Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Integrated Bioinformatics Identifies NLRP3 Inflammasome Hub Genes and Therapeutic Targets in Schizophrenia
Authors Ma J, Cui Y, Li X, Liu R, Wang J, Liu M ![]() , Kong L, Ren Y
, Kong L, Ren Y
Received 8 August 2025
Accepted for publication 24 November 2025
Published 1 December 2025 Volume 2025:21 Pages 2655—2670
DOI https://doi.org/10.2147/NDT.S555802
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yu-Ping Ning
Jiawei Ma,1 Yongchun Cui,1 Xinying Li,2 Ruiyuan Liu,3 Jinhui Wang,3,4 Mengdi Liu,2 Linping Kong,5 Yan Ren6
1Department of Psychiatry, Third Hospital of Shanxi Medical University, Shanxi Bethune Hospital, Shanxi Academy of Medical Sciences, Tongji Shanxi Hospital, Taiyuan, 030032, People’s Republic of China; 2School of Humanities and Social Sciences, Shanxi Medical University, Taiyuan, People’s Republic of China; 3Academy of Medical Sciences, Shanxi Medical University, Taiyuan, 030001, People’s Republic of China; 4Department of Pharmacy, Shanxi Medical University, Taiyuan, 030001, People’s Republic of China; 5Department of Medical Psychology, Taiyuan Psychiatric Hospital, Taiyuan, 030000, People’s Republic of China; 6Department of Psychiatry, The Fifth Hospital of Shanxi Medical University, The Fifth Clinical Medical College of Shanxi Medical University, Shanxi Provincial People’s Hospital, Taiyuan, 030012, People’s Republic of China
Correspondence: Yan Ren, Department of Psychiatry, The Fifth Hospital of Shanxi Medical University, The Fifth Clinical Medical College of Shanxi Medical University, Shanxi Provincial People’s Hospital, Taiyuan, 030012, People’s Republic of China, Email [email protected]
Background: The pathogenesis of schizophrenia (SZ) remains incompletely understood; although neuroinflammation and the NLRP3 inflammasome have been implicated, the key regulatory genes involved are still unidentified.
Objective: To investigate the association between SZ and NLRP3 inflammasome-related genes, and to screen for hub genes as potential biomarkers and therapeutic targets.
Methods: We analyzed the GEO dataset GSE27383, comprising 43 SZ patients and 29 controls, and identified 1,672 differentially expressed genes (DEGs). NLRP3-related genes were obtained from GeneCards, and weighted gene co-expression network analysis (WGCNA) highlighted the green, yellow, and red modules. The intersection of DEGs, NLRP3-related genes, and module genes was further refined using LASSO and Random Forest algorithms. Immune cell infiltration was profiled with CIBERSORT, and the diagnostic utility of candidate genes was evaluated using ROC curves. Molecular docking was performed to predict compound binding, and hub gene expression was validated in an independent cohort of 20 SZ patients and 20 controls using RT-qPCR on PBMCs.
Results: Five hub genes—HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS—were significantly down-regulated in SZ (P < 0.05). The combined ROC-AUC reached 0.883. Molecular docking revealed strong binding affinities of ellagic acid to FLNA (− 4.70 kcal mol−1), and of hydralazine to HSPA8 (− 4.27), TRAF2 (− 4.84), and SCAP (− 4.98). Differential expression was confirmed in PBMCs. Additionally, SZ patients exhibited increased naive B cells and neutrophils, along with reduced resting NK cells and M2 macrophages.
Conclusion: Five genes (HSPA8, SCAP, FLNA, TRAF2, PINK1-AS) were identified as potential novel biomarkers and therapeutic targets for SZ, providing a theoretical foundation for elucidating disease mechanisms and advancing precision medicine in SZ.
Keywords: schizophrenia, NLRP3 inflammasome, molecular docking, bioinformatics, biomarkers, precision psychiatry
Introduction
Schizophrenia (SZ) is a highly heterogeneous severe mental disorder characterized by a core symptom cluster encompassing disorganized thought structure, perceptual abnormalities, blunted affect, social withdrawal, and pervasive cognitive impairment.1 There is significant variability in individual symptoms, onset patterns, course, and outcomes. Additionally, positive, negative, and disorganized symptoms are often accompanied by other related features such as cognitive impairments, emotional symptoms, and motor disorders.2 Notably, the current clinical diagnostic system primarily relies on subjective symptom assessments and rating scales, lacking established, clinically translatable biomarker standards. The absence of such objective indicators not only poses challenges for early screening and differential diagnosis but also hinders the development of targeted therapies and personalized intervention strategies. In this context, exploring reliable biomarkers has become a critical breakthrough for addressing the current limitations in psychiatric clinical practice.
In recent years, the role of neuroinflammation mechanisms in the pathophysiological processes of SZ has increasingly become a research hotspot.3 Recent studies have shown that dysregulation of the NLRP3 inflammasome-mediated signaling pathway can activate downstream pro-inflammatory factors, leading to abnormal activation of microglia and triggering a neuroinflammatory cascade reaction.4 This pathological mechanism is not only closely related to the onset and progression of SZ but may also be involved in the common pathogenic pathways of various neuropsychiatric disorders, including bipolar disorder, Alzheimer’s disease, amyotrophic lateral sclerosis, and Parkinson’s disease.5 Building on prior evidence of elevated peripheral IL-1β and post-mortem cortical NLRP3 protein in schizophrenia patients,6,7 this study extends these findings by systematically analyzing multi-cohort transcriptomic data to identify central drivers of inflammasome dysregulation rather than isolated cytokine alterations. Although previous investigations have explored polygenic risk scores and functional MRI markers, these approaches remain constrained by modest effect sizes or high costs and have not yet produced clinically applicable assays; a transcriptome-based inflammasome module could therefore address a critical need for biomarkers. Therefore, this study aims to systematically identify hub genes associated with NLRP3 inflammasome activation in SZ through integrated bioinformatics approaches, with the goal of providing a theoretical basis for the development of novel diagnostic biomarkers and targeted therapeutic strategies.The workflow of this study is presented in Figure 1.
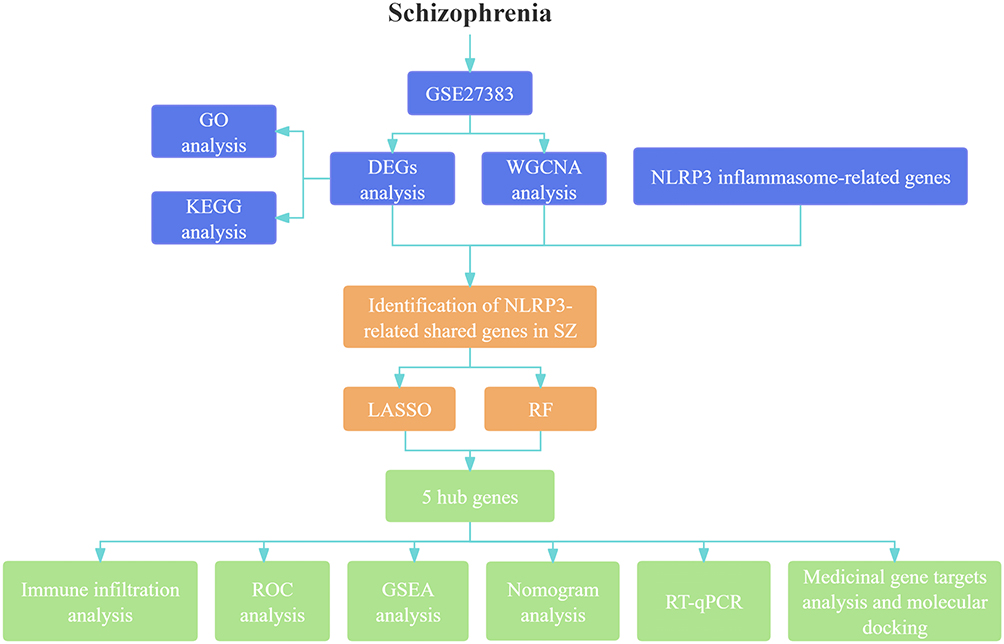 |
Figure 1 The flowchart of the study. |
Methods
Subject Recruitment
The research protocols were approved by the medical ethics committee of the Third Hospital of Shanxi Medical University. All participants voluntarily participated in the study and provided informed consent. Each procedure adhered to applicable rules and regulations. Patient recruitment was conducted from November 1st to December 31th, 2024. A total of 20 hospitalized patients with schizophrenia (SZ) were recruited through a systematic screening of admission records at the Department of Psychiatry of our hospital. Potential participants were initially identified by their attending physicians based on preliminary diagnosis. Eligible patients were then approached by a research coordinator who explained the study in detail.
The diagnosis of SZ patients was established according to the Diagnostic and Statistical Manual of Mental Disorders (Fifth Edition) (DSM-5). To ensure that patients possessed adequate cognitive capacity to understand the study and provide valid consent, all potential schizophrenia participants underwent Mini-Mental State Examination (MMSE) assessment before inclusion. Only patients with MMSE scores ≥24 were enrolled in this study. Exclusion criteria included significant physical comorbidities, other psychiatric disorders, or substance abuse. All participants provided written informed consent voluntarily after a thorough explanation of the study procedures. Each participant’s capacity to provide informed consent was formally assessed and confirmed by two independent attending psychiatrists, who evaluated their clinical stability and cognitive ability to comprehend the study’s purpose, risks, and benefits. Only individuals deemed capable of providing informed consent were included in the study.
Additionally, 20 healthy controls (HCs) were recruited during the same period (March 1st to April 30th, 2025). HCs were recruited from the health screening center of the same hospital via flyers and announcements. Potential HCs underwent an initial telephone screening to confirm the absence of personal or family history of psychiatric disorders. Eligible individuals were then invited for a structured clinical interview conducted by a trained psychiatrist to definitively rule out any DSM-5 disorders. All HCs were confirmed to be free of neurological, systemic medical conditions, and any DSM-5 disorders.
Data Collection and Processing
The gene expression data and clinical information used in this study were obtained from existing SZ datasets in the GEO database (https://www.ncbi.nlm.nih.gov/geo/). GSE27383 comprises gene expression profiles derived from peripheral blood mononuclear cells (PBMCs) of 43 schizophrenia patients and 29 healthy controls. All tissue samples were analyzed using the GPL570 [HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 array.
Additionally, NLRP3 inflammasome-related genes were obtained from GeneCards (https://www.genecards.org/). We searched for the term “NLRP3 inflammasome” to identify genes associated with the NLRP3 inflammasome. Genes with a GeneCards association score >1.7 were defined as NLRP3 inflammasome-related.
Differential Expression Analysis
The Limma package in R software was used to screen differentially expressed genes (DEGs) based on p < 0.05 and |log FC| > 0.1. DEGs with logFC < 0 were classified as downregulated; those with logFC > 0 were classified as upregulated. Volcano plots and heatmaps were then generated using the “ggplot2” and “pheatmap” packages.8
Enrichment Analysis of Differentially Expressed Genes
The “clusterProfiler” package in R was used to perform Gene Ontology (GO) analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis on the differentially expressed genes.9 GO analysis includes biological processes (BP), cellular components (CC), and molecular functions (MF). KEGG is a well-known reference database containing information on genomes, biological pathways, diseases, and drugs. The significance level was set at P < 0.05.
Weighted Gene Co-Expression Network Analysis (WGCNA)
WGCNA can identify co-expressed gene modules and explore the relationship between gene sets and phenotypes, which is useful for studying core genes in gene expression data.10 In this study, we used the WGCNA package to construct a scale-free co-expression gene network based on peripheral blood data from SZ patient samples and healthy control samples (GSE27383). We selected the top 25% of genes by variance for WGCNA analysis. Then, we used the goodSamplesGenes function in the WGCNA R package to remove outliers, ensuring a refined dataset for network construction. The soft threshold power β was set to 8, representing the lowest value where the scale-free topology fit index (R2) reached 0.9. After dynamic pruning and TOM hierarchical clustering analysis, modules showing the strongest correlation with the SZ phenotype were identified for further analysis. Genes were clustered into modules using hierarchical clustering with a deepSplit parameter of 2, and modules with a correlation less than 0.25 were merged. Ultimately, 19 co-expression gene modules of different colors were obtained. Correlation analysis showed that the green, yellow, and red modules were highly correlated with SZ. Therefore, genes from the green, yellow, and red modules were selected for further analysis.
Identification of NLRP3 Inflammasome-Associated Genes in Schizophrenia
We identified NLRP3 inflammasome overlap genes in SZ by taking the intersection of three gene sets: (1) the DEGs, (2) genes from WGCNA co-expression modules highly associated with SZ, and (3) the NLRP3 inflammasome gene set.
Machine Learning
To further investigate key potential genes, we used the glmnet package for LASSO logistic regression11 and the randomForest package for the RF algorithm.12 These two machine learning methods were employed to narrow down the range of candidate biomarkers. Finally, the intersection of the results from both algorithms was identified as the hub genes for diagnosis.
Validation of Hub Gene Expression in Schizophrenia
The expression of hub genes obtained from LASSO regression analysis and RF analysis was validated in the schizophrenia dataset GSE27383. The ggplot2 package was used to compare and visualize the expression levels of hub genes.
Construction of Nomogram and ROC Curve Plotting
To determine the importance of hub genes in SZ diagnosis and enable clinical prediction, we used the rms package to construct a nomogram based on the hub genes.13 The nomogram consists of scales for each predictor variable (contributing to a “Points” score), a “Total Points” scale, and a scale for the predicted probability of SZ. ROC curves were then used to assess the diagnostic performance of this nomogram in predicting SZ. An AUC value >0.7 is commonly considered to indicate good diagnostic capability.14 The diagnostic efficacy of the model incorporating the five key genes for SZ was evaluated accordingly.
Gsea
In the SZ dataset GSE27383, we used the org.Hs.eg.db and clusterProfiler packages to perform gene set enrichment analysis on each hub gene to compare the differential signaling pathways between SZ and healthy controls.15 The Enrichplot package was used to display the top 5 activated and inhibited pathways in SZ.
Immune Infiltration Analysis
In the SZ dataset GSE27383, we used CIBERSORT to describe the differences in the distribution of 22 immune cell types between SZ patients and HCs.16 The ggplot2 package was used to visualize the infiltration of the 22 immune cell types.
Quantitative Reverse Transcription-PCR
Venous blood was collected from all participants using a blood collection tube containing ethylenediaminetetraacetic acid (EDTA). Peripheral blood mononuclear cells (PBMCs) were subsequently isolated from whole blood samples using Ficoll solution (Solarbio Life Sciences, China). Total RNA was extracted using Trizol reagent (TransGen, China) according to the manufacturer’s instructions. The quality of the RNA was assessed using a NanoVue Plus Spectrophotometer (Biochrom, UK). Subsequently, qPCR was performed on a CFX96 Real-Time PCR Detection System (Bio-Rad, USA). GAPDH (glyceraldehyde-3-phosphate dehydrogenase), was used as a reference gene for data normalization. qPCR experiments were conducted using the primers listed in the Table 1. Fold changes in the expression levels of the indicated genes were calculated using the 2^-(ΔΔCt) method.17
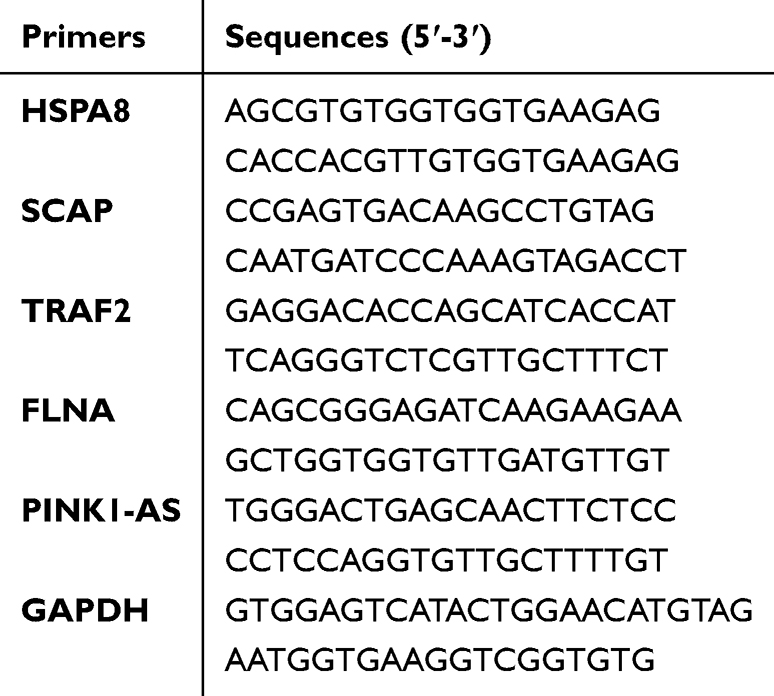 |
Table 1 RT-qPCR Primers |
Predicting Therapeutic Drugs for Hub Genes Through Molecular Docking
Further molecular docking experiments were conducted to explore the interactions between potential therapeutic drugs and the selected hub genes. First, the Drug Signature Database (https://maayanlab.cloud/Enrichr/DSigDB) on the Enrichr website (https://maayanlab.cloud/Enrichr/) was accessed to obtain potential drugs targeting the core genes.18,19 Subsequently, obtained protein structures corresponding to hub genes and small-molecule drugs from the UniProt database (https://www.uniprot.org/) and the PubChem website (https://pubchem.ncbi.nlm.nih.gov/), respectively.20 Finally, AutoDuck Vina (version 1.5.6) (https://ccsb.scripps.edu/mgltools/downloads/) and PyMOL software (version 2.5.5) were used to simulate the binding of the target protein with the small-molecule drug.21
Statistical Analysis
All analyses were performed in R software. The normality of data distribution was assessed using the Shapiro–Wilk test. A p-value ≥ 0.05 was considered to indicate a normal distribution. Based on this, the Student’s t-test was used for normally distributed data, and the Mann–Whitney U-test was applied for non-normally distributed data. Significance was defined as a P-value < 0.05.
Results
Differential Gene Expression in Schizophrenia
R software was used to analyze the GSE27383 dataset and screen for DEGs associated with SZ. Based on the criteria of p < 0.05 and |log FC| > 0.1, a total of 1,672 DEGs were obtained, including 617 upregulated genes and 1,055 downregulated genes. These DEGs are shown in the volcano plot (Figure 2A). A heatmap of the top 30 differentially expressed genes was drawn (Figure 2B).
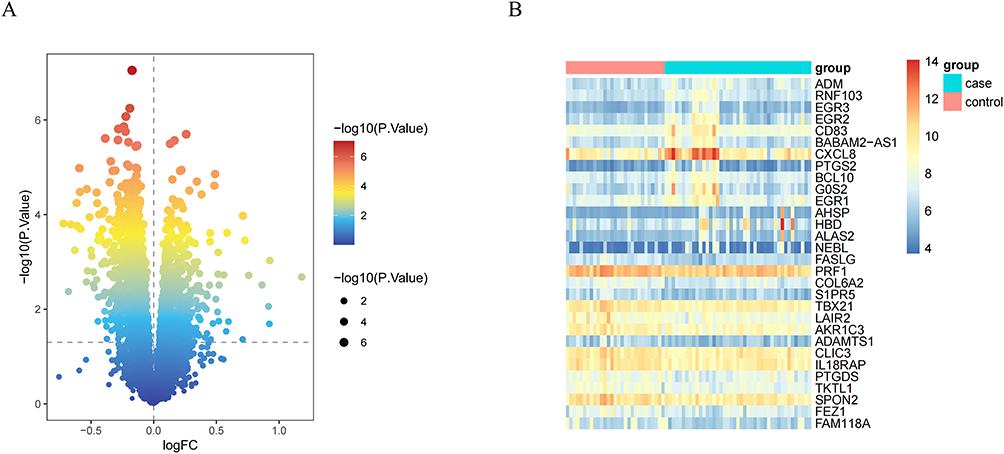 |
Figure 2 (A) Shows the volcano map of differentially expressed genes. (B) Shows the heat map of differentially expressed genes. |
Enrichment Analysis of Differentially Expressed Genes
GO and KEGG enrichment analysis were performed on differentially expressed genes. Biological process(BP) was mainly related to innate immune responses, cellular responses to pathogen stimuli, and signal pathway regulation. Cellular Component (CC) terms were primarily enriched in structures related to cell migration, phagocytosis, and signal transduction platforms. Molecular function(MF) covered functions such as protein modification and signal molecule binding (Figure 3A and B). KEGG analysis mainly involved inflammatory responses, pathogen recognition, cytokine signal transduction, and immune activation (Figure 3C and D).
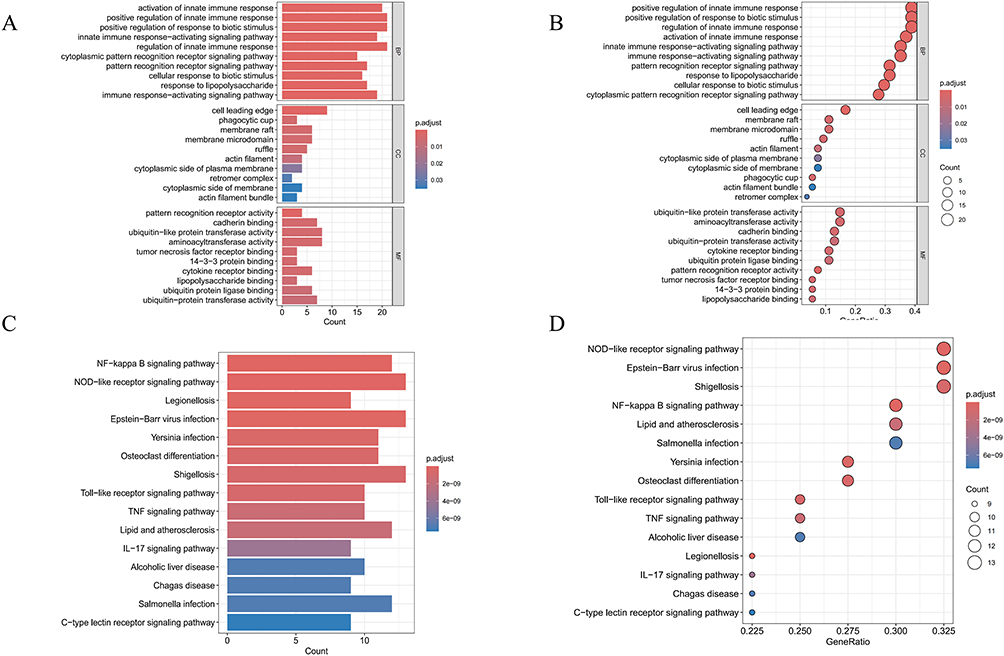 |
Figure 3 GO functional and KEGG pathway analysis of DEGs. (A) Shows the bar graph of GO enrichment analysis.(B) Shows the bubble graph of GO enrichment analysis.(C) Shows the bar graph of KEGG enrichment analysis.(D) Shows the bubble graph of KEGG enrichment analysis. |
Construction of a Weighted Gene Co-Expression Network and Identification of Key Modules in Schizophrenia
To further explore hub genes in SZ, weighted gene co-expression network analysis (WGCNA) was performed to identify the most relevant gene modules in SZ samples. A soft threshold of β = 8 was used to ensure average connectivity and scale-free topology in the co-expression network (Figure 4A and B). Ultimately, 19 co-expression gene modules of different colors were obtained. Module-trait association analysis revealed that the green, yellow, and red modules exhibited the strongest correlations with SZ status (Figure 4C). Therefore, 906 module genes from the green module (r = −0.43, p = 2e-04, n=283), yellow module (r = −0.39, p = 6e-04, n=375), and red module (r = 0.35, p = 0.003, n=248) were selected for further analysis (Figure 4D–F).
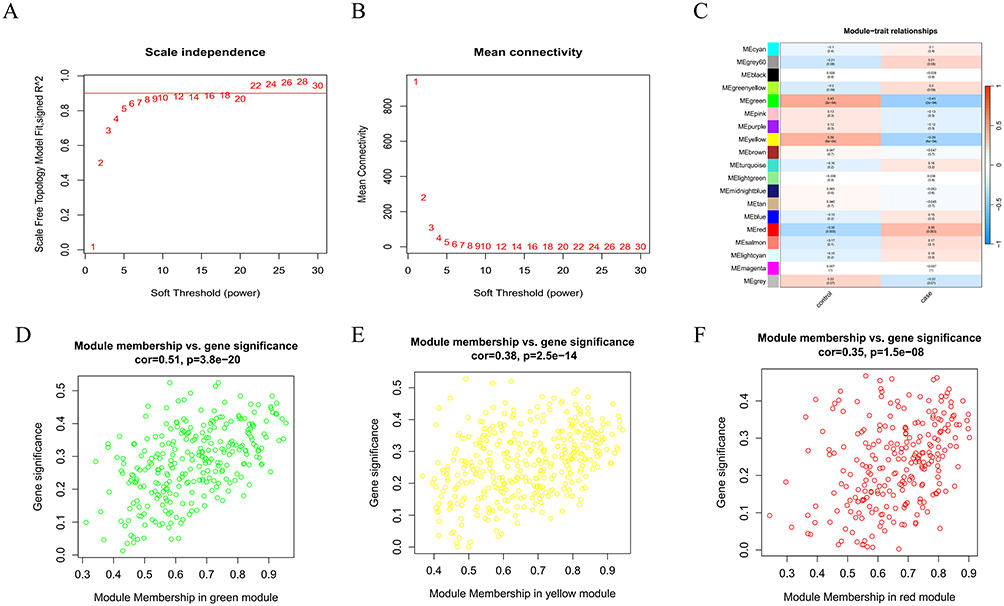 |
Figure 4 (A and B) Determination of the optimal β value using a scale-free topology model, with β=8 selected as the soft threshold based on average connectivity and scale Independence. (C) Heatmap revealing the relationship between module-feature genes and SZ. (D–F):Correlation plots of green, yellow, and red module members with gene significance. |
NLRP3 Inflammasome Gene Overlap Screening in Schizophrenia
Based on the GeneCards (https://www.genecards.org/) database, we obtained 458 genes related to the NLRP3 inflammasome. The intersection of these 458 NLRP3 inflammasome-related genes with the 906 SZ-associated module genes (from WGCNA) and the 1,672 DEGs yielded overlapping genes associated with SZ and the NLRP3 inflammasome (Figure 5). This multi-tiered filtering approach, integrating differential expression, co-expression networks, and a specific pathological pathway, was designed to prioritize genes with higher confidence and biological relevance beyond conventional single-method analyses.
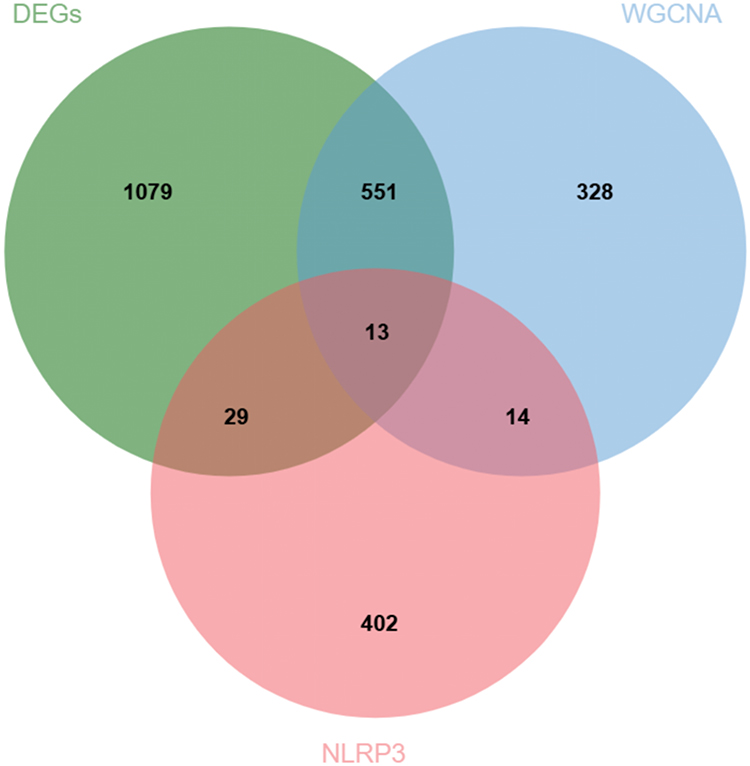 |
Figure 5 Analysis of overlapping genes associated with SZ and the NLRP3 inflammasome. |
Screening Hub Genes Using Machine Learning
We further screened hub genes using two machine learning algorithms: LASSO and RF. LASSO regression identified 17 genes with non-zero coefficients (Figure 6A and B). The RF algorithm ranked genes based on their importance, and we selected the top 8 genes as candidate genes (Figure 6C and D). The intersection of genes selected by LASSO (17 genes) and RF (top 8 genes) yielded 5 common hub genes: HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS (Figure 6E).
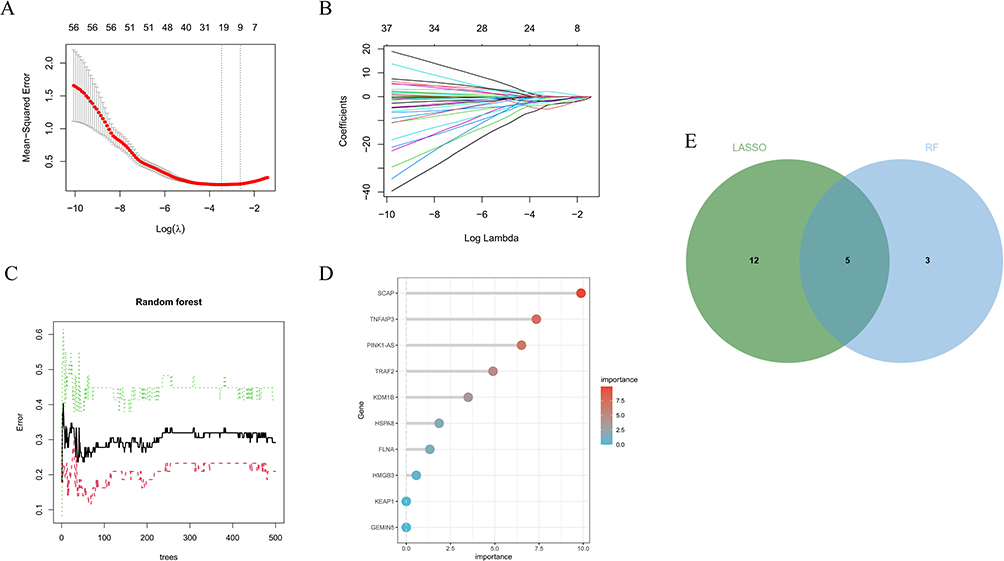 |
Figure 6 (A and B) Biomarkers screening in the LASSO model. (C and D):The error rate confidence intervals and the relative importance of genes for Random Forest algorithm. (E):The Venn diagram of the intersection of LASSO and Random Forest signature genes. |
Validating Hub Gene Expression in Schizophrenia
In the schizophrenia dataset (GSE27383), compared with the control group, the expression levels of HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS were all downregulated in SZ patients (P < 0.05). These results confirm the downregulation of HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS expression in SZ patients compared to HCs (Figure 7).
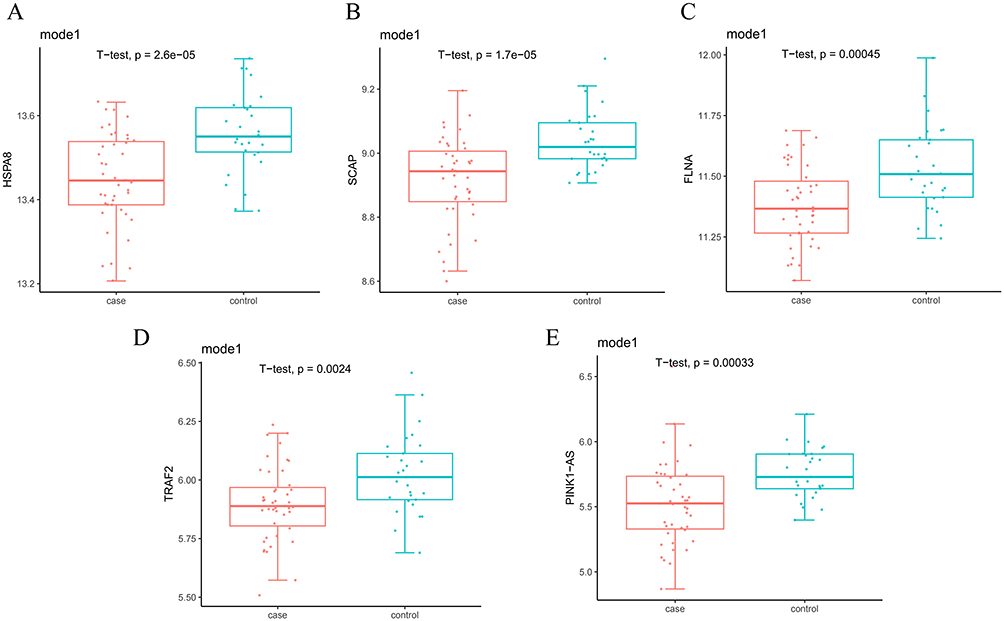 |
Figure 7 Expression levels of the five hub genes in the SZ dataset (GSE27383). (A) HSPA8; (B) SCAP; (C) FLNA; (D) TRAF2; (E) PINK1-AS. |
Construction of a Nomogram and ROC Curve
To improve the diagnosis and prediction of SZ, we constructed a nomogram based on the five key genes. By summing the scores corresponding to each key gene, we obtained a total score, which corresponds to the disease risk of SZ (Figure 8A and B). ROC curve analysis was used to assess the area under the curve (AUC) of the nomogram model to determine its sensitivity and specificity in diagnosing SZ. The AUC value for dataset GSE27383 was 0.883, indicating that the nomogram model has good diagnostic performance for SZ. Furthermore, the individual diagnostic efficacy of each of the five hub genes for SZ was also evaluated. The results demonstrated that all five hub genes exhibited significant diagnostic value (AUC > 0.7): HSPA8 (AUC = 0.764), SCAP (AUC = 0.771), FLNA (AUC = 0.735), TRAF2 (AUC = 0.702), and PINK1-AS (AUC = 0.750) (Figure 8C).
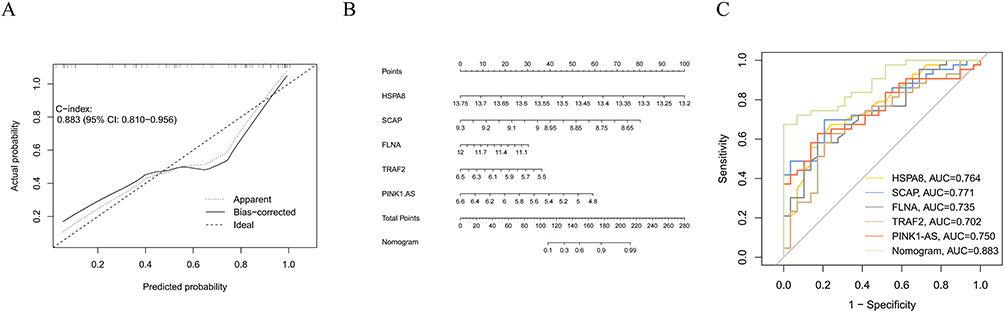 |
Figure 8 (A) Calibration. (B) nomogram constructed based on diagnostic biomarkers curve of the nomogram model predicting SZ. The dashed line labeled “Ideal” represents the standard curve, indicating perfect prediction by an ideal model. The dashed line labeled “Apparent” represents the uncalibrated prediction curve, while the solid line labeled “Bias-Corrected”represents the calibrated prediction curve. (C) The nomogram model diagnostic performance of SZ and the diagnostic performance of 5 hub genes were predicted by receiver operating characteristic (ROC) curves in the internal dataset. |
Gsea
To elucidate the role of these five hub genes in the SZ dataset GSE27383, we performed GSEA analysis. The results showed that GO enrichment was primarily concentrated in multiple important pathways, including detection of bacterial-derived molecules, processing and presentation of exogenous antigens, mitochondrial fusion, membrane repolarization during ventricular cardiomyocyte action potentials, and positive regulation of mRNA metabolic processes, indicating that these five hub genes are involved in the regulation of SZ (Figure 9).
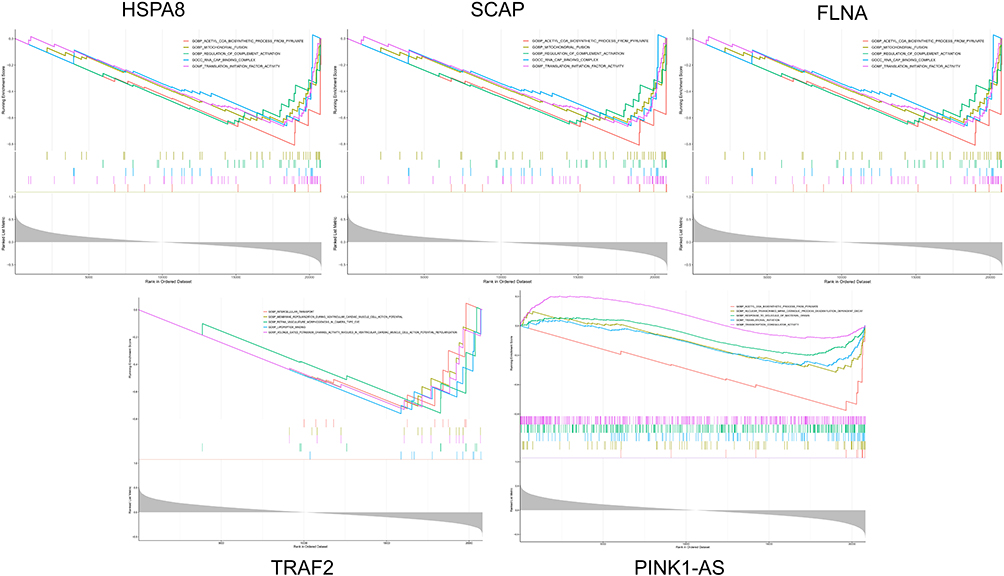 |
Figure 9 GSEA analysis and GO enrichment of the five hub genes. |
Immune Infiltration Analysis
We used the Cibersort algorithm to perform immune infiltration analysis on peripheral blood mononuclear cells from 43 SZ patients and 29 healthy controls. Analysis of the 22 infiltrating immune cell types in the SZ dataset GSE27383 revealed significant differences in 6 cell types. Compared to the healthy controls (HCs), SZ patients showed significantly higher proportions of naive B cells, activated memory B cells, plasma cells, and neutrophils, while the proportions of resting NK cells and M2 macrophages were significantly lower. Additionally, we further explored the relationship between the expression of five hub genes and the proportions of different infiltrating immune cell types. HSPA8 was positively correlated with resting NK cells but negatively correlated with M1 macrophages, activated mast cells, and monocytes; SCAP was positively correlated with resting NK cells and M2 macrophages; FLNA was positively correlated with resting NK cells and CD8+ T cells, but negatively correlated with resting memory CD4+ T cells, CD4+ naive T cells, monocytes, and eosinophils; TRAF2 was positively correlated with resting NK cells but negatively correlated with monocytes; PINK1-AS was positively correlated with resting NK cells but negatively correlated with naive B cells and resting memory CD4+ T cells (Figure 10).
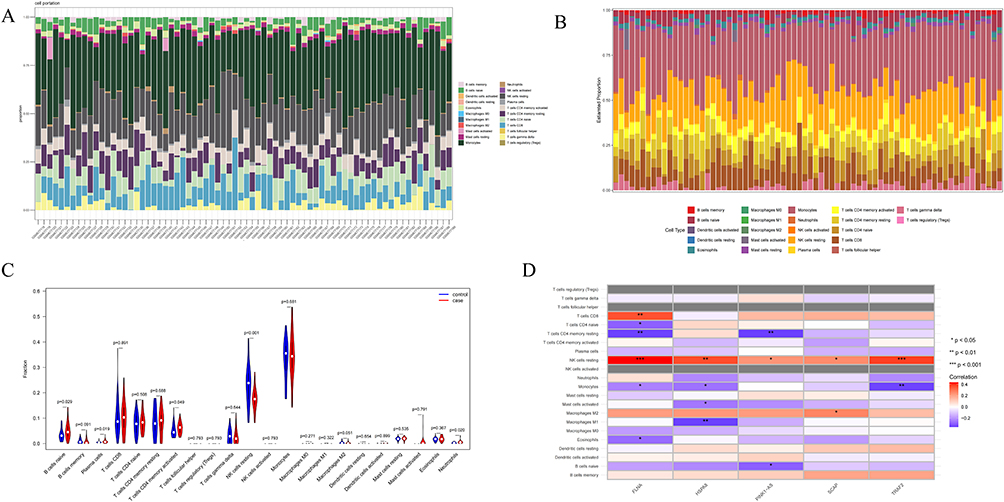 |
Figure 10 (A and B) The relative percentage of 22 immune cells in each sample. (C) Differences in immune infiltration between SZ and HCs samples. (D) Correlation of hub genes with immune cells and different immune factors. Red represents a positive correlation, and blue represents a negative correlation. The darker the color is, the stronger correlation is. Significant differences were supposed at *p-value < 0.05, **p-value < 0.01, ***P-value < 0.001 compared with the control. |
Expression Validation of Hub Genes by RT-qPCR
To more comprehensively assess the levels of HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS in SZ patients, we analyzed the mRNA expression of these hub genes in PBMCs from healthy individuals and SZ patients using RT-qPCR. The primer information for RT-qPCR is shown in Table 1. RT-qPCR analysis in the independent cohort confirmed significant downregulation of HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS mRNA expression in PBMCs from SZ patients compared to HCs (P < 0.05) (Figure 11).
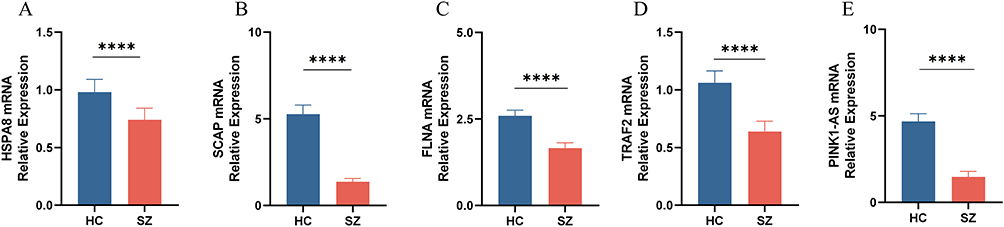 |
Figure 11 Relative mRNA expression of the following genes in SZ v. HC. (A) HSPA8. (B) SCAP. (C) FLNA. (D) TRAF2. (E) PINK1-AS. Significant differences were supposed at *p-value < 0.05, **p-value < 0.01, ***P-value < 0.001, ****P-value < 0.0001 compared with the control. |
Visualization of Molecular Docking Simulations Targeting Hub Genes
To explore the therapeutic effects of drugs targeting the five hub genes HSPA8, FLNA, TRAF2, SCAP, and PINK1-AS on SZ, we first identified candidate drugs using the DSigDB database. As shown in Table 2, ellagic acid and hydralazine were among the top predicted candidates, both with a nominal P-value of 0.001 and an adjusted P-value of 0.051, associated with the gene sets HSPA8;FLNA and HSPA8;SCAP;TRAF2, respectively. Based on these predictions, we downloaded the high-resolution crystal structures of HSPA8, FLNA, TRAF2, SCAP. Molecular docking simulations predicted favorable binding energies between candidate compounds and hub gene products: specifically, ellagic acid bound to FLNA with an energy of −4.70 kcal/mol, while hydralazine bound to HSPA8 (−4.27 kcal/mol), TRAF2 (−4.84 kcal/mol), and SCAP (−4.98 kcal/mol) (Figure 12).
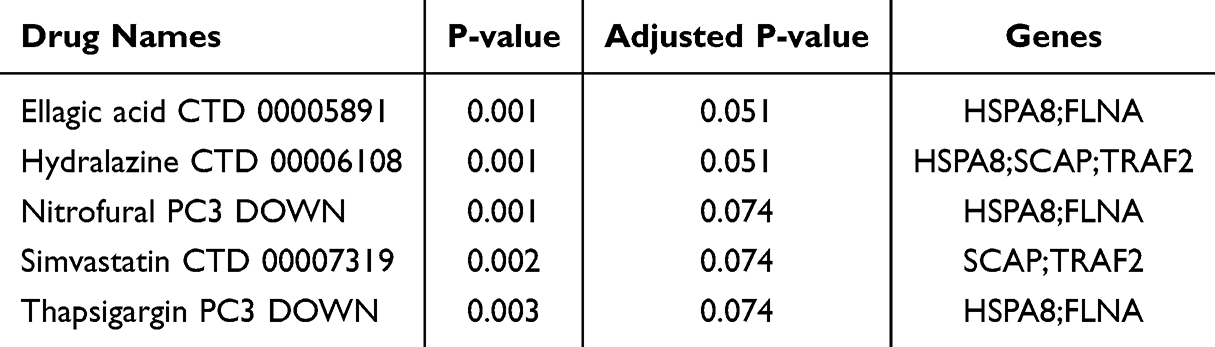 |
Table 2 Candidate Drug Predicted Using DSigDB |
 |
Figure 12 (A) HSPA8 complexed with hydralazine. (B) SCAP complexed with hydralazine. (C) FLNA complexed with ellagic acid. (D) TRAF2 complexed with hydralazine. |
Discussion
Schizophrenia is a chronic and complex mental illness that can cause various cognitive impairments, including memory, thinking, perception, and volition.22 Despite extensive research, effective biomarkers are still lacking. Increasing evidence suggests that inflammation contributes to the pathophysiology of SZ, particularly negative symptoms, by disrupting brain structure.23 Against this backdrop, the present study was designed to identify and validate novel inflammasome-related biomarkers through a multi-method approach, thereby directly addressing the critical need for reliable diagnostic markers in SZ. This study systematically investigated five hub genes (HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS) associated with the NLRP3 inflammasome pathway in SZ through the integration of bioinformatics analysis and experimental validation. The study found that the five genes HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS were downregulated in peripheral blood mononuclear cells (PBMCs) from SZ patients and demonstrated good diagnostic value (AUC > 0.7), with the AUC of the multi-gene-based nomogram model reaching 0.883. This multi-gene panel exhibits superior diagnostic performance over many previously reported single-gene biomarkers for SZ, underscoring the benefit of combinatorial biomarker approaches.
A key advancement of our study is its multi-tiered analytical strategy, which integrates differential expression, co-expression network analysis (WGCNA), and pathway-focused filtering for the NLRP3 inflammasome, culminating in machine learning. This integrated approach prioritizes genes with greater confidence and biological relevance than conventional analyses based on a single method.
Heat shock homolog 71 kDa (HSC70), also known as HSPA8, is a member of the heat shock protein 70 family. As an important molecular chaperone, HSPA8 participates in protein folding, transport, and degradation, maintaining cellular homeostasis.24 Studies have shown that HSPA8 can negatively regulate inflammasome activation by modulating the conformational stability of NLRP3 or inhibiting its oligomerization.25 In SZ, abnormal expression of HSPA8 may weaken its inhibitory effect on NLRP3, leading to excessive release of inflammatory factors and exacerbating neuroinflammatory damage.26,27 SCAP is a key transport molecule for cholesterol regulatory element-binding proteins, regulating lipid synthesis and metabolism.28 Abnormal cholesterol metabolism has been reported to be associated with SZ pathology.29 SCAP may promote SREBP activation, drive lipid accumulation, and thereby activate the NLRP3 inflammasome.30 FLNA is a key protein in cytoskeletal remodeling, involved in cell migration, signal integration, and membrane receptor localization. Studies have found that FLNA can directly interact with NLRP3 to promote inflammasome spatial assembly. Abnormal FLNA may disrupt neuronal synaptic stability while promoting NLRP3 activation.31 TRAF2 is a core adaptor protein in the NF-κB pathway, involved in inflammatory and apoptotic signal transduction.32 This study found that PINK1-AS is abnormally expressed in SZ, and PINK1-AS, as the antisense RNA of the PINK1 gene, may participate in the mitochondrial autophagy dysfunction mechanism of SZ by regulating PINK1 expression.33 The identification of these five genes aligns with the growing recognition of neuroinflammatory mechanisms in SZ and expands the current biomarker landscape by incorporating both protein-coding genes and regulatory RNAs.
Regarding the robustness of validation, we acknowledge that the sample size of the primary dataset (GSE27383) is modest. However, the subsequent experimental validation of the hub genes using RT-qPCR in an independent cohort of patient PBMCs significantly strengthens the reliability and reproducibility of our findings, mitigating concerns related to the initial sample size limitation.
Immune infiltration analysis showed that the proportions of naive B cells, memory B cells, plasma cells, and neutrophils were elevated, while the proportions of resting NK cells and M2 macrophages were reduced in SZ patients, suggesting that the peripheral immune system of patients is in a pro-inflammatory state. Correlation analysis between hub genes and immune cells further revealed that genes such as HSPA8 may participate in immune homeostasis imbalance by regulating NK cell and macrophage function.These findings strengthen the immunological dimension of SZ pathophysiology and suggest that the identified hub genes may serve as mediators between immune dysfunction and neural impairment.
Molecular docking predictions of potential interactions between candidate drugs and the proteins encoded by the hub genes provide a theoretical foundation for exploring NLRP3 inflammasome-targeted therapies. Notably, hydralazine demonstrated multi-target binding capabilities: it exhibited binding affinity for HSPA8 (−4.27 kcal/mol), suggesting a potential role in stabilizing chaperone-mediated stress responses, and showed even stronger binding to TRAF2 (−4.84 kcal/mol) and SCAP (−4.98 kcal/mol), potentially modulating inflammatory signaling and lipid metabolism, respectively. Ellagic acid also exhibited binding affinity with FLNA (−4.70 kcal/mol), indicating possible cytoskeletal regulation. These computational findings offer promising leads for developing novel therapeutic strategies; however, it is crucial to emphasize that these are preliminary in silico predictions that require thorough experimental validation.
From a clinical perspective, our findings hold significant implications. The identified five-gene signature could potentially be developed into a blood-based diagnostic test, providing a much-needed objective tool for SZ diagnosis. Moreover, the association of these genes with inflammasome pathways opens new avenues for targeted therapies, particularly for patients with prominent inflammatory features. The differential immune cell profiles further suggest potential for immunomodulatory interventions in subset of SZ patients.
Our study also has certain limitations. First, the data used in our study were obtained from the GEO database, and the limited sample size of the primary dataset (GSE27383) increases the risk of false discoveries and requires validation in larger cohorts. Therefore, further studies with larger sample sizes are needed to elucidate the exact pathogenesis of NLRP3 inflammasomes associated with SZ. Second, while we validated gene expression in PBMCs, which are clinically accessible, it remains to be determined whether these peripheral findings fully reflect pathological processes within the central nervous system. Third, although the predicted binding affinity between drugs and targets shows potential, these predicted interactions require experimental validation through in vitro binding assays and cellular models.The binding energies reported (−4.27 to −4.98 kcal/mol) indicate moderate affinity; further pharmacological assays are needed to confirm functional efficacy. Finally, the cross-sectional nature of our data limits inferences regarding the temporal dynamics of these biomarkers throughout disease progression. Future longitudinal studies tracking these markers across illness stages would offer valuable insights into their potential as state versus trait markers.
In conclusion, by employing an integrated strategy, we identified and validated a set of key genes associated with SZ and the NLRP3 inflammasome pathway. We have highlighted the methodological advancements of our approach, discussed the robustness of validation despite initial sample size constraints, and acknowledged the limitations concerning generalizability. These findings provide new insights into the inflammatory mechanisms of SZ and offer promising candidates for diagnostic development and therapeutic exploration.
Conclusion
This study identified five NLRP3 inflammasome-related genes—HSPA8, SCAP, FLNA, TRAF2, and PINK1-AS—as potential biomarkers for SZ. Their downregulation is closely associated with NLRP3 inflammasome-mediated neuroinflammation and peripheral immune dysfunction. The diagnostic model constructed based on these genes shows promising predictive performance and suggests potential for future clinical translation, pending further validation. These findings contribute to the growing field of neuroinflammatory research in SZ and provide a foundational framework for developing precision diagnostic tools and targeted therapeutic strategies aimed at the NLRP3 inflammasome pathway. Future studies should focus on independent validation in larger cohorts and further exploration of the mechanistic roles of these genes to fully assess their clinical applicability.
Data Sharing Statement
All data generated or analysed during this study are included in this published article. The public datasets analysed are available in the NCBI Gene Expression Omnibus (GEO) repository (https://www.ncbi.nlm.nih.gov/geo/). Any additional raw data that are not publicly available can be obtained upon reasonable request to the two Corresponding Authors.
Ethics Statement
This study protocol was in accordance with the Declaration of Helsinki of the World Medical Association. The studies involving humans were approved by the medical ethics committee of Shanxi Bethune Hospital, with the Approval Notice Number: 2021GLL147. Informed consent was obtained from all the study subjects before enrollment.
Acknowledgments
An unauthorized version of the Chinese MMSE was used by the study team without permission, however this has now been rectified with PAR. The MMSE is a copyrighted instrument and may not be used or reproduced in whole or in part, in any form or language, or by any means without written permission of PAR (www.parinc.com).
Funding
The authors declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (No. 82101579), Funding project for Science and Technology Activities of Overseas Students in Shanxi Province (No.20240055), Applied basic research projects of Shanxi Province, China (Grant No. 202403021211045).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Jauhar S, Johnstone M, McKenna PJ. Schizophrenia. Lancet. 2022;399(10323):473–486. doi:10.1016/S0140-6736(21)01730-X
2. Faden J, Citrome L. Schizophrenia: one name, many different manifestations. Med Clin North Am. 2023;107(1):61–72. doi:10.1016/j.mcna.2022.05.005
3. Péladeau C, Sandhu JK. Aberrant NLRP3 inflammasome activation ignites the fire of inflammation in neuromuscular diseases. Int J Mol Sci. 2021;22(11):6068. doi:10.3390/ijms22116068
4. Jia C, Zhang M, Wu X, et al. HERV-W env induces neuron pyroptosis via the NLRP3-CASP1-GSDMD pathway in recent-onset schizophrenia. Int J Mol Sci. 2025;26(2):520. doi:10.3390/ijms26020520
5. Leng F, Edison P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol. 2021;17(3):157–172. doi:10.1038/s41582-020-00435-y
6. Goldsmith DR, Bekhbat M, Mehta ND, Felger JC. Inflammation-related functional and structural dysconnectivity as a pathway to psychopathology. Biol Psychiatry. 2023;93(5):405–418. doi:10.1016/j.biopsych.2022.11.003
7. Szabo A, Akkouh I, Osete JR, et al. NLRP3 inflammasome mediates astroglial dysregulation of innate and adaptive immune responses in schizophrenia. Brain Behav Immun. 2025;124:144–156. doi:10.1016/j.bbi.2024.11.030
8. Ritchie ME, Phipson B, Wu D, et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015;43(7):e47. doi:10.1093/nar/gkv007
9. Kanehisa M, Furumichi M, Sato Y, Kawashima M, Ishiguro-Watanabe M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023;51(D1):D587–D592. doi:10.1093/nar/gkac963
10. Xu M, Zhou H, Hu P, et al. Identification and validation of immune and oxidative stress-related diagnostic markers for diabetic nephropathy by WGCNA and machine learning. Front Immunol. 2023;14:1084531. doi:10.3389/fimmu.2023.1084531
11. Engebretsen S, Bohlin J. Statistical predictions with glmnet. Clin Clin Epigenet. 2019;11(1):123. doi:10.1186/s13148-019-0730-1
12. Alderden J, Pepper GA, Wilson A, et al. Predicting pressure injury in critical care patients: a machine-learning model. Am J Crit Care. 2018;27(6):461–468. doi:10.4037/ajcc2018525
13. Zhong M, Li J, Zhang LZ, Nie R, Zheng Z. Screening of immune-related secretory proteins linking chronic kidney disease with calcific aortic valve disease based on comprehensive bioinformatics analysis and machine learning. J Transl Med. 2023;21(1):359. doi:10.1186/s12967-023-04171-x
14. Sing T, Sander O, Beerenwinkel N, Lengauer T. ROCR: visualizing classifier performance in R. Bioinformatics. 2005;21(20):3940–3941. doi:10.1093/bioinformatics/bti623
15. Wu T, Hu E, Xu S, et al. A universal enrichment tool for interpreting omics data. Innovation. 2021;2(3):100141. doi:10.1016/j.xinn.2021.100141
16. Xu J, Zhou H, Cheng Y, Xiang G. Identifying potential signatures for atherosclerosis in the context of predictive, preventive, and personalized medicine using integrative bioinformatics approaches and machine-learning strategies. EPMA J. 2022;13(3):433–449. doi:10.1007/s13167-022-00289-y
17. Biray Avci C, Sezgin B, Goker Bagca B, Karci HB, Gode S. PI3K/AKT/mTOR pathway and autophagy regulator genes in paranasal squamous cell carcinoma metastasis. Mol Biol Rep. 2020;47(5):3641–3651. doi:10.1007/s11033-020-05458-8
18. Yoo M, Shin J, Kim J, et al. DSigDB: drug signatures database for gene set analysis. Bioinformatics. 2015;31(18):3069–3071. doi:10.1093/bioinformatics/btv313
19. Kuleshov MV, Jones MR, Rouillard AD, et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016;44(W1):W90–7. doi:10.1093/nar/gkw377
20. Magrane M, UniProt Consortium. UniProt knowledgebase: a hub of integrated protein data. Database. 2011;2011:bar009. doi:10.1093/database/bar009.
21. Ferreira LG, Dos Santos RN, Oliva G, Andricopulo AD. Molecular docking and structure-based drug design strategies. Molecules. 2015;20(7):13384–13421. doi:10.3390/molecules200713384
22. Patlola SR, Donohoe G, McKernan DP. The relationship between inflammatory biomarkers and cognitive dysfunction in patients with schizophrenia: a systematic review and meta-analysis. Prog Neuropsychopharmacol Biol Psychiatry. 2023;121:110668. doi:10.1016/j.pnpbp.2022.110668
23. Goldsmith DR, Rapaport MH. Inflammation and negative symptoms of schizophrenia: implications for reward processing and motivational deficits. Front Psychiatry. 2020;11:46. doi:10.3389/fpsyt.2020.00046
24. Wu E, He W, Wu C, et al. HSPA8 acts as an amyloidase to suppress necroptosis by inhibiting and reversing functional amyloid formation. Cell Res. 2023;33(11):851–866. doi:10.1038/s41422-023-00859-3
25. Liu J, Song K, Lin B, et al. The suppression of HSPA8 attenuates NLRP3 ubiquitination through SKP2 to promote pyroptosis in sepsis-induced lung injury. Cell Biosci. 2024;14(1):56. doi:10.1186/s13578-024-01239-z
26. Kowalczyk M, Owczarek A, Suchanek-Raif R, Kucia K, Kowalski J. An association study of the HSPA8 gene polymorphisms with schizophrenia in a Polish population. Cell Stress Chaperones. 2022;27(1):71–82. doi:10.1007/s12192-021-01249-7
27. Mi J, Yang Y, Yao H, et al. Inhibition of heat shock protein family A member 8 attenuates spinal cord ischemia-reperfusion injury via astrocyte NF-κB/NLRP3 inflammasome pathway: HSPA8 inhibition protects spinal ischemia-reperfusion injury. J Neuroinflammation. 2021;18(1):170. doi:10.1186/s12974-021-02220-0
28. Lee SH, Lee JH, Im SS. The cellular function of SCAP in metabolic signaling. Exp Mol Med. 2020;52(5):724–729. doi:10.1038/s12276-020-0430-0
29. Xu CX, Huang W, Shi XJ, et al. Dysregulation of serum exosomal lipid metabolism in schizophrenia: a biomarker perspective. Mol Neurobiol. 2025;62(3):3556–3567. doi:10.1007/s12035-024-04477-x
30. Guo C, Chi Z, Jiang D, et al. Cholesterol homeostatic regulator SCAP-SREBP2 integrates NLRP3 inflammasome activation and cholesterol biosynthetic signaling in macrophages. Immunity. 2018;49(5):842–856.e7. doi:10.1016/j.immuni.2018.08.021
31. Shi Y, Qiu T, Wu C, et al. Filamin A facilitates NLRP3 inflammasome activation during arsenic-induced nonalcoholic steatohepatitis. Environ Sci Pollut Res Int. 2023;30(49):107703–107715. doi:10.1007/s11356-023-29702-3
32. Albini A, Di Paola L, Mei G, et al. Inflammation and cancer cell survival: TRAF2 as a key player. Cell Death Dis. 2025;16(1):292. doi:10.1038/s41419-025-07609-w
33. Chu H, Zhu H, Ma J, et al. Mitochondrial dysfunction and metabolic indicators in patients with drug-naive first-episode schizophrenia: a case-control study. Neuropsychiatr Dis Treat. 2024;20:2433–2442. doi:10.2147/NDT.S501527
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification of Critical Biomarkers and Mechanisms of Fructus Ligustri Lucidi on Vitiligo Using Integrated Bioinformatics Analysis

Liang TS, Tang N, Xian MH, Wen WL, Huang CJ, Cai LH, Li QL, Wu YH
Clinical, Cosmetic and Investigational Dermatology 2023, 16:2061-2071
Published Date: 7 August 2023
Network Analysis of Osteoarthritis Progression Using a Steiner Minimal Tree Algorithm
Xie Y, Shao F, Ji Y, Feng D, Wang L, Huang Z, Wu S, Sun F, Jiang H, Miyamoto A, Wang H, Zhang C
Journal of Inflammation Research 2024, 17:3201-3209
Published Date: 18 May 2024
ITGA4 Contributes to 5-Fluorouracil Resistance by Up-Regulating PI3K/AKT Signaling: Evidence from Network Pharmacology, Molecular Docking and Experimental Verification

Yan S, Hu X, Wu Y, Ye W, Zhu Y, He Y, Zhan F, Wu W, Ma Z
Drug Design, Development and Therapy 2025, 19:4105-4122
Published Date: 20 May 2025
Identification of Bioactive Compounds and Molecular Targets of Compound Herbs Against Stomach Adenocarcinoma: A Network Pharmacology Approach
Liu L, Liu Y, Wu M
International Journal of General Medicine 2026, 19:549743
Published Date: 29 January 2026
Integrated Machine Learning and Multi-Omics Analysis Identifies Mitophagy-Related Core Genes and Mechanisms in Non-Alcoholic Fatty Liver Disease
Yuan Y, Zhang T, Song C, Lin C, Sun Y, Tang H
Journal of Inflammation Research 2026, 19:575586
Published Date: 24 March 2026