Back to Journals » ImmunoTargets and Therapy » Volume 15
Irinotecan Modulates Immune Checkpoints in Neuroblastoma
Authors Lapidus T ![]() , Zorde-Khvalevsky E, Jolani A, Goldstein G, Peled A
, Zorde-Khvalevsky E, Jolani A, Goldstein G, Peled A
Received 25 January 2026
Accepted for publication 12 May 2026
Published 12 June 2026 Volume 2026:15 598469
DOI https://doi.org/10.2147/ITT.S598469
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Michael Shurin
Tom Lapidus,1 Elina Zorde-Khvalevsky,1 Alaa Jolani,1 Gal Goldstein,2 Amnon Peled1
1Goldyne Savad Institute of Gene Therapy, Hadassah-Hebrew University Medical Center, Jerusalem, Israel; 2The Pediatric Hemato-Oncology Department, Hadassah- Hebrew University Medical Center, Jerusalem, Israel
Correspondence: Tom Lapidus, Goldyne Savad Institute of Gene Therapy, Hadassah-Hebrew university Medical Center, Jerusalem, Hebrew University Medical School, Jerusalem, Israel, Email [email protected]
Background: Neuroblastoma (NBL) is the most common extracranial solid tumor in children, with a 50% survival rate in high-risk cases. Chemotherapy combination with immunotherapy is central to HR-NBL treatment.
Aim: This study aimed to explore how Irinotecan influences the immune landscape of NBL cells, particularly focusing on the modulation of immune checkpoints relevant to future chemo-immunotherapy strategies.
Methods: Two neuroblastoma cell lines, SK-N-BE(2) and SH-SY5Y, were exposed to sub-lethal doses of Irinotecan and IFN-γ. Flow cytometry assessed the expression of immune markers, including GD2, CD47, MHC I, and PD-L1. We also examined phagocytosis by macrophages and T-cell-mediated killing. In vivo studies using a neuroblastoma mouse model assessed the effects of Irinotecan and PBMC injections. Soluble CD47 was also measured in cell culture supernatants and exploratory patient plasma samples.
Results: Treatment exposure was associated with modulation of several immune-related markers, including GD2, CD47, PD-L1, and MHC class I. While Irinotecan reduced macrophage-mediated phagocytosis, this effect was reversed by the addition of anti-CD47 antibodies. T-cell-mediated killing was enhanced when NBL cells were pre-treated with Irinotecan. In vivo studies showed that the combination of PBMCs and Irinotecan resulted in reduced tumor burden compared to either treatment alone. Furthermore, CD47 expression was detectable in-patient plasma, suggesting that soluble CD47 may warrant exploration in larger future cohorts.
Conclusion: This pilot study suggests that irinotecan may modulate key immune checkpoints in neuroblastoma. These results supports further investigation of rational chemo-immunotherapy combinations, including GD2 and CD47-targeted combination strategies.
Keywords: neuroblastoma, chemotherapy, immunotherapy, irinotecan, CD47
Introduction
Despite advancements in pediatric cancer care, malignant neoplasms remain the third leading cause of pediatric death in the US. Immunotherapy is still a relatively minor component of the standard treatment for pediatric solid tumors, this is due to various challenges these therapies face in addressing the complexities of pediatric solid tumors such as low tumor mutational burden, reduced neoantigen expression, and an immunosuppressive tumor microenvironment.
Neuroblastoma (NBL) is the most common extracranial solid tumor of childhood and accounts for approximately 15% of pediatric cancer mortality. Survival for patients with high-risk neuroblastoma remains poor with 5 year overall survival rates of around 50% despite multimodal therapy.1
Current treatment of high-risk neuroblastoma includes intensive chemotherapy, surgery, radiotherapy, autologous stem cell transplantation, differentiation therapy, and anti-GD2 immunotherapy, which has become a mainstay of frontline treatment. Nevertheless, relapse and refractory disease remain major clinical challenges, highlighting the need for improved therapeutic strategies.
Increasing evidence suggests that conventional chemotherapy may influence anti-tumor immunity in addition to its direct cytotoxic effects. These immunomodulatory effects may alter antigen presentation, immune checkpoint expression, and susceptibility to immune-mediated killing, thereby affecting the activity of combination chemo-immunotherapy approaches.
Although we are witnessing a renaissance of new therapies and targets being searched for NBL, chemotherapy remains the cornerstone of both standard induction and post induction treatment for HR-NBL.
Neuroblastoma is considered an immunological “cold” cancer, characterized by a tumor microenvironment (TME) with limited activated immune cell infiltration, making it difficult for the immune system to target.2 Notably, the use of checkpoint inhibitors, which have shown remarkable success in some adult cancers, has not been effective in NBL.3
Several studies have examined the impact of chemotherapies on the response to immunotherapy across various cancers.4 However, there is still much to learn about the specific effects of chemotherapies on surviving cancer cells. In particular, ongoing clinical studies are investigating chemo-immunotherapy combinations in NBL.5
We chose to investigate irinotecan, a topoisomerase I inhibitor widely used in relapsed or refractory neuroblastoma and in combination regimens with immunotherapy. Beyond its cytotoxic activity, prior studies in other tumor models have suggested that irinotecan may modulate the tumor immune environment, including effects on regulatory T cells, MHC class I expression, and PD-L1 signaling.6
However, whether similar immunologic effects occur in neuroblastoma remains insufficiently characterized.
GD2 is a disialoganglioside highly expressed in NBL,7 and is the target of the first approved immunotherapy against NBL, Dinutuximab. This treatment has improved survival in the HR group,8 and several clinical studies are underway to determine the optimal combination of anti-GD2 therapy with chemotherapy. Unfortunately, a significant number of patients relapse despite anti-GD2 treatment,9 and side effects such as severe visceral pain remain important dose-limiting toxicities.10,11
CD47 is a transmembrane immune checkpoint molecule frequently overexpressed by malignant cells. Through binding to signal regulatory protein alpha (SIRPα) on macrophages and other myeloid cells,12–14 CD47 delivers a “don’t eat me” signal that suppresses phagocytosis and contributes to immune escape.
The CD47–SIRPα axis has emerged as a promising therapeutic target in multiple cancers, including pediatric malignancies. In neuroblastoma, preclinical studies have demonstrated anti-tumor activity with CD47 blockade, particularly when combined with anti-GD2 therapy or other immune-based approaches.15,16
In this pilot exploratory study, we investigated immune-related changes following exposure of neuroblastoma cells to sub-lethal irinotecan. We then examined potential functional consequences in macrophage phagocytosis, T cell–mediated killing, a xenogeneic in vivo model, and exploratory patient-derived CD47 measurements. Our goal was to generate hypothesis-forming data regarding how chemotherapy may influence immune targeting strategies in neuroblastoma.
Materials and Methods
Cell Lines and Cell Culture Conditions
SK-N-BE(2) (CRL-2271, RRID:CVCL_0528) and SH-SY5Y (CRL-2266, RRID:CVCL_0019) cell lines were obtained from American Type Culture Collection (ATCC). Both cell lines were cultured in DMEM high glucose medium, supplemented with 10% (vol/vol) heat-inactivated FCS, 2 mM glutamine, 100 units/mL penicillin, and 100 µg/mL Peniciline/Streptomycin. The cell cultures were maintained in a humidified atmosphere of 5% (vol/vol) CO2 at 37 °C. GFP and Luciferase co-expressing clones were prepared by transfection of pYOE-LV-luc-GFP plasmid (Ubigene) and followed by sorting of GFP-expressing cells. Irinotecan (Irinotecan HCl, Pfizer) was added directly to the media at noted concentrations. IFN γ (Recombinant Human IFN-γ, cat# 300–02, PeproTech) was added directly to the media, 20ng/mL. Cell growth follow up was done using Incucyte machine and cell confluence analysis software.
Flow Cytometry and Cell Sorting
Cells were detached and mixed via pipetting to create a single-cell solution. For flow cytometry, cells were analysed for fluorescence using a CytoFLEX S flow cytometer (Beckman Coulter) and analyzed using CytExpert program.
aPD-L1 APC anti-human CD274 (B7-H1, PD-L1), (BioLegend Cat# 329708, RRID:AB_940360)
MHC I, PE anti-human HLA-A,B,C, (BioLegend Cat# 311406, RRID:AB_314875)
GD2, PE anti-human Ganglioside GD2, (BioLegend Cat# 357303, RRID:AB_2561884)
CD47, APC anti-human CD47, (BioLegend Cat# 323124, RRID:AB_2716203)
CD11b, APC anti-human CD11b, (BioLegend Cat# 301310, RRID:AB_314162)
CD45, FITC anti-human CD45, (BioLegend Cat# 368508, RRID:AB_2566368)
CD3, APC anti-human CD3, (BioLegend Cat# 344812, RRID:AB_10645473)
CD8, PE anti-human CD8, (BioLegend Cat# 300908, RRID:AB_314112)
All antibodies were used at 1:100 dilution.
For sorting, cells were sorted using a SH800S cell sorter (Sony) using GFP as a marker for positive cells.
The results shown are representative experiment of results, with another two replicates performed with similar results.
Macrophage-Mediated Phagocytosis
Normal human PBMCs were purified from buffy coat by Ficoll (Histopaque-1077, Cat# 10771, Sigma Aldrich), according to the manufacturer’s instructions. Buffy coat was obtained from blood Bank in Hadassah-Hebrew university hospital, Institutional review board (IRB) approval number HMO-21-0553. Purified PBMCs were seeded in RPMI 1640 media (cat# 21875034, Gibco) without supplements for 50 minutes, 60 million cells/10-cm dish. Then the non-adherent cells were washed out and cells were incubated in media supplemented with 10% Fetal Bovine Serum (cat# 12657–029, Gibco), Penicillin / Streptomycin (cat# 15140–122, Gibco), L-Glutamine 200mM (cat# 25030–024, Gibco), Sodium Pyruvate Solution 100mM (cat# 03–042-1B, Biological Industries), MEM Non-Essential Amino Acids Solution (cat# 11140050, GIBCO) in addition of 50ng/mL GM-CSF (cat# 300–03-20, PeproTech). Media was changed every 3 days. Macrophages were defined by FACS using CD11b as a marker. Phagocytosis assay was performed on day 6 of the Macrophage differentiation. For the assay, GFP-expressing NBL cells were pre-treated with sub-lethal doses of Irinotecan, 1µM, for 24 hours. NBL cells and Macrophages were co-cultured and noted E:T ratios for 4–6 hours. Anti-CD47 Antibody (InVivoMab anti-human CD47, (Bio X Cell Cat# BE0019-1, RRID:AB 1107655)) was added directly to the mixture at 10µg/mL concentration. For analysis of phagocytosis using flow cytometry, cells were stained with CD11b-APC antibody to detect Macrophages. Phagocytic Macrophages were defined as CD11b+GFP+ cells. Phagocytic efficacy was defined and Mean Fluorescent Intensity (MFI) of the FITC channel of phagocytic Macrophages. The results shown are representative experiment of results, with another replicate performed with similar results.
Cytotoxicity Assay (Killing)
Normal human PBMCs were purified from buffy coat by Ficoll (Histopaque-1077, Cat# 10771, Sigma Aldrich), according to the manufacturer’s instructions. For T cell enrichment, purified PBMCs were seeded in RPMI 1640 media (cat# 21875034, Gibco) supplemented with 10% Fetal Bovine Serum (cat# 12657–029, Gibco), Penicillin / Streptomycin (cat# 15140–122, Gibco), L-Glutamine 200mM (cat# 25030–024, Gibco), Sodium Pyruvate Solution 100mM (cat# 03–042-1B, Biological Industries), MEM Non-Essential Amino Acids Solution (cat# 11140050, GIBCO), 2-mercaptoethanol 50mM (cat: 31350–010 Gibco), in addition of 30 ng/mL Anti-human CD3/CD28 (ImmunoCult™ Human CD3/CD28 T Cell Activator, (STEMCELL Technologies Cat# 10971, RRID:AB_2827806)) and 1000 IU/mL Recombinant Human IL-2 (Recombinant Human IL-2, cat# 202-IL-500, R&D systems). PBMCs were seeded at concentration of 1 million PBMCs per 1 mL media, for 4 days. For the assay, GFP-expressing NBL cells were pre-treated with Irinotecan for 72 hours. NBL cells and T-cells were co-cultured and noted E:T ratios for over-night. For analysis of dying NBK cells using flow cytometry, cells were stained with 20ng/mL PI (Propidium Iodide, cat# P4864, Sigma Aldrich). Dying NBL cells were defined as GFP and PI positive. The results shown are representative experiment of results, with another replicate performed with similar results.
ELISA
CD47 protein levels of cell supernatants or tumor lysates were determined using sandwich-type ELISA commercially available kit according to the manufacturer’s protocol (R&D Systems, catalog #DY4670-05). The absorbance was read at 450 nm. Institutional review board (IRB) approval number 0495–23-HMO was granted prior to studying patients plasma according to the Hadassah-Hebrew university hospital ethics committee.
Immunohistochemistry (IHC)
Neuroblastoma tissue array (NB642c) representing 27 cases of neuroblastoma and 5 peripheral nerve tissue was purchased through TissueArray.com.
For immunohistochemical staining the slide subjected to heat induced epitope retrieval (HIER) in Tris-EDTA buffer TE (pH 8.0). After HIER the slide was incubated over-night at 4°C with anti-CD47 (Abcam Cat# ab226837, RRID:AB_3674726) diluted 1:100 in CAS-Block solution (cat#008120, Invitrogen). Peroxidase labeled anti-rabbit polymeric reagent was used for the detection of the section bound primary antibody. Sections processed without the primary antibody served as a negative control.
In vivo
All mice were kept in a specific pathogen-free facility. Mice were handled according to the criteria outlined in the Guide for the Care and Use of Laboratory Animals prepared by the National Academy of Sciences and published by the National Institutes of Health. Mice had free access to water and chow at all times. NSG mice couple was purchased from the Jackson Laboratory and bred in the animal unit in the animal facility. Maintenance of mice and all experimental procedures were approved by and performed in accordance with the Institutional Animal Care and Use Committee-approved animal treatment protocols (NIH approval number: OPRR-A01-5011). Approval for the animal experiments were given by the Animal Care Committee of Hebrew University, permit number MD-23-17,149-5. All animal experiments performed in this study were conducted in accordance with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines, and the study adheres to the ARRIVE checklist to ensure transparent and rigorous reporting of animal research.
NBL IP Model
Xenograft models of human neuroblastoma cancer were generated by intraperitoneal (IP) implantation of 2 × 106 SH-SY5Y-Luc cells in male NSG mice aged between 6 to 7 weeks.
9 days after the tumor induction, mice were exposed to total-body gamma irradiation at a dose of 2 Gray; luciferase expression was assessed using in vivo imaging system (IVIS) spectrum imaging (Perkin Elmer, Cat# 124262). For Luciferase expression measurement, D-luciferin (cat# E1605, Promega) at 150mg/kg was injected Intraperitoneally (IP) to mice 10 min prior IVIS spectrum imaging. Based on the image data, mice were divided into four similar treatment cohorts: untreated (3 mice); treated with PBMCs: 2milion PBMCs per mouse injected IP, 5 mice per group; treated with Irinotecan: 400ug/mouse, injected IV, 5 mice per group; or treated with combination of PBMCs with Irinotecan, 5 mice per group. On day 10 mice were treated according to the treatment groups. Tumor growth follow up was done by luciferase expression according to the IVIS imaging.
In vivo Sample Treatment – Spleen Isolation and FACS
To define human cells in the spleens of treated mice, mice were anesthetized and killed, spleens were weighted and then smashed via 70um cell strainer. Red blood cells were removed using ACK Lysis buffer (cat# A10492-01, Gibco). Cells from each spleen were dissolved in 1mL PBS, then 100ul of cell solution was taken for each of noted staining for FACS analysis.
Statistical Analysis
All data were subjected to statistical analysis using the Excel software package (Microsoft). Statistical analysis was performed using a two-way multivariate ANOVA to assess differences between treatment groups and experimental conditions.
Differences were considered significant at P < 0.05. Data are given as mean ± SEM.
Results
Neuroblastoma Cells Exhibit Immunologic Modulation Following Exposure to Irinotecan and IFN-γ
Our objective was to characterize the immunologic modulation of neuroblastoma (NBL) cells following exposure to the chemotherapeutic agent Irinotecan. In this study we used two NBL cell-lines, SK-N-BE(2) and SH-SY5Y, which are established neuroblastoma models with distinct phenotypic features and GD2 expression.
To mimic the behavior of NBL cells that survive chemotherapy in patients, we defined a sub-lethal dose of Irinotecan. To define sub-lethal doses, SK-N-BE(2) and SH-SY5Y cells were treated with escalating doses of Irinotecan. Cell growth was assessed using the Incucyte imaging system and cell confluence analysis software (Figure 1A and B). Sub-lethal doses were defined as doses that preserve at least 70% cell viability after a 40-hour incubation period. Based on these data, we selected 1 µM Irinotecan as a sub-lethal dose for both cell lines.
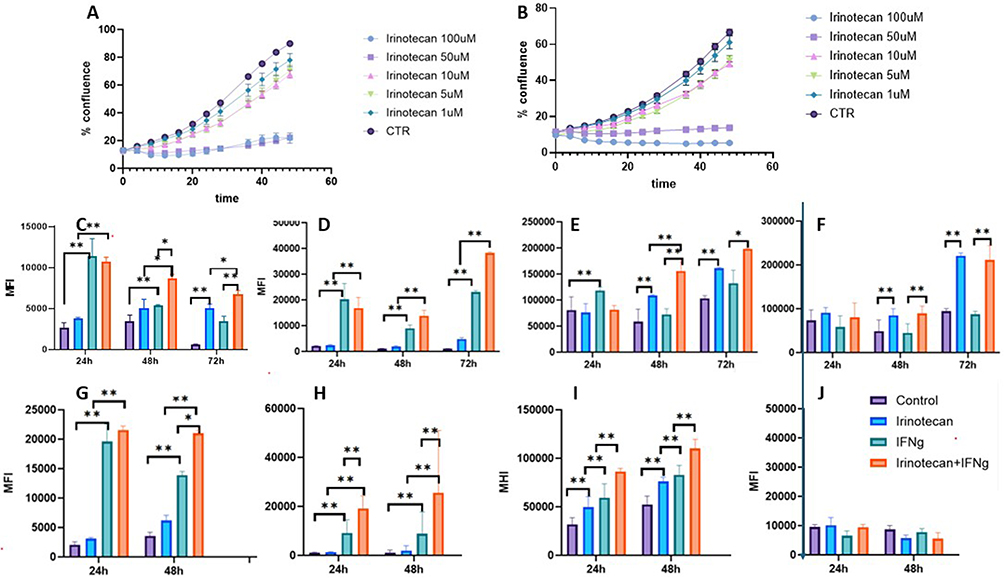 |
Figure 1 Irinotecan and IFN-ɣ modulate immune checkpoint expression in neuroblastoma cells. Determination of sub-lethal irinotecan concentrations (A and B). SK-N-BE(2) (A) and SH-SY5Y (B) cells were treated with increasing concentrations of irinotecan, and cell growth was monitored using the IncuCyte live-cell imaging system. Sub-lethal dose was defined as ≥70% viability after 48 hours. Surface expression of immune-related markers following treatment (C-J). SK-N-BE(2) (C–F) or SH-SY5Y (G–J) cells were treated with irinotecan (1 µM), IFN-γ (20 ng/mL), or their combination for the indicated time points. Expression of PD-L1 (C,G), MHC class I (D,H), CD47 (E,I), and GD2 (F and J) was assessed by flow cytometry and presented as mean fluorescence intensity (MFI). Treatment groups: control (purple), irinotecan (blue), IFN-γ (green), irinotecan + IFN-γ (Orange). Data are presented as mean ± SEM. *p < 0.05; **p < 0.01. |
Initially, we examined selected immune checkpoints and modulators implicated in NBL immune escape. Given the multitude of immune checkpoints and other interacting molecules within the tumor microenvironment (TME) that operate through receptor–ligand mechanisms, we focused on several targets with therapeutic relevance. The change in the expression of immune markers was assessed by FACS over time.
To evaluate treatment-associated changes in immune marker expression, we compared untreated cells with cells exposed to irinotecan, IFN-γ, or both conditions. IFN-γ was included as a biologically relevant immune stimulus and comparator because of its known effects on antigen presentation and checkpoint regulation. Marker expression was assessed by flow cytometry over time and is presented as mean fluorescence intensity (MFI) (Figure 1C–J).
Assessment of GD2, surface antigen expressed on NBL cells, showed an abundant expression in SK-N-BE(2), but not in SH-SY5Y cells (Figure 1F and J). Irinotecan further increased GD2 expression in SK-N-BE(2) cells. The effect was significantly more pronounced after 72 hours. An addition of IFN-γ had no effect. A possible loss of GD2 expression in SH-SY5Y cells may suggest a phenotypic transformation.
The initial PD-L1 levels were relatively low in both tested cell lines. Under the tested conditions, irinotecan and IFN-γ were both associated with time-dependent changes in PD-L1 expression, in SH-SY5Y the effect of IFN-γ was more pronounced. Combination treatment with Irinotecan and IFN-γ was associated with higher expression in PD-L1 expression in both cell lines (Figure 1C and G). In SK-N-BE(2) cells, basal PD-L1 expression transiently increased at 24 and 48 hours, but declined by 72 hours, whereas irinotecan and IFN-γ treatment preserved higher PD-L1 levels at this later time point (Figure 1C).
Assessment of the “don’t eat me” CD47 protein (Figure 1E and I) demonstrated high initial levels. Similar to the effect on PD-L1, Irinotecan or IFN-γ alone increased CD47 expression, while the combination of both further enhanced CD47 levels in both cell lines.
Measurement of MHC I expression revealed that untreated cells expressed relatively low levels of MHC I. Irinotecan alone led to a slight increase, while IFN-γ caused a marked elevation in MHC I levels. Interestingly, the combination of Irinotecan and IFN-γ further increased MHC I expression compared with untreated cells under the tested conditions. (Figure 1D and H).
Irinotecan Disturbs Phagocytosis, an Effect Which Might Be Reversed Through the Adjunctive Application of Anti-CD47 Antibodies
Upon observing the overexpression of CD47 on NBL cells, which undergoes further upregulation upon exposure to Irinotecan, and considering the established role of the CD47- SIRPα axis in mediating “don’t eat me” signals to immune cells, particularly Macrophages, we investigated its influence on phagocytic activity.
We conducted an analysis of Macrophage activity against neuroblastoma (NBL) cells utilizing a phagocytosis assay. This process involved the incubation of Macrophages with GFP-positive NBL cells. Phagocytic Macrophages were defined as Macrophages that engulfed cancer NBL-GFP positive cells and therefore GFP signal was detected inside those Macrophages. Phagocytic efficacy was assessed by FACS, were cancer NBL-GFP cells were defined as FITC-positive; Macrophages were defined by CD11b (APC) staining and phagocytic Macrophages were selected as CD11b(APC)+GFP(FITC)+ population.
We first assessed the effect of sub-lethal concentrations of Irinotecan on phagocytosis of NBL cells. We compared the percentage of phagocytic Macrophages in untreated versus Irinotecan-treated combination. The percentage of phagocytic Macrophages in the Macrophage-untreated NBL cells was defined as 100% (Figure 2A). Contrary to expectations that chemotherapy-treated cells would be more susceptible to phagocytosis, we observed a decrease in macrophage activity following treatment of NBL cell lines with sub-lethal doses of Irinotecan (Figure 2A). This phenomenon suggests that NBL cells, having been exposed to therapeutic interventions and survived, may acquire mechanisms to further evade immune detection. One possible escape route is through the upregulation of CD47 expression, as observed previously (Figure 1E–I).
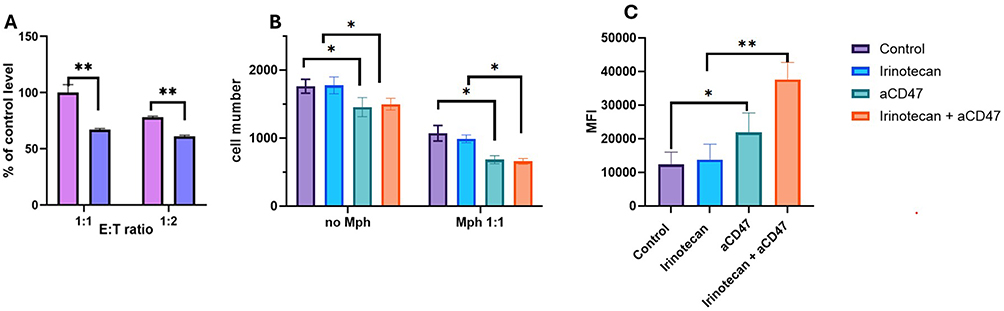 |
Figure 2 Irinotecan reduces macrophage-mediated phagocytosis, which is restored by CD47 blockade. GFP-expressing neuroblastoma cells (SK-N-BE(2) or SH-SY5Y) were pre-treated with irinotecan (1 µM) for 24 hours and co-cultured with human macrophages derived from PBMCs at a 1:1 ratio. Anti-CD47 antibody (αCD47, 10 µg/mL) was added where indicated. (A) Phagocytosis efficiency, defined as the percentage of CD11b⁺GFP⁺ macrophages, normalized to untreated control (set as 100%). (B) Quantification of live tumor cells, defined as GFP⁺ viable cells by flow cytometry. (C) Phagocytic activity, assessed as GFP mean fluorescence intensity (MFI) within CD11b⁺GFP⁺ macrophages. Treatment groups: control (purple), irinotecan (blue), αCD47 (green), irinotecan + αCD47 (Orange). Data are presented as mean ± SEM.*p < 0.05; **p < 0.01. |
CD47, also known as do not-eat-me signal was shown to inhibit phagocytosis, Based on this result, we extended our investigation to evaluate the effects of anti-CD47 (αCD47) on phagocytosis (Figure 2B and C). Interestingly, incubation of NBL cells with αCD47 alone led to a decrease in the number of live NBL cells. The combination of macrophages, NBL cells, and αCD47 resulted in a significant reduction in the number of viable cancer cells, and addition of Irinotecan did not reverse this effect.
We then assessed macrophage phagocytic efficacy by measuring the average GFP intensity in phagocytic macrophages (Figure 2C). While αCD47 alone significantly elevated macrophage phagocytic efficacy, the combination of αCD47 with sub-lethal doses of Irinotecan showed an additive effect. Our results revealed that not only does αCD47 alone enhance phagocytosis of NBL cells, but its combination with Irinotecan treatment results in the most pronounced phagocytic activity (Figure 2B and C).
Combination of Activated T Cells with Irinotecan Improves Killing of Neuroblastoma Cells
Subsequently, our aim was to characterize the effect of sub-lethal doses of Irinotecan on T-cell mediated cytotoxicity against NBL cells.
We conducted a killing assay, an immunological test to examine specifically T-cell killing of NBL cells (SK-N-BE(2)) (Figure 3). NBL cells were pre-treated with 1 µM Irinotecan or left untreated.We found that seeding activated T cells alone with NBL cells did not result in meaningful cancer cell killing. However, pre-incubation of NBL cells with sub-lethal doses of Irinotecan induced a significant T cell–mediated cancer cell killing (Figure 3).
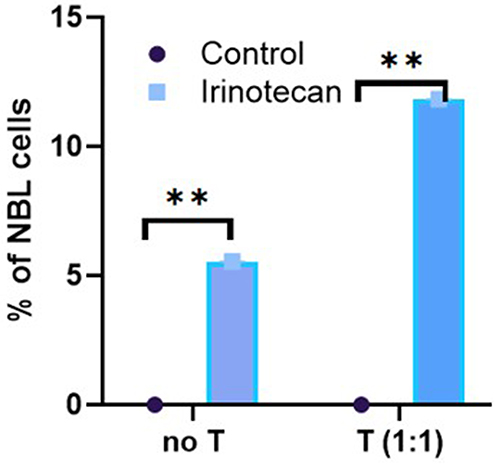 |
Figure 3 Irinotecan pre-treatment enhances T cell–mediated cytotoxicity against neuroblastoma cells. Activated T cells were induced from human PBMCs by CD3 and CD28 co-stimulation. NBL cells, SK-N-BE(2) and SH-SY5Y, were pre-treated with 1µM of Irinotecan for 24hrs. NBL and T cells were co-cultured for 24hrs. Live NBL cells were assessed by FACS. Data are presented as the change in the percentage of tumor cell death compared to untreated control cells. Representative data from SK-N-BE(2) cells are shown. Treatment groups: Control (dot), T cells (blue). Data are shown as mean ± SEM. **p < 0.01. |
The combination of irinotecan pre-treatment and activated T cells produced the greatest tumor cell killing under the tested conditions. This phenomenon may be explained by the fact that IFN γ, which is secreted by activated T cells,16 especially in combination with sub-lethal doses of Irinotecan, led to a significant elevation of MHC class 1 molecule on the cells (Figure 1D and H), which in turn induced cytotoxic effect. However, our earlier finding showing sub-lethal doses of Irinotecan elevated cell surface levels of PD-L1 comes in contrary with this observation. Our findings suggest that anti-PD-1 or anti-PD-L1 treatment may further enhance the cytotoxic effect.
Tumor Immunostaining of Patient Tissues Shows Variable CD47 Overexpression
Following the in vitro observation that CD47 expression may be modulated under treatment conditions, we next examined whether CD47 is expressed in human neuroblastoma tissues. To address this, we performed immunohistochemical analysis using an archival tissue microarray composed of baseline tumor specimens and normal peripheral nerve controls (Figure 4 and Table 1).
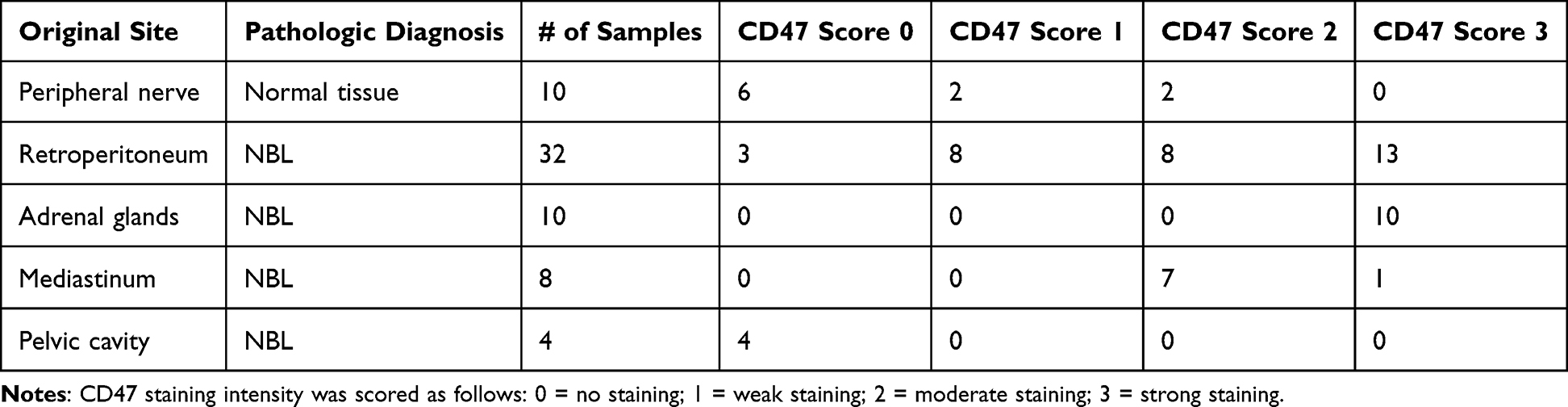 |
Table 1 Distribution of CD47 Immunohistochemical Staining Intensity in Neuroblastoma Samples |
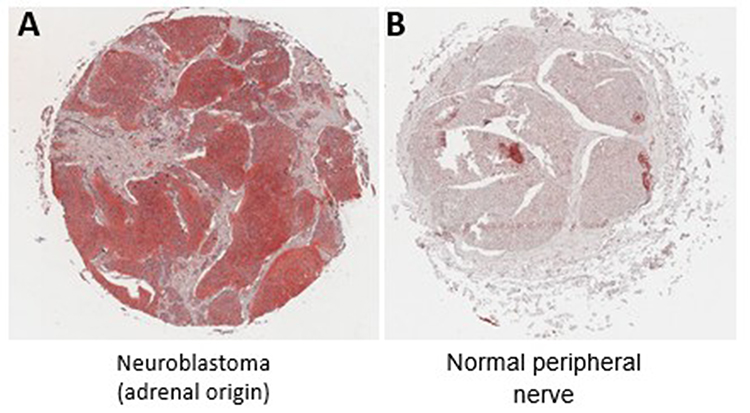 |
Figure 4 CD47 expression in human neuroblastoma tissues. (A) Representative immunohistochemical staining of CD47 in neuroblastoma tumor tissue (adrenal origin), demonstrating strong staining intensity (score 3). (B) Representative staining of normal peripheral nerve tissue, demonstrating absence of CD47 expression (score 0). CD47 expression was evaluated using a scoring system based on staining intensity: 0 (no staining), 1 (weak), 2 (moderate), and 3 (strong). The distribution of staining scores across all samples is presented in.Table 1. |
The immunostaining of tumor sections from NBL patients revealed a pronounced overexpression of CD47 in these tumors when contrasted with normal peripheral nerve tissue. The analyzed NBL samples originated from a diversity of anatomical locations, including the retroperitoneum, adrenal glands, mediastinum, and pelvic cavity.
We implemented a quantification scale to assess the intensity of CD47 staining, ranging from 0 (indicative of no staining) to 3 (denoting maximal staining)., Representative images corresponding to the scoring categories (0 and 3) are shown in Figure 4. Observations included minimal to weak expression predominantly in healthy peripheral nerve tissue and in NBL specimens from the retroperitoneum and pelvic cavity. Conversely, all NBL samples from the adrenal glands exhibited the maximal staining intensity of 3, while those from the mediastinum and retroperitoneum displayed variable scores (Table 1).
CD47 Might Be Shed from NBL Cells to the Surrounding Environment
To determine whether CD47 expression can serve as a diagnostic marker for neuroblastoma, we investigated the shedding of CD47 protein from neuroblastoma cells. We began by examining the SH-SY5Y cell line under various conditions (Figure 5A). Our findings indicated that pre-treatment with sub-lethal doses of irinotecan for 48 hours resulted in elevated levels of CD47, as detected by ELISA. Notably, pre-treatment with IFN-γ produced the most pronounced increase in CD47 levels. Additionally, CD47 was detectable in the medium of untreated neuroblastoma cells. This phenomenon had not been previously documented in the literature, prompting us to extend our investigation to patient samples.
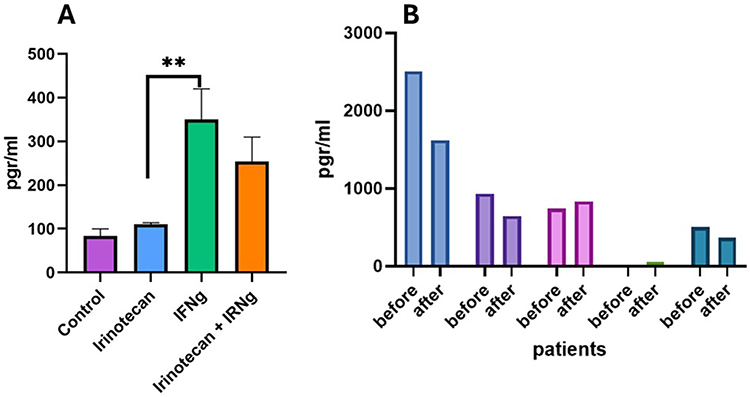 |
Figure 5 Soluble CD47 is detectable in neuroblastoma cell culture supernatants and patient plasma. (A) SH-SY5Y cells were treated with irinotecan (1 µM) and/or IFN-γ (20 ng/mL) for 48 hours. CD47 levels in cell culture supernatants were measured by ELISA. (B) Plasma levels of soluble CD47 in neuroblastoma patients (n=5) were measured by ELISA. Samples were obtained at indicated clinical time points (pre-treatment and post-treatment where available). Patient 1 (light blue), patient 2 (purple), patient 3 (pink), patient 4 (green), patient 5 (dark blue). Data are presented as mean ± SEM. **p < 0.01. |
Following approval from the hospital ethics committee, we analyzed plasma samples from five patients with different stages of Neuroblastoma (Figure 5B and Table 2). The cohort included two patients with high-risk neuroblastoma (HR-NBL), two with metastatic neuroblastoma in children under 18 months (M-NBL), and one with localized disease. All patients were NMYC-negative. The initial three samples were collected before the first treatment, while Patient 4’s samples were taken before and after relapse treatment with Topotecan and Cyclophosphamide. Patient 4 was the only individual who had received irinotecan in this cohort (a few weeks prior to sample collection). Patient 5’s samples was obtained post-chemotherapy. Detailed patient characteristics are presented in Table 2.
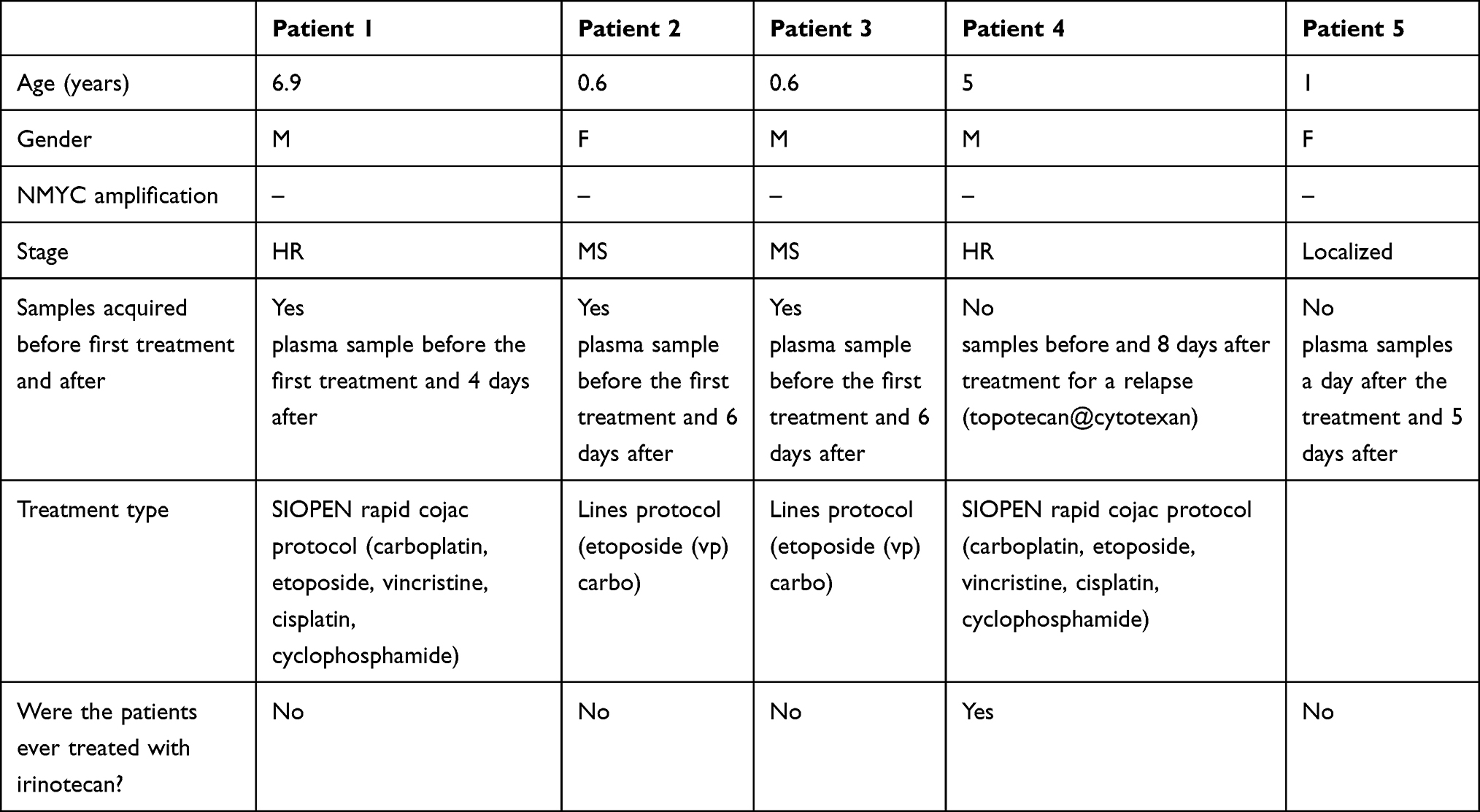 |
Table 2 Neuroblastoma Patients Characteristics |
Our analysis revealed the presence of CD47 in the plasma of all patients. Notably, Patient 4, who had no detectable CD47 in his plasma before treatment, exhibited low levels of CD47 following relapse therapy. However, no consistent trend was observed among the patients. These results suggest that neuroblastoma cells may express CD47 on their surface and release soluble forms of CD47 into the extracellular environment.
In vivo NBL Humanized Mice Model: Combination of PBMCs and Irinotecan Efficiently Eliminates NBL
To study the development of immunity against human neuroblastoma, we established a humanized mouse NBL model. The system comprised human NBL cells that develop tumors in the intraperitoneal (IP) cavity of immune-deficient NSG mice. We used luciferase-expressing NBL cells, which allowed us to monitor tumor growth using a bioluminescent camera (Bioluminescent & Fluorescent In Vivo Systems, IVIS). As we explored the effect of Irinotecan as an immunomodulator, we assessed the activity of human xenogeneic peripheral blood mononuclear cells (PBMCs) in combination with Irinotecan.
PBMCs were injected one day pre-conditioned gamma irradiation. The pre-conditioned total body irradiation (TBI) irradiation of 2 Gray was performed to enhance the engraftment of immune cells.17 We investigated the effect of our PBMCs treatment on the tumor growth without or in addition to Irinotecan treatment.
Mice were pre-injected with cells on day 1 and cancer disease burden was followed with IVIS machine. PBMCs were injected (2 x106) IP into the neuroblastoma bearing NSG mice on day 10, one day following pre-conditioned irradiation. Irinotecan treatment was given IV on the same day.
We found that intraperitoneal PBMC treatment was associated with marked reduction in tumor signal in this model (Figure 6). A single irinotecan treatment alone showed a more limited and less sustained effect. The combination of PBMCs with irinotecan showed numerically lower tumor burden at selected time points and a potentially more sustained response pattern; however, given the exploratory design and group sizes, these findings should be interpreted cautiously.
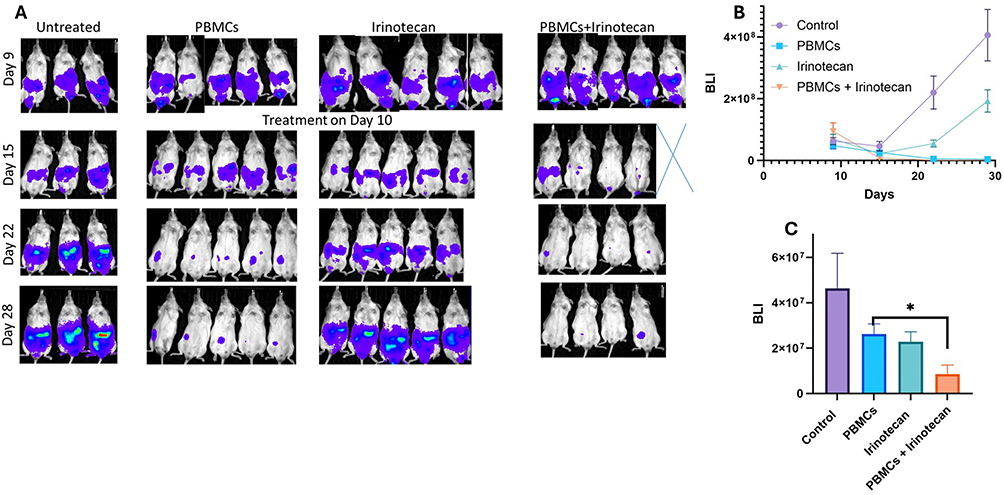 |
Figure 6 Effects of irinotecan and PBMCs on tumor burden in a humanized neuroblastoma mouse model. NSG mice were intraperitoneally injected with luciferase-expressing SH-SY5Y cells. On day 9, mice received total body irradiation (2 Gy) and were randomized into treatment groups: control, PBMCs alone, irinotecan alone, or irinotecan + PBMCs. PBMCs (2 × 106 cells) were administered intraperitoneally, and irinotecan (400 µg/mouse) intravenously. (A) Representative bioluminescence imaging (BLI) of tumor burden.(B) Quantification of BLI signal over time. (C) Tumor burden at day 15. Treatment groups: control (purple), irinotecan (green), PBMCs (blue), irinotecan + PBMCs (Orange).Data are presented as mean ± SEM. *p < 0.05. |
To explore the nature of the obtained effect, we defined the composition of the human effector cells in spleens on day 31, 21 days after the single treatment.
Irinotecan treatment alone did not lead to an increase in spleen weight, probably due to the lack of an intact adaptive immune system in the mice (Figure 7). Injecting PBMCs alone increased spleen weights, and combining PBMCs with Irinotecan led to the greatest increase in spleen weight, a result that may suggest a relationship between Irinotecan treatment and T cell activation (Figure 7A). We found that PBMC-treated spleen samples were composed of about 20% human cells; of these, 10–15% were human T cells, and approximately 40% of human T cells were CD8 effector cells (Figure 7B–D). These findings are consistent with the involvement of cytotoxic CD8+ T cells in mediating the observed tumor eradication. However, direct CD8 depletion studies would be required to conclusively establish casuality.
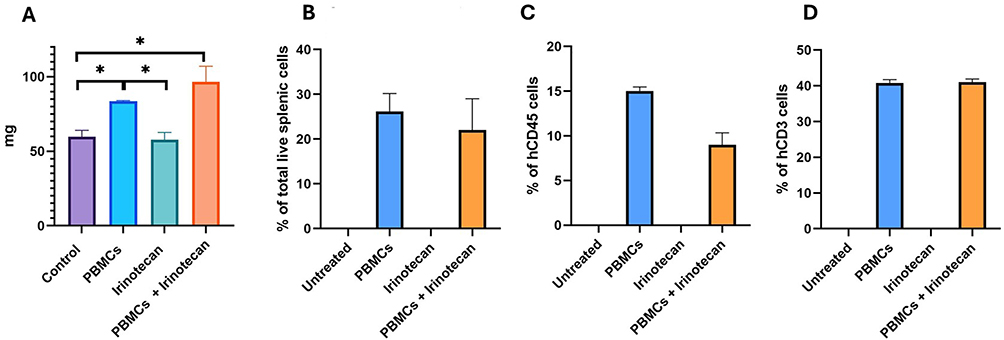 |
Figure 7 Combined irinotecan and PBMC treatment is associated with increased human T-cell representation in spleen. Neuroblastoma tumors were induced by intra-peritoneal injection of NBL SH-SY5Y-LUC cells. On day 10 mice were treated with either PBMCs or Irinotecan or a combination of both. At day 31 post-treatment, spleens were harvested and analyzed by flow cytometry. (A) Spleen weight. (B) Percentage of human leukocytes (hCD45⁺) among total splenocytes. (C) Percentage of human T cells (hCD3⁺) among human leukocytes. (D) Percentage of CD8⁺ T cells among human T cells. Treatment groups: control (purple), irinotecan (blue), PBMCs (green), irinotecan + PBMCs (Orange). Data are presented as mean ± SEM. |
Discussion
The treatment landscape for high-risk neuroblastoma continues to rely heavily on chemotherapy, particularly during the induction and consolidation therapy phases. Radiotherapy, hematopoietic stem cell transplantation (HSCT), and immunotherapy are important components of multimodal therapy. In particular, anti-GD2 immunotherapy has become a mainstay of treatment for patients with high-risk neuroblastoma. There is an urgent need for novel therapeutic strategies. Enhancing our understanding of existing therapies is important for optimizing their use and integrating them with new targeted treatments for improved outcomes.
In our study we looked at the immunomodulatory effects of Irinotecan on NBL cells, and its combination with PBMC’s injection in mice, offering a possible advantage of chemo - immunotherapy combination therapy. We investigated the immunomodulatory effects of irinotecan on neuroblastoma cells and in complementary experimental models. Among the immune-related pathways evaluated, the checkpoint molecule CD47 emerged as one important mechanistic and translational focus of the study.
Our findings suggest that neuroblastoma expresses several key immune modulators and checkpoint inhibitors, including GD2, MHC I, PD-L1, and CD47. We assessed the impact of Irinotecan and interferon-gamma (IFN-γ) on their expression levels. Consistent with the literature, neuroblastomas typically express GD2 and CD47,18 while PD-L1 expression is not uniformly high but is associated with relapse.19 MHC I expression is generally low, particularly in high-risk patients.20 Our baseline pre-treatment findings aligned with these observations. We observed that Irinotecan was associated with higher GD2, suggesting a potential approach for chemo-immunotherapy combination strategies in NBL. The initial PD-L1 levels were relatively low in both tested cell lines. Irinotecan and IFN-γ, separately led to time-dependent increases in PD-L1 expression. While IFN-γ induces PD-L1 primarily through JAK/STAT1/IRF-1 signaling, Irinotecan activates DNA damage response pathways that independently drive IRF-1 and NF-κB-mediated PD-L1 transcription. Combination treatment with irinotecan and IFN-γ was associated with higher PD-L1 expression than untreated cells under the tested conditions, possibly reflecting convergent activation of multiple transcription factor binding sites on the PD-L1 promoter through both IFN-γ-dependent and DNA damage-dependent signaling cascades.3,21–23
MHC I expression increased following IFN-γ treatment, and its combination with Irinotecan had an additive effect. Notably, CD47 expression was elevated after treatment with Irinotecan and IFN-γ, with the combination yielding the highest expression levels. Given that neuroblastomas are often infiltrated with myeloid cells and lymphocytes, which are frequently inactivated,19 supporting the hypothesis that high-risk neuroblastoma is an immunologically cold tumor, we explored the effect of Irinotecan treatment on immune cell-mediated killing. Because IFN-γ has well-established effects on these pathways, the present experiments do not allow definitive separation of IFN-γ-driven effects from irinotecan-specific effects in all settings. Accordingly, these findings should be interpreted as treatment-associated observations rather than formal evidence of synergy.
We began by examining the phagocytic activity of macrophages in NBL cells treated with Irinotecan. Counterintuitively, our results indicated that macrophage phagocytic activity decreases in irinotecan pre-treated NBL cells despite the drug’s direct cytotoxic effects. This apparent paradox can be mechanistically explained by CD47 upregulation observed in our initial findings. Irinotecan, a topoisomerase I inhibitor, induces double-strand breaks (DSBs) and activates the DNA damage response (DDR) pathway, which induces CD47 upregulation through an mre-11-dependent mechanism.3,24 We propose that DNA damage in irinotecan-exposed NBL cells triggers sustained CD47 expression as an adaptive immune escape response. While irinotecan initiates immunogenic effects through myeloid microenvironment remodeling, it simultaneously induces CD47 upregulation as a compensatory immune checkpoint activation that counteracts macrophage-mediated phagocytosis. Notably, the addition of an anti-CD47 antibody to the phagocytic assay was associated with increased macrophage-mediated tumor cell clearance, consistent with a role for CD47 in this assay. This finding suggests that CD47 upregulation represents a compensatory immune checkpoint activation following chemotherapy exposure, a potential mechanism of therapeutic resistance that may be overcome through combination strategies targeting both the myeloid microenvironment and CD47-SIRPα signaling. CD47 tissue microarray analysis of NBL patient samples revealed location-dependent overexpression, consistent with spatial heterogeneity of tumor immune microenvironment regulation and supporting the clinical relevance of CD47.
Recent analyses have underscored the biological and therapeutic relevance of the CD47–SIRPα axis in pediatric tumors. this pathway is particularly active in the myeloid-dominant tumor microenvironment of neuroblastoma, functioning as a “don’t-eat-me” signal that restricts macrophage and dendritic-cell activation.25 CD47 interacts with other immune checkpoints, including PD-1/PD-L1 and MHC I pathways, suggesting that its blockade may convert otherwise “cold” tumors into immunologically responsive ones.26 Tang et al (2024)13 provided a comprehensive perspective emphasizing that CD47 expression correlates with poor prognosis and therapeutic resistance in high-risk neuroblastoma. Our finding that Irinotecan induces CD47 expression extends this literature, suggesting that chemotherapy may enhance this inhibitory axis, thus creating an opportunity and a necessity for combination approaches integrating CD47 blockade with cytotoxic therapy. However, the present study was not designed to determine whether this effect is unique to irinotecan or shared by other cytotoxic agents, including other topoisomerase I inhibitors such as topotecan.
Next, we explored the effect of Irinotecan on T cell-mediated killing of NBL cells.
Despite increased PD-L1 expression following Irinotecan treatment in our initial results, this change did not translate into enhanced T-cell–mediated killing, mirroring the limited clinical success of PD-1/PD-L1 inhibitors in pediatric solid tumors. Neuroblastoma remains an immunologically “cold” malignancy characterized by low tumor mutational burden, scarce effector T-cell infiltration, and dominance of immunosuppressive myeloid populations.27 Unlike adult malignancies, where immune checkpoint inhibitors have revolutionized outcomes, pediatric tumors often lack the pre-existing immune activation required for PD-1 blockade to work effectively.28 Consistent with this biology clinical studies demonstrated minimal objective responses to nivolumab in children with relapsed or refractory solid tumors, including neuroblastoma.29 Together, these observations suggest that PD-1/PD-L1 targeting is unlikely to produce meaningful benefit in neuroblastoma unless combined with strategies that increase antigen presentation and remodel the myeloid-dominant microenvironment, mechanisms potentially engaged by Irinotecan in our study, future studies may explore whether checkpoint blockade could further modulate these responses in rational combination strategies.
Given our results indicating that CD47 is a potential therapeutic target for NBL, coupled with recent studies highlighting its promise in combination therapy, we explored whether soluble CD47 (sCD47) could serve as a circulating diagnostic and prognostic marker. We first detected sCD47 in the supernatant of NBL cell cultures using an ELISA assay, warranting investigation of CD47 levels in plasma from NBL patients before and after treatment. Our preliminary results demonstrated detectable sCD47 in small cohort of patient plasma, suggesting that soluble CD47 may warrant exploration in larger future cohorts. Soluble CD47 arises through two mechanistically distinct pathways: proteolytic shedding via metalloproteinases that generates soluble ectodomains retaining SIRPα-binding capacity, and exosomal release packaging CD47 into extracellular vesicles that circulate as CD47-expressing nanoparticles capable of immune suppression through CD47 cross-dressing.30 Neuroblastoma cells actively secrete CD47-expressing EVs, facilitating systemic immune evasion and metastatic dissemination.31
Finally, we assessed the impact of Irinotecan on PBMC function using our laboratory’s novel intraperitoneal injection model in mice. We found that sub-lethal doses of Irinotecan, when combined with irradiation and PBMC injections, yielded the best therapeutic outcomes in the NBL mouse model. Interestingly, spleen weight analysis revealed that this treatment combination resulted in heavier spleens compared to PBMC and irradiation alone, suggesting enhanced T cell activation. This could be explained by several mechanisms, one of them is Upregulation of MHC Class I Expression, as we shown in our study.
Our findings should be interpreted in the context of several limitations.
First, it represents a pilot exploratory study using two established neuroblastoma cell lines, which do not fully capture tumor heterogeneity. Second, although IFN-γ was included as a biologically relevant comparator, its relative contribution versus irinotecan cannot be fully separated in the current design. Also, we did not establish whether CD47 modulation is unique to irinotecan or shared by other chemotherapeutic agents, including other topoisomerase I inhibitors such as topotecan.
A limitation of the phagocytosis assay is the absence of an isotype IgG control antibody, which would have strengthened exclusion of Fc receptor–mediated effects independent of CD47 blockade.
Functional characterization of the T-cell mediated cytotoxicity part was not performed in the current study and would be valuable in future mechanistic investigations.
Regarding the tissue array, detailed clinicopathologic correlations, including MYCN status and stage-based analyses, were limited by the available metadata of the commercial tissue array.
The in vivo PBMC model relies on xenogeneic immune interactions that may amplify anti-tumor effects beyond those expected in human disease.
Our NBL cells environment findings require cautious interpretation given our small preliminary cohort size, which limits generalizability. Future neuroblastoma-specific studies must establish: (1) reference ranges in healthy pediatric controls; (2) whether sCD47 stratifies patients by risk category (MYCN, 1p deletion); (3) the relative contribution of membrane-derived versus EV-associated CD47; and (4) whether serial sCD47 dynamics during treatment predict therapeutic response or recurrence.
Accordingly, these results should be viewed as preliminary and hypothesis-generating. The broader value of the study lies in the integrated exploratory framework combining in vitro observations with in vivo and patient-derived data.
In conclusion, this pilot exploratory study suggests that irinotecan may influence several immune-related pathways in neuroblastoma, including checkpoint expression and interactions with innate and adaptive immune cells. These preliminary findings support further mechanistic investigation of chemo-immunotherapy combinations and CD47-targeted strategies in future neuroblastoma research.
AI
Generative Artificial Intelligence (AI) was used solely for language refinement during manuscript preparation. Specifically, ChatGPT (OpenAI, GPT-5.0 model) was utilized to improve clarity, grammar, and flow of the written text. The tool was not used for research planning, study design, data generation, data analysis, interpretation of results, or any other scientific or methodological component of this work. All scientific content, conclusions, and interpretations were developed exclusively by the authors. The use of AI-assisted language editing was performed to enhance readability and ensure clear communication of the research findings.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request. Public sharing of the data is restricted due to ethical approvals governing the use of human-derived biological materials and institutional biobank regulations. Anonymized data can be made available to qualified researchers upon reasonable request and with appropriate ethical approval.
Ethics Statement
Approval for using patients plasma samples were given by Institutional Review Board (IRB) approval (approval number 0495-23-HMO) from the Hadassah–Hebrew University Medical Center Ethics Committee prior to the initiation of the study. The research involved retrospective analysis of archived plasma samples obtained from the hospital biobank. Informed consent for participation in the study was waived by the IRB.
Approval for the animal experiments were given by the Animal Care Committee of Hebrew University. Permit number MD-23-17149-5. Application title: Establishment of the murine model for neuroblastoma and investigation of the effect of treatment using the combination of chemotherapy and immunotherapy. Maintenance of mice and all experimental procedures were approved by and performed in accordance with the Institutional Animal Care and Use Committee-approved animal treatment protocols (NIH approval number: OPRR-A01-5011).
Acknowledgments
We acknowledge and appreciate the collaboration between the Goldyne Savad gene therapy institute at Hadassah Hospital and the pediatric Hemato-Oncology department at the hospital. We thank Tali Liberman and Iris Aynor of the pediatric Hemato- Oncology department for helping us with the human ethics and Helsinki approval, and for helping us obtain samples. Thank you for professor Itay Galun for his notes concerning scientific research model.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Nong J, Su C, Li C, et al. Global, regional, and national epidemiology of childhood neuroblastoma (1990-2021): a statistical analysis of incidence, mortality, and DALYs. EClinicalMedicine. 2024;79:102964. doi:10.1016/j.eclinm.2024.102964
2. Voeller J, Erbe AK, Slowinski J, et al. Combined innate and adaptive immunotherapy overcomes resistance of immunologically cold syngeneic murine neuroblastoma to checkpoint inhibition. J ImmunoTher Cancer. 2019;7(1). doi:10.1186/s40425-019-0823-6
3. Iwai T, Sugimoto M, Wakita D, et al. Topoisomerase I inhibitor, irinotecan, depletes regulatory T cells and up-regulates MHC class I and PD-L1 expression, resulting in a supra-additive antitumor effect when combined with anti-PD-L1 antibodies. Oncotarget. 2018;9(59):31411–15. doi:10.18632/oncotarget.25830
4. Kushner BH. Chemoimmunotherapy for high-risk neuroblastoma. Lancet Oncol. 2017;18(7):845–846. doi:10.1016/s1470-2045(17)30337-6
5. Qiu B, Katherine KM. Advancing therapy for neuroblastoma. Nat Rev Clin Oncol. 2022;19(8):515–533. doi:10.1038/s41571-022-00643-z
6. Galluzzi L, Buque A, Kepp O, Zitvogel L, Kroemer G. Immunological effects of conventional chemotherapy and targeted anticancer agents. Cancer Cell. 2015;28(6):690–714. doi:10.1016/j.ccell.2015.10.012
7. Paret C, Ustjanzew A, Ersali S, et al. GD2 expression in medulloblastoma and neuroblastoma for personalized immunotherapy: a matter of subtype. Cancers. 2022;14(24):6051. doi:10.3390/cancers14246051
8. Sait S, Modak S. Anti-GD2 immunotherapy for neuroblastoma. Expert Rev Anticancer Ther. 2017;17(10):889–904. doi:10.1080/14737140.2017.1364995
9. Terzic T, Cordeau M, Herblot S, et al. Expression of disialoganglioside (GD2) in neuroblastic tumors: a prognostic value for patients treated with anti-GD2 immunotherapy. Pediatr Dev Pathol. 2017;21(4):355–362. doi:10.1177/1093526617723972
10. Mastrangelo S, Rivetti S, Triarico S, et al. Mechanisms, characteristics, and treatment of neuropathic pain and peripheral neuropathy associated with dinutuximab in neuroblastoma patients. Int J Mol Sci. 2021;22(23):12648. doi:10.3390/ijms222312648
11. Mora J, Climent A, Roldán M, et al. Desensitizing the autonomic nervous system to mitigate anti-GD2 monoclonal antibody side effects. Front Oncol. 2024;14:1380917. doi:10.3389/fonc.2024.1380917
12. Jaiswal S, Jamieson CH, Pang WW, et al. CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell. 2009;138(2):271–285. doi:10.1016/j.cell.2009.05.046
13. Tang XX, Shimada H, Ikegaki N. A perspective on the CD47-sirpa axis in high-risk neuroblastoma. Current Oncol. 2024;31(6):3212–3226. doi:10.3390/curroncol31060243
14. Tang XX, Shimada H, Ikegaki N. Macrophage-mediated anti-tumor immunity against high-risk neuroblastoma. Genes Immun. 2022;23(3–4):129–140. doi:10.1038/s41435-022-00172-w
15. Sikic BI, Lakhani N, Patnaik A, et al. First-in-human, first-in-class Phase I trial of the anti-cd47 antibody Hu5F9-G4 in patients with advanced cancers. J clin oncol. 2019;37(12):946–953. doi:10.1200/jco.18.02018
16. Theruvath J, Menard M, Smith BA, et al. Anti-GD2 synergizes with CD47 blockade to mediate tumor eradication. Nature Med. 2022;28(2):333–344. doi:10.1038/s41591-021-01625-x
17. Andrade J, Ge S, Symbatyan G, Rosol MS, Olch AJ, Crooks GM. Effects of sublethal irradiation on patterns of engraftment after murine bone marrow transplantation. Biol Blood Marrow Transplant. 2011;17(5):608–619. PMID: 29868020; PMCID: PMC5964139. doi:10.1016/j.bbmt.2010.12.697
18. Stip MC, Teeuwen L, Dierselhuis MP, Leusen JH, Krijgsman D. Targeting the myeloid microenvironment in neuroblastoma. J Exp Clin Cancer Res. 2023;42(1). PMID: 21176787; PMCID: PMC3086732. doi:10.1186/s13046-023-02913-9
19. Saletta F, Vilain RE, Gupta AK, et al. Programmed death-ligand 1 expression in a large cohort of pediatric patients with solid tumor and association with clinicopathologic features in neuroblastoma. JCO Precis Oncol. 2017;1:1–12. doi:10.1200/po.16.00049
20. Haworth KB, Leddon JL, Chen CY, Horwitz EM, Mackall CL, Cripe TP. Going back to class I: MHC and immunotherapies for childhood cancer. Pediatr Blood Cancer. 2014;62(4):571–576. doi:10.1002/pbc.25359
21. Sato H, Niimi A, Yasuhara T, et al. DNA double-strand break repair pathway regulates PD-L1 expression in cancer cells. Nat Commun. 2017;8(1):1751. doi:10.1038/s41467-017-01883-9
22. Garcia-Diaz A, Shin DS, Moreno BH, et al. Interferon receptor signaling pathways regulating PD-L1 and PD-L2 expression. Cell Rep. 2017;19(6):1189–1201. doi:10.1016/j.celrep.2017.04.031
23. Numata Y, Akutsu N, Ishigami K, et al. Synergistic effect of IFN-γ and IL-1β on PD-L1 expression in hepatocellular carcinoma. Biochem Biophys Rep. 2022;30:101270. doi:10.1016/j.bbrep.2022.101270
24. Ghantous L, Volman Y, Hefez R, et al. The DNA damage response pathway regulates the expression of the immune checkpoint CD47. Commun Biol. 2023;6(1):245. doi:10.1038/s42003-023-04615-6
25. Gupta A, Taslim C, Tullius BP, Cripe TP. Therapeutic modulation of the CD47-SIRPα axis in the pediatric tumor microenvironment: working up an appetite. Cancer Drug Resist. 2020;3(3):550–562. doi:10.20517/cdr.2020.12
26. Cocco C, Morandi F, Airoldi I. Immune checkpoints in pediatric solid tumors: targetable pathways for advanced therapeutic purposes. Cells. 2021;10(4):927. PMID: 35582455; PMCID: PMC8992496. doi:10.3390/cells10040927
27. Kennedy PT, Zannoupa D, Son MH, Dahal LN, Woolley JF. Neuroblastoma: an ongoing cold front for cancer immunotherapy. J Immunother Cancer. 2023;11(11):e007798. PMID: 33920505; PMCID: PMC8074115. doi:10.1136/jitc-2023-007798
28. Mora J, Modak S. Nivolumab in paediatric cancer: children are not little adults. Lancet Oncol. 2020;21(4):474–476. PMID: 37993280; PMCID: PMC10668262. doi:10.1016/S1470-2045(20)30086-3
29. Pasqualini C, Rubino J, Brard C, et al. Phase II and biomarker study of programmed cell death protein 1 inhibitor nivolumab and metronomic cyclophosphamide in paediatric relapsed/refractory solid tumours: arm G of AcSé-ESMART, a trial of the European innovative therapies for children with cancer consortium. Eur J Cancer. 2021;150:53–62. PMID: 32192574doi:10.1016/j.ejca.2021.03.032
30. He J, Ren W, Wang W, et al. Exosomal targeting and its potential clinical application. Drug Deliv Transl Res. 2022;12(10):2385–2402. PMID: 33892407. doi:10.1007/s13346-021-01087-1
31. Dhamdhere MR, Spiegelman VS. Extracellular vesicles in neuroblastoma: role in progression, resistance to therapy and diagnostics. Front Immunol. 2024;15:1385875. PMID: 34973131; PMCID: PMC9458566. doi:10.3389/fimmu.2024.1385875
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Pathologic Complete Response Prediction to Neoadjuvant Immunotherapy Combined with Chemotherapy in Resectable Locally Advanced Esophageal Squamous Cell Carcinoma: Real-World Evidence from Integrative Inflammatory and Nutritional Scores
Feng J, Wang L, Yang X, Chen Q, Cheng X
Journal of Inflammation Research 2022, 15:3783-3796
Published Date: 6 July 2022
Nanoparticles for Chemoimmunotherapy Against Triple-Negative Breast Cancer
Liu S, Li J, Gu L, Wu K, Xing H
International Journal of Nanomedicine 2022, 17:5209-5227
Published Date: 7 November 2022
Pembrolizumab in Lymphopenic Metastatic Breast Cancer Patients Treated with Metronomic Cyclophosphamide: A Clinical and Translational Prospective Study
Mery B, Ménétrier-Caux C, Montané L, Heudel PE, Ray-Coquard I, Bachelot T, Derbel O, Augereau P, Treilleux I, Berthet J, Nkodia A, Bardin-Dit-Courageot C, Attignon V, Ferrari A, Garin G, Perol D, Caux C, Dubois B, Trédan O
Breast Cancer: Targets and Therapy 2023, 15:311-325
Published Date: 27 April 2023
Breast Cancer: An Overview of Current Therapeutic Strategies, Challenge, and Perspectives
Wang J, Wu SG
Breast Cancer: Targets and Therapy 2023, 15:721-730
Published Date: 20 October 2023
Diagnosis, Prognosis, and Treatment of Triple-Negative Breast Cancer: A Review
Jie H, Ma W, Huang C
Breast Cancer: Targets and Therapy 2025, 17:265-274
Published Date: 17 March 2025