Back to Journals » ImmunoTargets and Therapy » Volume 13
Harnessing Variability Signatures and Biological Noise May Enhance Immunotherapies’ Efficacy and Act as Novel Biomarkers for Diagnosing and Monitoring Immune-Associated Disorders
Authors Sigawi T, Israeli A, Ilan Y ![]()
Received 12 July 2024
Accepted for publication 27 September 2024
Published 14 October 2024 Volume 2024:13 Pages 525—539
DOI https://doi.org/10.2147/ITT.S477841
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Tal Sigawi, Adir Israeli, Yaron Ilan
Faculty of Medicine, Hebrew University and Department of Medicine, Hadassah Medical Center, Jerusalem, Israel
Correspondence: Yaron Ilan, Faculty of Medicine, Hebrew University and Department of Medicine, Hadassah Medical Center, Jerusalem, Israel, Email [email protected]
Abstract: Lack of response to immunotherapies poses a significant challenge in treating immune-mediated disorders and cancers. While the mechanisms associated with poor responsiveness are not well defined and change between and among subjects, the current methods for overcoming the loss of response are insufficient. The Constrained Disorder Principle (CDP) explains biological systems based on their inherent variability, bounded by dynamic boundaries that change in response to internal and external perturbations. Inter and intra-subject variability characterize the immune system, making it difficult to provide a single therapeutic regimen to all patients and even the same patients over time. The dynamicity of the immune variability is also a significant challenge for personalizing immunotherapies. The CDP-based second-generation artificial intelligence system is an outcome-based dynamic platform that incorporates personalized variability signatures into the therapeutic regimen and may provide methods for improving the response and overcoming the loss of response to treatments. The signatures of immune variability may also offer a method for identifying new biomarkers for early diagnosis, monitoring immune-related disorders, and evaluating the response to treatments.
Keywords: immune system, variability, artificial intelligence, immunotherapy
Introduction
There has been a growing interest in immunotherapy in the last decades. Immunotherapy, including immune cell therapy and targeted therapy, is gradually developed through the ongoing discovery of molecular compounds or immune cells.1 Selecting the best approach or combination of target compounds and immune-cell therapy is challenging for clinical scientists and clinicians. Colorectal cancer (CRC) ranks among the most prevalent malignancies affecting the gastrointestinal tract.2 The infiltration of CD8+ T cells significantly influences the prognosis and progression of tumor patients.2
Passive immunotherapy involves using anti-CD20 and anti-TNF agents, bispecific antibodies with well-defined specificity and subclasses, and antibody-drug conjugates (ADCs). Another approach involves T cells transduced with chimeric antigen receptors (CAR) to expand tumor-infiltrating lymphocytes. In active immunotherapy, patients are given B- or T-cell-based immunity against selected antigens or induced active tolerance against allergens, autoantigens, and alloantigens. Combining both approaches is utilized to release the brakes on T cells that have already responded to antigens through T cell checkpoint control blockers.3 However, the current response rate to immunotherapies could be more satisfactory. Many patients suffer from primary non-responsiveness, and among those who initially respond, there is a large segment of secondary non-responders.4–6 The lack of response is associated with the dynamicity of both the host and the disease.
Introducing variability patterns was proposed to enhance the clinical response to immunomodulatory agents.7 The current paper aims to introduce the concept of constrained disorder principle (CDP) in light of recent data on the immune system’s natural variability. Developing second-generation treatments based on artificial intelligence (AI) is essential to enhance the response to immunotherapies.
The Principle of Constrained Disorder Explains How Complex Systems Operate Within Specific Boundaries
The CDP defines systems based on their inherent variability.” It differentiates living organisms by their variability, which is bounded by dynamic boundaries enabling systems to respond to perturbations.8 The CDP is based on the variability that characterizes biological systems at the gene, cell, and whole organ levels.9–33 In homeostasis, systems aim at returning to their setpoint post perturbations, and in allostasis, systems change their setpoint in response to triggers. The CDP views noise as inherent to the proper function of biological systems. Perturbations are answered by widening or tightening the noise boundaries in a system. Per the CDP, diseased states are described by a reduction in noise or an increase beyond the borders.8,33–35
Variability is Inherent in the Immune Response: Mechanisms of Intra- and Inter-Individual Variability in the Immune System
The immune system exhibits remarkable inter-individual variability as well as intra-individual variability.36 Evolution maintains diversity within the immune system, which provides a powerful defense against pathogens. Nevertheless, immune-associated diseases may develop based on immune variation.37,38 The dynamic nature of the immune system is influenced by various internal and external factors, including genetic variations, age, sex, environmental influences, microbiome, and disease states.36 Many of these variables are inextricably intertwined, making it challenging to assign definitive contributions to them.37,39,40
Genetic polymorphism in immune-related genes can lead to diverse immune phenotypes.37 The heritable contribution to the variability in immune cell frequencies is estimated to be between 20–50%.37,41–44 As a result of the polymorphic genes that control the immune system and the sensitive environmental sensors that shape it, immunity can be pushed into various functional configurations.44 One of these configurations is an intrinsic bias toward a particular type of immune response. Most healthy humans can produce type I interferons, type II and type III cells, Th17 cells, type I helper T cells, and inflammasome activation. However, the extent to which individuals are predisposed to a particular functional configuration varies significantly from person to person.45,46 The interindividual differences continuously expand with age and are somewhat stable and resilient to perturbation.36,37,47,48
Genetic variations in human leukocyte antigen (HLA) genes have been linked to varying susceptibility to autoimmune diseases, infectious diseases, and response to immunotherapies.37 Specific HLA alleles, such as HLA-DRB1 in rheumatoid arthritis (RA) and HLA-DQB1 in type 1 diabetes, are lined with disease susceptibility.37,49 Interferon α (IFNα) levels are increased in systemic lupus erythematosus (SLE) and their healthy first-degree relatives, implying genetic influences on cytokine expression.36
Sexual dimorphism is observed in various aspects of the immune system, leading to inter-individual variability.42 Overall, women appear to have higher immune cell counts and immunoglobulin levels, although some studies inconsistently demonstrated higher counts of specific immune cells and immunoglobulin subgroups in males.42,50–52 Sex-specific differences in disease prevalence, clinical outcomes, and response to immunotherapies have been observed, highlighting the importance of considering sex-based differences in immune profiles.1,2 Clinically, many autoimmune and inflammatory diseases show female predominance, while ankylosing spondylitis is more prevalent in male patients.36 The response to vaccines is often more intense in women, attributed to the immune-enhancing effects of estrogen and immune-depressing effects of testosterone.36,37,42,53 Several studies did not show a significant sex-related inter-individual immune variability.50,54
The immune system is relatively stable within a healthy individual, as implied by the steady immune cell and protein profiles of blood samples drawn weeks to years apart from healthy subjects.36,50,54–56 Variations in immune function over time are an individual feature.41,57 Individual variations not caused by reported infections are a feature of the immune system, causing more significant shifts in phenotypes from one point to another.41,50,58 A few cellular subpopulations showed intra-individual variability in consecutive tests of healthy individuals.55 Healthy older individuals may show enhanced intra-individual immune variability over time.47,54 This highly variable phenotype may be linked to a diminished flu vaccine response.54 Also, high intra-individual variability in immune components was associated with a morbid metabolic profile and markers of cardiovascular morbidity and possibly mortality.41,47
A year-long study was conducted on healthy adults aged 50 to 65 years. The study monitored 750 plasma proteins and 115 immune cell populations every three months. It helped in understanding the relationships and fluctuations in the blood-immune system. Immune systems are usually stable, but the level of longitudinal variability varies from individual to individual. In the absence of apparent infections, individuals with the most variation exhibited variances in metabolic health markers, suggesting a link between immunologic and metabolic regulation.41 Cell composition changes radically during early life due to environmental exposures, suggesting a distinctive cell composition developed during this critical period.59–61
Neutrophils are more abundant in the circulation of older individuals but exhibit impaired migration and phagocytosis.54,62 Cytotoxic NK cell count rises with age, with diminished cytotoxic capabilities.54,62 Self vs non-self antigen recognition disruptions due to altered function and amount of the antigen-presenting dendritic cells are also noted.62 Involving the thymus leads to a decrease in naïve T cells, a reduction in peripheral T cell diversity, and an increase in memory and regulatory T cells.42,43,54,55,62 Lymph nodes, splenic degeneration, and bone marrow dysfunction may also interrupt the homeostasis of lymphocyte populations and the humoral response of older individuals.42,50,54,62 These changes, recognized as immunosenescence, increase susceptibility to infections, cancer, and inflammatory conditions and decrease vaccine responsiveness in older adults.36,37,43,53,62,63 Aging is linked to low-grade inflammatory phenotype (CLIP), inflammaging.62 It is characterized by elevated levels of inflammatory cytokines such as IL-6, IL-18, tumor necrosis factor (TNF), and CRP.36,50,52,62 This chronic inflammation disrupts immune homeostasis, impairs immune cell function, and underlies the development of age-related diseases.52 Age is also associated with inter-individual immune variability, as younger individuals show more inter-individual similarity of immunological traits than the diverse phenotypes of older adults.37,41,54,63 Monozygotic twin pairs show higher inter-individual variability with older age, attributed to non-heritable factors.43 This age-related variability is attributed to an increased cumulative environmental exposure and to direct age effects.37,41
Non-heritable factors are the dominant source of immune variability.37,41–44 These factors occur throughout an individual’s life course, thus contributing to intra-individual variability on various time scales. Additionally, they constitute a distinct input reflecting an individual’s lifestyle, consequently influencing inter-individual variability.
Several studies demonstrated the seasonal variability of immune cell frequencies and plasma protein concentrations,42,51,52 while others showed no seasonal effects.41 Clinically, the incidence of type-1 diabetes mellitus is higher during winter;34 allergies and viral infections show seasonal variations;37,48 RA symptoms exhibit seasonal variations, as well as a circadian pattern with morning symptoms, correlated to a spike in serum IL-6 levels.36
Variations in microbiota composition can contribute to immune system variability and influence susceptibility to immune-related disorders and vaccine response.36 The imbalance in the microbial community, bacterial dysbiosis, is typically linked to reduced microbial diversity and is related to various immune disorders such as inflammatory bowel disease(IBD) and asthma.36 Diet can alter immune system modulation. Deficiencies or nutrient imbalances can compromise immune responses and increase susceptibility to infections.37 Nonetheless, the impact of normal dietary variations on the immune response is uncertain.37 Dietary alterations may also exert their effects indirectly through modifications in the microbiome or body weight.37,52 Nutritional supplementations may influence immune response, as was demonstrated in measles-vaccinated iron-deficient children treated with iron.37 Salt-rich diets are associated with increased autoimmunity.37 Obesity has been linked to increased inflammatory markers such as IL-6 and altered NK cell count and function, attributed to the lipid-rich environment.37,50 Long-term exposure to pollutants may lead to chronic inflammation, immune dysregulation, and increased susceptibility to immune-related disorders.36,37
Cigarette smoking increases leukocyte counts, reduces total immunoglobulin levels, and increases autoantibody titers.36,37 Exposure to various infectious agents can shape the immune system. CMV-seropositivity contributed to inter-individual variability significantly.64,65 Monozygotic twins with discordant CMV seropositivity showed higher inter-individual variability in many immune parameters than concordant seropositive or seronegative twin pairs.44 Certain medications, vaccines, and immune therapies can affect the immune system, leading to variability within and between individuals.36
The data suggest that internal and external factors affect the immune system’s inter and intra-individual variabilities. This variability emphasizes the need for personalized medicine to address immune-mediated pathologies effectively. Furthermore, fluctuating immune function may indicate the aging process and may be associated with mortality, suggesting the potential clinical relevance of its monitoring.66 The design of variability-based digital twins is being developed to improve the accuracy of diagnosis and monitoring.67
Variability is Essential to the Proper Function of the Immune System
The immune system’s primary purpose is to protect the host from the uncountable diversity of constantly changing and evolving pathogens. Therefore, dynamicity, plasticity, and variability are required characteristics of the immune system.64 However, variability is not confined solely to the whole system or organism level. Instead, the immune cell itself can be perceived as a complex system that is subjected to constantly changing various inputs (eg, cytokines, chemokines, medications, toxins, and pollutants) aiming to generate a specific output or behavior (cytokine production, receptor expression, migration, and differentiation). Intercellular variability is a characteristic of all major immune cells.65,68,69 Characterization of single-cell “omics”, quantifying intra-individual intercellular variability, even among a well-defined, allegedly homogenous cellular subset, unravel previously concealed variability.70 The high-resolution single-cell analysis identified intra-individual daily variability of immune cellular composition among healthy individuals, challenging the previous dogma of intra-individual stability over time.68,71
The immunological inputs fluctuate, manifesting noise and stochasticity.68 Noise is a variation in a specific input signal that does not induce a different output by itself.68 However, accumulating noisy signals may alter the cellular output, demonstrating noisy properties.68 Stochasticity is another expression of randomness in cellular behavior that regards the choice of a distinct cellular outcome from a restricted repertoire of possibilities in response to a fixed input.68 Noise and stochasticity are evident at the level of intracellular molecules, various cellular processes, and intercellular interplay and account for intercellular variability.70
Intracellular molecular stochasticity mechanisms are fundamental in B and T cells.65,72 Through V(D)J recombination, these cells produce diverse B cell receptor (BCR) and T cell receptor (TCR) chains, allowing recognition of a wide range of antigen-major histocompatibility complex (MHC) combinations.65,72 Epigenetic modifications generate variability in the single-cell level phenotypes and show stochastic properties.65 Inter-cellular heterogeneity in interleukins and interferons mRNA expression in response to pathogens or vaccines was observed, stemming from stochasticity and probabilistic events in gene expression processes.64,65 Protein level variation was also noted for intracellular signaling molecules and surface receptors, contributing to intercellular diversity.65,70 Multiple intracellular stochastic events may determine the differentiation trajectory of a single immune cell.64,65 Cellular variability creates heterogeneity within a single immune cell type population that enables a gradual activation of the immune response.64,65
For an effective threat-appropriate and dose-dependent response, the immune response necessitates the maintenance of a varied array of responsive cell states.68 Variations in protective antibody production following vaccination have been linked to high plasmablast activity within a week of vaccination.73 CD38+ B cell subsets are a strong predictor across many cohorts and studies, accompanied by specific gene-expression signatures.74 Different activations of the interferon pathway have consistently been linked to increased antibody production in subsequent immune responses, as demonstrated across various vaccines.75 Positive innate responses at baseline, mediated primarily by plasmacytoid dendritic cells, are also possible.74 It is intriguing to note that a similar interferon signature predicts clinical flare-ups of SLE, indicating a common immunological variation depending on the context of activation.37,76
Elevated intercellular phenotypic variability can serve as a biomarker of aging and a quantitative trait, facilitating individual comparisons and intra-individual surveillance.70 Uncontrolled cellular variability can detrimentally impact tissue function, notably in cancer development and autoreactivity.64,65 With age, there is an increase in transcriptional variability, which has been demonstrated to disrupt synchronized immune responses.77 Single-cell examination has been used in studying immune disorders, such as RA,78,79 SLE,79,80 multiple sclerosis,81 IBD,82 asthma,83 and cancers.84,85 Single-cell proteomics showed that alterations in macrophage prevalence have predictive value for RA treatment effects.69 These methods revealed new immune cellular subtypes, advanced understanding of disease pathogenesis and clinical diversity, and potential individual therapeutic targets. Single-cell-based research established the boundaries between normal and pathological cellular heterogeneity of the ongoingly recognized immune cells, focusing on inter-individual variability.51 Intra-individual variability analysis utilizing these novel markers is a potential tool for monitoring disease progression and treatment response.86,87
Per the CDP, the variability kept within dynamic boundaries is essential for the proper function of systems.35 Variability characterizes biological systems and can be seen at the level of the DNA,9 tissues,12,13 and whole organs, including heart rate,10,11 blood pressure,14 respiratory,15 gait,16 and brain functions.17–19 In several of these systems, it was proposed that they be associated with improved function by providing biological systems with an ability to adapt to perturbations in their continuously changing environments.9,36,88
Similarly, variation can confer unique advantages on the immune system.37,89 As a protective mechanism, evolution has been selected for immune diversity rather than a homogeneous state of infection resistance.90,91 Possessing an immune system wired differently from the previous host can provide an evolutionary advantage when pathogens can quickly specialize in taking advantage of a fixed niche.34,37,92 Therefore, immune variation during immune responses is an essential mechanism mandated for effectiveness.34,93 Both antibodies’ structure, with a variable part of their structure, the multi types of cells cellular immunity, and cytokines panel, which are involved in all types of responses, are examples of the inherent variability that enables a relatively rapid response to perturbations.29,94,95
Thus, the variability is altered under changing host and environmental conditions, allowing the immune response to better deal with foreign antigens. Per the CDP, immune-associated diseases may evolve from a lack of variability or form a higher degree of variability.8 It is possible to develop treatments targeting these mechanisms to modulate the immune response by understanding how individuals’ immune systems differ, improving the immune response to vaccines, pathogens, and tumors, or alleviating immune-mediated disorders.96,97 By optimizing modifiable environmental conditions, it may be possible to enhance the long-term immunological health of all populations by gaining a better understanding of how and when the immune system achieves a stable state.36,98,99
The Challenge to Achieve a Sustained Effect of Immune-Mediated Therapies: Primary and Secondary Non-Responsiveness: Time Dimension in Disease Pathogenesis and Host Response
Lack of response to immunotherapies poses a significant challenge in treating immune-mediated disorders and cancer.100,101 Patients may not respond to initial therapy, primary non-response (PNR), or may lose response over time after initial improvement, with secondary loss of response (SLR).102
Tolerance against immunotherapies can result from forming anti-drug antibodies (ADA). Using drugs with high immunogenicity can generate ADA in up to 70% of the patients, depending on the drug.103 ADAs can be generated by T-cell-dependent or independent B-cell activation pathways. ADAs are produced in the T-cell-dependent pathway when a T helper cell (Th) differentiates into a Th1 or Th2 phenotype, ultimately creating plasma cells that secrete ADAs. It is clinically essential as infliximab-specific T-lymphocytes have been detected in the serum of subjects who received the drug, and there has been a correlation to ADA formation in those patients.104 Recent research examined risk factors for ADA formation against Infliximab. Smoking and RA encouraged ADA creation, whereas higher infliximab doses and higher serum infliximab concentrations reduced the risk of immunogenicity.105
PNR occurs in 10–40% of IBD patients, and it is usually declared after 12–14 weeks of therapy.102 Switching treatments between anti-TNF formulations can be effective in over 50% of PNR patients, suggesting that it is not entirely a class-effect phenomenon.106 SLR may result in treatment intensification or discontinuation in up to 50% of patients after 12 months. It may exacerbate symptoms of active IBD during maintenance therapy in a patient with controlled disease following induction treatment.107 A confirmation of SLR usually requires the recurrence of symptoms mediated by the inflammatory disease exacerbation of.7,108 The recent PANTS study tested 1610 active Crohn’s disease subjects receiving adalimumab or Infliximab to predict clinical factors leading to PNR and SLR. PNR at 14 weeks of treatment occurred in 23%. Non-remission after 54 weeks occurred in 63% of patients. In multivariable analysis, low drug levels at week 14 were an independent factor related to PNR and SLR. Low drug levels were also a predictive value for the immunogenicity of both drugs. Obesity, smoking, low albumin concentrations, higher baseline disease activity markers, and immunogenicity development were associated with low drug concentrations, which mediated non-remission.109
Among patients with rheumatoid arthritis, anti-TNF drugs are the first-choice treatment for patients who fail to respond to methotrexate therapy.110 Nonetheless, it is estimated that only 60% of subjects will attain a long-term response to these agents, while 30% of patients do not have an adequate primary response.7 In the DREAM registry, 6% of patients achieved remission according to the European League Against Rheumatism (EULAR) and the American College of Rheumatology (ACR) criteria. In comparison, 27% achieved a Disease Activity Score (DAS28) of less than 2.6, a less strict measure.111 The heterogeneity of patients with RA makes it challenging to predict who benefits from anti-TNF treatment. A recent study tried to predict response before treatment with anti-TNF, using machine learning models to assess DNA methylation and gene expression profiling in peripheral mononuclear cells. An accuracy of 79–88% in predicting response, based on the EULAR criteria for disease response, was reported.112 Unlike PLR, there is a lack of consensus on the definition of SLR in RA. SLR can be considered when there is an increase in EULAR response or DAS28>0.6 during the last six months.7,113 The average survival on anti-TNF in subjects with RA is estimated to be 47 months.114
Using immune-mediated therapy for cancer treatment also encounters challenges in achieving long-term effects. The reasons for treatment suspension vary from side effects to NPR and SLR. Despite performing a preliminary selection of patients who are candidates for Immune checkpoint inhibitor (ICI) treatment according to histo-pathology characteristics of the tumor tissue, only 13% of subjects are responsive.115–117 Prediction of ICI response is a complicated and evolving endeavor. It includes biomarkers such as intensity of PD-1 expression, tumor mutational burden, and analyzing signaling pathways in tumors as Interferon-γ (IFN-γ). More recent attempts for prediction include microbiome analysis using machine learning.118 One of the latest attempts is to examine the effects of epigenetics on NPR. Several studies have demonstrated that mutations in genes associated with the SWI-SNF chromatin remodeling complex can increase the sensitivity of human tumors to ICIs.
Additionally, treatment with an epigenetic modulator enhances the ICI-mediated anti-tumor effect.115,118 Resistance to ICIs is not fully understood due to the complexity of the immune response and its reliance on the host. While the response to ICIs varies depending on the specific disease, approximately 70% of patients are considered non-responders or experience disease progression after initially responding to these treatments.119
One suspected significant factor in resistance to ICI among cancer patients is the evolution or selection of tumors acquiring mutations in crucial pathways involved in the checkpoint blockade response.115 A tumor microenvironment (TME), which comprises factors extrinsic to cancer cells, can also prevent the ICI effect. It was shown that the TME can obtain Immune-suppressive cells and inhibitory cytokines, thus undermining the ICI effect.120,121 What was mentioned is only a fraction of how an SLR can occur in cancer patients, adding more challenges to finding an effective bypass to regain response.
Current Methods for Overcoming Immunotherapy Non-Responsiveness
Overcoming immunotherapy’s non-responsiveness is a significant challenge, considering the heterogeneity of the disease and multiple mechanisms of PNR and SLR. In cancer, non-response can occur through TME, creating a non-favorable condition around the tumor and preventing the drug effect. There are various strategies to overcome the TME effect. In triple-negative breast cancer, suppressing assays targeting annexin A1 (ANXA1) in mice in vivo experiment led to Treg function impairment and helped reduce the tumor size.122 Hypoxia also plays an essential role in TME deregulation and resistance to therapy. Recent research on HER2+ breast cancer in a pre-clinical model demonstrated that interfering with AXL tyrosine kinase receptor reduces It was also found that a pharmacological combination of inhibition of Axl, with anti–PD–1, assisted in reducing the size of the tumor and the spread of cancer.123 It is now essential to use additional drugs and biochemical agents to help CAR-T cells. It implies that compounds that affect different signaling pathways are given along with CAR-T cells. For example, Olaparib, a PARP inhibitor, significantly improved the activity of EGFR-specific CAR-T cells in a breast cancer model.124 Combining drugs is also an essential way to improve response to ICIs. Cytotoxic drugs are a pillar in treating advanced cancer and metastases. Essential cytotoxic drugs such as carboplatin, cisplatin, fluorouracil, and oxaliplatin can upregulate PD-L1 in malignant cells by generating danger signals.125
They enhance anti-tumor immunity via cytotoxic T cell activation, antigen-presenting cells maturing, immunosuppressive regulatory T cell depletion, and myeloid-derived suppressor cell expansion.126 Recent attempts to overcome ICI resistance include a gut microbiome therapeutic strategy. Several studies have demonstrated that microbial metabolites regulate anti-tumor immunity.127,128 Short-chain fatty acids (SCFAs) are produced when gut microbes ferment dietary fiber. These acids interact directly with CD8+ T cells, boosting their ability to fight tumors. Clinical studies have revealed that solid cancer patients treated with nivolumab had higher levels of fecal SCFAs if they responded positively to the treatment, suggesting a longer time without the cancer progressing.129 The benefits of SCFAs-producing microbiota can become another option for improving the response rate to ICI treatments.
Immune-mediated therapies, especially anti-TNF, are essential for many immunological diseases such as RA. However, as mentioned, 30–40% of patients cease the treatment due to PNR or SLR. Nonetheless, there are still no defined rules which indicate what to do in case of a treatment failure. European League Against Rheumatism (EULAR) recommends that a different biological disease-modifying anti-rheumatic drug (bDMARD) and anti-TNF be used instead in case of failure. Results of randomized controlled trials and observational data indicate that switching to a bDMARD with a different mechanism of action increases the likelihood of clinically significant improvements and improves drug retention rates.130 The GO-AFTER study discovered that golimumab reduced the symptoms of active RA and improved physical function in patients previously treated with TNFα inhibitors.131 An NJM study found that using abatacept in subjects with insufficient response to anti-TNF was clinically effective and had an acceptable safety profile.132 A recent study compared the effectiveness of second-line therapies for rheumatoid arthritis (RA), precisely anti-TNF drugs, and other biologics (abatacept, BDMA, rituximab, tocilizumab). The study found that BDMA showed greater sustainability as a second-line treatment than TNFi, especially in seropositive RA patients. However, seronegative patients did not experience the same drug survival advantage with BDMA. The study also highlighted that the failure and adverse effects of TNFi could be attributed to the development of anti-TNF antibodies, leading to a higher rate of treatment withdrawal.
Nonetheless, either BDMA or TNF can be chosen in seronegative and seropositive RA subjects exposed to anti-TNF or if the duration of TNFi was shorter than 2 years.133 This study is consistent with a previous study that determined the effectiveness of using a second-line anti-TNF drug compared with a non-TNF group. A non-TNF biologic agent achieved non-statistically better results than a second-line anti-TNF agent at 24 weeks.134
In IBD, dose intensification (DI) is often necessary due to loss of response. It is typically used in the context of SLR in case of low therapeutic drug levels and low ADA levels.7 In a meta-analysis, the drug intervention requirement rate was 28% in treatment-naïve patients and 39% in non-naïve patients. The short-term response to the empirically prescribed drug intervention was 63% and 58% in naïve and non-naïve subjects, respectively.
Additionally, no differences were found in comparing UC vs CD, the use of anti-TNF drugs, or even between intensification regimens.135 Another option is switching between anti-TNF drugs with success rates of about 50%. Shifting from adalimumab to Infliximab was beneficial in subjects with LOR and untraceable adalimumab levels, and half of them mandated DI after six months and 75% after 12 months of infliximab therapy. At 12 months, 81% still were on infliximab.136 Besides a change of anti-TNF, there is a question of whether adding an immunosuppressive drug will increase response. A study compared switching to a new anti-TNF in IBD subjects after experiencing LOR versus switching to a new anti-TNF treatment with the addition of Azathioprine. The study found that combining Azathioprine with the new anti-TNF treatment was more effective.137
A Second-Generation AI Platform Based on the CDP Can Overcome Drug Tolerance
Immune variation during health and disease must be considered when developing methods to overcome the loss of responsiveness to immunotherapies.36,37,138 In precision medicine or personalized therapy, patients are treated differently based on their disease mechanisms and specific treatment needs. It is possible to achieve better outcomes by determining the requirements for each patient.139 Based on the CDP, the second-generation AI systems are designed to incorporate personalized variability signatures into treatment regimens.140–142 These systems quantify inherent variabilities using biological sensors (eg, heart rate, blood pressure, and respiratory variabilities) or test variabilities (eg, cytokines secretion and subsets of inflammatory cells variabilities) and implement them into therapeutic regimens. This method may overcome the loss of response to chronic treatments.7,66,143–160
Implementing variability signatures in treating patients with congestive heart failure who have developed resistance to diuretics has successfully overcome the resistance. Treated patients showed improved clinical and laboratory parameters, significantly reducing hospitalizations and emergency room admissions.161 Similar beneficial effects were described for patients suffering from chronic pain who developed tolerance to painkillers and those with multiple sclerosis receiving immunotherapy.35 This platform improves the response to Lenvatinib in head and neck cancer patients.162 Long-term studies are required to ensure the sustainability of these effects.
The data shows potential for addressing medication response challenges through personalized variability signatures in treatment plans. Personalization of the treatment plans needs to be dynamic as both the host response and the disease pathogenesis keep changing. It mandates using algorithms, which can alter the variability in a way that deals with changing internal and external environments.34,140–142,163
Changes in Immune Variations as Early Signs for Disease and Monitoring: The Development of Variability-Based Biomarkers for Immune-Associated Diseases
Currently used laboratory tests and clinical scores are insufficient for early diagnosis and monitoring of the response to therapies in subjects with immune-associated diseases. The intra- and intersubject variability of the immune systems creates a significant challenge in identifying early diagnostic and valid monitoring biomarkers.
The lack of proper early diagnostic tools impacts the prognosis of many immunological diseases as it is associated with the late start of therapies.164–167 Similarly, the lack of valid biomarkers provides a challenge for personalizing immunotherapies when faced with a dynamic immune system that alters the pathogenesis of the disease over time within and among patients treated with immunotherapies.168 The current biomarkers are insufficient to select the ideal immunotherapy from the available immunotherapies. Moreover, as the host and the disease change, a dynamic system must provide time-dependent personalizing immunotherapies.169,170
The CDP-based early signs system may provide a variability-based method for the early diagnosis of immune-mediated disorders. Variances in the immune system, if too low or high, can predict disease. Alerts in variability levels can also monitor disease progression and therapy response. Both autoimmune diseases and cancers have a time factor in their pathogenesis and response to immunotherapies, which evolve from host and disease alterations. Therefore, immune-variability signatures are anticipated to offer a more accurate way to follow up on these patients and determine their treatment response.67
Long-term studies using CDP-based AI systems are required to show their use in monitoring the improvement of outcomes.
Using CDP-Based Platforms for Improving the Response to Immune-Based Treatments
The second-generation AI system based on CDP is designed to address the lack of response to immunotherapies and the loss of response. Figure 1 shows a schematic presentation of the changes in variability signatures that can serve as a method for early signs of disease (A) and an schematic presentation of a CDP-based treatment plan based on changing the degree of variability (B). The changes in the variability signatures may enable the selection of immunotherapies and change them over time while serving as biomarkers for disease progression, response to therapies, and prognosis. Box 1 shows several potential biomarkers that can serve as biomarkers.
 |
Box 1 Potential variability signatures as novel biomarkers |
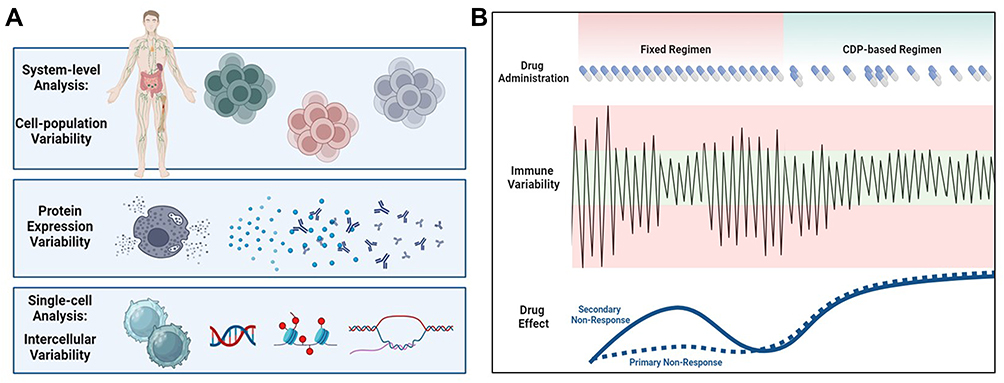 |
Figure 1 (A) Schematic presentation of the hypothesis of how changes in variability signatures can serve as a method for early signs of disease. (B) Schematic presentation of the hypothesized CDP-based treatment plan based on changing the degree of variability. The changes in the variability signatures may enable the selection of immunotherapies and change them over time while serving as biomarkers for disease progression, response to therapies, and prognosis. |
The platform is being developed in three stages. Initially, a pseudorandom number generator picks the administration time and dose from a pre-defined dosing scheme. This open-loop system operates independently of the outcome and is not personalized. For using anti-TNF Adalimumab in colitis or arthritis, the algorithm can select between 40 and 80mg, and a range between ten and sixteen days between administrations, rather than a fixed 80mg every fourteen days, is proposed to improve the response. In the second stage, a closed-loop system is implemented. Clinical and laboratory endpoints provide feedback to select dosing and administration times dynamically. In the third step, the quantification of biological signatures of variability is implemented into the algorithm to improve the response further.140–142
Future studies are designed to evaluate the overarching framework and assess the necessity of potential mergers to improve the clinical outcomes of using this method.
Summary
The immune system’s intra and inter-subject variability is a significant challenge for immunotherapies and current diagnostic and monitoring modalities. The CDP-based second-generation AI system may offer a new platform to address these challenges. Ongoing studies determine the ability to implement these modalities into clinical practice to improve the diagnosis, monitoring, and responsiveness to therapies.
Abbreviations
CDP, constrained disorder principle; ADCs, antibody-drug conjugates; HLA, human leukocyte antigen; TNF, tumor necrosis factor; RA, rheumatoid arthritis; PNR, primary non-response; SLR, secondary loss of response; LOR, loss of response; ADA, anti-drugs antibodies; IBD, inflammatory bowel disease; ICI, immune checkpoint inhibitor; EULAR, European League Against Rheumatism; bDMARD, biologic disease-modifying anti-rheumatic drugs; DI, dose intensification.
Funding
There is no funding to report.
Disclosure
YI is the founder of Oberon Sciences. The authors report no conflicts of interest in this work.
References
1. Li B. Personalized Immunotherapy of Patients: defining by Single-cell RNA-seq with Artificial Intelligence. Med Res Arch. 2023;11:1.
2. Gong Z, Huang X, Cao Q, et al. A CLRN3-Based CD8+ T-Related Gene Signature Predicts Prognosis and Immunotherapy Response in Colorectal Cancer. Biomolecules. 2024;14(8):891. doi:10.3390/biom14080891
3. Melief CJM. Special Review: the future of Immunotherapy. Immunother Adv. 2020;1:ltaa005.
4. Pilard C, Ancion M, Delvenne P, et al. Cancer immunotherapy: it’s time to better predict patients’ response. Br J Cancer. 2021;125(7):927–938. doi:10.1038/s41416-021-01413-x
5. Li J, Liu Z, Hu P, et al. Indicators of suboptimal response to anti-tumor necrosis factor therapy in patients from China with inflammatory bowel disease: results from the EXPLORE study. BMC Gastroenterol. 2022;22(1):44. doi:10.1186/s12876-021-02074-z
6. Gisbert JP, Chaparro M. Primary Failure to an Anti-TNF Agent in Inflammatory Bowel Disease: switch (to a Second Anti-TNF Agent) or Swap (for Another Mechanism of Action)? J Clin Med. 2021;11(1):10. doi:10.3390/jcm11010010
7. Khoury T, Ilan Y. Introducing Patterns of Variability for Overcoming Compensatory Adaptation of the Immune System to Immunomodulatory Agents: a Novel Method for Improving Clinical Response to Anti-TNF Therapies. Front Immunol. 2019;10:10. doi:10.3389/fimmu.2019.00010
8. Ilan Y. The constrained disorder principle defines living organisms and provides a method for correcting disturbed biological systems. Comput Struct Biotechnol J. 2022;20:6087–6096. doi:10.1016/j.csbj.2022.11.015
9. Finn EH, Misteli T. Molecular basis and biological function of variability in spatial genome organization. Science. 2019;365(6457). doi:10.1126/science.aaw9498
10. Chiera M, Cerritelli F, Casini A, et al. Heart Rate Variability in the Perinatal Period: a Critical and Conceptual Review. Front Neurosci. 2020;14:561186. doi:10.3389/fnins.2020.561186
11. Forte G, Favieri F, Casagrande M. Heart Rate Variability and Cognitive Function: a Systematic Review. Front Neurosci. 2019;13:710. doi:10.3389/fnins.2019.00710
12. Mitchison T, Kirschner M. Dynamic instability of microtubule growth. Nature. 1984;312(5991):237–242. doi:10.1038/312237a0
13. Kirschner MW, Mitchison T. Microtubule dynamics. Nature. 1986;324(6098):621. doi:10.1038/324621a0
14. Schutte AE, Kollias A, Stergiou GS. Blood pressure and its variability: classic and novel measurement techniques. Nat Rev Cardiol. 2022;19(10):643–654. doi:10.1038/s41569-022-00690-0
15. van den Bosch OFC, Alvarez-Jimenez R, de Grooth HJ, et al. Breathing variability-implications for anaesthesiology and intensive care. Crit Care. 2021;25(1):280. doi:10.1186/s13054-021-03716-0
16. Boripuntakul S, Kamnardsiri T, Lord SR, et al. Gait variability during abrupt slow and fast speed transitions in older adults with mild cognitive impairment. PLoS One. 2022;17(10):e0276658. doi:10.1371/journal.pone.0276658
17. Genon S, Eickhoff SB, Kharabian S. Linking interindividual variability in brain structure to behaviour. Nat Rev Neurosci. 2022;23(5):307–318. doi:10.1038/s41583-022-00584-7
18. Saha S, Baumert M. Intra- and Inter-subject Variability in EEG-Based Sensorimotor Brain Computer Interface: a Review. Front Comput Neurosci. 2020;13:87. doi:10.3389/fncom.2019.00087
19. Crawford L, Mills E, Meylakh N, et al. Brain activity changes associated with pain perception variability. Cereb Cortex. 2022;33:4145–4155.
20. Ilan Y. Overcoming randomness does not rule out the importance of inherent randomness for functionality. J Biosci. 2019;44(2):44.
21. Ilan Y. Generating randomness: making the most out of disordering a false order into a real one. J Transl Med. 2019;17(1):49. doi:10.1186/s12967-019-1798-2
22. Ilan Y. Advanced Tailored Randomness: a Novel Approach for Improving the Efficacy of Biological Systems. J Comput Biol. 2020;27(1):20–29. doi:10.1089/cmb.2019.0231
23. Ilan Y. Order Through Disorder: the Characteristic Variability of Systems. Front Cell Dev Biol. 2020;8:186. doi:10.3389/fcell.2020.00186
24. El-Haj M, Kanovitch D, Ilan Y. Personalized inherent randomness of the immune system is manifested by an individualized response to immune triggers and immunomodulatory therapies: a novel platform for designing personalized immunotherapies. Immunol Res. 2019;67(4–5):337–347. doi:10.1007/s12026-019-09101-y
25. Ilan Y. Randomness in microtubule dynamics: an error that requires correction or an inherent plasticity required for normal cellular function? Cell Biol Int. 2019;43(7):739–748. doi:10.1002/cbin.11157
26. Ilan Y. Microtubules: from understanding their dynamics to using them as potential therapeutic targets. J Cell Physiol. 2019;234(6):7923–7937. doi:10.1002/jcp.27978
27. Ilan-Ber T, Ilan Y. The role of microtubules in the immune system and as potential targets for gut-based immunotherapy. Mol Immunol. 2019;111:73–82. doi:10.1016/j.molimm.2019.04.014
28. Forkosh E, Kenig A, Ilan Y. Introducing variability in targeting the microtubules: review of current mechanisms and future directions in colchicine therapy. Pharmacol Res Perspect. 2020;8(4):e00616. doi:10.1002/prp2.616
29. Ilan Y. β-Glycosphingolipids as Mediators of Both Inflammation and Immune Tolerance: a Manifestation of Randomness in Biological Systems. Front Immunol. 2019;10:1143. doi:10.3389/fimmu.2019.01143
30. Ilan Y. Microtubules as a potential platform for energy transfer in biological systems: a target for implementing individualized, dynamic variability patterns to improve organ function. Mol Cell Biochem. 2022;2022:1.
31. Ilan Y. Enhancing the plasticity, proper function and efficient use of energy of the Sun, genes and microtubules using variability. Clini Transl Discovery. 2022;2(4):e103. doi:10.1002/ctd2.103
32. Shabat Y, Lichtenstein Y, Ilan Y. Short-Term Cohousing of Sick with Healthy or Treated Mice Alleviates the Inflammatory Response and Liver Damage. Inflammation. 2021;44(2):518–525. doi:10.1007/s10753-020-01348-0
33. Ilan Y. Making use of noise in biological systems. Prog Biophys Mol Biol. 2023;178:83–90. doi:10.1016/j.pbiomolbio.2023.01.001
34. Ilan Y. Constrained disorder principle-based variability is fundamental for biological processes: beyond biological relativity and physiological regulatory networks. Prog Biophys Mol Biol. 2023;180–181:37–48. doi:10.1016/j.pbiomolbio.2023.04.003
35. Sigawi T, Lehmann H, Hurvitz N, et al. Constrained disorder principle-based second-generation algorithms implement quantified variability signatures to improve the function of complex systems. J Bioinform Syst Biol. 2023;6(01):82–89. doi:10.26502/jbsb.5107051
36. Brodin P, Davis MM. Human immune system variation. Nat Rev Immunol. 2017;17(1):21–29. doi:10.1038/nri.2016.125
37. Liston A, Humblet-Baron S, Duffy D, et al. Human immune diversity: from evolution to modernity. Nat Immunol. 2021;22(12):1479–1489. doi:10.1038/s41590-021-01058-1
38. Barreiro LB, Quintana-Murci L. Evolutionary and population (epi)genetics of immunity to infection. Hum Genet. 2020;139(6–7):723–732. doi:10.1007/s00439-020-02167-x
39. Liston A, Carr EJ, Linterman MA. Shaping Variation in the Human Immune System. Trend Immunol. 2016;37(10):637–646. doi:10.1016/j.it.2016.08.002
40. Wiertsema SP, Van bergenhenegouwen J, Garssen J, et al. The Interplay between the Gut Microbiome and the Immune System in the Context of Infectious Diseases throughout Life and the Role of Nutrition in Optimizing Treatment Strategies. Nutrients. 2021;13(3):886. doi:10.3390/nu13030886
41. Lakshmikanth T, Muhammad SA, Olin A, et al. Human Immune System Variation during 1 Year. Cell Rep. 2020;32(3):107923. doi:10.1016/j.celrep.2020.107923
42. Aguirre-Gamboa R, Joosten I, Urbano PCM, et al. Differential Effects of Environmental and Genetic Factors on T and B Cell Immune Traits. Cell Rep. 2016;17(9):2474–2487. doi:10.1016/j.celrep.2016.10.053
43. Booth JS, Toapanta FR. B and T Cell Immunity in Tissues and Across the Ages. Vaccines. 2021;9:24.
44. Brodin P, Jojic V, Gao T, et al. Variation in the human immune system is largely driven by non-heritable influences. Cell. 2015;160(1–2):37–47. doi:10.1016/j.cell.2014.12.020
45. Li Y, Oosting M, Deelen P, et al. Inter-individual variability and genetic influences on cytokine responses to bacteria and fungi. Nature Med. 2016;22(8):952–960. doi:10.1038/nm.4139
46. Quintana M, Palicki O, Lucchi G, et al. Inter-individual variability of protein patterns in saliva of healthy adults. J Proteomics. 2009;72(5):822–830. doi:10.1016/j.jprot.2009.05.004
47. Alpert A, Pickman Y, Leipold M, et al. A clinically meaningful metric of immune age derived from high-dimensional longitudinal monitoring. Nat Med. 2019;25(3):487–495. doi:10.1038/s41591-019-0381-y
48. Simon AK, Hollander GA, McMichael A. Evolution of the immune system in humans from infancy to old age. Proc R Soc B. 2015;282(1821):20143085. doi:10.1098/rspb.2014.3085
49. Zhao J, Guo S, Schrodi SJ, et al. Molecular and Cellular Heterogeneity in Rheumatoid Arthritis: mechanisms and Clinical Implications. Front Immunol. 2021;12:790122. doi:10.3389/fimmu.2021.790122
50. Carr EJ, Dooley J, Garcia-Perez JE, et al. The cellular composition of the human immune system is shaped by age and cohabitation. Nat Immunol. 2016;17(4):461–468. doi:10.1038/ni.3371
51. Liefferinckx C, De Greve Z, Toubeau JF, et al. New approach to determine the healthy immune variations by combining clustering methods. Sci Rep. 2021;11(1):8917. doi:10.1038/s41598-021-88272-x
52. Ter Horst R, Jaeger M, Smeekens SP, et al. Host and Environmental Factors Influencing Individual Human Cytokine Responses. Cell. 2016;167(4):1111–1124e13. doi:10.1016/j.cell.2016.10.018
53. Tsang JS. Utilizing population variation, vaccination, and systems biology to study human immunology. Trends Immunol. 2015;36(8):479–493. doi:10.1016/j.it.2015.06.005
54. Cevirgel A, Shetty SA, Vos M, et al. Identification of aging-associated immunotypes and immune stability as indicators of post-vaccination immune activation. Aging Cell. 2022;21(10):e13703. doi:10.1111/acel.13703
55. Burel JG, Qian Y, Lindestam Arlehamn C, et al. An Integrated Workflow To Assess Technical and Biological Variability of Cell Population Frequencies in Human Peripheral Blood by Flow Cytometry. J Immunol. 2017;198(4):1748–1758. doi:10.4049/jimmunol.1601750
56. Mueller SC, Marz R, Schmolz M, et al. Intraindividual long term stability and response corridors of cytokines in healthy volunteers detected by a standardized whole-blood culture system for bed-side application. BMC Med Res Methodol. 2012;12(1):112. doi:10.1186/1471-2288-12-112
57. Kaczorowski KJ, Shekhar K, Nkulikiyimfura D, et al. Continuous immunotypes describe human immune variation and predict diverse responses. Proc Natl Acad Sci. 2017;114(30):E6097–E6106. doi:10.1073/pnas.1705065114
58. Ryan FJ, Hope CM, Masavuli MG, et al. Long-term perturbation of the peripheral immune system months after SARS-CoV-2 infection. BMC Med. 2022;20(1):26. doi:10.1186/s12916-021-02228-6
59. Olin A, Henckel E, Chen Y, et al. Stereotypic Immune System Development in Newborn Children. Cell. 2018;174(5):1277–1292.e14. doi:10.1016/j.cell.2018.06.045
60. Kanodia P, Kaur G, Coshic P, et al. Characterization of biological variation of peripheral blood immune cytome in an Indian cohort. Sci Rep. 2019;9(1):14735. doi:10.1038/s41598-019-51294-7
61. Pinti M, Appay V, Campisi J, et al. Aging of the immune system: focus on inflammation and vaccination. Eur J Immunol. 2016;46(10):2286–2301. doi:10.1002/eji.201546178
62. Aging Biomarker C, Bao H, Cao J, et al. Biomarkers of aging. Sci China Life Sci. 2023;66(5):893–1066. doi:10.1007/s11427-023-2305-0
63. Shen-Orr SS, Furman D. Variability in the immune system: of vaccine responses and immune states. Curr Opin Immunol. 2013;25(4):542–547. doi:10.1016/j.coi.2013.07.009
64. Shaffer SM, Dunagin MC, Torborg SR, et al. Rare cell variability and drug-induced reprogramming as a mode of cancer drug resistance. Nature. 2017;546(7658):431–435. doi:10.1038/nature22794
65. Satija R, Shalek AK. Heterogeneity in immune responses: from populations to single cells. Trends Immunol. 2014;35(5):219–229. doi:10.1016/j.it.2014.03.004
66. Hurvitz N, Elkhateeb N, Sigawi T, et al. Improving the effectiveness of anti-aging modalities by using the constrained disorder principle-based management algorithms. Front Aging. 2022;3:1044038. doi:10.3389/fragi.2022.1044038
67. Sigawi T, Ilan Y. Using Constrained-Disorder Principle-Based Systems to Improve the Performance of Digital Twins in Biological Systems. Biomimetics. 2023;8:359.
68. Van Eyndhoven LC, Tel J. Revising immune cell coordination: origins and importance of single-cell variation. Eur J Immunol. 2022;52(12):1889–1897. doi:10.1002/eji.202250073
69. Lee S, Vu HM, Lee JH, et al. Advances in Mass Spectrometry-Based Single Cell Analysis. Biology. 2023;12:395.
70. Lu Y, Biancotto A, Cheung F, et al. Systematic Analysis of Cell-to-Cell Expression Variation of T Lymphocytes in a Human Cohort Identifies Aging and Genetic Associations. Immunity. 2016;45(5):1162–1175. doi:10.1016/j.immuni.2016.10.025
71. Kashima Y, Kaneko K, Reteng P, et al. Intensive single-cell analysis reveals immune-cell diversity among healthy individuals. Life Sci Alliance. 2022;5:1.
72. Papalexi E, Satija R. Single-cell RNA sequencing to explore immune cell heterogeneity. Nat Rev Immunol. 2018;18(1):35–45. doi:10.1038/nri.2017.76
73. Amanat F, Thapa M, Lei T, et al. The plasmablast response to SARS-CoV-2 mRNA vaccination is dominated by non-neutralizing antibodies and targets both the NTD and the RBD. medRxiv. 2021;2021:1.
74. Henn AD, Wu S, Qiu X, et al. High-resolution temporal response patterns to influenza vaccine reveal a distinct human plasma cell gene signature. Sci Rep. 2013;3(1):2327. doi:10.1038/srep02327
75. Mettelman RC, Allen EK, Thomas PG. Mucosal immune responses to infection and vaccination in the respiratory tract. Immunity. 2022;55(5):749–780. doi:10.1016/j.immuni.2022.04.013
76. Crow MK. Interferon-alpha: a therapeutic target in systemic lupus erythematosus. Rheum Dis Clin North Am. 2010;36(1):173–186. doi:10.1016/j.rdc.2009.12.008
77. Martinez-Jimenez CP, Eling N, Chen HC, et al. Aging increases cell-to-cell transcriptional variability upon immune stimulation. Science. 2017;355(6332):1433–1436. doi:10.1126/science.aah4115
78. Nair N, Mei HE, Chen SY, et al. Mass cytometry as a platform for the discovery of cellular biomarkers to guide effective rheumatic disease therapy. Arthritis Res Ther. 2015;17(1):127. doi:10.1186/s13075-015-0644-z
79. Zhang Y, Lee TY. Revealing the Immune Heterogeneity between Systemic Lupus Erythematosus and Rheumatoid Arthritis Based on Multi-Omics Data Analysis. Int J Mol Sci. 2022;23:5166.
80. Zheng F, Xu H, Zhang C, et al. Immune cell and TCR/BCR repertoire profiling in systemic lupus erythematosus patients by single-cell sequencing. Aging. 2021;13(21):24432–24448. doi:10.18632/aging.203695
81. Ochocka N, Kaminska B. Microglia Diversity in Healthy and Diseased Brain: insights from Single-Cell Omics. Int J Mol Sci. 2021;22:3027.
82. van Unen V, Li N, Molendijk I, et al. Mass Cytometry of the Human Mucosal Immune System Identifies Tissue- and Disease-Associated Immune Subsets. Immunity. 2016;44(5):1227–1239. doi:10.1016/j.immuni.2016.04.014
83. Stewart E, Wang X, Chupp GL, et al. Profiling cellular heterogeneity in asthma with single cell multiparameter CyTOF. J Leukoc Biol. 2020;108(5):1555–1564. doi:10.1002/JLB.5MA0720-770RR
84. Martinez-Morilla S, Villarroel-Espindola F, Wong PF, et al. Biomarker Discovery in Patients with Immunotherapy-Treated Melanoma with Imaging Mass Cytometry. Clin Cancer Res. 2021;27(7):1987–1996. doi:10.1158/1078-0432.CCR-20-3340
85. Zhu YP, Eggert T, Araujo DJ, et al. CyTOF mass cytometry reveals phenotypically distinct human blood neutrophil populations differentially correlated with melanoma stage. J Immunother Cancer. 2020;8:1.
86. Altschuler SJ, Wu LF. Cellular heterogeneity: do differences make a difference? Cell. 2010;141:559–563. doi:10.1016/j.cell.2010.04.033
87. Kuret T, Sodin-Semrl S, Leskosek B, et al. Single Cell RNA Sequencing in Autoimmune Inflammatory Rheumatic Diseases: current Applications, Challenges and a Step Toward Precision Medicine. Front Med Lausanne. 2021;8:822804. doi:10.3389/fmed.2021.822804
88. Summers RL. Entropic Dynamics in a Theoretical Framework for Biosystems. Entropy. 2023;25:528.
89. Phillips KP, Cable J, Mohammed RS, et al. Immunogenetic novelty confers a selective advantage in host–pathogen coevolution. Proc Natl Acad Sci. 2018;115(7):1552–1557. doi:10.1073/pnas.1708597115
90. Muraille E. Generation of individual diversity: a too neglected fundamental property of adaptive immune system. Front Immunol. 2014;5:208. doi:10.3389/fimmu.2014.00208
91. van Houte S, Buckling A, Westra ER. Evolutionary Ecology of Prokaryotic Immune Mechanisms. Microbiol Mol Biol Rev. 2016;80(3):745–763. doi:10.1128/MMBR.00011-16
92. Cobey S. Pathogen evolution and the immunological niche. Ann N Y Acad Sci. 2014;1320(1):1–15. doi:10.1111/nyas.12493
93. Noble D. The role of stochasticity in biological communication processes. Prog Biophys Mol Biol. 2021;162:122–128. doi:10.1016/j.pbiomolbio.2020.09.008
94. Jawa V, Terry F, Gokemeijer J, et al. T-Cell Dependent Immunogenicity of Protein Therapeutics Pre-clinical Assessment and Mitigation–Updated Consensus and Review 2020. Front Immunol. 2020;11:1301. doi:10.3389/fimmu.2020.01301
95. Vaidehi Narayanan H, Hoffmann A. From Antibody Repertoires to Cell-Cell Interactions to Molecular Networks: bridging Scales in the Germinal Center. Front Immunol. 2022;13:1.
96. Schijns V, Fernández-Tejada A, Barjaktarović Ž, et al. Modulation of immune responses using adjuvants to facilitate therapeutic vaccination. Immunol Rev. 2020;296(1):169–190. doi:10.1111/imr.12889
97. Pulendran B, Arunachalam S, O’Hagan DT. Emerging concepts in the science of vaccine adjuvants. Nat Rev Drug Discov. 2021;20(6):454–475. doi:10.1038/s41573-021-00163-y
98. Kreitinger JM, Beamer CA, Shepherd DM. Environmental Immunology: lessons Learned from Exposure to a Select Panel of Immunotoxicants. J Immunol. 2016;196(8):3217–3225. doi:10.4049/jimmunol.1502149
99. Netea MG, Domínguez-Andrés J, Barreiro LB, et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol. 2020;20(6):375–388. doi:10.1038/s41577-020-0285-6
100. Bai R, Chen N, Li L, et al. Mechanisms of Cancer Resistance to Immunotherapy. Front Oncol. 2020;10:1290. doi:10.3389/fonc.2020.01290
101. Restifo NP, Smyth MJ, Snyder A. Acquired resistance to immunotherapy and future challenges. Nat Rev Cancer. 2016;16(2):121–126. doi:10.1038/nrc.2016.2
102. Roda G, Jharap B, Neeraj N, et al. Loss of Response to Anti-TNFs: definition, Epidemiology, and Management. Clin Transl Gastroenterol. 2016;7(1):e135. doi:10.1038/ctg.2015.63
103. Vaisman-Mentesh A, Gutierrez-Gonzalez M, DeKosky BJ, et al. The Molecular Mechanisms That Underlie the Immune Biology of Anti-drug Antibody Formation Following Treatment With Monoclonal Antibodies. Front Immunol. 2020;11:1951. doi:10.3389/fimmu.2020.01951
104. Vultaggio A, Petroni G, Pratesi S, et al. Circulating T cells to infliximab are detectable mainly in treated patients developing anti-drug antibodies and hypersensitivity reactions. Clin Exp Immunol. 2016;186(3):364–372. doi:10.1111/cei.12858
105. Brun MK, Goll GL, Jorgensen KK, et al. Risk factors for anti-drug antibody formation to infliximab: secondary analyses of a randomised controlled trial. J Intern Med. 2022;292(3):477–491. doi:10.1111/joim.13495
106. Ben-Horin S, Kopylov U, Chowers Y. Optimizing anti-TNF treatments in inflammatory bowel disease. Autoimmun Rev. 2014;13(1):24–30. doi:10.1016/j.autrev.2013.06.002
107. Fine S, Papamichael K, Cheifetz AS. Etiology and Management of Lack or Loss of Response to Anti-Tumor Necrosis Factor Therapy in Patients With Inflammatory Bowel Disease. Gastroenterol Hepatol. 2019;15(12):656–665.
108. Allez M, Karmiris K, Louis E, et al. Report of the ECCO pathogenesis workshop on anti-TNF therapy failures in inflammatory bowel diseases: definitions, frequency and pharmacological aspects. J Crohns Colitis. 2010;4(4):355–366. doi:10.1016/j.crohns.2010.04.004
109. Kennedy NA, Heap GA, Green HD, et al. Predictors of anti-TNF treatment failure in anti-TNF-naive patients with active luminal Crohn’s disease: a prospective, multicentre, cohort study. Lancet Gastroenterol Hepatol. 2019;4(5):341–353. doi:10.1016/S2468-1253(19)30012-3
110. Johnson KJ, Sanchez HN, Schoenbrunner N. Defining response to TNF-inhibitors in rheumatoid arthritis: the negative impact of anti-TNF cycling and the need for a personalized medicine approach to identify primary non-responders. Clin Rheumatol. 2019;38(11):2967–2976. doi:10.1007/s10067-019-04684-1
111. de Punder YM, Fransen J, Kievit W, et al. The prevalence of clinical remission in RA patients treated with anti-TNF: results from the Dutch Rheumatoid Arthritis Monitoring (DREAM) registry. Rheumatology. 2012;51(9):1610–1617. doi:10.1093/rheumatology/kes078
112. Tao W, Concepcion AN, Vianen M, et al. Multiomics and Machine Learning Accurately Predict Clinical Response to Adalimumab and Etanercept Therapy in Patients With Rheumatoid Arthritis. Arthritis Rheumatol. 2021;73(2):212–222. doi:10.1002/art.41516
113. Kalden JR, Schulze-Koops H. Immunogenicity and loss of response to TNF inhibitors: implications for rheumatoid arthritis treatment. Nat Rev Rheumatol. 2017;13(12):707–718. doi:10.1038/nrrheum.2017.187
114. Fafa BP, Louzada-Junior P, Titton DC, et al. Drug survival and causes of discontinuation of the first anti-TNF in ankylosing spondylitis compared with rheumatoid arthritis: analysis from BIOBADABRASIL. Clin Rheumatol. 2015;34(5):921–927. doi:10.1007/s10067-015-2929-7
115. Bagchi S, Yuan R, Engleman EG. Immune Checkpoint Inhibitors for the Treatment of Cancer: clinical Impact and Mechanisms of Response and Resistance. Annu Rev Pathol. 2021;16(1):223–249. doi:10.1146/annurev-pathol-042020-042741
116. Garon EB, Rizvi NA, Hui R, et al. Pembrolizumab for the treatment of non-small-cell lung cancer. N Engl J Med. 2015;372(21):2018–2028. doi:10.1056/NEJMoa1501824
117. Haslam A, Prasad V. Estimation of the Percentage of US Patients With Cancer Who Are Eligible for and Respond to Checkpoint Inhibitor Immunotherapy Drugs. JAMA Netw Open. 2019;2(5):e192535. doi:10.1001/jamanetworkopen.2019.2535
118. Lee KA, Thomas AM, Bolte LA, et al. Cross-cohort gut microbiome associations with immune checkpoint inhibitor response in advanced melanoma. Nat Med. 2022;28(3):535–544. doi:10.1038/s41591-022-01695-5
119. Perez-Ruiz E, Melero I, Kopecka J, et al. Cancer immunotherapy resistance based on immune checkpoints inhibitors: targets, biomarkers, and remedies. Drug Resist Updat. 2020;53:100718. doi:10.1016/j.drup.2020.100718
120. Fares CM, Van Allen EM, Drake CG, et al. Mechanisms of Resistance to Immune Checkpoint Blockade: why Does Checkpoint Inhibitor Immunotherapy Not Work for All Patients? Am Soc Clin Oncol Educ Book. 2019;39(39):147–164. doi:10.1200/EDBK_240837
121. Walsh RJ, Soo RA. Resistance to immune checkpoint inhibitors in non-small cell lung cancer: biomarkers and therapeutic strategies. Ther Adv Med Oncol. 2020;12:1758835920937902. doi:10.1177/1758835920937902
122. Bai F, Zhang P, Fu Y, et al. Targeting ANXA1 abrogates Treg-mediated immune suppression in triple-negative breast cancer. J Immunother Cancer. 2020;8:1.
123. Goyette MA, Elkholi IE, Apcher C, et al. Targeting Axl favors an antitumorigenic microenvironment that enhances immunotherapy responses by decreasing Hif-1alpha levels. Proc Natl Acad Sci U S A. 2021;118:e2023868118.
124. Retecki K, Seweryn M, Graczyk-Jarzynka A, et al. The Immune Landscape of Breast Cancer: strategies for Overcoming Immunotherapy Resistance. Cancers. 2021;14(1):13. doi:10.3390/cancers14010013
125. Gilad Y, Eliaz Y, Yu Y, et al. Drug-induced PD-L1 expression and cell stress response in breast cancer cells can be balanced by drug combination. Sci Rep. 2019;9(1):15099. doi:10.1038/s41598-019-51537-7
126. Bailly C, Thuru X, Quesnel B. Combined cytotoxic chemotherapy and immunotherapy of cancer: modern times. NAR Cancer. 2020;2(1):zcaa002. doi:10.1093/narcan/zcaa002
127. Bachem A, Makhlouf C, Binger KJ, et al. Microbiota-Derived Short-Chain Fatty Acids Promote the Memory Potential of Antigen-Activated CD8(+) T Cells. Immunity. 2019;51(2):285–297e5. doi:10.1016/j.immuni.2019.06.002
128. Skelly AN, Sato Y, Kearney S, et al. Mining the microbiota for microbial and metabolite-based immunotherapies. Nat Rev Immunol. 2019;19(5):305–323. doi:10.1038/s41577-019-0144-5
129. Huang J, Jiang Z, Wang Y, et al. Modulation of gut microbiota to overcome resistance to immune checkpoint blockade in cancer immunotherapy. Curr Opin Pharmacol. 2020;54:1–10. doi:10.1016/j.coph.2020.06.004
130. Rubbert-Roth A, Szabo MZ, Kedves M, et al. Failure of anti-TNF treatment in patients with rheumatoid arthritis: the pros and cons of the early use of alternative biological agents. Autoimmun Rev. 2019;18(12):102398. doi:10.1016/j.autrev.2019.102398
131. Smolen JS, Kay J, Doyle MK, et al. Golimumab in patients with active rheumatoid arthritis after treatment with tumour necrosis factor alpha inhibitors (GO-AFTER study): a multicentre, randomised, double-blind, placebo-controlled, Phase III trial. Lancet. 2009;374(9685):210–221. doi:10.1016/S0140-6736(09)60506-7
132. Genovese MC, Becker JC, Schiff M, et al. Abatacept for rheumatoid arthritis refractory to tumor necrosis factor alpha inhibition. N Engl J Med. 2005;353(11):1114–1123. doi:10.1056/NEJMoa050524
133. Shipa MRA, Di Cicco M, Balogh E, et al. Drug-survival profiling of second-line biologic therapy in rheumatoid arthritis: choice of another tumour necrosis factor inhibitor or a biologic of different mode of action? Mod Rheumatol. 2023;33(4):700–707. doi:10.1093/mr/roac086
134. Gottenberg JE, Brocq O, Perdriger A, et al. Non-TNF-Targeted Biologic vs a Second Anti-TNF Drug to Treat Rheumatoid Arthritis in Patients With Insufficient Response to a First Anti-TNF Drug: a Randomized Clinical Trial. JAMA. 2016;316(11):1172–1180. doi:10.1001/jama.2016.13512
135. Guberna L, Nyssen OP, Chaparro M, et al. Frequency and Effectiveness of Empirical Anti-TNF Dose Intensification in Inflammatory Bowel Disease: systematic Review with Meta-Analysis. J Clin Med. 2021;10:2132.
136. Peeters H, Louis E, Baert F, et al. Efficacy of switching to infliximab in patients with Crohn’s disease with loss of response to Adalimumab. Acta Gastroenterol Belg. 2018;81:15–21.
137. Roblin X, Williet N, Boschetti G, et al. Addition of azathioprine to the switch of anti-TNF in patients with IBD in clinical relapse with undetectable anti-TNF trough levels and antidrug antibodies: a prospective randomised trial. Gut. 2020;69(7):1206–1212. doi:10.1136/gutjnl-2019-319758
138. Wagar LE, DiFazio RM, Davis MM. Advanced model systems and tools for basic and translational human immunology. Genome Med. 2018;10(1):73. doi:10.1186/s13073-018-0584-8
139. Strianese O, Rizzo F, Ciccarelli M, et al. Precision and Personalized Medicine: how Genomic Approach Improves the Management of Cardiovascular and Neurodegenerative Disease. Genes. 2020;12(1):11. doi:10.3390/genes12010011
140. Ilan Y. Second-Generation Digital Health Platforms: placing the Patient at the Center and Focusing on Clinical Outcomes. Front Digit Health. 2020;2:569178. doi:10.3389/fdgth.2020.569178
141. Ilan Y. Improving Global Healthcare and Reducing Costs Using Second-Generation Artificial Intelligence-Based Digital Pills: a Market Disruptor. Int J Environ Res Public Health. 2021;19(1):18. doi:10.3390/ijerph19010018
142. Ilan Y. Next-Generation Personalized Medicine: implementation of Variability Patterns for Overcoming Drug Resistance in Chronic Diseases. J Pers Med. 2022;12:1303.
143. Kessler A, Weksler-Zangen S, Ilan Y. Role of the Immune System and the Circadian Rhythm in the Pathogenesis of Chronic Pancreatitis: establishing a Personalized Signature for Improving the Effect of Immunotherapies for Chronic Pancreatitis. Pancreas. 2020;49(8):1024–1032. doi:10.1097/MPA.0000000000001626
144. Ishay Y, Kolben Y, Kessler A, et al. Role of circadian rhythm and autonomic nervous system in liver function: a hypothetical basis for improving the management of hepatic encephalopathy. Am J Physiol Gastrointest Liver Physiol. 2021;321(4):G400–G412. doi:10.1152/ajpgi.00186.2021
145. Kolben Y, Weksler-Zangen S, Ilan Y. Adropin as a potential mediator of the metabolic system-autonomic nervous system-chronobiology axis: implementing a personalized signature-based platform for chronotherapy. Obes Rev. 2021;22(2):e13108. doi:10.1111/obr.13108
146. Kenig A, Kolben Y, Asleh R, et al. Improving Diuretic Response in Heart Failure by Implementing a Patient-Tailored Variability and Chronotherapy-Guided Algorithm. Front Cardiovasc Med. 2021;8:695547. doi:10.3389/fcvm.2021.695547
147. Azmanov H, Ross EL, Ilan Y. Establishment of an Individualized Chronotherapy, Autonomic Nervous System, and Variability-Based Dynamic Platform for Overcoming the Loss of Response to Analgesics. Pain Physician. 2021;24:243–252.
148. Potruch A, Khoury ST, Ilan Y. The role of chronobiology in drug-resistance epilepsy: the potential use of a variability and chronotherapy-based individualized platform for improving the response to anti-seizure drugs. Seizure. 2020;80:201–211. doi:10.1016/j.seizure.2020.06.032
149. Isahy Y, Ilan Y. Improving the long-term response to antidepressants by establishing an individualized platform based on variability and chronotherapy. Int J Clin Pharmacol Ther. 2021;59(12):768–774. doi:10.5414/CP204000
150. Khoury T, Ilan Y. Platform introducing individually tailored variability in nerve stimulations and dietary regimen to prevent weight regain following weight loss in patients with obesity. Obes Res Clin Pract. 2021;15(2):114–123. doi:10.1016/j.orcp.2021.02.003
151. Kenig A, Ilan Y. A Personalized Signature and Chronotherapy-Based Platform for Improving the Efficacy of Sepsis Treatment. Front Physiol. 2019;10:1542. doi:10.3389/fphys.2019.01542
152. Ilan Y. Why targeting the microbiome is not so successful: can randomness overcome the adaptation that occurs following gut manipulation? Clin Exp Gastroenterol. 2019;12:209–217. doi:10.2147/CEG.S203823
153. Gelman R, Bayatra A, Kessler A, et al. Targeting SARS-CoV-2 receptors as a means for reducing infectivity and improving antiviral and immune response: an algorithm-based method for overcoming resistance to antiviral agents. Emerg Microbes Infect. 2020;9(1):1397–1406. doi:10.1080/22221751.2020.1776161
154. Ishay Y, Potruch A, Schwartz A, et al. A digital health platform for assisting the diagnosis and monitoring of COVID-19 progression: an adjuvant approach for augmenting the antiviral response and mitigating the immune-mediated target organ damage. Biomed Pharmacother. 2021;143:112228. doi:10.1016/j.biopha.2021.112228
155. Ilan Y, Spigelman Z. Establishing patient-tailored variability-based paradigms for anti-cancer therapy: using the inherent trajectories which underlie cancer for overcoming drug resistance. Cancer Treat Res Commun. 2020;25:100240. doi:10.1016/j.ctarc.2020.100240
156. Hurvitz N, Azmanov H, Kesler A, et al. Establishing a second-generation artificial intelligence-based system for improving diagnosis, treatment, and monitoring of patients with rare diseases. Eur J Hum Genet. 2021;29(10):1485–1490. doi:10.1038/s41431-021-00928-4
157. Ilan Y. Digital Medical Cannabis as Market Differentiator: second-Generation Artificial Intelligence Systems to Improve Response. Front Med Lausanne. 2021;8:788777. doi:10.3389/fmed.2021.788777
158. Gelman R, Berg M, Ilan Y. A Subject-Tailored Variability-Based Platform for Overcoming the Plateau Effect in Sports Training: a Narrative Review. Int J Environ Res Public Health. 2022;19(3):1722. doi:10.3390/ijerph19031722
159. Azmanov H, Bayatra A, Ilan Y. Digital Analgesic Comprising a Second-Generation Digital Health System: increasing Effectiveness by Optimizing the Dosing and Minimizing Side Effects. J Pain Res. 2022;15:1051–1060. doi:10.2147/JPR.S356319
160. Kolben Y, Azmanov H, Gelman R, et al. Using chronobiology-based second-generation artificial intelligence digital system for overcoming antimicrobial drug resistance in chronic infections. Ann Med. 2023;55(1):311–318. doi:10.1080/07853890.2022.2163053
161. Gelman R, Hurvitz N, Nesserat R, et al. A second-generation artificial intelligence-based therapeutic regimen improves diuretic resistance in heart failure: results of a feasibility open-labeled clinical trial. Biomed Pharmacother. 2023;161:114334. doi:10.1016/j.biopha.2023.114334
162. Sigawi T, Gelman R, Maimon O, et al. Improving the response to lenvatinib in partial responders using a Constrained-Disorder-Principle-based second-generation artificial intelligence-therapeutic regimen: a proof-of-concept open-labeled clinical trial. Front Oncol. 2024;14:1426426. doi:10.3389/fonc.2024.1426426
163. Ilan Y. Overcoming Compensatory Mechanisms toward Chronic Drug Administration to Ensure Long-Term, Sustainable Beneficial Effects. Mol Ther Methods Clin Dev. 2020;18:335–344. doi:10.1016/j.omtm.2020.06.006
164. Castro C, Gourley M. Diagnostic testing and interpretation of tests for autoimmunity. J Allergy Clin Immunol. 2010;125(2):S238–47. doi:10.1016/j.jaci.2009.09.041
165. Lenti MV, Miceli E, Cococcia S, et al. Determinants of diagnostic delay in autoimmune atrophic gastritis. Aliment Pharmacol Ther. 2019;50(2):167–175. doi:10.1111/apt.15317
166. Rosa JE, García MV, Luissi A, et al. Rheumatoid Arthritis Patient’s Journey: delay in Diagnosis and Treatment. J Clinic Rheumatol. 2020;26:s148–s152.
167. Lee DW, Koo JS, Choe JW, et al. Diagnostic delay in inflammatory bowel disease increases the risk of intestinal surgery. World J Gastroenterol. 2017;23(35):6474–6481. doi:10.3748/wjg.v23.i35.6474
168. Wang D-R, X-L W, Sun Y-L. Therapeutic targets and biomarkers of tumor immunotherapy: response versus non-response. Signal Transduc Target Ther. 2022;7(1):331. doi:10.1038/s41392-022-01136-2
169. Silverstein A, Dudaev A, Studneva M, et al. Chapter Eight - Evolution of biomarker research in autoimmunity conditions for health professionals and clinical practice†Deceased. In: Teplow DB, editor. Progress in Molecular Biology and Translational Science. Academic Press; 2022: 219–276.
170. Harris KM, Lu T, Lim N, et al. Challenges and Opportunities for Biomarkers of Clinical Response to AHSCT in Autoimmunity. Front Immunol. 2018;9. 10.3389/fimmu.2018.00100
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Emerging Predictive Biomarkers of Immunotherapy Sensitivity in Patients with Non-Small Cell Lung Cancer
Gariazzo E, Colamartini F, Ubaldi M, Santo V, Brunetti L, Tomarelli C, Ognissanti D, Nassar J, Costabile S, Romano L, De Vita E, Scorpiniti I, Macrì M, Porreca R, Currà MF, Cortellini A, Ricciuti B, Metro G
ImmunoTargets and Therapy 2026, 15:567238
Published Date: 2 February 2026
Artificial Intelligence–Enabled Multi-Omics for Predicting Immune Checkpoint Inhibitor Response and Resistance
Wang X, He J, Ding G, Tang Y, Wang Q
Journal of Multidisciplinary Healthcare 2026, 19:572089
Published Date: 26 February 2026