Back to Journals » Journal of Inflammation Research » Volume 19
Isopropyl 3-(3,4-Dihydroxyphenyl)-2-Hydroxypropanoate Attenuates Experimental Renal Ischemia-Reperfusion Injury by Limiting Sterile Inflammation and Preserving Mitochondrial Function: Involvement of an Nrf2-Associated Cytoprotective Response
Authors Yan X ![]() , Miao Y, Zhang Y, Li Y, Wang Z, Pu Z, Zheng X, Zheng J
, Miao Y, Zhang Y, Li Y, Wang Z, Pu Z, Zheng X, Zheng J ![]()
Received 28 April 2026
Accepted for publication 16 June 2026
Published 10 July 2026 Volume 2026:19 616533
DOI https://doi.org/10.2147/JIR.S616533
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Wenjian Li
Xiaofei Yan,1,2 Yuli Miao,1 Ying Zhang,2 Yixuan Li,2 Ziyi Wang,1 Zhaoqi Pu,1 Xiaohui Zheng,3 Jin Zheng2
1Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China; 2Department of Renal Transplantation, Nephrology Hospital, The First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi, People’s Republic of China; 3Shaanxi Provincial Innovative Drug Research Center, Northwest University, Xi’an, Shaanxi, People’s Republic of China
Correspondence: Xiaofei Yan, Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Department of Renal Transplantation, Nephrology Hospital, The First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi, 710061, People’s Republic of China, Email [email protected] Jin Zheng, Department of Renal Transplantation, Nephrology Hospital, The First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi, 710061, People’s Republic of China, Email [email protected]
Purpose: Renal ischemia-reperfusion injury (IRI) is a major cause of acute kidney injury and a critical determinant of delayed graft dysfunction after kidney transplantation. Sterile inflammation, mitochondrial dysfunction, and oxidative stress are central drivers of renal IRI, yet effective pharmacologic interventions remain limited. Isopropyl 3-(3,4-dihydroxyphenyl)-2-hydroxypropanoate (IDHP), an esterified derivative of Danshensu, has shown anti-inflammatory and antioxidant activity in other injury models, but its role in renal IRI remains unclear.
Methods: A murine bilateral renal IRI model and a hypoxia/reoxygenation (H/R) model in HK-2 cells were used to evaluate the protective effects of IDHP. In the in vivo preventive treatment model, mice received IDHP at 15 mg/kg before ischemia and shortly after reperfusion. In vitro, HK-2 cells were pretreated with IDHP, and 10 μM was selected for subsequent experiments based on its protective efficacy and acceptable tolerability. Renal injury was assessed by serum creatinine (Scr), blood urea nitrogen (BUN), PAS staining, tubular injury scoring, and KIM-1 expression. Inflammation, mitochondrial function, oxidative stress, cytosolic mitochondrial DNA (mtDNA) release, and Nrf2-associated signaling were further examined. The involvement of Nrf2 was explored in HK-2 cells using the pharmacologic inhibitor ML385.
Results: IDHP reduced Scr and BUN by 40.2% and 39.3%, respectively, compared with the IRI group, and alleviated histological features of tubular injury after renal IRI. IDHP also reduced macrophage and neutrophil accumulation, suppressed renal TNF-α, IL-6, and IL-1β expression, and increased IL-10 expression. IDHP preserved mitochondrial complex activity, restored ATP levels, reduced cytosolic mtDNA accumulation, attenuated oxidative stress, and enhanced Nrf2-associated antioxidant signaling in injured kidneys. In H/R-treated HK-2 cells, IDHP improved cell viability, reduced ROS accumulation, limited cytosolic mtDNA release, and decreased secretion of TNF-α, IL-6, MCP-1, and CXCL-8. In this in vitro model, ML385 partially blunted the protective effects of IDHP, suggesting that Nrf2-associated signaling may contribute to its cytoprotective actions.
Conclusion: IDHP attenuates experimental renal IRI and H/R-induced tubular epithelial injury, potentially through coordinated suppression of sterile inflammation, preservation of mitochondrial function, reduction of oxidative stress, and engagement of Nrf2-associated cytoprotective signaling. These findings support further preclinical investigation of IDHP as a candidate renoprotective agent. However, dose-response, therapeutic window, long-term outcomes, and in vivo mechanistic studies remain necessary before clinical translation can be considered.
Keywords: renal ischemia-reperfusion injury, inflammation, mitochondria, Nrf2, oxidative stress, IDHP
Introduction
Renal ischemia-reperfusion injury (IRI) is a central pathogenic process in kidney transplantation and ischemic acute kidney injury.1 It directly contributes to delayed graft function, increases the risk of acute rejection, and adversely affects long-term graft survival.2 Beyond transplantation, renal IRI also occurs in clinical settings such as cardiac surgery, severe trauma, shock, and other conditions involving transient renal hypoperfusion.3 Despite advances in perioperative management and supportive care, pharmacologic strategies that directly target renal IRI remain limited.4 Because renal IRI often occurs in clinically predictable settings, identifying pharmacologic agents or intervention strategies suitable for prophylactic or peri-reperfusion use remains necessary.
Sterile inflammation, oxidative stress, and mitochondrial dysfunction are major drivers of renal IRI.5 During ischemia and reperfusion, injured tubular epithelial cells release danger-associated signals and promote neutrophil and macrophage infiltration.6,7 These immune cells amplify tissue damage through cytokine release, protease production, and oxidant generation.8 Mitochondria are deeply involved in this process. Mitochondrial respiratory dysfunction leads to excessive reactive oxygen species (ROS) generation, bioenergetic failure, and release of mitochondrial DNA (mtDNA) into the cytosol.9 Cytosolic mtDNA can act as a danger-associated molecular pattern and contribute to innate inflammatory activation.10 Therefore, an intervention strategy that simultaneously restrains inflammation, limits oxidative stress, and preserves mitochondrial integrity may be particularly valuable in renal IRI.
Salvia miltiorrhiza contains multiple bioactive compounds with antioxidant and anti-inflammatory activities.11 Several Salvia miltiorrhiza-derived compounds have shown protective effects in experimental renal IRI. Nrf2-associated antioxidant responses are relevant to renal IRI because they contribute to redox homeostasis, inflammatory modulation, and mitochondrial stress adaptation.12–14 Compounds such as tanshinone IIA, salvianolic acid B, and cryptotanshinone have been reported to attenuate renal IRI through mechanisms involving oxidative stress, apoptosis, inflammation, pyroptosis, or Nrf2-related signaling.15–17 These studies support the general renoprotective potential of Salvia miltiorrhiza-derived compounds.
IDHP is an esterified derivative of Danshensu, a phenolic acid associated with Salvia miltiorrhiza.11,18 Compared with Danshensu, this esterified structure has been reported to improve lipophilicity and metabolic stability, which may facilitate cellular uptake and intracellular activity.18,19 Previous studies have suggested that IDHP exerts anti-inflammatory, antioxidant, anti-ischemia/reperfusion, and mitochondria-protective effects in inflammatory and stress-related injury models, including cardiomyocyte injury, pulmonary injury, and activated microglia.19–22 Thus, IDHP provides a distinct pharmacologic rationale relative to other previously studied Salvia miltiorrhiza-derived compounds. Nevertheless, whether IDHP protects against renal IRI remains unknown, and its potential effects on mitochondrial preservation, cytosolic mtDNA accumulation, sterile inflammation, and Nrf2-associated cytoprotective signaling have not been fully explored.
In the present study, we evaluated the renoprotective effects of IDHP in a murine renal IRI model and an H/R-induced tubular epithelial cell model. We tested whether IDHP pretreatment attenuates renal IRI by suppressing sterile inflammation, preserving mitochondrial function, and reducing oxidative stress, with accompanying changes in Nrf2-associated cytoprotective signaling. By combining in vivo renal injury assessment with in vitro pharmacologic inhibition of Nrf2, this study aimed to define the renoprotective profile of IDHP and to clarify whether Nrf2-associated signaling may contribute to its protective effects.
Materials and Methods
Reagents and Antibodies
Anti-KIM-1 (NBP1-76701ss) was purchased from Novus Biologicals (Centennial, CO, USA), anti-F4/80 (70076T) from Cell Signaling Technology (Danvers, MA, USA), anti-Nrf2 (66,504-1-Ig) from Proteintech (Wuhan, China), and anti-Ly6G (GB11229) from Servicebio (Wuhan, China). Cy3-conjugated goat anti-rabbit IgG (GB21303), horseradish peroxidase-conjugated goat anti-mouse IgG (GB23301), goat anti-rabbit IgG (GB23303), and the bicinchoninic acid protein assay kit were obtained from Servicebio (Wuhan, China). ML385 (HY-100523) was purchased from MedChemExpress (Monmouth Junction, NJ, USA). IDHP (purity >98% by high-performance liquid chromatography) was provided by the Hong-Taoism Research Institute of Analytical Science and Technology (China). For in vivo experiments, IDHP was weighed immediately before use and freshly prepared in sterile normal saline. For in vitro experiments, IDHP was freshly prepared in sterile phosphate-buffered saline (PBS) and diluted into the culture medium to the indicated final concentrations. The corresponding vehicle controls received the same volume of normal saline or PBS.
Cell Culture and H/R Treatment
The human tubular epithelial HK-2 cell line was obtained from the Cell Center of Shanghai Institutes for Biological Sciences and has since been maintained in our laboratory. Cells were routinely tested for mycoplasma and used within 15 passages. HK-2 cells were cultured in DMEM/F12 supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin at 37°C in a humidified atmosphere containing 5% CO2. To induce H/R injury, cells were exposed to 0.5% O2, 94.5% N2, and 5% CO2 for 24 h and then returned to normoxic conditions for 6 h. Control cells were maintained under normoxic conditions. IDHP was freshly prepared in sterile PBS, diluted into the culture medium to the indicated final concentrations, and added 30 min before hypoxia. The control and H/R groups received the same volume of sterile PBS. For Nrf2 inhibition experiments, ML385 was added at a final concentration of 5 μM 30 min before IDHP treatment.
Cell Viability Assay
HK-2 cell viability was determined by MTT assay. Cells were seeded into 96-well plates at 1×10^4 cells/well and incubated overnight. After treatment, cells were incubated with MTT solution (0.5 mg/mL) for 4 h. Formazan crystals were dissolved in dimethyl sulfoxide, and absorbance was measured at 570 nm.
Animals and Experimental Design
Specific pathogen-free male C57BL/6J mice aged 9–10 weeks were obtained from Dossy Experimental Animals Co., Ltd. (Chengdu, China). All mice were acclimatized to the animal facility for 7 days before experimentation and housed under standard conditions, including a temperature of 19–21°C, a 12 h light/dark cycle, and ad libitum access to food and water. All animal procedures were approved by the Animal Care and Use Committee of Xi’an Jiaotong University (Approval No. XJTU-IAUC/765/2019, #19765) and were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals of Xi’an Jiaotong University, the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and the ARRIVE 2.0 guidelines.
To investigate the effect of IDHP on renal IRI, a total of 18 mice were randomly assigned to three groups (n = 6 per group) using a block randomization method with a block size of 6. A formal a priori power calculation was not performed; the sample size was selected based on previous renal IRI studies using similar experimental designs and endpoints.16,17 The random sequence was generated in Microsoft Excel by assigning random numbers to group labels and sorting accordingly. The allocation sequence was concealed in sequentially numbered opaque envelopes, which were opened only at the time of assignment. The three groups were as follows: (1) sham-operated group, (2) IRI + Vehicle (saline) group, and (3) IRI + IDHP group. All surgeries, drug administrations, sample collections, and outcome assessments were performed by investigators blinded to group allocation. Group allocation was performed by an independent researcher who was not involved in subsequent procedures or outcome assessments. The group key was revealed only after all data had been collected.
The IRI model was established by bilaterally clamping the renal pedicles for 27 min under anesthesia induced by intraperitoneal injection of 1.25% tribromoethanol at 0.2 mL/10 g. The sham-operated group underwent the same surgical procedure without vascular occlusion. For the treatment group, IDHP was freshly prepared in sterile normal saline and administered intraperitoneally at 15 mg/kg 30 min before ischemia and again 30 min after reperfusion. The vehicle group received an equivalent volume of sterile normal saline on the same schedule. The 15 mg/kg dose was selected based on previous preclinical evidence showing protective effects of IDHP in inflammatory or ischemia-related injury models at comparable dosing ranges.19 Because no renal IRI-specific dose-response data for IDHP were available, this dose was used to evaluate whether IDHP confers renal protection in the present experimental model.
Throughout surgery and recovery, body temperature was maintained at 37°C using a heating pad. Postoperative analgesia was provided by subcutaneous injection of carprofen at 10 mg/kg. The order of surgeries alternated among groups. Before, during, and after surgery, all mice were housed under identical conditions, operated on by the same surgeon, maintained under controlled temperature, and sampled at identical time points. Each individual mouse was considered an experimental unit. No specific inclusion or exclusion criteria were established a priori. All mice that successfully underwent the surgical procedure without intraoperative death were included in the study, and no animals were excluded from the analysis. At 24 h after reperfusion, mice were anesthetized with tribromoethanol (1.25%, 0.2 mL/10 g, i.p). Blood was collected by cardiac puncture using a 1 mL syringe. Immediately after blood collection, mice were euthanized by cervical dislocation. Kidney tissues were then rapidly harvested for subsequent analysis.
Assessment of Renal Function and Histologic Injury
Serum creatinine (Scr) and blood urea nitrogen (BUN) were measured using a Hitachi 7060 automatic biochemical analyzer. Kidney tissues were fixed, embedded in paraffin, and sectioned at 3 μm. Sections were stained with periodic acid-Schiff. Tubular injury was scored in a blinded manner in at least 10 fields per sample according to a previously described semiquantitative method.23 KIM-1 expression was assessed by immunofluorescence staining.
Immunostaining
Paraffin-embedded kidney sections were deparaffinized, rehydrated, subjected to antigen retrieval, and blocked according to standard immunostaining procedures as previously described.24 Primary antibodies were used at the following dilutions: KIM-1, 1:100; Ly6G, 1:100; F4/80, 1:50; and Nrf2, 1:100. KIM-1 expression was assessed by immunofluorescence staining. After incubation with the primary antibody, sections were incubated with a species-appropriate fluorophore-conjugated secondary antibody in the dark, counterstained with DAPI, and imaged using a fluorescence microscope. F4/80, Ly6G, and Nrf2 were assessed by immunohistochemistry using standard chromogenic detection. F4/80 and Ly6G staining were used to evaluate inflammatory cell infiltration, and Nrf2 staining was used to assess nuclear localization.
Quantitative Real-Time Polymerase Chain Reaction
Total RNA was isolated with TRIzol reagent and reverse-transcribed using the MightyScript RT reagent kit. Quantitative polymerase chain reaction was performed with SYBR Green master mix on a QuantStudio 3 system. Relative expression was calculated by the 2^-ΔΔCt method using β-actin as the internal control. Primer sequences are listed in Table 1.
 |
Table 1 Primer Sequences Used for qPCR |
Mitochondrial Complex Activity, ATP, and Cytosolic Mitochondrial DNA
Mitochondria were isolated from renal tissues or HK-2 cells using commercial kits from Abcam (Cambridge, UK). Complex II activity was measured using kit ab109908, and complex IV activity was measured using kit ab109911 according to the manufacturer’s instructions. ATP levels were determined using a luciferase-based ATP assay kit (Beyotime, Shanghai, China).
For cytosolic mtDNA measurement, cytosolic fractions were prepared using a mitochondria/cytosol fractionation procedure under ice-cold conditions. Briefly, kidney tissues or HK-2 cells were homogenized and subjected to differential centrifugation to remove nuclei, cell debris, and intact mitochondria. The supernatant was collected as the cytosolic fraction. An aliquot of cytosolic fraction was reserved for fractionation quality control by Western blotting before mtDNA extraction. β-actin was used as a cytosolic marker, and TOMM20 was used as a mitochondrial marker. Cytosolic fractions showing clear β-actin enrichment and absent or minimal TOMM20 signal were considered acceptable for subsequent cytosolic mtDNA extraction and quantitative PCR analysis. The fractionation quality-control result is shown in Supplementary Figure S1. DNA from the cytosolic fraction was extracted using the TIANamp Genomic DNA Kit and quantified by quantitative polymerase chain reaction.
Oxidative Stress and Reactive Oxygen Species Assays
Malondialdehyde (MDA), oxidized glutathione (GSSG), and total superoxide dismutase (t-SOD) were measured using commercial kits according to the manufacturers’ instructions. Intracellular reactive oxygen species in HK-2 cells were quantified with H2DCFDA. Fluorescence was measured at excitation and emission wavelengths of 490 and 520 nm, respectively.
Enzyme-Linked Immunosorbent Assay
Supernatant concentrations of IL-6, TNF-α, MCP-1, and CXCL-8 were determined using commercially available enzyme-linked immunosorbent assay (ELISA) kits according to the manufacturers’ instructions.
Statistical Analysis
Statistical analyses were performed using SPSS 22.0 (IBM Corp., Armonk, NY, USA). Continuous variables are presented as mean ± SD. Semiquantitative tubular injury scores were treated as ordinal variables and are presented as median with interquartile range. For continuous variables, data distribution and homogeneity of variance were assessed using the Shapiro–Wilk test, Levene’s test, and visual inspection of data distribution. When parametric assumptions were considered acceptable, comparisons among multiple groups were performed using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test. Semiquantitative tubular injury scores were treated as ordinal variables and analyzed using the Kruskal–Wallis test followed by Dunn’s multiple-comparison test. For in vitro experiments, data were obtained from at least three independent biological replicates unless otherwise stated. Molecular and cell-based endpoints were considered exploratory and were interpreted cautiously in the context of multiple comparisons. A two-sided p-value < 0.05 was considered statistically significant.
Results
IDHP Reduced Tubular Injury in H/R-Treated HK-2 Cells and in Mice Subjected to Renal IRI
We first examined whether IDHP exhibited intrinsic cytotoxicity in HK-2 cells. IDHP did not significantly reduce cell viability at concentrations up to 100 μM after 24 h of exposure, indicating acceptable in vitro tolerability (Figure 1A). Under H/R conditions, IDHP improved HK-2 cell viability across the tested concentration range (Figure 1B). Although several concentrations showed protective effects, 10 μM produced a robust response that approached the apparent efficacy plateau, whereas higher concentrations did not provide further obvious improvement. Therefore, 10 μM was selected as the working concentration for subsequent mechanistic experiments.
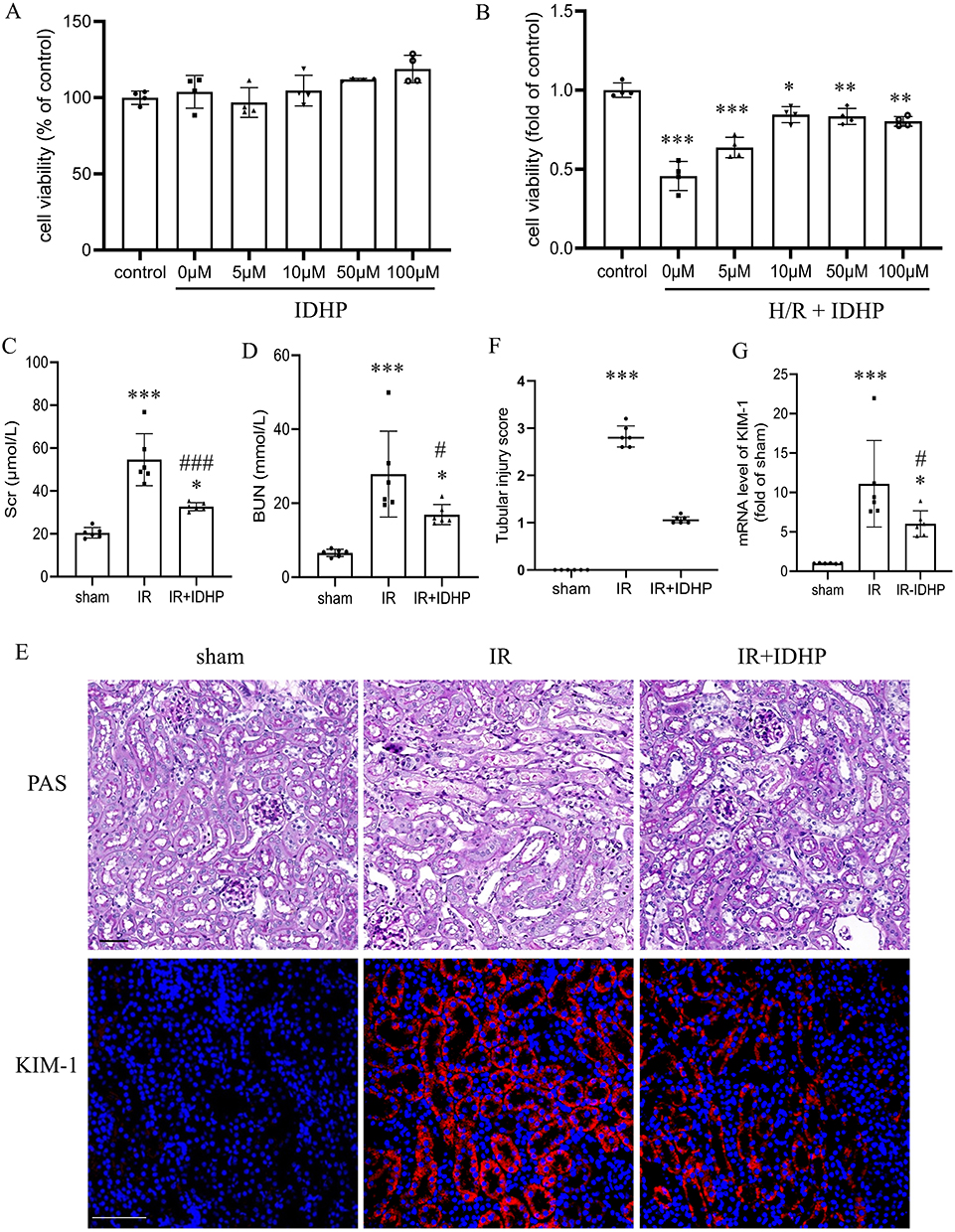 |
Figure 1 IDHP ameliorates renal tubular injury in vitro and in vivo. (A) Effect of IDHP alone (0–100 μM) on HK-2 cell viability. (B) Effect of IDHP pretreatment on the viability of HK-2 cells subjected to hypoxia/reoxygenation (H/R). (C) Scr levels. (D) BUN levels. (E) Representative PAS staining and immunofluorescence staining for KIM-1 in kidney sections from the sham, IRI, and IRI + IDHP groups. (F) Semiquantitative tubular injury scores. Each point represents one mouse. Tubular injury scores are presented as median with interquartile range and were analyzed using the Kruskal–Wallis test followed by Dunn’s multiple-comparison test. (G) Renal KIM-1 mRNA expression. Data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001 vs. control or sham group; #p < 0.05, ###p < 0.001 vs. IR group. |
We next evaluated the in vivo effect of IDHP. Compared with the sham group, mice subjected to renal IRI showed marked increases in Scr and BUN (Figure 1C and D), confirming successful induction of acute kidney injury. IDHP treatment significantly reduced both indicators of renal dysfunction. Histologically, IRI caused substantial tubular injury characterized by tubular dilation, cast formation, and loss of brush border architecture, whereas these pathological changes appeared attenuated in IDHP-treated mice (Figure 1E). Tubular injury scores increased markedly after IRI and were numerically lower after IDHP treatment, although the difference between the IRI and IDHP-treated groups did not reach statistical significance (Figure 1F). In parallel, renal KIM-1 expression was significantly reduced in the IDHP-treated group (Figure 1G).
IDHP Suppressed Sterile Inflammation After Renal IRI
Because inflammatory cell recruitment is a major determinant of post-ischemic tissue damage, we examined inflammatory infiltration in kidney sections. IRI caused pronounced accumulation of F4/80-positive macrophages and Ly6G-positive neutrophils in the renal interstitium. IDHP significantly reduced both macrophage and neutrophil infiltration, with a more pronounced reduction in Ly6G-positive cells (Figure 2A–C).
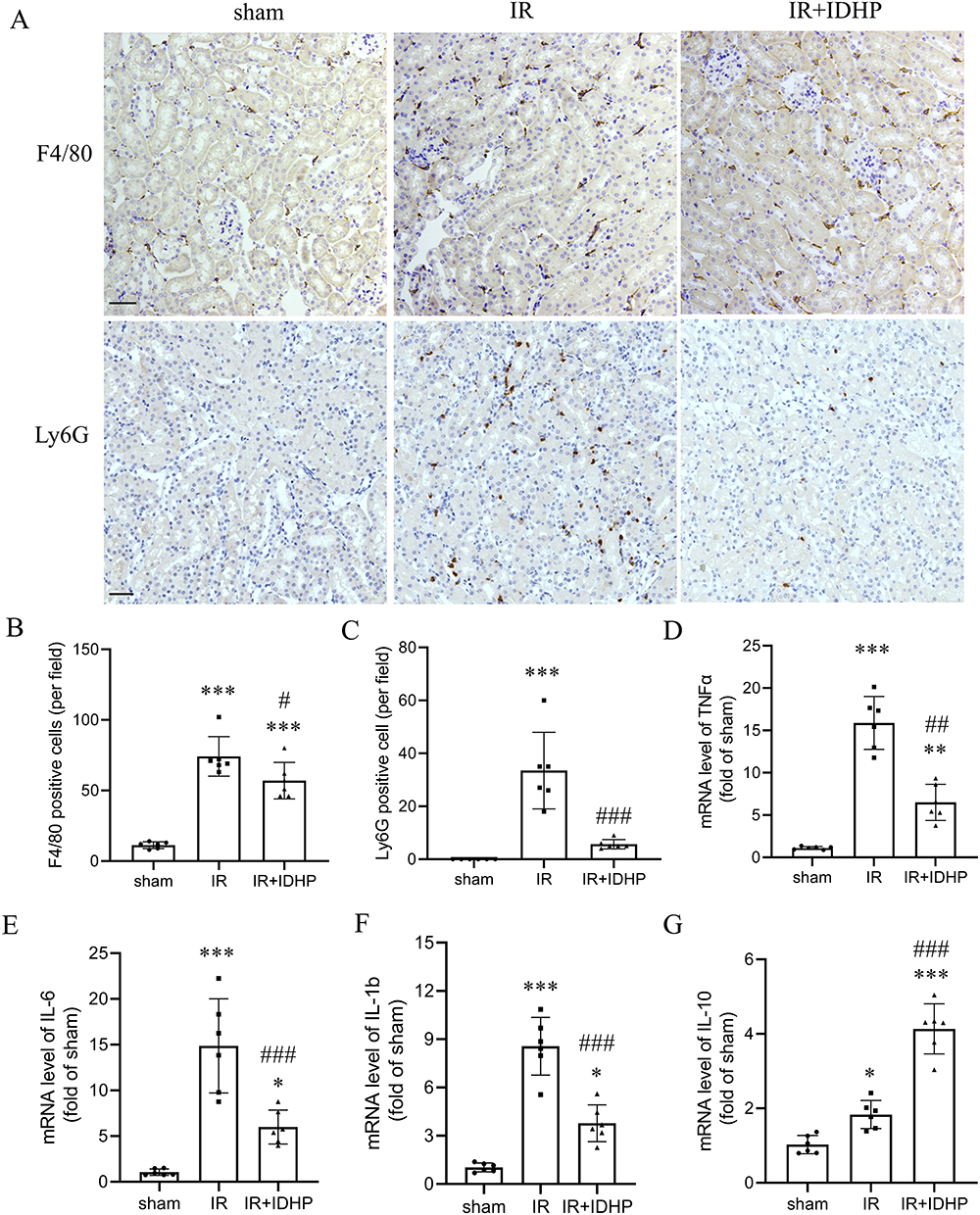 |
Figure 2 IDHP suppresses inflammatory cell infiltration and inflammatory cytokine expression after renal IR. (A) Representative immunohistochemical staining for F4/80 and Ly6G in kidney sections from the sham, IRI, and IRI + IDHP groups. (B) Quantification of F4/80-positive cells per field. (C) Quantification of Ly6G-positive cells per field. (D–G) Renal mRNA expression of TNF-α, IL-6, IL-1β, and IL-10. Data are presented as mean ± SD. *p < 0.05, ***p < 0.001 vs. sham group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. IR group. |
At the transcriptional level, renal IRI markedly increased the expression of the pro-inflammatory cytokines TNF-α, IL-6, and IL-1β. IDHP significantly blunted these increases (Figure 2D–F). In contrast, renal IL-10 expression was increased in the IDHP-treated group (Figure 2G). Together, these data suggest that IDHP alleviated the sterile inflammatory response triggered by renal IRI.
IDHP Preserved Mitochondrial Function and Reduced Cytosolic Mitochondrial DNA Release in Injured Kidneys
Given the central role of mitochondria in ischemic renal injury, we next examined mitochondrial function. Renal IRI significantly reduced mitochondrial complex II and complex IV activities and decreased ATP content. IDHP significantly improved these parameters, indicating preservation of mitochondrial bioenergetic function (Figure 3A–C).
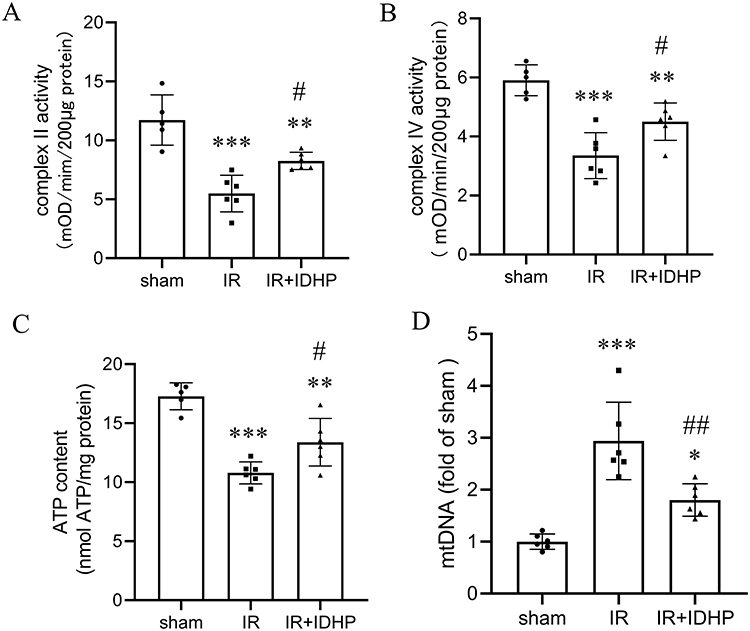 |
Figure 3 IDHP preserves mitochondrial function and reduces cytosolic mitochondrial DNA release in kidneys subjected to IR. (A) Mitochondrial complex II activity in renal tissue. (B) Mitochondrial complex IV activity in renal tissue. (C) ATP content in renal tissue. (D) Cytosolic mitochondrial DNA (mtDNA) levels in renal tissue. Data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001 vs. sham group; #p < 0.05, ##p < 0.01, vs. IR group. |
We also examined cytosolic mtDNA as an index of mitochondrial damage-associated inflammatory signaling. Cytosolic mtDNA levels were markedly increased after renal IRI, whereas IDHP significantly reduced mtDNA accumulation in the cytosolic fraction (Figure 3D). These findings suggest that IDHP preserves mitochondrial bioenergetic function and reduces cytosolic mtDNA accumulation, which is consistent with attenuation of mitochondrial danger signaling.
IDHP Attenuated Oxidative Stress and Enhanced Nrf2-Associated Antioxidant Signaling in the Injured Kidney
Because mitochondrial injury and inflammation are closely linked to oxidative stress, we measured several oxidative stress-related indicators. Renal IRI increased malondialdehyde and oxidized glutathione levels and reduced total superoxide dismutase activity, consistent with substantial oxidative damage. IDHP reversed these changes, indicating improved redox homeostasis (Figure 4A–C).
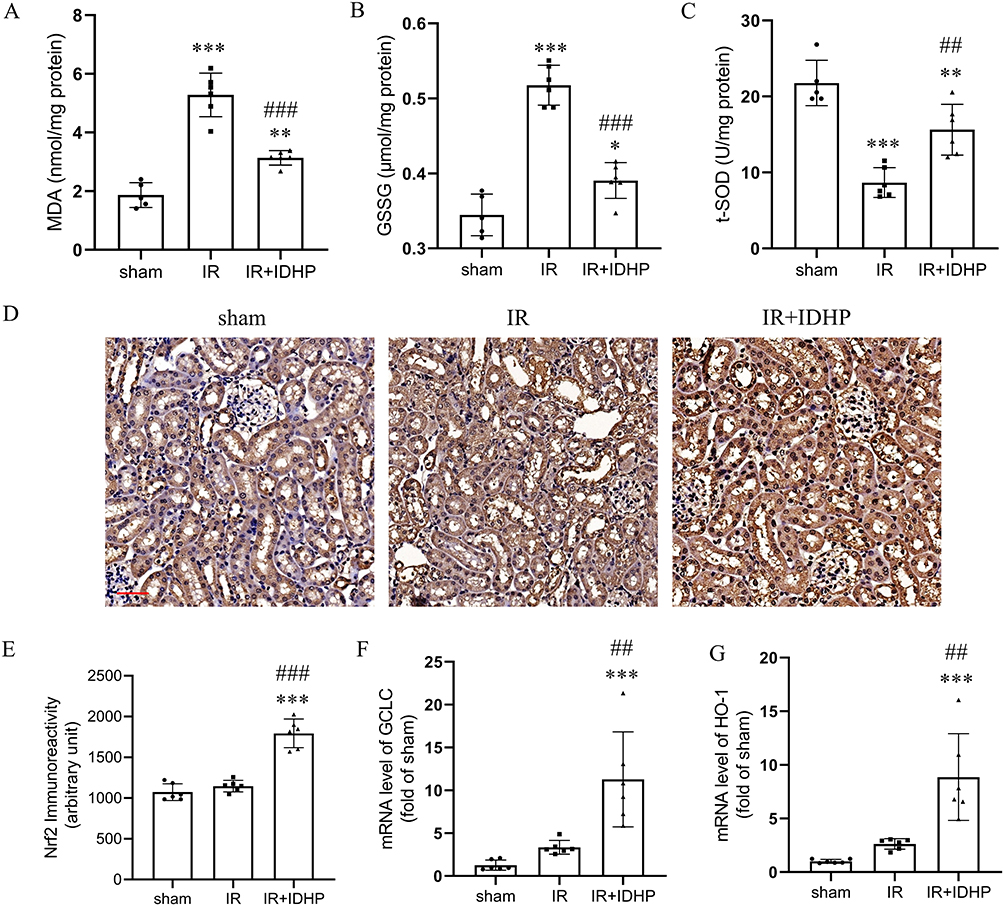 |
Figure 4 IDHP attenuates oxidative stress and enhances Nrf2-associated signaling in kidneys subjected to IR. (A) MDA levels in renal tissue. (B) GSSG levels in renal tissue. (C) T-SOD activity in renal tissue. (D) Representative immunohistochemical staining for Nrf2 in kidney sections from the sham, IR, and IR + IDHP groups. (E) Quantification of renal Nrf2 immunoreactivity. (F) Renal mRNA expression of GCLC. (G) Renal mRNA expression of HO-1. Data are presented as mean ± SD. **p < 0.01, ***p < 0.001 vs. sham group; ##p < 0.01, ###p < 0.001 vs. IR group. |
Nrf2 is a master regulator of antioxidant defense and a known modulator of inflammatory responses. In sham and untreated IRI kidneys, Nrf2 staining was weak and predominantly cytoplasmic. IDHP increased overall Nrf2 expression and promoted its nuclear localization. In parallel, the Nrf2 target genes HO-1 and GCLC subunit were significantly upregulated in IDHP-treated kidneys (Figure 4D–G). These results are consistent with enhanced Nrf2-associated antioxidant signaling in IDHP-treated kidneys after renal IRI.
IDHP Reduced Mitochondrial Dysfunction, Oxidative Stress, and Inflammatory Mediator Release in H/R-Treated HK-2 Cells
To further explore the direct effects of IDHP on tubular epithelial cells, we analyzed mitochondrial and inflammatory outcomes in H/R-treated HK-2 cells. IDHP significantly restored complex II activity and ATP production in H/R-injured cells. It also reduced intracellular reactive oxygen species accumulation and decreased cytosolic mitochondrial DNA levels (Figure 5A–D).
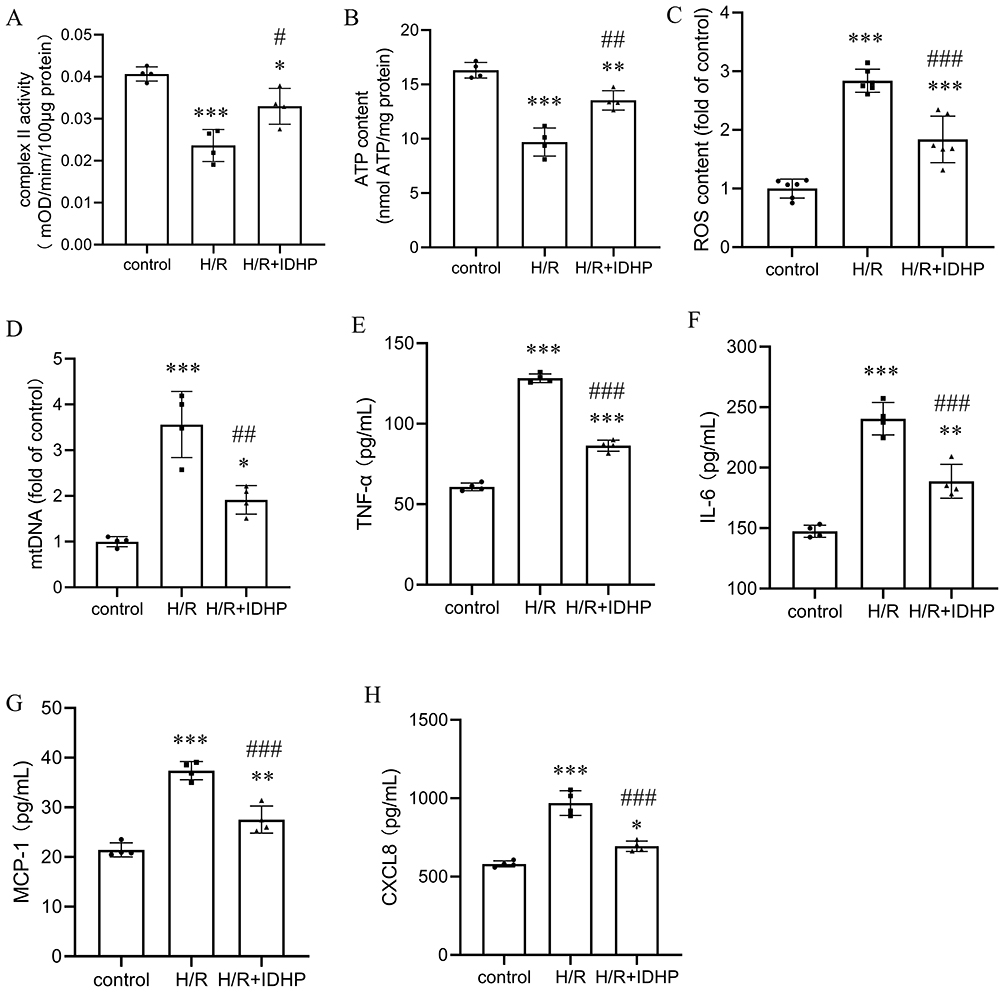 |
Figure 5 IDHP directly improves mitochondrial function and suppresses inflammatory mediator release in H/R-treated HK-2 cells. (A) Mitochondrial complex II activity. (B) ATP content. (C) Intracellular ROS levels. (D) Cytosolic mtDNA levels. (E–H) Secretion of TNF-α, IL-6, MCP-1, and CXCL-8 measured by ELISA. Data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001 vs. control group; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. H/R group. |
Because injured tubular epithelial cells can actively contribute to renal inflammation, we measured inflammatory mediators in cell supernatants. H/R significantly increased secretion of TNF-α, IL-6, MCP-1, and CXCL-8, whereas pretreatment with IDHP significantly suppressed all four mediators (Figure 5E–H). These data suggest that IDHP can attenuate inflammatory mediator release from tubular epithelial cells under ischemic stress.
Nrf2 Inhibition Blunted the Protective Effects of IDHP in HK-2 Cells
To examine whether Nrf2 participates in the protective actions of IDHP, HK-2 cells were treated with the Nrf2 inhibitor ML385. ML385 partially attenuated the ability of IDHP to restore complex II activity and ATP content after H/R. ML385 also partially weakened the suppressive effects of IDHP on ROS accumulation and cytosolic mtDNA release (Figure 6A–D).
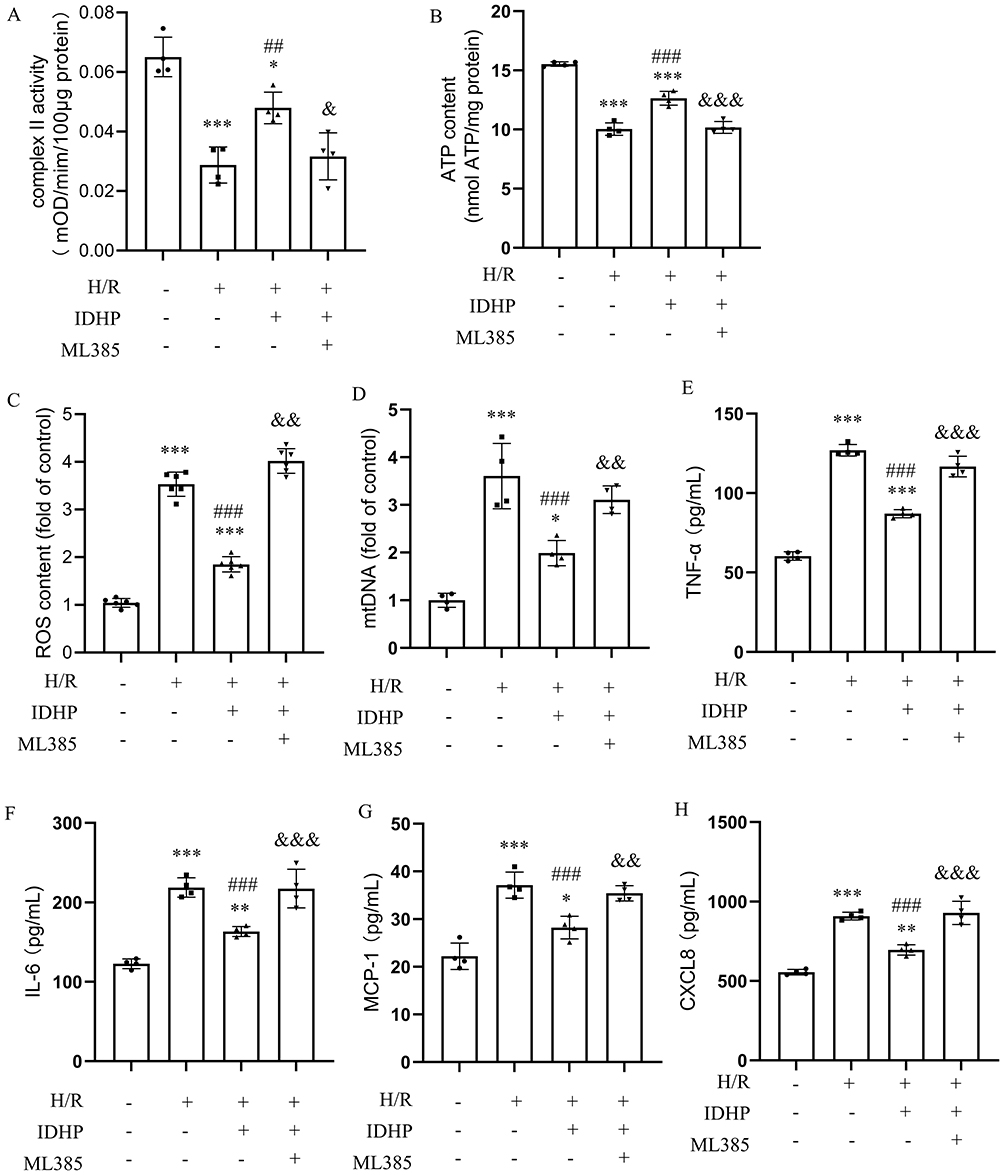 |
Figure 6 Pharmacologic inhibition of Nrf2 partially attenuates the protective effects of IDHP in H/R-treated HK-2 cells. (A) Mitochondrial complex II activity. (B) ATP content. (C) Intracellular ROS levels. (D) Cytosolic mtDNA levels. (E–H) Secretion of TNF-α, IL-6, MCP-1, and CXCL-8 measured by ELISA in control cells and cells treated with H/R, IDHP, and ML385 as indicated. Data are presented as mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001 vs. control group; ##p < 0.01, ### p< 0.001 vs. H/R group; &p < 0.05, &&p < 0.01, &&&p < 0.001 vs. H/R+IDHP group. |
Similarly, the IDHP-mediated reductions in TNF-α, IL-6, MCP-1, and CXCL-8 secretion were partially attenuated by ML385 (Figure 6E–H). These findings suggest that Nrf2-associated signaling may contribute, at least in part, to the anti-inflammatory and mitochondria-preserving effects of IDHP in H/R-injured tubular epithelial cells.
Discussion
This study shows that IDHP confers protection against experimental renal ischemia-reperfusion injury. In both animal and cell models, IDHP improved renal injury phenotypes and was associated with reduced inflammatory burden, lower oxidative stress, and better preservation of mitochondrial function. These findings support further preclinical evaluation of IDHP in ischemic kidney injury, while the mechanistic and translational implications should be interpreted cautiously.
Inflammation is a major amplifier of tissue injury after ischemia and reperfusion.25 Tubular epithelial cell injury initiates a cascade of danger signaling that rapidly recruits neutrophils and macrophages to the kidney. These cells release cytokines, chemokines, ROS, and proteases, thereby worsening local injury.26 In the present study, IDHP reduced both macrophage and neutrophil infiltration and suppressed renal TNF-α, IL-6, and IL-1β expression, while enhancing IL-10 expression. The stronger reduction observed in neutrophil accumulation is noteworthy because neutrophils are among the earliest inflammatory effectors to infiltrate the post-ischemic kidney and can intensify tissue injury through multiple mechanisms.27 The in vitro findings further support an anti-inflammatory profile for IDHP, as it reduced secretion of TNF-α, IL-6, MCP-1, and CXCL-8 from H/R-injured HK-2 cells. Together, these results suggest that suppression of inflammatory responses is an important component of IDHP-associated protection in renal IRI.
A second important finding is that IDHP preserved mitochondrial function. Renal tubular epithelial cells depend heavily on mitochondrial oxidative metabolism and are therefore particularly vulnerable to ischemic injury.28 In the current study, IDHP improved complex II and IV activities, increased ATP content, and reduced cytosolic mtDNA release in vivo. Similar protective effects on complex II activity, ATP production, ROS accumulation, and cytosolic mtDNA release were observed in HK-2 cells. Because cytosolic mtDNA can function as a danger-associated molecular pattern and activate innate inflammatory pathways,29,30 the reduced cytosolic mtDNA accumulation observed after IDHP treatment may be linked to the attenuated inflammatory response. However, because downstream mtDNA-sensing pathways were not directly examined, this finding should not be interpreted as direct causal evidence.
Oxidative stress links mitochondrial dysfunction to inflammatory amplification.31 IDHP lowered MDA and GSSG levels and increased t-SOD activity in injured kidneys, indicating an improved redox state. We further found that IDHP promoted Nrf2 nuclear translocation and increased expression of the downstream antioxidant genes HO-1 and GCLC. Nrf2 is increasingly recognized not only as an antioxidant transcription factor but also as a broader regulator of inflammation-associated tissue protection.13,32 In HK-2 cells, pharmacologic inhibition of Nrf2 with ML385 partially attenuated the effects of IDHP on mitochondrial function, ROS accumulation, cytosolic mtDNA accumulation, and inflammatory mediator production. These data suggest that Nrf2-associated signaling may participate, at least in part, in the protective actions of IDHP. Nevertheless, alternative interpretations should be considered. The partial reversal observed after ML385 treatment suggests that IDHP may also act through Nrf2-independent antioxidant or mitochondria-protective pathways. Moreover, because ML385 is a pharmacologic inhibitor, potential off-target effects cannot be fully excluded. It is also possible that the reduction in inflammatory mediator production resulted, at least in part, from improved mitochondrial preservation rather than representing the initiating protective mechanism. Therefore, the observed relationship among Nrf2-associated signaling, mitochondrial preservation, cytosolic mtDNA accumulation, and sterile inflammation should be regarded as a plausible mechanistic association rather than definitive in vivo causal evidence.
The upstream mechanism by which IDHP enhances Nrf2-associated signaling remains unclear. Potential mechanisms may involve Keap1 cysteine modification, GSK-3β inhibition, PKCδ-related signaling, or other redox-sensitive pathways, but these possibilities were not directly examined in this study. Likewise, whether Nrf2-associated signaling limits cytosolic mtDNA accumulation by preserving mitochondrial integrity remains to be determined. Future studies using in vivo Nrf2 loss-of-function models, together with assays of mitochondrial quality control and mtDNA-sensing pathways, will be needed to clarify this mechanism.
Our findings extend previous evidence that Danshen-related compounds can confer renal or ischemia-related cytoprotection through antioxidant and anti-inflammatory mechanisms.33–35 Although tanshinone IIA, salvianolic acid B, and cryptotanshinone have been evaluated in renal IRI,15–17 IDHP differs from these compounds as a Danshensu-related esterified metabolite with a distinct chemical scaffold and reported anti-inflammatory, antioxidant, and mitochondria-protective activities.19–22 This esterified structure may improve lipophilicity and metabolic stability, thereby facilitating cellular penetration, bioavailability, and intracellular or mitochondrial protective effects.18,19 The present findings also align with studies of classical Nrf2-activating agents, including sulforaphane, dimethyl fumarate, and bardoxolone methyl, in renal IRI or ischemic AKI contexts.36–38 However, because direct comparisons with these agents or with other Salvia miltiorrhiza-derived compounds were not performed, this study does not establish the superiority of IDHP. Instead, it supports IDHP as a distinct candidate for further preclinical evaluation in renal IRI, while emphasizing the need for careful safety assessment before translation.39
From a translational perspective, renal IRI is particularly relevant to kidney transplantation, where injury occurs in a clinically predictable setting. IDHP may therefore be worth exploring in future studies as a donor-preparation strategy, perioperative recipient therapy, organ-preservation additive, or early reperfusion intervention. Nevertheless, these possibilities remain speculative at present because pharmacokinetic, toxicological, dose-response, long-term outcome, and actual transplantation-model data are not yet available.
The present study has several limitations. First, only a single in vivo dose of IDHP was tested, and formal dose-response analysis was not performed; therefore, the optimal effective dose and dose-related safety profile remain to be defined. Second, because IDHP was administered before ischemia and shortly after reperfusion, the present findings mainly support a prophylactic or peri-reperfusion protective strategy rather than treatment of established renal injury, and future studies should define its therapeutic window. Third, the 24 h reperfusion period allowed assessment of acute injury and early inflammatory responses but did not address sustained functional recovery, long-term structural preservation, or delayed mitochondrial adaptation. In addition, only male mice were used. Fourth, Nrf2 involvement was supported mainly by in vitro pharmacologic inhibition, and downstream mtDNA-sensing pathways, such as cGAS-STING or TLR9 signaling, were not directly examined. Finally, tribromoethanol anesthesia and carprofen analgesia were applied consistently across groups according to animal welfare requirements; however, their potential influence on inflammatory readouts cannot be entirely excluded, particularly because carprofen is a nonsteroidal anti-inflammatory analgesic.
Despite these limitations, the data support further preclinical evaluation of IDHP as an inflammation-modulating and mitochondria-protective candidate in renal IRI. Its potential applications may include prophylactic or peri-reperfusion protection in predictable ischemic settings, but dose-response, therapeutic-window, long-term outcome, pharmacokinetic, toxicological, and in vivo mechanistic studies are needed before clinical translation can be considered.
Conclusion
IDHP attenuates experimental renal IRI and H/R-induced tubular epithelial injury, potentially through coordinated suppression of sterile inflammation, preservation of mitochondrial function, reduction of oxidative stress, and engagement of Nrf2-associated cytoprotective signaling. These findings support IDHP as a preclinical candidate renoprotective agent. Further dose-response, therapeutic-window, long-term outcome, pharmacokinetic, toxicological, and in vivo mechanistic studies are required before clinical translation can be considered.
Protocol Registration
No formal study protocol was prepared or registered before the study. The experimental design and analysis plan were established before data collection based on preliminary findings and published literature. This study adheres to the ARRIVE 2.0 guidelines.
Abbreviations
AKI, acute kidney injury; BUN, blood urea nitrogen; CXCL-8, C-X-C motif chemokine ligand 8; GCLC, glutamate-cysteine ligase catalytic subunit; GSSG, oxidized glutathione; H/R, hypoxia/reoxygenation; HO-1, heme oxygenase-1; IDHP, isopropyl 3-(3,4-dihydroxyphenyl)-2-hydroxypropanoate; IRI, ischemia-reperfusion injury; MCP-1, monocyte chemoattractant protein-1; MDA, malondialdehyde; mtDNA, mitochondrial DNA; Nrf2, nuclear factor erythroid 2-related factor 2; PAS, periodic acid-Schiff; ROS, reactive oxygen species; Scr, serum creatinine; t-SOD, total superoxide dismutase.
Data Sharing Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval
The animal study was approved by the Animal Care and Use Committee of Xi’an Jiaotong University (approval no. XJTU-IAUC/765/2019, #19765) and was performed in accordance with the Guidelines for the Care and Use of Laboratory Animals of Xi’an Jiaotong University, the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and the ARRIVE 2.0 guidelines.
Acknowledgments
The authors thank the members of their laboratories for technical assistance and helpful discussions.
Author Contributions
Xiaofei Yan: Conceptualization; Methodology; Investigation; Formal analysis; Data curation; Writing-original draft; Writing-review & editing.
Yuli Miao: Investigation; Validation; Data curation; Writing-review & editing.
Ying Zhang: Investigation; Validation; Data curation; Writing-review & editing.
Yixuan Li: Investigation; Validation; Data curation; Writing-review & editing.
Ziyi Wang: Investigation; Methodology; Validation; Writing-review & editing.
Zhaoqi Pu: Investigation; Methodology; Validation; Writing-review & editing.
Xiaohui Zheng: Resources; Conceptualization; Writing-review & editing.
Jin Zheng: Conceptualization; Methodology; Supervision; Project administration; Writing-review & editing; Funding acquisition.
All authors made substantial contributions to the conception and design of the study, acquisition of data, analysis and interpretation of data, drafting of the manuscript, or critical revision of the manuscript. All authors reviewed and approved the final version of the manuscript, agreed to submit the manuscript to the current journal, and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (82170768), the Postdoctoral Research Project of Shaanxi Province (2023BSHEDZZ04), and the Natural Science Basic Research Program of Shaanxi Province (2024JC-YBQN-0972 and 2024JC-YBQN-0826). The funders had no role in study design, data collection, data analysis, interpretation, or manuscript preparation.
Disclosure
The authors report no conflicts of interest in this article.
References
1. Nieuwenhuijs-Moeke GJ, Pischke SE, Berger SP, et al. Ischemia and reperfusion injury in kidney transplantation: relevant mechanisms in injury and repair. J Clin Med. 2020;9(1):253. doi:10.3390/jcm9010253
2. Zhao H, Alam A, Soo AP, et al. Ischemia-reperfusion injury reduces long-term renal graft survival: mechanism and beyond. EBioMedicine. 2018;28:31–16. doi:10.1016/j.ebiom.2018.01.025
3. Kim YH, Lee TK, Lee JC, et al. Risperidone administration attenuates renal ischemia and reperfusion injury following cardiac arrest by anti-inflammatory effects in rats. Vet Sci. 2023;10(3):184. doi:10.3390/vetsci10030184
4. Yan R, Zhang Y, Li Z, et al. Core signaling pathways in renal ischemia-reperfusion injury: network regulation and therapeutic prospects. Cell Signal. 2025;136:112185. doi:10.1016/j.cellsig.2025.112185
5. Zhou L, Tang S, Li F, et al. Ceria nanoparticles prophylaxis for renal ischemia-reperfusion injury by attenuating oxidative stress and inflammatory response. Biomaterials. 2022;287:121686. doi:10.1016/j.biomaterials.2022.121686
6. Riley JS, Tait SW. Mitochondrial DNA in inflammation and immunity. EMBO Rep. 2020;21(4):e49799. doi:10.15252/embr.201949799
7. Piantadosi CA. Mitochondrial DNA, oxidants, and innate immunity. Free Radic Biol Med. 2020;152:455–461. doi:10.1016/j.freeradbiomed.2020.01.013
8. Zhao M, Liu S, Wang C, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate mitochondrial damage and inflammation by stabilizing mitochondrial DNA. ACS Nano. 2021;15(1):1519–1538. doi:10.1021/acsnano.0c08947
9. Chen Y, Li Z, Zhang H, et al. Mitochondrial metabolism and targeted treatment strategies in ischemia-induced acute kidney injury. Cell Death Discov. 2024;10(1):69. doi:10.1038/s41420-024-01843-5
10. Vringer E, Tait SWG. Mitochondria and cell death-associated inflammation. Cell Death Differ. 2023;30(2):304–312. doi:10.1038/s41418-022-01094-w
11. Mei XD, Cao YF, Che YY, et al. Danshen: a phytochemical and pharmacological overview. Chin J Nat Med. 2019;17(1):59–80. doi:10.1016/S1875-5364(19)30010-X
12. Liu M, Grigoryev DN, Crow MT, et al. Transcription factor Nrf2 is protective during ischemic and nephrotoxic acute kidney injury. Kidney Int. 2009;76(3):277–285. doi:10.1038/ki.2009.157
13. Ahmed SMU, Luo L, Namani A, Wang XJ, Tang X. Nrf2 signaling pathway: pivotal roles in inflammation. Biochim Biophys Acta Mol Basis Dis. 2017;1863(2):585–597. doi:10.1016/j.bbadis.2016.11.005
14. Dinkova-Kostova AT, Abramov AY. The emerging role of Nrf2 in mitochondrial function. Free Radic Biol Med. 2015;88(Pt B):179–188. doi:10.1016/j.freeradbiomed.2015.04.036
15. Xu YM, Ding GH, Huang J, et al. Tanshinone IIA pretreatment attenuates ischemia/reperfusion-induced renal injury. Exp Ther Med. 2016;12(4):2741–2746. doi:10.3892/etm.2016.3674
16. Pang Y, Zhang PC, Lu RR, et al. Salvianolic acid B modulates caspase-1-mediated pyroptosis in renal ischemia-reperfusion injury via the Nrf2 pathway. Front Pharmacol. 2020;11:541426. doi:10.3389/fphar.2020.541426
17. Bai T, Yang K, Qin C, et al. Cryptotanshinone ameliorates renal ischaemia-reperfusion injury by inhibiting apoptosis and inflammatory response. Basic Clin Pharmacol Toxicol. 2019;125(5):420–429. doi:10.1111/bcpt.13275
18. Zheng X, Zhang X, Zhang X, et al. Determination of the main bioactive metabolites of Radix salvia miltiorrhizae in compound Danshen dripping pills and the tissue distribution of Danshensu in rabbit by SPE-HPLC-MSn. J Sep Sci. 2007;30(6):851–857. doi:10.1002/jssc.200600287
19. Lei W, Xu X, Li N, et al. Isopropyl 3-(3,4-dihydroxyphenyl)-2-hydroxypropanoate protects septic myocardial injury via regulating the GAS6/Axl-AMPK signaling pathway. Biochem Pharmacol. 2024;221:116035. doi:10.1016/j.bcp.2024.116035
20. Chen J, Wang Y, Li X, et al. IDHP mitigates LPS-induced cardiomyocyte injury via the GAS6/Axl-AMPK axis: a multi-target strategy counteracting inflammation, oxidative stress, and apoptosis. Pharmaceuticals. 2025;18(8):1188. doi:10.3390/ph18081188
21. Wang S, Wang H, Jing H, et al. Anti-inflammatory effects of isopropyl 3-(3,4-dihydroxyphenyl)-2-hydroxypropanoate, a novel metabolite from Danshen, on activated microglia. Chin J Physiol. 2012;55(6):428–434. doi:10.4077/CJP.2012.BAA045
22. Zhang ML, Wang M, Chen J, et al. Isopropyl 3-(3,4-dihydroxyphenyl)-2-hydroxypropanoate protects lipopolysaccharide-induced acute lung injury in mice by attenuating pyroptosis. Eur J Pharmacol. 2023;942:175545. doi:10.1016/j.ejphar.2023.175545
23. Tavares MB, Chagas de Almeida MC, Martins RT, et al. Acute tubular necrosis and renal failure in patients with glomerular disease. Ren Fail. 2012;34:1252–1257. doi:10.3109/0886022X.2012.723582
24. Zheng J, Lan P, Li M, et al. Anti-Na+/K+-ATPase DR antibody attenuates UUO-induced renal fibrosis through inhibition of Na+/K+-ATPase α1-dependent HMGB1 release. Int Immunopharmacol. 2023;116:109826. doi:10.1016/j.intimp.2023.109826
25. Algoet M, Janssens S, Himmelreich U, et al. Myocardial ischemia-reperfusion injury and the influence of inflammation. Trends Cardiovasc Med. 2023;33(6):357–366. doi:10.1016/j.tcm.2022.02.005
26. Daemen MA, de Vries B, Van’t Veer C, et al. Apoptosis and chemokine induction after renal ischemia-reperfusion. Transplantation. 2001;71(7):1007–1011. doi:10.1097/00007890-200104150-00032
27. Heinzelmann M, Mercer-Jones MA, Passmore JC. Neutrophils and renal failure. Am J Kidney Dis. 1999;34(2):384–399. doi:10.1016/S0272-6386(99)70375-6
28. Bhargava P, Schnellmann RG. Mitochondrial energetics in the kidney. Nat Rev Nephrol. 2017;13:629–646. doi:10.1038/nrneph.2017.107
29. West AP, Khoury-Hanold W, Staron M, et al. Mitochondrial DNA stress primes the antiviral innate immune response. Nature. 2015;520(7548):553–557. doi:10.1038/nature14156
30. McArthur K, Whitehead LW, Heddleston JM, et al. BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science. 2018;359(6378):eaao6047. doi:10.1126/science.aao6047
31. Xu X, Pang Y, Fan X. Mitochondria in oxidative stress, inflammation and aging: from mechanisms to therapeutic advances. Signal Transduct Target Ther. 2025;10(1):190. doi:10.1038/s41392-025-02253-4
32. Zhang DD. Thirty years of NRF2: advances and therapeutic challenges. Nat Rev Drug Discov. 2025;24(6):421–444. doi:10.1038/s41573-025-01145-0
33. Chen Y, Wei Q, Zhang M, et al. Danshensu attenuates cisplatin-induced acute kidney injury through anti-oxidation and anti-apoptosis in vivo and in vitro. Cell Physiol Biochem. 2013;31(1):79–90. doi:10.1159/000343351
34. Dai X, Huo J, Wang X, et al. Salvianolic acid B and Danshensu activate the Nrf2 antioxidant pathway to alleviate cerebral ischemia/reperfusion injury. Phytomedicine. 2021;91:153675. doi:10.1016/j.phymed.2021.153675
35. Xu Z, Cai K, Su SL, et al. Salvianolic acid B and tanshinone IIA synergistically improve early diabetic nephropathy through regulating PI3K/Akt/NF-kappaB signaling pathway. J Ethnopharmacol. 2024;319(Pt 3):117356. doi:10.1016/j.jep.2023.117356
36. Yoon HY, Kang NI, Lee HK, Jang KY, Park JW, Park BH. Sulforaphane protects kidneys against ischemia-reperfusion injury through induction of the Nrf2-dependent Phase 2 enzyme. Biochem Pharmacol. 2008;75(11):2214–2223. doi:10.1016/j.bcp.2008.02.029
37. Zhen X, Li J, Zhou Y, et al. Activation of Nrf2 pathway by dimethyl fumarate attenuates renal ischemia-reperfusion injury. Transplant Proc. 2021;53(7):2133–2139. doi:10.1016/j.transproceed.2021.07.017
38. Wu QQ, Wang Y, Senitko M, et al. Bardoxolone methyl (BARD) ameliorates ischemic AKI and increases expression of protective genes Nrf2, PPARγ, and HO-1. Am J Physiol Renal Physiol. 2011;300(5):F1180–F1192. doi:10.1152/ajprenal.00353.2010
39. Chin MP, Wrolstad D, Bakris GL, et al. Risk factors for heart failure in patients with type 2 diabetes mellitus and stage 4 chronic kidney disease treated with bardoxolone methyl. J Card Fail. 2014;20(12):953–958. doi:10.1016/j.cardfail.2014.10.001
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Effects and Pathogenesis of PM2.5 and Its Components on Chronic Obstructive Pulmonary Disease

Wang Q, Liu S
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:493-506
Published Date: 6 April 2023
Dental Pulp Stem Cells Ameliorate Elastase-Induced Pulmonary Emphysema by Regulating Inflammation and Oxidative Stress
Gao X, Liu Z, Wang Z
Journal of Inflammation Research 2023, 16:1497-1508
Published Date: 8 April 2023
Quercetin: A Flavonoid with Potential for Treating Acute Lung Injury

Huang M, Liu X, Ren Y, Huang Q, Shi Y, Yuan P, Chen M
Drug Design, Development and Therapy 2024, 18:5709-5728
Published Date: 6 December 2024
Exercise Prescription Training in Chronic Obstructive Pulmonary Disease: Benefits and Mechanisms
Liu S, Yang A, Yu Y, Xu B, Yu G, Wang H
International Journal of Chronic Obstructive Pulmonary Disease 2025, 20:1071-1082
Published Date: 15 April 2025
The Nrf Family and Its Cardioprotective Potential: Mechanisms, Functions, and Therapeutic Perspectives
Cui C, Song H, Guo Y, Shi J, Geng B, Wang G
Drug Design, Development and Therapy 2025, 19:8339-8373
Published Date: 15 September 2025