humanity,174 the Medi-Diet has been endorsed in the 2015–2020 American Dietary Guidelines as a model of a healthy dietary pattern.175 The Medi-Diet is predominantly plant-based, specified by high consumption of vegetables, particularly leafy greens, pulses, fruits, legumes, nuts, whole-grain cereals and extra virgin olive oil (EVOO) as the primary fat source. Traditional recipes often incorporate sauces such as sofrito, made with tomato, olive oil, onion, garlic or leek, which contain carotenoids and phenolic compounds like naringenin, lycopene, hydroxy-tyrosol, and β-carotene.176 Moderate consumption of fermented alcoholic drinks (red wine), typically consumed with meals, is another hallmark of the Medi-Diet, alongside low to moderate consumption of poultry and fish, and minimal intake of pastries, butter, red meat, sweets, and soft drinks.177,178 The traditional Medi-Diet is characterized by a low-carbohydrate, high-fat dietary composition, with fat contributing to total daily energy intake of 35–45%, approximately 40–45% from carbohydrates and 15% from protein.177 EVOO, a cornerstone of the Medi-Diet, is the prime source of monounsaturated fatty-acids (MUFAs) in Mediterranean countries, with oleic acid as its major component. Numerous studies have associated MUFA consumption with enhanced insulin sensitivity, improved blood lipid levels, and reductions in blood pressure levels (both systolic and diastolic), addressing critical risk factors for MetS.179 In addition to its MUFA content, EVOO contains abundant polyphenols, which exhibit antioxidant anti-inflammatory properties, further supporting endothelial function and improved lipid profiles.180 Overall, this dietary pattern provides a wide array of bioactive compounds, including antioxidant vitamins (β-carotene, C, and E), folates, phytochemicals, and essential minerals, which collectively contribute to its protective health effects.
Di Daniele et al documented a review examining the influence of adherence to the Medi-Diet on MetS criteria, dysfunction of adipose tissue, and obesity. Study results indicated that prescribing the Medi-Diet could serve as a therapeutic approach for MetS by reducing excess fat and mitigating inflammatory responses associated to obesity.171 Franquesa et al also highlighted strong evidence supporting the Medi-Diet’s efficacy in hampering obesity and MetS in both healthy individuals and those at high CVD risk, as well as decreasing mortality risk among obese individuals.172 A meta-analysis of 12 prospective and cross-sectional cohorts demonstrated that greater adherence to the Medi-Diet was related with a 19% reduction in MetS risk. Additionally, individual MetS components, such as blood pressure and waist circumference, showed improvements.169 Comparable protective effects have been identified in subsequent trials conducted in both Mediterranean and non-Mediterranean populations.181,182 In the CARDIA (Coronary Artery Risk Development in Young Adults) study, a prospective cohort of 4,713 individuals in the US comprising white and black populations, higher Medi-Diet consumption was linked to a reduced incidence of MetS compared to lower adherence subjects, with a significant linear trend across five adherence score categories.181 Similarly, in the SU.VI.MAX trial, a 6-year follow-up of 3,232 participants, Kesse-Guyot et al found that individuals in the highest tertile of Medi-Diet adherence had a 53% lower risk of developing MetS compared to the lowest tertile.182 Medi-Diet adherence was also linked with favorable changes in individual MetS components, such as, blood pressure, waist circumference, HDL-c and TG levels.182 Furthermore, studies in Korean subjects found lower MetS prevalence among those with medium to high adherence to Medi-Diet adherence.183
Furthermore, the Medi-Diet has been reported to reduce T2DM and CVD incidences, while also mitigating disease severity and complications in patients already diagnosed with these conditions.172,178 Given its demonstrated health benefits and simplicity of adherence, the Medi-Diet is recommended as a primary strategy for MetS management. A 2025 systematic review showed that dietary patterns influence gut microbiota composition in ways that align with cardiometabolic benefits, including reduced inflammation and improved lipid profiles, though heterogeneity in adherence and populations persists.184
DASH Diet
Dietary Approaches to Stop Hypertension (DASH) diet, in 1997, emerged as a promising intervention for managing high blood pressure,185 with subsequent randomized clinical trials (RCTs) reinforcing its efficacy.186 This dietary pattern emphasizes the consumption of whole grains, fruits, vegetables, low- or fat-free dairy products, nuts and legumes, while limiting sugar-sweetened beverages and red and processed meats consumption.185 The DASH diet features a low fat content (27% of daily caloric intake from fat), particularly saturated fats, and a cholesterol intake of approximately 150 mg/day. Additionally, it reduces sodium intake (ranging from 1500 to 2300 mg/day) while providing high levels of fiber (>30 g/day), calcium, magnesium, and potassium, with respect to other dietary approaches.187,188 It has demonstrated effectiveness as a therapeutic approach for hypertension,188–190 and numerous epidemiological analysis have linked DASH diet adherence to improved cardiometabolic health.191–195 A meta-analysis of multiple cohort studies by Schwingshackl et al found that increased DASH diet adherence was significantly correlated with reduced risks of all-cause mortality, T2DM incidence, and CVD incidence or mortality.196
Investigating the use of the DASH dietary approach for hypertension management, a meta-analysis of 30 randomized controlled trials involving 5,545 participants (both hypertensive and non-hypertensive), demonstrated that the DASH diet, combined with lifestyle modifications, significantly reduced blood pressure, compared to a control diet.186 These effects were more pronounced in individuals with sodium intake below 2,400 mg/day, participants under the age of 50, and hypertensive individuals not receiving antihypertensive medications.186 Furthermore, on comparing the DASH diet to 13 other dietary patterns (including the Mediterranean diet, low-sodium diet, Paleolithic diet, Nordic diet and low-fat diet) for their antihypertensive efficacy, the DASH diet emerged as the most effective in managing hypertension, particularly when compared to low-fat diets.190
The DASH diet has demonstrated potential benefits in reducing abdominal obesity and excess body weight.197 Adherence to the DASH diet by overweight or obese subjects resulted in greater weight reduction compared to other dietary patterns, with an average reduction of −3.63 kg. However, after one year of DASH intervention, the weight loss was slightly attenuated, averaging −3.08 kg.188 The results correlating the impact of the DASH diet on blood lipoproteins are inconsistent.188,189 Ge et al reported no significant alteration in LDL-c or HDL-c levels following a DASH dietary approach compared to a usual diet.185 Conversely, a meta-analysis by Siervo et al involving 1,917 subjects with CVD risk factors found reductions in total cholesterol and LDL-c levels, but observed no significant changes in TGs or HDL-c levels.189 Similar outcomes were noted in a controlled trial involving 80 patients with T2DM, where adherence to the DASH diet for 12 weeks resulted in significant reductions in TGs, total cholesterol, and VLDL levels, comparable to those achieved with an antidiabetic diet recommended by American Diabetes Association.198
Epidemiological studies indicate that following the DASH dietary approach vigorously is linked to an improved cardiometabolic profile, and a reduced risk of CVDs.194,199 A cross-sectional study involving 1,493 subjects found that following the DASH dietary routine was linked to 48% lower risk of MetS development. Indeed, humans with greater adherence exhibited lower pro-inflammatory markers, BMI, waist circumference, and measures of adiposity compared to those with lower adherence.191 Similarly, Asghari et al reported that in a cohort of 425 healthy adolescents and children aged 6–18 years, higher DASH diet adherence corresponded to a 64% reduced risk of MetS. This study also identified inverse relationships between adherence to the DASH diet and hypertension, abdominal obesity, and fasting plasma glucose levels.200 Furthermore, modifying the DASH diet to accommodate the glucose management needs of individuals with T1DM (by reducing carbohydrate content by ~10% and increasing fat intake by ~15%), improved glucose control and enhanced overall dietary quality, characterized by increased consumption of leafy vegetables, fiber, fruits, and protein compared to typical dietary intake.201
The health advantages of the DASH diet are likely attributed to its nutritional composition and distribution. This diet is abundant in fruits and vegetables, leading to increased intake of magnesium, potassium, and fiber—nutrients that have been shown to regulate insulin response, blood pressure and glucose metabolism.
Plant-Based Diets
Diets that emphasize plant-based products including vegetables, nuts, fruits, legumes, and grains while reducing or eliminating animal-based meals are known as plant-based diets (PBDs). Within this category, strict vegetarian diets, or vegan diets, exclude all animal-based meals, including eggs, honey and dairy. Lacto-vegetarian diets limit animal food intake but allow dairy products, while lacto-ovo-vegetarian foods exclude seafood, poultry and meat, but include dairy products and eggs. Pescatarians or pesco-vegetarians, follow a diet similar to lacto-ovo-vegetarians but also include fish.202
PBDs have been linked to favorable cardiometabolic outcomes, particularly a reduced risk of MetS development and its individual factors.203 These eating habits are linked to a decreased risk of T2DM, obesity, and CVDs, as well as a lower all-cause mortality.204,205 Several investigations have reported a reduced risk of ischemic heart disease induced mortality in vegetarians compared to non-vegetarians.206 Furthermore, a meta-analysis of seven controlled clinical trials demonstrated that individuals following a vegetarian diet sustained a mean decrease of 4.8 mmHg and 2.2 mmHg in systolic and in diastolic blood pressure, respectively, in comparison to those who followed an omnivorous diet.204 These findings were supported by another meta-analysis conducted by the same authors, which included 32 clinical studies with a total of 604 participants. This analysis showed significant associations between adherence to PBDs and decrease in both systolic (−6.9 mmHg) and diastolic blood pressure (−4.7 mmHg).207,208
The impact of PBDs on serum lipid concentrations remains a subject of debate. Wang et al documented a meta-analysis of 11 randomized controlled trials to assess the impact of a PBD on TGs, HDL-c, LDL-c and non-HDL-c levels.209 The results showed significant reductions in total cholesterol, HDL-c and LDL-c after adhering to vegetarian diet compared to an omnivorous control diet. Notably, no significant changes in TG levels were reported. This investigation also reported significant weight reduction in subjects following the vegetarian diet, with a mean reduction of −2.88 kg compared to those following an omnivorous diet. Similar results were documented in another meta-analysis study of 12 RCTs involving 1,151 participants, where individuals on the vegetarian diet demonstrated a significant reduction in body weight, compared to non-vegetarian subjects (mean difference: −2.02 kg).210
In summary, recent research highlights the protective effects of PBDs against CVDs, MetS and their associated risk components. However, the health benefits are contingent on the selection of nutrient-rich plant-based foods. Therefore, dietary guidelines ought to promote PBDs as a promising approach for the management of MetS.
Low-Carbohydrate Diet
Low-carbohydrate (low-CH) dietary patterns are defined by a reduction in CH intake to less than 50% of daily caloric consumption. These diets typically involve limiting the acquisition of processed foods, starches, refined grains, and foods high in simple or added sugars.211 The relationship between CH consumption and the incidence and/or management of MetS remains inconsistent.212 A meta-analysis of 18 clinical studies encompassing 69,554 individuals with MetS, consuming high intake of CH, by Liu et al reported a 2.5% elevation in MetS risk for every 5% rise in energy derived from CH.212 High intake of CH was also associated with adverse effects on the lipid profile, including elevated blood pressure, LDL-c, and TGs along with decreased HDL-c levels213. The health advantages reported in low-CH diets are partly attributed to the avoidance of rapidly absorbed CHs, such as refined grains and glucose, which can exacerbate insulin resistance and increase insulin demand. In management of T2DM, clinical guidelines emphasize individualized dietary approaches rather than prescribing specific carbohydrate restrictions.214
Bazzano et al performed a randomized controlled trial, in obese patients (BMI 30–45 kg/m2), to compare the weight lowering benefits of a low-CH diet with a low-fat diet.215 The study did not include energy restriction or physical activity guidance. Participants on the low-CH diet lost a considerably higher amount of weight after a year (−3.5 kg), mostly due to decreases in fat mass. Additionally, the low-CH group demonstrated improvements in CVD risk factors, including lower TG levels, increased HDL-c, and an improved total cholesterol-to-HDL ratio.215 In the context of T2DM, low-CH dietary interventions were associated with greater reductions in body weight, TGs, glycosylated hemoglobin (HbA1c), and blood pressure, along with increases in HDL-c, compared to low-fat diets. These changes were also accompanied by modifications in glucose-lowering medication regimens.216 Several studies have identified a correlation between modified CH intake and an increased risk of CVD events, including myocardial infarction and stroke.217,218 However, there remains insufficient scientific evidence to conclusively support the metabolic benefits of low-carbohydrate diets (<50–55% of total energy intake). More studies are needed to investigate the impact of low-CH diet on cardiometabolic events.
Among low-CH diets, very-low-CH ketogenic diets are being proposed to have a therapeutic role in managing MetS and associated factors including obesity, and CVDs.219,220 Although there is no universally accepted definition, ketogenic diets are typically characterized by CH intake comprising less than 10% of daily energy (equivalent to approximately 30–50 g /day) accompanied by a relative elevation in fat intake, with a fat-to- protein and CH ratio of 4:1 and 3:1, respectively.221 This highly restrictive dietary approach has demonstrated protective effects against CVD and obesity, primarily through improvements in lipid profiles and reductions in body weight.219,222,223
The meta-analysis conducted by Bueno et al reviewed that adherence to ketogenic diet caused significant weight loss, as well as reductions in TGs and blood pressure, in comparison to low-fat diets. Additionally, HDL-c levels increased in individuals following a ketogenic diet.224 The mechanisms underlying these benefits include the reduction of insulin secretion due to the absence of dietary CH intake, which inhibits lipogenesis and fat accumulation while promoting lipolysis. Other contributing factors are the satiating effects of protein consumption, which influence appetite-regulating hormones (ghrelin and leptin), and the modulation of ketone body and insulin production, which may enhance metabolic processes, particularly insulin signaling.219,225 Furthermore, the glycogen depletion and CH restriction associated with a ketogenic diet promotes the utilization of ketone bodies as a primary energy source. However, sustained production of ketone bodies requires energy restriction. Recent evidence indicates that the observed benefits of ketogenic diets on body weight and cardiovascular health are primarily attributable to energy restriction rather than the diet’s macronutrient composition.226 Healthcare professionals should also consider the challenges of adhering to a ketogenic diet, as well as the potential exclusion of nutrient-rich foods such as fruits, vegetables and whole grains. These foods are strongly linked with a lower risk of chronic diseases, including, T2DM, CVDs and certain cancers.
Low-Fat Diets
Low-fat diets (LFDs), are characterized by fat intake comprising less than 30% of total energy, with saturated fats (SFA) contributing less than 10%, moderate PUFA content, and minimal trans fatty acids.227 LFDs typically emphasize higher CH intake and moderate protein intake (approximately 15–17% of total energy). Foods included in LFDs often feature reduced-fat options (low-fat dairy), in place of whole-fat alternatives. In weight-loss interventions, LFDs have been associated with a reduced risk of premature death in overweight subjects.228 A meta-analysis of 34 randomized controlled trials reported an 18% reduced risk of all-cause mortality in such interventions, although no significant influence on CVD incidence or mortality were reported.228 Additionally, a network meta-analysis highlighted the potential of LFDs in reducing body weight compared to normal dietary habits or other dietary patterns, particularly after short-term interventions (6 months). However, this effect was found to diminish after one year.188
Clinical trials examining the impact of LFDs on the prevalence of MetS have produced inconsistent findings.229–232 Dietary interventions emphasizing low fat intake (approximately 20% of total energy from fat) demonstrated modest reductions in certain components of MetS, but showed no significant effects on CVDs outcomes or the prevalence of coronary heart disease in postmenopausal women compared to normal diets.229 Additionally, adherence to a LFD did not correlate with a reduced incidence of MetS in older individuals at high risk for CVD.232
Veum et al evaluated the impact of a high-CH, low-fat diet versus a low-CH, very high-fat diet (~73% of total energy from fat) on MetS components.233 The study reported no significant changes in MetS components, body composition or body weight, over the medium term. Similarly, the DIETFITS trial, conducted by Gardner et al, found that both low-CH and low-fat dietary interventions resulted in significant reduction in body weight, with no notable changes between the two approaches.234 LFDs intervention demonstrated short-term potential for managing hypertension and blood lipoproteins, including reduction in blood pressure (systolic and diastolic) and HDL-c and LDL-c level improvement levels, compared to usual diets. However, these effects diminish over the long term.188,190 A meta-analysis of RCTs involving 17,230 participants with pre-hypertension or hypertension by Schwingshackl et al found that the DASH diet and Medi-Diet are more potent than LFDs for long-term management of blood pressure.190 Regarding, T2DM management, a study by Basterra-Gortari et al documented that LFDs did not influence the need for managing glucose-lowering medications, whereas a Medi-Diet supplemented with EVOO was associated with a delayed requirement for such medications in older adults at high CVD risk.235 Notably, various studies have reported that LFD interventions may improve microbiome dysbiosis associated with MetS by enhancing microbial α-diversity.236 However, the available evidence is limited, and further investigations are required to understand the long-term impact of these interventions.
High-Protein Diet
Recent evidence indicates that dietary patterns rich in protein may result in significant weight reduction and improvements in CVD progression, compared to standard protein diets, which typically provide 0.8g of protein (per kg of body weight). In high-protein diets (HPDs), protein intake contributes around 20–30% of daily energy, equivalent to approximately 1.3–1.5 g of protein (per kg of body weight).237 These diets have been proposed for the treatment of MetS and associated factors including obesity and T2DM.238,239 However, the efficacy of HPDs for weight reduction remains debatable A meta-analysis report of 18 studies evaluating the impact of HPDs in T2DM subjects found no significant reductions in body weight in comparison to a standard protein diet.237 Similarly, no significant improvements were observed in glycemic indexes (fasting glucose, HbA1c and insulin), blood pressure and blood lipid levels. Despite this, a significant reduction in TG levels was reported among participants on HPDs. In individuals with MetS, HPDs combined with CH restriction have demonstrated effectiveness in weight management in obese adults with MetS.237,238
The Optimal Macronutrient Intake Trial to Prevent Heart Disease (OmniHeart) was a controlled, three-period crossover randomized study involving 164 subjects who were overweight or obese and had pre-hypertension or stage 1 hypertension, but were not diabetics.240 The trial sought to assess insulin sensitivity using the quantitative insulin sensitivity check index across three dietary interventions: a high-CH diet (58% of daily calories from carbohydrates, 27% from fat, and 15% from protein): a HPD (replacing 10% of total CH intake with protein, resulting in 25% of daily calories from protein, primarily plant-based); an unsaturated-fat diet (replacing 10% of total CH intake with unsaturated fats, leading to 37% of daily calories from fat, primarily sourced from seeds, olive oil, canola oil, safflower oil, and nuts). The study found no significant impact of the high-protein or high-CH diets on insulin sensitivity, whereas the unsaturated-fat diet improved insulin sensitivity. This suggests that replacing CHs with unsaturated fats, as seen in Medi-Diet patterns, may be a potent approach to enhance insulin sensitivity. The health benefits of HPDs are attributed to their ability to increase satiety, reducing subsequent energy intake.241 Additionally, intake of a HPD helps preserve muscle mass during restrictive dietary intake aimed at weight loss.242 Protein sources such as meat and related products have been linked to an elevated risk of CVD, T2DM, and MetS development.243 Consequently, dietary guidelines advocate for utilizing plant-based protein sources, including legumes, soy, beans, seeds and nuts, as healthier alternatives to meat and processed meat products.244
Collectively, dietary strategies such as the Mediterranean, DASH, and plant-based diets demonstrate consistent benefits in improving metabolic markers, often in parallel with favorable shifts in gut microbiota. Yet, results differ by population, adherence, and diet composition, and not all interventions translate into sustained microbiome changes. Importantly, many findings come from short-term or observational studies, underscoring the need for long-term, controlled trials that link dietary modulation of the microbiome to durable clinical outcomes.
Critical Evaluation of Current Evidences
While the preceding sections detail the association between gut microbiota (GM) dysbiosis and Metabolic Syndrome (MetS), the critical question is how dietary interventions can modulate this relationship. A mere listing of studies, however, is insufficient. A rigorous synthesis reveals that the efficacy of dietary strategies is not equal and is fundamentally linked to their capacity to restore a healthy GM and its functional output.
The most compelling evidence supports holistic dietary patterns over approaches focused on isolated macronutrient restriction. The Mediterranean and DASH diets consistently demonstrate superior, sustainable benefits across MetS components. Their effectiveness is likely rooted in their high content of fermentable fibers and polyphenols, which serve as substrates for commensal bacteria, stimulating the production of beneficial metabolites like short-chain fatty acids (SCFAs). These SCFAs, particularly butyrate, are not merely end-products but critical signaling molecules that enhance gut barrier integrity, reduce systemic inflammation, and improve insulin sensitivity. In contrast, macronutrient-manipulative diets like strict Low-Fat or Ketogenic diets show inconsistent results. While ketogenic diets may induce rapid weight loss, their severe carbohydrate restriction may paradoxically starve key SCFA-producing bacteria, potentially undermining long-term GM health and metabolic resilience. The failure of many low-fat diets to significantly improve MetS outcomes highlights that the type of fat—with a priority on unsaturated fats as in the Mediterranean diet—is more critical than its quantity alone.
Furthermore, the concept of “crosstalk” necessitates a critical evaluation of food quality and personalization. The benefits of a Plant-Based Diet are contingent on the consumption of nutrient-dense, minimally processed plant foods; a diet high in refined carbohydrates and plant-based ultra-processed foods will not confer the same GM and metabolic advantages. Emerging evidence from trials like PREDICT suggests that inter-individual variation in GM composition (eg, high Prevotella vs Bacteroides) determines responses to specific dietary fibers and fats. This challenges the one-size-fits-all model and positions the GM as a key biomarker for personalizing nutritional therapy. Therefore, future research must pivot from describing dietary outcomes to delineating the causal mechanisms by which these diets reshape the GM. Integrating multi-omics data from well-controlled dietary interventions is essential to move beyond association and toward a mechanistic, personalized prescription for MetS management through the gut microbiome.
Gut Microbiota Modulation as Therapeutic Approach for Metabolic Syndrome
The GM has emerged as a key target for live bacterial cell-based therapies in managing chronic diseases, including MetS.245 These therapies aim to establish healthy gut conditions by restoring bacterial balance and enhancing beneficial metabolic activities. Some of the primary therapeutic interventions used for modulating GM composition are discussed below.
Probiotics
Probiotics include live microorganisms, typically bacteria or fungi, that benefit the host’s health when taken in recommended quantities.246 Fermented foods contain probiotic microorganisms, which have long been a part of human diets. However, Western-style diets, which are frequently linked to the current obesity crisis, no longer include them as much.247 Meta-analyses have demonstrated that probiotic supplementation effectively improves key characteristics of T2DM, including pre-prandial plasma glucose levels, HbA1c, insulin concentration, and insulin resistance.248,249 Additionally, probiotics have been shown to enhance lipid profile parameters.250 Probiotic interventions have also been linked with modest improvements in BMI, body weight, and body fat percentage in individuals who are overweight or obese.251,252 Consequently, probiotics may hold potential as an adjunctive strategy for managing MetS, and related factors including diabetes and obesity. However, the existing data are inconsistent due to variability in study designs, probiotic formulations, intervention durations, and measured outcomes, making it difficult to reach definitive conclusions. Additionally, the underlying mechanisms responsible for the observed effects, as well as the role of the microbiome and its modulation, remain poorly understood. Probiotics have been evaluated in both single-strain and multi-strain or multi-species formulations, and in some cases, combined with a prebiotic component to form a symbiotic.32
A number of studies demonstrated the therapeutic impact of single and multi-strain probiotics in MetS. A single-strain formulation containing Lactobacillus reuteri V3401 demonstrated reductions in sVCAM, IL-6, and insulin levels in patients newly diagnosed with MetS following 12 weeks of consumption. Additionally, it increased the prevalence of Akkermansia muciniphila/Verrucomicrobia.253 In treatment-naive patients with T2DM, Lactobacillus reuteri ADR-1 reduced HbA1c, while heat-killed L. reuteri ADR-3 lowered mean arterial pressure, and IL-1β levels, and raised the Bifidobacteria abundance. Both interventions enriched the gut microbiome with the administered probiotic strain, but no additional effects on the overall microbiome composition were observed.254 Moreover, in a 12-week intervention involving obese adults, Lactobacillus plantarum K50, a probiotic strain obtained from kimchi, significantly reduced total cholesterol and TG levels but did not affect body composition or body weight. The intervention enhanced the abundance of L. plantarum, the genus Enterococcus hirae and Enterococcus, a bacterium with potential health benefits,255 while decreasing the abundance of Actinobacteria.256 In prediabetic adults, Lactobacillus plantarum HAC01 decreased 2-hour postprandial glucose (2h-PPG) and HbA1c levels without altering the GM. In contrast, L. plantarum Dad-13 decreased BMI and body weight in obese individuals while modulating the microbiome by decreasing the abundance of Firmicutes and increasing Bacteroidetes.
Regarding multi-strain/multi-species probiotics, a combination of Lactobacillus rhamnosus HN001 and Bifidobacterium longum BB536, when paired with a Medi-Diet, resulted in reduced waist-to-hip ratio, waist circumference and insulin levels, in obese female breast cancer survivors. Additionally, it increased alpha diversity and the abundance of L-Ruminococcus, and Eubacterium, while decreasing the levels of Butyricicoccus and Bacteroides in the microbiome, compared to the Medi-Diet alone.257 The combination of two Bifidobacterium breve strains (B632 and BR03) led to reductions in waist circumference, BMI standard deviation score, pre-meal insulin, and ALT levels in obese children and adolescents participating in a weight loss program. However, the changes observed in the microbiome were comparable to those in the placebo group.258
Prebiotics
Prebiotics, are characterized as substrates which are utilized selectively by host microorganisms to confer a health benefit.259 Their beneficial physiological effects were first recognized over two decades ago, however, recently interest in prebiotics has increased significantly.260 The most well-known prebiotics today include inulin, fructo-oligosaccharides, oligofructose, galacto-oligosaccharides, and resistant starch, all of which for example, have been reported to stimulate the growth of Lactobacilli and Bifidobacteria261. Given that individuals with excess weight typically exhibit a reduced bacterial diversity in their gut microbiome, particularly a decrease in Bifidobacteria abundance, the use of prebiotics to restore the gut microbiome’s balance and function has been proposed as a promising therapeutic approach.262 Resistant starches, inulin, and fructo-oligosaccharide-enriched inulin may provide beneficial effects on limiting the components of MetS.262 Supplementation with trans-galactooligosaccharides (GOS) has been shown to improve insulin and lipid metabolism, increase Bifidobacteria abundance, and decrease the levels of Gram-negative bacteria such as Desulfovibrio spp., Bacteroides spp., and the Clostridium histolyticum group.263 Alpha-GOS resulted in a dose-dependent reduction in food intake and appetite.264 GOS supplementation specifically raised the number of Bifidobacterium species, but it had no discernible effect on pre-diabetic patients’ energy and substrate metabolism or peripheral insulin sensitivity.265 Notably, supplementation with inulin-type fructans resulted in a slight reduction in fat mass, but did not affect lipid levels glucose homeostasis and BMI.266 Additionally, inulin had a positive impact on the GM composition.267 Supplementation with resistant starch type 4 (RS4) positively influenced lipid metabolism, likely through the modulation of bile acid metabolism and changes in composition of GM, but did not affect glucose metabolism.268,269 The inclusion of polyphenolic food components enhanced the prevalence of butyrate-producing gut bacteria and altered the microbiome composition; however, these effects were influenced mostly by the concurrent medication of the patients.270,271 Overall, prebiotics appear to alter the composition of the gut microbiome, but their effects on markers of body weight and MetS are infrequent. The factors that may influence their effectiveness such as timing, dosage, duration of treatment, the type of intervention, and potential combinations with other established interventions remain unclear. A 2023 meta-regression of randomized clinical trials confirmed that microbiome-modulating therapies modestly improve fasting glucose and insulin, though results for HbA1c and insulin resistance remain inconsistent.272
Fecal Microbiota Transplantation
Fecal microbiota transplantation (FMT), which involves transferring fecal matter from one human to another to enhance or restore the gut microbiome composition and treat a disease, has garnered significant interest both in the scientific community and among the general public. The first evidence of FMT effectiveness in MetS was reported in 2012. A pilot clinical trial conducted in the Netherlands revealed that insulin resistance was reduced in nine male subjects with MetS, who received FMT from a lean donor, while no changes were reported in the nine control participants who underwent autologous FMT.273 Following the initial proof-of-concept trials, several clinical investigations have been carried out to assess the potential of FMT in improving metabolism and reducing weight in conditions such as obesity, diabetes, non-alcoholic fatty liver disease, and MetS. However, in randomized, placebo-controlled trials, results regarding improvements in metabolic parameters have been inconsistent. While most studies observed alterations in microbiome composition, prominent effects on clinically significant outcomes, such as insulin resistance or body weight remain lacking. Despite FMT’s exceptional capacity to transmit the entire GM and its byproducts from healthy, lean donors to recipients, opportunistic pathogenic microorganisms may be introduced due to inadequate donor screening and fecal material analysis, potentially exacerbating dysbiosis in obese recipients and leading to serious side effects, such as vomiting and sepsis. These safety issues raise questions about the use of FMT.274,275 Despite its promising therapeutic effect, recent clinical studies have reported that FMT does not produce significant changes in metabolic profiles or induce weight loss.98,276 These contradicting results highlight the need for further research. Beyond diet and microbial therapies, a 2025 meta-analysis demonstrated that structured exercise interventions significantly increased microbiota diversity in individuals with obesity and T2DM, suggesting synergistic benefits when combined with dietary approaches.277
In summary, probiotics, prebiotics, and FMT show promise for modulating gut microbiota in metabolic syndrome, but clinical results remain inconsistent. Variability in strains, formulations, and study design limits comparability, and mechanistic understanding is still incomplete. Current evidence supports adjunctive use rather than stand-alone therapy, and more rigorously designed trials are required to confirm efficacy and safety.
Critical Evaluation of Current Evidence
The exploration of direct microbiome modulation as a therapeutic strategy for MetS is a logical progression from dietary insights, yet the evidence demands a critical and nuanced interpretation. While the potential of probiotics, prebiotics, and fecal microbiota transplantation (FMT) is considerable, the field is currently characterized by promising yet inconsistent results, underscoring a transition from proof-of-concept to a need for precision.
Probiotic interventions demonstrate that specific microbial strains can impart targeted metabolic benefits, such as improvements in glycemic control or lipid profiles. However, a critical synthesis reveals that these effects are often strain-specific, dose-dependent, and transient, with minimal long-term impact on the overall ecosystem of the native gut microbiota. The common failure of administered probiotics to durably colonize the gut suggests their mode of action may be more pharmacological than ecological, exerting effects through direct interaction with host immunity or the gut epithelium rather than through permanent restructuring of the microbial community. Similarly, prebiotics, by providing selective substrates, can enrich beneficial native taxa like Bifidobacterium and Lactobacillus. Yet, the clinical translation of these shifts into robust, reproducible improvements in MetS components remains inconsistent. The variable outcomes highlight a fundamental challenge: the individualized nature of the baseline gut microbiome, which dictates whether a given prebiotic will find the necessary bacterial strains to utilize it effectively, leading to the desired metabolic outcome.
FMT represents the most ambitious approach, attempting to reset the entire microbial community. The initial, dramatic results in insulin-sensitive subjects receiving lean donor FMT generated warranted excitement. However, subsequent randomized controlled trials have largely failed to replicate significant, sustained metabolic improvements, particularly regarding body weight. This discrepancy critically exposes our limited understanding of the complex ecology involved. Simply transferring a “healthy” microbiome may be insufficient if the host’s diet, genetics, and immune system do not support the engraftment and function of the keystone taxa. Furthermore, the safety concerns and logistical hurdles of FMT render it an impractical broad-scale solution for MetS. The collective evidence suggests that future therapies must move beyond broad-stroke.
Conflicting Evidence and Variability Across Studies
Despite the accumulating evidence linking gut microbiota dysbiosis to MetS, findings across studies are not always consistent. Variability in microbial signatures has been observed across populations, with some cohorts reporting phylum-level shifts (eg, Firmicutes-to-Bacteroidetes ratio) while others fail to replicate these associations. Such discrepancies likely stem from geographic and ethnic differences, dietary exposures, host genetic background, and variation in sequencing platforms and bioinformatics pipelines. These inconsistencies highlight the challenges in defining universal microbial biomarkers for MetS and underscore the need for harmonized protocols, standardized reference databases, and multi-cohort longitudinal studies. Acknowledging such heterogeneity is crucial for translating microbiome discoveries into clinically meaningful interventions.
Conclusion and Future Perspectives
Metabolic syndrome (MetS) and its associated comorbidities represent a substantial global health and economic burden. Accumulating evidence underscores a critical role of gut microbiota (GM) dysbiosis in the onset and progression of MetS and its key components, including type 2 diabetes mellitus (T2DM), obesity, and cardiovascular diseases (CVDs). Mechanistically, the gut microbiome contributes to MetS pathophysiology through modulation of lipid metabolism, chronic low-grade inflammation, host nutrient absorption, and the production of bioactive microbial metabolites. Notably, dietary patterns are major modulators of GM composition and function. Comparative studies indicate that the Mediterranean diet (Medi-Diet) represents a well-supported dietary strategy for the prevention and management of MetS.
Therapeutic manipulation of the GM—including probiotics, prebiotics, and fecal microbiota transplantation (FMT)—emerges as a promising avenue for targeted interventions. However, these approaches remain in the experimental stage, and robust clinical validation is required before integration into standard care. The complexity and functional breadth of the intestinal microbiome are only partially understood, and current insights have yet to be fully translated into evidence-based clinical applications.
Current literature highlights that the field of GM–metabolism interactions remains nascent, both at the mechanistic and translational levels. Key knowledge gaps persist, including the characterization of numerous unannotated metabolites and peptides in various biofluids, and the differentiation of host-, diet-, or microbiota-derived compounds. Most studies provide static “snapshots” of microbiome composition, offering limited insight into temporal dynamics and the long-term stability of microbial communities. Furthermore, translational utility is constrained by the lack of animal models that accurately recapitulate human genetic susceptibilities.
Bridging these gaps will require integrative approaches combining longitudinal multi-omics analyses with bio-clinical variables to define causal mechanisms. In particular, elucidating specific microbiome-driven pathogenic pathways in MetS could enable the rational design of precision microbiome-targeted therapies. Overall, advancing our understanding of GM–host interactions holds transformative potential for metabolic disease prevention and treatment, but achieving clinical translation demands rigorous, mechanistically informed research.
Acknowledgments
Parts of the figures were generated using Servier Medical Art, provided by Servier, licensed under CC BY 4.0 (available at https://creativecommons.org/licenses/by/4.0/) and NIH BIOART Source (https://bioart.niaid.nih.gov/).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Li W, Chen D, Peng Y, Lu Z, Kwan M-P, Tse LA. Association Between Metabolic Syndrome and Mortality: prospective Cohort Study. JMIR Public Health and Surveillance. 2023;9:e44073. doi:10.2196/44073
2. Pucci G, Alcidi R, Tap L, Battista F, Mattace-Raso F, Schillaci G. Sex- and gender-related prevalence, cardiovascular risk and therapeutic approach in metabolic syndrome: a review of the literature. Pharmacol Res. 2017;120:34–42. doi:10.1016/j.phrs.2017.03.008
3. Alberti KGMM, Zimmet P, Shaw J. The metabolic syndrome—a new worldwide definition. Lancet. 2005;366(9491):1059–1062. doi:10.1016/S0140-6736(05)67402-8
4. Swarup S, Ahmed I, Grigorova Y, Zeltser R. Metabolic Syndrome. In: StatPearls. Treasure Island (FL): StatPearls Publishing; 2024.
5. Rizzo M, Rizvi AA. New Advances in Metabolic Syndrome. Int J Mol Sci. 2024;25(15):8311. doi:10.3390/ijms25158311
6. Pérez-Martínez P, Mikhailidis DP, Athyros VG, et al. Lifestyle recommendations for the prevention and management of metabolic syndrome: an international panel recommendation. Nutr Rev. 2017;75(5):307–326. doi:10.1093/nutrit/nux014
7. Noubiap JJ, Nansseu JR, Lontchi-Yimagou E, et al. Global, regional, and country estimates of metabolic syndrome burden in children and adolescents in 2020: a systematic review and modelling analysis. Lancet Child Adolesc Health. 2022;6(3):158–170. doi:10.1016/S2352-4642(21)00374-6
8. Noubiap JJ, Nansseu JR, Lontchi-Yimagou E, et al. Geographic distribution of metabolic syndrome and its components in the general adult population: a meta-analysis of global data from 28 million individuals. Diabet Res Clin Pract. 2022;188:109924. doi:10.1016/j.diabres.2022.109924
9. Cani PD, Delzenne NM. Gut microflora as a target for energy and metabolic homeostasis. Curr Opin Clin Nutr Metab Care. 2007;10(6):729–734. doi:10.1097/MCO.0b013e3282efdebb
10. Dupont HL, Jiang Z-D, Dupont AW, Utay NS. THE INTESTINAL MICROBIOME IN HUMAN HEALTH AND DISEASE. Transact Am Clin Climatol Assoc. 2020;131:178.
11. Cӑtoi AF, Corina A, Katsiki N, et al. Gut microbiota and aging-A focus on centenarians. Biochim Biophys Acta Mol Basis Dis. 2020;1866(7):165765. doi:10.1016/j.bbadis.2020.165765
12. Dabke K, Hendrick G, Devkota S. The gut microbiome and metabolic syndrome. J Clin Invest. 2019;129(10):4050. doi:10.1172/JCI129194
13. Rochlani Y, Pothineni NV, Kovelamudi S, Mehta JL. Metabolic syndrome: pathophysiology, management, and modulation by natural compounds. Therap Adv Cardiovascular Dis. 2017;11(8):215. doi:10.1177/1753944717711379
14. Cani PD, Amar J, Iglesias MA, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56(7):1761–1772. doi:10.2337/db06-1491
15. Ursell LK, Haiser HJ, Treuren WV, et al. The Intestinal Metabolome: an Intersection Between Microbiota and Host. Gastroenterology. 2014;146(6):1470. doi:10.1053/j.gastro.2014.03.001
16. Hou K, Wu Z-X, Chen X-Y, et al. Microbiota in health and diseases. Sig Transduct Target Ther. 2022;7:1–28. doi:10.1038/s41392-022-00974-4
17. Donia MS, Fischbach MA. Small Molecules from the Human Microbiota. Science. 2015;349(6246):1254766. doi:10.1126/science.1254766
18. Han S, Van Treuren W, Fischer CR, et al. A metabolomics pipeline for the mechanistic interrogation of the gut microbiome. Nature. 2021;595(7867):415–420. doi:10.1038/s41586-021-03707-9
19. Laterza L, Rizzatti G, Gaetani E, Chiusolo P, Gasbarrini A. The Gut Microbiota and Immune System Relationship in Human Graft-versus-Host Disease. Mediterr J Hematol Infect Dis. 2016;8:e2016025. doi:10.4084/MJHID.2016.025
20. Auchtung TA, Fofanova TY, Stewart CJ, et al. Investigating Colonization of the Healthy Adult Gastrointestinal Tract by Fungi. mSphere. 2018;3(2):e00092–18. doi:10.1128/mSphere.00092-18
21. Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022–1023. doi:10.1038/4441022a
22. Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021;19(1):55–71. doi:10.1038/s41579-020-0433-9
23. Hillman ET, Lu H, Yao T, Nakatsu CH. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017;32(4):300. doi:10.1264/jsme2.ME17017
24. Crudele L, Gadaleta RM, Cariello M, Moschetta A. Gut microbiota in the pathogenesis and therapeutic approaches of diabetes. EBioMedicine. 2023;97:104821. doi:10.1016/j.ebiom.2023.104821
25. Flint HJ, Scott KP, Louis P, Duncan SH. The role of the gut microbiota in nutrition and health. Nat Rev Gastroenterol Hepatol. 2012;9:577–589. doi:10.1038/nrgastro.2012.156
26. Milani C, Duranti S, Bottacini F, et al. The First Microbial Colonizers of the Human Gut: composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev. 2017;81:e00036–17. doi:10.1128/MMBR.00036-17
27. Rinninella E, Raoul P, Cintoni M, et al. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms. 2019;7:14. doi:10.3390/microorganisms7010014
28. Amabebe E, Robert FO, Agbalalah T, Orubu ESF. Microbial dysbiosis-induced obesity: role of gut microbiota in homoeostasis of energy metabolism. Br J Nutr. 2020;123:1127–1137. doi:10.1017/S0007114520000380
29. Yatsunenko T, Rey FE, Manary MJ, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486:222–227. doi:10.1038/nature11053
30. Guigoz Y, Doré J, Schiffrin EJ. The inflammatory status of old age can be nurtured from the intestinal environment. Curr Opin Clin Nutr Metab Care. 2008;11:13–20. doi:10.1097/MCO.0b013e3282f2bfdf
31. Fujisaka S, Watanabe Y, Tobe K. The gut microbiome: a core regulator of metabolism. J Endocrinol. 2023;256. doi:10.1530/JOE-22-0111
32. Horvath A, Zukauskaite K, Hazia O, Balazs I, Stadlbauer V. Human gut microbiome: therapeutic opportunities for metabolic syndrome—Hype or hope? Endocrinol Diabetes Metabol. 2024;7:e436. doi:10.1002/edm2.436
33. El-Tanani M, Rabbani SA, El-Tanani Y, Matalka II. Metabolic vulnerabilities in cancer: a new therapeutic strategy. Crit rev oncol/hematol. 2024;201:104438. doi:10.1016/j.critrevonc.2024.104438
34. Zimmet P, Alberti KG, Magliano DJ, Bennett PH. Diabetes mellitus statistics on prevalence and mortality: facts and fallacies. Nat Rev Endocrinol. 2016;12:616–622. doi:10.1038/nrendo.2016.105
35. Pereira SG, Moura J, Carvalho E, Empadinhas N. Microbiota of Chronic Diabetic Wounds: ecology, Impact, and Potential for Innovative Treatment Strategies. Front Microbiol. 2017;8:1791. doi:10.3389/fmicb.2017.01791
36. Ojo OA, Ibrahim HS, Rotimi DE, Ogunlakin AD, Ojo AB. Diabetes mellitus: from molecular mechanism to pathophysiology and pharmacology. Medicine Novel Technol Devices. 2023;19:100247. doi:10.1016/j.medntd.2023.100247
37. Janez A, Muzurovic E, Bogdanski P, et al. Modern Management of Cardiometabolic Continuum: from Overweight/Obesity to Prediabetes/Type 2 Diabetes Mellitus. Recommendations from the Eastern and Southern Europe Diabetes and Obesity Expert Group. Diabetes Ther. 2024;15:1865–1892. doi:10.1007/s13300-024-01615-5
38. Auvinen A-M, Luiro K, Jokelainen J, et al. Type 1 and type 2 diabetes after gestational diabetes: a 23 year cohort study. Diabetologia. 2020;63:2123. doi:10.1007/s00125-020-05215-3
39. Kang N, Chen G, Tu R, et al. Adverse associations of different obesity measures and the interactions with long-term exposure to air pollutants with prevalent type 2 diabetes mellitus: the Henan Rural Cohort study. Environ Res. 2022;207:112640. doi:10.1016/j.envres.2021.112640
40. Zhao L, Lou H, Peng Y, Chen S, Zhang Y, Li X. Comprehensive relationships between gut microbiome and faecal metabolome in individuals with type 2 diabetes and its complications. Endocrine. 2019;66:526–537. doi:10.1007/s12020-019-02103-8
41. Zhernakova A, Bonder MJ, Tigchelaar EF, et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science. 2016;352:565–569. doi:10.1126/science.aad3369
42. Larsen N, Vogensen FK, van den Berg FWJ, et al. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One. 2010;5:e9085. doi:10.1371/journal.pone.0009085
43. Qin J, Li Y, Cai Z, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. 2012;490:55–60. doi:10.1038/nature11450
44. Allin KH, Tremaroli V, Caesar R, et al. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia. 2018;61:810–820. doi:10.1007/s00125-018-4550-1
45. Everard A, Belzer C, Geurts L, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci U S A. 2013;110:9066–9071. doi:10.1073/pnas.1219451110
46. Gomes JMG, Costa JDA, Alfenas RDCG. Metabolic endotoxemia and diabetes mellitus: a systematic review. Metabolism. 2017;68:133–144. doi:10.1016/j.metabol.2016.12.009
47. Boroni Moreira AP, de Cássia Gonçalves Alfenas R. The influence of endotoxemia on the molecular mechanisms of insulin resistance. Nutr Hosp. 2012;27:382–390. doi:10.1590/S0212-16112012000200007
48. Shapiro H, Kolodziejczyk AA, Halstuch D, Elinav E. Bile acids in glucose metabolism in health and disease. J Exp Med. 2018;215:383. doi:10.1084/jem.20171965
49. Psichas A, Sleeth ML, Murphy KG, et al. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int J Obes Lond. 2015;39:424–429. doi:10.1038/ijo.2014.153
50. Takahashi D, Hoshina N, Kabumoto Y, et al. Microbiota-derived butyrate limits the autoimmune response by promoting the differentiation of follicular regulatory T cells. EBioMedicine. 2020;58:102913. doi:10.1016/j.ebiom.2020.102913
51. Zeng Y, Mtintsilana A, Goedecke JH, Micklesfield LK, Olsson T, Chorell E. Alterations in the metabolism of phospholipids, bile acids and branched-chain amino acids predicts development of type 2 diabetes in black South African women: a prospective cohort study. Metabolism. 2019;95:57–64. doi:10.1016/j.metabol.2019.04.001
52. Pedersen HK, Gudmundsdottir V, Nielsen HB, et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature. 2016;535:376–381. doi:10.1038/nature18646
53. Maguire M, Maguire G. The role of microbiota, and probiotics and prebiotics in skin health. Arch Dermatol Res. 2017;309:411–421. doi:10.1007/s00403-017-1750-3
54. Cox AJ, Zhang P, Bowden DW, et al. Increased intestinal permeability as a risk factor for type 2 diabetes. Diabetes Metab. 2017;43:163–166. doi:10.1016/j.diabet.2016.09.004
55. Sudo N, Chida Y, Aiba Y, et al. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol. 2004;558:263–275. doi:10.1113/jphysiol.2004.063388
56. de Goffau MC, Fuentes S, van den Bogert B, et al. Aberrant gut microbiota composition at the onset of type 1 diabetes in young children. Diabetologia. 2014;57:1569–1577. doi:10.1007/s00125-014-3274-0
57. Pellegrini S, Sordi V, Bolla AM, et al. Duodenal Mucosa of Patients With Type 1 Diabetes Shows Distinctive Inflammatory Profile and Microbiota. J Clin Endocrinol Metab. 2017;102:1468–1477. doi:10.1210/jc.2016-3222
58. Vatanen T, Franzosa EA, Schwager R, et al. The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature. 2018;562:589–594. doi:10.1038/s41586-018-0620-2
59. Zheng P, Li Z, Zhou Z. Gut microbiome in type 1 diabetes: a comprehensive review. Diabetes/Metab Res Rev. 2018;34:e3043. doi:10.1002/dmrr.3043
60. Karlsson FH, Tremaroli V, Nookaew I, et al. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature. 2013;498:99–103. doi:10.1038/nature12198
61. Chen Y-L, Qiao Y-C, Pan Y-H, et al. Correlation between serum interleukin-6 level and type 1 diabetes mellitus: a systematic review and meta-analysis. Cytokine. 2017;94:14–20. doi:10.1016/j.cyto.2017.01.002
62. Leiva-Gea I, Sánchez-Alcoholado L, Martín-Tejedor B, et al. Gut Microbiota Differs in Composition and Functionality Between Children With Type 1 Diabetes and MODY2 and Healthy Control Subjects: a Case-Control Study. Diabetes Care. 2018;41:2385–2395. doi:10.2337/dc18-0253
63. Acosta-Montaño P, Rodríguez-Velázquez E, Frayde-Gómez H, et al. Fatty Acid and Lipopolysaccharide Effect on Beta Cells Proteostasis and its Impact on Insulin Secretion. Cells. 2019;8:884. doi:10.3390/cells8080884
64. Devaraj S, Dasu MR, Park SH, Jialal I. Increased levels of ligands of Toll-like receptors 2 and 4 in type 1 diabetes. Diabetologia. 2009;52:1665–1668. doi:10.1007/s00125-009-1394-8
65. Gülden E, Ihira M, Ohashi A, et al. Toll-like receptor 4 deficiency accelerates the development of insulin-deficient diabetes in non-obese diabetic mice. PLoS One. 2013;8:e75385. doi:10.1371/journal.pone.0075385
66. Li J, Wang X, Zhang F, Yin H. Toll-like receptors as therapeutic targets for autoimmune connective tissue diseases. Pharmacol Ther. 2013;138:441–451. doi:10.1016/j.pharmthera.2013.03.003
67. Wang J, Cao H, Wang H, et al. Multiple mechanisms involved in diabetes protection by lipopolysaccharide in non-obese diabetic mice. Toxicol Appl Pharmacol. 2015;285:149–158. doi:10.1016/j.taap.2015.04.006
68. Radzicka S, Pietryga M, Iciek R, Brązert J. The role of visfatin in pathogenesis of gestational diabetes (GDM). Ginekol Pol. 2018;89:518–521. doi:10.5603/GP.a2018.0088
69. Gomez-Arango LF, Barrett HL, McIntyre HD, et al. Connections Between the Gut Microbiome and Metabolic Hormones in Early Pregnancy in Overweight and Obese Women. Diabetes. 2016;65:2214–2223. doi:10.2337/db16-0278
70. Koren O, Goodrich JK, Cullender TC, et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell. 2012;150:470–480. doi:10.1016/j.cell.2012.07.008
71. Cortez RV, Taddei CR, Sparvoli LG, et al. Microbiome and its relation to gestational diabetes. Endocrine. 2019;64:254–264. doi:10.1007/s12020-018-1813-z
72. Crusell MKW, Hansen TH, Nielsen T, et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome. 2018;6:89. doi:10.1186/s40168-018-0472-x
73. Ponzo V, Ferrocino I, Zarovska A, et al. The microbiota composition of the offspring of patients with gestational diabetes mellitus (GDM). PLoS One. 2019;14:e0226545. doi:10.1371/journal.pone.0226545
74. Wang J, Shi W, Du N, et al. Dysbiosis of maternal and neonatal microbiota associated with gestational diabetes mellitus. Gut. 2018;67:1614–1625. doi:10.1136/gutjnl-2018-315988
75. GBD 2015 Obesity Collaborators. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N Engl J Med. 2017;377:13–27. doi:10.1056/NEJMoa1614362
76. Kelly T, Yang W, Chen C-S, Reynolds K, He J. Global burden of obesity in 2005 and projections to 2030. Int J Obes. 2008;32:1431–1437. doi:10.1038/ijo.2008.102
77. Islam ANMS, Sultana H, Nazmul Hassan Refat MD, Farhana Z, Abdulbasah Kamil A, Meshbahur Rahman M. The global burden of overweight-obesity and its association with economic status, benefiting from STEPs survey of WHO member states: a meta-analysis. Prevent Med Rep. 2024;46:102882. doi:10.1016/j.pmedr.2024.102882
78. Saltiel AR, Olefsky JM. Inflammatory mechanisms linking obesity and metabolic disease. J Clin Invest. 2017;127:1–4. doi:10.1172/JCI92035
79. El-Tanani M, Rabbani SA, Aljabali AA, et al. The Complex Connection between Obesity and Cancer: signaling Pathways and Therapeutic Implications. Nutr Cancer. 2024;76:683–706. doi:10.1080/01635581.2024.2361964
80. Sonnenburg JL, Bäckhed F. Diet–microbiota interactions as moderators of human metabolism. Nature. 2016;535:56–64. doi:10.1038/nature18846
81. Gomes AC, Hoffmann C, Mota JF. The human gut microbiota: metabolism and perspective in obesity. Gut Microbes. 2018;9:308–325. doi:10.1080/19490976.2018.1465157
82. Weiss GA, Hennet T. Mechanisms and consequences of intestinal dysbiosis. Cell Mol Life Sci. 2017;74:2959–2977. doi:10.1007/s00018-017-2509-x
83. Roland BC, Lee D, Miller LS, et al. Obesity increases the risk of small intestinal bacterial overgrowth (SIBO). Neurogastroenterol Motil. 2018;30. doi:10.1111/nmo.13199
84. Vetrani C, Di Nisio A, Paschou SA, et al. From Gut Microbiota through Low-Grade Inflammation to Obesity: key Players and Potential Targets. Nutrients. 2022;14:2103. doi:10.3390/nu14102103
85. Sarmiento-Andrade Y, Suárez R, Quintero B, Garrochamba K, Chapela SP. Gut microbiota and obesity: new insights. Front Nutr. 2022;9. doi:10.3389/fnut.2022.1018212
86. Schwiertz A, Taras D, Schäfer K, et al. Microbiota and SCFA in lean and overweight healthy subjects. Obesity. 2010;18:190–195. doi:10.1038/oby.2009.167
87. Magne F, Gotteland M, Gauthier L, et al. The Firmicutes/Bacteroidetes Ratio: a Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients. 2020;12:1474. doi:10.3390/nu12051474
88. Sze MA, Schloss PD. Looking for a Signal in the Noise: revisiting Obesity and the Microbiome. mBio. 2016;7:e01018–16. doi:10.1128/mBio.01018-16
89. Houtman TA, Eckermann HA, Smidt H, de Weerth C. Gut microbiota and BMI throughout childhood: the role of firmicutes, bacteroidetes, and short-chain fatty acid producers. Sci Rep. 2022;12:3140. doi:10.1038/s41598-022-07176-6
90. Wang T, Yu L, Xu C, et al. Chronic fatigue syndrome patients have alterations in their oral microbiome composition and function. PLoS One. 2018;13:e0203503. doi:10.1371/journal.pone.0203503
91. Dhanasekaran D, Venkatesan M, Sabarathinam S. Efficacy of microbiome-targeted interventions in obesity management-A comprehensive systematic review. Diabetes Metab Syndr. 2025;18:103208.
92. Waters JL, Ley RE. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019;17:83. doi:10.1186/s12915-019-0699-4
93. Depommier C, Everard A, Druart C, et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med. 2019;25:1096–1103. doi:10.1038/s41591-019-0495-2
94. Crovesy L, Ostrowski M, Ferreira DMTP, Rosado EL, Soares-Mota M. Effect of Lactobacillus on body weight and body fat in overweight subjects: a systematic review of randomized controlled clinical trials. Int J Obes Lond. 2017;41:1607–1614. doi:10.1038/ijo.2017.161
95. Yin Y-N, Yu Q-F, Fu N, Liu X-W, Lu F-G. Effects of four Bifidobacteria on obesity in high-fat diet induced rats. World J Gastroenterol. 2010;16:3394–3401. doi:10.3748/wjg.v16.i27.3394
96. Waldram A, Holmes E, Wang Y, et al. Top-down systems biology modeling of host metabotype-microbiome associations in obese rodents. J Proteome Res. 2009;8:2361–2375. doi:10.1021/pr8009885
97. Cӑtoi AF, Vodnar DC, Corina A, et al. Gut Microbiota, Obesity and Bariatric Surgery: current Knowledge and Future Perspectives. Curr Pharm Des. 2019;25:2038–2050. doi:10.2174/1381612825666190708190437
98. Zhuang Z, Zhou P, Wang J, Lu X, Chen Y. The Characteristics, Mechanisms and Therapeutics: exploring the Role of Gut Microbiota in Obesity. Diabetes Metab Syndr Obes. 2023;16:3691–3705. doi:10.2147/DMSO.S432344
99. Krajmalnik-Brown R, Ilhan Z-E, Kang D-W, DiBaise JK. Effects of gut microbes on nutrient absorption and energy regulation. Nutr Clin Pract. 2012;27:201–214. doi:10.1177/0884533611436116
100. Mithieux G. New data and concepts on glutamine and glucose metabolism in the gut. Curr Opin Clin Nutr Metab Care. 2001;4:267–271. doi:10.1097/00075197-200107000-00004
101. Brahe LK, Astrup A, Larsen LH. Is butyrate the link between diet, intestinal microbiota and obesity-related metabolic diseases? Obes Rev. 2013;14:950–959. doi:10.1111/obr.12068
102. Mandard S, Zandbergen F, van Straten E, et al. The fasting-induced adipose factor/angiopoietin-like protein 4 is physically associated with lipoproteins and governs plasma lipid levels and adiposity. J Biol Chem. 2006;281:934–944. doi:10.1074/jbc.M506519200
103. Lin HV, Frassetto A, Kowalik EJ, et al. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS One. 2012;7:e35240. doi:10.1371/journal.pone.0035240
104. Blaak EE, Theis S, Frost G, et al. Short chain fatty acids in human gut and metabolic health. Benef Microbes. 2020;11:411–455. doi:10.3920/BM2020.0057
105. De la cuesta-zuluaga J, Mueller NT, Álvarez-quintero R, et al. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients. 2018;11:51. doi:10.3390/nu11010051
106. Liu R, Hong J, Xu X, et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat Med. 2017;23:859–868. doi:10.1038/nm.4358
107. Virtue AT, McCright SJ, Wright JM, et al. The gut microbiota regulates white adipose tissue inflammation and obesity via a family of microRNAs. Sci Transl Med. 2019;11:eaav1892. doi:10.1126/scitranslmed.aav1892
108. Ridlon JM, Kang D-J, Hylemon PB. Bile salt biotransformations by human intestinal bacteria. J Lipid Res. 2006;47:241–259. doi:10.1194/jlr.R500013-JLR200
109. Trauner M, Claudel T, Fickert P, Moustafa T, Wagner M. Bile acids as regulators of hepatic lipid and glucose metabolism. Dig Dis. 2010;28:220–224. doi:10.1159/000282091
110. Rohm TV, Meier DT, Olefsky JM, Donath MY. Inflammation in obesity, diabetes, and related disorders. Immunity. 2022;55:31–55. doi:10.1016/j.immuni.2021.12.013
111. Saad MJA, Santos A, Prada PO. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology. 2016;31:283–293. doi:10.1152/physiol.00041.2015
112. Torres-Fuentes C, Schellekens H, Dinan TG, Cryan JF. The microbiota-gut-brain axis in obesity. Lancet Gastroenterol Hepatol. 2017;2:747–756. doi:10.1016/S2468-1253(17)30147-4
113. Perry RJ, Samuel VT, Petersen KF, Shulman GI. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature. 2014;510:84–91. doi:10.1038/nature13478
114. Gasmi A, Noor S, Menzel A, Doşa A, Pivina L, Bjørklund G. Obesity and Insulin Resistance: associations with Chronic Inflammation, Genetic and Epigenetic Factors. Curr Med Chem. 2021;28:800–826. doi:10.2174/0929867327666200824112056
115. Kawai T, Autieri MV, Scalia R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol. 2021;320:C375–91. doi:10.1152/ajpcell.00379.2020
116. Boulangé CL, Neves AL, Chilloux J, Nicholson JK, Dumas M-E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016;8:42. doi:10.1186/s13073-016-0303-2
117. Arkan MC, Hevener AL, Greten FR, et al. IKK-beta links inflammation to obesity-induced insulin resistance. Nat Med. 2005;11:191–198. doi:10.1038/nm1185
118. Lafferty RA, Flatt PR, Irwin N. Emerging therapeutic potential for peptide YY for obesity-diabetes. Peptides. 2018;100:269–274. doi:10.1016/j.peptides.2017.11.005
119. Khandekar N, Berning BA, Sainsbury A, Lin S. The role of pancreatic polypeptide in the regulation of energy homeostasis. Molecular and Cellular Endocrinology. 2015;418:33–41. doi:10.1016/j.mce.2015.06.028
120. Salehi M, Purnell JQ. The Role of Glucagon-Like Peptide-1 in Energy Homeostasis. Metab Syndr Relat Disord. 2019;17:183–191. doi:10.1089/met.2018.0088
121. Cheng Z, Zhang L, Yang L, Chu H. The critical role of gut microbiota in obesity. Front Endocrinol. 2022;13:1025706. doi:10.3389/fendo.2022.1025706
122. Federico A, Dallio M, Tolone S, et al. Gastrointestinal Hormones, Intestinal Microbiota and Metabolic Homeostasis in Obese Patients: effect of Bariatric Surgery. Vivo. 2016;30:321–330.
123. Schéle E, Grahnemo L, Anesten F, Hallén A, Bäckhed F, Jansson J-O. The gut microbiota reduces leptin sensitivity and the expression of the obesity-suppressing neuropeptides proglucagon (Gcg) and brain-derived neurotrophic factor (Bdnf) in the central nervous system. Endocrinology. 2013;154:3643–3651. doi:10.1210/en.2012-2151
124. Chimerel C, Emery E, Summers DK, Keyser U, Gribble FM, Reimann F. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep. 2014;9:1202–1208. doi:10.1016/j.celrep.2014.10.032
125. Tolhurst G, Heffron H, Lam YS, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes. 2012;61:364–371. doi:10.2337/db11-1019
126. Larraufie P, Martin-Gallausiaux C, Lapaque N, et al. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci Rep. 2018;8:74. doi:10.1038/s41598-017-18259-0
127. Wong DH-J, Beiko RG. Transfer of energy pathway genes in microbial enhanced biological phosphorus removal communities. BMC Genomics. 2015;16:526. doi:10.1186/s12864-015-1752-5
128. Silberbauer CJ, Surina-Baumgartner DM, Arnold M, Langhans W. Prandial lactate infusion inhibits spontaneous feeding in rats. Am J Physiol Regul Integr Comp Physiol. 2000;278:R646–653. doi:10.1152/ajpregu.2000.278.3.R646
129. Nesci A, Carnuccio C, Ruggieri V, et al. Gut Microbiota and Cardiovascular Disease: evidence on the Metabolic and Inflammatory Background of a Complex Relationship. Int J Mol Sci. 2023;24:9087. doi:10.3390/ijms24109087
130. Li J, Zhao F, Wang Y, et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome. 2017;5:14. doi:10.1186/s40168-016-0222-x
131. Sun S, Lulla A, Sioda M, et al. Gut Microbiota Composition and Blood Pressure. Hypertension. 2019;73:998–1006. doi:10.1161/HYPERTENSIONAHA.118.12109
132. Liu J, An N, Ma C, et al. Correlation analysis of intestinal flora with hypertension. Exp Ther Med. 2018;16:2325–2330. doi:10.3892/etm.2018.6500
133. Yan Q, Gu Y, Li X, et al. Alterations of the Gut Microbiome in Hypertension. Front Cell Infect Microbiol. 2017;7:381. doi:10.3389/fcimb.2017.00381
134. Kim S, Goel R, Kumar A, et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin Sci. 2018;132:701–718. doi:10.1042/CS20180087
135. Dinakis E, Nakai M, Gill P, et al. Association Between the Gut Microbiome and Their Metabolites With Human Blood Pressure Variability. Hypertension. 2022;79:1690–1701. doi:10.1161/HYPERTENSIONAHA.122.19350
136. Wan C, Jin G, Zhu M, Hua J, He Y. Analysis of Gut Microbiota in Patients with Coronary Artery Disease and Hypertension. Evid Based Complement Alternat Med. 2021;2021:7195082. doi:10.1155/2021/7195082
137. Allayee H, Hazen SL. Contribution of Gut Bacteria to Lipid Levels: another Metabolic Role for Microbes? Circ Res. 2015;117:750–754. doi:10.1161/CIRCRESAHA.115.307409
138. Rangraze IR, El-Tanani M, Arman Rabbani S, Babiker R, Matalka I, Rizzo M. Diabetes and its Silent Partner: a Critical Review of Hyperinsulinemia and its Complications. Curr Diabetes Rev. 2024. doi:10.2174/0115733998311738240813110032
139. Lei L, Zhao N, Zhang L, Chen J, Liu X, Piao S. Gut microbiota is a potential goalkeeper of dyslipidemia. Front Endocrinol. 2022;13:950826. doi:10.3389/fendo.2022.950826
140. Qi J, You T, Li J, et al. Circulating trimethylamine N-oxide and the risk of cardiovascular diseases: a systematic review and meta-analysis of 11 prospective cohort studies. J Cell Mol Med. 2018;22:185–194. doi:10.1111/jcmm.13307
141. Chaplin A, Parra P, Serra F, Palou A. Conjugated Linoleic Acid Supplementation under a High-Fat Diet Modulates Stomach Protein Expression and Intestinal Microbiota in Adult Mice. PLoS One. 2015;10:e0125091. doi:10.1371/journal.pone.0125091
142. Ryan KK, Tremaroli V, Clemmensen C, et al. FXR is a molecular target for the effects of vertical sleeve gastrectomy. Nature. 2014;509:183–188. doi:10.1038/nature13135
143. Gargari G, Deon V, Taverniti V, et al. Evidence of dysbiosis in the intestinal microbial ecosystem of children and adolescents with primary hyperlipidemia and the potential role of regular hazelnut intake. FEMS Microbiol Ecol. 2018;94. doi:10.1093/femsec/fiy045
144. Moreno-Indias I, Sánchez-Alcoholado L, Pérez-Martínez P, et al. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016;7:1775–1787. doi:10.1039/c5fo00886g
145. Koren O, Spor A, Felin J, et al. Human oral, gut, and plaque microbiota in patients with atherosclerosis. Proc Natl Acad Sci U S A. 2011;108 Suppl 1:4592–4598. doi:10.1073/pnas.1011383107
146. Karlsson FH, Fåk F, Nookaew I, et al. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nat Commun. 2012;3:1245. doi:10.1038/ncomms2266
147. Martins D, Silva C, Ferreira AC, et al. Unravelling the Gut Microbiome Role in Cardiovascular Disease: a Systematic Review and a Meta-Analysis. Biomolecules. 2024;14:731. doi:10.3390/biom14060731
148. Zhu Y, Li Q, Jiang H. Gut microbiota in atherosclerosis: focus on trimethylamine N‐oxide. APMIS. 2020;128:353–366. doi:10.1111/apm.13038
149. Tang WHW, Wang Z, Levison BS, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368:1575–1584. doi:10.1056/NEJMoa1109400
150. Tang WHW, Wang Z, Kennedy DJ, et al. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ Res. 2015;116:448–455. doi:10.1161/CIRCRESAHA.116.305360
151. Tang WHW, Wang Z, Fan Y, et al. Prognostic value of elevated levels of intestinal microbe-generated metabolite trimethylamine-N-oxide in patients with heart failure: refining the gut hypothesis. J Am Coll Cardiol. 2014;64:1908–1914. doi:10.1016/j.jacc.2014.02.617
152. Senthong V, Wang Z, Fan Y, Hazen SL, Tang WHW. Trimethylamine N-Oxide and Mortality Risk in Patients With Peripheral Artery Disease. J Am Heart Assoc. 2016;5:e004237. doi:10.1161/JAHA.116.004237
153. Fu Q, Zhao M, Wang D, et al. Coronary Plaque Characterization Assessed by Optical Coherence Tomography and Plasma Trimethylamine-N-oxide Levels in Patients With Coronary Artery Disease. Am J Cardiol. 2016;118:1311–1315. doi:10.1016/j.amjcard.2016.07.071
154. Baker RG, Hayden MS, Ghosh S. NF-κB, inflammation and metabolic disease. Cell Metab. 2011;13:11–22. doi:10.1016/j.cmet.2010.12.008
155. Gorabi AM, Kiaie N, Khosrojerdi A, et al. Implications for the role of lipopolysaccharide in the development of atherosclerosis. Trends Cardiovasc Med. 2022;32:525–533. doi:10.1016/j.tcm.2021.08.015
156. Yoshida N, Yamashita T, Kishino S, et al. A possible beneficial effect of Bacteroides on faecal lipopolysaccharide activity and cardiovascular diseases. Sci Rep. 2020;10:13009. doi:10.1038/s41598-020-69983-z
157. Kasubuchi M, Hasegawa S, Hiramatsu T, Ichimura A, Kimura I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients. 2015;7:2839–2849. doi:10.3390/nu7042839
158. Fu J, Bonder MJ, Cenit MC, et al. The Gut Microbiome Contributes to a Substantial Proportion of the Variation in Blood Lipids. Circ Res. 2015;117:817–824. doi:10.1161/CIRCRESAHA.115.306807
159. Jian C, Silvestre MP, Middleton D, et al. Gut microbiota predicts body fat change following a low-energy diet: a PREVIEW intervention study. Genome Med. 2022;14:54. doi:10.1186/s13073-022-01053-7
160. Zeevi D, Korem T, Zmora N, et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell. 2015;163:1079–1094. doi:10.1016/j.cell.2015.11.001
161. Berry SE, Valdes AM, Drew DA, et al. Human postprandial responses to food and potential for precision nutrition. Nat Med. 2020;26:964–973. doi:10.1038/s41591-020-0934-0
162. Asnicar F, Berry SE, Valdes AM, et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat Med. 2021;27:321–332. doi:10.1038/s41591-020-01183-8
163. Dao MC, Everard A, Aron-Wisnewsky J, et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut. 2016;65:426–436. doi:10.1136/gutjnl-2014-308778
164. Haro C, Montes-Borrego M, Rangel-Zúñiga OA, et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J Clin Endocrinol Metab. 2016;101:233–242. doi:10.1210/jc.2015-3351
165. Gradisteanu Pircalabioru G, Rizzo M, Pantea Stoian A, Chifiriuc MC. Editorial: diet-microbe-host interactions in metabolic syndrome. Front Nutr. 2023;10. doi:10.3389/fnut.2023.1129200
166. Katsiki N, Stoian AP, Rizzo M. Dietary patterns in non-alcoholic fatty liver disease (NAFLD): stay on the straight and narrow path! Clin Investig Arterioscler. 2022;34(Suppl 1):S24–31. doi:10.1016/j.arteri.2021.10.001
167. Corina A, Abrudan MB, Nikolic D, et al. Effects of Aging and Diet on Cardioprotection and Cardiometabolic Risk Markers. Curr Pharm Des. 2019;25:3704–3714. doi:10.2174/1381612825666191105111232
168. Steckhan N, Hohmann C-D, Kessler C, Dobos G, Michalsen A, Cramer H. Effects of different dietary approaches on inflammatory markers in patients with metabolic syndrome: a systematic review and meta-analysis. Nutrition. 2016;32:338–348. doi:10.1016/j.nut.2015.09.010
169. Godos J, Zappalà G, Bernardini S, Giambini I, Bes-Rastrollo M. Adherence to the Mediterranean diet is inversely associated with metabolic syndrome occurrence: a meta-analysis of observational studies. Int J Food Sci Nutr. 2017;68:138–148. doi:10.1080/09637486.2016.1221900
170. Lăcătușu C-M, Grigorescu E-D, Floria M, Onofriescu A, Mihai B-M. The Mediterranean Diet: from an Environment-Driven Food Culture to an Emerging Medical Prescription. Int J Environ Res Public Health. 2019;16:942. doi:10.3390/ijerph16060942
171. Di Daniele N, Noce A, Vidiri MF, et al. Impact of Mediterranean diet on metabolic syndrome, cancer and longevity. Oncotarget. 2017;8:8947–8979. doi:10.18632/oncotarget.13553
172. Franquesa M, Pujol-Busquets G, García-Fernández E, et al. Mediterranean Diet and Cardiodiabesity: a Systematic Review through Evidence-Based Answers to Key Clinical Questions. Nutrients. 2019;11:655. doi:10.3390/nu11030655
173. Dernini S, Berry EM, Serra-Majem L, et al. Med Diet 4.0: the Mediterranean diet with four sustainable benefits. Public Health Nutr. 2017;20:1322–1330. doi:10.1017/S1368980016003177
174. Xavier Medina F. Mediterranean diet, culture and heritage: challenges for a new conception. Public Health Nutr. 2009;12:1618–1620. doi:10.1017/S1368980009990450
175. McGuire S, Scientific Report of the 2015 Dietary Guidelines Advisory Committee. Washington, DC: US Departments of Agriculture and Health and Human Services, 2015. Adv Nutr. 2016;7:202–204. doi:10.3945/an.115.011684
176. Storniolo CE, Sacanella I, Mitjavila MT, Lamuela-Raventos RM, Moreno JJ. Bioactive Compounds of Cooked Tomato Sauce Modulate Oxidative Stress and Arachidonic Acid Cascade Induced by Oxidized LDL in Macrophage Cultures. Nutrients. 2019;11:1880. doi:10.3390/nu11081880
177. Davis C, Bryan J, Hodgson J, Murphy K. Definition of the Mediterranean Diet: a Literature Review. Nutrients. 2015;7:9139–9153. doi:10.3390/nu7115459
178. Finicelli M, Squillaro T, Di Cristo F, et al. Metabolic syndrome, Mediterranean diet, and polyphenols: evidence and perspectives. J Cell Physiol. 2019;234:5807–5826. doi:10.1002/jcp.27506
179. Widmer RJ, Flammer AJ, Lerman LO, Lerman A. The Mediterranean diet, its components, and cardiovascular disease. Am J Med. 2015;128:229–238. doi:10.1016/j.amjmed.2014.10.014
180. Chiva-Blanch G, Badimon L. Effects of Polyphenol Intake on Metabolic Syndrome: current Evidences from Human Trials. Oxid Med Cell Longev. 2017;2017:5812401. doi:10.1155/2017/5812401
181. Steffen LM, Van Horn L, Daviglus ML, et al. A modified Mediterranean diet score is associated with a lower risk of incident metabolic syndrome over 25 years among young adults: the CARDIA (Coronary Artery Risk Development in Young Adults) study. Br J Nutr. 2014;112:1654–1661. doi:10.1017/S0007114514002633
182. Kesse-Guyot E, Ahluwalia N, Lassale C, Hercberg S, Fezeu L, Lairon D. Adherence to Mediterranean diet reduces the risk of metabolic syndrome: a 6-year prospective study. Nutr, Metab Cardiovasc Dis. 2013;23:677–683. doi:10.1016/j.numecd.2012.02.005
183. Kim Y, Je Y. A modified Mediterranean diet score is inversely associated with metabolic syndrome in Korean adults. Eur J Clin Nutr. 2018;72:1682–1689. doi:10.1038/s41430-018-0156-4
184. Yu J, Wu Y, Zhu Z, et al. The impact of dietary patterns on gut microbiota for the primary and secondary prevention of cardiovascular disease: a systematic review. Nutr J. 2025;24:17. doi:10.1186/s12937-024-01060-x
185. Pilla SJ, Yeh H-C, Mitchell CM, et al. Dietary Patterns, Sodium Reduction, and Blood Pressure in Type 2 Diabetes: the DASH4D Randomized Clinical Trial. JAMA Intern Med. 2025;185(8):937–946. doi:10.1001/jamainternmed.2025.1580
186. Filippou CD, Tsioufis CP, Thomopoulos CG, et al. Dietary Approaches to Stop Hypertension (DASH) Diet and Blood Pressure Reduction in Adults with and without Hypertension: a Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv Nutr. 2020;11:1150–1160. doi:10.1093/advances/nmaa041
187. Said MS, El Sayed IT, Ibrahim EE, Khafagy GM. Effect of DASH Diet Versus Healthy Dietary Advice on the Estimated Atherosclerotic Cardiovascular Disease Risk. J Prim Care Community Health. 2021;12:2150132720980952. doi:10.1177/2150132720980952
188. Ge L, Sadeghirad B, Ball GDC, et al. Comparison of dietary macronutrient patterns of 14 popular named dietary programmes for weight and cardiovascular risk factor reduction in adults: systematic review and network meta-analysis of randomised trials. BMJ. 2020;369:m696. doi:10.1136/bmj.m696
189. Siervo M, Lara J, Chowdhury S, Ashor A, Oggioni C, Mathers JC. Effects of the Dietary Approach to Stop Hypertension (DASH) diet on cardiovascular risk factors: a systematic review and meta-analysis. Br J Nutr. 2015;113:1–15. doi:10.1017/S0007114514003341
190. Schwingshackl L, Chaimani A, Schwedhelm C, et al. Comparative effects of different dietary approaches on blood pressure in hypertensive and pre-hypertensive patients: a systematic review and network meta-analysis. Crit Rev Food Sci Nutr. 2019;59:2674–2687. doi:10.1080/10408398.2018.1463967
191. Phillips CM, Harrington JM, Perry IJ. Relationship between dietary quality, determined by DASH score, and cardiometabolic health biomarkers: a cross-sectional analysis in adults. Clin Nutr. 2019;38:1620–1628. doi:10.1016/j.clnu.2018.08.028
192. Drehmer M, Odegaard AO, Schmidt MI, et al. Brazilian dietary patterns and the dietary approaches to stop hypertension (DASH) diet-relationship with metabolic syndrome and newly diagnosed diabetes in the ELSA-Brasil study. Diabetol Metab Syndr. 2017;9:13. doi:10.1186/s13098-017-0211-7
193. Gibson R, Singh D, Vergnaud A-C, et al. A cross-sectional investigation into the occupational and socio-demographic characteristics of British police force employees reporting a dietary pattern associated with cardiometabolic risk: findings from the Airwave Health Monitoring Study. Eur J Nutr. 2018;57:2913–2926. doi:10.1007/s00394-017-1562-4
194. Jones NRV, Forouhi NG, Khaw K-T, Wareham NJ, Monsivais P. Accordance to the Dietary Approaches to Stop Hypertension diet pattern and cardiovascular disease in a British, population-based cohort. Eur J Epidemiol. 2018;33:235–244. doi:10.1007/s10654-017-0354-8
195. Djoussé L, Ho Y-L, Nguyen X-MT, et al. DASH Score and Subsequent Risk of Coronary Artery Disease: the Findings From Million Veteran Program. J Am Heart Assoc. 2018;7:e008089. doi:10.1161/JAHA.117.008089
196. Schwingshackl L, Bogensberger B, Hoffmann G. Diet Quality as Assessed by the Healthy Eating Index, Alternate Healthy Eating Index, Dietary Approaches to Stop Hypertension Score, and Health Outcomes: an Updated Systematic Review and Meta-Analysis of Cohort Studies. J Acad Nutr Diet. 2018;118:74–100.e11. doi:10.1016/j.jand.2017.08.024
197. Soltani S, Shirani F, Chitsazi MJ, Salehi-Abargouei A. The effect of dietary approaches to stop hypertension (DASH) diet on weight and body composition in adults: a systematic review and meta-analysis of randomized controlled clinical trials. Obes Rev. 2016;17:442–454. doi:10.1111/obr.12391
198. Hashemi R, Mehdizadeh Khalifani A, Rahimlou M. Comparison of the effect of Dietary Approaches to Stop Hypertension diet and American Diabetes Association nutrition guidelines on lipid profiles in patients with type 2 diabetes: a comparative clinical trial. Nutr Diet. 2020;77:204–211. doi:10.1111/1747-0080.12543
199. Mertens E, Markey O, Geleijnse JM, Lovegrove JA, Givens DI. Adherence to a healthy diet in relation to cardiovascular incidence and risk markers: evidence from the Caerphilly Prospective Study. Eur J Nutr. 2018;57:1245–1258. doi:10.1007/s00394-017-1408-0
200. Asghari G, Yuzbashian E, Mirmiran P, Hooshmand F, Najafi R, Azizi F. Dietary Approaches to Stop Hypertension (DASH) Dietary Pattern Is Associated with Reduced Incidence of Metabolic Syndrome in Children and Adolescents. J Pediatr. 2016;174:178–184.e1. doi:10.1016/j.jpeds.2016.03.077
201. Peairs AD, Shah AS, Hess M, Couch SC. Effects of the dietary approaches to stop hypertension (DASH) diet on glucose variability in youth with Type 1 diabetes. Diabetes Manag (Lond). 2017;7:383–391.
202. Satija A, Hu FB. Plant-based diets and cardiovascular health. Trends Cardiovasc Med. 2018;28:437–441. doi:10.1016/j.tcm.2018.02.004
203. Kahleova H, Levin S, Barnard N. Cardio-Metabolic Benefits of Plant-Based Diets. Nutrients. 2017;9:848. doi:10.3390/nu9080848
204. Kim H, Caulfield LE, Rebholz CM. Healthy Plant-Based Diets Are Associated with Lower Risk of All-Cause Mortality in US Adults. J Nutr. 2018;148:624–631. doi:10.1093/jn/nxy019
205. Enayati A, Butler AE, Rizzo M, Sahebkar A. The Effect of Curcumin on the Gut-Brain Axis: therapeutic Implications. J Neurogastroenterol Motil. 2023;29:409–418. doi:10.5056/jnm23065
206. Crowe FL, Appleby PN, Travis RC, Key TJ. Risk of hospitalization or death from ischemic heart disease among British vegetarians and nonvegetarians: results from the EPIC-Oxford cohort study123. The American Journal of Clinical Nutrition. 2013;97:597–603. doi:10.3945/ajcn.112.044073
207. Yokoyama Y, Nishimura K, Barnard ND, et al. Vegetarian diets and blood pressure: a meta-analysis. JAMA Intern Med. 2014;174:577–587. doi:10.1001/jamainternmed.2013.14547
208. Hemler EC, Hu FB. Plant-Based Diets for Cardiovascular Disease Prevention: all Plant Foods Are Not Created Equal. Curr Atheroscler Rep. 2019;21:18. doi:10.1007/s11883-019-0779-5
209. Wang F, Zheng J, Yang B, Jiang J, Fu Y, Li D. Effects of Vegetarian Diets on Blood Lipids: a Systematic Review and Meta-Analysis of Randomized Controlled Trials. J Am Heart Assoc. 2015;4:e002408. doi:10.1161/JAHA.115.002408
210. Huang R-Y, Huang -C-C, Hu FB, Chavarro JE. Vegetarian Diets and Weight Reduction: a Meta-Analysis of Randomized Controlled Trials. J Gen Intern Med. 2016;31:109–116. doi:10.1007/s11606-015-3390-7
211. Mozaffarian D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity: a Comprehensive Review. Circulation. 2016;133:187–225. doi:10.1161/CIRCULATIONAHA.115.018585
212. Liu Y-S, Wu Q-J, Xia Y, et al. Carbohydrate intake and risk of metabolic syndrome: a dose-response meta-analysis of observational studies. Nutr, Metab Cardiovasc Dis. 2019;29:1288–1298. doi:10.1016/j.numecd.2019.09.003
213. Dehghan M, Mente A, Zhang X, et al. Associations of fats and carbohydrate intake with cardiovascular disease and mortality in 18 countries from five continents (PURE): a prospective cohort study. Lancet. 2017;390:2050–2062. doi:10.1016/S0140-6736(17)32252-3
214. Evert AB, Dennison M, Gardner CD, et al. Nutrition Therapy for Adults With Diabetes or Prediabetes: a Consensus Report. Diabetes Care. 2019;42:731–754. doi:10.2337/dci19-0014
215. Bazzano LA, Hu T, Reynolds K, et al. Effects of low-carbohydrate and low-fat diets: a randomized trial. Ann Intern Med. 2014;161:309–318. doi:10.7326/M14-0180
216. van Zuuren EJ, Fedorowicz Z, Kuijpers T, Pijl H. Effects of low-carbohydrate- compared with low-fat-diet interventions on metabolic control in people with type 2 diabetes: a systematic review including GRADE assessments. Am J Clin Nutr. 2018;108:300–331. doi:10.1093/ajcn/nqy096
217. Puska P. Fat and heart disease: yes we can make a change--the case of North Karelia (Finland). Ann Nutr Metab. 2009;54(1):33–38. doi:10.1159/000220825
218. Yu E, Rimm E, Qi L, et al. Diet, Lifestyle, Biomarkers, Genetic Factors, and Risk of Cardiovascular Disease in the Nurses. Health Studies Am J Public Health. 2016;106:1616–1623. doi:10.2105/AJPH.2016.303316
219. Paoli A, Rubini A, Volek JS, Grimaldi KA. Beyond weight loss: a review of the therapeutic uses of very-low-carbohydrate (ketogenic) diets. Eur J Clin Nutr. 2013;67:789–796. doi:10.1038/ejcn.2013.116
220. Stoica RA, Diaconu CC, Rizzo M, et al. Weight loss programmes using low carbohydrate diets to control the cardiovascular risk in adolescents (Review). Exp Ther Med. 2021;21:90. doi:10.3892/etm.2020.9522
221. Jensen MD, Ryan DH, Apovian CM, et al. 2013 AHA/ACC/TOS guideline for the management of overweight and obesity in adults: a report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines and The Obesity Society. Circulation. 2014;129:S102–138. doi:10.1161/01.cir.0000437739.71477.ee
222. Moreno B, Bellido D, Sajoux I, et al. Comparison of a very low-calorie-ketogenic diet with a standard low-calorie diet in the treatment of obesity. Endocrine. 2014;47:793–805. doi:10.1007/s12020-014-0192-3
223. Choi H-R, Kim J, Lim H, Park YK. Two-Week Exclusive Supplementation of Modified Ketogenic Nutrition Drink Reserves Lean Body Mass and Improves Blood Lipid Profile in Obese Adults: a Randomized Clinical Trial. Nutrients. 2018;10:1895. doi:10.3390/nu10121895
224. Bueno NB, de Melo ISV, de Oliveira SL, da Rocha Ataide T. Very-low-carbohydrate ketogenic diet v. low-fat diet for long-term weight loss: a meta-analysis of randomised controlled trials. Br J Nutr. 2013;110:1178–1187. doi:10.1017/S0007114513000548
225. Corpeleijn E, Saris WHM, Blaak EE. Metabolic flexibility in the development of insulin resistance and type 2 diabetes: effects of lifestyle. Obes Rev. 2009;10:178–193. doi:10.1111/j.1467-789X.2008.00544.x
226. Westerterp-Plantenga MS, Nieuwenhuizen A, Tomé D, Soenen S, Westerterp KR. Dietary protein, weight loss, and weight maintenance. Annu Rev Nutr. 2009;29:21–41. doi:10.1146/annurev-nutr-080508-141056
227. Ajala O, English P, Pinkney J. Systematic review and meta-analysis of different dietary approaches to the management of type 2 diabetes. Am J Clin Nutr. 2013;97:505–516. doi:10.3945/ajcn.112.042457
228. Ma C, Avenell A, Bolland M, et al. Effects of weight loss interventions for adults who are obese on mortality, cardiovascular disease, and cancer: systematic review and meta-analysis. BMJ. 2017;359:j4849. doi:10.1136/bmj.j4849
229. Prentice RL, Aragaki AK, Van Horn L, et al. Low-fat dietary pattern and cardiovascular disease: results from the Women’s Health Initiative randomized controlled trial. Am J Clin Nutr. 2017;106:35–43. doi:10.3945/ajcn.117.153270
230. Neuhouser ML, Howard B, Lu J, et al. A low-fat dietary pattern and risk of metabolic syndrome in postmenopausal women: the Women’s Health Initiative. Metabolism. 2012;61:1572–1581. doi:10.1016/j.metabol.2012.04.007
231. Paniagua JA, Pérez-Martinez P, Gjelstad IMF, et al. A low-fat high-carbohydrate diet supplemented with long-chain n-3 PUFA reduces the risk of the metabolic syndrome. Atherosclerosis. 2011;218:443–450. doi:10.1016/j.atherosclerosis.2011.07.003
232. Babio N, Toledo E, Estruch R, et al. Mediterranean diets and metabolic syndrome status in the PREDIMED randomized trial. CMAJ. 2014;186:E649–657. doi:10.1503/cmaj.140764
233. Veum VL, Laupsa-Borge J, Eng Ø, et al. Visceral adiposity and metabolic syndrome after very high-fat and low-fat isocaloric diets: a randomized controlled trial. Am J Clin Nutr. 2017;105:85–99. doi:10.3945/ajcn.115.123463
234. Gardner CD, Trepanowski JF, Del Gobbo LC, et al. Effect of Low-Fat vs Low-Carbohydrate Diet on 12-Month Weight Loss in Overweight Adults and the Association With Genotype Pattern or Insulin Secretion: the DIETFITS Randomized Clinical Trial. JAMA. 2018;319:667–679. doi:10.1001/jama.2018.0245
235. Basterra-Gortari FJ, Ruiz-Canela M, Martínez-González MA, et al. Effects of a Mediterranean Eating Plan on the Need for Glucose-Lowering Medications in Participants With Type 2 Diabetes: a Subgroup Analysis of the PREDIMED Trial. Diabetes Care. 2019;42:1390–1397. doi:10.2337/dc18-2475
236. Santos-Marcos JA, Perez-Jimenez F, Camargo A. The role of diet and intestinal microbiota in the development of metabolic syndrome. J Nutr Biochem. 2019;70:1–27. doi:10.1016/j.jnutbio.2019.03.017
237. Campos-Nonato I, Hernandez L, Barquera S. Effect of a High-Protein Diet versus Standard-Protein Diet on Weight Loss and Biomarkers of Metabolic Syndrome: a Randomized Clinical Trial. Obes Facts. 2017;10:238–251. doi:10.1159/000471485
238. Rock CL, Flatt SW, Pakiz B, et al. Weight loss, glycemic control, and cardiovascular disease risk factors in response to differential diet composition in a weight loss program in type 2 diabetes: a randomized controlled trial. Diabetes Care. 2014;37:1573–1580. doi:10.2337/dc13-2900
239. Guasch-Ferré M, Satija A, Blondin SA, et al. Meta-Analysis of Randomized Controlled Trials of Red Meat Consumption in Comparison With Various Comparison Diets on Cardiovascular Risk Factors. Circulation. 2019;139:1828–1845. doi:10.1161/CIRCULATIONAHA.118.035225
240. Gadgil MD, Appel LJ, Yeung E, Anderson CAM, Sacks FM, Miller ER. The effects of carbohydrate, unsaturated fat, and protein intake on measures of insulin sensitivity: results from the OmniHeart trial. Diabetes Care. 2013;36:1132–1137. doi:10.2337/dc12-0869
241. Astrup A. The satiating power of protein--a key to obesity prevention? Am J Clin Nutr. 2005;82:1–2. doi:10.1093/ajcn.82.1.1
242. Wycherley TP, Moran LJ, Clifton PM, Noakes M, Brinkworth GD. Effects of energy-restricted high-protein, low-fat compared with standard-protein, low-fat diets: a meta-analysis of randomized controlled trials. Am J Clin Nutr. 2012;96:1281–1298. doi:10.3945/ajcn.112.044321
243. Pan A, Sun Q, Bernstein AM, Manson JE, Willett WC, Hu FB. Changes in red meat consumption and subsequent risk of type 2 diabetes mellitus: three cohorts of US men and women. JAMA Intern Med. 2013;173:1328–1335. doi:10.1001/jamainternmed.2013.6633
244. Castro-Barquero S, Ruiz-León AM, Sierra-Pérez M, Estruch R, Casas R. Dietary Strategies for Metabolic Syndrome: a Comprehensive Review. Nutrients. 2020;12:2983. doi:10.3390/nu12102983
245. Al-Dughaishi T, Nikolic D, Zadjali F, et al. Nutraceuticals as Lipid-Lowering Treatment in Pregnancy and Their Effects on the Metabolic Syndrome. Curr Pharm Biotechnol. 2016;17:614–623. doi:10.2174/1389201017666160401145817
246. Sanders ME. Probiotics: definition, sources, selection, and uses. Clin Infect Dis. 2008;46(Suppl 2):S58–61. doi:10.1086/523341
247. Chilton SN, Burton JP, Reid G. Inclusion of fermented foods in food guides around the world. Nutrients. 2015;7:390–404. doi:10.3390/nu7010390
248. Akbari V, Hendijani F. Effects of probiotic supplementation in patients with type 2 diabetes: systematic review and meta-analysis. Nutr Rev. 2016;74:774–784. doi:10.1093/nutrit/nuw039
249. Samah S, Ramasamy K, Lim SM, Neoh CF. Probiotics for the management of type 2 diabetes mellitus: a systematic review and meta-analysis. Diabet Res Clin Pract. 2016;118:172–182. doi:10.1016/j.diabres.2016.06.014
250. Li C, Li X, Han H, et al. Effect of probiotics on metabolic profiles in type 2 diabetes mellitus: a meta-analysis of randomized, controlled trials. Medicine. 2016;95:e4088. doi:10.1097/MD.0000000000004088
251. Borgeraas H, Johnson LK, Skattebu J, Hertel JK, Hjelmesaeth J. Effects of probiotics on body weight, body mass index, fat mass and fat percentage in subjects with overweight or obesity: a systematic review and meta-analysis of randomized controlled trials. Obes Rev. 2018;19:219–232. doi:10.1111/obr.12626
252. Cicero AFG, Colletti A, Bajraktari G, et al. Lipid lowering nutraceuticals in clinical practice: position paper from an International Lipid Expert Panel. Arch Med Sci. 2017;13:965–1005. doi:10.5114/aoms.2017.69326
253. Tenorio-Jiménez C, Martínez-Ramírez MJ, Del Castillo-Codes I, et al. Lactobacillus reuteri V3401 Reduces Inflammatory Biomarkers and Modifies the Gastrointestinal Microbiome in Adults with Metabolic Syndrome: the PROSIR Study. Nutrients. 2019;11:1761. doi:10.3390/nu11081761
254. Hsieh M-C, Tsai W-H, Jheng Y-P, et al. The beneficial effects of Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2 diabetes mellitus: a randomized, double-blinded, placebo-controlled trial. Sci Rep. 2018;8:16791. doi:10.1038/s41598-018-35014-1
255. Hanchi H, Mottawea W, Sebei K, Hammami R. The Genus Enterococcus: between Probiotic Potential and Safety Concerns-An Update. Front Microbiol. 2018;9:1791. doi:10.3389/fmicb.2018.01791
256. Sohn M, Na GY, Chu J, Joung H, Kim B-K, Lim S. Efficacy and Safety of Lactobacillus plantarum K50 on Lipids in Koreans With Obesity: a Randomized, Double-Blind Controlled Clinical Trial. Front Endocrinol. 2021;12:790046. doi:10.3389/fendo.2021.790046
257. Pellegrini M, Ippolito M, Monge T, et al. Gut microbiota composition after diet and probiotics in overweight breast cancer survivors: a randomized open-label pilot intervention trial. Nutrition. 2020;74:110749. doi:10.1016/j.nut.2020.110749
258. Solito A, Bozzi Cionci N, Calgaro M, et al. Supplementation with Bifidobacterium breve BR03 and B632 strains improved insulin sensitivity in children and adolescents with obesity in a cross-over, randomized double-blind placebo-controlled trial. Clin Nutr. 2021;40:4585–4594. doi:10.1016/j.clnu.2021.06.002
259. Gibson GR, Hutkins R, Sanders ME, et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. 2017;14:491–502. doi:10.1038/nrgastro.2017.75
260. Rizzo M, Colletti A, Penson PE, et al. Nutraceutical approaches to non-alcoholic fatty liver disease (NAFLD): a position paper from the International Lipid Expert Panel (ILEP). Pharmacol Res. 2023;189:106679. doi:10.1016/j.phrs.2023.106679
261. Kellow NJ, Coughlan MT, Reid CM. Metabolic benefits of dietary prebiotics in human subjects: a systematic review of randomised controlled trials. Br J Nutr. 2014;111:1147–1161. doi:10.1017/S0007114513003607
262. O’Connor S, Chouinard-Castonguay S, Gagnon C, Rudkowska I. Prebiotics in the management of components of the metabolic syndrome. Maturitas. 2017;104:11–18. doi:10.1016/j.maturitas.2017.07.005
263. Vulevic J, Juric A, Tzortzis G, Gibson GR. A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J Nutr. 2013;143:324–331. doi:10.3945/jn.112.166132
264. Morel FB, Dai Q, Ni J, Thomas D, Parnet P, Fança-Berthon P. α-Galacto-oligosaccharides Dose-Dependently Reduce Appetite and Decrease Inflammation in Overweight Adults. J Nutr. 2015;145:2052–2059. doi:10.3945/jn.114.204909
265. Canfora EE, van der Beek CM, Hermes GDA, et al. Supplementation of Diet With Galacto-oligosaccharides Increases Bifidobacteria, but Not Insulin Sensitivity, in Obese Prediabetic Individuals. Gastroenterology. 2017;153:87–97.e3. doi:10.1053/j.gastro.2017.03.051
266. Dewulf EM, Cani PD, Claus SP, et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut. 2013;62:1112–1121. doi:10.1136/gutjnl-2012-303304
267. Tian R, Hong J, Zhao J, et al. Overall Structural Alteration of Gut Microbiota and Relationships with Risk Factors in Patients with Metabolic Syndrome Treated with Inulin Alone and with Other Agents: an Open-Label Pilot Study. Mediators Inflamm. 2022;2022:2078520. doi:10.1155/2022/2078520
268. Dhakal S, Dey M. Resistant starch type-4 intake alters circulating bile acids in human subjects. Front Nutr. 2022;9:930414. doi:10.3389/fnut.2022.930414
269. Upadhyaya B, McCormack L, Fardin-Kia AR, et al. Impact of dietary resistant starch type 4 on human gut microbiota and immunometabolic functions. Sci Rep. 2016;6:28797. doi:10.1038/srep28797
270. Xu D, Feng M, Chu Y, et al. The Prebiotic Effects of Oats on Blood Lipids, Gut Microbiota, and Short-Chain Fatty Acids in Mildly Hypercholesterolemic Subjects Compared With Rice: a Randomized, Controlled Trial. Front Immunol. 2021;12:787797. doi:10.3389/fimmu.2021.787797
271. Cortés-Martín A, Iglesias-Aguirre CE, Meoro A, Selma MV, Espín JC. Pharmacological Therapy Determines the Gut Microbiota Modulation by a Pomegranate Extract Nutraceutical in Metabolic Syndrome: a Randomized Clinical Trial. Mol Nutr Food Res. 2021;65:e2001048. doi:10.1002/mnfr.202001048
272. Kaul R, Paul P, Harfouche M, Ayyan M, Laws S, Chaari A. The effect of microbiome-modulating therapeutics on glucose homeostasis in metabolic syndrome: a systematic review, meta-analysis, and meta-regression of clinical trials. Diabetes Metab Syndr. 2024;18(8):103118. doi:10.1016/j.dsx.2024.103118 Epub 2024 Sep 12. PMID: 39298907.
273. Vrieze A, Van Nood E, Holleman F, et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology. 2012;143:913–916.e7. doi:10.1053/j.gastro.2012.06.031
274. DeFilipp Z, Bloom PP, Soto MT, et al. Drug-Resistant E. coli Bacteremia Transmitted by Fecal Microbiota Transplant. N Engl J Med. 2019;381:2043–2050. doi:10.1056/NEJMoa1910437
275. Rapoport EA, Baig M, Puli SR. Adverse events in fecal microbiota transplantation: a systematic review and meta-analysis. Ann Gastroenterol. 2022;35:150. doi:10.20524/aog.2022.0695
276. Leong KSW, Jayasinghe TN, Wilson BC, et al. Effects of Fecal Microbiome Transfer in Adolescents With Obesity: the Gut Bugs Randomized Controlled Trial. JAMA Network Open. 2020;3:e2030415. doi:10.1001/jamanetworkopen.2020.30415
277. Lin W, Pu L, Qian X, Pan J, Cheng R, Sun P. Exercise-Induced Modulation of Gut Microbiota in Individuals With Obesity and Type 2 Diabetes: a Systematic Review and Meta-Analysis. Front Microbiol. 2025;16:1671975.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Understanding the Role of Metabolic Syndrome as a Risk Factor for Hepatocellular Carcinoma
Chavez-Tapia NC, Murúa-Beltrán Gall S, Ordoñez-Vázquez AL, Nuño-Lambarri N, Vidal-Cevallos P, Uribe M
Journal of Hepatocellular Carcinoma 2022, 9:583-593
Published Date: 5 July 2022
Knowledge of Cardiovascular Disease Risk Factors and Its Primary Prevention Practices Among the Saudi Public – A Questionnaire-Based Cross-Sectional Study
Bashatah A, Syed W, Al-Rawi MBA
International Journal of General Medicine 2023, 16:4745-4756
Published Date: 20 October 2023
Evaluating Glycemic Control Efficacy and Safety of the Oral Small Molecule Glucagon-Like Peptide 1 Receptor Agonist Danuglipron in Type 2 Diabetes Patients: A Systemic Review and Meta-Analysis
Fatima H, Rangwala HS, Mustafa MS, Shafique MA, Abbas SR, Rizwan A, Fadlalla Ahmed TK, Arshad A
Diabetes, Metabolic Syndrome and Obesity 2023, 16:3567-3578
Published Date: 7 November 2023
Improvement in Cutaneous Conditions Can Benefit Some Health Conditions in the Elderly
Yang B, Man MQ
Clinical Interventions in Aging 2023, 18:2031-2040
Published Date: 1 December 2023
Design and Protocol of the Biobank for Metabolic Syndrome Consequences (BMSC): A Prospective Cohort Study in Northwest China
Yang X, Li Q, He Y, Yin G, Li M, Zhu W, Fan X, Gong Y, Wang Y, Qiang W, Wang Y, Willems van Dijk K, Rensen PCN, Guo H, Shi B, Wang Y
Clinical Epidemiology 2025, 17:823-835
Published Date: 14 October 2025