Back to Journals » Journal of Inflammation Research » Volume 18
Knockdown of Long Noncoding RNA IPCRL1 Mitigates Myocardial Ischemia/Reperfusion Injury via miR-185-3p/JIP3 Axis and JNK Pathway
Authors Chen J, Zhang Y, Zhang Z, Yu Z, Zhang H, Zhao Q ![]()
Received 16 December 2024
Accepted for publication 10 July 2025
Published 8 August 2025 Volume 2025:18 Pages 10695—10709
DOI https://doi.org/10.2147/JIR.S512561
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rongxue Wu
Jingyu Chen,1,2 Yi Zhang,1,2 Zixin Zhang,1,2 Ziwei Yu,1,2 Hao Zhang,3 Qifeng Zhao1,2
1Department of Cardiovascular and Thoracic Surgery, The second Affiliated Hospital and Yuying Children’s Hospital of Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China; 2Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China; 3Heart Center and Shanghai Institute of Pediatric Congenital Heart Disease, Shanghai Children’s Medical Center, National Children’s Medical Center, School of Medicine, Shanghai Jiao tong University, Shanghai, People’s Republic of China
Correspondence: Qifeng Zhao, Department of Cardiovascular and Thoracic Surgery, The Second Affiliated Hospital and Yuying Children’s Hospital of Wenzhou Medical University, Wenzhou, Zhejiang Province, People’s Republic of China, Email [email protected]
Purpose: Myocardial ischemia/reperfusion injury (MIRI) represents a significant culprit leading to adverse consequences after cardiac surgery. This study aims to clarify the function and related pathway of ischemic preconditioning related lncRNA-1 (IPCRL1) in MIRI.
Methods: MIRI model was established in C57BL/6J mice via the myocardial reperfusion method, and hypoxia/reoxygenation (H/R) model was constructed using HL-1 cell. IPCRL1, miR-185-3p, JIP3, TNF-α were determined using RT-qPCR. JIP3, c-Jun, JNK phosphorylation, B-cell lymphoma 2(BCL2), Bcl-2-associated X protein (BAX), cleaved caspase-3 levels were measured using Western blot. ELISA was used to measured cardiomyocyte injury markers and TNF-α concentrations. IHC and flow cytometry investigated the trends in apoptosis. The binding relationships between IPCRL1, miR-185-3p, JIP3 were verified by Dual-luciferase reporter assay.
Results: IPCRL1 knockdown reduced infarct size, inflammation, and apoptosis. Additionally, knockdown of IPCRL1 downregulates the expression of JIP3 via sponge miRNA-185-3p, thereby affecting the JNK pathway, meanwhile inhibition of miRNA-185-3p reversed above effects. Knocking down IPCRL1 can counteract cardiomyocyte apoptosis through miR-185-3p/JIP3 axis, offering protection against MIRI.
Conclusion: IPCRL1/miRNA-185-3p/JIP3 axis mediates MIRI through JNK pathway and IPCRL1 may hold promise as a new noteworthy target for MIRI.
Keywords: myocardial reperfusion injury, long non-coding RNA, miR-185-3p, JNK, apoptosis, inflammation
Introduction
Cardiopulmonary bypass (CPB) is a crucial adjunct in cardiac surgery for procedures involving the heart and major blood vessels.1 Nonetheless, reperfusion of the myocardium is widely acknowledged to exacerbate myocardial ischemia/reperfusion injury (MIRI) during CPB.1–3 Mitigating MIRI remains a focal point of clinical and foundational investigation in cardiovascular surgery. The mechanisms underlying MIRI are multifaceted and involve cardiomyocyte apoptosis, necrosis, inflammatory damage, and mitochondrial dysfunction in disease progression.4,5 Remote ischemic preconditioning (RIPC) is a strategy that utilizes inherent protective mechanisms within the human body to combat ischemia/reperfusion injury (IRI), and it could suppress mitochondrial permeability transition-mediated apoptosis and effectively alleviate MIRI.6 Our preliminary research has confirmed the beneficial effects of RIPC in reducing MIRI.7 However, the translational efficacy from animal experimentation to clinical application has not been successful, and the cardioprotective effects of RIPC have been a subject of controversy.8 In certain studies, RIPC has not significantly improved clinical outcomes in patients undergoing primary percutaneous coronary intervention (PPCI) for myocardial infarction.9,10 Therefore, to enhance the clinical applicability of RIPC in MIRI to its fullest extent, it is imperative to identify the molecular functions of key players underlying the cardioprotective effects provided.
A wealth of studies indicates that long noncoding RNAs (lncRNAs) play a crucial role in the occurrence and progression of MIRI, serving as competitive endogenous RNAs (ceRNAs) to regulate certain RNA transcripts, including microRNAs (miRNAs).11–14 lncRNAs also play significant roles in mediating inflammation and cardiomyocyte apoptosis during MIRI.12,15–17 For example, lncRNA KCNQ1OT1 promotes hydrogen peroxide–induced myocardial cell apoptosis and inflammation by binding to miR-130a-3p and upregulating ZNF791.16 According to Bao et al, lncRNA JPX targets SERCA2a to alleviate MIRI through its interaction with EZH2.17 Interestingly, our previous study using microarray analysis identified a RIPC-related LncRNA, designated as IPCRL1 (ENSMUST00000123752), a murine LncRNA also known as Alox12e-002, Alox12-ps1, and Alox12-ps2. This 793 bp LncRNA, located on the reverse strand of chromosome 11, was upregulated in myocardial tissue following MIRI in mice, suggesting that it may play a crucial role in RIPC-mediated protection against MIRI.7 However, its function has not been previously discovered or confirmed, and its specific role and mechanisms require further investigation.
The role of miRNAs in cardiac protection has long been established.18,19 In particular, miR-126a-3p20 and miR-2421 have been identified to play a role in reducing MIRI through RIPC. Sun et al found reduced expression of miR-185-5p in mice with MIRI and in hypoxia/reoxygenation (H/R) cells and confirmed the targeting interaction between miR-185-5p and lncRNA ROR.22 Recent research has revealed that knockdown of lncRNA LINC00461 alleviates MIRI by upregulating the expression of miR-185-3p to suppress Myd88 expression.23 The apoptosis-related regulatory pathway involving JNK-interacting protein 3 (JIP3) has also been reported to be involved in MIRI.24 Xu et al demonstrated that JIP3 protein expression increases in H9c2 cells subjected to IR injury, and JIP3 overexpression enhances JNK activation and cell apoptosis after IR.24 JIP3, a direct substrate of Jun N-terminal kinases (JNK), plays a pivotal role in the JNK pathway,25 with some miRNAs and lncRNAs functionally associated with JNK.26 In our preliminary experiments, bioinformatics methods have demonstrated target binding sites for miR-185-3p and IPCRL1; however, their regulatory mechanisms in MIRI remain unclear. Therefore, the aim of the present study was to explore the potential molecular mechanisms and actions of IPCRL1, miR-185-3p, and JIP3 in MIRI, aiming to provide new therapeutic options for the treatment of MIRI.
Materials and Methods
Experimental Animals
Specific Pathogen Free (SPF) C57BL/6 male mice, aged 6–8 weeks and weighing 22–26g, were obtained from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). Mice were housed in standard barrier experimental cages at the laboratory animal center of Wenzhou Medical University with a 12/12-hour light/dark cycle at a constant temperature of 20–22°C, with ad libitum access to water and food. Thirty-six C57BL/6 mice were randomly divided into four groups, by one investigator not privy to treatment assignment, namely Sham, MIRI, MIRI+NC, and MIRI+KD-IPCRL1 groups (n = 9 per group). The sample size was determined based on previous studies involving models of MIRI,27,28 in which 6–10 mice per group were found to provide sufficient statistical power. This sample size also aligns with ethical principles aimed at minimizing animal use while ensuring scientific reliability. Adeno-associated virus 9 (AAV9) used in this study was provided by Genechem (Shanghai, China) with the sequence: 5′ GGAGAACAGAAGGAATGTGTA ′3. MIRI model carrying either knockdown IPCRL1 (KD-IPCRL1) or the scramble control (NC) was constructed following standard procedures.15 The mice were intravenously injected with AAV9 constructs (1×1012 vg/mL, 100 μL) via the tail vein, and a MIRI model was established about 2 weeks after the injection. The establishment of the MIRI mouse model was conducted as described in previous research.29 Following induction of anesthesia with isoflurane, the mice were intubated and placed in the right lateral decubitus position. A thoracotomy was performed at the third or fourth left intercostal space to expose the pericardium. The pericardium was then opened using micro-forceps, and the left anterior descending coronary artery was ligated with a 6–0 silk suture to induce myocardial ischemia. After 30 min of myocardial ischemia, the ligation was released to allow for myocardial reperfusion and then the chest was rapidly closed in layers. The mice were resuscitated with 100% oxygen ventilation and transferred to a warming pad. After 180 min, the mice were euthanized under anesthesia, and samples were collected for next experiments. Sham surgery involved all procedures except coronary artery ligation. The occlusion and reperfusion of the left anterior descending artery were confirmed by significant changes in myocardial color.
Cell Culture and Treatment
HL-1 cells were sourced from iCell Bioscience Inc. (Shanghai, China) and cultured in Claycomb medium (iCell Bioscience Inc., Shanghai, China) supplemented with 10% fetal bovine serum (FBS), 1% penicillin, 1% streptomycin, 0.1 mM epinephrine, and 2 mM L-glutamine. The cells were maintained under standard conditions at 37°C and 5% CO2. Cells were used between passages 3 to 8 and cultured for 48–72 h prior to each assay. All experiments were performed in triplicate. To induce H/R injury, HL-1 cells were cultured in MEM containing 1% O2, 94% N2, and 5% CO2 for 2 h followed by 24 h of reoxygenation. miR-185-3p inhibitor and NC were provided by RiboBio Co., Ltd (Guangzhou, China), and siRNA-IPCRL1 (5′ CCCUGCUGGAUGGAGUCAAACCUAA ′3 and 5′UUAGGUUUGACUCCAUCCAGCAGGG ′3) and NC (5′ UUCUCCGAACGUGUCACGUTT ′3 and 5′ ACGUGACACGUUCGGAGAATT ′3) was synthesized by Generay (Shanghai, China). Transfection was performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., USA) in line with the manufacturer’s instructions.
Western Blot Analysis
Total protein was extracted from cells or tissues using RIPA lysis buffer (containing phenylmethylsulfonyl fluoride, PMSF) from Sangon Biotech (Shanghai). The concentration of total protein was determined using a BCA assay kit (Sangon Biotech, Shanghai). The protein samples were separated by 10% SDS-PAGE and transferred to PVDF membranes (Bio-Rad, USA). The membranes were blocked with 5% skimmed milk for 2 h and then incubated at 4°C overnight with the following primary antibodies: anti-JIP3, anti-JNK (Affinity Biosciences, USA), anti-BAX, anti-BCL2, anti–cleaved caspase-3, anti–P-JNK, anti–c-Jun (Cell Signaling technology, USA) and anti–β-Actin (Proteintech Group, USA). The membranes were washed with Tris buffered saline with Tween-20 (TBST) buffer and incubated with HRP-conjugated secondary antibodies at 37°C for 1 h. Protein bands were visualized using ECL (APExBIO, USA) and analyzed for band intensities using ImageJ software.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
Total RNA was extracted using Trizol (Invitrogen, Thermo Fisher Scientific, USA), and its concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA). Samples with A260/A280 ratios between 1.8 and 2.1 were considered of acceptable purity and used for downstream applications. Reverse transcription was performed using RT SuperMix for qPCR (APExBIO, USA) according to the manufacturer’s instructions. qPCR was performed using the SYBR Green method with a LightCycler-480 instrument (Roche, Switzerland) to quantify target gene expression levels. GAPDH and U6 were used as an endogenous control, and data were analyzed using the 2−ΔΔCt method. The PCR cycling program was as follows: Initial denaturation at 95°C for 2 min, 40 cycles of: Denaturation at 95°C for 15s, annealing at 60°C for 30–60s, extension at 72°C for 30s. Primer sequences, accession numbers, annealing temperatures (Tm), and product sizes are summarized in Table S1 (Sangon, Shanghai, China).
Determination of Myocardial Infarct Size by TTC/EB Dual Staining
At the end of reperfusion, the mice were anesthetized, and the chest was reopened at the same location for re-ligation of the coronary artery. A 2% solution of Evans Blue dye (Solarbio, China) was injected intravenously. Rapid bluing of the heart indicated successful perfusion below the ligature. The heart was then excised, washed with phosphate-buffered saline (PBS) solution, and frozen at −20°C. Five 1mm thick heart sections were made, incubated with 2% 2,3,5-triphenyltetrazolium chloride (TTC) solution (Solarbio, China) at 37°C for 40 min, and photographed. Image Pro-Plus 6.0 software was used to analyze the infarct area, where white areas represent infarcted myocardial tissue, and blue areas represent normal myocardial tissue.
Hematoxylin–Eosin (HE)
Cardiac tissue was fixed in 4% paraformaldehyde (Servicebio Technology Co., Ltd., China), dehydrated in ethanol gradient, embedded in paraffin, and sectioned into 5-μm-thick slices for HE staining. The sections were observed under an optical microscope (NE910, Nexcope, USA).
Immunohistochemical (IHC) Detection
IHC analysis was performed to assess the expression levels of JIP3 in cardiac tissue. The sections were deparaffinized by baking and subjected to antigen retrieval by heat. Subsequently, they were immersed in 1% periodic acid for 10 min. Non-strain-specific primary antibodies against JIP3 (1:100, Affinity Biosciences, USA) was then applied and incubated at 4°C overnight. Secondary antibodies were added and incubated at 37°C for 30 min. The sections were sequentially processed with DAB, HE staining, and ethanol treatment. Afterward, they were dehydrated in xylene, sealed with neutral resin, and observed under a microscope (NE910, Nexcope, USA). Morphological analysis was performed using ImageJ software, evaluating six randomly selected high-power fields (200×) per section.
TUNEL Staining
TdT-mediated dUTP-biotin nick end labeling (TUNEL) staining was performed to detect apoptosis in myocardial cells. Sections of the myocardial tissue were fixed in 4% paraformaldehyde, permeabilized with 0.5% Triton X-100, and blocked with 5% bovine serum albumin for 1 h. TUNEL staining was carried out using the TUNEL fluorescein isothiocyanate (FITC) apoptosis detection kit (Vazyme, China) in accordance with the manufacturer’s instructions, followed by counterstaining with DAPI for nuclear visualization. The stained cells were observed under a fluorescence microscope (CKX41, Olympus, Japan).
Enzyme-Linked Immunosorbent Assay (ELISA)
Serum concentrations of cardiac and inflammatory markers were measured using enzyme-linked immunosorbent assay (ELISA) kits, following the manufacturer’s protocols. ELISA kits were as following: CK-MB (SEKM-0152, Solarbio, China), cTnI (SEKM-0153, Solarbio), IL-1β (PI301, Beyotime, China), IL-6 (PI326, Beyotime), and TNF-α (PT512, Beyotime).
CCK-8 Assay
After transfection, logarithmically growing cells were harvested and adjusted to a concentration of about 4,000 cells/well in a 96-well plate. The cells were cultured at 37°C for 24 h, followed by addition of 10 µL of CCK-8 reagent and further incubation for 2 h. Absorbance at 450 nm was measured using a microplate reader (Thermo Fisher Scientific Inc., USA). Each experiment was independently repeated three times.
Flow Cytometry Analysis
Following the treatment, the cells were collected in tubes, centrifuged at 4°C, washed with PBS, resuspended, and incubated with Annexin V–FITC/propidium iodide (PI). Annexin V–FITC and PI fluorescence was measured using a CytoFLEX (Beckman Coulter, USA).
Immunofluorescence Detection
HL-1 cardiomyocytes were fixed in 4% paraformaldehyde (Servicebio Technology Co., Ltd., China), washed with PBS three times, permeabilized with 0.5% Triton X-100 for 1 h at room temperature, and blocked with 10% normal goat serum (Merck, USA, G9023) for 1 h at 37°C. Next, the cells were incubated with JNK antibody (1:500, Affinity Biosciences, USA) at 4°C overnight, followed by incubation with secondary antibodies (Cat#4412S, Cell Signaling Technology, USA) at 37°C. Cell nuclei were stained with DAPI. Immunofluorescence was observed using a fluorescence microscope (Nikon, Japan).
Fluorescence in situ Hybridization (FISH)
FISH was performed using the RiboBio fluorescence in situ hybridization kit (RiboBio, Guangzhou, China) in accordance with the manufacturer’s protocol to detect the localization of IPCRL1 in HL-1 cells. The cells (2×104 cells/well) were seeded onto round coverslips in a 24-well cell culture plate and cultured in DMEM supplemented with 10% FBS for about 24 h, followed by fixation with 4% paraformaldehyde at room temperature. The fixed cells were permeabilized with pre-cooled 1% Triton X-100, washed with 1×PBS, and subjected to hybridization using probes designed against the target RNA. Subsequently, secondary antibodies were used to amplify the signals from IPCRL1 hybridized probes (RiboBio, China). Nuclei were counterstained with DAPI. After mounting with a mounting medium, the stained slides were observed under a confocal microscope.
Dual-Luciferase Reporter Assay
The binding relationship between miR-185-3p and IPCRL1 or JIP3 was predicted through relevant database (https://mirdb.org/ and https://www.targetscan.org/vert_80/). The relationship between miR-185-3p and IPCRL1 or JIP3 was evaluated using dual-luciferase assay. Wild-type (WT) or mutant (MUT) sequences containing the miR-185-3p–binding sites within the 3′-UTR of IPCRL1 or JIP3 were cloned into plasmid vector, and the dual-luciferase reporter plasmids were synthesized by Generay Biotech (Shanghai, China). In accordance with the manufacturer’s instructions, Lipofectamine 2000 (Invitrogen,Thermo Fisher Scientific, USA) was used to co-transfect miRNA mimics (or miR-NC) with IPCRL1 or JIP3 (WT/mut). After about 48h of transfection, HEK293T cells (Wuhan Pricella Biotechnology, China) were lysed following the instructions of the Dual Luciferase Reporter Gene Assay Kit (Yeasen Biotechnology, China), and then luciferase activity was measured.
Statistical Analysis
Experimental data are presented as mean ± standard deviation (SD). Analysis of variance (ANOVA) was performed for comparisons among multiple groups after normality test, whereas comparisons between two groups were conducted using two-tailed Student’s t test. A P value less than 0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism Software (Version 9.5.1, USA).
Results
IPCRL1 Is Highly Expressed in MIRI Myocardium, and Knockdown of IPCRL1 Alleviates Myocardial Injury and Inflammation
To investigate the role of IPCRL1 in MIRI mice, we conducted knockdown experiments of IPCRL1 in mice and detected IPCRL1 levels by qRT-PCR. IPCRL1 expression was significantly upregulated following MIRI but significantly reduced after KD-IPCRL1 AAV injection (p < 0.05, Figure 1A). Subsequently, TTC staining was performed to measure the infarct area. Compared with the Sham group, the infarct area significantly increased in the MIRI group and the MIRI+NC group but decreased after IPCRL1 knockdown (p < 0.05, Figure 1B). Histopathological examination of myocardial cells using HE staining demonstrated cells in the Sham group were neatly arranged and intact, without typical pathological changes. In contrast, the MIRI group and the MIRI+NC group exhibited inflammatory cell infiltration, myocardial cell swelling, and fiber disorder, which were reduced by IPCRL1 knockdown (Figure 1C). Furthermore, we analyzed the effects of IPCRL1 knockdown on inflammatory factors and myocardial injury markers. The expression of serum TNF-α, IL-1β, IL-6, CK-MB, and cTnI increased in MIRI group and MIRI+NC group, along with upregulation of TNF-α mRNA in the tissues (p < 0.05, Figure 1E). IPCRL1 knockdown inhibited the upregulation of these inflammatory factors and myocardial injury markers induced by MIRI (p < 0.05, Figure 1D and F). These results preliminarily demonstrate the involvement of IPCRL1 in the regulation of myocardial injury and inflammation induced by MIRI.
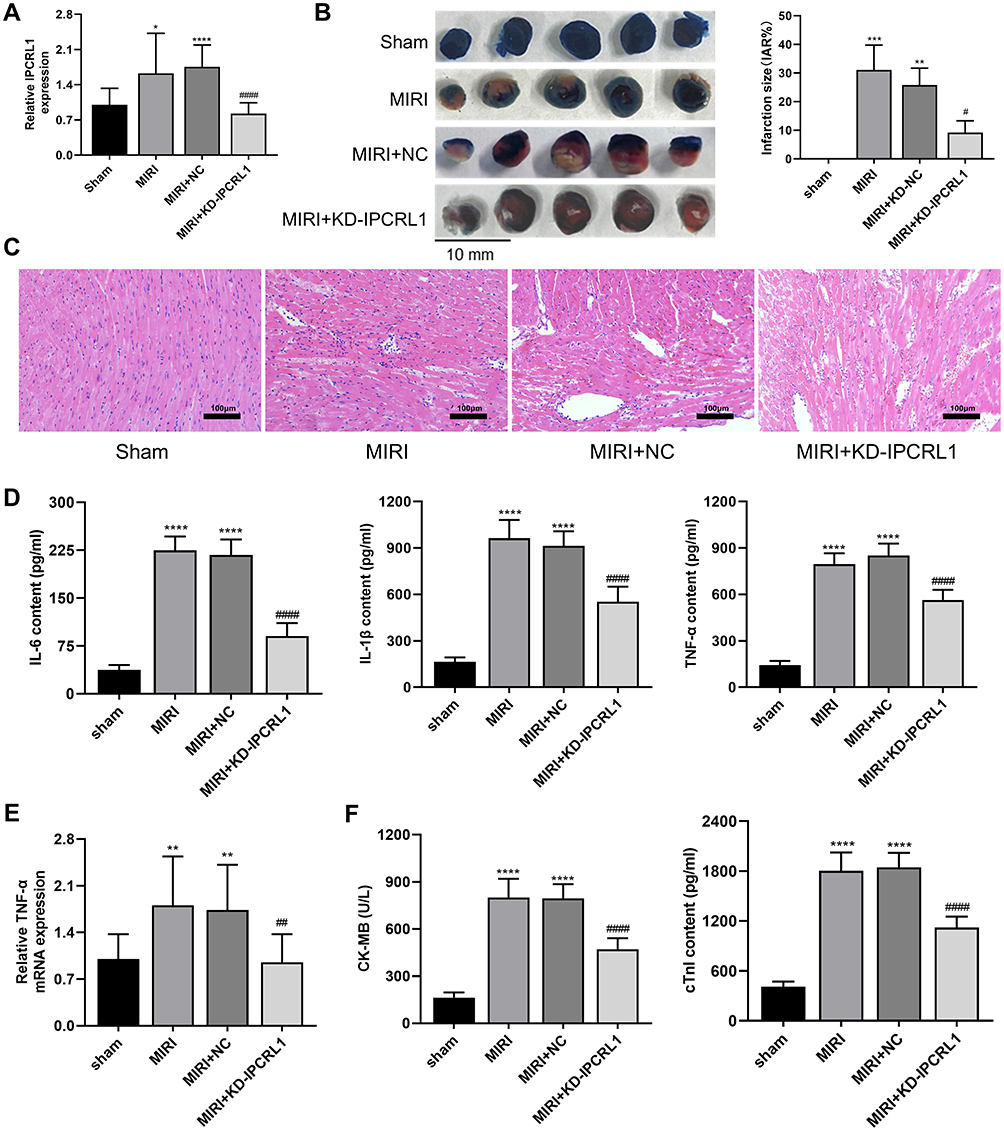 |
Figure 1 Knockdown of IPCRL1 alleviates infarct area, inflammation, and myocardial damage in vivo. (A) Quantification of IPCRL1 expression in myocardial tissue by qRT-PCR. (B) Representative images of TTC/EB staining of heart sections and quantification of myocardial infarct size. (C) Histopathological analysis of myocardial tissue by HE staining. 200×. (D) ELISA assessment of serum levels of inflammatory factors IL-6, IL-1β, and TNF-α in mice. (E) Evaluation of TNF-α expression levels in myocardial tissue by qRT-PCR. (F) ELISA assessment of the expression levels of myocardial injury markers CK-MB and cTnI in mouse serum. For TTC/EB staining, each group included three mice. For other experiments, each group consisted of six mice. Compared with the Sham group, *P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001; compared to the MIRI+NC group, # P < 0.05, ## P < 0.01, #### P < 0.0001. |
Knockdown of IPCRL1 Inhibits Cardiomyocyte Apoptosis After MIRI
To evaluate the effect of IPCRL1 on myocardial cell apoptosis in mice following MIRI, TUNEL staining was performed. The results revealed a significant increase in apoptotic cells in the MIRI group, whereas IPCRL1 knockdown markedly reduced this increase (Figure 2A and B). Similrly, Western blot results demonstrated increased expression of apoptosis-related proteins BAX and cleaved caspase-3 in the MIRI and MIRI+NC groups, along with decreased expression of the anti-apoptotic protein Bcl-2. These protein levels exhibited a trend toward restoration following IPCRL1 knockdown (p < 0.05, Figure 2C and D). These finding suggest that IPCRL1 knockdown inhibits myocardial apoptosis in mice following MIRI.
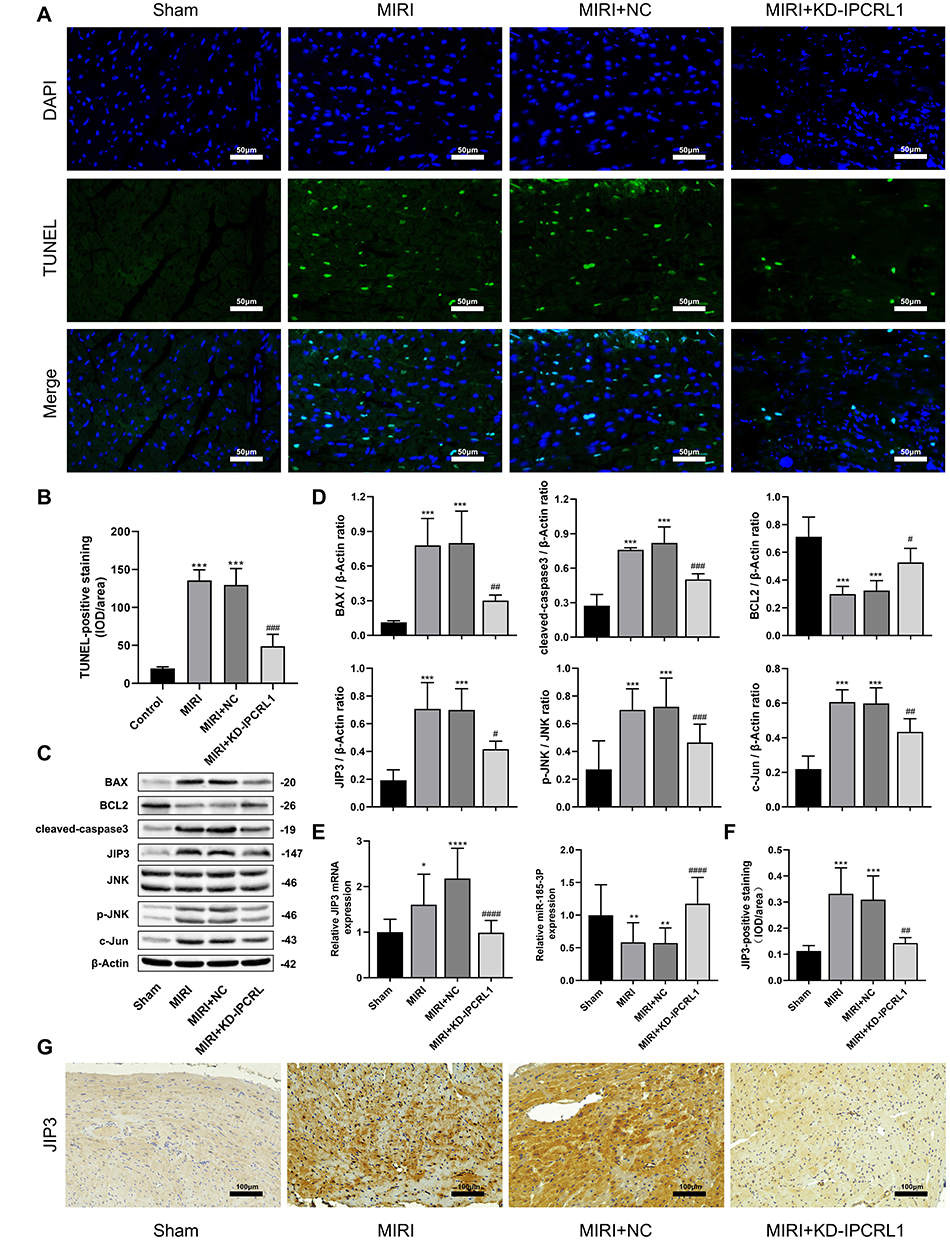 |
Figure 2 Downregulation of IPCRL1 attenuates myocardial ischemia/reperfusion injury in vivo by regulating the expression of miR-185-3p and JNK pathway. (A And B) Representative images and quantification of TUNEL staining of mouse heart sections to evaluate apoptosis. 200×. (C) Representative images and of protein expression levels of BAX, BCL2, cleaved-caspase-3, JIP3, JNK, P-JNK, and c-Jun by Western blot. (D) quantification of protein expression levels of BAX, BCL2, cleaved-caspase-3, JIP3, phosphorylation of JNK, and c-Jun in mouse myocardial tissue by Western blot. (E) Assessment of miR-185-3p and JIP3 mRNA expression level by qRT-PCR. (F and G) Representative images and quantification of immunohistochemical staining of JIP3 in mouse heart sections. 200×. For IHC and TUNEL, each group included three mice. For other experiments, each group consisted of six mice. Compared with the Sham group, *P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001; compared to the MIRI+NC group, # P < 0.05, ## P < 0.01, ### P < 0.001, #### P < 0.0001. |
Knockdown of IPCRL1 Modulates miR-185-3p and JIP3 to Participate in the JNK Pathway
After MIRI, the expression levels of JIP3, phosphorylation levels of JNK, and the downstream pathway-associated protein c-Jun increased, but these protein levels showed a trend toward restoration following IPCRL1 knockdown (p < 0.05, Figure 2C and D). qRT-PCR results revealed a significant increase in JIP3 mRNA expression and a significant decrease in miR-185-3p mRNA expression in the MIRI and MIRI+NC groups compared with the Sham group (p < 0.05, Figure 2E). These alterations were reversed by IPCRL1 knockdown. IHC data analysis further confirmed that IPCRL1 knockdown suppressed the elevated expression of JIP3 in myocardial tissues MIRI mice (p < 0.05, Figure 2F and G). These findings preliminarily suggest that IPCRL1 knockdown may alleviate myocardial injury and apoptosis induced by MIRI by upregulating miR-185-3p and inhibiting the JIP3 and JNK downstream pathways.
Subcellular Localization of IPCRL1 and Validation of the IPCRL1 Knockdown Effect in HL-1
The subcellular localization of lncRNA is closely associated with its biological functions and molecular mechanisms.30 Thus, we investigated the cellular distribution of lncRNA IPCRL1. Using U6 and 18S as positive controls for the nucleus and cytoplasm, respectively, through nuclear-cytoplasmic fractionation and FISH localization, IPCRL1 was found to primarily reside in the cytoplasm (Figure 3A). Furthermore, the efficiency of IPCRL1 knockdown was confirmed by qRT-PCR, showing reduced expression of IPCRL1 in cardiomyocytes after transfection with IPCRL1 knockdown (Figure 3B).
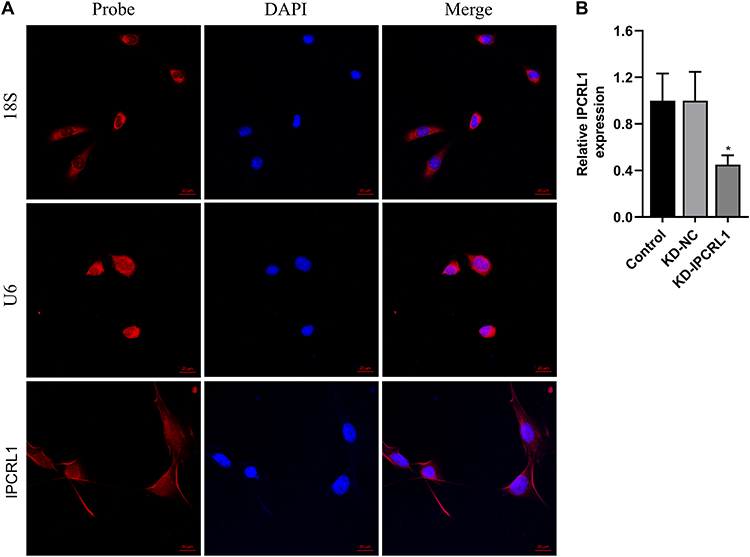 |
Figure 3 Subcellular Localization of IPCRL1 and IPCRL1 Knockdown effect. (A) Fluorescence in situ hybridization (FISH) confocal images depicting the subcellular localization of IPCRL1 (red) within cells. Cell nuclei were stained with DAPI (blue). U6 and 18S served as positive controls for nuclear and cytoplasmic localization, respectively. 18S, 18S rRNA probe utilized for cytoplasmic localization. U6, probe targeting U6 snRNA, employed as a representative for nuclear localization. Scale bar, 20 μm. (B) Assessment of IPCRL1 knockdown by qRT-PCR. Compared with the KD-NC group, * P < 0.05. |
Knockdown of IPCRL1 Attenuates Apoptosis and Inflammation in HL-1 Cells Induced by H/R
Subsequently, we conducted knockdown experiments to investigate the role of IPCRL1 in the H/R cell model. As shown in Figure 4A (p < 0.05), H/R treatment significantly upregulated the expression of IPCRL1 in HL-1 cells, whereas knockdown of IPCRL1 in H/R-induced cells markedly reduced its expression. H/R induction promoted cell apoptosis, which was attenuated upon IPCRL1 knockdown (p < 0.05, Figure 4B). Furthermore, IPCRL1 knockdown prevented the decrease in viability of H/R-induced HL-1 cells and mitigated the upregulation of the inflammatory factor TNF-α induced by H/R (p < 0.05, Figure 4A and C). Apoptosis-related proteins were then evaluated by Western blotting, revealing a significant downregulation of Bcl-2 and upregulation of pro-apoptotic proteins BAX and cleaved caspase-3 following H/R treatment. These alterations were reversed upon IPCRL1 knockdown (p < 0.05, Figure 4D and E).
 |
Figure 4 IPCRL1 regulates the expression of miR-185-3p and JNK pathway to participate in H/R-induced inflammation and apoptosis process in vitro. (A) Evaluation of IPCRL1, JIP3 mRNA, miR-185-3p, TNF-αexpression levels in HL-1 cells by qRT-PCR. (B) Assessment of apoptosis in HL-1 cells by flow cytometry. (C) Assessment of proliferation capacity of HL-1 cells by CCK-8 assay. (D and E) Representative images and Quantification of protein expression levels of BAX, BCL2, cleaved-caspase-3, JIP3, phosphorylation of JNK, and c-Jun in HL-1 cells by Western blot. (F) Immunofluorescence detection of nuclear translocation of JNK. 200×.Each experiment was repeated three times. Compared with the control group, *P < 0.05, ** P < 0.01, *** P < 0.001; compared with the H/R+KD-NC group, # P < 0.05, ## P < 0.01, ### P < 0.001. |
Knockdown of IPCRL1 Affects the Expression Levels of miR-185-3p and JIP3 and Participates in the JNK Pathway in vitro
It has been reported that the JNK signaling pathway is activated during ischemia/reperfusion, leading to the translocation of phosphorylated JNK from the cytoplasm to the nucleus, where it activates c-Jun/AP-1 through phosphorylation, thereby initiating apoptosis signaling and resulting in cell apoptosis.31,32 Western blot analysis revealed that the protein expression of JIP3, phosphorylation levels of JNK, and c-Jun increased after H/R treatment, and displayed a trend towards restoration following IPCRL1 knockdown (P < 0.05, Figure 4D and E). qRT-PCR results indicated that IPCRL1 knockdown increased miR-185-3p expression and decreased JIP3 mRNA expression (P < 0.05, Figure 4A). Immunofluorescence staining further demonstrated nuclear translocation of JNK in the H/R group, which was notably attenuated by IPCRL1 knockdown (Figure 4F). These results preliminarily suggest that IPCRL1 knockdown may alleviate cardiomyocyte apoptosis induced by H/R by upregulating miR-185-3p and inhibiting the downstream JIP3 and JNK pathways.
IPCRL1 Acts as a Sponge of miR-185-3p, and JIP3 is a Direct Target of miR-185-3p
We further explored the possibility of IPCRL1 acting as a ceRNA sponge for miRNA to regulate its target genes.30 Bioinformatics analysis identified complementary binding sites between IPCRL1 and miR-185-3p, while miR-185-3p also exhibited complementary binding sites with JIP3 (https://mirdb.org/ and https://www.targetscan.org/vert_80/) (Figure 5A). Validation was performed through dual-luciferase reporter gene assays, demonstrating that miR-185-3p mimic reduced the luciferase activity of IPCRL1-WT and JIP3-WT cells, with no observed differences in the MUT group (Figure 5B). The results suggested that JIP3 and IPCRL1 competitively bind to the same complementary binding sites with miR-185-3p, forming a ceRNA system, which may act on the downstream JNK pathway.
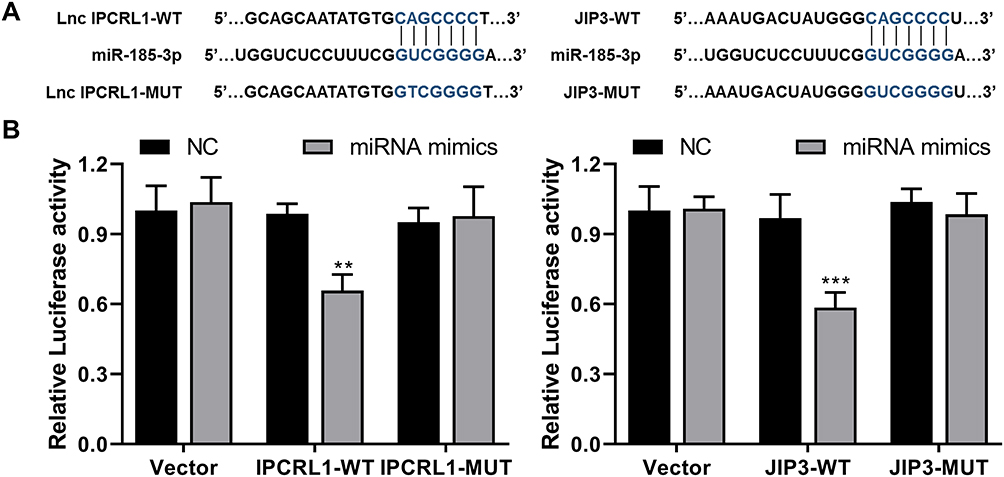 |
Figure 5 Subcellular Localization of IPCRL1 and IPCRL1 Knockdown effect. (A) Prediction of binding sites between miR-185-3p and lncRNA IPCRL1 or JIP3, with the sites indicated in blue. (B) Dual-luciferase validation of binding sites of miR-185-3p on IPCRL1 or JIP3. Compared with the NC group, ** P < 0.01, *** P < 0.001. |
IPCRL1/miR-185-3p/JIP3 Axis Mediates Apoptosis via the JNK Pathway
Next, we investigated whether IPCRL1 regulates cell apoptosis induced by H/R via the miR-185-3p/JIP3 axis. IPCRL1 was knocked down in HL-1 cells, and cell viability was assessed after transfection with miR-185-3p inhibitor. The results showed that the inhibition of miR-185-3p reversed the improvement in cell viability observed after IPCRL1 knockdown (Figure 6A). Additionally, compared to simply knocking down IPCRL1, the miR-185-3p inhibitor led to increased expression of apoptosis-related proteins BAX and cleaved caspase-3, JIP3, and pathway-associated proteins P-JNK and c-Jun, along with decreased expression of the anti-apoptotic protein Bcl-2 (Figure 6B and C). These results suggest that IPCRL1 regulates cell apoptosis during H/R via the miR-185-3p/JIP3 axis.
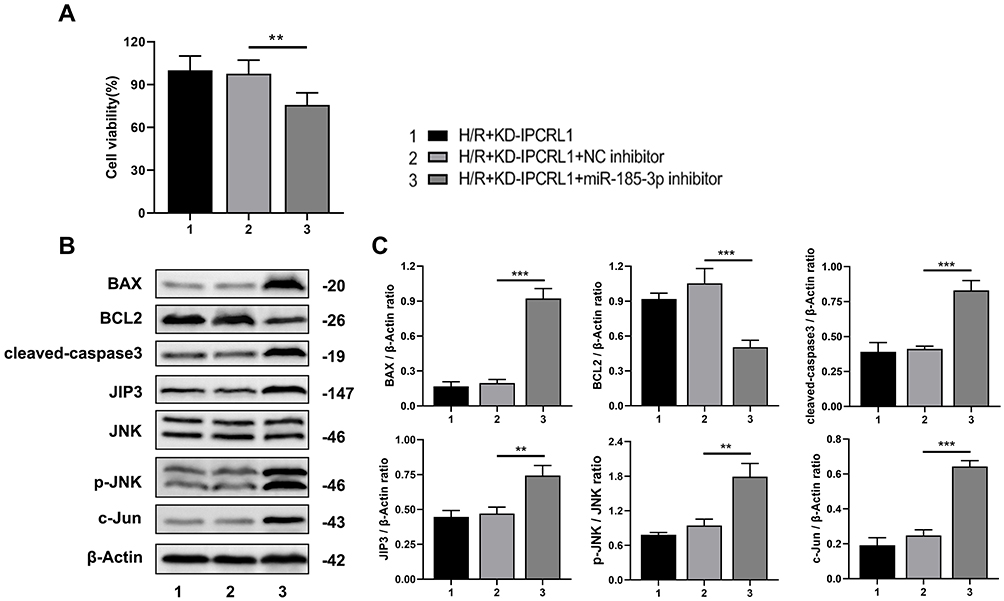 |
Figure 6 The IPCRL1/miR-185-3p/JIP3 axis regulates myocardial cell apoptosis induced by H/R. (A) Assessment of proliferation capacity of HL-1 cells by CCK-8 assay. (B and C) Representative images and quantification of protein expression levels of BAX, BCL2, cleaved-caspase-3, JIP3, JNK, P-JNK, and c-Jun in HL-1 cells by Western blot. Each experiment was repeated three times. ** P < 0.01, *** P < 0.001. |
Discussion
Cardiovascular diseases are the most common and deadly diseases globally, and MIRI can lead to adverse cardiovascular outcomes after myocardial ischemia, cardiac surgery, or CPB. RIPC has been demonstrated to be effective in protecting the heart against MIRI. Previous studies have confirmed significant changes in the expression of certain lncRNAs, including IPCRL1, due to RIPC.7 However, the function and molecular mechanism of IPCRL1 in the field of MIRI remain largely unknown. In this study, we found that inhibiting IPCRL1 could suppress MIRI-induced apoptosis and inflammation through interacting miR-185-3p to target JIP3, thereby reducing infarct size and improving MIRI. To our knowledge, this is the first study to report the role of IPCRL1 in MIRI and its related mechanism (Figure 7).
 |
Figure 7 The conception map of IPCRL1/miR-185-3p/JIP3 axis. IPCRL1 mediated myocardial cell apoptosis in MIRI through its role as a sponge for miR-185-3p, thereby impacting JIP3 expression and downstream JNK signaling pathway. |
Apoptosis and inflammation are important influencing factors in myocardial cell damage during MIRI.4,5,33 Various apoptotic signaling molecules such as BCL2, BAX, and caspase-3 and inflammatory factors such as IL-6, IL-1β, and TNF-α are dysregulated in MIRI myocardium.5,33–35 Thus far, numerous lncRNAs have been reported to participate in myocardial cell apoptosis and inflammation during MIRI.12,15–17 Liang et al reported that the lncRNA ROR/miR-124-3p/TRAF6 axis regulates the inflammatory response induced by MIRI.36 lncRNA CIRKIL binds to the Ku70 protein complex, reduces its nuclear translocation, and inhibits DNA double-strand break repair, exacerbating ischemic myocardial cell apoptosis.37 Similarly, our results indicate that IPCRL1 expression is significantly increased after MIRI, promoting myocardial cell apoptosis and infarct size and increasing the levels of apoptotic factors, inflammatory factors, and myocardial injury markers. Furthermore, we observed an improvement in these MIRI indicators after IPCRL1 downregulation. Similar results were obtained from the in vitro H/R model experiments using HL-1 cell line.
There is evidence suggesting that dysregulation of the lncRNA–miRNA–mRNA network based on ceRNA is implicated in the pathological processes of myocardium.38,39 In human cardiomyocytes subjected to H/R treatment, the expression of miR-185 is decreased and is negatively regulated by lncRNA ROR. The ROR/miR-185-5p/CDK6 axis modulates cellular apoptosis and H/R-induced inflammatory response.22 miRNAs participate in the pathogenesis of various cardiovascular diseases.40 Notably, a previous study showed that miRNA-185-3p overexpression in MIRI myocardium could alleviate myocardial damage.23 In the current study, the expression of miR-185-3p decreased after MIRI and H/R treatment and was confirmed to be regulated by IPCRL1. Inhibition of miR-185-3p countered the effect of IPCRL1 downregulation in HL-1 cells induced by H/R. Multiple signaling pathways are involved in the MIRI process, among which JIP3 is a direct substrate of JNK and plays a role in the JNK pathway, mediating cell apoptosis.25 Similarly, the JAK/STAT pathway participates in reducing MIRI through RIPC.41 Tan et al reported that lncRNA MIAT downregulation mitigates human cardiomyocyte damage induced by hypoxia and glucose deprivation through miR-181a-5p–mediated regulation of the JAK2/STAT3 pathway.42 lncRNA TALNEC2 regulates miR-19a-3p/JNK to alleviate rat cerebral ischemic injury.43 Bioinformatics analysis predicted that IPCRL1, miRNA-185-3p, and JIP3 mRNA have shared binding sites, suggesting the possibility of a ceRNA mechanism among them.44 Our experiments showed that IPCRL1 and JIP3 expression increased in MIRI myocardium, whereas miRNA-185-3p expression decreased, and IPCRL1 downregulation significantly increased miRNA-185-3p expression and decreased JIP3 expression. Furthermore, luciferase assays confirmed the competitive binding of lncRNA to JIP3 mRNA via miRNA-185-3p, affecting myocardial cell MIRI. To sum up, our work showed that the IPCRL1/miRNA-185-3p/JIP3 axis regulates myocardial ischemia reperfusion injury through the JNK pathway.
Due to significant differences of phenotype between humans and mice, researches on animal may not elucidate the complexity of humans MIRI pathological processes appropriately. Secondly, sequence homology analysis showed a high degree of conservation between the sequences dimension of ENST00000574727 and ENSMUST00000123752 (IPCRL1), but the role of ENST00000574727 in humans similar to IPCRL1 remains to be verified.7 Although our current findings are based on preclinical models, they provide a solid foundation for future translational research. From a clinical perspective, these results indicate that IPCRL1 may represent a novel and promising therapeutic target for MIRI. Given its pivotal role in regulating cardiomyocyte apoptosis and inflammatory responses, IPCRL1-targeted strategies—such as siRNA, antisense oligonucleotides, or small-molecule inhibitors—could be developed to effectively attenuate myocardial injury following ischemia-reperfusion. Future studies should further characterize the expression pattern of homologous LncRNA in human myocardial tissue and evaluate its correlation with MIRI severity. The clinical relevance and therapeutic potential of IPCRL1 should also be validated through clinical specimens and prospective studies.
Conclusions
In short, we established that the IPCRL1/miRNA-185-3p/JIP3 axis mediates apoptosis and inflammation in MIRI via the JNK pathway. IPCRL1 acts as a molecular sponge of miRNA-185-3p, increasing the expression of JIP3 and activating the JNK signaling pathway, thereby promoting myocardial cell apoptosis. This provides a new starting point for improving the prognosis of MIRI and may become a new clinical therapeutic target.
Abbreviations
AAV9, adeno-associated virus 9; ceRNA, competitive endogenous RNA; CK-MB, creatine kinase-MB; CPB, Cardiopulmonary bypass; cTnI, cardiac troponin I; ELISA, Enzyme-linked immunosorbent assay; FISH, Fluorescence in situ hybridization; HE, hematoxylin-eosin; IHC, Immunohistochemical; IPCRL1, ischemic preconditioning related lncRNA-1; JNK, c-Jun N-terminal kinase; JIP3, JNK-interacting protein 3; lncRNA, long non-coding RNA; miR, microRNA; MIRI, Myocardial ischemia-reperfusion injury; qRT-PCR, quantitative real-time polymerase chain reaction; RIPC, remote ischemic preconditioning; TNF-α, tumor necrosis factor-alpha; TTC/EB, 2,3,5-triphenyltetrazolium chloride / Evans Blue; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling.
Ethics Approval
All animal experiments procedures pertaining to mice were in strict compliance with the Guide for laboratory animals’ care and use (Eighth Edition,2011, USA). All procedures involving mice were approved by the Animal Research Committee of Wenzhou Medical University (wydw2023-0293).
Acknowledgments
We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (No. 82070274).
Disclosure
The authors report no potential conflicts of interest in this work.
References
1. Liang G, Li Y, Li S, Huang Z. Efficacy of dexmedetomidine on myocardial ischemia/reperfusion injury in patients undergoing cardiac surgery with cardiopulmonary bypass: a protocol for systematic review and meta-analysis. Medicine. 2023;102(9):e33025. doi:10.1097/MD.0000000000033025
2. Wu T, Shi G, Ji Z, Wang S, Geng L, Guo Z. Circulating small extracellular vesicle-encapsulated SEMA5A-IT1 attenuates myocardial ischemia-reperfusion injury after cardiac surgery with cardiopulmonary bypass. Cell Mol Biol Lett. 2022;27(1):95. doi:10.1186/s11658-022-00395-9
3. Gollmann-Tepekoylu C, Graber M, Polzl L, et al. Toll-like receptor 3 mediates ischaemia/reperfusion injury after cardiac transplantation. Eur J Cardiothorac Surg. 2020;57(5):826–835. doi:10.1093/ejcts/ezz383
4. Algoet M, Janssens S, Himmelreich U, et al. Myocardial ischemia-reperfusion injury and the influence of inflammation. Trends Cardiovasc Med. 2023;33(6):357–366. doi:10.1016/j.tcm.2022.02.005
5. Xia B, Ding J, Li Q, et al. Loganin protects against myocardial ischemia-reperfusion injury by modulating oxidative stress and cellular apoptosis via activation of JAK2/STAT3 signaling. Int J Cardiol. 2024;395:131426. doi:10.1016/j.ijcard.2023.131426
6. Liu G, Lv Y, Wang Y, et al. Remote ischemic preconditioning reduces mitochondrial apoptosis mediated by calpain 1 activation in myocardial ischemia-reperfusion injury through calcium channel subunit Cacna2d3. Free Radic Biol Med. 2024;212:80–93. doi:10.1016/j.freeradbiomed.2023.12.030
7. Lou Z, Wu W, Chen R, et al. Microarray analysis reveals a potential role of lncRNA expression in remote ischemic preconditioning in myocardial ischemia-reperfusion injury. Am J Transl Res. 2021;13(1):234–252.
8. Dang Y, Hua W, Zhang X, et al. Anti-angiogenic effect of exo-LncRNA TUG1 in myocardial infarction and modulation by remote ischemic conditioning. Basic Res Cardiol. 2023;118(1):1. doi:10.1007/s00395-022-00975-y
9. Verouhis D, Sorensson P, Gourine A, et al. Long-term effect of remote ischemic conditioning on infarct size and clinical outcomes in patients with anterior ST-elevation myocardial infarction. Catheter Cardiovasc Interv. 2021;97(3):386–392. doi:10.1002/ccd.28760
10. Hausenloy DJ, Kharbanda RK, Moller UK, et al. Effect of remote ischaemic conditioning on clinical outcomes in patients with acute myocardial infarction (CONDI-2/ERIC-PPCI): a single-blind randomised controlled trial. Lancet. 2019;394(10207):1415–1424. doi:10.1016/S0140-6736(19)32039-2
11. Wang F, Wu Y. lncRNA DLX6-AS1 promotes myocardial ischemia-reperfusion injury through mediating the miR-204-5p/FBXW7 axis. Mediators Inflamm. 2023;2023:9380398. doi:10.1155/2023/9380398
12. Tang RF, Li WJ, Lu Y, Wang XX, Gao SY. LncRNA SNHG1 alleviates myocardial ischaemia-reperfusion injury by regulating the miR-137-3p/KLF4/TRPV1 axis. ESC Heart Fail. 2024;11(2):1009–1021. doi:10.1002/ehf2.14660
13. Tan T, Tu L, Yu Y, He M, Zhou X, Yang L. Mechanisms by which silencing long-stranded noncoding RNA KCNQ1OT1 alleviates myocardial ischemia/reperfusion injury (MI/RI)-induced cardiac injury via miR-377-3p/HMOX1. BMC Cardiovasc Disord. 2024;24(1):19. doi:10.1186/s12872-023-03693-y
14. Samidurai A, Olex AL, Ockaili R, et al. Integrated analysis of lncRNA-miRNA-mRNA regulatory network in rapamycin-induced cardioprotection against ischemia/reperfusion injury in diabetic rabbits. Cells. 2023;12(24):2820. doi:10.3390/cells12242820
15. Gao R, Wang L, Bei Y, et al. Long noncoding RNA cardiac physiological hypertrophy-associated regulator induces cardiac physiological hypertrophy and promotes functional recovery after myocardial ischemia-reperfusion injury. Circulation. 2021;144(4):303–317. doi:10.1161/CIRCULATIONAHA.120.050446
16. Xin H, Li C, Cai T, Cao J, Wang M. LncRNA KCNQ1OT1 contributes to hydrogen peroxide-induced apoptosis, inflammation, and oxidative stress of cardiomyocytes via miR-130a-3p/ZNF791 axis. Cell Biol Int. 2022;46(12):2018–2027. doi:10.1002/cbin.11873
17. Bao J, Zhang C, Chen J, et al. LncRNA JPX targets SERCA2a to mitigate myocardial ischemia/reperfusion injury by binding to EZH2. Exp Cell Res. 2023;427(1):113572. doi:10.1016/j.yexcr.2023.113572
18. Bei Y, Lu D, Bar C, et al. miR-486 attenuates cardiac ischemia/reperfusion injury and mediates the beneficial effect of exercise for myocardial protection. Mol Ther. 2022;30(4):1675–1691. doi:10.1016/j.ymthe.2022.01.031
19. Marinescu MC, Lazar AL, Marta MM, Cozma A, Catana CS. Non-coding RNAs: prevention, diagnosis, and treatment in myocardial ischemia-reperfusion injury. Int J Mol Sci. 2022;23(5):2728. doi:10.3390/ijms23052728
20. Li D, Zhao Y, Zhang C, Wang F, Zhou Y, Jin S. Plasma exosomes at the late phase of remote ischemic pre-conditioning attenuate myocardial ischemia-reperfusion injury through transferring miR-126a-3p. Front Cardiovasc Med. 2021;8:736226. doi:10.3389/fcvm.2021.736226
21. Minghua W, Zhijian G, Chahua H, et al. Plasma exosomes induced by remote ischaemic preconditioning attenuate myocardial ischaemia/reperfusion injury by transferring miR-24. Cell Death Dis. 2018;9(3):320. doi:10.1038/s41419-018-0274-x
22. Sun J, Zhu YM, Liu Q, et al. LncRNA ROR modulates myocardial ischemia-reperfusion injury mediated by the miR-185-5p/CDK6 axis. Lab Invest. 2022;102(5):505–514. doi:10.1038/s41374-021-00722-2
23. Gao F, Wang X, Fan T, et al. LncRNA LINC00461 exacerbates myocardial ischemia-reperfusion injury via microRNA-185-3p/Myd88. Mol Med. 2022;28(1):33. doi:10.1186/s10020-022-00452-1
24. Xu B, Zhou Y, K O, Choy PC, Pierce GN, Siow YL. Regulation of stress-associated scaffold proteins JIP1 and JIP3 on the c-Jun NH2-terminal kinase in ischemia-reperfusion. Can J Physiol Pharmacol. 2010;88(11):1084–1092. doi:10.1139/Y10-088
25. Zeke A, Misheva M, Remenyi A, Bogoyevitch MA. JNK signaling: regulation and functions based on complex protein-protein partnerships. Microbiol Mol Biol Rev. 2016;80(3):793–835. doi:10.1128/MMBR.00043-14
26. Ghafouri-Fard S, Abak A, Mohaqiq M, Shoorei H, Taheri M. Interaction between non-coding RNAs and JNK in human disorders. Biomed Pharmacother. 2021;138:111497. doi:10.1016/j.biopha.2021.111497
27. She H, Hu Y, Zhao G, et al. Dexmedetomidine ameliorates myocardial ischemia-reperfusion injury by inhibiting MDH2 lactylation via regulating metabolic reprogramming. Adv Sci. 2024;11(48):e2409499. doi:10.1002/advs.202409499
28. Ge C, Peng Y, Li J, et al. Hydroxysafflor Yellow A alleviates acute myocardial ischemia/reperfusion injury in mice by inhibiting ferroptosis via the activation of the HIF-1alpha/SLC7A11/GPX4 signaling pathway. Nutrients. 2023;15(15):3411. doi:10.3390/nu15153411
29. Dong Y, Kang Z, Zhang Z, et al. Single-cell profile reveals the landscape of cardiac immunity and identifies a cardio-protective Ym-1(hi) neutrophil in myocardial ischemia-reperfusion injury. Sci Bull. 2024;69(7):949–967. doi:10.1016/j.scib.2024.02.003
30. Statello L, Guo CJ, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118. doi:10.1038/s41580-020-00315-9
31. Shvedova M, Anfinogenova Y, Atochina-Vasserman EN, Schepetkin IA, Atochin DN. c-Jun N-terminal kinases (JNKs) in myocardial and cerebral ischemia/reperfusion injury. Front Pharmacol. 2018;9:715. doi:10.3389/fphar.2018.00715
32. Xue HM, Sun WT, Chen HX, He GW, Yang Q. Targeting IRE1alpha-JNK-c-Jun/AP-1-sEH signaling pathway improves myocardial and coronary endothelial function following global myocardial ischemia/reperfusion. Int J Med Sci. 2022;19(9):1460–1472. doi:10.7150/ijms.74533
33. Luo Q, Sun W, Li Z, et al. Biomaterials-mediated targeted therapeutics of myocardial ischemia-reperfusion injury. Biomaterials. 2023;303:122368. doi:10.1016/j.biomaterials.2023.122368
34. Qi B, Zheng Y, Gao W, et al. Alpha-lipoic acid impedes myocardial ischemia-reperfusion injury, myocardial apoptosis, and oxidative stress by regulating HMGB1 expression. Eur J Pharmacol. 2022;933:175295. doi:10.1016/j.ejphar.2022.175295
35. Cao Y, Luo F, Peng J, Fang Z, Liu Q, Zhou S. KMT2B-dependent RFK transcription activates the TNF-alpha/NOX2 pathway and enhances ferroptosis caused by myocardial ischemia-reperfusion. J Mol Cell Cardiol. 2022;173:75–91. doi:10.1016/j.yjmcc.2022.09.003
36. Liang YP, Liu Q, Xu GH, et al. The lncRNA ROR/miR-124-3p/TRAF6 axis regulated the ischaemia reperfusion injury-induced inflammatory response in human cardiac myocytes. J Bioenerg Biomembr. 2019;51(6):381–392. doi:10.1007/s10863-019-09812-9
37. Xiao H, Zhang M, Wu H, et al. CIRKIL exacerbates cardiac ischemia/reperfusion injury by interacting with Ku70. Circ Res. 2022;130(5):e3–e17. doi:10.1161/CIRCRESAHA.121.318992
38. An L, Zhong Y, Tan J, et al. Sevoflurane exerts protection against myocardial ischemia-reperfusion injury and pyroptosis through the circular RNA PAN3/microRNA-29b-3p/stromal cell-derived factor 4 axis. Int Immunopharmacol. 2023;120:110219. doi:10.1016/j.intimp.2023.110219
39. Ye H, Li Y, Li L, Huang Y, Wang J, Gao Q. Construction of a ceRNA network of regulated ferroptosis in doxorubicin-induced myocardial injury. PeerJ. 2023;11:e14767.
40. Sessa F, Salerno M, Esposito M, Cocimano G, Pomara C. miRNA dysregulation in cardiovascular diseases: current opinion and future perspectives. Int J Mol Sci. 2023;24(6):5192. doi:10.3390/ijms24065192
41. Sawashita Y, Hirata N, Yoshikawa Y, Terada H, Tokinaga Y, Yamakage M. Remote ischemic preconditioning reduces myocardial ischemia-reperfusion injury through unacylated ghrelin-induced activation of the JAK/STAT pathway. Basic Res Cardiol. 2020;115(4):50. doi:10.1007/s00395-020-0809-z
42. Tan JK, Ma XF, Wang GN, Jiang CR, Gong HQ, Liu H. LncRNA MIAT knockdown alleviates oxygen-glucose deprivation‑induced cardiomyocyte injury by regulating JAK2/STAT3 pathway via miR-181a-5p. J Cardiol. 2021;78(6):586–597. doi:10.1016/j.jjcc.2021.08.018
43. Luan D, Jiang C. The mechanism of lncRNA TALNEC2 regulating miR-19a-3p/JNK to alleviate cerebral ischemia injury in rats with acute cerebral infarction. Cell Mol Biol. 2022;68(6):17–24. doi:10.14715/cmb/2022.68.6.3
44. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Vitamin D Ameliorates Apoptosis and Inflammation by Targeting the Mitochondrial and MEK1/2-ERK1/2 Pathways in Hyperoxia-Induced Bronchopulmonary Dysplasia
Hu J, Wu Z, Wang H, Geng H, Huo J, Zhu X, Zhu X
Journal of Inflammation Research 2022, 15:4891-4906
Published Date: 25 August 2022
Eriochloa villosa Alleviates Progression of Benign Prostatic Hyperplasia in vitro and in vivo
Baek EB, Hwang YH, Park S, Hong EJ, Won YS, Kwun HJ
Research and Reports in Urology 2022, 14:313-326
Published Date: 24 September 2022
Engeletin Alleviates the Inflammation and Apoptosis in Intervertebral Disc Degeneration via Inhibiting the NF-κB and MAPK Pathways
Li B, Yang X, Zhang P, Guo J, Rong K, Wang X, Cao X, Zhou T, Zhao J
Journal of Inflammation Research 2022, 15:5767-5783
Published Date: 10 October 2022
PANoptosis: A Cell Death Characterized by Pyroptosis, Apoptosis, and Necroptosis
Shi C, Cao P, Wang Y, Zhang Q, Zhang D, Wang Y, Wang L, Gong Z
Journal of Inflammation Research 2023, 16:1523-1532
Published Date: 12 April 2023
TP73-AS1 Regulates MPP+-Induced Cell Inflammation and Apoptosis in SH-SY5Y Cells
Zhang X, Xue L, Li H, Yu X, Dou K, Xie A
Degenerative Neurological and Neuromuscular Disease 2025, 15:81-94
Published Date: 5 September 2025