Back to Journals » Infection and Drug Resistance » Volume 19
Integrated UHPLC-QTOF-MS, Network Pharmacology, and Experimental Validation Reveal the Antiviral and Anti-Inflammatory Mechanisms of Xiaoer Fengreqing Against COVID-19-Associated Encephalitis
Authors Chen R, Li X ![]() , Wang M, Xiong Y, Xie P, Liu Y, Cui Z, Ye Y, Li R, Hon C, Ma Q
, Wang M, Xiong Y, Xie P, Liu Y, Cui Z, Ye Y, Li R, Hon C, Ma Q
Received 27 March 2026
Accepted for publication 21 June 2026
Published 15 July 2026 Volume 2026:19 610625
DOI https://doi.org/10.2147/IDR.S610625
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Alberto Ospina Stella
Ruihan Chen,1,* Xuanxuan Li,2,* Menglei Wang,3,* Yueyue Xiong,2,* Peifang Xie,2,4 Yan Liu,3 Zhanyu Cui,2 Yi Ye,2 Runfeng Li,2,* Chitin Hon,1 Qinhai Ma2
1Department of Engineering Science, Faculty of Innovation Engineering, Macau University of Science and Technology, Taipa, Macao SAR, People’s Republic of China; 2State Key Laboratory of Respiratory Disease, National Clinical Research Center for Respiratory Disease, Guangzhou Institute of Respiratory Health, The First Affiliated Hospital of Guangzhou Medical University, Guangzhou, Guangdong, People’s Republic of China; 3Research Department, Hebei Province (Handan) Chinese Medicine Industry Technology Research Institute, Handan, Hebei, People’s Republic of China; 4State Key Laboratory of Quality Research in Chinese Medicine, Macau University of Science and Technology, Taipa, Macau SAR, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qinhai Ma; Chitin Hon, Email [email protected]; [email protected]
Background: Xiaoer Fengreqing (XF), a traditional Chinese medicine, is included in pediatric COVID-19 guidelines, but its active components and mechanisms remain unclear. This study aimed to explore XF’s bioactive constituents and potential effects against COVID-19-related encephalitis.
Methods: UHPLC-QTOF-MS and network pharmacology were used to identify active compounds, targets, and pathways. Antiviral effects against Omicron were assessed by cytopathic inhibition assays. RT-qPCR and Western blot evaluated anti-inflammatory responses, and in vivo experiments tested antiviral and neuroprotective effects against HCoV-OC43 using an intracranial infection model.
Results: UHPLC-QTOF-MS identified 220 compounds, with 137 absorbed into plasma, yielding 774 potential targets and 242 KEGG pathways. Core targets included ACE2, EGFR, PARP1, JAK1, and SRC. Molecular docking showed isoalantolactone and isosakuranetin interacted with ACE2. XF exhibited an IC50 of 5.687 mg/mL and selectivity index of 7.06. It significantly reduced TNF-α, IL-6, MCP-1, and IP-10, and downregulated p-STAT1, p-AKT, and p-p38 MAPK. In vivo, XF prolonged survival, reduced HCoV-OC43 titers and brain index, and decreased IFN-γ, TNF-α, IP-10, and IL-6 levels. Pathology confirmed reduced brain necrosis and immune infiltration at high dose.
Conclusion: XF may exert antiviral and neuroprotective effects against COVID-19-related encephalitis through TNF-α/IL-6 suppression and modulation of MAPK, JAK/STAT, and PI3K/AKT immune regulatory pathways. Although XF showed potential for further development, its clinical translation remains limited by the complexity of its components, the lack of evaluation of individual active compounds, and its relatively modest in vitro antiviral potency. Therefore, in the absence of clinical trial evidence, XF should be regarded only as a candidate for further investigation.
Keywords: Xiaoer Fengreqing, XF, coronavirus, UHPLC-QTOF-MS, network pharmacology, molecular docking
Introduction
SARS-CoV-2 emerged in late 2019 and rapidly spread worldwide, profoundly affecting global health.1 Although most patients experience mild to moderate illness, approximately 5–10% develop severe or life-threatening conditions.2
With the rollout of COVID-19 vaccines, the relative burden of COVID-19 may shift toward younger age groups. Children and adolescents account for a substantial proportion of reported COVID-19 cases.3 In the United States, nearly 15.5 million pediatric COVID-19 cases have been reported, accounting for approximately 18% of all reported cases.4 It has been well established that children, including neonates, are vulnerable to long-term consequences of SARS-CoV-2 infection, such as multisystem inflammatory syndrome and prolonged neurological complications.5,6 Large-scale clinical investigations have further confirmed that a considerable proportion of patients experience persistent post-COVID-19 symptoms, indicating a substantial disease burden.7 Notably, COVID-19-associated encephalitis is a severe pediatric complication, characterized by systemic inflammation and blood–brain barrier disruption. This highlights the need for therapeutic agents capable of simultaneously exerting antiviral and neuroprotective effects. A multicenter study in the United States reported that 7.0% of hospitalized children experienced neurological manifestations, including seizures, which may be associated with systemic inflammation and blood–brain barrier disruption.8
Despite the availability of several licensed anti-COVID-19 drugs, therapeutic options for pediatric patients remain limited. Certain antivirals approved for adults, such as molnupiravir, are contraindicated in children due to potential adverse effects on bone development. Other agents, including Paxlovid, are restricted to children aged ≥12 years and weighing ≥40 kg, while remdesivir remains the only broadly applicable option.9 However, the efficacy and safety of these agents in children remain insufficiently established due to limited pharmacokinetic data and randomized clinical trials. Consequently, there is an urgent need to develop safe and effective COVID-19 therapies for children. Increasing evidence suggests that traditional Chinese medicine (TCM) has been widely used in pediatric COVID-19 treatment with favorable clinical outcomes and minimal adverse effects. In March 2022, the WHO convened an expert meeting to evaluate TCM for COVID-19 treatment. The meeting explicitly concluded that TCM is safe and effective in reducing disease progression, shortening viral shedding time, and decreasing hospitalization duration in mild to moderate cases, formally recommending its integration into COVID-19 management strategies. A recent review further summarized the clinical benefits of TCM and its role in modern therapeutic development.10 Multiple Chinese patent medicines, such as Liushen Capsules and Jinzhen granules, have demonstrated significant antiviral activities against SARS-CoV-2 and other human coronaviruses in vivo and in vitro, while alleviating virus-induced lung injury.11,12
Recent clinical and preclinical studies have highlighted the efficacy of various TCM interventions, such as thunder-fire moxibustion, Lianhua Qingwen, and Xuanfei Baidu Granules, on alleviating COVID-19 symptoms, reducing viral loads, and modulating immune responses, particularly in pediatric cohorts.13–17 Building on its extensive historical practice and ongoing innovation,18,19 the utilization of TCM represents a highly favorable clinical option. However, these previous studies have predominantly focused on pulmonary outcomes and general symptom relief, leaving the therapeutic mechanisms of TCM against pediatric COVID-19-associated neurological complications less explored.
Xiaoer Fengreqing (XF) is a TCM remedy frequently employed for treating colds in children.20 The formulation is derived from the TCM formula “Yinqiao Powder” and has been refined by incorporating syndrome distinction through the process of addition and subtraction. XF consists of Lonicera japonica Thunb (called Jinyinhua, JYH in Chinese), Forsythia suspensa (Thunb). Vahl (called Lianqiao, LQ in Chinese), Isatis tinctoria L. (called Banlangen, BLG in Chinese), Mentha haplocalyx Briq. (called Bohe, BH in Chinese), Bupleurum chinense DC. (called Chaihu, CH in Chinese), Arctium lappa L. (called Niubangzi, NBZ in Chinese), Schizonepeta tenuisfolia (Benth). Briq. (called Jingjiesui, JJS in Chinese), Gypsum Fibrosuum (called Shigao, SG in Chinese), Scutellaria baicalensis Georgi (called Huangqin, HQ in Chinese), Gardenia jasminoides J.Ellis (called Zhizi, ZZ in Chinese), Platycodon grandiflorus (Jacq). A.DC. (called Jiegeng, JG in Chinese), Paeonia lactiflora Pall. (called Chishao, CS in Chinese), Phragmites communis Trin. (called Lugen, LG in Chinese), Prunus armeniaca L. (called Kuxingren, KXR in Chinese), Lophatherum gracile Brongn. (called Danzhuye, DQY in Chinese), Citrus x aurantium L. (called Zhiqiao, ZQ in Chinese), Medicated Leaven (called liushenqu, LSQ in Chinese), Bombyx mori L. (called Jiangcan, JC in Chinese), Saposhnikovia divaricata (Turcz). Schischk. (called Fangfeng, FF in Chinese) and Glycyrrhiza uralensis Fisch. (called Gancao, GC in Chinese). All the plant names have been checked with “The Plant List” (http://www.theplantlist.org), with the exception of Gypsum Fibrosuum (called Shigao, SG in Chinese), Medicated Leaven (called liushenqu, LSQ in Chinese) and Bombyx mori L. (called Jiangcan, JC in Chinese). XF exhibits a therapeutic impact on viral pneumonia induced by the influenza virus, and its mechanism of action may be associated with the amelioration of inflammation and the mitigation of viral load.21 Moreover, TCM has been suggested to remodel immune responses in long COVID-19.22 The State Administration of TCM of China has currently incorporated XF into the therapy regimen for pediatric COVID-19. The potential therapeutic effects of XF on COVID-19 may be attributed to its ability to bind to ACE2 and 3CLpro, as well as its impact on various signaling pathways including PI3K-Akt, JAK-STAT, MAPK, and NF-κB.21 While several studies have examined the use of XF in treating SARS-CoV-2 in children, the underlying mechanisms that contribute to the effectiveness of XF in combating new coronavirus infections remain uncertain.
Hence, the primary objective of this study was to examine the potential effects of XF in experimental models of coronavirus-associated encephalitis, while also exploring its bioactive constituents and probable underlying mechanisms. In this study, the mechanism of XF multi-components in the treatment of COVID-19 were investigated using a combination of UHPLC-QTOF-MS and network pharmacology approaches, with core targets prediction highlighting potential regulation of multiple signaling pathways. Next, the Omicron variant’s cytotoxicity and antiviral activities were evaluated in vitro, and the virus’s anti-inflammatory capabilities were assessed by identifying inflammatory factors using RT-qPCR. Signaling biomarkers modulation were assessed by Western blot. Subsequently, in vivo experiments were conducted to assess the antiviral and anti-encephalitic characteristics of XF in the context of coronavirus HCoV-OC43 infection.
Materials and Methods
Ethics and Biosafety Statement
All animal experiments were reviewed and approved by the Ethics Committee of Animal Experiments at Guangzhou Medical University (approval No. 20230250) and were conducted in accordance with the National Research Council Guidelines and the Chinese Animal Protection Act. The serum pharmacochemistry experiment in rats was approved by the Laboratory Animal Management and Use Committee of Weitong Lihua Laboratory Animal Technology Co., Ltd. (ethical approval No. P2021096).
All experiments involving the SARS-CoV-2 Omicron variant were performed in the biosafety level 3 (BSL-3) laboratory at the Guangzhou Customs Inspection and Quarantine Technology Center. This laboratory was approved by the National Health Commission of the People’s Republic of China to conduct experimental activities involving highly pathogenic microorganisms, including SARS-CoV-2. The experimental protocol was approved under approval No. IQTC2026021-01. All procedures involving infectious SARS-CoV-2 were conducted in accordance with institutional biosafety regulations and national guidelines for BSL-3 laboratory operation.
Reagents
The XF (batch No. S03520001), which is commercially accessible, was supplied by Handan Pharmaceutical Co., Ltd. Its specification is 60 mL each bottle, with each 1 mL corresponding to 2.0 g of decoction pieces. The analysis using LC-MS/MS was conducted using a Shimadzu Nexera ultra-high performance liquid chromatography LC-30A system equipped with a Waters UPLC BEH C18 column measuring 1.7 μm 2.1*100 mm. The mobile phase was composed of a solution containing 0.1% formic acid in water (A) and in acetonitrile (B). The multi-step linear elution gradient program consisted of the following steps: 0–3.5 min, 95–85% A; 3.5–6 min, 85–70%A; 6–6.5 min, 70% A; 6.5 −12 min, 70–30% A; 12 −18 min, 30% A; 18–22 min, 100% B.
Virus, Cells, and Animals
Vero E6, Calu-3, HRT-18, and HMC3 cells were originally obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The epithelial cells of African green monkeys (Vero E6) were subjected to treatment with 10% fetal bovine serum (FBS) obtained from ExCell Biotech, China, along with an antibiotic concentration of 100 U/mL penicillin. The cultures were cultivated in Dulbecco’s modified Eagle medium (DMEM, USA) supplemented with 100 μg/mL of streptomycin (Gibco, USA). The cells were kept at a temperature of 37 °C in an atmosphere with 5% CO2. The propagation and adaptation of the SARS-CoV-2 Omicron variant, which was obtained from the First Affiliated Hospital of Guangzhou Medical University, followed the previously described methodology. The Reed-Muench method was employed to ascertain the 50% tissue culture infectious dose (TCID50) of the SARS-CoV-2 Omicron variant (TCID50 = 10−5/100 μL) using the Vero E6 cells. The working reservoir of the virus was collected and stored at a temperature of −80°C. Vero E6 cells were selected for cytotoxicity and CPE inhibition assays because they are highly permissive to SARS-CoV-2 infection and allow clear observation of virus-induced cytopathic effects, making them suitable for antiviral screening.23,24 Calu-3 cells, a human lung epithelial cell line with endogenous expression of ACE2 and TMPRSS2, were used for RT-qPCR and Western blot analyses because they are suitable for assessing host responses and signaling pathway activation following SARS-CoV-2 infection.25 Therefore, Vero E6 cells were used to evaluate antiviral activity, whereas Calu-3 cells were used to assess the anti-inflammatory and signaling-regulatory effects of XF.
The HRT-18 human colon cancer tumor cells and the HCoV-OC43 strain, which has been tailored for the mouse brain, were preserved in the Virus Laboratory at the State Key Laboratory of Respiratory Diseases, Guangzhou Institute of Respiratory Health. The process of HCoV-OC43 amplification was carried out. A volume of 30 μL of LD50 HCoV-OC43 was administered intracranially to healthy C57BL/6 mice within 10 days of their birth, with a weight range of 4 to 6 g. Four days following the test, the brain tissues were collected and subjected to homogenization. The HCoV-OC43 virus was purified using centrifugation at 5000 g/min for 15 minutes at a temperature of 4°C. To evaluate the LD50 (median lethal dosage) of HCoV-OC43 in suckling mice, a freshly amplified HCoV-OC43 viral solution was diluted in sterile PBS using a 10-fold gradient for seven dilution gradients. A total of 42 healthy suckling mice, all under the age of 10 days, were randomly allocated into 7 groups. Each group consisted of 6 mice and was subjected to a gradient dilution inoculation ranging from 10−1 to 10−7. Each experimental group received an intracranial injection of a 30 μL gradient dilution of the virus. Over a period of 7 days, the experiment was monitored, with daily measurements of body weight and number of deaths. The LD50 was determined using the Reed-Muench method. The LD50 value of the virus employed in this experimental study was reported as 10−5.45/30 μL. The 42 suckling mice were used exclusively for HCoV-OC43 LD50 determination rather than for the subsequent treatment-efficacy experiment. The sample size was based on the Reed–Muench endpoint-titration design rather than a conventional between-group efficacy comparison. Seven 10-fold serial virus dilutions were tested, with six mice allocated to each dilution group. The mortality data across the dilution groups were used to estimate the median lethal dose while minimizing animal use in accordance with the 3R principle. Similar HCoV-OC43 suckling-mouse models have used serial virus dilutions and the Reed–Muench method to determine the median lethal dose.26
Investigation of the Bioactive Ingredients of XF Through UHPLC-QTOF-MS Analysis
Experiment of Serum Drug Chemistry in vivo
The study utilized male SD rats with a weight of 200 ± 20 g, which were obtained from Beijing Weitong Lihua Experimental Animal Technology Co., Ltd. The animal production license number assigned to the rats was SCXK (Beijing) (No. 2021–0006). The animals were kept in a controlled environment characterized by a consistent temperature range of 20–26°C and humidity levels ranging from 50% to 65%. The experiment involved the provision of sterilized feed and purified water, as well as the implementation of a 12 h alternating light and dark lighting system. The experiment received approval from the Laboratory Animal Management and Use Committee of Weitong Lihua Laboratory Animal Technology Co., Ltd., under the ethical evaluation number P2021096. Following a five-day period of adaptive eating, the rats were allocated into two groups: a blank control group and a drug delivery group. Each group consisted of three rats. The administration group of rats received a dosage of 22.5 mL/kg XF via gavage, whereas the blank control group of rats received an equivalent volume of distilled water via gavage. Blood samples were obtained after a period of 7 d. Animals were given only water and no food for 12 h prior to sample collection. Pentobarbital sodium was administered intraperitoneally one hour after intragastric administration to induce anesthesia. Blood was collected from the abdominal aorta, and serum was recovered through centrifugation at 4 °C at 3000 r/min for 10 min. After blood collection, rats were humanely euthanized via intraperitoneal injection of an overdose of sodium pentobarbital (150 mg/kg) in compliance with the AVMA Guidelines for the Euthanasia of Animals.
Metabolites Extraction
For the purpose of mass spectrometry analysis, the commercially available XF was employed. The sample (30 mg) was extracted using 450 μL of 70% methanol (v/v) spiked with an internal standard at 1 μg/mL. Subsequently, the mixture was subjected to vortexing for a duration of 1 minute, followed by ultrasonication for 30 min in water at a temperature of 10°C. Following a refrigerated period of 1 h at −20 °C, the material underwent centrifugation at 12000 rpm for 15 min at 4 °C. Ultimately, the liquid portion below the sediment was collected and transferred into a new 2 mL tube for LC-MS/MS examination.
Mass spectrometry analysis was conducted using the serum of rats. 500 μL of plasma sample was combined with 50 μL of hydrochloric acid (2 mol/L). The resulting mixture was subjected to vortexing for a duration of 1 min, after which it was incubated at a temperature of 4 °C for a period of 15 min. The process of vortexing and incubating was done four times. Introduce 2 mL of acetonitrile into the mixture, followed by vortexing for 5 min. The sample was then subjected to centrifugation at 12000 rpm for 5 min at a temperature of 4 °C. 1800 μL of the liquid portion was transferred to a new tube and dried using nitrogen. The desiccated specimens were reconstituted in a solution of 70% methyl alcohol with 1 μg/mL of the internal standard using a vortexing technique for a duration of 5 min. Subsequently, the constitution underwent centrifugation at a speed of 12000 revolutions per minute for 5 minutes at a temperature of 4 °C. The resulting liquid was then transferred to a new glass vial for LC/MS analysis.
LC-MS/MS Conditions
Samples were analyzed by LC-MS/MS on a Shimadzu Nexera LC-30A UHPLC platform fitted with a Waters UPLC BEH C18 column (1.7 μm 2.1*100 mm). The experimental procedure involved configuring the flow rate to 0.4 mL/min and the sample injection volume to 3 μL. The mobile phase was composed of a solution containing 0.1% formic acid in water (A) and in acetonitrile (B). The software for linear elution gradient consisted of multiple steps. The following time intervals are represented by the following values: 0–3.5 min, 95–85% A; 3.5–6 min, 85–70% A; 6–6.5, 70–70% A; 6.5–12 min, 70–30% A; 12–12.5 min, 30–30% A; 12.5–18 min, 30–40% A; 18–22 min, 0–0% A.
The MS and MS/MS data were obtained using an AB SCIEX Triple TOF 5600 mass spectrometer in conjunction with an Analyst TF 1.7 software, employing the IDA acquisition mode. Throughout each acquisition cycle, the molecular ions with a strength over 100 were subjected to screening, and subsequently, the associated MS/MS data were obtained. The temperature of the ESI heater was consistent at 550 °C, while the pressures of the nebulizer gas, auxiliary gas, and curtain gas were adjusted to 55 psi, 55 psi, and 35 psi, respectively. The collision energy was established at 40 V, while the energy spread was fixed at 20 V. In addition, the Ion Spray Voltage Floating feature was configured to operate at 5500 V in positive ion mode and −4000 V in negative mode.
Network Pharmacology
Screening of Potential Targets Between XF and SARS-CoV-2 and Construction of Networks and Pathway Analysis
The active constituents of XF were initially evaluated using LC-MS/MS. Subsequently, the corresponding chemical structures were identified by a search in the Pubchem database (https://pubchem.ncbi.nlm.nih.gov, accessed on May 20, 2026). Additionally, the relevant targets were identified through a screening process in the SwissTargetPrediction database (http://www.swisstargetprediction.ch/, accessed on May 20, 2026).
Additionally, it is imperative to gather the therapeutic targets of SARS-CoV-2. The identification of therapeutic targets was achieved through a comprehensive search of various databases, including GeneCards (https://www.genecards.org/, accessed on April 2, 2024), DrugBank (https://go.drugbank.com/, accessed on April 2, 2024), TTD (https://db.idrblab.net/ttd/, accessed on April 2, 2024), and CTD (http://ctdbase.org/, accessed on April 2, 2024), using the phrase “COVID-19”. Next, use the Oebiotech mapping platform (https://cloud.oebiotech.com/task/, accessed on April 2, 2024) to create a Venn diagram representing the relationship between disease and medicine.
Furthermore, the examination of the Protein Protein Interaction (PPI) network, Gene Ontology (GO), and Kyoto Encyclopedia of Genes and Genomes (KEGG) was being conducted. The target protein-protein interaction (PPI) network was constructed using the probable targets obtained from the STRING database (https://string-db.org/, accessed on April 2, 2024). The biological species used for this network was “Homo sapiens”, and a minimum confidence level of 0.900 was employed. The PPI network analysis was conducted using the Mcode plug-in in Cytoscape3.7.2 software (http://www.cytoscape.org/, accessed on April 2, 2024). In order to examine the potential molecular processes of XF in treating SARS-COV-2, we utilized the online Metascape platform (http://metascape.org/gp/index.html, accessed on April 2, 2024) to conduct GO and KEGG enrichment analysis. The screening condition was set at a P value cutoff of 0.05.
Finally, to gain a thorough understanding of the molecular mechanisms of XF in treating SARS-CoV-2, a diagram titled “active ingredient - hub target - core route” was created using Cytoscape software.
Molecular Docking
The 3D structures of the chemicals were acquired from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/, accessed on May 20, 2026). The 3D conformation of ACE2 was extracted from the ACE2–SARS-CoV-2 spike receptor-binding domain complex structure in RCSB (https://www.rcsb.org/, accessed on May 20, 2026), with the Protein Data Bank (PDB) identifier 6M0J. For the purpose of converting molecular structure files, the OpenBabel software (Version 3.1.1) was utilized. The AutoDock software (Version 1.5.6) streamlined multiple procedures, such as eliminating protein ligands and water molecules, hydrogenating, calculating charges, docking molecules, and processing the results. The complete ACE2 molecule was used as the docking range during molecular docking analysis, and the findings were shown using Pymol software (Version 2.4.0).
Cytotoxicity and Cytopathic Effect (CPE) Inhibition Assays
The cytotoxicity of XF against Vero E6 cells was assessed by determining the 50% cytotoxic concentration (CC50) using the MTT (3-(4,5-dimethylthazol-2-yl)-2,5-diphenyltetrazolium bromide) test. The cytotoxicity assay involved incubating various dilutions of XF with Vero E6 at a concentration of 5×104 cells per well in 96-well plates. Subsequently, the CC50 values of XF were determined.
The efficacy of XF against the Omicron strain of SARS-CoV-2 was investigated using the 50% inhibitory concentration (IC50) of virus-induced cytopathic effect (CPE). Briefly, a single layer of Vero E6 cells was exposed to 100 TCID50 of virus and maintained at 37°C for a 2-h adsorption period. Cells were subsequently exposed to varying doses of XF following inoculum removal. The assessment of CPE involved the observation of infected cells under a microscope following a 72-h incubation period. The IC50 value was computed, and the selectivity index (SI) was identified by dividing the CC50 by the IC50.
RNA Isolation and Reverse Transcriptase-Quantitative PCR Analysis (RT-qPCR)
The Calu-3 cells, which were cultivated in 12-well plates, were washed with PBS and subsequently exposed to the Omicron version of SARS-CoV-2 at a concentration of 100 TCID50 for a duration of 2 h. Subsequently, the inocula were extracted and the cells were segregated into four distinct groups: the control group (NC) treated with no treatment, the group infected with the virus (Virus), and two groups including nontoxic quantities of XF. Cells were collected 48 h after infection. The extraction of total RNA from various groups was performed using Trizol reagent (Invitrogen, Carlsbad, MA, USA). cDNA synthesis was conducted using the PrimeScript™ RT Master Mix kit (Takara Bio, Tokyo, Japan), followed by RT-qPCR with SYBR Premix Ex Taq™ II (Takara Bio) on an ABI PRISM7500 system (Applied Biosystems Co., Foster, CA, USA). Relative quantification was conducted according to the 2− ΔΔCt method described previously. Primer 5.0 was utilized to build the primers for TNF-α, IP-10, IL-6, MCP-1, and GAPDH (Table 1).
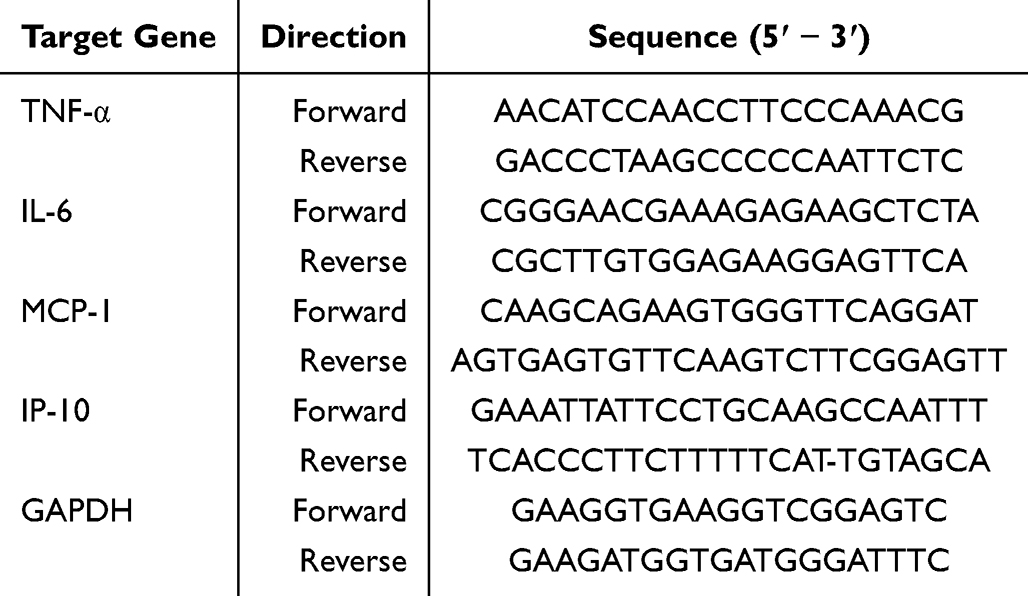 |
Table 1 Primer Sequence for RT-qPCR |
Western Blot Analysis
Calu-3 cells were treated as described in the “RNA Isolation and Reverse Transcriptase-Quantitative PCR (RT-qPCR) Analysis” section and subsequently harvested for Western blot analysis. Total protein was extracted in radioimmunoprecipitation assay (RIPA) buffer (Beyotime, China), quantified by bicinchoninic acid (BCA) assay kit (Beyotime, China, protein-grade purity), and 20 μg aliquots were subjected to 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) prior to transfer onto polyvinylidene fluoride (PVDF) membranes (Immobilon-PSQ, Ireland). The membranes were blocked with NcmBlot blocking buffer and incubated with specific primary antibodies (Table 2) overnight at 4°C, followed by incubation with species-specific secondary antibodies (Table 2) for 1 hour at room temperature. Immunodetection was performed using enhanced chemiluminescence (ECL) reagents (NCM Biotech, China).
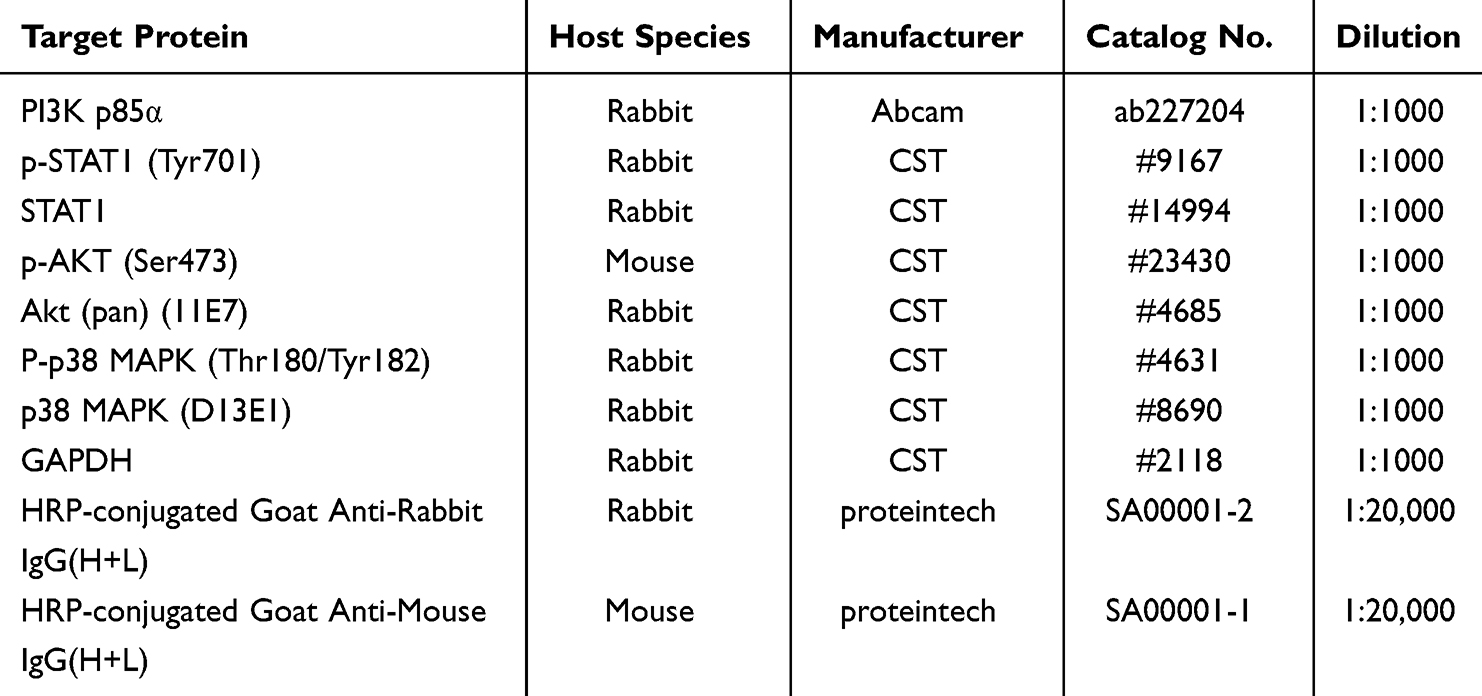 |
Table 2 Antibodies Used for Western Blot Analysis |
Death Protection and Viral Titer Assay of XF
Healthy C57BL/6 suckling mice, weighing 4–6 g, were randomly assigned to 5 groups within 10 days of age. The groups were as follows: control group (NC), HCoV-OC43 group (Virus), remdesivir group (RDV, 25 mg/kg), and Xiaoer Fengreqing group (XF) at doses of 2.9 and 11.6 mL/kg, respectively. Each group consisted of 6 suckling mice for tissue analysis. A 30 μL intracranial injection of HCoV-OC43 virus solution containing 1 LD50 was administered to the virus, RDV, and XF groups, whereas the normal group received an equivalent volume of PBS. A 12% sulfobutyl ether-β-cyclodextrin aqueous solution was used to solubilize RDV, which was subsequently adjusted to a pH of 5.0 using NaOH/HCl. After 2 h of the challenge, the XF groups were administered the corresponding XF dose by gavage, while the NC and Virus groups received deionized water. The RDV group, on the other hand, received a subcutaneous injection of RDV. On the fourth day, the mice were anesthetized by intraperitoneal injection of sodium pentobarbital (50 mg/kg), and then euthanized by cervical dislocation after deep anesthesia was confirmed. Subsequently, the brain tissues were extracted, and the brain index (measured as the ratio of brain weight to body weight) was determined. Subsequently, the left brains were homogenized with the addition of 1 mL of PBS (5000 × g, centrifuged at 4°C for 15 min). The liquid portion was utilized to measure the amount of virus present and the level of inflammatory factors being expressed. Once the right brains were immersed in neutral formalin, slices were prepared using H and E staining. For the survival analysis, a separate cohort of 8 suckling mice per group was used and monitored daily for 7 days post-infection.
Detection of the Expression of Inflammatory Factors by Using Bio-Plex Assay
The thin-bottom 96-well plate should be washed using 200 µL of a 1× wash solution each well. The washing process should be conducted at a speed of 300 rpm/min for a duration of 5 minutes over room temperature. Add 50 µL of the standard solution to the designated wells. 50 µL of detection buffer should be added to the background wells. Transfer 50 µL of the sample into the designated well. Incorporate 50 µL of 1° microspheres into each well. The cells should be incubated in a light-free environment at ambient temperature for a duration of 30 min, while employing a speed setting of approximately 850 ± 50 rpm/min. Utilize an automated washing machine to do three iterations of the washing process. Subsequently, distribute a cumulative volume of 25 µL of detection antibody into each well and subject them to incubation under conditions of darkness at ambient temperature for an additional duration of 30 min, employing a speed setting of approximately 850 ± 50 rpm/min. Utilize an automated washing machine to do three iterations of the washing process. Next, 50 µL of SA-PE solution was introduced into each well and allowed to incubate at room temperature in the absence of light for 15 minutes, with a rotational speed of 850 ± 50 rpm/min. To perform three washes, an automatic washing machine was utilized. Each well was treated with 125 µL of detection buffer, and the pellets were thoroughly resuspended prior to detection using Bio-Plex 200, with a density of 50 pellets per area. In the study of data quality control, the Bio-Plex Manager software was employed.
Observation of Pathological Changes
Following the fixation process, the brain tissues were dried off, embedded in paraffin, divided into sections, stained with hematoxylin and eosin (HE), sealed, and then analyzed for histopathological alterations using a light microscope.
Statistical Analysis
SPSS19.0 was utilized for data analysis. The mean and standard deviation (SD) values for brain index, virus titer, and inflammatory factor expression were reported. Group differences were examined using One-Way ANOVA. The Log-rank (Mantel-Cox) Test was employed to undertake survival curve analysis. A p-value below the threshold of 0.05 was deemed to possess statistical significance.
Results
Investigation of the Bioactive Ingredients of XF Through UHPLC-QTOF-MS Analysis
The XF was subjected to chromatographic analysis using UHPLC-QTOF-MS, as depicted in Figure 1A, Supplementary Figures S1 and S2. In the negative ion mode, a total of 99 compounds were discovered, whereas in the positive ion mode, 151 compounds were detected. A total of 220 compounds were obtained by combining the results of the two halves without any duplication. The substances under consideration could be classified into many groups, such as flavonoid glycosides, flavonoids, xanthones, fat acids, terpenoids, alkaloids, glycosides, phenylpropanoids, and coumarins. Detailed information on the identified compounds, including their classification, molecular weight, and retention time, is provided in Supplementary Table S1.
 |
Figure 1 Investigation of the bioactive ingredients and potential mechanism of XF for COVID-19 through UHPLC-QTOF-MS analysis. (A) The TIC diagram of XF sample1. (i) Negative ion mode. (ii) Positive ion mode; (B) The TIC diagram of serum sample1 (blank control). (i) Negative ion mode. (ii) Positive ion mode; (C) The TIC diagram of serum sample1 (drug treatment). (i) Negative ion mode. (ii) Positive ion mode; (D) The heat map of identified compounds from three XF samples. Red represents higher relative concentration and blue represents lower relative concentration. |
The chemical makeup of XF was effectively identified in the aforementioned data. The process of medication efficacy mostly relies on the absorption of TCM ingredients into the bloodstream. Consequently, rats were subjected to gavage administration of XF in order to investigate the chemicals that were absorbed into the bloodstream. Serum samples were obtained from rats in two groups: the blank control group and the drug administration group. The compound map was determined using UHPLC-QTOF-MS, as shown in Figure 1B, C and Supplementary Figures S3–S6. Based on the aforementioned procedure, a total of 90 compounds were identified in negative ion mode and 47 compounds were identified in positive ion mode, as per the selection criteria for chemicals taken into the blood. A total of 137 compounds were obtained by combining the results of the two portions, without any duplication. There were various distinct categories in which these molecules could be classified, including flavonoid glycosides, flavonoids, terpenoids, lipid acids, phenylpropanoids, and alkaloids. Detailed information on the plasma-absorbed compounds, including their classification, molecular weight, and retention time, is provided in Supplementary Table S2. Figure 1D utilized aheat map to visually represent the relative concentrations of the chemicals. The findings of the study indicated that the quantities of the identified chemicals in the three samples exhibited anotable degree of consistency, hence confirming the relative stability of each molecule in XF.
Network Pharmacology
Initially, a comprehensive investigation using UHPLC-QTOF-MS yielded a total of 137 bioactive constituents of XF. Subsequently, the chemical structures of these constituents revealed the presence of 774 matching targets, encompassing flavonoid glycosides, flavonoids, terpenoids, lipid acids, phenylpropanoids, and alkaloids. The term “COVID-19” was utilized to conduct searches across four databases. The GeneCards database yielded 2682 targets, the TTD database yielded 87 targets, the DrugBank database yielded 19 targets, and the CTD database yielded 4725 targets. The aforementioned targets were combined and subjected to deduplication, resulting in the acquisition of 6464 active targets. Through the utilization of Venn analysis, a total of 361 possible targets were identified between XF and SARS-CoV-2, as depicted in Figure 2A.
 |
Figure 2 Screening of potential targets between XF and SARS-CoV-2 and construction of networks and pathway analysis. (A) Venn diagram of Intersection Targets between XF and COVID-19; (B) Analysis results of PPI network and the core targets of the PPI network; (C) GO Biological Processes enrichment. (D) KEGG enrichment analysis; (E) “Active ingredients-potential targets-signaling pathways” Network of XF for treatment of SARS-CoV-2. (green: active ingredient’s degree ≥ 5; red: hub target’s degree ≥ 20); (F) 3D simulation diagrams of molecular docking between the six compounds with the highest binding affinities and the ACE2 chain extracted from 6M0J. The compounds are shown in yellow or blue according to the background color of the interaction site: (i) Isoalantolactone; (ii) Isosakuranetin; (iii) 3,6,3′-Trihydroxyflavone; (iv) Dihydronorwogonin; (v) Luteolin; and (vi) Baicalein; (G) Representative hydrogen-bonding interactions between selected top-ranked compounds and ACE2. The compounds are shown in red, hydrogen bonds are shown as cyan dashed lines, and the interacting amino acid residues are shown in yellow: (i) Isoalantolactone interacted with LYS562 and ASN394 of ACE2 through hydrogen bonds; (ii) Isosakuranetin interacted with LYS562 and LEU391 of ACE2 through hydrogen bonds. |
Furthermore, the PPI network was created by identifying shared potential targets of XF and SARS-CoV-2, with a minimum confidence level of 0.900. The Mcode plug-in was utilized to examine the PPI network and identify specific clusters (with a minimum score of 11.091), such as ACE2, EGFR, PARP1, JAK1, SRC, and others (Figure 2B).
Thirdly, the Gene Ontology (GO) annotations consisted of 361 predicted targets, which were classified into three categories: biological processes (2817 terms), molecular activities (268 terms), and cellular components (203 terms). The top 5 GO keywords were presented as bar charts (Figure 2C) based on the Log P value. The genes associated with Biological Processes exhibited a notable enrichment in phosphorylation and cellular response to nitrogen compounds. Similarly, the genes related to Cellular Components displayed a clear enrichment in membrane raft and membrane raft function. On the other hand, the genes associated with Molecular Functions displayed a distinct enrichment in phosphotransferase activity and protein kinase activity, among others. The KEGG pathway analysis yielded a total of 242 enrichment results. The KEGG database displayed 14 pathways associated with COVID-19 in the form of a bubble diagram (Figure 2D). The signaling pathways primarily implicated in this study include the PI3K-Akt signaling pathway, JAK-STAT signaling system, MAPK signaling pathway, NF-κB signaling circuit, among others. These findings suggested that the efficacious constituents of XF might have the potential to act against SARS-CoV-2 by modulating these processes.
Furthermore, the network diagram illustrating the relationship between the active component, hub target, and core route was created using CytoScape3.7.2. According to the plug-in Network analyzer, a total of 18 active substances were identified, namely luteolin, tangeritin, and Saikosaponin D. The network structure, depicted in Figure 2E, consisted of 52 nodes and 251 edges, with 18 hub targets and 14 core pathways.
Finally, in accordance with the aforementioned description, the chemicals that were consumed by XF were assessed. The 3D structures of 111 out of the 137 compounds that were identified were acquired from the PubChem database, while the ACE2 chain extracted from the ACE2–SARS-CoV-2 spike receptor-binding domain complex structure 6M0J was used for molecular docking. The link between these chemicals and the ACE2 protein was investigated using molecular docking technology in order to examine the compound foundation of XF for its anti-COVID-19 action. The binding energies of all compounds are provided in Supplementary Table S3. Figure 2F presents the outcomes of molecular docking for the six most promising molecules. The binding energies of Isoalantolactone, Isosakuranetin, 3,6,3′-Trihydroxyflavone, Dihydronorwogonin, Luteolin, and Baicalein were −6.45, −6.05, −6.04, −5.99, −5.97, and −5.81 kcal/mol, respectively. Representative hydrogen-bonding interactions of Isoalantolactone and Isosakuranetin with ACE2 are shown in Figure 2G. Isoalantolactone formed hydrogen bonds with LYS562 and ASN394, while Isosakuranetin formed hydrogen bonds with LYS562 and LEU391. This finding suggests that these compounds can form stable binding interactions with ACE2, hence highlighting their potential involvement in the anti-COVID-19 mechanism of XF.
Antiviral and Anti-Inflammatory Activities of XF on SARS-CoV-2 in vitro
The cytotoxic effects of XF on Vero E6 cells were assessed using a non-radioactive cell proliferation test (MTT). A CC50 value of 40.14 mg/mL was observed for XF in Vero E6 cells, as depicted in Figure 3A. To assess the antiviral properties of XF against the Omicron form of SARS-CoV-2, a CPE inhibition experiment was employed. The administration of XF at concentrations of 12.5 mg/mL, 6.25 mg/mL, and 3.125 mg/mL resulted in a considerable reduction in the CPE generated by infection in Vero E6 cells. The IC50 value of XF was determined to be 5.687 mg/mL, and the selectivity index (SI) of XF was found to be 7.06 (Figure 3B). The results demonstrated that XF exhibited antiviral activity against SARS-CoV-2 in vitro.
 |
Figure 3 Antiviral and anti-inflammatory activities of XF on SARS-CoV-2 in vitro. (A) The effect of XF on the survival rate of Vero E6 cells; (B) The antiviral effect of XF against SARS-CoV-2; (C–F) The results of XF induced inflammatory cytokines (IL-6 (C), IP-10 (D), MCP-1 (E), TNF-α (F)) in SARS-CoV-2 infected cells. Values represent the mean ± S.D. of three individual experiments. Comparisons with the viral control are denoted by **p < 0.01; ***p < 0.001. |
The infection caused by SARS-CoV-2 commonly triggers an inflammatory response and the generation of cytokines. Consequently, the levels of mRNA expression were assessed by RT-qPCR in vitro. According to the data presented in Figure 3C–F, the mRNA expression levels of TNF-α, IL-6, IP-10, and MCP-1 were considerably increased (p < 0.001 or p < 0.01) in the virus group following SARS-CoV-2 infection, compared to the NC group. Administration of XF at concentrations of 12.5 mg/mL and 6.25 mg/mL effectively suppressed this rise in a manner that was dependent on the dosage (p < 0.001 or p <0.01), indicating that XF has the potential as an anti-inflammatory agent.
Inhibitory Effect of XF on Lethality and Inflammation of Brain Tissues in HCoV-OC43 Infected Suckling Mice
The antiviral activity of XF was assessed by detecting the survival of mice that were fatally infected with HCoV-OC43. The survival of mice was depicted in Figure 4A following a period of 7 days of daily observation. In the virus group, all infected mice succumbed by day 6. However, the administration of XF (2.9 mL/kg) (p < 0.05) and RDV (p < 0.01) resulted in a considerable extension of their survival duration as compared to the viral control group.
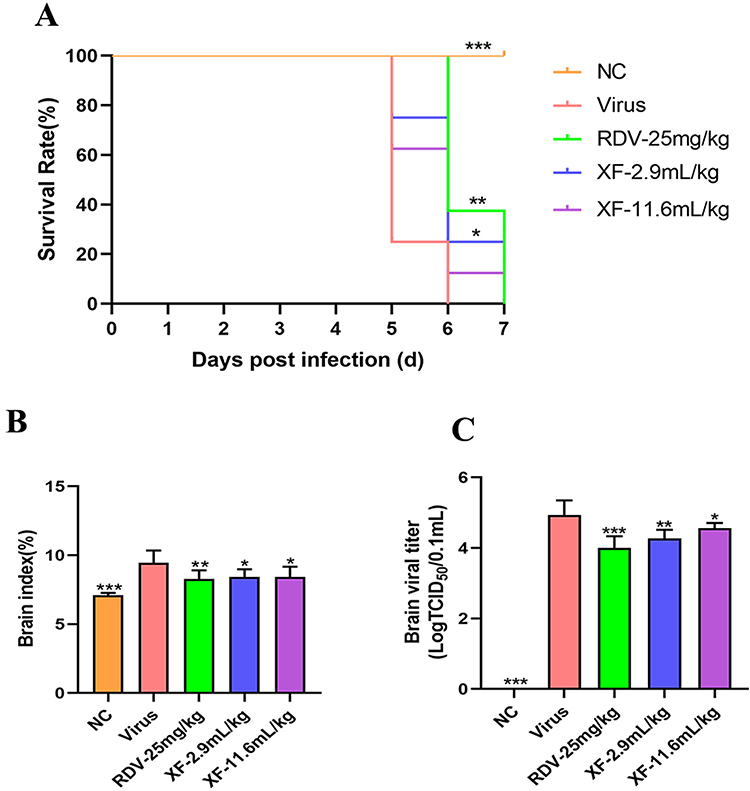 |
Figure 4 XF could reduce brain injury and inflammation in suckling mice infected with HCoV-OC43. (A) The survival of infected mice was recorded daily for 7 days, and the corresponding survival rate was calculated; (B) The inhibitory effect of XF on index in brain tissue of mice. (C) The inhibitory effect of XF on virus titer in brain tissue of mice. Eight suckling mice per group were used for survival analysis, and six suckling mice per group were used for tissue-based analyses. Comparisons with the viral control are denoted by *p < 0.05; **p < 0.01; ***p < 0.001. |
In this work, the brain index was computed to evaluate the level of viral inflammation caused by HCoV-OC43. In Figure 4B, it could be observed that the brain index of the virus group was 9.44 ± 0.90, a considerably greater value compared to the normal control group (p < 0.001). Nevertheless, the brain index significantly decreased after XF or RDV treatment, but there was no significant difference between the XF groups. Viral titers are indicative of the quantities of virus required to induce infection, as well as the capacity to withstand viral pathogens, rather than the capacity for survival. Consequently, the levels of brain virus titers were assessed across several groups. There was a significant drop in brain virus titers observed in the XF (2.9 mL/kg and 11.6 mL/kg) and RDV groups as compared to the viral group, as depicted in Figure 4C. The virus group exhibited a brain virus titer of 4.93 ± 0.42, significantly higher than that of the drug group. The findings of this study suggested that XF effectively suppressed both viral titers and virus-induced inflammation.
XF Inhibited Virus-Induced Expression of Inflammatory Cytokines in Brain Tissues in vivo
HCoV-OC43 infection significantly increased the protein levels of IFN-γ, IP-10, TNF-α, and IL-6 in brain tissues compared with the NC group (p < 0.001). The XF intervention had a substantial impact on the levels of IFN-γ (2.9 mL/kg and 11.6 mL/kg, p < 0.001) (Figure 5A), IP-10 (2.9 mL/kg, p < 0.001; 11.6 mL/kg, p < 0.01) (Figure 5B), TNF-α (2.9 and 11.6 mL/kg, p < 0.001) (Figure 5C), and IL-6 (2.9 mL/kg, p < 0.05; 11.6 mL/kg, p < 0.01) (Figure 5D). The findings of the study revealed that the administration of XF resulted in a noteworthy suppression of the expression of pro-inflammatory factors. We further validated the anti-neuroinflammatory effect of XF in HMC3 cells. XF significantly suppressed virus-induced upregulation of IL-6, TNF-α, IP-10, MCP-1, and IFN-γ (Supplementary Figures S7).
 |
Figure 5 XF inhibited virus-induced expression of inflammatory cytokines in brain in vivo. (A) IFN-γ protein in the brain of the infected mice were assessed using bio-plex analysis; (B) IP-10 protein in the brain of the infected mice were assessed using bio-plex analysis; (C) TNF-α protein in the brain of the infected mice were assessed using bio-plex analysis; (D) IL-6 protein in the brain of the infected mice were assessed using bio-plex analysis; The values are presented as the means ± S.D. of six individual experiments. *p < 0.05; **p < 0.01; ***p < 0.001, when compared to the viral control. |
Effects of XF on the Pathological Changes of Brain Tissue Induced by HCoV-OC43 Infection in Suckling Mice
Additional tests were conducted to investigate the pathological alterations in the brains that were infected, as depicted in Figure 6. The histology of the uninfected mice’s brains in the NC group exhibited no apparent alterations. Nevertheless, mice infected with HCoV-OC43 exhibited brain cell pyknosis and necrosis, with neutrophils and lymphocytes being the predominant inflammatory infiltrate. The administration of high-dose XF (11.6 mL/kg) resulted in marked amelioration of brain damage. Low-dose XF (2.9 mL/kg) also showed partial histological improvement compared with the infected group, although the effect was less pronounced than that observed in the high-dose XF group. RDV (25 mg/kg) showed limited histological improvement under the present experimental conditions. The pathological findings demonstrated that XF alleviated HCoV-OC43-induced brain cell necrosis and inflammatory infiltration, with more evident protection observed in the high-dose XF group.
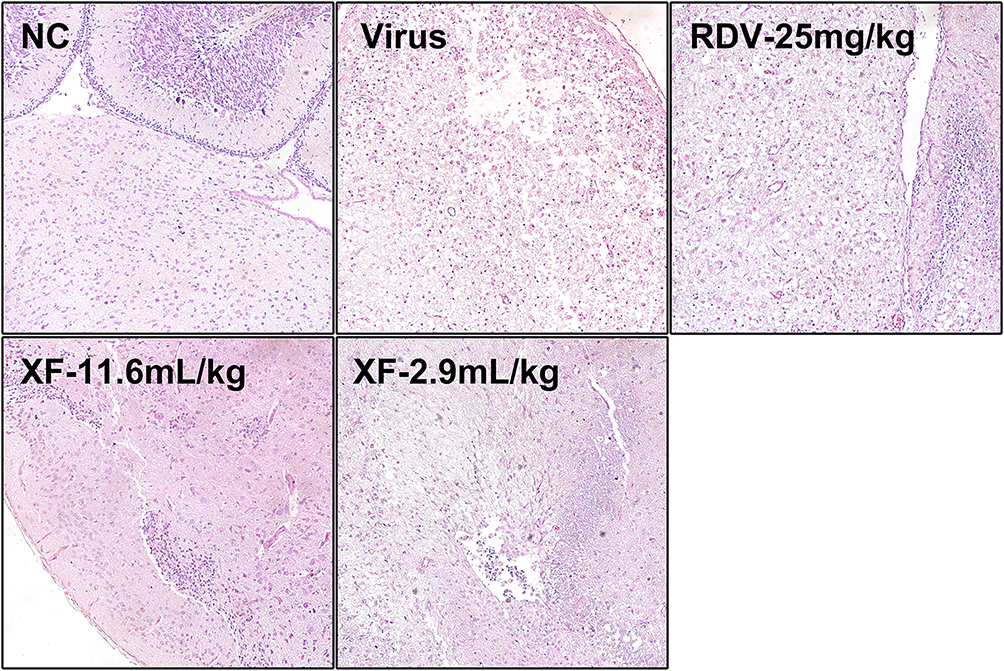 |
Figure 6 Effects of XF on the pathological changes of brain tissue induced by HCoV-OC43 infection in suckling mice. Histological observations of brain tissues for mice sacrificed at the 4th d.p.i. Scale bar =100 μm. |
Potential Anti-Inflammatory and Antiviral Mechanism of XF Against SARS-CoV-2
Based on KEGG pathway enrichment analysis, the compound Chinese medicine XF was predicted to target PI3K-Akt, JAK-STAT, and MAPK signaling pathways. Experimental validation through Western blot analysis (Supplementary Figures S8–S15) demonstrated, the protein levels of p-p38 MAPK (p < 0.001), p-STAT1 (p < 0.001), and p-AKT (p < 0.001) increased in the viral infection group compared to normal controls (NC). Subsequent XF treatment was found to significantly reduce the protein expression level of p-STAT1 (p < 0.001), p-p38 MAPK (p < 0.001), and p-AKT (p < 0.001). Among different groups, there was no significant difference in the total amounts of PI3K p85α STAT1, AKT and P38 MAPK proteins. (Figure 7).
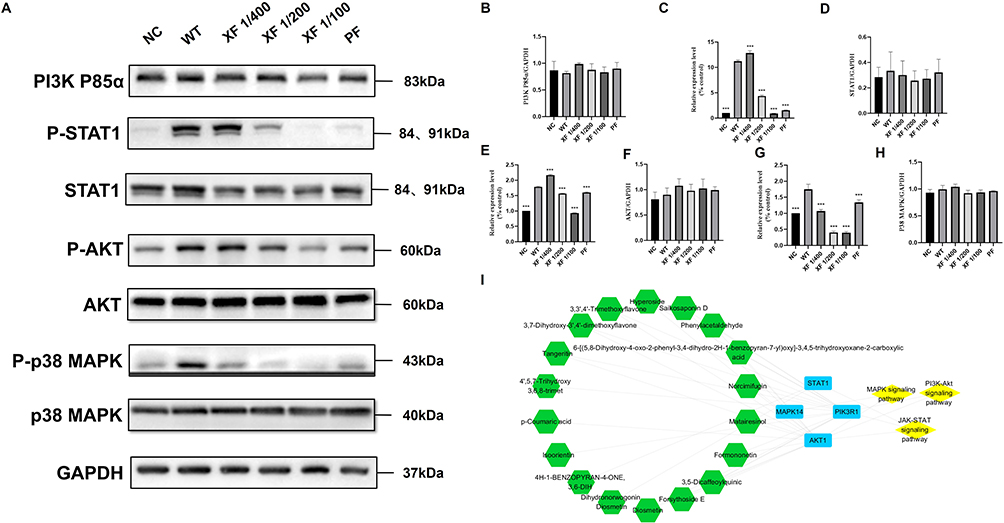 |
Figure 7 XF reverses coronavirus-induced hyperphosphorylation through selective pathway modulation. (A) The expression of the p-STAT1, p-AKT and p-p38 MAPK, PI3K p85, STAT1, AKT, p38 MAPK and GAPDH proteins in the Calu-3 cells was detected by Western blot analysis; (B–H) The quantitative analysis of the PI3K p85 (B), p-STAT1 (C), STAT1 (D), p-AKT (E), AKT (F), p-p38 MAPK (G) and p38 MAPK (H) proteins was analyzed by Image J. (I) Key Constituents-Core Targets-Pathways Network Pharmacology Diagram. Values represent the mean ± S.D. of three individual experiments. Comparisons with the viral control are denoted by ***p < 0.001. |
The primary compounds regulating signaling biomarkers included Diosmetin, 3,7-Dihydroxy-3′,4′-dimethoxyflavone, 3,3′,4′-Trimethoxyflavone, Hyperoside, Saikosaponin D, Phenylacetaldehyde, Tangeritin, Norcimifugin, Matairesinol, Formononetin, 3,5-Dicaffeoylquinic Acid, Forsythoside E, and 6-[(5,8-Dihydroxy-4-oxo-2-phenyl-3,4-dihydro-2H-1-benzopyran-7-yl)oxy]-3,4,5-trihydroxyoxane-2-carboxylic Acid in total. They exert anti-inflammatory and antiviral effects by regulating the key proteins STAT1, AKT and P38 in the PI3K AKT signaling pathway, JAK-STAT signaling pathway and MAPK signaling pathway respectively.
Discussion
In the present study, we investigated the antiviral and anti-neuroinflammatory effects of Xiaoer Fengreqing (XF) against coronavirus-associated encephalitis through an integrated analytical, in vitro, and in vivo framework. Our findings suggest that XF may downregulate key inflammation-related signaling pathways, partly supporting the network pharmacology predictions. By focusing on neurological manifestations rather than typical pulmonary injuries, this study provides experimental evidence for further investigation of XF in coronavirus-associated neuroinflammation. The scientific community faces limited accessibility for research purposes due to the categorization of SARS-CoV-2 as a biosafety level 3 (BSL-3) infection, primarily due to its relatively high death rate. The development of effective therapies against COVID-19 has been significantly hindered by this impediment.27 In relation to this matter, there has been a resurgence of interest in human coronavirus OC43 (HCoV-OC43) as a compelling and secure model for investigating coronaviruses inside low-risk research environments at BSL-2.28 In addition, HCoV-OC43 is a human betacoronavirus related to SARS-CoV-2. However, it can be studied under “routine” biosafety conditions. Furthermore, it has been demonstrated to be a significant pathogen in mild upper respiratory tract infections in children. Additionally, it can infect suckling mice, leading to neurotropic lethality and encephalitis-like manifestations.29 Therefore, the HCoV-OC43-infected suckling mouse model was selected for in vivo pharmacological evaluation. The in vivo tests were done to investigate the antiviral and anti-inflammatory effects of XF against coronavirus HCoV-OC43 infection, as well as its protective efficiency against mortality. The study findings indicated that the administration of XF resulted in a notable extension of the survival duration in the mice who were infected, as well as a decrease in the levels of the virus in the brain tissue of the mice that were fed. Furthermore, in our in vivo histopathological assessments, high-dose XF (11.6 mL/kg) showed the most evident improvement in brain injury, while low-dose XF (2.9 mL/kg) showed partial histological improvement. RDV showed limited histological protection under the present experimental conditions. This finding may be related to the severity and rapid progression of the intracranial HCoV-OC43 neonatal infection model, as well as the potentially limited CNS exposure to RDV and its metabolites due to blood-brain barrier constraints.30 Further pharmacokinetic studies are needed to clarify the distribution of RDV and XF-derived active components in brain tissue and to optimize the dosing regimen before clinical translation. We also observed that the brain index in the XF-treated groups was significantly different from the virus group, but no dose-dependent effect was detected. Since this measure is based solely on brain weight/body weight, it may not fully reflect changes in viral replication, inflammatory responses, or histopathological injury. Therefore, the similar brain index values observed in the two XF-treated groups do not exclude dose-related differences in other outcomes.26 XF exhibited a potential antiviral impact against typical coronavirus infections and demonstrated anti-inflammatory properties against encephalitis resulting from coronavirus infection. These findings suggest that XF may exert antiviral and anti-inflammatory effects through multiple targets and pathways in the experimental models used in this study. In addition, in vitro studies showed that XF inhibited infection by the SARS-CoV-2 Omicron variant, although its selectivity index (SI = 7.06) was below the commonly referenced threshold of 10, and the IC50 was relatively high (5.687 mg/mL). However, the SI benchmark should be regarded as a practical guide rather than an absolute criterion, as compounds with SI values below 10 may occasionally remain effective in animal infection models.31 Some clinically used traditional Chinese medicine formulas have also shown relatively modest SI values in experimental antiviral models. For example, Lianhuaqingwen capsule showed an SI value of 6.21 against the A/PR/8/34 (H1N1) influenza strain,32 while Liu Shen Wan showed an SI value of 5.21 against influenza A/PR/8/34 virus in an MDCK cell model.33 Lianhuaqingke also showed a relatively narrow in vitro antiviral window against SARS-CoV-2, with a TC50 of 972.2 μg/mL and an IC50 of 684.2 μg/mL; the authors reported an SI greater than 1.34 These findings suggest that the potential effects of complex traditional Chinese medicine formulas may not depend solely on direct viral inhibition, but may also involve anti-inflammatory and immunomodulatory mechanisms. Nevertheless, whether an effective in vivo concentration for the anti-SARS-CoV-2 activity of XF can be achieved remains unclear and requires further pharmacokinetic investigation.
A significant mechanism behind the development of nervous system disorders resulting from COVID-19 infection involves the unregulated growth and release of cytokines. Following SARS-CoV-2 infection, substantial alterations occur in host inflammatory signaling pathways, characterized by the activation and intricate crosstalk of multiple cascades that critically govern disease progression. The JAK/STAT, PI3K-Akt, and MAPK pathways play central roles in driving pathological hyperinflammation. The JAK/STAT phosphorylation cascade were activated by sustained high level of IL-6, leading to the excessive release of pro-inflammatory cytokines such as IL-1β, TNF-α.35 PI3K-Akt signaling is hyperactivated to facilitate viral entry via ACE2 endocytosis while amplifying mTORC1-mediated viral protein synthesis and proinflammatory cytokine production.36 The MAPK cascade, particularly p38 MAPK, synergizes with NF-κB through reciprocal phosphorylation events (eg, IKKβ/RelA), enhancing transcriptional upregulation of IL-6, TNF-α, and IL-1β.37 These pathways collectively establish a self-reinforcing network, where PI3K-Akt potentiates viral replication, JAK/STAT imbalance disrupts immune homeostasis, and MAPK/NF-κB crosstalk exacerbates cytokine storm severity. Meanwhile, the cytokine storm continues to exacerbate tissue damage through a positive feedback loop that changes the permeability of the endothelium, causes dysfunction in the blood-brain barrier, and ultimately activates microglia (macrophages that reside in the brain). As a result, macroglia experience secondary inflammation, which can lead to the onset of neurological disorders.38 The study found that XF effectively suppressed the increased levels of inflammatory factors caused by viral infection, both in vivo and vitro. In particular, the anti-inflammatory effect of XF was further confirmed in virus-infected HMC3 microglial cells, where XF markedly reduced the expression of IL-6, TNF-α, IP-10, MCP-1 and IFN-γ, providing additional evidence for its ability to modulate coronavirus-associated neuroinflammation. As described in the Methods section, Vero E6 cells were used for antiviral screening, whereas Calu-3 cells were used to assess inflammatory responses and signaling regulation because of their different biological suitability for these assays. Based on the KEGG pathway enrichment results of XF, we also focused on the PI3K/AKT, STAT, and p38 MAPK signaling pathways. Further Western blot (WB) results revealed that revealed that XF also decreased the expression of signaling biomarkers, including p-p38 MAPK, p-STAT1 and p-AKT, following viral infection, Thus, it may be deduced that XF exerted its anti-SARS-CoV-2 impact by suppressing the excessive production of inflammatory factors through down-regulation of PI3K/AKT, STAT and p38 MAPK signaling.
Subsequently, through network pharmacology analysis, the compounds that regulate these key pathways were further identified as Flavonoids, Flavonol glycosides, Triterpene saponin, Triterpene saponin Aldehyde, Volatile oils, Lignan, Phenylpropanoid. These might be the key compounds of XF in resisting COVID-19 (Figure 7). Studies have shown that most flavonoids, such as hyperoside, diosmetin, tangeritin and formononetin, can regulate and suppress Akt activation. Notably, 3,3′,4′-trimethoxyflavone additionally regulates the MAPK signaling pathway.39 Phenylacetaldehyde, a bioactive aldehyde, exerts anti-inflammatory effects by modulating the PI3K-AKT signaling pathway.40 Norcimifugin (a volatile oil derivative) and matairesinol (a lignan) both attenuate inflammation through inhibition of the PI3K-AKT signaling pathway; additionally, matairesinol exhibits dual regulatory activity by modulating the MAPK signaling cascade.41,42 Phenylpropanoid compounds, such as 3,5-dicaffeoylquinic acid and Forsythoside E, exert anti-inflammatory effects by suppressing TNFα-induced AKT phosphorylation.43
The ACE2 receptor, a carboxypeptidase, has been recognized as a crucial functional receptor inside the COVID-19 host. The invasion of SARS-CoV-2 into the brain is significantly influenced by its role. According to reports, there is a high expression of ACE2 in both olfactory epithelium cells and brain capillary endothelial cells. One potential mechanism by which SARS-CoV-2 can disseminate is through transsynaptic transfer, allowing it to reach the olfactory bulb and the central nervous system (CNS). Conversely, another mechanism involves the entry of SARS-CoV-2 into the brain through capillary endothelial cells. After excessive production of the virus, the endothelium becomes compromised, allowing the virus to traverse the underlying smooth muscle cells and ultimately infiltrate astrocytes, microglia, and neurons that also exhibit ACE2 expression.44 Molecular docking technology enables the simulation of interactions between ACE2 and other compounds in three-dimensional space, facilitating the identification of compounds capable of stable ACE2 binding as potential active pharmaceutical components. In the revised molecular docking analysis using the ACE2 chain extracted from 6M0J, Isoalantolactone, Isosakuranetin, 3,6,3′-Trihydroxyflavone, Dihydronorwogonin, Luteolin, and Baicalein showed the highest binding affinities among the candidate compounds. Furthermore, research has demonstrated that the administration of Isoalantolactone effectively inhibited the secretion of pro-inflammatory substances, including IL-6, IL-1β, and TNF-α, in the midbrain tissue.45 Despite the expanding pharmacological exploration of bioactive compounds against COVID-19, the antiviral efficacy of aldehyde derivatives and lignan-class phytochemicals against SARS-CoV-2 infection remains systematically underexplored. This knowledge gap underscores their untapped therapeutic potential, particularly given their documented anti-inflammatory properties in other viral pathologies.
Despite the promising findings, this study has several limitations that warrant consideration. First, as a traditional Chinese medicine, XF is composed of multiple herbs, and the inherent uncertainty and variability of its active components—influenced by factors such as geographic origin, harvest time, and preparation methods—may pose challenges to the reproducibility and translational applicability of these findings. To partially mitigate this, we utilized UHPLC-QTOF-MS to profile the chemical constituents; however, ensuring batch-to-batch consistency remains a long-term challenge. Second, the active components of XF were not separated and evaluated individually, meaning potential synergistic or antagonistic effects remain uncharacterized. Third, while molecular docking and pathway prediction provide mechanistic insights, further verification with gene knockdown or pathway inhibition experiments is needed to confirm causal relationships. Furthermore, the relatively modest selectivity index (SI = 7.06) and the relatively high IC50 value (5.687 mg/mL) observed in vitro raise important considerations regarding the clinical achievability of effective antiviral doses in vivo. Finally, the clinical relevance of the identified anti-inflammatory and antiviral mechanisms requires further validation in human pediatric populations through rigorous clinical trials. Given the complete absence of human clinical data and pediatric safety trials at this stage, XF should be qualified not as a definitive treatment, but rather as a potential complementary agent worthy of further investigation.
Conclusion
In brief, XF demonstrated notable antiviral and anti-inflammatory properties in vitro and in vivo. The network pharmacology and molecular docking analyses suggested that several XF-derived compounds may be involved in the regulation of ACE2-related and inflammation-related pathways. The main approach is to inhibit excessive inflammatory responses by regulating ACE2, AKT, STAT1, MAPK14 and their corresponding core signaling pathways, namely the PI3K-Akt signaling pathway, JAK-STAT signaling pathway and MAPK signaling pathway. These findings suggest that XF may inhibit excessive inflammatory responses through multiple components, targets, and pathways. Given the preliminary nature of the present findings, XF should be regarded as a potential complementary agent worthy of further investigation for coronavirus-associated neuroinflammation, rather than a confirmed treatment for pediatric COVID-19.
Data Sharing Statement
The data generated or analyzed during this study are not publicly available due to institutional policies and data confidentiality. However, they are available from the corresponding author, Qinhai Ma, upon reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by National Administration of Traditional Chinese Medicine (ZYYCXTD-D-202406); Natural Science Foundation of China (82474155); the Science and Technology Development Fund of Macau SAR (Grant FDCT 0002/2024/RDP); Special Research Project on Traditional Chinese Medicine Science and Technology of the National Administration of Traditional Chinese Medicine (GZY-KJS-2025-053); Guangdong Basic and Applied Basic Research Foundation (2023A1515140140); Science and Technology Project of Haihe Laboratory of Modern Chinese Medicine (25HHZYSS00004); Yangcheng Traditional Chinese Medicine Innovative Talent Team Project (2026RC010); Science and Technology Joint Development Project of the Department of Science and Technology, National Administration of Traditional Chinese Medicine (GZY-KJS-GD-2025-077). The funding agencies had no role in the study design, data collection, data analysis, interpretation of results, manuscript preparation, or decision to submit the manuscript for publication.
Disclosure
Menglei Wang and Yan Liu are affiliated with the Hebei Province (Handan) Chinese Medicine Industry Technology Research Institute. The authors declare that they have no financial ties to the manufacturer of XF. The authors report no other conflicts of interest in this work.
References
1. Kannan S, Shaik Syed Ali Pakeer P, Sheeza Ali A, Hemalatha K. Reply letter—COVID-19 (novel coronavirus 2019)—recent trends. Eur Rev Med Pharmacol Sci. 2020;24:6482–21. doi:10.26355/eurrev_202006_21629
2. Grainger R, Kim AHJ, Conway R, Yazdany J, Robinson PC. COVID-19 in people with rheumatic diseases: risks, outcomes, treatment considerations. Nat Rev Rheumatol. 2022;18:191–204. doi:10.1038/s41584-022-00755-x
3. Oberg C, Hodges HR, Gander S, Nathawad R, Cutts D. The impact of COVID-19 on children’s lives in the United States: amplified inequities and a just path to recovery. Curr Probl Pediatr Adolesc Health Care. 2022;52(7):101181. doi:10.1016/j.cppeds.2022.101181
4. Children’s Hospital Association. The facts about COVID-19 and children. Available from: https://www.childrenshospitals.org/content/public-policy/fact-sheet/the-facts-about-covid-19-and-children.
5. Molloy EJ, Nakra N, Gale C, Dimitriades VR, Lakshminrusimha S. Multisystem inflammatory syndrome in children and neonates associated with COVID-19: optimizing definition and management. Pediatr Res. 2023;93:1499–1508. doi:10.1038/s41390-022-02263-w
6. Stafstrom CE. Neurological effects of COVID-19 in infants and children. Dev Med Child Neurol. 2022;64:818–829. doi:10.1111/dmcn.15185
7. Cao L, Wu X, Zhang C, et al. Short-term post-COVID-19 symptoms in 21,012 patients: a cross-sectional study. Acupunct Herb Med. 2025;5(3):291–300. doi:10.1097/HM9.0000000000000166
8. Antoon JW, Hall M, Howard LM, et al. COVID-19 and acute neurologic complications in children. Pediatrics. 2022:150e2022058167. doi:10.1542/peds.2022-058167
9. Esposito S, Autore G, Argentiero A, Ramundo G, Perrone S, Principi N. Update on COVID-19 therapy in pediatric age. Pharmaceuticals. 2022;15:1512. doi:10.3390/ph15121512
10. Chen X, Wang M. Traditional Chinese medicine during the COVID-19 pandemic: recent successes and future perspectives. Acupunct Herb Med. 2023;3:357–359. doi:10.1097/HM9.0000000000000084
11. Ma Q, Lei B, Chen R, et al. Liushen capsules, a promising clinical candidate for COVID-19, alleviates SARS-CoV-2-induced pulmonary in vivo and inhibits the proliferation of the variant virus strains in vitro. Chin Med. 2022;17(1):40. doi:10.1186/s13020-022-00598-4
12. Ma Q, Wang Z, Chen R, et al. Effect of Jinzhen granule on two coronaviruses: the novel SARS-CoV-2 and the HCoV-229E and the evidences for their mechanisms of action. Phytomedicine. 2022;95:153874. doi:10.1016/j.phymed.2021.153874
13. Duan N, Liu B, Li X, et al. Traditional Chinese medicine in treating children with coronavirus disease 2019: a scoping review. Front Pediatr. 2022;10:935551. doi:10.3389/fped.2022.935551
14. Fang F, Yang L, Qin SC, et al. Clinical efficacy of traditional Chinese medicine Lianhua Qingwen Granules in 42 suspected cases of children with coronavirus disease 2019. Chin J New Drugs. 2020;29(24):2809–2812. [In Chinese].
15. Zhan H, Bai SX.Clinical characteristics and diagnosis and treatment of coronavirus disease 2019 in children. J Hubei Univ Med. 2020;39(3):26–30. [In Chinese].
16. Zhao Q, Zhao R, Geng Z, et al. Xuanfei Baidu granule alleviates coronavirus-induced pneumonia in low-temperature and high-humidity environments. Acupunct Herb Med. 2023;3:200. doi:10.1097/HM9.0000000000000068
17. Ma Q, Xie P, Zhan Y, et al. Elucidating forsythin’s anti-inflammatory action through modulation of the p38 MAPK pathway in SARS-CoV-2 infection. Engineering. 2025;54:187–201. doi:10.1016/j.eng.2025.01.020
18. Zhang B, Dong E, Liu L, et al. Ten recommendations for the high-quality development of traditional Chinese medicine in the new era. Acupunct Herb Med. 2025;5:131. doi:10.1097/HM9.0000000000000162
19. Zhang J, Chen X, Huang L, et al. Traditional Chinese medicine + artificial intelligence: Wuzhen consensus. Acupunct Herb Med. 2025;5:134–135. doi:10.1097/HM9.0000000000000163
20. Wang XF, Shang YX. Expert consensus in integrated traditional Chinese and western medicine prevention and treatment for influenza in children. Chin Pediatr Integr Tradit Western Med. 2021;13(5):369–374. [In Chinese]. doi:10.3969/j.issn.1674-3865.2021.05.001
21. Sun JH, Hao LY, Li ZY, et al. Effect of Xiaoer Fengreqing Mixture (Oral Liquid) on viral pneumonia mice model infected with influenza virus. Chin Tradit Herb Drugs. 2022;53(9):2740–2746. [In Chinese]. doi:10.7501/j.issn.0253-2670.2022.09.017
22. Zhao Q, Bucchi A, Duan X, et al. Traditional Chinese medicine for long COVID-19: the potential roles of immunity remodeling. Acupunct Herb Med. 2025;5(4):400–419. doi:10.1097/HM9.0000000000000175
23. Ogando NS, Dalebout TJ, Zevenhoven-Dobbe JC, et al. SARS-coronavirus-2 replication in VeroE6cells: replication kinetics, rapid adaptation and cytopathology. J Gen Virol. 2020;101(9):925–940. doi:10.1099/jgv.0.001453
24. Zaliani A, Vangeel L, Reinshagen J, et al. Cytopathic SARS-CoV-2 screening on VERO-E6cells in a large-scale repurposing effort. Sci Data. 2022;9:405. doi:10.1038/s41597-022-01532-x
25. Biering SB, Sarnik SA, Wang E, et al. Genome-wide bidirectional CRISPR screens identify mucins as host factors modulating SARS-CoV-2 infection. Nat Genet. 2022;54(8):1078–1089. doi:10.1038/s41588-022-01131-x
26. Xie P, Fang Y, Baloch Z, et al. A mouse-adapted model of HCoV-OC43 and its usage to the evaluation of antiviral drugs. Front Microbiol. 2022;13:845269. doi:10.3389/fmicb.2022.845269
27. Rehman SU, Rehman SU, Yoo HH. COVID-19 challenges and its therapeutics. Biomed Pharmacother. 2021;142:112015. doi:10.1016/j.biopha.2021.112015
28. Kim MI, Lee C. Human coronavirus OC43 as a low-risk model to study COVID-19. Viruses. 2023;15:578. doi:10.3390/v15020578
29. Kim J, Hickerson BT, Ilyushina NA. Coinfection of influenza A and B and human OC43 coronavirus in normal human bronchial epithelial cells. Influenza Other Respir Viruses. 2024;18e13279. doi:10.1111/irv.13279
30. Tempestilli M, Caputi P, Avataneo V, et al. Pharmacokinetics of remdesivir and GS-441524 in two critically ill patients who recovered from COVID-19. J Antimicrob Chemother. 2020;75(10):2977–2980. doi:10.1093/jac/dkaa239
31. Smee DF, Hurst BL, Evans WJ, et al. Evaluation of cell viability dyes in antiviral assays with RNA viruses that exhibit different cytopathogenic properties. J Virol Methods. 2017;246:51–57. doi:10.1016/j.jviromet.2017.03.012
32. Ding Y, Zeng L, Li R, et al. The Chinese prescription lianhuaqingwen capsule exerts anti-influenza activity through the inhibition of viral propagation and impacts immune function. BMC Complement Altern Med. 2017;17:130. doi:10.1186/s12906-017-1585-7
33. Ma Q, Huang W, Zhao J, et al. Liu Shen Wan inhibits influenza A virus and excessive virus-induced inflammatory response via suppression of TLR4/NF-κB signaling pathway in vitro and in vivo. J Ethnopharmacol. 2020;252:112584. doi:10.1016/j.jep.2020.112584
34. Wang M, Li W, Cui W, et al. The therapeutic promises of Lianhuaqingke in the mice model of coronavirus pneumonia (HCoV-229E and SARS-CoV-2). Chin Med. 2021;16:124. doi:10.1186/s13020-021-00513-3
35. Goker Bagca B, Biray Avci C. The potential of JAK/STAT pathway inhibition by ruxolitinib in the treatment of COVID-19. Cytokine Growth Factor Rev. 2020;54:51–62. doi:10.1016/j.cytogfr.2020.06.013
36. Borchers C, Thyagarajan A, Rapp CM, Travers JB, Sahu RP. Evaluation of SARS-CoV-2 spike S1 protein response on PI3K-mediated IL-8 release. Med Sci. 2021;9:30. doi:10.3390/medsci9020030
37. Grimes JM, Grimes KV. p38 MAPK inhibition: a promising therapeutic approach for COVID-19. J Mol Cell Cardiol. 2020;144:63–65. doi:10.1016/j.yjmcc.2020.05.007
38. Perrin P, Collongues N, Baloglu S, et al. Cytokine release syndrome-associated encephalopathy in patients with COVID-19. Eur J Neurol. 2021;28:248–258. doi:10.1111/ene.14491
39. Lee DH, Lee CS. Flavonoid myricetin inhibits TNF-α-stimulated production of inflammatory mediators by suppressing the Akt, mTOR and NF-κB pathways in human keratinocytes. Eur J Pharmacol. 2016;784:164–172. doi:10.1016/j.ejphar.2016.05.025
40. Zheng B, Qi J, Yang Y, et al. Mechanisms of cinnamic aldehyde against myocardial ischemia/hypoxia injury in vivo and in vitro: involvement of regulating PI3K/AKT signaling pathway. Biomed Pharmacother. 2022;147:112674. doi:10.1016/j.biopha.2022.112674
41. Jang WY, Kim MY, Cho JY. Antioxidant, anti-inflammatory, anti-menopausal, and anti-cancer effects of lignans and their metabolites. Int J Mol Sci. 2022;23:15482. doi:10.3390/ijms232415482
42. Zhang YL, Peng HM, Li JJ, et al. The volatile oil of Hyssopus cuspidatus Boriss. ameliorates OVA-induced allergic asthma via inhibiting PI3K/Akt/JNK/P38 signaling pathway and maintaining airway barrier integrity. J Ethnopharmacol. 2024;334:118568. doi:10.1016/j.jep.2024.118568
43. Yang L, Liu R, Fang Y, He J. Anti-inflammatory effect of phenylpropanoids from Dendropanax dentiger in TNF-α-induced MH7A cells via inhibition of NF-κB, Akt and JNK signaling pathways. Int Immunopharmacol. 2021;94:107463. doi:10.1016/j.intimp.2021.107463
44. Karuppan MKM, Devadoss D, Nair M, Chand HS, Lakshmana MK. SARS-CoV-2 infection in the central and peripheral nervous system-associated morbidities and their potential mechanism. Mol Neurobiol. 2021;58:2465–2480. doi:10.1007/s12035-020-02245-1
45. He D, Liu Y, Li J, et al. Isoalantolactone regulates neuro-inflammation and neuronal apoptosis to curb pathology of Parkinson’s disease. Cells. 2022;11:2927. doi:10.3390/cells11182927
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Anti-Inflammatory Effects and Molecular Mechanisms of Shenmai Injection in Treating Acute Pancreatitis: Network Pharmacology Analysis and Experimental Verification
He Y, Hu C, Liu S, Xu M, Liang G, Du D, Liu T, Cai F, Chen Z, Tan Q, Deng L, Xia Q
Drug Design, Development and Therapy 2022, 16:2479-2495
Published Date: 2 August 2022
A Novel Approach Based on Gut Microbiota Analysis and Network Pharmacology to Explain the Mechanisms of Action of Cichorium intybus L. Formula in the Improvement of Hyperuricemic Nephropathy in Rats

Amatjan M, Li N, He P, Zhang B, Mai X, Jiang Q, Xie H, Shao X
Drug Design, Development and Therapy 2023, 17:107-128
Published Date: 20 January 2023
Network Pharmacology and Experimental Validation to Explore That Celastrol Targeting PTEN is the Potential Mechanism of Tripterygium wilfordii (Lév.) Hutch Against IgA Nephropathy

Zhao J, Liu H, Xia M, Chen Q, Wan L, Leng B, Tang C, Chen G, Liu Y, Zhang L, Liu H
Drug Design, Development and Therapy 2023, 17:887-900
Published Date: 23 March 2023
Investigating the Mechanism of Action of Schisandra chinensis Combined with Coenzyme Q10 in the Treatment of Heart Failure Based on PI3K-AKT Pathway

Wen S, Yang K, Bai Y, Wu Y, Liu D, Wu X, Zhang X, Sun J
Drug Design, Development and Therapy 2023, 17:939-957
Published Date: 27 March 2023
Network Pharmacology Analysis of the Mechanisms Underlying the Therapeutic Effects of Yangjing Zhongyu Tang on Thin Endometrium
Zhang L, Li H, Zhang L, Zu Z, Xu D, Zhang J
Drug Design, Development and Therapy 2023, 17:1805-1818
Published Date: 17 June 2023