Back to Journals » International Journal of Nanomedicine » Volume 21
EV-Mediated Oncogenic Regulation in Lung Cancer and Clinical Translation: From Liquid Biopsy to Targeted Delivery Systems
Authors Fang J, Li J, Yan L, Yang Y, Chen Y, Zhang X, Zhang C
Received 16 January 2026
Accepted for publication 3 July 2026
Published 16 July 2026 Volume 2026:21 596841
DOI https://doi.org/10.2147/IJN.S596841
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Kamakhya Prakash Misra
Jianjun Fang,1– 3,* Jing Li,1– 3,* Lun Yan,1– 3 Yang Yang,1– 3 Ying Chen,1– 3 Xi Zhang,1– 3 Cheng Zhang1– 3
1Medical Center of Hematology, Xinqiao Hospital, Army Medical University, Chongqing, 400037, People’s Republic of China; 2Chongqing Key Laboratory of Hematology and Microenvironment, Army Medical University, Chongqing, 400037, People’s Republic of China; 3State Key Laboratory of Trauma and Chemical Poisoning, Army Medical University, Chongqing, 400037, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xi Zhang, Email [email protected] Cheng Zhang, Email [email protected]
Abstract: Lung cancer (LC) remains the leading cause of cancer-associated mortality globally. Delayed diagnosis, therapeutic resistance and high postoperative recurrence are major hurdles that compromise its clinical outcomes. Extracellular vesicles (EVs) are cell-secreted nanoscale lipid vesicles that mediate intercellular crosstalk via the transfer of bioactive cargo, such as nucleic acids and proteins. In the lung cancer microenvironment, EVs remodel core signaling axes including PI3K/AKT and NF-κB, consequently driving hallmark malignant phenotypes: angiogenesis, distant metastasis, immune evasion and multidrug resistance. Boasting robust physicochemical stability, complete molecular cargo signatures and minimally invasive sampling feasibility, EVs represent ideal tumor biomarkers for liquid biopsy. Additionally, their intrinsic low immunogenicity renders EVs versatile nanovehicles capable of amplifying anti-tumor effects elicited by conventional radiotherapy, chemotherapy and immunotherapy. However, substantial inherent heterogeneity, a lack of unified standards for EVs isolation and characterization, obstacles to scalable production and inadequate tumor-targeting efficiency collectively impede their clinical translation. This review systematically dissects the molecular mechanisms through which EVs orchestrate lung cancer malignant progression, comprehensively outlines state-of-the-art progress of EVs applications in liquid biopsy-based early screening, prognostic stratification, targeted drug delivery and cellular immunotherapy, thoroughly discusses prevailing translational bottlenecks, and envisions future directions of EVs in precision oncology for lung cancer. The work offers theoretical guidance for developing next-generation EV-centric diagnostic and therapeutic platforms.
Keywords: extracellular vesicles, lung cancer, cancer biomarkers, targeted drug delivery, liquid biopsy
Introduction
Despite advances in oncology, LC remains a dominant contributor to global cancer mortality and continues to impose a substantial healthcare burden worldwide.1 Recent epidemiological estimates from the International Agency for Research on Cancer (IARC) indicate that LC accounts for approximately 12.4% of all newly diagnosed malignancies, representing a substantial and persistent global health burden.2,3 From a histopathological perspective, LC is broadly categorized into small cell LC (SCLC) and non-small cell LC (NSCLC), with NSCLC comprising nearly 85%–90% of cases. The major NSCLC subtypes include lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC).4 At the molecular level, LC development is driven by a heterogeneous landscape of genetic and signaling alterations. Clinically actionable driver events, including EGFR mutations, ROS1 rearrangements, and KRAS mutations, play central roles in NSCLC pathogenesis. Dysregulation of key oncogenic pathways such as PI3K/AKT, together with loss of tumor suppressor function in genes including TP53 and RB1, contributes to tumor initiation and progression.5 Environmental and lifestyle-related exposures further modulate disease risk, with tobacco smoking, air pollution, and occupational exposure to asbestos and particulate matter representing well-established risk factors.6 Despite advances in molecular diagnostics and therapeutic strategies, the clinical management of LC continues to face substantial limitations. Early-stage disease is often clinically silent, and current screening modalities, including imaging and serum-based biomarkers, lack sufficient sensitivity for reliable early detection. As a result, a significant proportion of patients are diagnosed at advanced stages. Therapeutic resistance to radiotherapy, chemotherapy, and targeted agents remains a major challenge, frequently accompanied by disease recurrence, metastatic progression, and immune evasion, collectively contributing to poor long-term survival outcomes.7 These limitations underscore the urgent need for more effective, minimally invasive diagnostic strategies and improved therapeutic approaches with higher precision.
In this context, EVs are nanoscale, lipid-bilayer-enclosed vesicles actively secreted by virtually all cell types and are widely detectable in diverse biological fluids. They carry a complex cargo of bioactive molecules, including nucleic acids, proteins, and lipids, and participate in the dynamic regulation of the tumor microenvironment. In LC, EVs have attracted increasing attention due to their inherent stability, molecular specificity, and accessibility through minimally invasive sampling, making them promising candidates for applications in early detection, disease monitoring, and therapeutic response assessment.8,9 Furthermore, EVs are being actively investigated as natural nanocarriers for drug delivery and as engineered platforms for targeted therapeutic intervention in LC.10 This review provides a comprehensive overview of EV-mediated mechanisms involved in LC progression, evaluates current liquid biopsy approaches, and summarizes recent advances in EV-based diagnostic and therapeutic strategies. Finally, it discusses key challenges limiting clinical translation and highlights future perspectives for the development of EV-based precision oncology approaches.
Overview of EVs
In accordance with the MISEV 2023 guidelines, EVs are defined and characterized based on their biogenesis, size distribution, and molecular composition, although no single definitive marker uniquely distinguishes EVs subtypes.11 EVs are heterogeneous, lipid bilayer-enclosed vesicles that are conventionally classified into three major categories: exosomes, microvesicles (MVs), and apoptotic bodies, reflecting distinct biogenetic pathways and biophysical properties (Figure 1).12
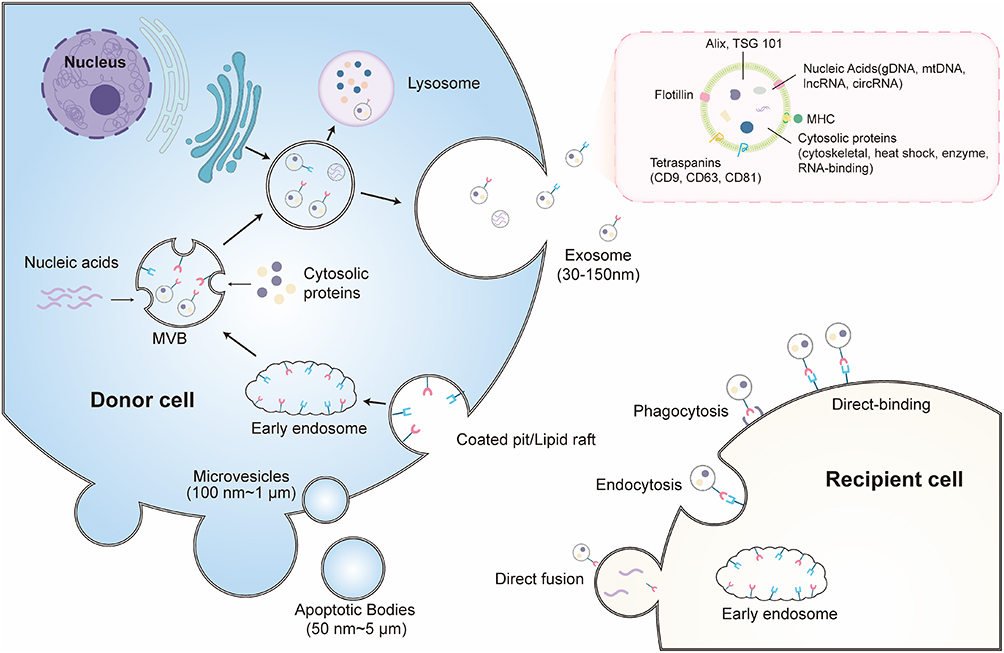 |
Figure 1 Biogenesis, secretion, and cellular uptake pathways of EVs: EVs are broadly classified into exosomes, microvesicles (MVs), and apoptotic bodies based on their biogenesis and size distribution. Exosomes originate from the endosomal system, where early endosomes undergo inward budding to generate intraluminal vesicles (ILVs), forming multivesicular bodies (MVBs). MVBs subsequently follow two distinct fates: fusion with lysosomes leading to cargo degradation, or fusion with the plasma membrane, resulting in the release of mature exosomes into the extracellular space. Microvesicles are generated through direct outward budding and fission of the plasma membrane, whereas apoptotic bodies are formed as membrane-enclosed vesicles released during programmed cell death. Following their release, EVs are internalized by recipient cells via multiple mechanisms, including endocytosis, phagocytosis, and direct membrane fusion, enabling the transfer of bioactive cargo that modulates recipient cell function. |
Exosomes
Exosomes are nanoscale EVs actively released by virtually all cell types into the extracellular microenvironment, with a typical diameter of approximately 30–150 nm.13 Their biogenesis originates from the endosomal system, beginning with plasma membrane invagination and the formation of early endosomes that internalize extracellular and membrane-associated cargo. During endosomal maturation, selected cargo is either directed toward recycling pathways or retained within intraluminal vesicles (ILVs) formed by inward budding of the endosomal membrane, leading to the generation of multivesicular bodies (MVBs).14 ILV formation is predominantly regulated by the endosomal sorting complex required for transport (ESCRT), comprising ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III complexes. ESCRT-0 is responsible for cargo recognition and clustering; ESCRT-I and ESCRT-II facilitate membrane deformation and budding, while ESCRT-III mediates membrane scission and vesicle release.15,16 In addition to ESCRT-dependent mechanisms, ESCRT-independent pathways involving lipids, tetraspanins, and heat shock proteins (HSPs) also contribute to ILV biogenesis.17 MVBs follow two main fates: fusion with lysosomes for degradation or fusion with the plasma membrane, resulting in the extracellular release of ILVs as exosomes. Exosomes are enriched in proteins such as CD63, LAMP1, and LAMP2, and commonly express conserved markers including Alix, TSG101, HSC70, and HSP90β, although no single universal marker definitively defines their origin.18,19 They appear as round to oval vesicles and often exhibit a cup-shaped structure under an electron microscope.20
MVs
MVs are a heterogeneous population of EVs typically ranging from 150 to 1000 nm in diameter.16 Unlike exosomes, MVs are formed through direct outward budding of the plasma membrane, a process driven by alterations in membrane lipid composition, intracellular calcium (Ca2⁺) flux, and cytoskeletal reorganization. A key early event in MV biogenesis is the translocation of phosphatidylserine (PS) from the inner to the outer leaflet of the plasma membrane, which promotes membrane curvature and budding.14 This process is tightly regulated by cytoskeletal remodeling and signaling pathways involving Rho family GTPases and Rho-associated protein kinase (ROCK) activity.21 Cargo sorting into MVs occurs through multiple mechanisms, including lipid raft-associated localization, glycosylphosphatidylinositol anchoring, and sequence-dependent RNA and protein recruitment.22,23 Subsequent actin– myosin contraction and ATP-dependent cytoskeletal forces facilitate membrane scission and release of MVs into the extracellular space.24 Intracellular Ca2⁺ signaling, CDC42 activation, and actin dynamics further regulate this shedding process.25
Apoptotic Bodies
Apoptotic bodies are large EVs generated during programmed cell death, with a broad size range of approximately 50–5000 nm.26 Their formation is closely linked to apoptotic membrane remodeling and reflects the structural and molecular composition of the parent dying cell. During apoptosis, increased intracellular pressure and cytoskeletal breakdown lead to membrane blebbing, resulting in the formation of dynamic membrane protrusions surrounding nuclear and cytoplasmic material.27 Progressive nuclear condensation and fragmentation further contribute to the encapsulation of cellular components into discrete vesicular structures.28 These fragmented cellular components are ultimately packaged into apoptotic bodies, which contain proteins, nucleic acids, and organelle remnants derived from the parent cell.29 Apoptotic bodies may express markers such as caspase-3-associated components and histones, reflecting their apoptotic origin.30 Under physiological conditions, they are efficiently cleared by phagocytic cells, particularly macrophages, through phosphatidylserine-dependent recognition pathways involving Annexin V binding.31
Exosomes, microvesicles, and apoptotic bodies represent distinct yet partially overlapping populations of EVs that differ in their biogenetic origin, size distribution, and molecular composition. In LC, these EV populations participate in multiple aspects of tumor progression by shaping the tumor microenvironment and influencing cellular behavior. Their biological diversity also has important implications for the development of EV-based diagnostic and therapeutic approaches, while simultaneously creating challenges for standardization and clinical translation.
The Role of EVs in LC Progression
Tumor cells exhibit high proliferative activity and release abundant EVs, which serve as key mediators of intercellular communication within the tumor microenvironment. In LC, EVs are implicated in multiple hallmarks of disease progression, including tumor proliferation, immune modulation, and the development of therapeutic resistance.32 Through both paracrine and systemic signaling, EVs facilitate bidirectional communication between malignant and stromal cells, contributing to tumor microenvironment remodeling and establishing a permissive niche for tumor growth and metastatic dissemination.33 Building on these concepts, the following sections explore the key mechanistic pathways through which EVs regulate LC progression.
Angiogenesis
Tumor angiogenesis is a critical pathological process that supports cancer cell survival and promotes tumor growth, invasion, and metastatic dissemination.34 In the early stages of tumor development, hypoxic stress and microenvironmental alterations stimulate tumor cells to release abundant EVs.35 These EVs circulate through biological fluids and are subsequently taken up by vascular endothelial cells within the tumor microenvironment, where they modulate key cellular functions including proliferation, migration, and vascular remodeling, facilitating angiogenic activation.36 At the mechanistic level, EV-derived bioactive cargo such as non-coding RNAs and functional proteins disrupt endothelial junction integrity by downregulating cadherin and β-catenin expression, resulting in reduced intercellular adhesion and enhanced endothelial motility.37 EVs signaling activates transcriptional regulation associated with angiogenesis, leading to upregulation of pro-angiogenic factors and receptors including VEGFA, FGF2, PDGFB, and ANGPT2.38 This molecular reprogramming promotes endothelial cell proliferation and contributes to neovessel formation. Subsequently, activated endothelial cells secrete cytokines and growth factors that further amplify angiogenic signaling. This process is reinforced through the PI3K/Akt signaling axis, which facilitates pericyte activation and stabilization of newly formed vascular structures, sustaining progressive vascular expansion.39 In LC, multiple EV-associated molecular regulators have been implicated in angiogenic modulation. Tumor-derived EVs enriched with miR-671-3p suppress KLF2 expression in endothelial cells, leading to increased vascular endothelial growth factor receptor 2 (VEGFR2) expression and enhanced angiogenic signaling; inhibition of miR-671-3p effectively disrupts this regulatory axis.40 Similarly, EVs carrying miR-197-3p, miR-23a, and miR-141 promote angiogenesis through distinct mechanisms: miR-197-3p downregulates TIMP2 and TIMP3 expression to activate the ERK pathway, miR-23a suppresses PTEN signaling, and miR-141 targets KLF12 to induce abnormal endothelial proliferation and vascular remodeling.41,42 EVs containing YAP protein enhance angiogenic potential in a concentration-dependent manner and significantly increase endothelial tube formation when co-cultured with HUVECs.43 EVs orchestrate angiogenic processes in LC through coordinated effects on endothelial signaling, gene regulation, and microenvironmental remodeling, contributing to sustained tumor vascularization and disease progression.
Metastasis
The invasion and metastasis of LC represent a highly complex, multi-step cascade process that includes local invasion of the primary tumor, intravasation into the vasculature, survival in circulation, extravasation into distant tissues, and eventual colonization within the microenvironment of secondary organs to establish metastatic lesions.44 Accumulating evidence indicates that EVs, derived from both tumor and stromal cells within the tumor microenvironment, act as critical mediators of intercellular communication and play a central role in regulating both local invasion and distant metastatic colonization in LC.45 During the early phases of metastasis, tumor-derived EVs (TDEs) promote metastatic dissemination through complementary mechanisms.46 On one hand, they enhance tumor angiogenesis, while on the other hand, they compromise vascular integrity by disrupting tight junctions and adherens junctions in vascular endothelial cells, degrading the vascular basement membrane, and weakening extracellular matrix barriers. These alterations increase vascular permeability, facilitating tumor cell intravasation and entry into the systemic circulation, establishing the structural basis for distant metastasis.47 Beyond vascular remodeling, TDEs actively contribute to the formation of a pre-metastatic niche in distant organs. This is achieved through the activation of key signaling pathways, including NLRP6/NF-κB, OSMR/JAK/STAT3, and PTPRD/STAT3, which collectively promote immunosuppressive and tumor-supportive microenvironmental conditions that favor metastatic colonization.48 TDEs also enhance the invasive potential of tumor cells by inducing epithelial–mesenchymal transition (EMT) and stemness-associated phenotypes through activation of the NF-κB/SOX2 axis, accompanied by upregulation of EMT-related markers such as VIM, CDH2, ZEB2, TWIST, and SOX family genes.47 In LC-specific regulatory contexts, EV-associated miRNAs and non-coding RNAs further amplify metastatic progression. For instance, EV-enriched miR-499a promotes EMT through modulation of the mTOR pathway, while miR-31 facilitates metastasis by targeting and silencing SATB2 and activating MEK/ERK signaling.49 Similarly, non-coding RNAs including LINC00963, circ_0001715, miR-19b-3p, and miR-770 contribute to metastatic niche remodeling by regulating macrophage polarization toward an M2 phenotype, reinforcing immunosuppression and enhancing LC invasion and metastasis.50
Immune Evasion
The programmed cell death protein 1 (PD-1)/programmed death-ligand 1 (PD-L1) axis represents a central immune checkpoint pathway and a major focus in tumor immunotherapy. PD-1 is predominantly expressed on activated T lymphocytes and interacts with its ligands, PD-L1 and PD-L2, which are present on tumor cells as well as immune cells, including T and B cells. This interaction leads to functional exhaustion of T cells, impairing anti-tumor immune surveillance and cytotoxic activity.51 PD-1 engagement triggers phosphorylation of the intracellular immunoreceptor tyrosine-based switch motif (ITSM), leading to recruitment and activation of SHP2 phosphatase. This event suppresses downstream signaling cascades, including PI3K/AKT, RAS/Raf, and PLCγ pathways, ultimately resulting in reduced T cell proliferation, diminished survival capacity, and decreased production of key anti-tumor cytokines.52,53 Immune checkpoint inhibitors such as atezolizumab and durvalumab targeting the PD-1/PD-L1 pathway have demonstrated significant therapeutic efficacy across multiple malignancies.54,55 Within the tumor microenvironment, TDEs have emerged as important regulators of immune evasion in LC, particularly through the transfer of functional PD-L1. Multiple experimental studies have confirmed that LC-derived EVs carry biologically active PD-L1 and contribute directly to immune suppression.56 PD-L1⁺ EVs inhibit CD8⁺ T cell proliferation and activation, reduce the secretion of anti-tumor cytokines such as IL-2 and IFN-γ, and suppress NF-κB signaling activity. They promote PD-L1 expression in macrophages through glycolysis-associated metabolic reprogramming, reinforcing an immunosuppressive tumor microenvironment through coordinated cellular crosstalk.57 In addition to local effects, PD-L1⁺ EVs can disseminate systemically via lymphatic drainage and circulation, leading to widespread suppression of anti-tumor immune responses and facilitating immune evasion as well as metastatic progression in LC.57 Furthermore, LC-derived EVs exhibit elevated expression of the transmembrane protein CD47, which interacts with signal regulatory protein alpha (SIRPα) on macrophages to deliver a “don’t eat me” signal, inhibiting phagocytic clearance and further enabling tumor immune escape.58
Drug Resistance
Therapeutic resistance represents a major clinical barrier in oncology and is responsible for a substantial proportion of cancer-related mortality, estimated at approximately 90%.59 Increasing evidence indicates that EVs play a central role in the development and propagation of drug-resistant tumor phenotypes.60 One key mechanism involves intercellular transfer of resistance traits from drug-resistant to drug-sensitive tumor cells via EV-mediated delivery of functional biomolecules, including proteins and non-coding RNAs. For instance, tumor cells can secrete EVs enriched with the anti-apoptotic protein Survivin, which suppresses caspase activity in recipient cells and promotes resistance to apoptosis.61 In LC, chemotherapy-induced stress has been shown to modulate EV cargo composition. Cisplatin exposure downregulates miR-100 in EVs derived from A549 cells, and these EVs regulate mTOR signaling by targeting its 3′-UTR, altering cisplatin sensitivity and influencing downstream processes such as proliferation, apoptosis, and invasion in recipient cells.62 Similarly, treatment with chidamide induces upregulation of the long non-coding RNA SNHG7 in lung adenocarcinoma-derived EVs, which promotes doxorubicin resistance by enhancing autophagy-related gene expression, including ATG5 and ATG12, in recipient cells.63 Moreover, mesenchymal-like LC cells can transmit ZEB1 mRNA via EVs, inducing epithelial–mesenchymal transition-associated drug resistance and conferring reduced sensitivity to cisplatin, gemcitabine, and combination chemotherapy regimens.64 In addition to cargo-mediated reprogramming, TDEs also contribute to drug resistance through efflux-based mechanisms. These EVs frequently exhibit elevated expression of ATP-binding cassette (ABC) transporters, including ABCG2 (BCRP, breast cancer resistance protein) and ABCC1 (MRP1, multidrug resistance-associated protein 1), which actively export chemotherapeutic agents from tumor cells, reducing intracellular drug accumulation and promoting multidrug resistance (Figure 2).65
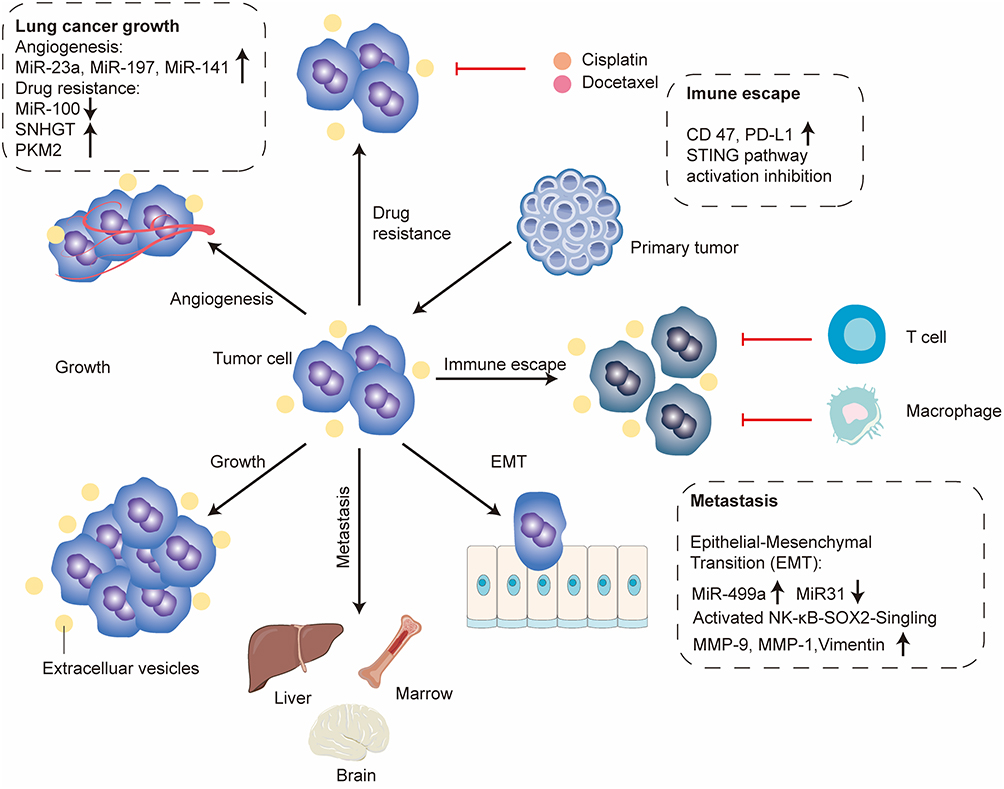 |
Figure 2 Components such as miR-23a, miR-197, miR-141, miR-100, SNHGT and PKM2 carried by extracellular vesicles regulate tumor angiogenesis and mediate cisplatin and docetaxel resistance in lung cancer. Surface molecules CD47 and PD-L1 on extracellular vesicles facilitate tumor immune escape, while miR-499a and miR-31 induce tumor epithelial-mesenchymal transition. These multiple mechanisms synergistically drive tumor progression and ultimately lead to distant organ metastasis.↑: upregulated expression; ↓: downregulated expression. |
Application of EVs in the Diagnosis and Prognosis of LC
The lack of effective early detection strategies and the frequently asymptomatic nature of early-stage disease result in a substantial proportion of LC cases being diagnosed at advanced stages, contributing to a 5-year survival rate of approximately 19.7%.66 Imaging modalities remain central to clinical diagnosis. Chest radiography is commonly used as an initial diagnostic tool, with findings such as irregular pulmonary margins, lobulated masses, pleural retraction, and patchy opacities raising suspicion of malignancy. However, its diagnostic specificity remains limited.67 When radiographic findings suggest a malignant lesion, contrast-enhanced computed tomography (CT) provides a more detailed assessment of tumor morphology, extent, and anatomical relationships with adjacent structures.1 Despite these advances, current diagnostic approaches remain insufficient for reliable early detection. Therefore, the development of accurate, minimally invasive, and reproducible diagnostic strategies remains a clinical priority. Liquid biopsy has emerged as a promising alternative owing to its minimal invasiveness and ability to facilitate longitudinal disease monitoring. Among the major liquid biopsy analytes, circulating tumor cells (CTCs), circulating tumor DNA (ctDNA), and EVs have received considerable attention as potential biomarkers for LC diagnosis and prognosis.68 The following sections summarize their diagnostic applications and associated limitations.
CTCs
CTCs are malignant cells that detach from primary tumors or metastatic lesions and enter the peripheral circulation either spontaneously or during clinical interventions. Because they provide direct evidence of tumor dissemination, CTCs have attracted considerable interest as biomarkers for early cancer detection, prognostic evaluation, and risk stratification.69 Current CTC enrichment strategies primarily include density-gradient centrifugation and antigen-dependent immunomagnetic separation, while detection approaches include the CellSearch system, fiber-optic array scanning technology (FAST), flow cytometry, and immunofluorescence-based assays.70 Early clinical investigations demonstrated the feasibility of CTC detection in LC. For example, Tanaka et al identified CTCs in 17 of 88 patients with suspected or confirmed primary LC using the CellSearch platform, corresponding to a detection rate of 19.3%.71 Similarly, a study involving 101 untreated patients with stage III–IV NSCLC reported detectable CTCs in 21 individuals.72 Despite their clinical potential, several challenges limit the routine application of CTCs. Their extremely low abundance in peripheral blood complicates reliable detection, and a large proportion of CTCs are rapidly eliminated from circulation following tumor cell dissemination.73 Currently employed detection technologies display important technical limitations. The CellSearch system relies largely on EpCAM expression and may fail to identify EpCAM-negative tumor cells, whereas FAST requires relatively large sample volumes. Flow cytometry and immunofluorescence-based approaches may also be affected by suboptimal specificity. These limitations continue to hinder the widespread clinical implementation of CTC-based diagnostics in LC.74
ctDNA
cfDNA comprises extracellular DNA fragments released into the circulation primarily through apoptosis and necrosis. The fraction of cfDNA derived specifically from tumor cells is termed ctDNA, which can be detected in peripheral blood and other body fluids, including cerebrospinal fluid and urine.75,76 Because ctDNA retains tumor-specific molecular alterations, including somatic mutations, DNA methylation patterns, and microsatellite instability, it provides a valuable non-invasive biomarker for monitoring tumor evolution and disease progression in real time.77 Furthermore, ctDNA levels gradually increase with tumor burden and disease progression.78 Advances in molecular detection technologies, including amplification refractory mutation system (ARMS), quantitative PCR (qPCR), digital PCR, and next-generation sequencing (NGS), have substantially improved the sensitivity and reliability of ctDNA analysis since 2016. Thus, ctDNA has become an important tool for assessing tumor heterogeneity, monitoring disease dynamics, and evaluating treatment responses across multiple malignancies, including LC, breast cancer, and colorectal cancer.79 Clinical studies have demonstrated the diagnostic value of ctDNA in LC. In one investigation involving 68 patients with NSCLC, ctDNA was detected in more than 80% of serum samples, with TP53, KRAS, and EGFR being the most frequently mutated genes.80 A subsequent meta-analysis reported a pooled sensitivity of 68% and a specificity of 98% for ctDNA-based EGFR mutation detection.81 Aberrant methylation of genes including MGMT, p16INK4a, RASSF1A, DAPK, and RAR-β has been proposed as a potential biomarker panel for LC detection and risk assessment.82 Although ctDNA offers high specificity and clinically meaningful sensitivity, its application remains constrained by biological and technical challenges. ctDNA typically constitutes only a small fraction of total circulating nucleic acids and exhibits a short half-life, often less than two hours, making its detection susceptible to pre-analytical and analytical variability. These limitations continue to affect its accessibility and routine clinical implementation.83
EVs
EVs have emerged as promising liquid biopsy biomarkers for LC because they encapsulate diverse molecular cargo that reflects the biological characteristics of their cells of origin. Among these cargo molecules, miRNAs, long non-coding RNAs (lncRNAs), and proteins have demonstrated considerable potential for early detection and disease monitoring.84 As early as 2013, Harvey I. Pass et al employed the ExoQuick precipitation method to isolate peripheral blood-derived EVs from 10 patients with lung adenocarcinoma, 10 patients with lung granulomas, and 10 age- and sex-matched healthy smokers, followed by miRNA expression profiling using RT-PCR. Expression profiling identified significant upregulation of miR-151a-5p, miR-30a-3p, miR-200b-5p, miR-629, miR-100, and miR-154-3p in TDEs. The resulting biomarker panel achieved high diagnostic performance, with reported sensitivities exceeding 95% and an area under the receiver operating characteristic (ROC) curve greater than 0.90.85 Subsequent investigations further supported the diagnostic value of EV-associated miRNAs in NSCLC. In a cohort comprising 100 patients with NSCLC and 90 healthy controls, elevated expression of several EV-derived miRNAs was observed. Among these candidates, miR-17-5p demonstrated an area under the ROC curve (AUC) of 0.746, with a sensitivity of 70.0% and specificity of 82.2% for NSCLC detection. Its diagnostic performance exceeded that of several routinely used serum biomarkers, including carcinoembryonic antigen (CEA), cytokeratin 19 fragment (CYFRA21-1), and squamous cell carcinoma antigen (SCCA), highlighting the potential clinical value of EV-associated biomarkers in LC detection.86 Beyond individual miRNA candidates, numerous studies have identified additional EV-derived non-coding RNAs with potential diagnostic relevance, including miR-23a, miR-30b/30c, let-7 family members, miR-23a-3p, miR-486-5p, PITPNA-AS1, Mir100hg, TBILA, and AGAP2-AS1.87–89 Protein cargoes carried by TDEs have also attracted attention as candidate biomarkers, with PD-L1, CD317, EGFR, A-kinase anchoring protein 4, and CD9 among the most frequently reported markers associated with LC.90 Although peripheral blood remains the most commonly investigated source of EVs, other biological fluids may provide additional diagnostic opportunities. Bronchoalveolar lavage fluid (BALF) and sputum are enriched in tumor-associated EVs and may offer greater proximity to the primary tumor site. For example, comparative RNA sequencing of EVs isolated from BALF revealed significantly elevated miR-1246 expression in patients with LC compared with individuals with benign pulmonary nodules. Diagnostic analysis yielded an AUC of 0.743, with a sensitivity of 80% and specificity of 60%. Functional studies further demonstrated that miR-1246 promotes tumor cell proliferation, migration, and invasion, whereas its inhibition suppresses these malignant phenotypes.91 Other biomarkers identified in BALF- and sputum-derived EVs, including miR-126, Let-7a, miR-21, miR-155, miR-210, and E-cadherin, have similarly shown potential utility for LC detection92,93 (Figure 3).
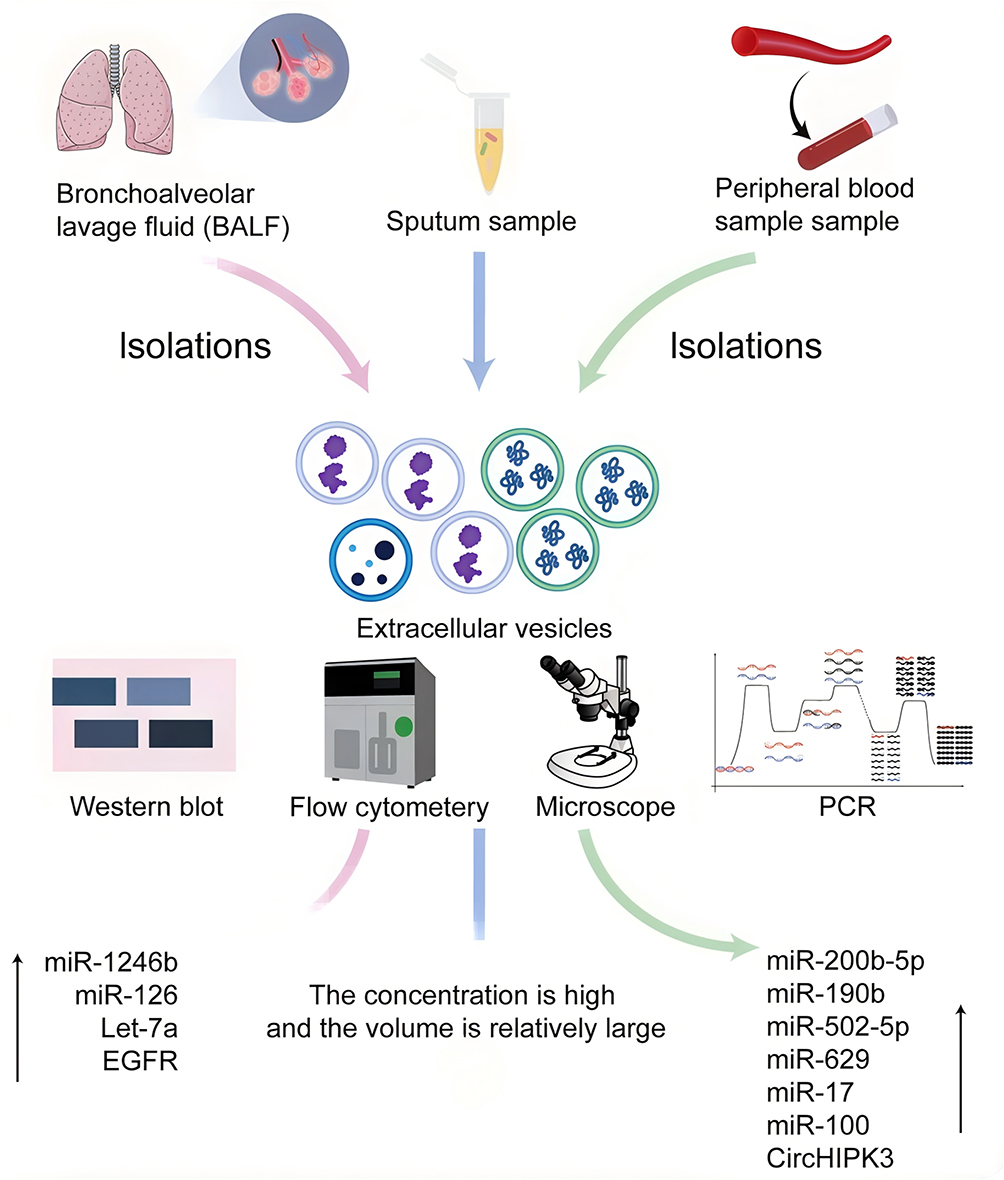 |
Figure 3 EVs as diagnostic biomarkers in lung cancer. EV-based diagnostic strategies employ multiple analytical platforms, including Western blot (WB), flow cytometry, PCR-based assays, nanoparticle tracking analysis, and morphological characterization, to evaluate EVs cargo derived from clinical specimens. EVs isolated from sputum, peripheral blood, and BALF of lung cancer patients exhibit distinct molecular signatures. BALF-derived EVs show elevated expression of miR-1246b, miR-190b, and miR-126, while blood-derived EVs are enriched in miR-502-5p, miR-629, and miR-17. Sputum-derived EVs display comparatively larger particle size distributions relative to non-malignant controls, highlighting their diagnostic heterogeneity and clinical utility. ↑: upregulated expression. |
CTCs, ctDNA, and EVs each provide unique advantages for liquid biopsy applications. However, the low abundance of CTCs and the limited stability of ctDNA present important analytical challenges. EVs are released in large quantities, possess a protective lipid bilayer that preserves their molecular cargo, and are widely distributed across biological fluids. These characteristics facilitate the detection of diverse nucleic acid and protein biomarkers and support applications in early diagnosis, disease monitoring, therapeutic response assessment, and drug resistance surveillance. As methodologies for EV isolation and characterization continue to improve, EV-based liquid biopsy approaches are expected to play an increasingly important role in precision diagnostics and personalized management of LC (Table 1).
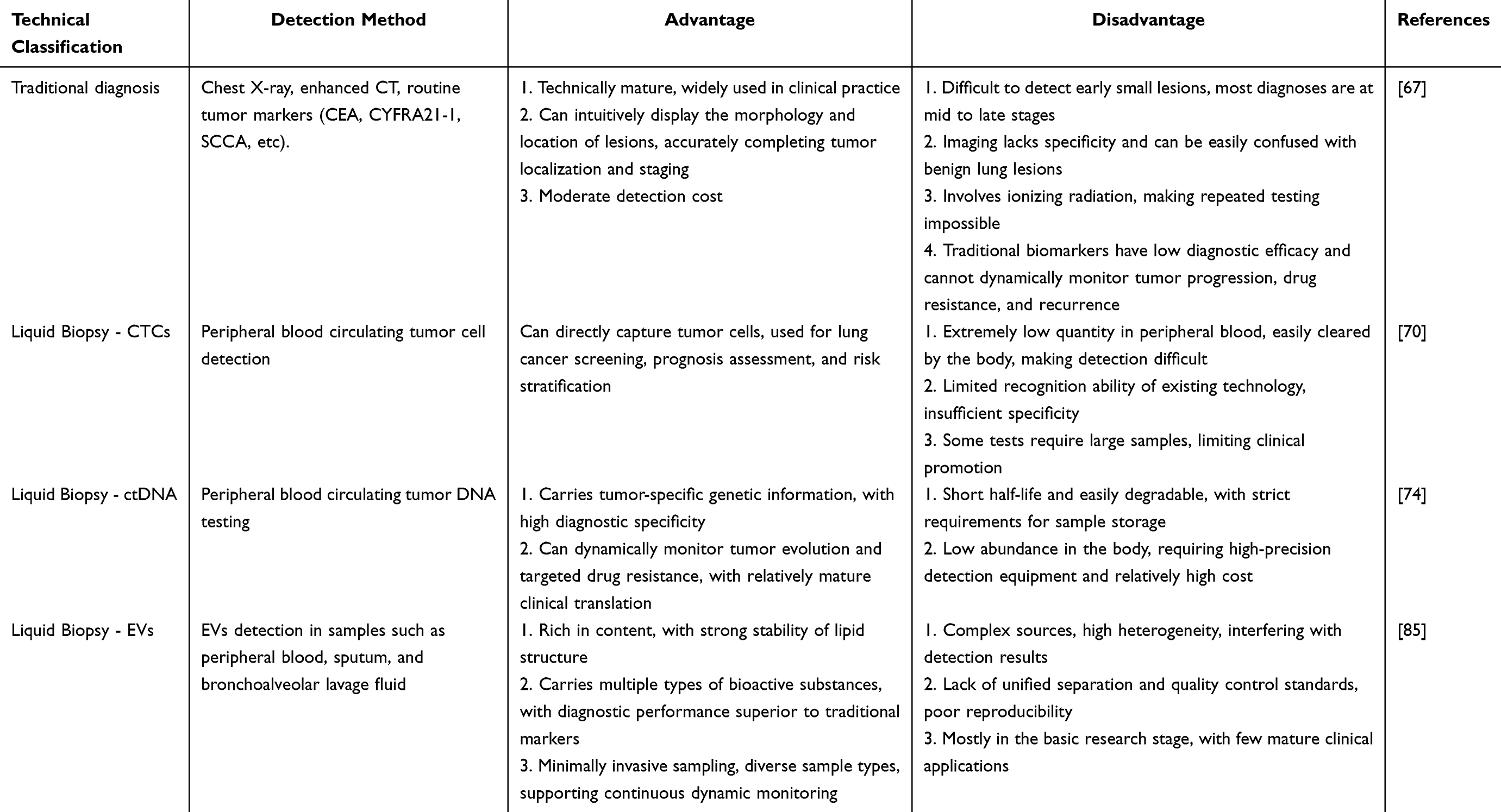 |
Table 1 Comparison of Advantages and Disadvantages Between Traditional Lung Cancer Diagnosis and Liquid Biopsy Diagnostic Techniques |
Application of EVs in LC Treatment
Current EV-based therapeutic strategies in LC can be broadly categorized into two complementary approaches. The first approach focuses on mitigating the pathological effects of tumor-derived EVs by reducing their production, release, or functional interaction with recipient cells. This includes pharmacological inhibition of EV biogenesis, such as using sphingomyelinase inhibitors like GW4869, which suppress the production of pro-tumorigenic EVs. Other strategies involve blocking EV uptake or neutralizing circulating EVs to limit their ability to transfer oncogenic cargo and promote malignant progression.94 The second approach involves the therapeutic engineering of EVs as natural nanocarriers for drug delivery. In this context, EVs can be loaded with diverse therapeutic agents, including small-molecule chemotherapeutic drugs, nucleic acid-based therapeutics, and immune checkpoint inhibitors. Owing to their intrinsic biocompatibility and targeting capability, engineered EVs can preferentially accumulate within the tumor microenvironment, where they exert anti-tumor effects through inhibition of tumor cell proliferation and induction of cancer cell death.95 These strategies highlight the dual role of EVs in LC therapy, encompassing both inhibition of pathogenic EV signaling and exploitation of ECs as precision drug delivery systems.
Inhibition of Pathogenic EV Secretion
EVs serve as essential mediators of intercellular communication by facilitating the transfer of bioactive molecules between donor and recipient cells.96 EV biogenesis and function depend on two key processes: the regulated formation and release of vesicles from donor cells and subsequent uptake of EVs by recipient cells through receptor-mediated binding, membrane fusion, or endocytosis, enabling intercellular signal transmission.97 Accordingly, therapeutic strategies aimed at disrupting EV-mediated communication in LC primarily focus on interfering with vesicle biogenesis, secretion, or cellular uptake. One major pharmacological approach involves inhibition of EVs biogenesis and release. GW4869, a neutral sphingomyelinase inhibitor, suppresses ceramide-dependent EVs formation and has been widely used to attenuate the secretion of TDEs within the tumor microenvironment.98 In LC models, GW4869 has been shown to reduce the release of EVs carrying oncogenic miRNAs, suppress aberrant signaling pathways, and reverse drug-resistant phenotypes. For example, inhibition of miR-7-containing EVs has been associated with restoration of Hippo pathway regulation and reversal of gefitinib resistance in LC cells.99 EV suppression via GW4869 enhances the cytotoxic effects of chemotherapeutic agents such as cisplatin and etoposide in vitro.100 Other small molecules, including pantethine and imipramine, have also been reported to modulate EV secretion indirectly through the regulation of lipid metabolic pathways. A second strategy targets intracellular trafficking and cytoskeletal regulation involved in EV release. Inhibition of Rho-associated coiled-coil-containing protein kinase (ROCK) using agents such as Y27632 disrupts actin cytoskeleton dynamics, impairing membrane budding and reducing EV secretion.101 Similarly, Manumycin A, a Ras pathway inhibitor, interferes with ESCRT-associated EV biogenesis and has been reported to reduce tumor-derived EV release by approximately 50%–60% under experimental conditions.102 In addition to pharmacological approaches, genetic modulation of Rab GTPases represents a key regulatory strategy for suppressing EV production and trafficking. Proteins such as Rab4, Rab11, Rab15, Rab35, and Rab27A regulate multiple stages of EV biogenesis, intracellular transport, and vesicle secretion. Their inhibition has been shown to significantly impair EV release in tumor systems.103 In LC models, Rab27A knockdown reduces EV-mediated systemic effects, including adipocyte reprogramming and cancer-associated cachexia, by limiting EV uptake and downstream PKA signaling activation in adipose tissue.104 These results indicate that pharmacological and genetic targeting of EVs biogenesis and secretion pathways may provide a promising strategy to attenuate tumor-promoting intercellular communication in LC. However, further studies are required to determine the translational feasibility, specificity, and systemic safety of these approaches in clinical settings.
Using EVs Directly as Therapeutic Agents
Because EVs selectively incorporate proteins, lipids, nucleic acids, and other bioactive molecules from their parent cells during biogenesis, they often retain important functional characteristics of their cellular origin. Consequently, in addition to strategies aimed at suppressing pathogenic EV signaling, increasing attention has been directed toward the therapeutic use of EVs as cell-free biological agents capable of modulating tumor progression in LC. Among the most extensively studied candidates are mesenchymal stem cell-derived EVs (MSC-EVs). Mesenchymal stem cells (MSCs) possess immunomodulatory, anti-inflammatory, and tissue repair properties while exhibiting relatively low immunogenicity due to limited expression of major histocompatibility complex class II (MHC-II) molecules and co-stimulatory proteins.105 These biological characteristics are partially retained by MSC-EVs, which offer further advantages including improved stability, lower toxicity, and reduced risk of immune rejection compared with cell-based therapies. As a result, MSC-EVs have emerged as promising cell-free therapeutic platforms for cancer treatment. Experimental studies have demonstrated that MSC-EVs can suppress malignant behaviors in LC through the delivery of functional regulatory molecules. For example, human umbilical cord mesenchymal stem cell-derived EVs (hUCMSC-EVs) enriched in miR-320a inhibit LC progression through negative regulation of the SOX4/Wnt/β-catenin signaling pathway.106 Similarly, bone marrow MSC-derived EVs carrying let-7i suppress LC cell proliferation, migration, and invasion through downregulation of lysine demethylase 3A (KDM3A), whereas inhibition of let-7i largely abolishes these anti-tumor effects.107 However, the biological activity of MSC-EVs is not uniformly tumor suppressive. Emerging evidence indicates that certain MSC-derived populations may facilitate tumor progression depending on their molecular cargo. EVs carrying miR-21-5p, miR-410, and other regulatory molecules have been reported to enhance LC cell proliferation, promote EMT, increase migratory capacity, and induce macrophage polarization toward a pro-tumorigenic M2 phenotype.108 These observations highlight the importance of rigorous cargo characterization, functional validation, and quality control when developing MSC-EV-based therapeutic strategies.
Beyond stem cell-derived vesicles, EVs released by immune cells have also attracted considerable interest as therapeutic agents capable of reversing the immunosuppressive tumor microenvironment characteristic of LC.109 Dendritic cell-derived EVs contain functional MHC-peptide complexes, co-stimulatory molecules, cytokines, and chemokines that support antigen presentation and activation of both innate and adaptive immune responses.110 Natural killer (NK) cell-derived EVs carry cytotoxic effector molecules, including perforin, granzymes, Fas ligand, and NKp46, enabling them to induce oxidative stress, DNA damage, and apoptosis in tumor cells.111 T cell-derived EVs contribute to antitumor immunity through multiple mechanisms. EVs released from activated CD8⁺ T cells can suppress tumor invasion and metastatic dissemination, whereas EVs secreted by IL-2-stimulated CD4⁺ T cells are enriched in regulatory miRNAs, including miR-25-3p, miR-155-5p, miR-215-5p, and miR-375. Through paracrine transfer of these molecules, CD4⁺ T cell-derived EVs enhance CD8⁺ T cell proliferation and cytotoxic activity, strengthening anti-tumor immune responses.109
Using EVs as Drug Delivery Systems
Conventional therapeutic modalities for LC, including chemotherapy, radiotherapy, immunotherapy, and gene-based therapies, are frequently limited by inefficient drug accumulation at tumor sites and dose-limiting systemic toxicity. Although increasing drug dose may enhance local therapeutic exposure, it often results in undesirable off-target effects in healthy tissues and organs. Consequently, the development of safe and efficient drug delivery platforms has become a major focus of contemporary cancer research. Among emerging delivery systems, EVs have attracted considerable attention because of their intrinsic biocompatibility, low immunogenicity, prolonged circulation stability, and natural ability to mediate intercellular cargo transfer. Compared with conventional nanocarriers, including liposomes, polymeric nanoparticles, mesoporous materials, and metallic nanoparticles, EVs possess unique biological properties that facilitate cellular uptake and improve delivery efficiency within complex tumor microenvironments. As a result, EV-based drug delivery platforms have become an increasingly active area of investigation in LC therapy (Figure 4).
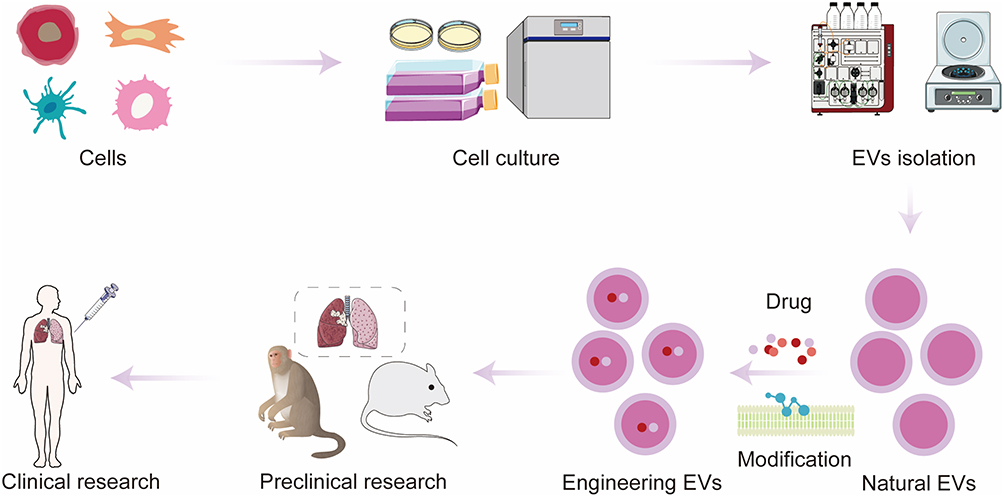 |
Figure 4 EVs have emerged as promising nanocarriers for targeted therapeutic delivery against lung cancer, with substantial efforts advancing their clinical translation. To generate clinical-grade EVs, these nanoparticles are initially purified from the conditioned culture medium of donor cells via three mainstream isolation approaches: ultracentrifugation, size-exclusion chromatography, and microfluidic separation. Following isolation, therapeutic cargoes encompassing chemotherapeutics, nucleic acids, and functional proteins are loaded into EVs using either endogenous biogenesis-mediated packaging or exogenous post-isolation loading techniques. Genetic manipulation of parent cells and chemical bioconjugation (eg, click chemistry) are two primary strategies to engineer EVs surface moieties, which drastically improves their specific tropism toward lung tumor lesions. Accumulating preclinical investigations across mouse, rabbit, and non-human primate animal models have validated the potent anti-tumor performance of EV-based therapeutics, laying robust preclinical evidence to support the clinical advancement of EVs delivery systems for lung cancer intervention. |
Chemotherapy
LC is broadly classified into SCLC and NSCLC based on histopathological characteristics.112 Surgical resection remains the preferred treatment for early-stage disease, whereas chemotherapy and radiotherapy constitute the principal therapeutic approaches for advanced-stage tumors.113 Platinum-based regimens, particularly combinations of etoposide with cisplatin or carboplatin, continue to serve as standard treatment options for many patients with LC.114 However, their clinical efficacy is frequently constrained by poor tumor selectivity, limited bioavailability, systemic toxicity, and the emergence of drug resistance.115 EV-based delivery systems have shown potential for overcoming several of these limitations. In an animal study conducted by Cui et al, cisplatin-loaded EVs derived from the breast cancer cell line MDA-MB-231 were preferentially internalized by A549 lung cancer cells following co-culture. No significant cytotoxicity was observed in non-malignant 293T or dendritic cells (DCs). In vivo administration of drug-loaded EVs significantly reduced tumor burden and prolonged survival in tumor-bearing mice, supporting the feasibility of EVs as targeted chemotherapy carriers.116 In addition to enhancing drug delivery, EVs may contribute to overcoming chemotherapy resistance. Jin et al reported that airway administration of paclitaxel-loaded macrophage-derived exosomes significantly increased the sensitivity of Lewis lung carcinoma cells expressing high levels of P-glycoprotein (Pgp) to paclitaxel treatment. Furthermore, cellular uptake of exosome-loaded paclitaxel was approximately 30-fold higher than that observed with paclitaxel-loaded liposomes and polystyrene nanoparticles, highlighting the superior delivery efficiency of EV-based systems.117
Radiation Therapy
Radiotherapy represents a fundamental component of LC management, with a substantial proportion of patients receiving radiation treatment during the course of their disease.118 Stereotactic ablative radiotherapy (SABR) has demonstrated favorable outcomes in patients with medically inoperable early-stage disease, while integration of radiotherapy with systemic treatment has improved progression-free survival (PFS) in selected patients with advanced-stage LC.119,120 Despite these clinical benefits, several factors limit the effectiveness of radiotherapy. Tumor heterogeneity, hypoxia, physiological respiratory motion, and the proximity of radiosensitive organs increase the difficulty of delivering therapeutic radiation while minimizing damage to surrounding tissues.121 Prolonged radiation exposure can induce adaptive resistance mechanisms involving altered DNA damage responses, cell-cycle regulation, signaling pathway activation, and metabolic reprogramming.122 Emerging evidence suggests that EVs participate in radiotherapy resistance and may serve as vehicles for radiosensitizing agents. For example, irradiation of A549 cells has been shown to increase the expression of miR-1246, which promotes radioresistance through suppression of death receptor 5 (DR5) and facilitates resistance transfer to neighboring cells.123 Radiation exposure has also been reported to stimulate EV release and influence immune-cell polarization within the tumor microenvironment.124 To improve radiosensitivity, investigators have explored EV-mediated delivery of therapeutic molecules. Microvesicles derived from bone marrow MSCs carrying miR-101-3p were shown to enhance the radiosensitivity of NSCLC cells through suppression of enhancer of zeste homolog 2 (EZH2) and subsequent inhibition of the PI3K/AKT/mTOR signaling pathway.125 Similarly, Liang et al developed M1 macrophage-derived EVs loaded with catalase (CAT), anti-PD-L1 (programmed death ligand-1) nanobodies, and DNA damage repair inhibitors (DDRi). This multifunctional platform alleviated tumor hypoxia, impaired DNA repair, relieved immune suppression, and enhanced radiotherapeutic efficacy through coordinated modulation of multiple resistance mechanisms.126
Immunotherapy
Immunotherapy has transformed the therapeutic landscape of LC by harnessing host immune responses to recognize and eliminate malignant cells.12 Immune checkpoint inhibitors targeting PD-1, PD-L1, and anti-cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) have demonstrated substantial clinical benefits and are now incorporated into standard treatment strategies for multiple LC subtypes.127 The PD-1/PD-L1 axis plays a central role in tumor immune evasion. Binding of PD-L1 expressed on tumor cells to PD-1 receptors on T lymphocytes suppresses T-cell activation, proliferation, effector function, and survival, weakening anti-tumor immunity.128 Therapeutic blockade of this pathway can restore immune activity and inhibit tumor progression.129 However, systemic administration of immune checkpoint inhibitors may be associated with immune-related adverse events (irAEs), including autoimmune complications. To improve therapeutic specificity and reduce systemic toxicity, EVs have increasingly been investigated as carriers for immunotherapeutic agents and cancer vaccines. A Phase II clinical study demonstrated that EVs derived from IFN-γ-stimulated DCs enhanced NK cell and T cell responses and were associated with improvements in PFS and overall survival (OS) among patients with LC.130 Similarly, EVs derived from CD4+ T cells and enriched with miR-25-3p, miR-155-5p, miR-215-5p, and miR-375 have been shown to stimulate CD8+ T-cell activity without promoting regulatory T-cell expansion, supporting their potential application in cancer immunotherapy.131 Advanced EV engineering strategies have further expanded the therapeutic potential of these vesicles. Huang et al developed chimeric antigen receptor T-cell-derived exosomes co-expressing mesothelin (MSLN) and PD-L1 targeting single-chain variable fragments. In preclinical models, these engineered EVs enhanced anti-tumor activity and prolonged survival. To address limitations associated with drug loading and delivery efficiency, the investigators subsequently integrated lung-targeting liposomes with engineered exosomes to create the Lip-CExo platform, combining the advantages of both delivery systems.132 In another study, Ning et al developed an EV-based multifunctional therapeutic platform by encapsulating indocyanine green-loaded manganese dioxide nanoparticles within anti-PD-L1-modified exosomes. Within the acidic tumor microenvironment, controlled release of anti-PD-L1 enhanced T-cell activation, while manganese dioxide relieved hypoxia through catalytic oxygen generation. Upon near-infrared (NIR) irradiation, indocyanine green generated reactive oxygen species, resulting in synergistic integration of immunotherapy and photodynamic therapy for LC treatment.133 These studies demonstrate the versatility of EVs as drug delivery platforms capable of integrating chemotherapy, radiotherapy sensitization, immunotherapy, and combination treatment strategies. Continued development of multifunctional EV-based systems that incorporate targeted delivery, stimulus-responsive release mechanisms, and complementary therapeutic modalities is expected to further advance precision treatment approaches for LC (Table 2).
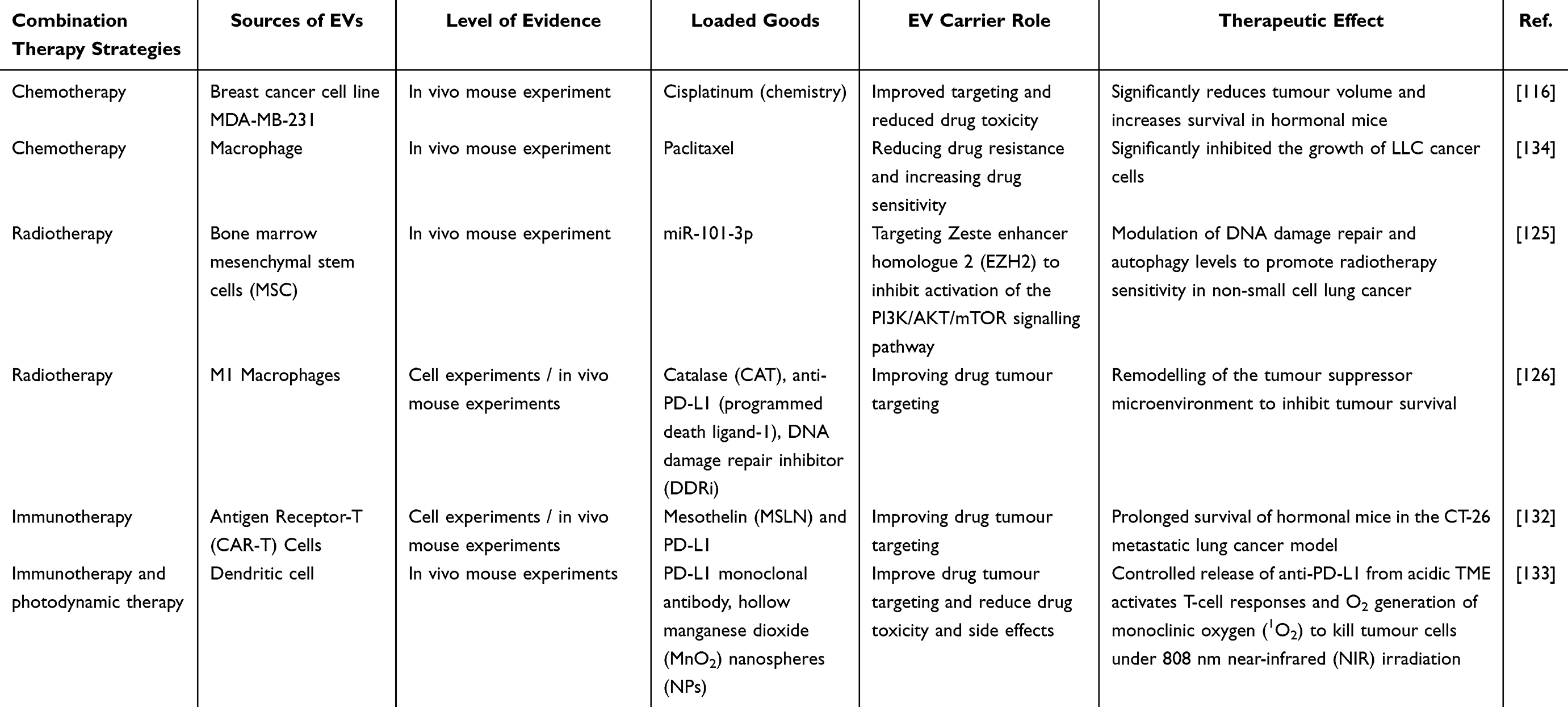 |
Table 2 EVs as Drug Delivery Systems in the Treatment of LC |
Existing Bottlenecks in Clinical Translation
Despite the considerable promise of EVs in LC diagnosis and therapy, their clinical translation remains largely confined to experimental and preclinical settings.135 Several scientific, technological, and manufacturing challenges continue to impede the development of standardized, scalable, and clinically applicable EV-based platforms.136 Major obstacles include EV heterogeneity, limitations in large-scale production and purification, storage instability, suboptimal drug-loading methodologies, the absence of standardized quality-control frameworks, and insufficient in vivo tracking technologies.137 One of the most significant challenges is the intrinsic heterogeneity of EVs populations. The molecular composition, biological activity, and functional properties of EVs are strongly influenced by the cellular origin, physiological state, and microenvironmental conditions of donor cells. Consequently, EVs derived from different sources often exhibit substantial variability in cargo composition and biological behavior. TDEs, while possessing inherent tumor-targeting capabilities, may also carry oncogenic proteins, pro-angiogenic factors, and immunosuppressive molecules that could potentially promote tumor progression and immune escape.138 Similarly, EVs derived from stem cells and immune cells may exert either tumor-suppressive or tumor-promoting effects depending on the biological status of the parent cells, raising important concerns regarding therapeutic consistency and safety.13,139 Additional complexity arises from the diversity of EVs subtypes. Exosomes, microvesicles, and apoptotic bodies differ considerably in their biogenesis, physicochemical characteristics, biodistribution, and clearance kinetics. For example, phosphatidylserine exposure on the surface of microvesicles and apoptotic bodies facilitates rapid recognition and elimination by the mononuclear phagocyte system, whereas exosomes generally demonstrate greater suitability for drug delivery applications.140 However, universally accepted criteria for selecting specific EV subtypes for clinical applications remain unavailable. Large-scale production of clinical-grade EVs also presents a major challenge. Differential ultracentrifugation remains the most widely employed isolation method, yet it often yields preparations contaminated with protein aggregates and cellular debris and may compromise vesicle integrity through exposure to high centrifugal forces.141 Size-exclusion chromatography preserves EVs structure more effectively but is limited by relatively low throughput.142 Immunoaffinity-based approaches offer high specificity but are associated with elevated costs, restricted marker availability, and potential alterations in EVs biological activity.143 Current isolation strategies suffer from limitations in yield, purity, reproducibility, and scalability, restricting their suitability for industrial and clinical implementation. Storage and long-term preservation represent another critical bottleneck. Current consensus recommends storage at 4 °C for short-term use and −80 °C for long-term preservation.144 However, prolonged storage under either condition can adversely affect EVs quality. Reductions in particle concentration, RNA content, protein stability, and surface marker integrity have been reported, together with alterations in physicochemical properties such as zeta potential.145 Furthermore, ice-crystal formation during freezing may disrupt membrane integrity and compromise biological function.145 Although cryoprotectants such as trehalose and dimethyl sulfoxide have shown potential in mitigating cryoinjury,134 and lyophilization has been explored as an alternative preservation strategy, long-term stability remains a significant unresolved issue.146 Drug loading technologies also require substantial optimization. Existing approaches, including passive incubation, electroporation, sonication, and lipid-based transfection, frequently exhibit limited loading efficiency and may compromise EVs integrity or biological activity.34,147 Physical loading procedures can induce membrane disruption and cargo leakage, whereas chemical modifications may alter surface proteins and impair natural targeting properties. Moreover, the diverse physicochemical characteristics of therapeutic cargoes, including small molecules, nucleic acids, and proteins, make the development of universally applicable loading strategies particularly challenging.148–150 Variability introduced during loading procedures further complicates batch standardization and large-scale manufacturing.151 Another major limitation is the absence of standardized quality-control systems. Current characterization approaches largely focus on particle size, concentration, morphology, surface markers, and loading efficiency. While informative, these parameters do not adequately assess critical attributes such as structural integrity, cargo stability, biodistribution, pharmacokinetics, targeting efficiency, and biological activity.152 The establishment of comprehensive and universally accepted quality standards remains essential for regulatory approval and clinical implementation. Finally, technologies for monitoring EVs biodistribution and fate in vivo remain relatively limited. Current strategies primarily rely on direct labeling methods using lipophilic fluorescent dyes such as KH26, DiD, DiR, or radioactive isotopes or bioluminescent reporter systems.153 Although these techniques have advanced understanding of EVs trafficking, challenges related to signal persistence, labeling specificity, imaging sensitivity, and quantitative interpretation continue to hinder accurate assessment of EV behavior in vivo. EVs heterogeneity, manufacturing challenges, preservation instability, inefficient drug-loading technologies, inadequate quality-control frameworks, and limitations in in vivo tracking collectively represent the principal barriers to clinical translation. Addressing these issues will be critical for the successful integration of EV-based technologies into future LC diagnostic and therapeutic strategies (Table 3).
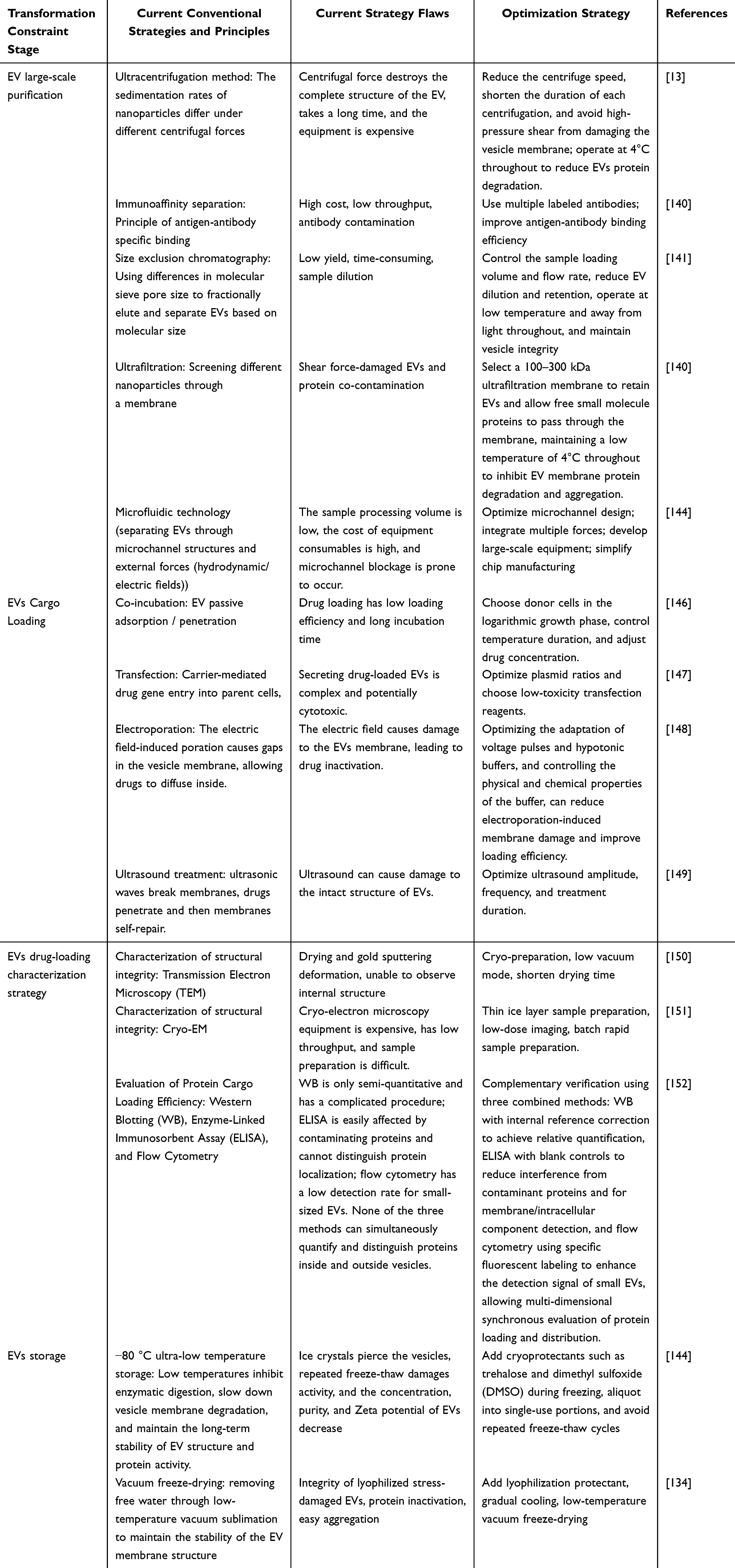 |
Table 3 Clinical Translation Constraints of EV-Mediated Drug Delivery Strategies for LC Treatment |
Conclusion
EVs have emerged as pivotal regulators of LC initiation, progression, and therapeutic response. Through the transfer of diverse bioactive cargoes, EVs mediate extensive intercellular communication within the tumor microenvironment and influence multiple hallmarks of malignancy, including angiogenesis, EMT, immune evasion, metastatic dissemination, and therapeutic resistance. Consequently, EVs occupy a central position in the molecular networks that drive LC progression. From a diagnostic perspective, EVs offer several advantages over liquid biopsy components, including CTCs and ctDNA. Their abundance in biological fluids, structural stability conferred by a lipid bilayer membrane, and capacity to protect nucleic acids and proteins from degradation make them attractive candidates for non-invasive disease detection and monitoring. The molecular cargo carried by EVs provides valuable information regarding tumor biology and has demonstrated considerable potential as a source of biomarkers for early diagnosis, prognostic evaluation, treatment monitoring, and assessment of therapeutic resistance. EVs represent a versatile platform with multiple potential applications. Strategies aimed at suppressing the production or activity of pathogenic EVs may help disrupt tumor-promoting communication networks within the tumor microenvironment. Functional EVs derived from stem cells or immune cells offer opportunities for cell-free therapeutic interventions. Moreover, the development of EV-based drug delivery systems has created new possibilities for improving the precision, efficacy, and safety of radiotherapy, chemotherapy, immunotherapy, and nucleic acid-based treatments. Despite these advances, substantial challenges continue to impede clinical translation. Key limitations include intrinsic EVs heterogeneity, difficulties in standardized large-scale production, instability during long-term storage, limited drug-loading efficiency, the absence of universally accepted quality-control standards, and insufficient understanding of EVs biodistribution and targeting behavior in vivo. Overcoming these challenges will require coordinated efforts across basic research, translational medicine, and regulatory science. Future investigations should focus on elucidating the molecular mechanisms governing EVs biogenesis and function in LC, identifying robust multi-omics biomarker signatures, and establishing standardized workflows for EV isolation, characterization, and clinical application. Advances in EVs engineering, targeted cargo delivery, and multimodal therapeutic design may further enhance their clinical utility. Equally important will be the development of rigorous quality-control frameworks and safety assessment standards to support regulatory approval and clinical adoption. With continued technological and methodological progress, EV-based approaches have the potential to become an integral component of precision oncology and to contribute substantially to improved outcomes for patients with LC.
Abbreviation
SCLC, Small cell lung cancer; NSCLC, Non-small cell lung cancer; LUAD, Lung adenocarcinoma; PI3K, Phosphatidylinositol 3-kinase; EVs, Extracellular vesicles; MVB, Multivesicular bodies; MVs, Microvesicles; ESCRT, Endosomal sorting complex required for transport; TME, Tumor microenvironment; ECs, Endothelial cells; HUVECs, Human umbilical vein endothelial cells; KLF, Krüppel-like factor; EMT, Epithelial-mesenchymal transition; ALDH, Aldehyde dehydrogenase; CSCs, Cancer stem cells; APCs, Antigen-presenting cells; TAMs, Tumor-associated macrophages; DCs, Dendritic cells; CAFs, Cancer-associated fibroblasts; CT, Computed tomography; ctDNA, Circulating tumor DNA; cfDNA, Cell-free DNA; TEM, Transmission electron microscopy; SEM, Scanning electron microscopy; HIPK3, Homologous domain interaction protein kinase 3; BALF, Bronchoalveolar lavage fluid; EGFR, Epidermal growth factor receptor; COPD, Chronic obstructive pulmonary disease; LLC, Lewis lung cancer; NSMase, Neutral sphingomyelinase; DCLK1, Dual corticotrophin-like kinase 1; NK, Natural killer; PD-1, Programmed cell death 1; PD-L1, Programmed cell death ligand 1; TDEs, Tumor-derived EVs.
Funding
This study was supported by grants from the National Natural Science Foundation (No. 82170212, No. 82370181). The Second Affiliated Hospital of the Army Medical University is a special project for clinical research (No. 2024F010).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Kim SY, Park HS, Chiang AC. Small cell lung cancer: a review. JAMA. 2025;333:1906–24.
2. Smolarz B, Łukasiewicz H, Samulak D, Piekarska E, Kołaciński R, Romanowicz H. Lung cancer-epidemiology, pathogenesis, treatment and molecular aspect (review of literature). Int J Mol Sci. 2025;26:2049.
3. Xiao Q, Tan M, Yan G, Peng L. Revolutionizing lung cancer treatment: harnessing exosomes as early diagnostic biomarkers, therapeutics and nano-delivery platforms. J Nanobiotechnol. 2025;23:232.
4. Li Y, Wu X, Yang P, Jiang G, Luo Y. Machine learning for lung cancer diagnosis, treatment, and prognosis. Genomics Proteomics Bioinf. 2022;20:850–866.
5. Yin X, Li Y, Wang H, et al. Small cell lung cancer transformation: from pathogenesis to treatment. Semin Cancer Biol. 2022;86:595–606.
6. Cao M, Chen W. Epidemiology of lung cancer in China. Thorac Cancer. 2019;10:3–7.
7. Skřičková J, Nebeský T, Kadlec B, et al. Lung cancer - dia nosis and early detection. Klin Onkol. 2021;34:6–19. doi:10.48095/ccko2021S6
8. Yang Y, Hong Y, Cho E, Kim GB, Kim IS. Extracellular vesicles as a platform for membrane-associated therapeutic protein delivery. J Extracell Vesicles. 2018;7:1440131. doi:10.1080/20013078.2018.1440131
9. Liu S, Wu X, Chandra S, et al. Extracellular vesicles: emerging tools as therapeutic agent carriers. Acta Pharm Sin B. 2022;12:3822–3842.
10. Kirian RD, Steinman D, Jewell CM, Zierden HC. Extracellular vesicles as carriers of mRNA: opportunities and challenges in diagnosis and treatment. Theranostics. 2024;14:2265–2289.
11. Miceli RT, Chen TY, Nose Y, et al. Extracellular vesicles, RNA sequencing, and bioinformatic analyses: challenges, solutions, and recommendations. J Extracell Vesicles. 2024;13:e70005.
12. Peng X, Fang J, Lou C, et al. Engineered nanoparticles for precise targeted drug delivery and enhanced therapeutic efficacy in cancer immunotherapy. Acta Pharm Sin B. 2024;14:3432–3456.
13. Wang Y, Chang YJ, Chen J, et al. Consensus on the monitoring, treatment, and prevention of leukaemia relapse after allogeneic haematopoietic stem cell transplantation in China: 2024 update. Cancer Lett. 2024;605:217264.
14. Zhang K, Fang J. Extracellular vesicles in drug delivery: from quality assurance to therapeutic application. Int J Nanomed. 2026;21:579695. doi:10.2147/IJN.S579695
15. Liu J, Ren L, Li S, et al. The biology, function, and applications of exosomes in cancer. Acta Pharm Sin B. 2021;11:2783–2797.
16. Lin W, Fang J, Wei S, et al. Extracellular vesicle-cell adhesion molecules in tumours: biofunctions and clinical applications. Cell Commun Signal. 2023;21:246.
17. Guo H, Xu Z-L, Pei X-Y, Zhao X-Y, Huang X-J. HLA haploidentical HSCT: from immune imbalance to a platform for immune homeostasis restoration. Immunity Inflam. 2025;1:12.
18. Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci. 2013;126(Pt 24):5553–5565. doi:10.1242/jcs.128868
19. Doyle LM, Wang MZ. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells. 2019;8:727.
20. Li J, Wang J, Chen Z. Emerging role of exosomes in cancer therapy: progress and challenges. Mol Cancer. 2025;24:13.
21. Li B, Antonyak MA, Zhang J, Cerione RA. RhoA triggers a specific signaling pathway that generates transforming microvesicles in cancer cells. Oncogene. 2012;31:4740–4749.
22. Shen Q, Huang Z, Yao J, Jin Y. Extracellular vesicles-mediated interaction within intestinal microenvironment in inflammatory bowel disease. J Adv Res. 2022;37:221–233.
23. Shen B, Fang Y, Wu N, Gould SJ. Biogenesis of the posterior pole is mediated by the exosome/microvesicle protein-sorting pathway. J Biol Chem. 2011;286(51):44162–44176. doi:10.1074/jbc.M111.274803
24. Faini M, Beck R, Wieland FT, Briggs JAG. Vesicle coats: structure, function, and general principles of assembly. Trends Cell Biol. 2013;23(6):279–288. doi:10.1016/j.tcb.2013.01.005
25. Sedgwick AE, Clancy JW, Olivia Balmert M, D’Souza-Schorey C. Extracellular microvesicles and invadopodia mediate non-overlapping modes of tumor cell invasion. Sci Rep. 2015;5:14748.
26. Zhou M, Li YJ, Tang YC, et al. Apoptotic bodies for advanced drug delivery and therapy. J Control Release. 2022;351:394–406.
27. Yu L, Zhu G, Zhang Z, et al. Apoptotic bodies: bioactive treasure left behind by the dying cells with robust diagnostic and therapeutic application potentials. J Nanobiotechnol. 2023;21:218.
28. Rahimian S, Mirkazemi K, Kamalinejad A, Doroudian M. Exosome-based advances in pancreatic cancer: the potential of mesenchymal stem cells. Crit Rev Oncol Hematol. 2025;207:104594.
29. Santavanond JP, Rutter SF, Atkin-Smith GK, Poon IKH. Apoptotic bodies: mechanism of formation, isolation and functional relevance. Subcell Biochem. 2021;97:61–88.
30. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). doi:10.1126/science.aau6977
31. Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol. 2013;113(1):1–11. doi:10.1007/s11060-013-1084-8
32. Beetler DJ, Di Florio DN, Bruno KA, et al. Extracellular vesicles as personalized medicine. Mol Aspects Med. 2023;91:101155. doi:10.1016/j.mam.2022.101155
33. Urabe F, Patil K, Ramm GA, Ochiya T, Soekmadji C. Extracellular vesicles in the development of organ-specific metastasis. J Extracell Vesicles. 2021;10:e12125. doi:10.1002/jev2.12125
34. Li M, Liu Y, Liu F, et al. Extracellular vesicle-based antitumor nanomedicines. Adv Healthc Mater. 2025;14(9):e2403903. doi:10.1002/adhm.202403903
35. Zhang S, Yang J, Shen L. Extracellular vesicle-mediated regulation of tumor angiogenesis— implications for anti-angiogenesis therapy. J Cell Mol Med. 2021;25(6):2776–2785. doi:10.1111/jcmm.16359
36. Yang K, Zhou Q, Qiao B, et al. Exosome-derived noncoding RNAs: function, mechanism, and application in tumor angiogenesis. Mol Ther Nucleic Acids. 2022;27:983–997. doi:10.1016/j.omtn.2022.01.009
37. Yi W, Shi Q, Wu J, Zhou B, Li Y. The impact of m6A modified cargo conveyed by exosomes on the tumor microenvironment. Discov Oncol. 2026. doi:10.1007/s12672-026-05060-7
38. Xie JY, Wei JX, Lv LH, et al. Angiopoietin-2 induces angiogenesis via exosomes in human hepatocellular carcinoma. Cell Commun Signal. 2020;18:46. doi:10.1186/s12964-020-00535-8
39. Zhao S, Mi Y, Guan B, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13:156. doi:10.1186/s13045-020-00991-2
40. Liu K, Wang Y, Li Q, et al. Hypoxia LUAD H1975 cell-derived exosomal miR-671-3p promotes angiogenesis via regulating KLF2-VEGFR2 axis. Sci Rep. 2025;15:13148. doi:10.1038/s41598-025-97488-0
41. Chang RM, Fu Y, Zeng J, Zhu XY, Gao Y. Cancer-derived exosomal miR-197-3p confers angiogenesis via targeting TIMP2/3 in lung adenocarcinoma metastasis. Cell Death Dis. 2022;13:1032. doi:10.1038/s41419-022-05420-5
42. Mao S, Lu Z, Zheng S, et al. Exosomal miR-141 promotes tumor angiogenesis via KLF12 in small cell lung cancer. J Exp Clin Cancer Res. 2020;39:193. doi:10.1186/s13046-020-01680-1
43. Wang Y, Dong L, Zhong H, et al. Extracellular vesicles (EVs) from lung adenocarcinoma cells promote human umbilical vein endothelial cell (HUVEC) angiogenesis through yes kinase-associated protein (YAP) transport. Int J Biol Sci. 2019;15:2110–2118. doi:10.7150/ijbs.31605
44. Ko J, Winslow MM, Sage J. Mechanisms of small cell lung cancer metastasis. EMBO Mol Med. 2021;13:e13122. doi:10.15252/emmm.202013122
45. Castaneda M, den Hollander P, Kuburich NA, Rosen JM, Mani SA. Mechanisms of cancer metastasis. Semin Cancer Biol. 2022;87:17–31. doi:10.1016/j.semcancer.2022.10.006
46. Zhang C, Qin C, Dewanjee S, et al. Tumor-derived small extracellular vesicles in cancer invasion and metastasis: molecular mechanisms, and clinical significance. Mol Cancer. 2024;23:18. doi:10.1186/s12943-024-01932-0
47. Yu F, Liang M, Huang Y, Wu W, Zheng B, Chen C. Hypoxic tumor-derived exosomal miR-31-5p promotes lung adenocarcinoma metastasis by negatively regulating SATB2-reversed EMT and activating MEK/ERK signaling. J Exp Clin Cancer Res. 2021;40:179. doi:10.1186/s13046-021-01979-7
48. Chen H, Liu L, Xing G, et al. Exosome tropism and various pathways in lung cancer metastasis. Front Immunol. 2025;16:1517495. doi:10.3389/fimmu.2025.1517495
49. He S, Li Z, Yu Y, et al. Exosomal miR-499a-5p promotes cell proliferation, migration and EMT via mTOR signaling pathway in lung adenocarcinoma. Exp Cell Res. 2019;379:203–213. doi:10.1016/j.yexcr.2019.03.035
50. Yin L, Liu X, Shao X, et al. The role of exosomes in lung cancer metastasis and clinical applications: an updated review. J Transl Med. 2021;19:312. doi:10.1186/s12967-021-02985-1
51. Tarin M, Oryani MA, Javid H, Karimi-Shahri M. Exosomal PD-L1 in non-small cell lung cancer: implications for immune evasion and resistance to immunotherapy. Int Immunopharmacol. 2025;155:114519. doi:10.1016/j.intimp.2025.114519
52. Hui E, Cheung J, Zhu J, et al. T cell costimulatory receptor CD28 is a primary target for PD-1-mediated inhibition. Science. 2017;355:1428–1433. doi:10.1126/science.aaf1292
53. Yokosuka T, Takamatsu M, Kobayashi-Imanishi W, Hashimoto-Tane A, Azuma M, Saito T. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J Exp Med. 2012;209:1201–1217. doi:10.1084/jem.20112741
54. Guo H, Zhang J, Qin C, Yan H, Luo X, Zhou H. Advances and challenges of first-line immunotherapy for non-small cell lung cancer: a review. Medicine. 2024;103:e36861. doi:10.1097/MD.0000000000036861
55. Hamid O, Robert C, Daud A, et al. Five-year survival outcomes for patients with advanced melanoma treated with pembrolizumab in KEYNOTE-001. Ann Oncol. 2019;30:582–588. doi:10.1093/annonc/mdz011
56. Kim DH, Kim H, Choi YJ, et al. Exosomal PD-L1 promotes tumor growth through immune escape in non-small cell lung cancer. Exp Mol Med. 2019;51:1–13.
57. Liu J, Peng X, Yang S, et al. Extracellular vesicle PD-L1 in reshaping tumor immune microenvironment: biological function and potential therapy strategies. Cell Commun Signal. 2022;20:14. doi:10.1186/s12964-021-00816-w
58. Tong S, Hua S, Wu Y, et al. miR-96-5p-mediated inhibition of CD47 contributes to pancreatic tumor regression via activating both innate and adaptive anti-tumor immunity. Cell Commun Signal. 2025;24:16. doi:10.1186/s12964-025-02582-5
59. Huang Q, Li Y, Huang Y, et al. Advances in molecular pathology and therapy of non-small cell lung cancer. Signal Transduct Target Ther. 2025;10:186. doi:10.1038/s41392-025-02243-6
60. Havryliuk V, Wojtowicz K, Gagat M, Żuryń A. Exosome-mediated mechanisms of drug resistance in lung cancer: molecular mechanisms and therapeutic strategies. Cell Physiol Biochem. 2025;59:358–374. doi:10.33594/000000779
61. Garg H, Suri P, Gupta JC, Talwar GP, Dubey S. Survivin: a unique target for tumor therapy. Cancer Cell Int. 2016;16:49. doi:10.1186/s12935-016-0326-1
62. Zhu Z, Wang CP, Zhang YF, Nie L. MicroRNA-100 resensitizes resistant chondrosarcoma cells to cisplatin through direct targeting of mTOR. Asian Pac J Cancer Prev. 2014;15:917–923. doi:10.7314/APJCP.2014.15.2.917
63. Zhang K, Chen J, Li C, et al. Exosome-mediated transfer of SNHG7 enhances docetaxel resistance in lung adenocarcinoma. Cancer Lett. 2022;526:142–154. doi:10.1016/j.canlet.2021.10.029
64. Lobb RJ, van Amerongen R, Wiegmans A, Ham S, Larsen JE, Möller A. Exosomes derived from mesenchymal non-small cell lung cancer cells promote chemoresistance. Int J Cancer. 2017;141:614–620. doi:10.1002/ijc.30752
65. Mir R, Baba SK, Elfaki I, et al. Unlocking the secrets of extracellular vesicles: orchestrating tumor microenvironment dynamics in metastasis, drug resistance, and immune evasion. J Cancer. 2024;15:6383–6415. doi:10.7150/jca.98426
66. Gridelli C, Rossi A, Carbone DP, et al. Non-small-cell lung cancer. Nat Rev Dis Primers. 2015;1:15009. doi:10.1038/nrdp.2015.9
67. Panunzio A, Sartori P. Lung cancer and radiological imaging. Curr Radiopharm. 2020;13:238–242. doi:10.2174/1874471013666200523161849
68. Behrouzi R, Clipson A, Simpson KL, et al. Cell-free and extrachromosomal DNA profiling of small cell lung cancer. Trends Mol Med. 2025;31:64–78. doi:10.1016/j.molmed.2024.08.004
69. Ren F, Fei Q, Qiu K, Zhang Y, Zhang H, Sun L. Liquid biopsy techniques and lung cancer: diagnosis, monitoring and evaluation. J Exp Clin Cancer Res. 2024;43(1):96. doi:10.1186/s13046-024-03026-7
70. Yousefi M, Ghaffari P, Nosrati R, et al. Prognostic and therapeutic significance of circulating tumor cells in patients with lung cancer. Cellular Oncology. 2020;43(1):31–49. doi:10.1007/s13402-019-00470-y
71. Tanaka F, Yoneda K, Kondo N, et al. Circulating tumor cell as a diagnostic marker in primary lung cancer. Clin Cancer Res. 2009;15:6980–6986. doi:10.1158/1078-0432.CCR-09-1095
72. Krebs MG, Sloane R, Priest L, et al. Evaluation and prognostic significance of circulating tumor cells in patients with non-small-cell lung cancer. J Clin Oncol. 2011;29:1556–1563. doi:10.1200/JCO.2010.28.7045
73. Park Y, Kitahara T, Urita T, Yoshida Y, Kato R. Expected clinical applications of circulating tumor cells in breast cancer. World J Clin Oncol. 2011;2:303–310. doi:10.5306/wjco.v2.i8.303
74. Van der Auwera I, Peeters D, Benoy IH, et al. Circulating tumour cell detection: a direct comparison between the CellSearch system, the AdnaTest and CK-19/mammaglobin RT-PCR in patients with metastatic breast cancer. Br J Cancer. 2010;102:276–284. doi:10.1038/sj.bjc.6605472
75. Landon BV, Annapragada AV, Niknafs N, Velculescu VE, Anagnostou V. Liquid biopsies across the cancer care continuum. Nat Med. 2025;31:4006–4021. doi:10.1038/s41591-025-04093-9
76. Li C, Shao J, Li P, Feng J, Li J, Wang C. Circulating tumor DNA as liquid biopsy in lung cancer: biological characteristics and clinical integration. Cancer Lett. 2023;577:216365. doi:10.1016/j.canlet.2023.216365
77. Wen X, Pu H, Liu Q, Guo Z, Luo D. Circulating tumor DNA-A novel biomarker of tumor progression and its favorable detection techniques. Cancers. 2022;14(24):6025. doi:10.3390/cancers14246025
78. Normanno N, Morabito A, Rachiglio AM, et al. Circulating tumour DNA in early stage and locally advanced NSCLC: ready for clinical implementation? Nat Rev Clin Oncol. 2025;22(3):215–231. doi:10.1038/s41571-024-00985-w
79. Ernst SM, Aldea M, von der Thüsen JH, et al. Utilizing ctDNA to discover mechanisms of resistance to targeted therapies in patients with metastatic NSCLC: towards more informative trials. Nat Rev Clin Oncol. 2025;22:371–378. doi:10.1038/s41571-025-01011-3
80. Villaflor V, Won B, Nagy R, et al. Biopsy-free circulating tumor DNA assay identifies actionable mutations in lung cancer. Oncotarget. 2016;7(41):66880–66891. doi:10.18632/oncotarget.11801
81. Wang N, Zhang X, Wang F, et al. The diagnostic accuracy of liquid biopsy in EGFR-mutated NSCLC: a systematic review and meta-analysis of 40 studies. SLAS Technol. 2021;26(1):42–54. doi:10.1177/2472630320939565
82. Wu TH, Hsiue EH, Yang JC. Opportunities of circulating tumor DNA in lung cancer. Cancer Treat Rev. 2019;78:31–41. doi:10.1016/j.ctrv.2019.07.002
83. Wang W, Zheng Z, Lei J. CTC, ctDNA, and exosome in thyroid cancers: a review. Int J Mol Sci. 2023;24:13767.
84. Khan A, Raza F, He N. Nanoscale extracellular vesicle-enabled liquid biopsy: advances and challenges for lung cancer detection. Micromachines. 2024;15:1181.
85. Cazzoli R, Buttitta F, Di Nicola M, et al. microRNAs derived from circulating exosomes as noninvasive biomarkers for screening and diagnosing lung cancer. J Thorac Oncol. 2013;8(9):1156–1162. doi:10.1097/JTO.0b013e318299ac32
86. Zhou X, Wen W, Shan X, et al. A six-microRNA panel in plasma was identified as a potential biomarker for lung adenocarcinoma diagnosis. Oncotarget. 2017;8:6513–6525. doi:10.18632/oncotarget.14311
87. Jin X, Chen Y, Chen H, et al. Evaluation of tumor-derived exosomal miRNA as potential diagnostic biomarkers for early-stage non-small cell lung cancer using next-generation sequencing. Clin Cancer Res. 2017;23:5311–5319. doi:10.1158/1078-0432.CCR-17-0577
88. Chen M, Feng X, Liu C, et al. Diagnostic value of exosome-derived lncRNA PITPNA-AS1 in lung cancer. Front Immunol. 2025;16:1539557. doi:10.3389/fimmu.2025.1539557
89. Shi L, Li B, Zhang Y, et al. Exosomal lncRNA Mir100hg derived from cancer stem cells enhance glycolysis and promote metastasis of lung adenocarcinoma through mircroRNA-15a-5p/31-5p. Cell Commun Signal. 2023;21:248. doi:10.1186/s12964-023-01281-3
90. Liu C, Xiang X, Han S, et al. Blood-based liquid biopsy: insights into early detection and clinical management of lung cancer. Cancer Lett. 2022;524:91–102. doi:10.1016/j.canlet.2021.10.013
91. Huang J, Ding M, Lu Y, et al. MiR-1246b, a novel miRNA molecule of extracellular vesicles in bronchoalveolar lavage fluid, promotes nodule growth through FGF14 in patients with lung cancer. Cell Death Dis. 2023;14:789. doi:10.1038/s41419-023-06218-9
92. Hubers AJ, Prinsen CF, Sozzi G, Witte BI, Thunnissen E. Molecular sputum analysis for the diagnosis of lung cancer. Br J Cancer. 2013;109(3):530–537. doi:10.1038/bjc.2013.393
93. Kim JE, Eom JS, Kim WY, et al. Diagnostic value of microRNAs derived from exosomes in bronchoalveolar lavage fluid of early-stage lung adenocarcinoma: a pilot study. Thorac Cancer. 2018;9:911–915. doi:10.1111/1759-7714.12756
94. Kadota T, Fujita Y, Yoshioka Y, Araya J, Kuwano K, Ochiya T. Extracellular vesicles in chronic obstructive pulmonary disease. Int J Mol Sci. 2016;17(11):1801. doi:10.3390/ijms17111801
95. Pullan JE, Confeld MI, Osborn JK, Kim J, Sarkar K, Mallik S. Exosomes as drug carriers for cancer therapy. Mol Pharm. 2019;16(5):1789–1798. doi:10.1021/acs.molpharmaceut.9b00104
96. Yan L, Li J, Zhang C. The role of MSCs and CAR-MSCs in cellular immunotherapy. Cell Commun Signal. 2023;21(1):187. doi:10.1186/s12964-023-01191-4
97. Goberdhan DCI, Harris AL, Wilson C. Generating exosome subtypes: diverse membrane origins and mergers. Trends Cell Biol. 2026. doi:10.1016/j.tcb.2026.03.009
98. Catalano M, O’Driscoll L. Inhibiting extracellular vesicles formation and release: a review of EV inhibitors. J Extracell Vesicles. 2020;9(1):1703244. doi:10.1080/20013078.2019.1703244
99. Chen R, Qian Z, Xu X, et al. Exosomes-transmitted miR-7 reverses gefitinib resistance by targeting YAP in non-small-cell lung cancer. Pharmacol Res. 2021;165:105442. doi:10.1016/j.phrs.2021.105442
100. Irep N, Inci K, Tokgun PE, Tokgun O. Exosome inhibition improves response to first-line therapy in small cell lung cancer. J Cell Mol Med. 2024;28(4):e18138. doi:10.1111/jcmm.18138
101. Ren W, Hou J, Yang C, et al. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J Exp Clin Cancer Res. 2019;38(1):62. doi:10.1186/s13046-019-1027-0
102. Datta A, Kim H, Lal M, et al. Manumycin A suppresses exosome biogenesis and secretion via targeted inhibition of Ras/Raf/ERK1/2 signaling and hnRNP H1 in castration-resistant prostate cancer cells. Cancer Lett. 2017;408:73–81. doi:10.1016/j.canlet.2017.08.020
103. Menaceur C, Dusailly O, Gosselet F, Fenart L, Saint-Pol J. Vesicular trafficking, a mechanism controlled by cascade activation of rab proteins: focus on Rab27. Biology. 2023;12:1530.
104. Hu W, Xiong H, Ru Z, et al. Extracellular vesicles-released parathyroid hormone-related protein from Lewis lung carcinoma induces lipolysis and adipose tissue browning in cancer cachexia. Cell Death Dis. 2021;12(1):134. doi:10.1038/s41419-020-03382-0
105. Chen Y, Li J, Ma Y, et al. Recent advances in CAR-MSCs: the new engine of cellular immunotherapy evolution. J Hematol Oncol. 2025;18:92. doi:10.1186/s13045-025-01746-7
106. Xie H, Wang J. MicroRNA-320a-containing exosomes from human umbilical cord mesenchymal stem cells curtail proliferation and metastasis in lung cancer by binding to SOX4. J Recept Signal Transduction Res. 2022;42:268–278. doi:10.1080/10799893.2021.1918166
107. Liu J, Feng Y, Zeng X, He M, Gong Y, Liu Y. Extracellular vesicles-encapsulated let-7i shed from bone mesenchymal stem cells suppress lung cancer via KDM3A/DCLK1/FXYD3 axis. J Cell Mol Med. 2021;25:1911–1926. doi:10.1111/jcmm.15866
108. Dong L, Pu Y, Zhang L, et al. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles promote lung adenocarcinoma growth by transferring miR-410. Cell Death Dis. 2018;9:218. doi:10.1038/s41419-018-0323-5
109. Li S, Li W, Wu X, Zhang B, Liu L, Yin L. Immune cell-derived extracellular vesicles for precision therapy of inflammatory-related diseases. J Control Release. 2024;368:533–547. doi:10.1016/j.jconrel.2024.03.007
110. Kalluri R. The biology and function of extracellular vesicles in immune response and immunity. Immunity. 2024;57:1752–1768. doi:10.1016/j.immuni.2024.07.009
111. Buzas EI. The roles of extracellular vesicles in the immune system. Nat Rev Immunol. 2023;23(4):236–250. doi:10.1038/s41577-022-00763-8
112. Li Y, Yan B, He S. Advances and challenges in the treatment of lung cancer. Biomed Pharmacother. 2023;169:115891. doi:10.1016/j.biopha.2023.115891
113. Wathoni N, Puluhulawa LE, Joni IM, et al. Monoclonal antibody as a targeting mediator for nanoparticle targeted delivery system for lung cancer. Drug Deliv. 2022;29:2959–2970. doi:10.1080/10717544.2022.2120566
114. Wang S, Zimmermann S, Parikh K, Mansfield AS, Adjei AA. Current diagnosis and management of small-cell lung cancer. Mayo Clin Proc. 2019;94(8):1599–1622. doi:10.1016/j.mayocp.2019.01.034
115. Xu Y, Hsu JC, Xu L, Chen W, Cai W, Wang K. Nanomedicine-based adjuvant therapy: a promising solution for lung cancer. J Nanobiotechnol. 2023;21(1):211. doi:10.1186/s12951-023-01958-4
116. Cui Z, Ruan Z, Zeng J, et al. Lung-specific exosomes for co-delivery of CD47 blockade and cisplatin for the treatment of non-small cell lung cancer. Thorac Cancer. 2022;13:2723–2731. doi:10.1111/1759-7714.14606
117. Kim MS, Haney MJ, Zhao Y, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2016;12(3):655–664. doi:10.1016/j.nano.2015.10.012
118. Vinod SK, Hau E. Radiotherapy treatment for lung cancer: current status and future directions. Respirology. 2020;25 Suppl 2:61–71. doi:10.1111/resp.13870
119. Brown S, Banfill K, Aznar MC, Whitehurst P, Faivre Finn C. The evolving role of radiotherapy in non-small cell lung cancer. Br J Radiol. 2019;92(1104):20190524. doi:10.1259/bjr.20190524
120. Iyengar P, Wardak Z, Gerber DE, et al. Consolidative radiotherapy for limited metastatic non-small-cell lung cancer: a Phase 2 randomized clinical trial. JAMA Oncol. 2018;4:e173501. doi:10.1001/jamaoncol.2017.3501
121. Christodoulou M, Bayman N, McCloskey P, Rowbottom C, Faivre-Finn C. New radiotherapy approaches in locally advanced non-small cell lung cancer. Eur J Cancer. 2014;50(3):525–534. doi:10.1016/j.ejca.2013.11.027
122. Wu Y, Song Y, Wang R, Wang T. Molecular mechanisms of tumor resistance to radiotherapy. Mol Cancer. 2023;22(1):96. doi:10.1186/s12943-023-01801-2
123. Yuan D, Xu J, Wang J, et al. Extracellular miR-1246 promotes lung cancer cell proliferation and enhances radioresistance by directly targeting DR5. Oncotarget. 2016;7(22):32707–32722. doi:10.18632/oncotarget.9017
124. Lei QQ, Sui JD, Jin F, et al. Impact of high-dose rate radiotherapy on B and natural killer (NK) cell polarization in peripheral blood mononuclear cells (PBMCs) via inducing non-small cell lung cancer (NSCLC)-derived exosomes. Transl Cancer Res. 2021;10:3538–3547. doi:10.21037/tcr-21-536
125. Sun H, Zhu R, Guo X, et al. Exosome miR-101-3p derived from bone marrow mesenchymal stem cells promotes radiotherapy sensitivity in non-small cell lung cancer by regulating DNA damage repair and autophagy levels through EZH2. Pathol Res Pract. 2024;256:155271. doi:10.1016/j.prp.2024.155271
126. Ma X, Yao M, Gao Y, et al. Functional immune cell-derived exosomes engineered for the trilogy of radiotherapy sensitization. Adv Sci. 2022;9(23):e2106031. doi:10.1002/advs.202106031
127. Wang N, Song X. Research advances of immunotherapy of exosome PD-L1 in non-small cell lung cancer. Zhongguo Fei Ai Za Zhi. 2022;25:689–695. doi:10.3779/j.issn.1009-3419.2022.102.33
128. Kamimura N, Wolf AM, Iwai Y. Development of cancer immunotherapy targeting the PD-1 pathway. J Nippon Med Sch. 2019;86:10–14. doi:10.1272/jnms.JNMS.2019_86-2
129. Ricklefs FL, Alayo Q, Krenzlin H, et al. Immune evasion mediated by PD-L1 on glioblastoma-derived extracellular vesicles. Sci Adv. 2018;4:eaar2766. doi:10.1126/sciadv.aar2766
130. Besse B, Charrier M, Lapierre V, et al. Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. Oncoimmunology. 2016;5:e1071008. doi:10.1080/2162402X.2015.1071008
131. Shin S, Jung I, Jung D, et al. Novel antitumor therapeutic strategy using CD4(+) T cell-derived extracellular vesicles. Biomaterials. 2022;289:121765. doi:10.1016/j.biomaterials.2022.121765
132. Zhu T, Chen Z, Jiang G, Huang X. Sequential targeting hybrid nanovesicles composed of chimeric antigen receptor T-cell-derived exosomes and liposomes for enhanced cancer immunochemotherapy. ACS Nano. 2023;17:16770–16786. doi:10.1021/acsnano.3c03456
133. Guo J, Zhao W, Xiao X, et al. Reprogramming exosomes for immunity-remodeled photodynamic therapy against non-small cell lung cancer. Bioact Mater. 2024;39:206–223. doi:10.1016/j.bioactmat.2024.05.030
134. Tegegn TZ, De Paoli SH, Orecna M, et al. Characterization of procoagulant extracellular vesicles and platelet membrane disintegration in DMSO-cryopreserved platelets. J Extracell Vesicles. 2016;5:30422. doi:10.3402/jev.v5.30422
135. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
136. Zhang G, Huang X, Liu S, et al. Demystifying EV heterogeneity: emerging microfluidic technologies for isolation and multiplexed profiling of extracellular vesicles. Lab Chip. 2025;25:1228–1255. doi:10.1039/D4LC00777H
137. Tenchov R, Sasso JM, Wang X, Liaw WS, Chen CA, Zhou QA. Exosomes─nature’s lipid nanoparticles, a rising star in drug delivery and diagnostics. ACS Nano. 2022;16:17802–17846. doi:10.1021/acsnano.2c08774
138. Wan M, Ning B, Spiegel S, Lyon CJ, Hu TY. Tumor-derived exosomes (TDEs): how to avoid the sting in the tail. Med Res Rev. 2020;40:385–412. doi:10.1002/med.21623
139. Mizenko RR, Feaver M, Bozkurt BT, et al. A critical systematic review of extracellular vesicle clinical trials. J Extracell Vesicles. 2024;13:e12510. doi:10.1002/jev2.12510
140. Wen J, Chen Y, Liao C, et al. Engineered mesenchymal stem cell exosomes loaded with miR-34c-5p selectively promote eradication of acute myeloid leukemia stem cells. Cancer Lett. 2023;575:216407. doi:10.1016/j.canlet.2023.216407
141. Auquière M, Muccioli GG, Des Rieux A. Methods and challenges in purifying drug-loaded extracellular vesicles. J Extracell Vesicles. 2025;14:e70097. doi:10.1002/jev2.70097
142. Tiszbein K, Koss-Mikołajczyk I, Martysiak-żurowska D. Unlocking the secrets of human milk: isolation and characterization of extracellular vesicles. Adv Nutr. 2025;16:100430. doi:10.1016/j.advnut.2025.100430
143. Xylaki M, Chopra A, Weber S, Bartl M, Outeiro TF, Mollenhauer B. Extracellular vesicles for the diagnosis of Parkinson’s disease: systematic review and meta-analysis. Mov Disord. 2023;38:1585–1597.
144. Yuan F, Li YM, Wang Z. Preserving extracellular vesicles for biomedical applications: consideration of storage stability before and after isolation. Drug Deliv. 2021;28:1501–1509. doi:10.1080/10717544.2021.1951896
145. Jia Y, Yu L, Ma T, et al. Small extracellular vesicles isolation and separation: current techniques, pending questions and clinical applications. Theranostics. 2022;12:6548–6575. doi:10.7150/thno.74305
146. Wakayama S, Ito D, Kamada Y, et al. Tolerance of the freeze-dried mouse sperm nucleus to temperatures ranging from −196 °C to 150 °C. Sci Rep. 2019;9:5719. doi:10.1038/s41598-019-42062-8
147. Dave KM, Pinky PP, D SM. Molecular engineering of extracellular vesicles for drug delivery: strategies, challenges, and perspectives. J Control Release. 2025;386:114068. doi:10.1016/j.jconrel.2025.114068
148. Wu J, Jin Z, Fu T, et al. Extracellular vesicle-based drug delivery systems in cancer therapy. Int J Mol Sci. 2025;26:4835.
149. Luo X, McAndrews KM, Kalluri R. Natural and bioengineered extracellular vesicles in diagnosis, monitoring and treatment of cancer. ACS Nano. 2025;19:5871–5896. doi:10.1021/acsnano.4c11630
150. Wu X, Meng Y, Yao Z, et al. Extracellular vesicles as nature’s nano carriers in cancer therapy: insights toward preclinical studies and clinical applications. Pharmacol Res. 2025;217:107751. doi:10.1016/j.phrs.2025.107751
151. Fan L, Yao L, Li Z, et al. Exosome-based mitochondrial delivery of circRNA mSCAR alleviates sepsis by orchestrating macrophage activation. Adv Sci. 2023;10:e2205692. doi:10.1002/advs.202205692
152. Li D, Fang JJ, Jiang ZL. Extracellular vesicles in pancreatic cancer function and potential clinical applications. Onco Targets Ther. 2026;19:591622. doi:10.2147/OTT.S591622
153. van der Koog L, Gandek TB, Nagelkerke A. Liposomes and extracellular vesicles as drug delivery systems: a comparison of composition, pharmacokinetics, and functionalization. Adv Healthc Mater. 2022;11:e2100639.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Clinical Utility and Application of Liquid Biopsy Genotyping in Lung Cancer: A Comprehensive Review
Nigro MC, Marchese PV, Deiana C, Casadio C, Galvani L, Di Federico A, De Giglio A
Lung Cancer: Targets and Therapy 2023, 14:11-25
Published Date: 3 February 2023
The Cervical and Meningeal Lymphatic Network as a Pathway for Retrograde Nanoparticle Transport to the Brain
Ramos-Zaldívar HM, Polakovicova I, Salas-Huenuleo E, Yefi CP, Silva-Ancahuail D, Jara-Guajardo P, Oyarzún JE, Neira-Troncoso Á, Burgos PV, Cavieres VA, Arias-Muñoz E, Martínez C, Riveros AL, Corvalán AH, Kogan MJ, Andia ME
International Journal of Nanomedicine 2024, 19:10725-10743
Published Date: 24 October 2024
Drug Delivery System Targeting Cancer-Associated Fibroblast for Improving Immunotherapy
Zhang Z, Wang R, Chen L
International Journal of Nanomedicine 2025, 20:483-503
Published Date: 11 January 2025
Extracellular Vesicles for Disease Treatment

Wang F, Feng J, Jin A, Shao Y, Shen M, Ma J, Lei L, Liu L
International Journal of Nanomedicine 2025, 20:3303-3337
Published Date: 17 March 2025
Extracellular Vesicles in Pancreatic Cancer Function and Potential Clinical Applications
Li D, Fang JJ, Jiang ZL
OncoTargets and Therapy 2026, 19:591622
Published Date: 8 April 2026