Back to Journals » Neuropsychiatric Disease and Treatment » Volume 22
Exosomes Regulate the NLRP3/Caspase-1/IL-1β Signaling Pathway in Parkinson’s Disease: Mechanisms of Neuroinflammation Modulation and α-Synuclein Propagation
Received 10 December 2025
Accepted for publication 19 February 2026
Published 28 February 2026 Volume 2026:22 587802
DOI https://doi.org/10.2147/NDT.S587802
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Roger Pinder
Tieru Zhang,1 Hao Du,2 Shun Wang3
1Graduate School, Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China; 2Acupuncture Department, Fangta TCM Hospital of Songjiang District, Shanghai, People’s Republic of China; 3The Second Affiliated Hospital, Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China
Correspondence: Shun Wang, The Second Affiliated Hospital, Heilongjiang University of Chinese Medicine, Harbin, People’s Republic of China, Email [email protected]
Objective: Parkinson’s disease (PD) is a progressive neurodegenerative disorder, with neuroinflammation as core pathological drivers. The NLRP3/Caspase-1/IL-1β signaling pathway acts as a pivotal mediator of PD-related neuroinflammation, while exosome serves as key regulatory mediators of this pathway. This review systematically synthesizes the molecular mechanisms underlying exosome-mediated modulation of the NLRP3/Caspase-1/IL-1β axis in PD.
Methods: We screened PubMed and Embase databases from January 2010 to January 2025 to search for published studies. The search keywords used are as follows: [“Parkinsonl” or “PD”], [“exosome”], [“NLRP3” or “inflammation”], [“acupuncture” or “electroacupuncture”]. Studies on human/animal models were included, and articles that did not meet the requirements were excluded.
Results: Exosomes exert dual regulatory effects on the NLRP3/Caspase-1/IL-1β axis, with functional divergence determined by their cellular origin. From a pro-inflammatory perspective, exosomes derived from microglia and neurons are enriched in NLRP3, ASC, α-syn oligomers, and pro-IL-1β. After endocytosis by target dopaminergic neurons or surrounding microglia, these exosomes trigger mitochondrial ROS overproduction and intracellular K⁺ efflux—two critical signals for NLRP3 inflammasome activation. This leads to the assembly of the NLRP3-ASC-Caspase-1 complex, subsequent cleavage of pro-IL-1β/pro-IL-18 into mature cytokines, and exacerbation of dopaminergic neuronal pyroptosis. Notably, α-syn oligomers carried by these exosomes also enhance fibril formation in recipient cells, further amplifying NLRP3 activation and α-syn propagation; for example, microglial exosomes from MPTP-induced PD mice show 2– 3-fold higher NLRP3 expression compared to wild-type controls.
Conclusion: The exosome-NLRP3/Caspase-1/IL-1β axis mediates PD pathology. Targeting this axis holds promise for PD, and future research ought to optimize its clinical translation.
Keywords: exosome, NLRP3/Caspase-1/IL-1β, Parkinson’s disease, neuroinflammation, α-synuclein
Introduction
Parkinson’s disease (PD) is the second most common neurodegenerative disease worldwide. It is characterized by the progressive loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc), abnormal aggregation of α-synuclein (α-syn) forming Lewy bodies, and chronic neuroinflammation in the brain.1,2 As a key driving factor for PD progression, neuroinflammation forms a vicious cycle with α-syn pathology: abnormally aggregated α-syn activates microglia, triggering the release of pro-inflammatory factors, which further exacerbates neuronal damage and α-syn aggregation.3,4 Among numerous inflammatory regulatory pathways, the NLRP3 inflammasome plays a central role—its activation cleaves caspase-1, promotes the release of pro-inflammatory cytokines IL-1β and IL-18, and induces pyroptosis, directly contributing to dopaminergic neuronal damage and neuroinflammation amplification.5,6
As extracellular vesicles with a diameter of 30–150 nm, exosomes are key mediators of intercellular communication. They can carry bioactive molecules such as proteins (eg, α-syn, NLRP3), nucleic acids (microRNAs [miRNAs], messenger RNAs [mRNAs]), and lipids, crossing the blood–brain barrier (BBB) to transmit signals in the brain microenvironment.7,8 Recent studies have confirmed that exosomes play dual roles in the PD inflammatory microenvironment. On the one hand, pro-inflammatory cell-derived exosomes can mediate the activation of the NLRP3 inflammasome and α-syn propagation, exacerbating neuroinflammation; on the other hand, anti-inflammatory cell-derived exosomes can remodel the brain’s anti-inflammatory microenvironment by inhibiting the NLRP3 pathway and clearing α-syn aggregation.9,10
Given the central role of exosomes in NLRP3 inflammation regulation and PD pathological progression, this review comprehensively elaborates on the regulatory network of the exosome-NLRP3 inflammatory axis in PD from perspectives including cell source specificity, inflammatory regulatory mechanisms, complication relevance, and therapeutic translational potential, aiming to provide novel insights into disease-modifying therapy for PD.
Overview of Exosomes and Parkinson’s Disease-Associated Neuroinflammation
Exosome Biology and Secretion in PD
Exosomes are released through the fusion of intracellular multivesicular bodies (MVBs) with the plasma membrane and are widely present in the blood, cerebrospinal fluid (CSF), and brain tissues.11 Their biogenesis is regulated by the endosomal sorting complex required for transport (ESCRT) pathway, Rab GTPase family, and lipid metabolism. Notably, the pathological microenvironment of PD (eg, oxidative stress, mitochondrial dysfunction) can significantly affect exosome secretion efficiency and cargo composition.12,13 For instance, in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced PD mouse models, exosome secretion from microglia and neurons in the substantia nigra region is increased by 2–3 folds compared with wild-type mice, with significantly elevated levels of α-syn oligomers, NLRP3, and ASC proteins in exosomes.9,14
The cargo of exosomes exhibits cell source specificity. Among these, microglia-derived exosomes (MDEs) are enriched in inflammation-related molecules (eg, NLRP3, IL-1β, inducible nitric oxide synthase [iNOS]). In contrast, mesenchymal stem cell (MSC)-derived exosomes are enriched in anti-inflammatory factors (eg, IL-10, transforming growth factor [TGF]-β) and neurotrophic factors (eg, brain-derived neurotrophic factor [BDNF], glial cell line-derived neurotrophic factor [GDNF]). Additionally, neuron-derived exosomes carry α-syn, synapse-associated proteins (eg, postsynaptic density protein 95 [PSD95]), and mitochondrial fragments.9,10,15 These characteristics render exosomes “carriers” of intercellular signal transmission in the PD inflammatory microenvironment, mediating inflammatory crosstalk between neurons, microglia, and astrocytes.7,16
PD Neuroinflammation and NLRP3-α-Syn Vicious Cycle
Neuroinflammation in PD is centered on sustained microglial activation, characterized by M1 polarization (high expression of Iba-1, CD68), increased release of pro-inflammatory cytokines (IL-1β, IL-6, tumor necrosis factor [TNF]-α), and reactive astrogliosis.3,17 As a “molecular switch” for inflammatory activation, the NLRP3 inflammasome requires two signals for its activation: the first (priming signal) is triggered by LPS, α-syn, etc, activating the NF-κB pathway and upregulating the expression of NLRP3 and pro-IL-1β; the second (activation signal) is mediated by mitochondrial ROS, K⁺ efflux, and lysosomal damage, inducing the assembly of the NLRP3-ASC-caspase-1 complex.5,18
In the substantia nigra region of the brain in PD patients and animal models, biomarkers of NLRP3 inflammasome activation (cleaved caspase-1, IL-1β, GSDMD-N) are all significantly upregulated and positively correlated with the degree of dopaminergic neuronal loss.5,19 More importantly, NLRP3 activation and α-syn pathology form a vicious cycle: α-syn aggregation primes the NLRP3 inflammasome via the Toll-like receptor 4 (TLR4)/NF-κB pathway, while activated NLRP3 inhibits autophagy (via the IL-1β/mammalian target of rapamycin [mTOR] signaling pathway), further promoting α-syn aggregation and propagation.3,4
Bidirectional Exosome Regulation in PD Inflammation
Exosomes exhibit bidirectional regulatory properties of “pro-inflammatory-anti-inflammatory” in PD-related inflammation, with the pro-inflammatory effect manifested by microglia- and neuron-derived exosomes serving as key mediators of inflammatory propagation (as shown in Figure 1). For instance, after BV2 microglia is stimulated by α-syn, the secreted exosomes carry NLRP3 and ASC. Following endocytosis by dopaminergic neurons, these exosomes activate the intracellular NLRP3 pathway, promoting IL-1β release and α-syn aggregation.9 Neuron-derived exosomes are capable of transporting α-syn oligomers, which activate the TLR4/NF-κB/NLRP3 axis in microglia and induce a vicious cycle: neuronal damage triggers exosome release, which in turn activates microglia, and this microglial activation subsequently leads to further neuronal damage.14
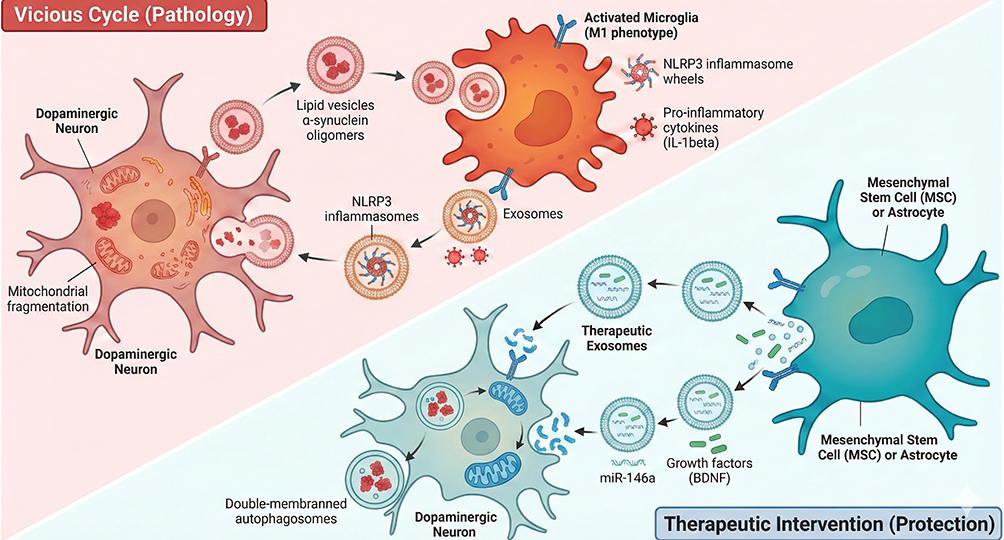 |
Figure 1 Dual Roles of Exosomes in Parkinson’s Disease Mediating Neuroinflammation vs Neuroprotection. Top Left (Vicious Cycle): The red-shaded region depicts the neurodegenerative feedback loop. Damaged Dopaminergic Neurons (left) exhibiting mitochondrial fragmentation release lipid vesicles containing α-synuclein oligomers (depicted as red clusters). These vesicles are internalized by Activated Microglia (M1 phenotype, Orange cell), triggering the assembly of NLRP3 inflammasomes (spoked wheel structures). The microglia subsequently release exosomes containing these inflammasomes and pro-inflammatory cytokines (eg, IL-1β, shown as red spiky spheres), which cycle back to further damage the neuron, inducing the formation of double-membraned autophagosomes. Bottom Right (Therapeutic Intervention): The blue-shaded region illustrates the protective mechanism. Mesenchymal Stem Cells (MSCs) or Astrocytes (teal cell) secrete Therapeutic Exosomes (blue vesicles). These exosomes are loaded with protective cargo, specifically microRNA-146a (miR-146a) (green strands) and Growth Factors such as BDNF (green rods). Upon uptake by the dopaminergic neuron, these factors restore mitochondrial health and interrupt the apoptotic pathway. Bold text: Highlights critical cell types (eg, Activated Microglia, Macrophage) and key molecular components (eg, NLRP3, NF-κB) central to the pathway; Red/Orange text and structures: Indicate pathological elements, pro-inflammatory cytokines, and damaged cellular states (eg, Damaged \α-syn, Mitochondrial Dysfunction); Arrows (→): Indicate the directional flow of exosomes and the sequence of signal transduction events (eg, Priming → Activation); Horizontal line (—): explanatory function. |
Meanwhile, the anti-inflammatory effect is reflected by mesenchymal stem cell (MSC)- and astrocyte-derived exosomes exerting neuroprotective effects via mechanisms including direct inhibition of NLRP3 inflammasome activation, regulation of microglial polarization (M1→M2), and facilitation of α-syn clearance.9,10 For instance, astrocyte-derived exosomes can upregulate the expression of IL-10 and BDNF, inhibit NLRP3 activation in microglia, and reduce dopaminergic neuronal loss.15
These bidirectional effects depend on the cellular origin of exosomes, the disease stage of PD, and the inflammatory state of the microenvironment. In the early stage of PD, exosomes may exert protective effects by clearing α-syn. Whereas in the progressive stage of the disease, pro-inflammatory exosomes predominate, exacerbating inflammation and pathological progression.7,16
Cell-Specific Exosome Mechanisms in PD Inflammation
Immune Cell Exosomes: NLRP3 Activation and α-Syn Propagation
Immune cells (microglia, macrophages) are the primary propagators of NLRP3 inflammation, and their derived exosomes serve as core carriers for pro-inflammatory signal transduction in PD (Figure 2). Their mechanisms focus on NLRP3 inflammasome activation and α-syn propagation. Studies have demonstrated that microglia-derived exosomes (MDEs) are the most widely studied pro-inflammatory exosomes in PD, and their regulation of inflammation depends on cargo composition and target cell types. NLRP3 and apoptosis-associated speck-like protein (ASC) carried by MDEs can directly enter neurons. By promoting mitochondrial ROS production and K⁺ efflux in neurons, MDEs activate the neuron-intrinsic NLRP3 inflammasome, leading to IL-1β release and neuronal pyroptosis.9 In MPTP-induced mouse models, inhibition of MDE release (via GW4869 treatment) reduced NLRP3 activation in substantia nigra neurons by 50% and increased dopaminergic neuron survival rate by 40%.9
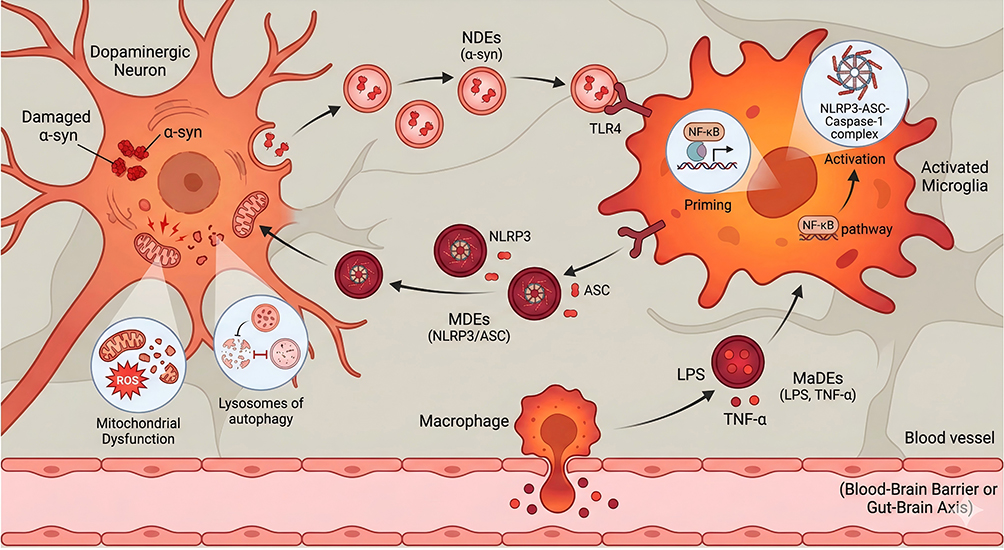 |
Figure 2 Immune Cell Exosomes mediated NLRP3 inflammatory cycle in Parkinson’s disease. Bold text: Highlights critical cell types (eg, Activated Microglia, Macrophage) and key molecular components (eg, NLRP3, NF-κB) central to the pathway; Red and Orange text and structures: Indicate pathological elements, pro-inflammatory cytokines, and damaged cellular states (eg, Damaged \α-syn, Mitochondrial Dysfunction); The red dots near macrophages are inflammatory factors, such as TNF - α; Arrows (→): Indicate the directional flow of exosomes and the sequence of signal transduction events (eg, Priming → Activation); Horizontal line (—): explanatory function. |
Furthermore, MDEs can encapsulate α-syn oligomers and facilitate α-syn spreading in the brain through intercellular transmission. After binding to MDEs, α-syn exhibits enhanced fibril-forming capacity, which further activates microglial NLRP3.7,16 Notably, released MDEs can be internalized by surrounding microglia and amplify NLRP3 inflammasome activation via the TLR4/NF-κB pathway, forming a “cascade reaction” of microglial population activation.20
Peripheral macrophage-derived exosomes (MaDEs) can cross the BBB to enter the brain, participating in the regulation of PD-related inflammation. For instance, exosomes secreted by peripheral blood macrophages from PD patients are enriched in TNF-α and IL-6, which can activate the NLRP3 inflammasome in brain microglia. In contrast, knockdown of macrophage exosome secretion alleviates neuroinflammation in MPTP-induced mice.21 Additionally, intestinal macrophage-derived exosomes can carry gut microbiota metabolites (eg, lipopolysaccharide [LPS]), activating the intracerebral NLRP3 pathway via the gut-brain axis and exacerbating PD-related inflammation.22
Neurocyte Exosomes: Heterogeneous Inflammation Regulation
Neurocyte-derived exosomes (from neurons and astrocytes) exhibit functional heterogeneity in PD-related inflammation. Neuron-derived exosomes are predominantly pro-inflammatory, while astrocyte-derived exosomes are predominantly anti-inflammatory.
Neuron-derived exosomes (NDEs) are key mediators of α-syn propagation and inflammation initiation, with their pro-inflammatory mechanisms involving two pathways. First, activating microglial NLRP3. α-syn oligomers carried by NDEs bind to toll-like receptor 2 (TLR2)/TLR4 on microglia, initiating the NF-κB pathway and upregulating the expression of NLRP3 and pro-IL-1β; meanwhile, mitochondrial fragments in NDEs can induce mitochondrial reactive oxygen species (ROS) production in microglia, triggering NLRP3 inflammasome activation.14,23 In A53T transgenic mice, inhibition of NDE secretion reduced IL-1β release from microglia by 35% and α-syn aggregation by 42%.14 Second, exacerbating neuron–neuron inflammatory propagation. NDEs released by damaged neurons can be internalized by adjacent healthy neurons, delivering α-syn and NLRP3-activated fragments, which leads to inflammatory spreading and neuronal population damage [52, 94].
Astrocyte-derived exosomes (ADEs) are key anti-inflammatory exosomes in the brain, with their regulatory mechanisms focusing on NLRP3 inhibition and neuroprotection. Studies have demonstrated that ADEs upregulate the expression of IL-10 and BDNF, inhibit microglial M1 polarization as well as the assembly and activation of the NLRP3 inflammasome, and simultaneously promote the phagocytic clearance of α-syn by microglia.15 In MPTP-induced mice, intravenous injection of ADEs reduced NLRP3 expression in the substantia nigra by 45% and increased the survival rate of tyrosine hydroxylase (TH)-positive neurons by 38%.15 Additionally, ADEs can upregulate the expression of tight junction proteins (eg, occludin, ZO-1), reduce the entry of peripheral inflammatory factors into the brain, and indirectly inhibit NLRP3 inflammasome activation.24
Stem Cell Exosomes: Anti-Inflammation and Neuroprotection
Stem cell-derived exosomes (from mesenchymal stem cells and neural stem cells) have emerged as promising candidates for PD treatment due to their potent anti-inflammatory and neuroprotective activities, exhibiting multi-targeted regulation of NLRP3 inflammation.
Mesenchymal stem cell-derived exosomes (MSC-Exo) are the most extensively studied anti-inflammatory exosomes. They inhibit NLRP3 inflammation through three pathways: direct inhibition of NLRP3 assembly, autophagy-mediated NLRP3 degradation promotion, and regulation of microglial polarization. MiR-146a and miR-29c carried by MSC-Exo can directly target the 3’ untranslated region (3’UTR) of NLRP3 mRNA to inhibit its expression; meanwhile, TGF-β in MSC-Exo activates the Smad pathway, downregulates nuclear factor-κB (NF-κB) activity, and blocks the priming signal of the NLRP3 inflammasome [97, S1].24,25 MSC-Exo can activate the autophagic pathway in neurons and microglia (upregulating the LC3-II/LC3-I ratio and reducing p62 expression), promote NLRP3 protein degradation, and simultaneously enhance α-syn clearance.10 Additionally, MSC-Exo can induce microglial polarization toward the M2 phenotype (high expression of arginase-1 [Arg-1] and CD206), reduce the release of IL-1β and TNF-α, and increase the secretion of IL-4 and IL-10.26 In MPTP-induced PD mice, intranasal administration of MSC-Exo significantly improved motor dysfunction, reduced the expression of NLRP3 and IL-1β in the substantia nigra by 60%, and decreased dopaminergic neuronal loss by 55%.10
Neural stem cell-derived exosomes (NSC-Exo) possess both anti-inflammatory and neuroregenerative activities, with their regulation of NLRP3 inflammation relying on neurotrophic factors and miRNA networks. BDNF secreted by NSC-Exo can activate the TrkB pathway in neurons, enhance mitochondrial function, and reduce ROS-mediated NLRP3 activation.26 Additionally, miR-124 carried by NSC-Exo can target TLR4, inhibit the NF-κB/NLRP3 pathway, and simultaneously promote microglial M2 polarization.8 We have summarized the inflammatory regulatory characteristics of extracellular vesicles from different sources, as shown in Table 1.
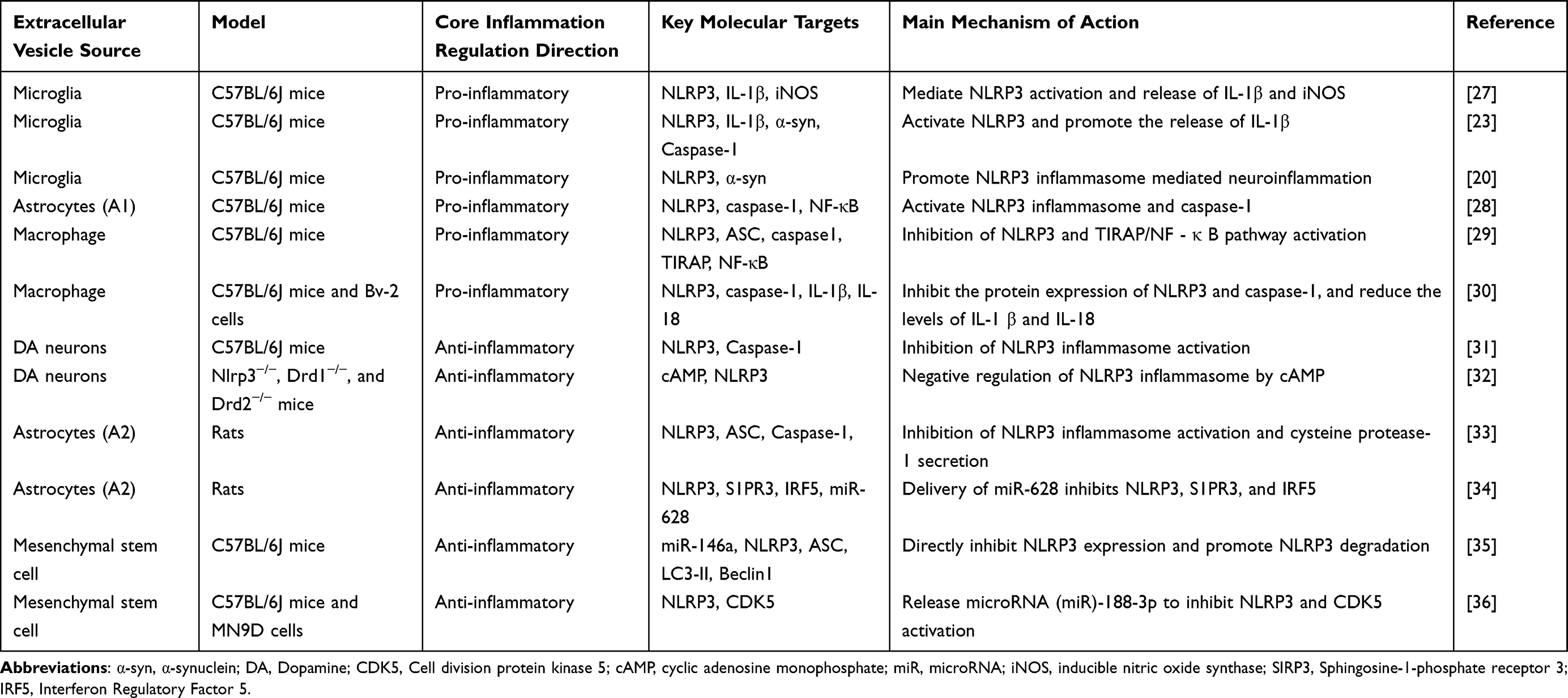 |
Table 1 Comparison of Inflammatory Regulatory Characteristics of Extracellular Vesicles from Different Sources |
PD Complications and the Exosome-NLRP3 Axis
Complications of PD are categorized into motor-related and non-motor-related types. The exosome-NLRP3 inflammatory axis exhibits specificity in its roles across different complications, which is closely associated with differences in affected brain regions and inflammatory networks.
Motor Complications: Nigrostriatal NLRP3 Activation
Freezing of gait (FOG) and postural instability (core affected brain region: nigrostriatal pathway) are the most common motor complications of PD, with their core inflammatory mechanism being exosome-mediated NLRP3 inflammation activation in the nigrostriatal pathway. The TLR4/NF-κB/NLRP3 axis and α-syn/TLRs/NF-κB/NLRP3 axis are key axes driving inflammatory progression in the disease.3,9 NLRP3, IL-1β, TNF-α, inducible nitric oxide synthase (iNOS), and α-syn oligomers carried by exosomes are core biomarkers of inflammatory progression.37 MDEs carry NLRP3 and α-syn, target and damage substantia nigra dopaminergic neurons and striatal synapses, leading to impaired motor coordination. In MPTP-induced mice, the level of NLRP3 in MDEs is positively correlated with the incidence of FOG, and inhibition of exosome release prolongs the rotarod test latency by 30%.9
Dysphagia (core affected brain region: brainstem swallowing center) has been rarely studied, but preliminary evidence indicates that the NLRP3/IL-1β axis is a key inflammatory axis driving the occurrence and progression of the condition.37 After brainstem microglia are stimulated by α-syn carried by exosomes, NLRP3 is activated to release IL-1β, damaging dysphagia-related neurons. Subsequently, exosomes can spread from the substantia nigra to the brainstem, exacerbating local inflammation.37
Non-Motor Complications: Hippocampal/Amygdalar/Gut-Brain Axis
Cognitive impairment (core-affected brain region: hippocampus) is an important non-motor complication of PD, with the exosome-NLRP3 inflammatory axis focusing on hippocampal synaptic damage. The cGAS-STING-NLRP3 axis and NLRP3/IL-1β/PSD95 axis are key inflammatory axes.38,39 NLRP3, IL-1β, PSD95 (synaptic plasticity marker), and BrdU⁺/NeuN⁺ (neurogenesis marker) are critical biomarkers of inflammatory progression.38 MDEs carry NLRP3 into hippocampal neurons, activate the inflammasome, reduce synaptic density (by decreasing PSD95 expression) and neurogenesis, leading to spatial memory impairment. In the Morris water maze test, mice with regulated NLRP3 in exosomes showed a 40% increase in time spent in the target quadrant.38
Depression and anxiety (core affected brain region: amygdala) are associated with NLRP3 activation in the amygdala and neurotrophic factor imbalance as their core inflammatory mechanisms. The amygdalar NLRP3/IL-1β/BDNF axis is a key axis driving inflammatory progression.37 NLRP3, IL-1β, and BDNF are core inflammatory biomarkers.37,40 Studies have shown that after NLRP3 activation in amygdalar MDEs, BDNF secretion is inhibited, leading to depressive-like behaviors. Following exosome regulation, the immobility time of mice in the forced swim test was reduced by 28%, and the exploration time in the central zone of the open field test was increased by 32%.37,41
Autonomic dysfunction (core affected brain region and organ: autonomic nervous center + intestine) is closely associated with inflammatory propagation via the gut-brain axis. The gut-brain axis NLRP3 pathway is a key pathway influencing inflammatory progression.22 NLRP3, LPS, and short-chain fatty acids (SCFA) are core biomarkers of inflammation induction.42 Gut microbiota dysbiosis leads to increased LPS carried by exosomes, which enter the central nervous system (CNS) via the gut-brain axis and activate NLRP3 inflammation in the autonomic nervous center.43 After gut microbiota modulation by electroacupuncture, exosomal LPS is reduced, and autonomic functions (eg, defecation frequency) are improved.44,45
The exosome-NLRP3 inflammatory axis in different complications is centered on NLRP3 inflammasome activation, with exosomes serving as carriers of inflammatory signals to mediate α-syn propagation and pro-inflammatory cytokine release.46 However, affected brain regions determine the specificity of inflammatory networks (the nigrostriatal pathway focuses on motor function, the hippocampus on cognition, and the amygdala on emotion), and biomarker expression profiles vary with brain region functions (eg, decreased PSD95 in cognitive impairment and reduced BDNF in depression and anxiety).
Exosome-Targeted Interventions for PD Inflammation
Based on the core role of exosomes in PD-related inflammation, current intervention strategies are mainly categorized into four types: regulation of exosome origin, modification of exosome cargo, enhancement of exosome delivery, and non-pharmacological combined intervention.47–51 All strategies are in the preclinical research stage, with some showing promising translational potential, as shown in Figure 3.
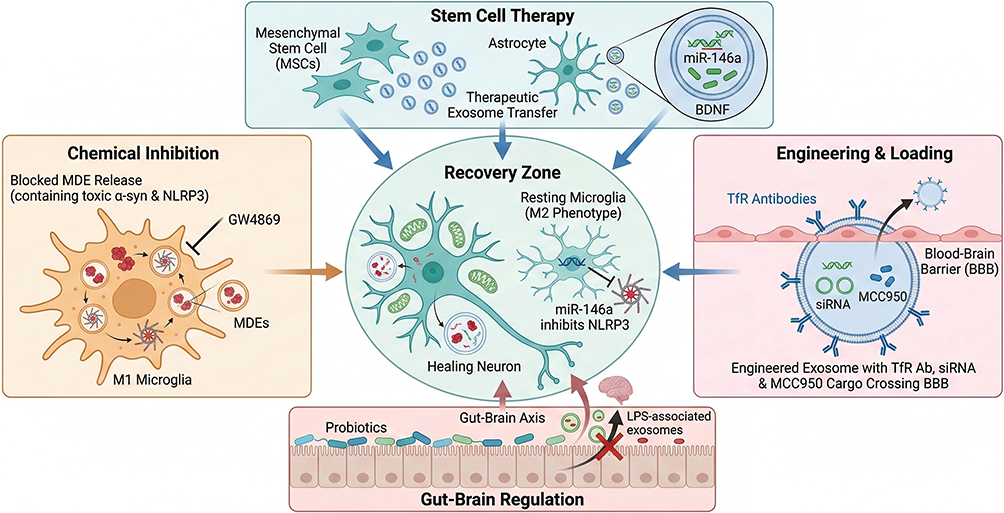 |
Figure 3 Therapeutic Interventions Targeting the Exosome-NLRP3 Aixs in Parkinson’s Disease. Top Panel (Stem Cell Therapy): Mesenchymal Stem Cells (MSCs) and Astrocytes are utilized to secrete therapeutic exosomes. These vesicles are naturally enriched with miR-146a (genetic regulator) and BDNF (Brain-Derived Neurotrophic Factor), which are transferred to recipient cells to promote survival and anti-inflammatory states. Left Panel (Chemical Inhibition): The pharmacological agent GW4869 is depicted inhibiting the exosomal secretory machinery of pro-inflammatory M1 Microglia. This blockade prevents the release of “MDEs” (Microglial-Derived Exosomes) carrying toxic α-synuclein and NLRP3 inflammasome components. Right Panel (Engineering & Loading): Engineered exosomes are modified with surface Transferrin Receptor (TfR) Antibodies (Y-shaped markers) to facilitate transport across the Blood-Brain Barrier (BBB). These vesicles serve as delivery vehicles for siRNA and MCC950 (a specific NLRP3 inhibitor) to directly target microglial inflammation. Bottom Panel (Gut-Brain Regulation): Probiotics are used to modulate the intestinal environment, strengthening the gut barrier and reducing the release of Lipopolysaccharide (LPS)-associated exosomes into the Gut-Brain Axis. Center Panel (Recovery Zone): The cumulative effect of these interventions is the promotion of a Healing Neuron phenotype (characterized by restored mitochondrial function and enhanced autophagy) and the repolarization of microglia to the protective Resting (M2) phenotype. Bold text: Indicates specific biological structures, molecules, or labels explicitly visible in the figure; Blue and Green text elements: Correspond to therapeutic agents (eg, “miR-146a”, “BDNF”) and neuroprotective states (eg, “Healing Neuron”) depicted in cool tones in the diagram; Red and Orange text elements: Correspond to pathological drivers (eg, “LPS”, “α-syn”) and inflammatory states (eg, “M1 Microglia”) depicted in warm tones in the diagram; Arrows (→): Indicate promotion, secretion, or transfer pathways; T-bars (T): Indicate inhibition or suppression of a pathway; Red Cross (×): Indicates the blockage or prohibition of a pathological process. |
Inhibit Pro-Inflammatory Exosome Secretion
This strategy is mainly achieved by small-molecule inhibitors blocking exosome secretion from pro-inflammatory cells (microglia, neurons), thereby reducing NLRP3 inflammatory propagation. GW4869 (a ceramide synthase inhibitor) and DMA (an exosome release inhibitor) are representative agents.9,52,53 Studies have demonstrated that, after treating MPTP-induced mice with GW4869, exosome secretion from brain microglia was reduced by 50%, the expression of NLRP3 and IL-1β was decreased by 45%, and the survival rate of dopaminergic neurons was increased by 40%.9 Small-molecule inhibitors offer simple operation and rapid inhibition of inflammatory propagation, but they lack cell specificity and may simultaneously suppress the secretion of anti-inflammatory exosomes.
Exosome Cargo Modification (Engineered/Drug-Loaded)
Currently, there are two main approaches for the modification of exosome cargo. The first is genetically modified exosomes, whose core mechanism involves modifying exosome donor cells via approaches such as small interfering RNA (siRNA) and lentiviral transfection, enabling the secreted exosomes to carry anti-inflammatory molecules (eg, NLRP3 siRNA, miR-146a).54,55 Studies have demonstrated that lentiviral transfection of BV2 microglia to silence NLRP3 resulted in exosomes that reduced α-syn aggregation in neurons by 35%.9,55 Additionally, after transfecting mesenchymal stem cells (MSCs) with miR-146a, the efficiency of exosomes in inhibiting NLRP3 activation was doubled.56 Genetically modified exosomes exhibit high targeting specificity, allowing precise regulation of inflammatory pathways, but the efficiency and safety of gene transfection require further optimization.
The second approach is drug-loaded exosomes, which involves loading NLRP3 inhibitors (eg, MCC950) and antioxidants (eg, curcumin) into exosomes to enhance drug brain penetrability and targeting specificity.57,58 When MCC950 is loaded into MSC-derived exosomes and administered intranasally, the intracerebral drug concentration is increased by 5-fold compared with free MCC950, and NLRP3 activation is reduced by 60%.59 Drug-loaded exosomes can synergistically exert the anti-inflammatory effects of exosomes and the inhibitory effects of drugs. However, the loading efficiency and drug release kinetics require further optimization.
Enhance Exosome Delivery (BBB Penetration)
Enhancement of exosome delivery involves the use of nanocarriers (eg, PEGylated liposomes, metal-organic frameworks [MOFs]) or ligand modification (eg, transferrin receptor [TfR] antibodies, CD11b ligands) to improve the BBB penetrability and microglial targeting specificity of exosomes.59–61
Studies have demonstrated that PEGylated liposome-encapsulated MSC-derived exosomes exhibit an 8-fold increase in intracerebral uptake compared with free exosomes and can reduce NLRP3 inflammation in the substantia nigra by 70%.59,62 Exosome delivery enhancement can address the key issue of low exosome brain delivery efficiency; however, the long-term safety of nanocarriers requires evaluation.
Non-Pharmacological Interventions (EA/Probiotics)
Electroacupuncture is a major clinical approach for regulating exosomes to treat inflammation. It modulates gut microbiota to increase SCFA, reduce LPS content in intestine-derived exosomes, and inhibit gut-brain axis NLRP3 inflammation.22,63 In an animal study, after electroacupuncture treatment of 6-hydroxydopamine (6-OHDA)-induced rats, fecal SCFA was increased by 2-fold, exosomal LPS was reduced by 45%, motor dysfunction was improved, and NLRP3 expression in the substantia nigra was decreased by 38%.22,64
Probiotic intervention can also regulate exosomes to inhibit inflammation. Studies have demonstrated that supplementation with probiotics such as Bifidobacteria and Lactobacilli modulates gut microbiota balance and reduces the secretion of pro-inflammatory exosomes.21,65 After probiotic intervention in MPTP-induced mice, intestinal barrier function was enhanced (occludin upregulated by 30%), NLRP3 activation in brain microglia was reduced by 35%, and α-syn aggregation was decreased by 40%.21,66 We compared the conversion potential of different intervention strategies, as shown in Table 2.
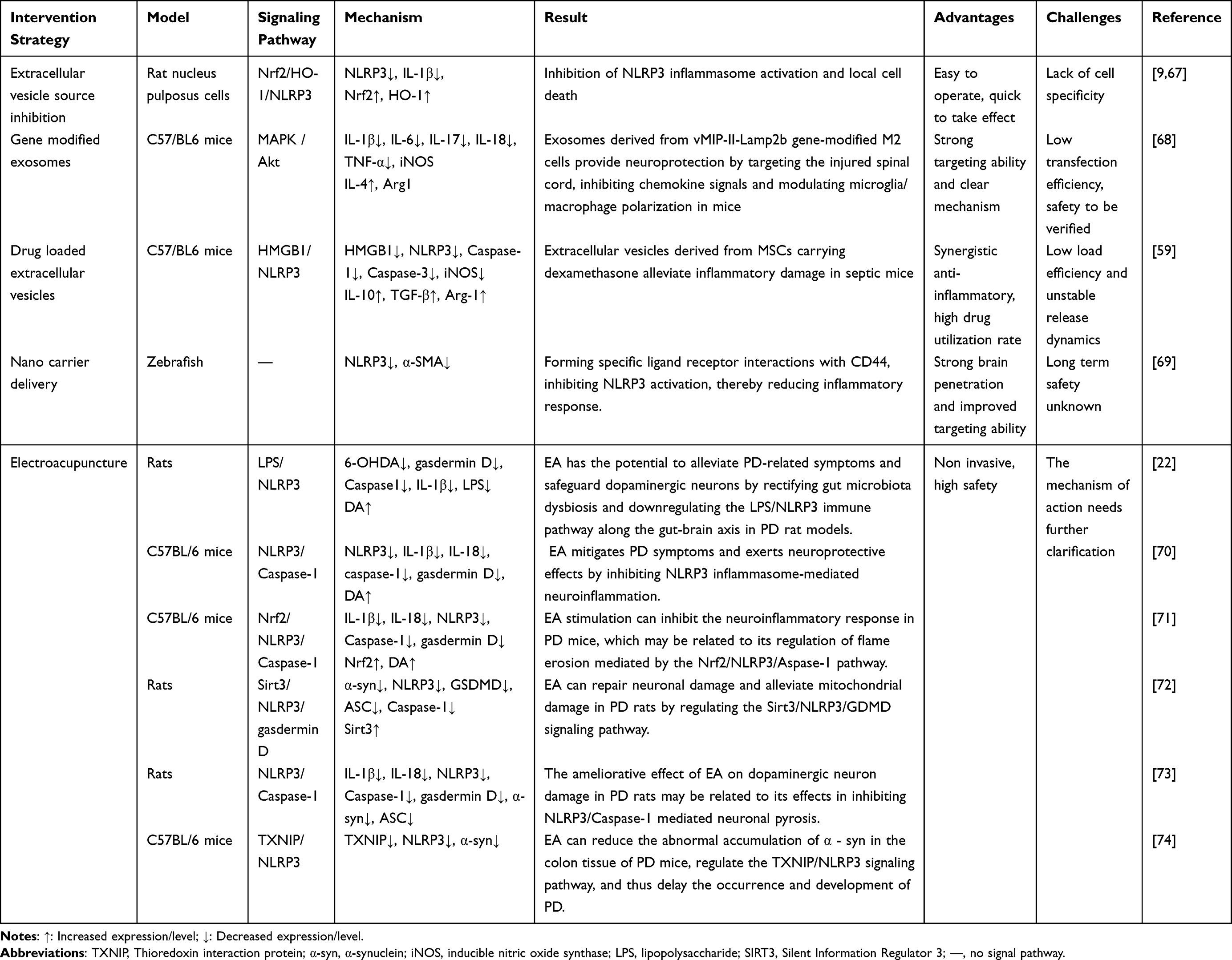 |
Table 2 Comparison of Conversion Potential of Different Intervention Strategies |
Preclinical Progress and Translational Challenges
PD heterogeneity demands a nuanced understanding of NLRP3’s functional role in disease pathogenesis sporadic PD which accounts for 90% of all clinical cases displays robust NLRP3 activation associated with gut microbiota dysbiosis and exosomal LPS transmission,21 while monogenic PD variants such as LRRK2 G2019S exhibit milder NLRP3 involvement owing to the predominance of kinase-mediated pathological processes.39 Human clinical evidence corroborates the clinical relevance of NLRP3 in PD the levels of NLRP3 and IL-1β in the cerebrospinal fluid and peripheral exosomes of PD patients are two to three times higher than those in healthy controls, and these elevations correlate with UPDRS scores and α-syn aggregation.50,75 Postmortem pathological studies have confirmed the colocalization of NLRP3 with α-syn in dopaminergic neurons and activated microglia within the substantia nigra.23,28 These lines of evidence establish a critical link between preclinical experimental findings and clinical patient populations and further highlight the subtype- and stage-dependent contributions of NLRP3 to PD pathogenesis.
Currently, all exosome-targeted interventions for PD-related inflammation are in the preclinical stage and have not yet entered clinical trials. The efficacy of exosome regulatory strategies has been validated in various PD animal models, such as MPTP-induced, A53T transgenic, and 6-hydroxydopamine (6-OHDA)-induced models, including improvements in motor function, protection of dopaminergic neurons, and inhibition of inflammation.10,59,76,77 In terms of technical breakthroughs, nanocarrier delivery systems (eg, PEGylated liposomes, metal-organic frameworks [MOFs]) have significantly enhanced exosome brain penetrability, enabling intracerebral drug concentrations to reach the therapeutic threshold.59,78 Additionally, engineered exosomes (eg, siRNA-modified, ligand-targeted) exhibit significantly superior targeting specificity and anti-inflammatory activity compared with natural exosomes.9,56
Numerous core challenges remain for the clinical translation of exosome-targeted therapeutic strategies. First, exosome heterogeneity and standardization. The cargo and functions of exosomes are significantly influenced by donor cell type, culture conditions, and disease stage, leading to poor reproducibility of experimental results among different research teams and a lack of unified exosome preparation and quality control standards.79,80 Studies have shown that the content of miR-146a in MSC-derived exosomes from different sources can vary by 3-fold, directly affecting their NLRP3 inhibitory efficiency.56 Second, insufficient brain penetrability and targeting specificity. The efficiency of natural exosomes crossing the BBB is only 0.1%-1%, making it difficult to achieve effective intracerebral concentrations.81,82 Although nanocarriers and ligand modification can improve delivery efficiency, issues such as carrier toxicity and immunogenicity persist.59,83 Additionally, there is a lack of clinical biomarkers for monitoring the activity of the exosome-NLRP3 inflammatory axis, making it difficult to evaluate the efficacy of intervention strategies.2 Although levels of NLRP3 and IL-1β in peripheral blood are associated with PD severity, whether they can reflect intracerebral inflammatory status remains to be verified.39 Most importantly, safety assessment is lacking. The long-term safety of exosome-targeted interventions is unclear: inhibition of exosome origin may disrupt normal intercellular communication; genetically modified exosomes may carry the risk of insertional mutations; and nanocarriers may accumulate in the brain, triggering long-term inflammation.8,39
Limitations and Future Directions
The existing evidence has the following limitations. First, unclear cell-specific mechanisms. Most studies focus on the overall effects of exosomes, lacking specific sorting and functional validation of exosomes derived from different cell types such as microglia and neurons.7,16 Second, insufficient research on disease stage dependency. The functional switching mechanism of exosomes (anti-inflammatory in the early stage of PD and pro-inflammatory in the progressive stage) has not been clarified, making it difficult to develop stage-specific intervention strategies.7 Third, lack of clinical data. All studies are based on cell and animal models, with a shortage of human sample validation (eg, the correlation between exosomal NLRP3 levels in CSF of PD patients and disease severity).39 Fourth, inadequate research on the crosstalk between exosomes and other inflammatory pathways. For instance, the interactions between the exosome-NLRP3 axis and pathways such as TLR4 and cGAS-STING remain unclear.18
Future research should focus on the following aspects. First, it is necessary to decipher the functional specificity of exosome subpopulations. Using techniques such as single-cell sequencing and flow cytometry sorting, we isolate exosome subpopulations from different cell sources with distinct cargo compositions, and clarify their specific roles in inflammation regulation.84,85 Second, we explore disease stage-dependent mechanisms. Construct progressive PD models (eg, different age stages of A53T transgenic mice) to analyze changes in exosome secretion and function with disease progression.7,75,86
Additionally, there is a need to develop clinically applicable biomarkers: validate the correlation between biomarkers of the “exosome-NLRP3-IL-1β” axis in CSF/peripheral blood and PD severity as well as complications, providing tools for clinical efficacy monitoring.87,88 Furthermore, we design combined intervention strategies, such as “nanocarrier-delivered engineered exosomes + NLRP3 inhibitors”, to synergistically exert targeted delivery and inflammation inhibitory effects.89–91 Most importantly, conduct preclinical safety assessments. Systematically evaluate the long-term toxicity, immunogenicity, and BBB accumulation risk of exosome-targeted intervention strategies.92,93
Conclusion
PD progression is tightly linked to the exosome-NLRP3 inflammatory axis, which mediates the vicious cycle between neuroinflammation and α-synuclein pathology. This review highlights exosomes as dual-function mediators. Pro-inflammatory exosomes from microglia/neurons propagate NLRP3 activation and α-syn spreading, while anti-inflammatory counterparts from MSCs/astrocytes inhibit NLRP3, clear α-syn aggregates, and protect dopaminergic neurons. Notably, this axis exhibits brain region specificity in PD complications—targeting the nigrostriatal pathway in motor deficits, hippocampus in cognitive impairment, and amygdala in neuropsychiatric symptoms—providing a foundation for precision therapy.
Preclinical advances support the translational feasibility of NLRP3 targeting with nanocarrier-modified exosomes such as PEGylated liposomes and RVG29 peptide-conjugated variants achieving an 8-fold higher brain penetration efficiency than free therapeutic agents,56,93 while cell-specifically engineered exosomes including miR-146a-loaded MSC-derived exosomes exert inhibitory effects on microglial NLRP3 activation without inducing systemic immunosuppression.34,35,54 Critically, NLRP3 acts as a causal driver of Parkinson’s disease with NLRP3 knockout mice exhibiting attenuated α-syn aggregation and dopaminergic neuron loss in MPTP/A53T mouse models and human genetic investigations establishing a direct link between the NLRP3 variant rs10754558 and elevated PD risk alongside accelerated disease progression.29 These experimental and clinical genetic data confirm that NLRP3 functions not merely as a secondary pathological response but as an upstream mediator of α-syn-related pathology thus validating its potential as a disease-modifying therapeutic target for PD.
Our work advances existing knowledge by three distinct novelties: First, we establish exosomes as non-redundant mediators bridging α-syn propagation and NLRP3 activation, whereas prior studies focused on direct protein–pathway interaction.3,5 Second, we uncover a disease-stage-dependent switch in exosome function—protective in early PD via α-syn clearance but pathogenic in progressive stages by amplifying NLRP3 inflammasome cascades,87 which explains the conflicting anti-/pro-inflammatory roles of exosomes reported in isolated studies. Third, we extend NLRP3 regulation to the gut-brain axis, demonstrating that intestinal exosomal LPS/α-syn activates central inflammation, a systemic mechanism rarely explored in PD.21,22 These findings clarify exosomes as upstream triggers and regulatory checkpoints of the α-syn-NLRP3 axis, distinguishing from prior work limited to brain-localized signaling.
Key challenges include exosome heterogeneity, insufficient BBB penetration, and lack of clinical biomarkers. Future research should focus on deciphering exosome subpopulation functions, developing stage-specific interventions, and validating peripheral exosomal NLRP3/IL-1β as clinical efficacy monitors. Overall, the exosome-NLRP3 axis unlocks a novel paradigm for PD disease-modifying therapy, with translational potential to transform clinical management by leveraging exosomes’ unique communication and targeting capabilities.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors have no acknowledgments and funding to report.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Yan YQ, Zheng R, Liu Y, et al. Parkin regulates microglial NLRP3 and represses neurodegeneration in Parkinson’s disease. Aging Cell. 2023;22:e13834. doi:10.1111/acel.13834
2. Liu TW, Chen CM, Chang KH. Biomarker of neuroinflammation in Parkinson’s disease. Int J Mol Sci. 2022;23. doi:10.3390/ijms23084148
3. Li Y, Xia Y, Yin S, et al. Targeting Microglial α-Synuclein/TLRs/NF-kappaB/NLRP3 Inflammasome Axis in Parkinson’s Disease. Front Immunol. 2021;12:719807. doi:10.3389/fimmu.2021.719807
4. Huang Q, Yang P, Liu Y, Ding J, Lu M, Hu G. The interplay between α-Synuclein and NLRP3 inflammasome in Parkinson’s disease. Biomed Pharmacother. 2023;168:115735. doi:10.1016/j.biopha.2023.115735
5. Gordon R, Albornoz EA, Christie DC, et al. Inflammasome inhibition prevents α-synuclein pathology and dopaminergic neurodegeneration in mice. Sci Transl Med. 2018;10. doi:10.1126/scitranslmed.aah4066
6. Wang S, Yuan YH, Chen NH, Wang HB. The mechanisms of NLRP3 inflammasome/pyroptosis activation and their role in Parkinson’s disease. Int Immunopharmacol. 2019;67:458–16. doi:10.1016/j.intimp.2018.12.019
7. Si XL, Fang YJ, Li LF, et al. From inflammasome to Parkinson’s disease: does the NLRP3 inflammasome facilitate exosome secretion and exosomal alpha-synuclein transmission in Parkinson’s disease? Exp Neurol. 2021;336:113525. doi:10.1016/j.expneurol.2020.113525
8. Saad HM, Atef E, Elsayed AE. New insights on the potential role of pyroptosis in Parkinson’s neuropathology and therapeutic targeting of NLRP3 inflammasome with recent advances in nanoparticle-based miRNA therapeutics. Mol Neurobiol. 2025;62:9365–9384. doi:10.1007/s12035-025-04818-4
9. Zengeler KE, Lukens JR. Taking the parkin brakes off of neuronal NLRP3 drives inflammasome activation and neurodegeneration in Parkinson’s disease. Neuron. 2022;110:2356–2358. doi:10.1016/j.neuron.2022.06.015
10. Xue J, Tao K, Wang W, Wang X. What can inflammation tell us about therapeutic strategies for Parkinson’s disease? Int J Mol Sci. 2024;25:1641. doi:10.3390/ijms25031641
11. Lee JH, Kim HJ, Kim JU, et al. A novel treatment strategy by natural products in NLRP3 inflammasome-mediated neuroinflammation in Alzheimer’s and Parkinson’s disease. Int J Mol Sci. 2021;22(3):1324.
12. Cheng J, Liao Y, Dong Y, et al. Microglial autophagy defect causes Parkinson disease-like symptoms by accelerating inflammasome activation in mice. Autophagy. 2020;16:2193–2205. doi:10.1080/15548627.2020.1719723
13. Chen J, Xu G, Shen R, et al. Communications among neurocytes in Parkinson’s disease regulated by differential metabolism and blood-brain barrier traversing of chiral gold cluster-MOF integrated nanoparticles. Adv Sci. 2025;12:e2500026. doi:10.1002/advs.202500026
14. Abitbol A, Mallard B, Tiralongo E, Tiralongo J. Mushroom natural products in neurodegenerative disease drug discovery. Cells. 2022;11:3938. doi:10.3390/cells11233938
15. Jiang L, Zhang X, Wang S, et al. Functional monomers equipped microgel system for managing Parkinson’s disease by intervening chemokine axis-mediated nerve cell communications. Adv Sci. 2025;12:e2410070. doi:10.1002/advs.202410070
16. Wang A, Zhong G, Ying M, et al. Inhibition of NLRP3 inflammasome ameliorates LPS-induced neuroinflammatory injury in mice via PINK1/Parkin pathway. Neuropharmacology. 2024;257:110063. doi:10.1016/j.neuropharm.2024.110063
17. de Araújo FM, Cuenca-Bermejo L, Fernández-Villalba E, Costa SL, Silva VDA, Herrero MT. Role of microgliosis and NLRP3 inflammasome in Parkinson’s disease pathogenesis and therapy. Cell Mol Neurobiol. 2022;42:1283–1300. doi:10.1007/s10571-020-01027-6
18. Han QQ, Le W. NLRP3 inflammasome-mediated neuroinflammation and related mitochondrial impairment in Parkinson’s disease. Neurosci Bull. 2023;39:832–844. doi:10.1007/s12264-023-01023-y
19. Lee E, Hwang I, Park S, et al. MPTP-driven NLRP3 inflammasome activation in microglia plays a central role in dopaminergic neurodegeneration. Cell Death Differ. 2019;26:213–228. doi:10.1038/s41418-018-0124-5
20. Kumari M, Bisht KS, Ahuja K, Motiani RK, Maiti TK. Glycation produces topologically different α-synuclein oligomeric strains and modulates microglia response via the NLRP3-inflammasome pathway. ACS Chem Neurosci. 2024. doi:10.1021/acschemneuro.4c00057
21. Yang H, Shao Y, Hu Y, et al. Fecal microbiota from patients with Parkinson’s disease intensifies inflammation and neurodegeneration in A53T mice. CNS Neurosci Ther. 2024;30:e70003. doi:10.1111/cns.70003
22. Quan J, Liu X, Liang S, et al. Electroacupuncture suppresses motor impairments via microbiota-metabolized LPS/NLRP3 signaling in 6-OHDA induced Parkinson’s disease rats. Int Immunopharmacol. 2025;162:115089. doi:10.1016/j.intimp.2025.115089
23. Pike AF, Varanita T, Herrebout MAC, et al. α-Synuclein evokes NLRP3 inflammasome-mediated IL-1β secretion from primary human microglia. Glia. 2021;69:1413–1428. doi:10.1002/glia.23970
24. Liu T, Wei J. FGF21 confers neuroprotection in Parkinson’s disease by activating the FGFR1-sirt1 pathway. Cell Mol Biol Lett. 2025;30:127. doi:10.1186/s11658-025-00807-6
25. Harrell CR, Djonov V, Antonijevic A, Volarevic V. NLRP3 inflammasome as a potentially new therapeutic target of mesenchymal stem cells and their exosomes in the treatment of inflammatory eye diseases. Cells. 2023;12:2327. doi:10.3390/cells12182327
26. Chiarini A, Gui L, Viviani C, Armato U, Dal Prà I. NLRP3 inflammasome’s activation in acute and chronic brain diseases-an update on pathogenetic mechanisms and therapeutic perspectives with respect to other inflammasomes. Biomedicines. 2023;11. doi:10.3390/biomedicines11040999
27. Li J, Wang H, Liu D, et al. CB2R activation ameliorates late adolescent chronic alcohol exposure-induced anxiety-like behaviors during withdrawal by preventing morphological changes and suppressing NLRP3 inflammasome activation in prefrontal cortex microglia in mice. Brain Behav Immun. 2023;110:60–79. doi:10.1016/j.bbi.2023.02.001
28. Li S, Fang Y, Zhang Y, et al. Microglial NLRP3 inflammasome activates neurotoxic astrocytes in depression-like mice. Cell Rep. 2022;41:111532. doi:10.1016/j.celrep.2022.111532
29. Yang Y, Huang G, Xu Q, et al. miR-146a-5p attenuates allergic airway inflammation by inhibiting the NLRP3 inflammasome activation in macrophages. Int Arch Allergy Immunol. 2022;183:919–930. doi:10.1159/000524718
30. Huang H, Gao Y, Nie K, Wang L. [Macrophage migration inhibitory factor meditates MPP+/MPTP-induced NLRP3 inflammasome activation in microglia cells]. Nan Fang Yi Ke Da Xue Xue Bao. 2021;41:972–979. Sesotho. doi:10.12122/j.issn.1673-4254.2021.07.02
31. Panicker N, Kam TI, Wang H, et al. Neuronal NLRP3 is a parkin substrate that drives neurodegeneration in Parkinson’s disease. Neuron. 2022;110:2422–2437.e2429. doi:10.1016/j.neuron.2022.05.009
32. Yan Y, Jiang W, Liu L, et al. Dopamine controls systemic inflammation through inhibition of NLRP3 inflammasome. Cell. 2015;160:62–73. doi:10.1016/j.cell.2014.11.047
33. Liu Y, Zhao W, Lv X, et al. Herkinorin ameliorates neuronal damage in a pentylenetetrazol-induced epilepsy rat model through altering microglial and astrocytic activation by inhibiting PARP1 and NF-κB. Int Immunopharmacol. 2025;155:114588.
34. Wang Y, Li H, Sun H, et al. A2 reactive astrocyte-derived exosomes alleviate cerebral ischemia-reperfusion injury by delivering miR-628. J Cell Mol Med. 2024;28:e70004. doi:10.1111/jcmm.70004
35. Hua T, Yang M, Song H, et al. Huc-MSCs-derived exosomes attenuate inflammatory pain by regulating microglia pyroptosis and autophagy via the miR-146a-5p/TRAF6 axis. J Nanobiotechnology. 2022;20:324. doi:10.1186/s12951-022-01522-6
36. Li Q, Wang Z, Xing H, Wang Y, Guo Y. Exosomes derived from miR-188-3p-modified adipose-derived mesenchymal stem cells protect Parkinson’s disease. Mol Ther Nucleic Acids. 2021;23:1334–1344. doi:10.1016/j.omtn.2021.01.022
37. Sun Q, Wang S, Chen J, et al. MicroRNA-190 alleviates neuronal damage and inhibits neuroinflammation via Nlrp3 in MPTP-induced Parkinson’s disease mouse model. J Cell Physiol. 2019;234:23379–23387. doi:10.1002/jcp.28907
38. Sandhu JK, Kulka M. Decoding mast cell-microglia communication in neurodegenerative diseases. Int J Mol Sci. 2021;22:1093. doi:10.3390/ijms22031093
39. Guan Y, Han F. Key mechanisms and potential targets of the NLRP3 inflammasome in neurodegenerative diseases. Front Integr Neurosci. 2020;14:37. doi:10.3389/fnint.2020.00037
40. Xia CY, Guo YX, Lian WW, et al. The NLRP3 inflammasome in depression: potential mechanisms and therapies. Pharmacol Res. 2023;187:106625. doi:10.1016/j.phrs.2022.106625
41. Wang Y, Liu J, Wang H, et al. Mesenchymal stem cell-derived exosomes ameliorate diabetic kidney disease through the nlrp3 signaling pathway. Stem Cells. 2023;41:368–383. doi:10.1093/stmcls/sxad010
42. An PP, Liang XY, Yao XY, et al. Bifidobacterium breve BBr60 enhances SCFA levels and restores NLRP3/NLRP6 balance in the gut to improve motor deficits in PD mice. Probiotics Antimicrob Proteins. 2025. doi:10.1007/s12602-025-10576-5
43. Li C, Qi X, Xu L, et al. Preventive effect of the total polyphenols from nymphaea candida on sepsis-induced acute lung injury in mice via gut microbiota and NLRP3, TLR-4/NF-κB pathway. Int J Mol Sci. 2024;25(8):4276.
44. Liu X, She C, Li X, et al. Electroacupuncture improves lipid metabolism via proteome and gut microbiota profiling in obese rats. Am J Transl Res. 2025;17:4008–4022. doi:10.62347/ZQZS9458
45. Xu MM, Guo Y, Chen Y, Zhang W, Wang L, Li Y. Electro-acupuncture promotes gut motility and alleviates functional constipation by regulating gut microbiota and increasing butyric acid generation in mice. J Integr Med. 2023;21:397–406. doi:10.1016/j.joim.2023.05.003
46. Sun J, Xu G. Mesenchymal stem cell-derived exosomal miR-150-3p affects intracerebral hemorrhage by regulating TRAF6/NF-κB axis, gut microbiota and metabolism. Stem Cell Rev Rep. 2023;19:1907–1921. doi:10.1007/s12015-023-10541-1
47. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
48. Heris RM, Shirvaliloo M, Abbaspour-Aghdam S, et al. The potential use of mesenchymal stem cells and their exosomes in Parkinson’s disease treatment. Stem Cell Res Ther. 2022;13:371. doi:10.1186/s13287-022-03050-4
49. Wang X, Chen Y, Zhao Z, et al. Engineered exosomes with ischemic myocardium-targeting peptide for targeted therapy in myocardial infarction. J Am Heart Assoc. 2018;7:e008737. doi:10.1161/JAHA.118.008737
50. Jiang C, Hopfner F, Katsikoudi A, et al. Serum neuronal exosomes predict and differentiate Parkinson’s disease from atypical parkinsonism. J Neurol Neurosurg Psychiatry. 2020;91:720–729. doi:10.1136/jnnp-2019-322588
51. Zhu Y, Xi Q, Liu Y, Zhou Y, Liao J, Wu Q. Recent advances in exosome-based nanodelivery systems for Parkinson’s disease. Biomaterials. 2026;325:123548. doi:10.1016/j.biomaterials.2025.123548
52. Takahashi E, Inanami O, Asanuma T, Kuwabara M. Effects of ceramide inhibition on radiation-induced apoptosis in human leukemia MOLT-4 cells. J Radiat Res. 2006;47:19–25. doi:10.1269/jrr.47.19
53. Lyu L, Wang H, Li B, et al. A critical role of cardiac fibroblast-derived exosomes in activating renin angiotensin system in cardiomyocytes. J Mol Cell Cardiol. 2015;89:268–279. doi:10.1016/j.yjmcc.2015.10.022
54. Meng WT, Zhu J, Wang YC, et al. Targeting delivery of miR-146a via IMTP modified milk exosomes exerted cardioprotective effects by inhibiting NF-κB signaling pathway after myocardial ischemia-reperfusion injury. J Nanobiotechnology. 2024;22:382. doi:10.1186/s12951-024-02631-0
55. Lai X, Wang Y, Wang X, Liu B, Rong L. miR-146a-5p-modified hUCMSC-derived exosomes facilitate spinal cord function recovery by targeting neurotoxic astrocytes. Stem Cell Res Ther. 2022;13:487. doi:10.1186/s13287-022-03116-3
56. Li Q, Ding X, Chang Z, et al. Metal-organic framework based nanozyme system for NLRP3 inflammasome-mediated neuroinflammatory regulation in Parkinson’s disease. Adv Healthc Mater. 2024;13:e2303454. doi:10.1002/adhm.202303454
57. Li H, Guan Y, Liang B, et al. Therapeutic potential of MCC950, a specific inhibitor of NLRP3 inflammasome. Eur J Pharmacol. 2022;928:175091. doi:10.1016/j.ejphar.2022.175091
58. Naeem A, Prakash R, Kumari N, et al. MCC950 reduces autophagy and improves cognitive function by inhibiting NLRP3-dependent neuroinflammation in a rat model of Alzheimer’s disease. Brain Behav Immun. 2024;116:70–84. doi:10.1016/j.bbi.2023.11.031
59. Nizami S, Hall-Roberts H, Warrier S, Cowley SA, Di daniel E. Microglial inflammation and phagocytosis in Alzheimer’s disease: potential therapeutic targets. Br J Pharmacol. 2019;176:3515–3532. doi:10.1111/bph.14618
60. Mondal J, Pillarisetti S, Junnuthula V, et al. Hybrid exosomes, exosome-like nanovesicles and engineered exosomes for therapeutic applications. J Control Release. 2023;353:1127–1149. doi:10.1016/j.jconrel.2022.12.027
61. Engin AB. Message transmission between adipocyte and macrophage in obesity. Adv Exp Med Biol. 2024;1460:273–295.
62. Lotfy A, AboQuella NM, Wang H. Mesenchymal stromal/stem cell (MSC)-derived exosomes in clinical trials. Stem Cell Res Ther. 2023;14:66. doi:10.1186/s13287-023-03287-7
63. Guo L, Hu H, Jiang N, et al. Electroacupuncture blocked motor dysfunction and gut barrier damage by modulating intestinal NLRP3 inflammasome in MPTP-induced Parkinson’s disease mice. Heliyon. 2024;10:e30819. doi:10.1016/j.heliyon.2024.e30819
64. Li M, Wang K, Su WT, Jia J, Wang XM. Effects of electroacupuncture on metabolic changes in motor cortex and striatum of 6-hydroxydopamine-induced parkinsonian rats. Chin J Integr Med. 2020;26:701–708. doi:10.1007/s11655-017-2975-x
65. Niechcial A, Schwarzfischer M, Wawrzyniak P, et al. Probiotic administration modulates gut microbiota and suppresses tumor growth in murine models of colorectal cancer. Int J Mol Sci. 2025;26:4404. doi:10.3390/ijms26094404
66. Ou Z, Zhou Y, Wang L, et al. NLRP3 inflammasome inhibition prevents α-synuclein pathology by relieving autophagy dysfunction in chronic MPTP-treated NLRP3 knockout mice. Mol Neurobiol. 2021;58:1303–1311. doi:10.1007/s12035-020-02198-5
67. Xi H, Weng Y, Zheng Y, Wu L, Han D. Diacetoxy-6-gingerdiol protects the extracellular matrix of nucleus pulposus cells and ameliorates intervertebral disc degeneration by inhibiting the IL-1β-mediated NLRP3 pathway. Heliyon. 2024;10:e37877. doi:10.1016/j.heliyon.2024.e37877
68. Fu GQ, Wang YY, Xu YM, et al. Exosomes derived from vMIP-II-Lamp2b gene-modified M2 cells provide neuroprotection by targeting the injured spinal cord, inhibiting chemokine signals and modulating microglia/macrophage polarization in mice. Exp Neurol. 2024;377:114784. doi:10.1016/j.expneurol.2024.114784
69. Gong L, Zhou H, Zhang S, et al. CD44-targeting drug delivery system of exosomes loading forsythiaside A combats liver fibrosis via regulating NLRP3-mediated pyroptosis. Adv Healthc Mater. 2023;12:e2202228. doi:10.1002/adhm.202202228
70. Kang M, Cai J, Yi W, et al. Electroacupuncture alleviates Parkinson’s disease by inhibiting the NLRP3 inflammasome pathway. Am J Transl Res. 2025;17:3619–3629. doi:10.62347/PGKC2376
71. Zhang XL, Hu MN, Rong Z, Li YN, Wang Y, Ma J. Effect of electroacupuncture on Nrf2/NLRP3/Caspase-1 pathway mediated-pyroptosis in mice with Parkinson’s disease. Zhen Ci Yan Jiu. 2024;49:15–22. doi:10.13702/j.1000-0607.20230407
72. Wang Y, Wang YC, Ma J. [Effects of electroacupuncture on Sirt3/NLRP3/GSDMD signaling pathway in the substantia nigra of midbrain of rats with Parkinson’s disease]. Zhen Ci Yan Jiu. 2024;49:384–390. Hausa. doi:10.13702/j.1000-0607.20230024
73. Liu YY, Guo YB, Zhai HY, et al. [Effect of electroacupuncture regulating NLRP3/Caspase-1 pathway on pyroptosis of dopaminergic neurons in rats with Parkinson’s disease]. Zhen Ci Yan Jiu. 2022;47:983–992. Hausa. doi:10.13702/j.1000-0607.20211016
74. Wang Y, Wang YC, Ma J, et al. Effects of electroacupuncture on the intestinal thioredoxin interaction protein/Nod-like receptor 3 signaling pathway in mice with Parkinson’s disease. Zhen Ci Yan Jiu. 2023;48:1041–1047. doi:10.13702/j.1000-0607.20220554
75. Huang Y, Liu Z, Li N, et al. Parkinson’s disease derived exosomes aggravate neuropathology in SNCA*A53T mice. Ann Neurol. 2022;92:230–245. doi:10.1002/ana.26421
76. Huang S, Liu H, Lin Y, et al. Berberine protects against NLRP3 Inflammasome via ameliorating autophagic impairment in MPTP-induced Parkinson’s disease model. Front Pharmacol. 2020;11:618787. doi:10.3389/fphar.2020.618787
77. Lee S, Oh ST, Jeong HJ, et al. MPTP-induced vulnerability of dopamine neurons in A53T α-synuclein overexpressed mice with the potential involvement of DJ-1 downregulation. Korean J Physiol Pharmacol. 2017;21:625–632. doi:10.4196/kjpp.2017.21.6.625
78. Barzin M, Bagheri AM, Ohadi M, Abhaji AM, Salarpour S, Dehghannoudeh G. Application of plant-derived exosome-like nanoparticles in drug delivery. Pharm Dev Technol. 2023;28:383–402. doi:10.1080/10837450.2023.2202242
79. Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomed. 2020;15:6917–6934. doi:10.2147/IJN.S264498
80. Jeppesen DK, Fenix AM, Franklin JL, et al. Reassessment of Exosome Composition. Cell. 2019;177:428–445.e418. doi:10.1016/j.cell.2019.02.029
81. Li Y, Liu B, Zhao T, et al. Comparative study of extracellular vesicles derived from mesenchymal stem cells and brain endothelial cells attenuating blood-brain barrier permeability via regulating Caveolin-1-dependent ZO-1 and Claudin-5 endocytosis in acute ischemic stroke. J Nanobiotechnology. 2023;21:70. doi:10.1186/s12951-023-01828-z
82. Morad G, Carman CV, Hagedorn EJ, et al. Tumor-derived extracellular vesicles breach the intact blood-brain barrier via transcytosis. ACS Nano. 2019;13:13853–13865. doi:10.1021/acsnano.9b04397
83. Ying N, Lin X, Xie M, Zeng D. Effect of surface ligand modification on the properties of anti-tumor nanocarrier. Colloids Surf B Biointerfaces. 2022;220:112944. doi:10.1016/j.colsurfb.2022.112944
84. Feng L, Lo H, Zheng J, Weng W, Sun Y, Pan X. Cycloastragenol reduces microglial NLRP3 inflammasome activation in Parkinson’s disease models by promoting autophagy and reducing Scrib-driven ROS. Phytomedicine. 2024;135:156210. doi:10.1016/j.phymed.2024.156210
85. Wang SY, Li MM, Wu JT, et al. Lignans of Schisandra chinensis (Turcz.) Baill inhibits Parkinson’s disease progression through mediated neuroinflammation-TRPV1 expression in microglia. Phytomedicine. 2024;135:156146. doi:10.1016/j.phymed.2024.156146
86. Papadopoulos VE, Nikolopoulou G, Antoniadou I, et al. Modulation of β-glucocerebrosidase increases α-synuclein secretion and exosome release in mouse models of Parkinson’s disease. Hum Mol Genet. 2018;27:1696–1710. doi:10.1093/hmg/ddy075
87. Ouerdane Y, Hassaballah MY, Nagah A, et al. Exosomes in Parkinson: revisiting their pathologic role and potential applications. Pharmaceuticals. 2022;15:76. doi:10.3390/ph15010076
88. Nazari S, Pourmand SM, Motevaseli E, Hassanzadeh G. Mesenchymal stem cells (MSCs) and MSC-derived exosomes in animal models of central nervous system diseases: targeting the NLRP3 inflammasome. IUBMB Life. 2023;75:794–810. doi:10.1002/iub.2759
89. Zhao JW, Dyson SC, Kriegel C, et al. Modelling of a targeted nanotherapeutic ‘stroma’ to deliver the cytokine LIF, or XAV939, a potent inhibitor of Wnt-β-catenin signalling, for use in human fetal dopaminergic grafts in Parkinson’s disease. Dis Model Mech. 2014;7:1193–1203. doi:10.1242/dmm.015859
90. van Dommelen SM, Vader P, Lakhal S, et al. Microvesicles and exosomes: opportunities for cell-derived membrane vesicles in drug delivery. J Control Release. 2012;161:635–644. doi:10.1016/j.jconrel.2011.11.021
91. Jiang Y, Wang F, Wang K, et al. Engineered exosomes: a promising drug delivery strategy for brain diseases. Curr Med Chem. 2022;29:3111–3124. doi:10.2174/0929867328666210902142015
92. Segura-Aguilar J, Mannervik B. A preclinical model for Parkinson’s disease based on transcriptional gene activation via KEAP1/NRF2 to develop new antioxidant therapies. Antioxidants. 2023;12:673. doi:10.3390/antiox12030673
93. Jiang P, Xiao Y, Hu X, et al. RVG29 peptide-modified exosomes loaded with Mir-133b mediate the RhoA-ROCK pathway to improve motor and neurological symptoms in Parkinson’s disease. ACS Biomater Sci Eng. 2024;10:3069–3085. doi:10.1021/acsbiomaterials.3c01622
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Co-Application of C16 and Ang-1 Improves the Effects of Levodopa in Parkinson Disease Treatment
Fu XX, Wang J, Cai HY, Jiang H, Jiang JZ, Chen HH, Han S
Journal of Inflammation Research 2022, 15:3797-3814
Published Date: 7 July 2022
Neural Stem Cell-Derived Exosomes Improve Neurological Function in Rats with Cerebral Ischemia-Reperfusion Injury by Regulating Microglia-Mediated Inflammatory Response

Zhao X, Zhu J, Chen S, Liu R, Long T
Journal of Inflammation Research 2023, 16:3079-3092
Published Date: 24 July 2023
Exosome-Mediated Mitochondrial Regulation: A Promising Therapeutic Tool for Alzheimer’s Disease and Parkinson’s Disease

Jung YH, Jo HY, Kim DH, Oh YJ, Kim M, Na S, Song HY, Lee HJ
International Journal of Nanomedicine 2025, 20:4903-4917
Published Date: 17 April 2025
Lifting the Veil on Neuroinflammation in Parkinson’s Disease: A Bibliometric Analysis
Dong S, Li X, Wang S, Gong Y, Wu L, Hu J, Chen X
Journal of Inflammation Research 2026, 19:607710
Published Date: 29 May 2026