Back to Journals » OncoTargets and Therapy » Volume 19
Extracellular Vesicles in Pancreatic Cancer Function and Potential Clinical Applications
Authors Li D, Fang JJ, Jiang ZL
Received 25 December 2025
Accepted for publication 30 March 2026
Published 8 April 2026 Volume 2026:19 591622
DOI https://doi.org/10.2147/OTT.S591622
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr John Riches
Dai Li,1,* Jian-jun Fang,2,* Zhi-Long Jiang1
1Department of General Surgery, Taixing People’s Hospital Affiliated to Yangzhou University, Taixing, Jiangsu, 225400, People’s Republic of China; 2Department of Medical Center of Hematology, Xinqiao Hospital of Army Medical University, Chongqing, 400037, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhi-Long Jiang, Email [email protected]
Abstract: Pancreatic cancer (PC) is one of the most aggressive gastrointestinal malignancies, characterized by a dismal 5-year survival rate. This poor prognosis is primarily attributed to delayed early detection, rapid disease progression, surgical complexities, and the limited efficacy of conventional oncological therapies. Extracellular vesicles (EVs) are nanoscale, cell-secreted vesicles that transport bioactive cargoes, including nucleic acids, proteins, and lipids. Upon release into the extracellular space, EVs facilitate short and long-distance intercellular communication and molecular transport via multiple pathways. In this review, we elucidate the multifaceted roles of EVs within the highly malignant PC microenvironment, specifically focusing on their mediation of intricate crosstalk between tumor and stromal cells. Furthermore, we summarize potential EV-based biomarkers for PC diagnosis and the recent advances in leveraging EVs as therapeutic platforms across radiotherapy, gene therapy, and immunotherapy. Ultimately, this review aims to provide novel insights into the clinical management of PC to improve patient outcomes and quality of life.
Keywords: pancreatic cancer, extracellular vesicles, nanomedicine, drug delivery, diagnosis
Introduction
PC is a highly malignant tumor of the digestive system, and epidemiological statistics have found that the 5-year survival rate of patients with PC is less than 9%.1 Point mutations in oncogenes kirsten rat sarcoma viral oncogene homolog (KRAS), tumor protein p53 (TP53), cyclin-dependent kinase inhibitor 2A (CDKN2A), and SMAD family member 4 (SMAD4), smoking, diabetes mellitus, obesity, and a history of chronic pancreatitis are the main risk factors for PC.2 The insidious origin, highly aggressive nature, and lack of concrete detection tools lead to late diagnosis and missed opportunities for surgical treatment.3 Currently, it is widely accepted that surgery is the only chance to cure PC, and the vast majority of patients have already developed lymph node and vascular infiltration and even metastases to the liver, lungs, and peritoneum by the time of definitive diagnosis.4 Most patients can only be treated with systemic palliative chemotherapy. Systemic palliative chemotherapy, ie, the use of drugs such as gemcitabine (GEM) and paclitaxel (PTX) to inhibit further tumorigenesis development, but the Treatment’s efficacy is minimal.5 Therefore, there is an urgent need to find diagnostic markers for the early diagnosis of PC that are reliable, as well as high-quality therapeutic strategies to improve the prospects for the grim survival of patients with PC.
EVs are membrane-enclosed, nanoscale vesicles originally considered cellular “waste products” intended to maintain intracellular homeostasis.6 Subsequent research has revealed that EVs are highly enriched in nucleic acids, proteins, and lipids, playing a pivotal role in intercellular communication by transferring these bioactive molecules and consequently altering the biological behavior of recipient cells.7 Both tumor cells and stromal cells within the tumor microenvironment (TME) abundantly secrete EVs. The resulting EV-mediated signaling crosstalk significantly promotes tumor invasion, metastasis, chemoresistance, and immunosuppression.8 In the present review, we focus on EV-mediated crosstalk within the PC microenvironment, highlighting their dual utility as diagnostic biomarkers and targeted delivery vehicles for therapeutics. Through this comprehensive synthesis, we aim to further enhance the understanding of the critical role EVs play in PC progression and clinical management.
General Introduction of EVs
EVs are heterogeneous collections of nanoscale vesicles secreted by cells into the extracellular space. Earlier, microvesicles, exosomes (Exos), and apoptotic vesicles were thought to be the leading entities of EVs.9 In recent years, several new subgroups of EVs have been identified, including secretory autophagosomes/amphibian, retractile vesicles, and migratory vesicles.10 EVs can be detected in various body fluids, including blood, cerebrospinal fluid, saliva, and sweat.11 Their heterogeneity is reflected in their particle size, shape, and charge, as well as the type and number of nucleic acid and lipid molecules they contain.12 Factors affecting the heterogeneity of EVs include but are not limited to, the type of parent cell, its health status, and the environment in which the cell is living.13 Release into the external environment can be achieved through receptor-ligand interactions, EVs interaction with plasma membranes (eg, the presence of a receptor), and the presence of a receptor. Interactions, a direct fusion of EVs with the plasma membrane (PM), and complete internalization of EVs by target cells through phagocytosis, lattice protein-mediated endocytosis, and megaloblast efflux.14 In recent years, based on its widespread presence in various body fluids and carrying a wide range of constituents, the EVs has been used as a liquid biopsy tool for diagnosing various diseases and as an essential indicator for prognostic analyses.14 Also, many studies have been conducted to modify EVs to make them ideal drug delivery vehicles for delivering therapeutic agents to malignant tumor cells, damaged cells, and immune cells15 (Figure 1).
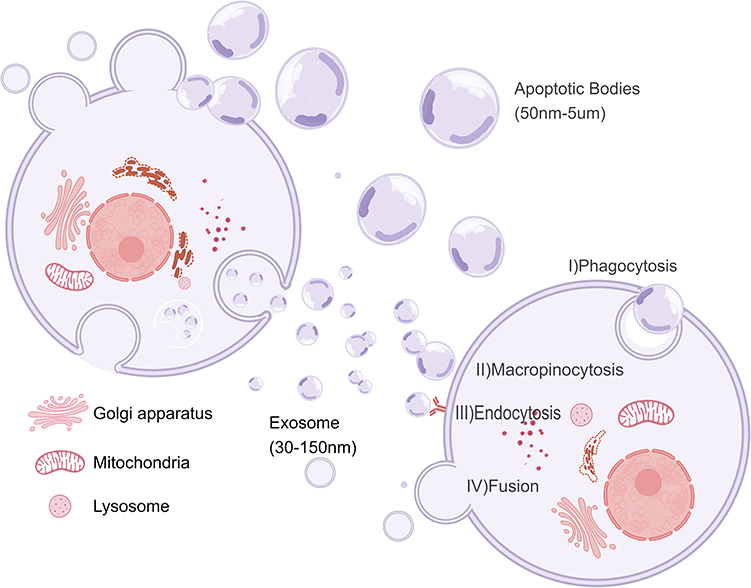 |
Figure 1 Biogenesis, secretion and cell entry of extracellular vesicles (EVs): EVs are categorized as exosomes, apoptotic bodies, and multivesiculars (MVs) based on their biological origin and size. Exosomes produce intraluminal vesicles (ILVs) through inward budding of early endosomes to form multivesicular Bodies (MVBs), which subsequently fuse with lysosomes or cell membranes and are released into the extracellular environment of the exosome, MVs are produced by outward budding from the plasma membrane, Apoptotic bodies are released during cell death through plasma membrane vesicles. EVs can deliver cargo to recipient cells through phagocytosis, megakaryocytosis, endocytosis, direct fusion, and other means, and subsequently modulate biological behavior. |
EVs Mediate Signaling Crosstalk Between Cells in the PC Microenvironment
The PC microenvironment is densely populated with diverse stromal components, including pancreatic stellate cells (PSCs), cancer-associated fibroblasts (CAFs), mesenchymal stem cells (MSCs), tumor-associated macrophages (TAMs), and the tumor cells themselves.16 These diverse cell populations secrete abundant EVs, utilizing them as crucial mediators for bidirectional intercellular communication. This complex interplay ultimately drives cancer progression, metastasis, therapeutic resistance, and immune evasion.17 Herein, we delineate the specific roles of EVs released by tumor and stromal cells, elucidating the intricate molecular interactions between PC cells and the stromal compartment within the TME.
Tumor Cells
Hypoxia, reduced extracellular pH, and elevated lactate concentrations are common features of the solid TMEs, which can be involved in cancer cell survival, metastasis, immune evasion, and chemoresistance by regulating the secretion of important chemokines and cytokines by tumor cells.18 In the PC microenvironment, the thickening of stromal fibrosis compresses tumor tissue, leading to the occlusion of lymphatic and vascular vessels within the tumor. The conflict between supply and demand caused by the high oxygen demand of rapid tumor growth and the limited oxygen supply from poorly opened blood vessels leads to prolonged, persistent, and severe hypoxia in many regions of PC tumor cells.19 In response to this hypoxic stress condition, PC tumor cells can construct a microenvironment conducive to their development by secreting EVs that act as inter-cellular crosstalk mediators.20 The migration and tube formation of human umbilical vein endothelial cells (HUVECs) are involved in tumor angiogenesis. Studies have found that PC secretes EVs, which induce the formation of PC angiogenesis through interactions with HUVECs. For example, EVs rich in miR-30b promote HUVEC migration and tube formation by inhibiting GJA1.21 Secretion of vesicles containing the LncRNA UCA1 promotes angiogenesis by modulating the miR-96-5p/AMOTL2/ERK1/2 signalling pathway in HUVECs.22 Whereas to promote tumor invasion and metastasis, secreted EVs containing a high expression of circ-IARS into HUVECs cause endothelial barrier dysfunction and enhance endothelial monolayer permeability by increasing RhoA expression and activity, increasing the level of F-actin expression, and decreasing the expression of tightly linked zonula occludens-1 (ZO-1), causing distant PC metastasis.23 Concomitant secretion of FGD5-AS1-containing EVs leads to the acetylation of signal transducer and activator of transcription factor 3 (STAT3) by interacting with p300, which promotes the nuclear localization and transcriptional activity of STAT3/NF-κB. The over-activation of the STAT3/NF-κB pathway leads to the development of an M2-type polarisation of TAMs, forming an inhibitory tumor microenvironment and increasing cancer cell proliferation and metastasis.24 On the other hand, tumor-immune tumor-secreted EVs are also involved in tumor drug resistance, and relevant studies have confirmed that PC-secreted EVs cause PC cell resistance to the standard chemotherapeutic agent GEM via transfer matrix metalloproteinase 14 (MMP14), miR-155, and hepatic ligand protein type A receptor 2 (EphA2)25 (Figure 2).
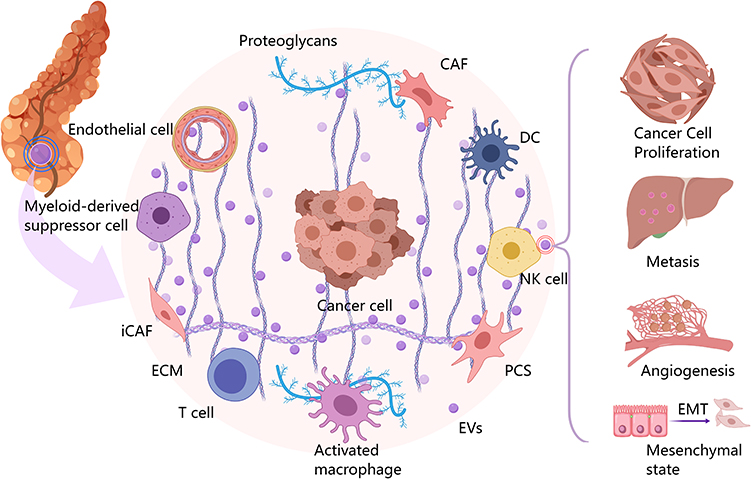 |
Figure 2 Pancreatic cancer (PC) microenvironment is composed of pancreatic tumor cells, pancreatic stellate cells, cancer fibroblasts, tumor-associated macrophages mesenchymal stem cells, and dendritic cells, all of which can realize information communication between cells in the microenvironment by secreting extracellular vesicles, forming a pro-tumor microenvironment, which ultimately leads to the rapid proliferation of tumor cells, invasion of distant and near organs, formation of neovascularization, and epithelial-mesenchymal transformation. |
PSCs
The physiological functions of PSCs under normal conditions include participation in the body’s immune response, phagocytosis, and amylase secretion.26 In the microenvironment of PC, PSCs can be further categorized into two main groups: quiescent PSCs and activated pancreatic stellate cells (aPSCs).27 The differences in the vitamin A content and the protein profile expression can be an essential indicator of the distinction between these cells. In resting PSCs, vitamin A droplets are stored in the cytoplasm and express nestin, vimentin and junctional proteins. In contrast, aPSCs are activated to a myofibroblast-like phenotype, resulting in a loss of vitamin A droplet storage and high expression of α-smooth muscle actin (α-SMA).28 In addition to the differences in constituents, the biological behaviors of the two types of cells also undergo significant changes. Cells also underwent significant changes in biological behavior, with aPSC mediating their crosstalk with pancreatic tumor cells and thus regulating the biological behavior of PC cells through the secretion of soluble factors (eg, cytokines, chemokines, growth factors) and EVs enriched in microRNAs (miRNAs), lncRNAs, proteins, mRNAs, lipids and metabolites. Here, we focus on reports of such crosstalk activities mediated by EVs secreted by aPSCs. miRNAs are small, constitutive, non-coding RNAs of approximately 21 nucleotides in length that control the activity of approximately 30% of protein-coding genes in mammals, specifically by binding to the 3ʹUTR of the mRNA of a target gene through translational repression or shear regulation of target gene expression by binding to the 3ʹUTR of the target gene mRNA target gene mRNA to regulate its expression through translational repression or cleavage. Moreover, the EVs secreted by PSCs and the microRNAs of PSCs themselves are heterogeneous. It has been found that 251 microRNAs, such as miR-1246, miR-21-5p and miR-1290, are enriched in EVs, while miR-199a-5p and miR-1260b, which are highly expressed in PSCs, are depleted in EVs.29 At the same time, different components of miRNAs in EVs can promote PC tumor cell proliferation, migration, and chemotherapy resistance by regulating the expression of crucial pathways.30 Compared to free miRNA-mediated crosstalk activities, miRNAs encapsulated in EVs phospholipid bilayers miRNAs can be protected from external environmental interference to a certain extent or degradation.31 It was found that miR-5703-containing EVs secreted by aPSC directly bind to and downregulate the 3ʹUTR of CMTM4, the subsequently downregulated CMTM4 promotes the proliferation of PC cells by inhibiting the activation of the activated kinase 4 (PAK4)-mediated PI3K/Akt pathway.32 Whereas, in the hypoxia condition, the miR-4465 and miR-616-3p-containing EVs secreted by aPSCs decreased the expression of PTEN and increased the expression of p-AKT by binding to the PTEN mRNA of the tumor cells, thereby promoting the proliferation, migration, and invasion of the cancer cells.33 In addition to the contained miRNA involved in the regulation of the malignant biological behaviour of PC cells, Liu et al reported that hypoxia stimulation can increase the secretion of EVs rich in LncRNA UCA1 by aPSC, which, after being taken up by PC cells, can reduce the histone methylation level in the SOCS3 gene region by recruiting EZH2, thereby enhancing PC’s resistance to GEM and inhibiting cancer cell apoptosis.34
Cancer Fibroblasts (CAFs)
CAFs make up the majority of the main body of the tumor stroma in PC, and their precursor cells of origin include, but are not limited to, quiescent fibroblasts, PSCs, MSCs, adipocytes, and others.35 The diverse origins have led to the heterogeneous CAFs in the microenvironment. Single-cell transcriptomics techniques have revealed that CAFs in the PC microenvironment can be divided into two subpopulations: myofibroblast CAFs (myCAFs) and inflammatory CAFs (iCAFs).36 Among them, myCAFs are spatially located adjacent to tumor cells, highly express αSMA, and cannot release inflammatory cytokines. In contrast, iCAFs are spatially located farther from tumor cells, low in αSMA expression, and release more inflammatory factors such as IL-6, IL-8, and IL-11.37 According to their roles in tumor development, they can be further classified into two subgroups: pro-cancer CAFs (pCAFs) and cancer-suppressor CAFs, and the majority of CAFs subgroups in the microenvironment usually exhibit pro-cancer effects.35 The cancer-suppressor CAFs subgroup releases a variety of mediators that can be used to inhibit cancer development. However, the pCAFs subpopulation releases a variety of mediators (eg, TGF-β, IL-6, and CXCL12) associated with rapid proliferation, neoangiogenesis, chemotherapy resistance, and the formation of a suppressive tumor microenvironment in PC.37 EVs, an essential medium for exchanging materials and information between cells, have been shown to benefit PC by directly or indirectly interacting with tumor cells or various subpopulations of immune cells in the microenvironment. Tumor cells have a metabolic symbiotic relationship with stromal cells in the microenvironment. Various metabolic waste intermediates of CAFs, such as lactate, acetate, amino acids, etc, can be effectively taken up by tumor cells through EVs transfer and used in central carbon metabolism to maintain the rapid proliferation of PC tumor cells even in the extreme lack of nutrients.38 The treatment of PC after surgery supplemented with chemotherapeutic agents is the gold standard of PC treatment. After a period of treatment, patients usually show drug resistance, significantly limiting the prognosis of the patients.39 Studies have found that treatment with GEM can increase the overall number of EVs secreted by CAFs to a certain degree and induce the secretion of EVs containing specific cargoes. For example, the study by Shi et al found that GEM-treated CAFs secreted highly expressed EVs containing miR-3173-5p, which significantly reduced Fe2+ and lipid ROS levels in tumor cells after uptake by pancreatic tumor cells following GEM treatment, causing increased resistance to ferroptosis in tumor cells, which in turn induced tumors to become resistant to GEM.40 In addition, the release of miR-21, miR-181a, miR-221, miR-222 and miR-92a have also been shown to promote tumor cell proliferation and chemotherapy resistance by inhibiting the tumor suppressor PTEN.41
MSCs
MSCs are pluripotent adult stem cells that can be derived from most tissues of the body, including but not limited to bone marrow (BM), adipose tissue (AD), umbilical cord (UC), and placenta.42 Properties such as self-renewal capacity, pluripotency, and immunomodulation have made MSCs an attractive tool for regenerative medicine.43 In a tumor context, MSCs are an essential component of the microenvironmental stromal cells and are closely associated with tumor progression. As early as the initiation of PC, MSCs can be recruited from the periphery to the vicinity of the tumor cells and participate in the formation of the “pre-metastatic ecological niche”.44 MSCs can be reprogrammed from normal trophic to tumor-promoting cells based on the membrane protein recognition system or the uptake of soluble factors released by the PC cells, such as chemokines, cytokines, and cargo-containing EVs, anchoring tumor cells as tumor stroma, secreting various mediators to enable crosstalk of material information with other cells in the microenvironment.45 As avid producers of EVs, MSCs secrete a considerable amount of EVs, and a study comparing the human acute monocytic leukemia cell line, human embryonic kidney (HEK) cell line and MSCs under equivalent treatment conditions found that MSCs produced 81 times the amount of EVs as compared to the other cell lines.46 Treatments such as hypoxia, C-MYC gene transfection, hydrogen peroxide, and lipopolysaccharide (LPS) further enhanced the amount of EVs secreted by MSCs.47 Here, we focus on the exchange of material and information in the PC microenvironment mediated by MSCs through the release of EVs. The Wnt signaling pathway, as a critical signaling pathway controlling the malignant progression of PC, is controlled upstream by the oncogene WIF1 and downstream of which regulates a variety of stemness-related genes in PC cells, such as Oct4, Nanog, and Sox2, etc. A study by Lu et al found that MSCs-derived EV containing circ_0030167 could sponge miR-338-5p and target the Wif1/Wnt8/β-catenin axis after uptake by PCs to achieve proliferation, invasion, and stemness of PC.48 In addition, a study by Li et al found that human umbilical cord blood-derived mesenchymal stem cell-secreted EVs could enhance the proliferation and migration of PDAC cells by transferring miR-100-5p to Panc-1 and BxPC3 cells.49
TAMs
TAMs, as the most abundant immune cells in the microenvironmental stroma of PC, account for approximately 15–20% of the total mass of tumor cells.50 In a tumor context, TAMs are usually activated into two distinct subpopulations, ie, classically activated macrophages (M1) and alternatively activated macrophages (M2), with the M1 macrophages having pro-inflammatory properties, whereas M2 macrophages express high levels of anti-inflammatory cytokines and a certain level of arginase-1 (Arg-1) activity, possessing pro-carcinogenic properties.51 EVs of tumor cells, CAFs, and other origins can activate macrophages through various mechanisms. For example, EVs containing miR-301a-3p and miR-320a secreted by PC cells and CAFs under hypoxic conditions promote M2-type polarisation through activation of the PTEN/PI3K-γ pathway.52,53 The release of EVs containing the lnc-FGD5-AS1 component a molecular bridge between p300 and STAT3, causing acetylation of STAT3 and enhanced transcriptional activity, followed by activation of the STAT3/NF-κB signaling pathway and M2 polarisation of macrophages.24 Tang et al also demonstrated that the release of CEACAM5-containing proteins from PC induced M2-type polarisation of TAMs through activation of the MAPK signaling pathway and increased TAMs secretion of matrix metalloproteinase-9 (MMP-9), which in turn enhances PC metastasis.54 In turn, polarised M2-type macrophages release EVs to participate in remodeling the tumor microenvironment to make it more suitable for tumor cell survival. The M2 type macrophages release EVs enriched in many types of miRNAs, and in the microenvironment, EVs can be used as a carrier for miRNA delivery to achieve intercellular information exchange.55 EVs containing miR-155-5p and miR-221-5p from M2 macrophages have been reported to promote PC angiogenesis by inhibiting the critical angiogenic transcription factor E2F2 after being transported to vascular endothelial cells.56 The expression level of miR-21-5p, an oncogene in various solid tumors, is highly correlated with the overall survival of patients. A study found that miR-21-5p was also highly expressed in EVs secreted by M2 macrophages, which could be involved in the regulation of PC stem cell differentiation and activity by targeting krüppel-like factor 3 (KLF3).57 In addition, Sheng et al found that EVs secreted by M2 macrophages containing miR-501-3p were highly expressed in PC stem cells and that the expression of miR-21-5p in EVs secreted by M2 macrophages could promote PC angiogenesis.58 miR-501-3p-containing EVs secreted by M2-type macrophages promoted migration, invasion, and tube formation of PC cells through down-regulation of Transforming growth factor β receptor 3 (TGFBR3).59 In addition to M2-type macrophage-derived EVs, Xia et al reported by single-cell sequencing coupled with spatial transcriptomics that RPS18-containing EVs of M1 macrophage origin were transferred to PC cells and resulted in PC cell activity, proliferation, migration and invasion to be inhibited.60
Dendritic Cells (DCs)
DCs, as antigen-presenting cells (APCs) at the core of the host adaptive immune response, are usually found in the blood and lymph nodes in three subpopulations: plasma cell-like DCs (pDCs), conventional type 1 DCs (cDC1s), and conventional type 2 DCs (cDC2s). pDCs exhibit a plasmacytoid morphology and produce large quantities of interferon I (IFN-α/β) after capturing viruses.61 cDC1 is a rare subset of DCs activated to produce a variety of inflammatory cytokines, including TNF-α, IL-6, IL-8, and IL-12, and cDC2, as a significant subset population of DCs, is the most potent APC for the activation and amplification of CD4 T-cells.62 In a tumor context, PC cells release a variety of EVs. Restricting peripheral DCs migration towards tumor cells and inhibiting microenvironmental DCs activation, antigen processing, and presentation functions cause defective T-cell initiation or inappropriate tolerance induction, ultimately leading to tumor cell immune escape.63 When PC cell-derived EVs were incubated with DCs, 3227 lncRNAs and 924 mRNAs were identified as differentially expressed in DCs, and many of these RNAs were involved in the regulation of key mutated genes in PC networks such as TP53, KRAS, SMAD4 and CDKN2A.64 In addition, Cao et al found that when miR-203-containing EVs were incubated with DCs, the release of tumor necrosis factor-α (TNF-α) and interleukin-12 (IL-12) from DCs was significantly reduced, the mechanism suggests that miR-203, which is carried by EVs after uptake by DCs, restricts NF-κB activation by targeting and inhibiting TLR4, whereas miR-212-3p, which is transferred to DCs, inhibits the expression of regulatory factor X associated protein (RFXAP), reduces the expression of major histocompatibility complex II (MHC II) and induces immune tolerance in DCs65 (Figure 3).
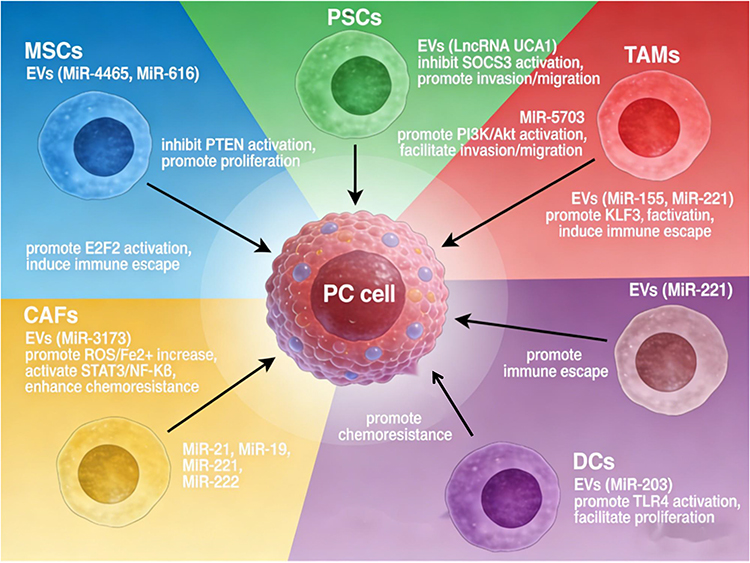 |
Figure 3 Chematic diagram of tumor microenvironment-derived extracellular vesicles (EVs) regulating pancreatic cancer progression: Various stromal cells, including mesenchymal stem cells (MSCs), pancreatic stellate cells (PSCs), tumor-associated macrophages (TAMs), cancer-associated fibroblasts (CAFs), and dendritic cells (DCs), secrete EVs carrying specific non-coding RNAs. These EVs deliver their cargo to pancreatic cancer cells, modulating key signaling pathways and leading to distinct malignant phenotypes. MSCs-derived EVs containing miR-4465 and miR-616 inhibit PTEN, promoting proliferation. DCs-derived EVs carrying miR-203 activate TLR4, also enhancing proliferation. PSCs-derived EVs deliver LncRNA UCA1 to suppress SOCS3, or miR-5703 to activate PI3K/Akt, both facilitating invasion and migration. TAMs-derived EVs loaded with miR-155/miR-221, miR-221, or miR-301a/miR-320a activate E2F2, KLF3, or directly induce immune escape. CAFs-derived EVs containing miR-3173 elevate ROS/Fe2⁺ levels and subsequently activate STAT3/NF-κB, while those carrying miR-21, miR-19, miR-221, and miR-222 contribute to drug resistance. These coordinated events collectively drive pancreatic cancer progression. |
Clinical Applications
EV-Based Diagnosis of PC
The high lethality of PC is closely related to the difficulty of making a definitive diagnosis at an early stage. Anatomically, the pancreas is located in the retroperitoneum and has no specific clinical manifestations at the early stage of development.12 Currently, endoscopic ultrasound-guided fine-needle aspiration (EUS-FAN) is considered to be the most accurate means of clinical diagnosis of PC. However, the puncture is highly, procedure is technically demanding for operators, and is prone to damaging to the nearby organs, leading to the risk of hemorrhage and cancer metastasis. It is not commonly used for universal screening of PC because of the risk of hemorrhage and metastasis.66 Instead, CA19-9 is currently a common and FDA-approved biomarker for early diagnosis of PC by detecting its levels in the patient’s serum. However, approximately 14% of the general population does not express it, and 25% of patients do not show elevated levels of it, Furthermore, serum levels of CA19-9 are falsely elevated in the following pathological conditions (eg, benign tumors, inflammatory masses, and diabetes).66 Serum levels of CA19-9 are also elevated in the following pathological conditions (eg, benign tumors, inflammatory masses, and diabetes mellitus), leading to a low specificity of CA19-9 in the diagnosis of PC.1 Therefore, there is an urgent need to find more suitable markers to clarify the diagnosis of PC as early as possible and to intervene accordingly in order to achieve a better quality of life for the patients. EVs, as nano-sized particles, can be obtained from a variety of body fluids, such as blood, saliva, sweat, etc, and carry a variety of proteins, lipids, DNA, RNA, and other important information of the parent cell.67 These two characteristics of EVs offer the possibility of various disease diagnoses. In the next section, we will discuss the application of the various EV-based cargoes in the early diagnosis of PC.
miRNA
The dysregulated expression of tumor-suppressive and oncogenic miRNAs is involved in the regulation of PC progression.68 A comparative study of miRNAs expression profiles in PC and normal pancreatic tissues revealed that miRNAs including, but not limited to, miR-31, miR-146a, miR-194, miR-210, and miR-223 were significantly upregulated in patients, while miR-92, miR-132, and miR-148a were significantly downregulated in patients.69 These aberrantly expressed miRNAs are sorted into EVs as parent cells secrete EVs. Current studies have found that miRNA cargoes from parent cells are loaded into EVs through four primary pathways: the neutral sphingomyelinase 2-dependent pathway, the miRNA motif and sumoylated heterogeneous nuclear ribonucleoprotein-dependent pathway, the 3′ end of the miRNA sequence-dependent pathway, and the miRNA-induced silencing complex-associated pathway, miRNA cargoes are loaded into EVs through four pathways.70 Moreover, miRNA expression profiling of secreted EVs reveals that in small RNAs, the proportion of miRNAs in EVs is higher than that in their parental cells.71 miRNAs are less susceptible to hydrolysis by nucleases in the external environment than free miRNAs under the protection of the lipid bilayer, making it easier for them to carry out the corresponding biological functions and broadening the possibility of early diagnosis of PC based on liquid biopsy.72 In the hypoxic microenvironment, PC secretes a large amount of miR-301a-containing EVs, and miR-301a expression in isolated serum from PC patients was found to be significantly higher than that in healthy individuals. Furthermore, the elevated expression of miR-301a in circulating EVs was closely correlated with the depth of tumor infiltration and lymph node metastasis.52 Michael House et al reported that miR-10-10a in EVs was found to be a significant contributor to the development of PC in the human body, the 100% sensitivity and specificity of miR-10b and miR-30c in EVs distinguish PC from normal control plasma.73 In addition to circulating sources of EVs, another study showed a significant elevation of miR-21 and miR-155 in pancreatic fluid-sourced EVs from patients with PC, and the diagnostic sensitivity specificity, and accuracy of the combination of these two miRNAs for PC were 96%, 75% and 91% respectively.74
LncRNA
LncRNAs are a class of endogenous RNAs with more than 200 nucleotides that do not code for proteins.75 lncRNAs play an important role in regulating transcriptional, post-transcriptional, and epigenetic levels during PC development through interactions with DNA, RNA, and proteins.76 A study using RNA sequencing of PC patients found significant differences in 43 lncRNAs compared to normal controls without cancer, including an approximately 6-fold upregulation of plasmacytoma variant translocation 1 (PVT1), a lncRNA that enhances GEM resistance, and a 5-fold overexpression of Lnc00152, a Lnc that controls invasion and metastasis.77 LncHOTTIP has also been reported to be highly expressed in PC patients and plays a significant role in regulating the WNT pathway.78 As tumor cells secrete EVs, differentially expressed lncRNAs are sorted into these vesicles and subsequently released into the external environment to participate in intercellular communication. In contrast, detecting “differential” lncRNAs in pancreatic fluid or blood provides new opportunities for pancreatic diagnosis.79 The lncRNA XIST, ROR, LINC01268, LINC02802, AC124854.1, and AL132657.1 have been highly expressed in PC-derived EVs, Among them, lncRNA XIST promoted perineural infiltration of PC cells through the miR-211-5p/GDNF axis, and pre-treatment of tumor cells with GW4869, an inhibitor of neutral sphingomyelinase, significantly suppressed GDNF-mediated neurotrophic signaling to inhibit tumor invasion.80 Compared with the normal pancreatic cell line (CCC-HPE-2), linc-ROR expression was elevated in EVs released from established PC cell lines (PANC-1, AsPC-1, MIA-PACA-2, CFPAC-1, and BxPC-3). Adipocytes induced the overexpression of linc-ROR via activation of the HIF1α/ZEB1 signaling pathway and interleukin-1β (IL-1β), which induced adipocyte dedifferentiation and promoted PC tumor invasion.81 A recent study used quantitative real-time reverse transcription polymerase chain reaction to detect the expression levels of lncRNAs in plasma-derived EVs from PC patients, compared with healthy subjects, the expression levels of LINC01268, LINC02802, AC124854.1 and AL132657.1 were significantly increased by 73.63-fold, 60.27-fold, 11.25-fold and 34.63-fold, respectively. The area under the curve (AUC) value for the four lncRNAs combined in the diagnosis of PC was 0.8476, with a sensitivity of 72% and a specificity of 89%.82
Protein
In addition to miRNAs in EVs, lncRNAs have great potential for PC diagnosis, and the analysis of EVs proteins and proteomic profiles is a powerful tool for PC diagnosis.83 The proteomic screening of 13 human PC cell lines and 2 non-tumor cell line sources of EVs by liquid chromatography-mass spectrometry identified CLDN4, EPCAM, CD151, LGALS3BP, HIST2H2BE, and HIST2H2BF could be used as specific candidate biomarkers for PC diagnosis.84 Glypican1 (GPC1), which is anchored to the outer PM via a glycosyl-phosphatidylinositol bond, is another critical biomarker for the diagnosis of PC. Indicators of great potential, it has been reported that circulating GPC1+ EVs is only present in mutant Kras transcripts, and GPC1+ EVs is not detected in EVs derived from wild-type Kras mRNA.7 Kras mutations are considered to be the initiating event driving PC development. Kras oncogenic mutations can be observed in approximately 40% of low-grade intraepithelial neoplasia lesions PanIN1 mutations and mutated Kras can drive pancreatic carcinogenesis by coordinating with CDKN2A, TP53, BRCA2, MLL3, and SMAD4.85 Based on this finding; Jiao et al found that by analyzing plasma-derived EVs from healthy subjects, 16 patients with benign pancreatic disease and 27 patients with PC that compared to healthy controls (HC) and patients with benign pancreatic disease, blood-derived EVs from patients with early and advanced PC had high expression of GPC1. The expression of GPC1 in EVs had an AUC of 0.59 for PC diagnosis, with a sensitivity of 74% and a specificity of 44%.86 To improve the study’s diagnostic efficiency, another study utilized the EVs developed using immune lipid nanoparticle (ILN) biobricks to identify GPC1 mRNA and proteins in the blood. The GPC1 mRNA and Protein co-diagnostic system was developed using an ILN biochip for EVs, which enabled the capture of GPC1 mRNA and Protein from a single EVs, increasing the AUC to 96%.87 In addition, ALIX in plasma-derived EVs can also be used to diagnose PC. In patients with stage I/II and stage III/IV PC, the AUC value of ALIX is 0.768, which is slightly higher than that of CA199 (0.756). The AUC of the two combined for diagnosing PC is 0.91, with a sensitivity of 90.6% and a specificity of 83.9%.88 Although no specific EVs protein has yet been established as the gold standard for PC diagnosis, proteins carried by PC-derived EVs provide significant diagnostic value, and their combination with CA19-9 can markedly improve diagnostic efficiency (Table 1 and Figure 4).
 |
Table 1 Extracellular Vesicle-Mediated Diagnostic Applications for Pancreatic Cancer |
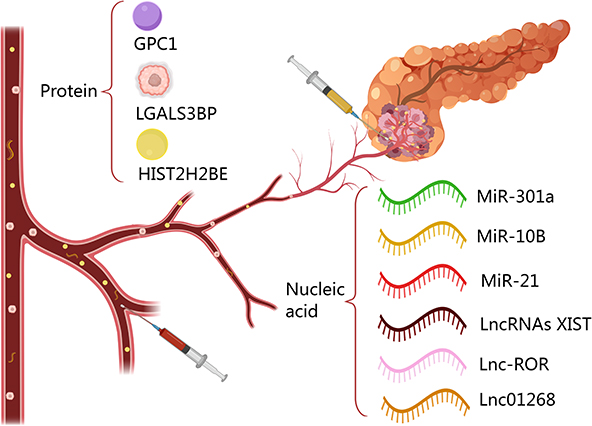 |
Figure 4 Proteins and nucleic acids carried in EVs in blood or pancreatic juice have been reported in the existing literature as biomarkers for the diagnosis of PC. |
Therapeutic Applications
Beyond their substantial potential in early diagnosis, EVs present novel avenues for PC therapeutic intervention. Current EV-based strategies encompass two primary approaches: (I) removing EVs containing nucleic acids or proteins related to the pathogenesis of the disease. Specifically, the secretion of EVs related to the progression of PC can be inhibited at the source by using sphingomyelinase inhibitors such as GW4869, or the effective binding of EVs to recipient cells can be inhibited, and circulating EVs that promote the progression of PC can be captured.89 (II) Using EVs as various therapeutic agents (small molecule chemotherapeutic drugs, miRNAs, EVs secretion or inhibit the effective binding of EV to receptor cells, and capture circulating EVs that promote PC progression).90 Next, we will further discuss the therapeutic application of EVs in PC in more detail.
Inhibit the Synthesis, Secretion, and Transport of Pro-Cancer EVs
The execution of EV-mediated intercellular communication necessitates several highly coordinated steps: biogenesis and secretion by the parent cell, release into the extracellular matrix, and subsequent uptake by recipient cells. Pharmacological blockade or inhibition of any of these stages can effectively disrupt EV-dependent signaling pathways.91 As previously described, within the highly heterogeneous PC microenvironment, multiple cell populations utilize diverse EVs cargoes to facilitate complex biomolecular crosstalk. This intricate signaling network ultimately accelerates tumor proliferation, distal metastasis, immune evasion, and the acquisition of chemoresistance.18 Consequently, exploring the application of pharmacological inhibitors targeting EV synthesis, secretion, or transport has emerged as a highly promising therapeutic frontier in PC management. Currently, the main inhibitors applied to limit EVs synthesis and secretion include GW4869 and Y27632, and compounds affecting EV transport include calreticulin, madecassicin A, and Y27632.92 GW4869 is a potent, specific, non-competitive inhibitor of membrane-associated neutral sphingomyelinase (nSMase). It inhibits the formation of the large lipid raft domain involved in EVs shedding by inhibiting the production of the bioactive lipid ceramide, thereby inhibiting EVs secretion.93 Y27632 is a potent, competitive inhibitor of Rho-associated protein kinase (ROCK) that inhibits EVs secretion through inhibition of reorganization of the cytoskeleton and actin filament-mediated contraction.94 Calcineurin is a calcineurin inhibitor, usually activated to inhibit G protein by binding to G protein. After activation, it can participate in the Regulation of cytoskeleton remodeling activity by binding to G and cytoskeletal proteins. In contrast, under the inhibitory effect of calpeptin, the remodeling activity of cytoskeletal proteins is reduced, and the secretion of EVs is significantly inhibited.95 In addition to the commonly used drugs, imatinib, glibenclamide, indomethacin, and chloroamidine have also been reported to modulate EVs synthesis and delivery.92 In PC, GW4869 has been more frequently used as a “harmful” EVs synthesis inhibitor to limit tumor development. For example, a study by Lei et al investigated the effect of GW4869 on EVs secreted by GEM-exposed PC CAFs and found that the addition of GW4869 could reduce the secretion of EV by 70%, which in turn could reduce the content of Snail and miR-146a in the microenvironmental EVs secreted by CAFs and inhibit the progression of tumor, while in vivo experiments found that the combination of GW4869 and GEM was more effective than PBS (control), treatment showed a significant reduction in growth rate compared to PBS and GEM group at 10 days after treatment.96 miR-29b-containing EVs secreted by PC cells protect HUVEC from PC cell-induced angiogenesis by attenuating the expression of ROBO1 and SRGAP2, and a study by Shi et al utilized the fact that GW4869 could suppress the miR-29b secreted by PC cells to prevent the progression of PC. A study by Shi et al using GW4869 to inhibit miR-29b-EV secreted by PC cells, co-cultured with GW4869 and tumor cells, found that GW4869 was involved in PC neoangiogenesis by controlling the expression of ROBO1 and SRGAP2.97 Similarly, another study also demonstrated that GW4869 could reverse EVs of PC cellular origin conferring resistance to GEM and cytokine TRAIL treatment.98
Treatment Mediated by EV as a Therapeutic Drug Delivery Vehicle
In recent years, EVs, initially regarded as cell-generated “waste products,” have been widely known as efficient drug delivery carriers due to their excellent biology, ease of artificial modification, low immunogenicity, and high-quality loading capacity.99 Relevant studies have reported that EVs can be used as vehicles for various therapeutic agents (eg, chemotherapeutic drugs, proteins, nucleic acids, and immunosuppressants) in the treatment of various malignant tumors. For example, EVs loaded with miR-451a human umbilical cord mesenchymal stem cells (hucMSCs) inhibit PTX resistance, cell cycle transition, proliferation, migration, and invasion and promote apoptosis of hepatocellular carcinoma cells (HCCs) by limiting epithelial-mesenchymal transition (EMT) of HCCs through the targeting of metalloproteinase 10 (ADAM10).100 Similarly, the study by Zhu et al found that EVs loaded with miR-129-5p can downregulate KCTD1 expression by targeting the 3′UTR region of the potassium channel tetramerization domain 1 (KCTD1), and further downregulate the HIF-1α/VEGF signaling pathway by inhibiting the interaction between KCTD1 and HIF-1α, ultimately suppressing the proliferation, migration, and metastasis of hepatocellular carcinoma.101 To overcome clinical bottlenecks such as insufficient targeting of small-molecule chemotherapy drugs and significant systemic toxicity, a recent study used electroporation technology to efficiently load Dox into extracellular vesicles derived from natural killer cells (NK-EVs). This delivery system significantly enhanced the uptake efficiency of Dox by liver cancer cells, effectively promoted tumor cell apoptosis, and demonstrated excellent anti-hepatocellular carcinoma efficacy both in vitro and in vivo. On the other hand, the antitumor active molecules carried by NK-EVs themselves can produce a synergistic killing effect with chemotherapy drugs, further inhibiting tumor proliferation and invasion, providing a new biomimetic delivery strategy for achieving precise and low-toxicity chemotherapy for HCC.102 In another study, exosomes derived from neutrophils (NEs-Exos) loaded with DOX a significant factor in treating malignancies. Neutrophil-derived Exos bind to integrin-related receptors on the blood-brain barrier (BBB), cross the BBB via, clathrin-mediated endocytosis and rapidly target C6-Luc gliomas under the chemotaxis of inflammatory factors, and fluorescence quantification revealed that the fluorescence intensity of brain tumors on day 15 in the NEs-Exos/DOX group was 2.74-fold lower than that of the free DOX group, and median survival increased to 27 days in the NEs-Exos/DOX group compared to only 22 days in the free DOX group.103,104 Here, we summarise the therapeutic applications of EVs in carrier-mediated PC over the last 5 years.
Chemotherapy
Postoperative adjuvant treatment with GEM, PTX, or doxorubicin (Dox) is the current conventional strategy for the treatment of PC.105 The primary mechanism of action of GEM and Dox is to inhibit the synthesis of nucleic acids in tumor cells, leading to the death of the tumor cells. In contrast, the primary mechanism of action of PTX is to block the mitotic division of tumor cells to exert an Anti-tumor effect.106 The long-term use of the above chemotherapeutic drugs brings a wide range of toxic side effects, including (cardiac, renal, neurotoxicity, etc) as well as PC cells inherent extracellular matrix abnormality proliferation, resulting in free chemotherapeutic drugs on the Treatment of PC has always been ineffective.107,108 In recent years, it has been found that the use of EVs as a delivery vehicle can increase the ability of the chemotherapy drug. For example, a study found that GEM-loaded Panc-1 EVs were more effective and safe than free GEM for PC treatment. The survival of the GEM EVs treatment group was 5 days longer than that of the free GEM treatment group. The safety assessment found that blood urea nitrogen (BUN) and alanine aminotransferase (ALT) in the GEM EVs treatment group were at normal levels, whereas these indicators were significantly elevated in the free GEM-treated group, indicating systemic toxicity.109 In addition, another study found that PC cell-derived EVs could be loaded with 14.06 ng/μg of DOX and that Treatment of PC cells for 48 hours with either 85.32nM DOX or different Exo-DOX preparations containing an equal amount of DOX resulted in an apoptotic index of tumor cells of up to 73.2%, which was significantly better than that of the free DOX-treated group.110 The current means of loading chemotherapeutic drugs into the EV generally includes two major categories, ie, EVs pre-separation loading and post-separation loading, in which pre-separation loading is the use of the mother cell co-incubated with the target drug and the drug enters into the cytoplasm of the mother cell under the action of concentration gradient. Then, the targeted drug is encapsulated into EVs during their synthesis and secretion.111 Post-isolation loading is operated at the level of the acquired EVs, including co-incubation, electroporation, sonication, and other technical means, wherein external forces induce electroporation or sonication to create pores in the EVs phospholipid bilayer membrane. Then, the target drug enters the EVs through the pores to complete loading.112,113
Gene Therapy
Dysregulation of the balance between oncogenic and tumor-suppressive non-coding RNA (ncRNA) expression is common in PC. On the one hand, overexpression of oncogenic ncRNAs inhibits the expression of oncogenic proteins, and on the other hand, overexpression of oncogenic ncRNAs leads to the overexpression of oncogenic proteins.114 Over the past two decades, the therapeutic use of small interfering RNAs (siRNA), miRNA mimics, and anti-miRNAs has made considerable achievements in treating PC based on the above theoretical foundations. Substances and anti-miRNAs have made considerable achievements in the Treatment of the pancreas. The specific mechanism of action is as follows: siRNA binds to cytoplasmic argonaute protein to form RISC, which induces translational inhibition of oncogenic mRNAs or mRNA decay; anti-miRNA binds to the untranslated region of oncogenic miRNAs or complementary target mRNAs and induces translation of the mRNAs into tumor suppressor proteins.115,116 miRNA mimics act as tumor-suppressor miRNAs, also known as tumor-suppressor mimics. Mimics act as substitutes for the absence of tumor suppressor miRNAs and can effectively inhibit the translation of oncogenes.117 The presence of multiple nuclease enzymes in the circulation and the extensive clearance of naked siRNAs and miRNAs by the liver and kidneys results in limited actual transfection of therapeutic miRNAs and siRNAs into tumor cells, severely diminishing the therapeutic efficacy.118 Therefore, there is an urgent need to develop high-quality miRNA and siRNA delivery platforms to improve term therapeutic efficacy. Liposomes, micelles, and polymer nanoparticles have been successively reported as carriers to deliver therapeutic miRNAs and siRNAs into PC.119–121 while potential toxic effects, difficulty in effective removal of carriers, and expensive cost of synthesis limit their further clinical translation.122 EVs, as cellular the good tolerance, low immunogenicity, and high loading capacity of EVs as an endogenous inter-cellular communication medium make them ideal vectors for the delivery of therapeutic miRNAs and siRNAs to pancreatic tumor cells.123 An innovative study by Xu et al successfully used electroporation to load siPAK4 into PANC-1 cell-derived EVs for PC treatment. In vivo and in vitro assays showed that the expression of PAK4 in the Exo-siPAK4 group was significantly lower than that in the Lipo-siPAK4 group (p < 0.001) and significantly increased the survival rate of the mice.124 In contrast, another study also loaded galectin-9 (gal-9) siRNAs into the PC cells by electroporation, making it challenging to apply them in PC treatment. In another study, gal-9 siRNA was loaded into bone marrow mesenchymal stem cell (BM-MSC)-derived EVs using an electroporation technique for treating PC. After administration, gal-9 siRNA EVs blocked the galectin-9/dectin-1 axis by lowering the gal-9 protein levels to reverse the profound immunosuppression induced by M2-TAMs and exert a therapeutic effect.125
Immunotherapy
Immunotherapy for PC exploits the host’s immune system to differentiate and eradicate malignant cells.50 Immunotherapy aims to increase the activity of intracellular cytotoxic T lymphocytes (CTLs) and to assist in initiating tumor-specific CTLs in lymphoid organs to build effective and long-lasting anticancer immunity.126 However, this is hampered by the immunosuppressive tumor microenvironment, the complex interplay of physiological and pathological barriers of complex interactions, etc, resulting in state-of-the-art immune checkpoint inhibitors currently having only a 30% success rate in present PC interventions, which highlights the importance of targeting the delivery of immune agents into the tumor microenvironment using the use of advanced drug delivery platforms.127 EVs mediate important cellular communication during tumor immunoediting. In the elimination phase, tumor-derived EVs containing contain immunomodulatory proteins, costimulatory molecules, and adhesion molecules that activate the immune system, immune cells secrete MHC-I, MHC-II, and CD86 components that can promote the activation of CD4+ and CD8+ T-cells.128 Although EVs hold promise for Immunotherapy, it has been found that EVs leaked as a therapeutic agent are readily eliminated from the body and have a very short circulating half-life.129 More studies of immunotherapy-engineered EVs are combining artificial modifications with Chemotherapy and gene therapy to improve therapeutic effects in PC further. For example, in a study by Chen et al gal-9 siRNA and oxaliplatin (OXA) were loaded into bone marrow MSCs via electroporation in combination for PC treatment. After intravenous injection, gal-9 siRNA down-regulated target genes to inhibit M2 polarisation effectively, OXA was released by the apoptotic cells through the induction of immunogenic cell death (ICD) in tumor cells. High mobility group box 1 (HMGB1) and adenosine triphosphate (ATP) act as immunostimulatory signals to initiate DCs maturation and antigen presentation, activate anti-tumour immune responses, and promote the infiltration of CTLs in the TME.125 Similarly, Chen et al utilized the use of electroporation to load CCL22 siRNAs into MART-1 peptide-modified tumor cell exosomes (referred to as the spMEXO system), which impedes the CCR4/CCL22 axis between DCs and Tregs, thereby inhibiting Treg amplification and thus suppressing tumor development130 (Figure 5 and Table 2).
 |
Table 2 Therapeutic Applications of Pancreatic Cancer Mediated by Extracellular Vesicles |
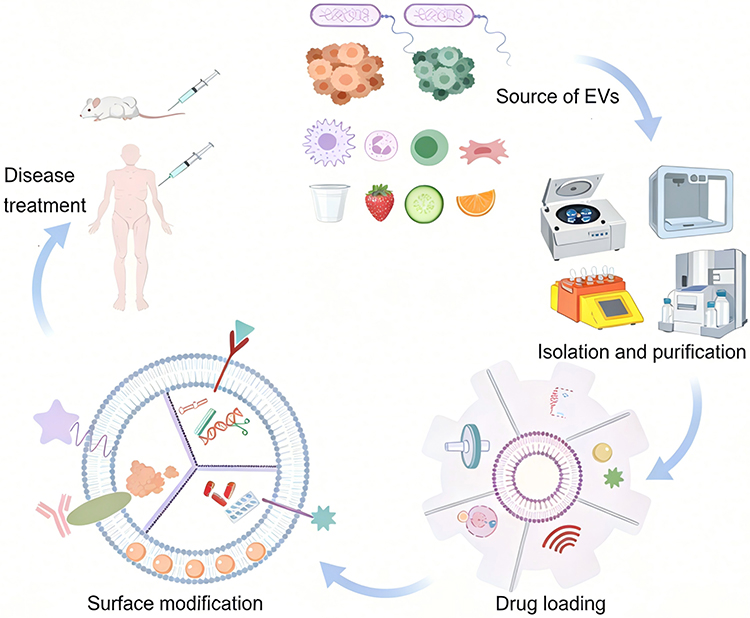 |
Figure 5 The workflow for the research and development of extracellular vesicle (EV)-based drug delivery systems and their therapeutic applications encompasses the multi-source preparation of EVs, efficient isolation and purification, targeted drug loading, and surface functional modification, ultimately realizing the full-chain technical route for disease treatment in animal models and preclinical settings. |
Current Limitations of Using EVs as Carriers
EVs making significant progress in PC targeted therapy due to their low immunogenicity, high drug-loading capacity, and excellent targeting ability, their clinical translation still faces a series of key bottlenecks. First, regarding the source of EVs carriers, although tumor-derived EVs are widely used as targeted drug delivery carriers because of their natural homing effect, increasing evidence shows that these vesicles carry pro-angiogenic and matrix remodeling factors such as VEGF and MMP-2/9, as well as oncogenic nucleic acids like miR-21 and miR-10b, posing potential tumorigenic risks.131 The long-term safety of using them as carriers for PC therapy still needs rigorous validation. Meanwhile, although EVs derived from plants, cow milk, and microbes have been preliminarily applied in PC research, their hepatotoxicity, nephrotoxicity, and cytotoxicity remain unclear, and head-to-head comparison data with human-derived EVs are lacking, seriously limiting their clinical advancement.132 Based on comprehensive safety assessment, EVs derived from MSCs and DCs remain ideal carriers with better translational prospects at the current stage. Secondly, large-scale production and standardized isolation of EVs remain core challenges. The currently mainstream separation strategies (differential centrifugation, density gradient centrifugation, size-exclusion chromatography, immunoaffinity enrichment, etc) all rely on physicochemical properties such as density, particle size, and surface markers, but they generally have defects such as structural damage, impurity contamination, insufficient purity, and poor reproducibility, and a standardized separation system has not yet been established.133 In addition, in the drug loading process, exogenous drug loading (electroporation, ultrasound, freeze-thaw) easily causes damage to the EVs membrane structure and leakage of contents; endogenous drug loading (co-incubation, transfection) has low drug loading efficiency and poor controllability. Both methods struggle to achieve efficient and stable drug encapsulation, which becomes an important limiting factor for clinical applications. In addition, the quality testing and quality control system for drug-loaded EVs is still in its early stages, lacking unified, standardized, and quantifiable evaluation criteria.134 Existing studies mostly rely on basic indicators such as particle size, concentration, morphology, surface markers, and simple drug loading rates, which are insufficient to comprehensively reflect the structural integrity, encapsulation stability, in vivo half-life, targeting efficiency, and biological activity of drug-loaded EVs.135 At the same time, there is a lack of functional validation systems specifically for the PC microenvironment, making it difficult to accurately predict their delivery efficiency, drug release behavior, and therapeutic effects in vivo. Furthermore, significant differences in detection methods, reagents, and equipment across different laboratories complicate data comparison and reproducibility, further hindering the standardized transition of EV-based drugs from basic research to clinical application.
Conclusions and Future Directions
As pivotal mediators of molecular exchange between tumor and stromal cells, EVs play a fundamental role in orchestrating PC invasion, metastasis, chemoresistance, and the immunosuppressive microenvironment. Notably, the isolation and detection of specific EV-encapsulated cargoes from peripheral blood or pancreatic fluid offer substantial diagnostic and prognostic value. Furthermore, EVs present transformative opportunities to enhance the precision of radiotherapy, chemotherapy, and gene therapy in PC.
Despite this profound potential, several critical hurdles must be prioritized to realize the clinical translation of EVs: (1) Technical Limitations in Isolation: Conventional isolation techniques—such as differential centrifugation, size-exclusion chromatography, and immunoaffinity capture—frequently suffer from compromised structural integrity, high operational costs, low purity, and insufficient yield. These constraints hinder the high-quality, large-scale isolation required for clinical applications, restricting current EVs platforms primarily to laboratory settings rather than commercial-grade manufacturing.136,137 (2) Standardization and Translation: Although relative to other synthetic carriers, EVs possess superior biocompatibility, standardizing their clinical use remains challenging.138,139 To overcome these bottlenecks, we propose that fostering robust multidisciplinary collaboration—encompassing materials science, clinical medicine, cell biology, and computational science—will expedite the resolution of current technical hurdles and catalyze the clinical translation of EV-based strategies. In summary, despite existing obstacles, EVs represent a transformative frontier with immense potential for the advanced diagnosis and therapeutic management of PC.
Abbreviations
PC, Pancreatic cancer; EVs, Extracellular vesicles; GEM, Gemcitabine; PTX, Paclitaxel; TMEs, Tumor microenvironments; Exos, Exosomes; HUVEC, Human umbilical vein endothelial cell; PSCs, Pancreatic stellate cells; aPSCs, Activated pancreatic stellate cells; α-SMA, α-smooth muscle actin; miRNAs, MicroRNAs; CAFs, Cancer-associated fibroblasts; MSCs, Mesenchymal stem cells; LPS, Lipopolysaccharide; MHC II, Major histocompatibility complex II; TAMs, Tumor-associated macrophages; GPC1, Glypican1; PM, Plasma membrane; DCs, Dendritic cells; HCCs, Hepatocellular carcinoma cells; AUC, Area under the curve; ILN, Immune lipid nanoparticle; BBB, Blood-brain barrier; Dox, Doxorubicin; SiRNA, Small interfering RNA; NcRNA, Non-coding RNA; CTLs, Cytotoxic T lymphocytes; OXA, Oxaliplatin; PAK4, P21-activated kinase 4; BUN, Blood urea nitrogen; ALT, Alanine aminotransferase; KRAS, kirsten rat sarcoma viral oncogene homolog; TP53, Tumor protein p53; CDKN2A, Cyclin-dependent kinase inhibitor 2A; SMAD4, SMAD family member 4.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Yang J, Xu R, Wang C, Qiu J, Ren B, You L. Early screening and diagnosis strategies of pancreatic cancer: a comprehensive review. Cancer Commun. 2021;41:1257–20. doi:10.1002/cac2.12204
2. Owens DK, Davidson KW, Krist AH, et al. Screening for pancreatic cancer: US preventive services task force reaffirmation recommendation statement. JAMA. 2019;322:438–444. doi:10.1001/jama.2019.10232
3. Qian L, Yu S, Chen Z, Meng Z, Huang S, Wang P. Functions and clinical implications of exosomes in pancreatic cancer. Biochim Biophys Acta Rev Cancer. 2019;1871:75–84. doi:10.1016/j.bbcan.2018.11.001
4. Giovannetti E, van der Borden CL, Frampton AE, Ali A, Firuzi O, Peters GJ. Never let it go: stopping key mechanisms underlying metastasis to fight pancreatic cancer. Semin Cancer Biol. 2017;44:43–59. doi:10.1016/j.semcancer.2017.04.006
5. Huang P, Gao W, Fu C, Tian R. Functional and clinical proteomic exploration of pancreatic cancer. Mol Cell Proteomics. 2023;22:100575. doi:10.1016/j.mcpro.2023.100575
6. Lai JJ, Chau ZL, Chen SY, et al. Exosome processing and characterization approaches for research and technology development. Adv Sci. 2022;9:e2103222. doi:10.1002/advs.202103222
7. Melo SA, Luecke LB, Kahlert C, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523:177–182. doi:10.1038/nature14581
8. Jiang Z, Wang H, Mou Y, Li L, Jin W. Functions and clinical applications of exosomes in pancreatic cancer. Mol Biol Rep. 2022;49:11037–11048. doi:10.1007/s11033-022-07765-8
9. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9:27. doi:10.1038/s41392-024-01735-1
10. He G, Liu J, Yu Y, et al. Revisiting the advances and challenges in the clinical applications of extracellular vesicles in cancer. Cancer Lett. 2024;593:216960. doi:10.1016/j.canlet.2024.216960
11. Cecchin R, Troyer Z, Witwer K, Morris KV. Extracellular vesicles: the next generation in gene therapy delivery. Mol Ther. 2023;31:1225–1230. doi:10.1016/j.ymthe.2023.01.021
12. Zhao Y, Tang J, Jiang K, Liu SY, Aicher A, Heeschen C. Liquid biopsy in pancreatic cancer - Current perspective and future outlook. Biochim Biophys Acta Rev Cancer. 2023;1878:188868. doi:10.1016/j.bbcan.2023.188868
13. Moghassemi S, Dadashzadeh A, Sousa MJ, et al. Extracellular vesicles in nanomedicine and regenerative medicine: a review over the last decade. Bioact Mater. 2024;36:126–156. doi:10.1016/j.bioactmat.2024.02.021
14. Zhao J, Zhu W, Mao Y, et al. Unignored intracellular journey and biomedical applications of extracellular vesicles. Adv Drug Deliv Rev. 2024;212:115388. doi:10.1016/j.addr.2024.115388
15. Cheng L, Hill AF. Therapeutically harnessing extracellular vesicles. Nat Rev Drug Discov. 2022;21:379–399. doi:10.1038/s41573-022-00410-w
16. Nannan L, Oudart JB, Monboisse JC, Ramont L, Brassart-Pasco S, Brassart B. Extracellular vesicle-dependent cross-talk in cancer-focus on pancreatic cancer. Front Oncol. 2020;10:1456. doi:10.3389/fonc.2020.01456
17. Bhatta B, Cooks T. Reshaping the tumor microenvironment: extracellular vesicles as messengers of cancer cells. Carcinogenesis. 2020;41:1461–1470. doi:10.1093/carcin/bgaa107
18. Han QF, Li WJ, Hu KS, et al. Exosome biogenesis: machinery, regulation, and therapeutic implications in cancer. Mol Cancer. 2022;21:207. doi:10.1186/s12943-022-01671-0
19. Tao J, Yang G, Zhou W, et al. Targeting hypoxic tumor microenvironment in pancreatic cancer. J Hematol Oncol. 2021;14:14. doi:10.1186/s13045-020-01030-w
20. Liu C, He D, Li L, et al. Extracellular vesicles in pancreatic cancer immune escape: emerging roles and mechanisms. Pharmacol Res. 2022;183:106364. doi:10.1016/j.phrs.2022.106364
21. Chen K, Wang Q, Liu X, Wang F, Yang Y, Tian X. Hypoxic pancreatic cancer derived exosomal miR-30b-5p promotes tumor angiogenesis by inhibiting GJA1 expression. Int J Biol Sci. 2022;18:1220–1237. doi:10.7150/ijbs.67675
22. Guo Z, Wang X, Yang Y, et al. Hypoxic tumor-derived exosomal long noncoding RNA UCA1 promotes angiogenesis via miR-96-5p/AMOTL2 in pancreatic cancer. Mol Ther Nucleic Acids. 2020;22:179–195. doi:10.1016/j.omtn.2020.08.021
23. Li J, Li Z, Jiang P, et al. Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis. J Exp Clin Cancer Res. 2018;37:177. doi:10.1186/s13046-018-0822-3
24. He Z, Wang J, Zhu C, et al. Exosome-derived FGD5-AS1 promotes tumor-associated macrophage M2 polarization-mediated pancreatic cancer cell proliferation and metastasis. Cancer Lett. 2022;548:215751. doi:10.1016/j.canlet.2022.215751
25. Li X, Li K, Li M, et al. Chemoresistance transmission via exosome-transferred MMP14 in pancreatic cancer. Front Oncol. 2022;12:844648. doi:10.3389/fonc.2022.844648
26. Li Y, Zhao W, Wang Y, Wang H, Liu S. Extracellular vesicle-mediated crosstalk between pancreatic cancer and stromal cells in the tumor microenvironment. J Nanobiotechnol. 2022;20:208. doi:10.1186/s12951-022-01382-0
27. Pereira BA, Vennin C, Papanicolaou M, et al. CAF subpopulations: a new reservoir of stromal targets in pancreatic cancer. Trends Cancer. 2019;5:724–741. doi:10.1016/j.trecan.2019.09.010
28. Xue R, Jia K, Wang J, et al. A rising star in pancreatic diseases: pancreatic stellate cells. Front Physiol. 2018;9:754. doi:10.3389/fphys.2018.00754
29. Takikawa T, Masamune A, Yoshida N, Hamada S, Kogure T, Shimosegawa T. Exosomes derived from pancreatic stellate cells: microRNA signature and effects on pancreatic cancer cells. Pancreas. 2017;46:19–27. doi:10.1097/MPA.0000000000000722
30. Rebelo R, Xavier CPR, Giovannetti E, Vasconcelos MH. Fibroblasts in pancreatic cancer: molecular and clinical perspectives. Trends Mol Med. 2023;29:439–453. doi:10.1016/j.molmed.2023.03.002
31. Yang Z, Zhao N, Cui J, Wu H, Xiong J, Peng T. Exosomes derived from cancer stem cells of gemcitabine-resistant pancreatic cancer cells enhance drug resistance by delivering miR-210. Cell Oncol. 2020;43:123–136. doi:10.1007/s13402-019-00476-6
32. Li M, Guo H, Wang Q, et al. Pancreatic stellate cells derived exosomal miR-5703 promotes pancreatic cancer by downregulating CMTM4 and activating PI3K/Akt pathway. Cancer Lett. 2020;490:20–30. doi:10.1016/j.canlet.2020.06.009
33. Cao W, Zeng Z, He Z, Lei S. Hypoxic pancreatic stellate cell-derived exosomal mirnas promote proliferation and invasion of pancreatic cancer through the PTEN/AKT pathway. Aging. 2021;13:7120–7132. doi:10.18632/aging.202569
34. Chi Y, Xin H, Liu Z. Exosomal lncRNA UCA1 derived from pancreatic stellate cells promotes gemcitabine resistance in pancreatic cancer via the SOCS3/EZH2 axis. Front Oncol. 2021;11:671082. doi:10.3389/fonc.2021.671082
35. Mao X, Xu J, Wang W, et al. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Mol Cancer. 2021;20:131. doi:10.1186/s12943-021-01428-1
36. Öhlund D, Handly-Santana A, Biffi G, et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J Exp Med. 2017;214:579–596. doi:10.1084/jem.20162024
37. Zhang F, Ma Y, Li D, et al. Cancer associated fibroblasts and metabolic reprogramming: unraveling the intricate crosstalk in tumor evolution. J Hematol Oncol. 2024;17:80. doi:10.1186/s13045-024-01600-2
38. Li Y, Zhang J, Xu J, Liu S. The metabolism symbiosis between pancreatic cancer and tumor microenvironment. Front Oncol. 2021;11:759376. doi:10.3389/fonc.2021.759376
39. Fang Y, Zhou W, Rong Y, et al. Exosomal miRNA-106b from cancer-associated fibroblast promotes gemcitabine resistance in pancreatic cancer. Exp Cell Res. 2019;383:111543. doi:10.1016/j.yexcr.2019.111543
40. Qi R, Bai Y, Li K, et al. Cancer-associated fibroblasts suppress ferroptosis and induce gemcitabine resistance in pancreatic cancer cells by secreting exosome-derived ACSL4-targeting miRNAs. Drug Resist Updat. 2023;68:100960. doi:10.1016/j.drup.2023.100960
41. Richards KE, Xiao W, Hill R; On Behalf of The Usc Pancreas Research Team. Cancer-associated fibroblasts confer gemcitabine resistance to pancreatic cancer cells through PTEN-targeting miRNAs in exosomes. Cancers. 2022;14:2812. doi:10.3390/cancers14112812
42. Chen L, Qu J, Kalyani FS, et al. Mesenchymal stem cell-based treatments for COVID-19: status and future perspectives for clinical applications. Cell Mol Life Sci. 2022;79:142.
43. Chamberlain G, Fox J, Ashton B, Middleton J. Concise review: mesenchymal stem cells: their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells. 2007;25:2739–2749.
44. Ludwig S, Floros T, Theodoraki MN, et al. Suppression of lymphocyte functions by plasma exosomes correlates with disease activity in patients with head and neck cancer. Clin Cancer Res. 2017;23:4843–4854.
45. Whiteside TL. Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment. Semin Immunol. 2018;35:69–79. doi:10.1016/j.smim.2017.12.003
46. Yeo RW, Lai RC, Zhang B, et al. Mesenchymal stem cell: an efficient mass producer of exosomes for drug delivery. Adv Drug Deliv Rev. 2013;65:336–341. doi:10.1016/j.addr.2012.07.001
47. Lai RC, Yeo RW, Padmanabhan J, Choo A, de Kleijn DP, Lim SK. Isolation and characterization of exosome from human embryonic stem cell-derived C-Myc-immortalized mesenchymal stem cells. Methods Mol Biol. 2016;1416:477–494.
48. Yao X, Mao Y, Wu D, et al. Exosomal circ_0030167 derived from BM-MSCs inhibits the invasion, migration, proliferation and stemness of pancreatic cancer cells by sponging miR-338-5p and targeting the Wif1/Wnt8/β-catenin axis. Cancer Lett. 2021;512:38–50.
49. Ding Y, Mei W, Zheng Z, et al. Exosomes secreted from human umbilical cord mesenchymal stem cells promote pancreatic ductal adenocarcinoma growth by transferring miR-100-5p. Tissue Cell. 2021;73:101623.
50. Peng X, Fang J, Lou C, et al. Engineered nanoparticles for precise targeted drug delivery and enhanced therapeutic efficacy in cancer immunotherapy. Acta Pharm Sin B. 2024;14:3432–3456. doi:10.1016/j.apsb.2024.05.010
51. Closa D. Pancreatic cancer, stroma, and exosomes. J Physiol Biochem. 2023;79:205–211. doi:10.1007/s13105-022-00898-1
52. Wang X, Luo G, Zhang K, et al. Hypoxic tumor-derived exosomal miR-301a mediates M2 macrophage polarization via PTEN/PI3Kγ to promote pancreatic cancer metastasis. Cancer Res. 2018;78:4586–4598. doi:10.1158/0008-5472.CAN-17-3841
53. Zhao M, Zhuang A, Fang Y. Cancer-associated fibroblast-derived exosomal miRNA-320a promotes macrophage M2 polarization in vitro by regulating PTEN/PI3Kγ signaling in pancreatic cancer. J Oncol. 2022;2022:9514697. doi:10.1155/2022/9514697
54. Ye M, Lu F, Gu D, et al. Hypoxia exosome derived CEACAM5 promotes tumor-associated macrophages M2 polarization to accelerate pancreatic neuroendocrine tumors metastasis via MMP9. FASEB J. 2024;38:e23762. doi:10.1096/fj.202302489RRR
55. Zheng P, Chen L, Yuan X, et al. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J Exp Clin Cancer Res. 2017;36:53. doi:10.1186/s13046-017-0528-y
56. Yang Y, Guo Z, Chen W, et al. M2 macrophage-derived exosomes promote angiogenesis and growth of pancreatic ductal adenocarcinoma by targeting E2F2. Mol Ther. 2021;29:1226–1238. doi:10.1016/j.ymthe.2020.11.024
57. Chang J, Li H, Zhu Z, et al. microRNA-21-5p from M2 macrophage-derived extracellular vesicles promotes the differentiation and activity of pancreatic cancer stem cells by mediating KLF3. Cell Biol Toxicol. 2022;38:577–590. doi:10.1002/advs.202306373
58. Li ZR, Luo XJ, Lin L, Zeng YH, Mai BX. Effect of laying sequence and selection of maternal tissues in assessment of maternal transfer of organohalogenated contaminants during chicken egg formation: a pilot study. Environ Pollut. 2021;270:116157. doi:10.1016/j.envpol.2020.116157
59. Yin Z, Ma T, Huang B, et al. Macrophage-derived exosomal microRNA-501-3p promotes progression of pancreatic ductal adenocarcinoma through the TGFBR3-mediated TGF-β signaling pathway. J Exp Clin Cancer Res. 2019;38:310. doi:10.1186/s13046-019-1313-x
60. Zhan T, Zou Y, Han Z, et al. Single-cell sequencing combined with spatial transcriptomics reveals that the IRF7 gene in M1 macrophages inhibits the occurrence of pancreatic cancer by regulating lipid metabolism-related mechanisms. Clin Transl Med. 2024;14:e1799. doi:10.1002/ctm2.1799
61. Gardner A, Ruffell B. Dendritic Cells and Cancer Immunity. Trends Immunol. 2016;37:855–865. doi:10.1016/j.it.2016.09.006
62. Verneau J, Sautés-Fridman C, Sun CM. Dendritic cells in the tumor microenvironment: prognostic and theranostic impact. Semin Immunol. 2020;48:101410. doi:10.1016/j.smim.2020.101410
63. Marciscano AE, Anandasabapathy N. The role of dendritic cells in cancer and anti-tumor immunity. Semin Immunol. 2021;52:101481. doi:10.1016/j.smim.2021.101481
64. Chen J, Wang S, Jia S, Ding G, Jiang G, Cao L. Integrated analysis of long non-coding RNA and mRNA expression profile in pancreatic cancer derived exosomes treated dendritic cells by microarray analysis. J Cancer. 2018;9:21–31. doi:10.7150/jca.21749
65. Ding G, Zhou L, Qian Y, et al. Pancreatic cancer-derived exosomes transfer miRNAs to dendritic cells and inhibit RFXAP expression via miR-212-3p. Oncotarget. 2015;6:29877–29888. doi:10.18632/oncotarget.4924
66. Chen T, Zhang MG, Xu HX, Wang WQ, Liu L, Yu XJ. Preoperative serum CA125 levels predict the prognosis in hyperbilirubinemia patients with resectable pancreatic ductal adenocarcinoma. Medicine. 2015;94:e751. doi:10.1097/MD.0000000000000751
67. Li W, Liu JB, Hou LK, et al. Liquid biopsy in lung cancer: significance in diagnostics, prediction, and treatment monitoring. Mol Cancer. 2022;21:25. doi:10.1186/s12943-022-01505-z
68. Zhang B, Pan X, Cobb GP, Anderson TA. microRNAs as oncogenes and tumor suppressors. Dev Biol. 2007;302:1–12. doi:10.1016/j.ydbio.2006.08.028
69. Sharma GG, Okada Y, Von Hoff D, Goel A. Non-coding RNA biomarkers in pancreatic ductal adenocarcinoma. Semin Cancer Biol. 2021;75:153–168. doi:10.1016/j.semcancer.2020.10.001
70. Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinf. 2015;13:17–24. doi:10.1016/j.gpb.2015.02.001
71. Goldie BJ, Dun MD, Lin M, et al. Activity-associated miRNA are packaged in Map1b-enriched exosomes released from depolarized neurons. Nucleic Acids Res. 2014;42:9195–9208. doi:10.1093/nar/gku594
72. Zhao R, Zhao T, He Z, Cai R, Pang W. Composition, isolation, identification and function of adipose tissue-derived exosomes. Adipocyte. 2021;10:587–604. doi:10.1080/21623945.2021.1983242
73. Lai X, Wang M, McElyea SD, Sherman S, House M, Korc M. A microRNA signature in circulating exosomes is superior to exosomal glypican-1 levels for diagnosing pancreatic cancer. Cancer Lett. 2017;393:86–93. doi:10.1016/j.canlet.2017.02.019
74. Nakamura S, Sadakari Y, Ohtsuka T, et al. Pancreatic juice exosomal MicroRNAs as biomarkers for detection of pancreatic ductal adenocarcinoma. Ann Surg Oncol. 2019;26:2104–2111. doi:10.1245/s10434-019-07269-z
75. Long Y, Wang X, Youmans DT, Cech TR. How do lncRNAs regulate transcription? Sci Adv. 2017;3:eaao2110. doi:10.1126/sciadv.aao2110
76. Robless EE, Howard JA, Casari I, Falasca M. Exosomal long non-coding RNAs in the diagnosis and oncogenesis of pancreatic cancer. Cancer Lett. 2021;501:55–65.
77. Müller S, Raulefs S, Bruns P, et al. Next-generation sequencing reveals novel differentially regulated mRNAs, lncRNAs, miRNAs, sdRNAs and a piRNA in pancreatic cancer. Mol Cancer. 2015;14:94.
78. Jiang Y, Li Z, Zheng S, et al. The long non-coding RNA HOTAIR affects the radiosensitivity of pancreatic ductal adenocarcinoma by regulating the expression of Wnt inhibitory factor 1. Tumour Biol. 2016;37:3957–3967.
79. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674.
80. Cheng K, Pan J, Liu Q, et al. Exosomal lncRNA XIST promotes perineural invasion of pancreatic cancer cells via miR-211-5p/GDNF. Oncogene. 2024;43:1341–1352.
81. Sun Z, Sun D, Feng Y, et al. Exosomal linc-ROR mediates crosstalk between cancer cells and adipocytes to promote tumor growth in pancreatic cancer. Mol Ther Nucleic Acids. 2021;26:253–268.
82. He X, Chen L, Di Y, et al. Plasma-derived exosomal long noncoding RNAs of pancreatic cancer patients as novel blood-based biomarkers of disease. BMC Cancer. 2024;24:961. doi:10.1186/s12885-024-12755-z
83. Yee NS, Zhang S, He HZ, Zheng SY. Extracellular vesicles as potential biomarkers for early detection and diagnosis of pancreatic cancer. Biomedicines. 2020;8:581. doi:10.3390/biomedicines8120581
84. Castillo J, Bernard V, San Lucas FA, et al. Surfaceome profiling enables isolation of cancer-specific exosomal cargo in liquid biopsies from pancreatic cancer patients. Ann Oncol. 2018;29:223–229. doi:10.1093/annonc/mdx542
85. Mann KM, Ying H, Juan J, Jenkins NA, Copeland NG. KRAS-related proteins in pancreatic cancer. Pharmacol Ther. 2016;168:29–42.
86. Frampton AE, Prado MM, López-Jiménez E, et al. Glypican-1 is enriched in circulating-exosomes in pancreatic cancer and correlates with tumor burden. Oncotarget. 2018;9:19006–19013. doi:10.18632/oncotarget.24873
87. Li H, Chiang CL, Kwak KJ, et al. Extracellular vesicular analysis of Glypican 1 mRNA and protein for pancreatic cancer diagnosis and prognosis. Adv Sci. 2024;11:e2306373.
88. Yang J, Zhang Y, Gao X, et al. Plasma-derived exosomal ALIX as a novel biomarker for diagnosis and classification of pancreatic cancer. Front Oncol. 2021;11:628346. doi:10.3389/fonc.2021.628346
89. Kadota T, Fujita Y, Yoshioka Y, Araya J, Kuwano K, Ochiya T. Extracellular Vesicles in Chronic Obstructive Pulmonary Disease. Int J Mol Sci. 2016;17:1801.
90. Xu K, Feng H, Zhao R, Huang Y. Targeting tetraspanins at cell interfaces: functional modulation and exosome-based drug delivery for precise disease treatment. ChemMedChem. 2024;20:e202400664. doi:10.1002/cmdc.202400664
91. Wredenberg A, Lagouge M, Bratic A, et al. MTERF3 regulates mitochondrial ribosome biogenesis in invertebrates and mammals. PLoS Genet. 2013;9:e1003178. doi:10.1371/journal.pgen.1003178
92. Catalano M, O’Driscoll L. Inhibiting extracellular vesicles formation and release: a review of EV inhibitors. J Extracell Vesicles. 2020;9:1703244. doi:10.1080/20013078.2019.1703244
93. Trajkovic K, Hsu C, Chiantia S, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319:1244–1247. doi:10.1126/science.1153124
94. Li B, Antonyak MA, Zhang J, Cerione RA. RhoA triggers a specific signaling pathway that generates transforming microvesicles in cancer cells. Oncogene. 2012;31:4740–4749. doi:10.1038/onc.2011.636
95. Siklos M, BenAissa M, Thatcher GR. Cysteine proteases as therapeutic targets: does selectivity matter? A systematic review of calpain and cathepsin inhibitors. Acta Pharm Sin B. 2015;5:506–519. doi:10.1016/j.apsb.2015.08.001
96. Richards KE, Zeleniak AE, Fishel ML, Wu J, Littlepage LE, Hill R. Cancer-associated fibroblast exosomes regulate survival and proliferation of pancreatic cancer cells. Oncogene. 2017;36:1770–1778. doi:10.1038/onc.2016.353
97. Wang L, Yang L, Zhuang T, Shi X. Tumor-derived exosomal miR-29b reduces angiogenesis in pancreatic cancer by silencing ROBO1 and SRGAP2. J Immunol Res. 2022;2022:4769385. doi:10.1155/2022/4769385
98. Rimmer E, Rashid S, Kraev I, Miralles F, Elia A. Extracellular vesicles inhibit the response of pancreatic ductal adenocarcinoma cells to gemcitabine and TRAIL treatment. Int J Mol Sci. 2022;23:7810. doi:10.3390/ijms23147810
99. Vahabi M, Comandatore A, Centra C, Blandino G, Morelli L, Giovannetti E. Thinking small to win big? A critical review on the potential application of extracellular vesicles for biomarker discovery and new therapeutic approaches in pancreatic cancer. Semin Cancer Biol. 2023;97:50–67. doi:10.1016/j.semcancer.2023.11.003
100. Xu Y, Lai Y, Cao L, et al. Human umbilical cord mesenchymal stem cells-derived exosomal microRNA-451a represses epithelial-mesenchymal transition of hepatocellular carcinoma cells by inhibiting ADAM10. RNA Biol. 2021;18:1408–1423. doi:10.1080/15476286.2020.1851540
101. Zhu X, Li Z, Chen L, et al. Exosomes delivering miR-129-5p combined with sorafenib ameliorate hepatocellular carcinoma progression via the KCTD1/HIF-1α/VEGF pathway. Cell Oncol. 2025;48:743–760. doi:10.1007/s13402-025-01044-x
102. Choi YH, Kim HY, Park JO, Choi E. Enhanced anti-tumor effects of natural killer cell-derived exosomes through doxorubicin delivery to hepatocellular carcinoma cells: cytotoxicity and apoptosis study. Int J Mol Sci. 2025;26:2234.
103. Wang J, Tang W, Yang M, et al. Inflammatory tumor microenvironment responsive neutrophil exosomes-based drug delivery system for targeted glioma therapy. Biomaterials. 2021;273:120784. doi:10.1016/j.biomaterials.2021.120784
104. Filipazzi P, Bürdek M, Villa A, Rivoltini L, Huber V. Recent advances on the role of tumor exosomes in immunosuppression and disease progression. Semin Cancer Biol. 2012;22:342–349. doi:10.1016/j.semcancer.2012.02.005
105. Pullan JE, Confeld MI, Osborn JK, Kim J, Sarkar K, Mallik S. Exosomes as Drug Carriers for Cancer Therapy. Mol Pharm. 2019;16:1789–1798. doi:10.1021/acs.molpharmaceut.9b00104
106. Xu B, Zuo C. Effect of exosomes as drug carriers in chemotherapy of pancreatic cancer. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2023;48:268–274. doi:10.11817/j.issn.1672-7347.2023.220439
107. Thomas D, Radhakrishnan P. Tumor-stromal crosstalk in pancreatic cancer and tissue fibrosis. Mol Cancer. 2019;18:14. doi:10.1186/s12943-018-0927-5
108. Wong A, Soo RA, Yong WP, Innocenti F. Clinical pharmacology and pharmacogenetics of gemcitabine. Drug Metab Rev. 2009;41:77–88. doi:10.1080/03602530902741828
109. Li YJ, Wu JY, Wang JM, Hu XB, Cai JX, Xiang DX. Gemcitabine loaded autologous exosomes for effective and safe chemotherapy of pancreatic cancer. Acta Biomater. 2020;101:519–530. doi:10.1016/j.actbio.2019.10.022
110. Kanchanapally R, Deshmukh SK, Chavva SR, et al. Drug-loaded exosomal preparations from different cell types exhibit distinctive loading capability, yield, and antitumor efficacies: a comparative analysis. Int J Nanomed. 2019;14:531–541. doi:10.2147/IJN.S191313
111. Lázaro-Ibáñez E, Faruqu FN, Saleh AF, et al. Selection of fluorescent, bioluminescent, and radioactive tracers to accurately reflect extracellular vesicle biodistribution in vivo. ACS Nano. 2021;15:3212–3227. doi:10.1021/acsnano.0c09873
112. Usman WM, Pham TC, Kwok YY, et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat Commun. 2018;9:2359. doi:10.1038/s41467-018-04791-8
113. Wang P, Wang H, Huang Q, et al. Exosomes from M1-polarized macrophages enhance paclitaxel antitumor activity by activating macrophages-mediated inflammation. Theranostics. 2019;9:1714–1727. doi:10.7150/thno.30716
114. Reese M, Dhayat SA. Small extracellular vesicle non-coding RNAs in pancreatic cancer: molecular mechanisms and clinical implications. J Hematol Oncol. 2021;14:141. doi:10.1186/s13045-021-01149-4
115. Krützfeldt J, Rajewsky N, Braich R, et al. Silencing of microRNAs in vivo with ‘antagomirs’. Nature. 2005;438:685–689. doi:10.1038/nature04303
116. Huang X, Schwind S, Yu B, et al. Targeted delivery of microRNA-29b by transferrin-conjugated anionic lipopolyplex nanoparticles: a novel therapeutic strategy in acute myeloid leukemia. Clin Cancer Res. 2013;19:2355–2367. doi:10.1158/1078-0432.CCR-12-3191
117. Ibrahim AF, Weirauch U, Thomas M, Grünweller A, Hartmann RK, Aigner A. MicroRNA replacement therapy for miR-145 and miR-33a is efficacious in a model of colon carcinoma. Cancer Res. 2011;71:5214–5224. doi:10.1158/0008-5472.CAN-10-4645
118. Gilleron J, Querbes W, Zeigerer A, et al. Image-based analysis of lipid nanoparticle-mediated siRNA delivery, intracellular trafficking and endosomal escape. Nat Biotechnol. 2013;31:638–646. doi:10.1038/nbt.2612
119. Younis MA, Tawfeek HM, Abdellatif AAH, Abdel-Aleem JA, Harashima H. Clinical translation of nanomedicines: challenges, opportunities, and keys. Adv Drug Deliv Rev. 2022;181:114083. doi:10.1016/j.addr.2021.114083
120. van der Meel R, Fens MH, Vader P, van Solinge WW, Eniola-Adefeso O, Schiffelers RM. Extracellular vesicles as drug delivery systems: lessons from the liposome field. J Control Release. 2014;195:72–85. doi:10.1016/j.jconrel.2014.07.049
121. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
122. Sercombe L, Veerati T, Moheimani F, Wu SY, Sood AK, Hua S. Advances and challenges of liposome assisted drug delivery. Front Pharmacol. 2015;6:286. doi:10.3389/fphar.2015.00286
123. Fang X, Lan H, Jin K, Qian J. Pancreatic cancer and exosomes: role in progression, diagnosis, monitoring, and treatment. Front Oncol. 2023;13:1149551. doi:10.3389/fonc.2023.1149551
124. Xu L, Faruqu FN, Lim YM, et al. Exosome-mediated RNAi of PAK4 prolongs survival of pancreatic cancer mouse model after loco-regional treatment. Biomaterials. 2021;264:120369. doi:10.1016/j.biomaterials.2020.120369
125. Zhou W, Zhou Y, Chen X, et al. Pancreatic cancer-targeting exosomes for enhancing immunotherapy and reprogramming tumor microenvironment. Biomaterials. 2021;268:120546. doi:10.1016/j.biomaterials.2020.120546
126. Xu Z, Zeng S, Gong Z, Yan Y. Exosome-based immunotherapy: a promising approach for cancer treatment. Mol Cancer. 2020;19:160. doi:10.1186/s12943-020-01278-3
127. Xu C, Hong H, Lee Y, et al. Efficient lymph node-targeted delivery of personalized cancer vaccines with reactive oxygen species-inducing reduced graphene oxide nanosheets. ACS Nano. 2020;14:13268–13278. doi:10.1021/acsnano.0c05062
128. Zhao Y, Liu L, Sun R, et al. Exosomes in cancer immunoediting and immunotherapy. Asian J Pharm Sci. 2022;17:193–205. doi:10.1016/j.ajps.2021.12.001
129. Jafari R, Rahbarghazi R, Ahmadi M, Hassanpour M, Rezaie J. Hypoxic exosomes orchestrate tumorigenesis: molecular mechanisms and therapeutic implications. J Transl Med. 2020;18:474. doi:10.1186/s12967-020-02662-9
130. Zhou W, Chen X, Zhou Y, et al. Exosomes derived from immunogenically dying tumor cells as a versatile tool for vaccination against pancreatic cancer. Biomaterials. 2022;280:121306. doi:10.1016/j.biomaterials.2021.121306
131. Shirejini SZ, Inci F. The Yin and Yang of exosome isolation methods: conventional practice, microfluidics, and commercial kits. Biotechnol Adv. 2022;54:107814. doi:10.1016/j.biotechadv.2021.107814
132. Shao H, Chung J, Balaj L, et al. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat Med. 2012;18:1835–1840. doi:10.1038/nm.2994
133. Giri PM, Banerjee A, Layek B. A recent review on cancer nanomedicine. Cancers. 2023;15:2256. doi:10.3390/cancers15082256
134. Giri PM, Banerjee A, Ghosal A, Layek B. Neuroinflammation in neurodegenerative disorders: current knowledge and therapeutic implications. Int J Mol Sci. 2024;25:3995. doi:10.3390/ijms25073995
135. Zhang K, Fang J. Extracellular vesicles in drug delivery: from quality assurance to therapeutic application. Int J Nanomed. 2026;21:579695. doi:10.2147/IJN.S579695
136. Zhang Q, Jeppesen DK, Higginbotham JN, Franklin JL, Coffey RJ. Comprehensive isolation of extracellular vesicles and nanoparticles. Nat Protoc. 2023;18:1462–1487. doi:10.1038/s41596-023-00811-0
137. Palmulli R, Couty M, Piontek MC, et al. CD63 sorts cholesterol into endosomes for storage and distribution via exosomes. Nat Cell Biol. 2024;26:1093–1109. doi:10.1038/s41556-024-01432-9
138. Qin B, Zhang Q, Hu XM, et al. How does temperature play a role in the storage of extracellular vesicles? J Cell Physiol. 2020;235:7663–7680. doi:10.1002/jcp.29700
139. Hill ML, Chung SJ, Woo HJ, et al. Exosome-coated prussian blue nanoparticles for specific targeting and treatment of glioblastoma. ACS Appl Mater Interfaces. 2024;16:20286–20301. doi:10.1021/acsami.4c02364
140. Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet. 2008;9:102–114. doi:10.1038/nrg2290
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
A Mini-Review of Diagnostic and Therapeutic Nano-Tools for Pancreatitis

Zhang Q, Li S, Yu Y, Zhu Y, Tong R
International Journal of Nanomedicine 2022, 17:4367-4381
Published Date: 19 September 2022
Advances and Prospects in the Treatment of Pancreatic Cancer
Duan H, Li L, He S
International Journal of Nanomedicine 2023, 18:3973-3988
Published Date: 19 July 2023
Combining Olaparib and Ascorbic Acid on Nanoparticles to Enhance the Drug Toxic Effects in Pancreatic Cancer

Quiñonero F, Parra-Torrejón B, Ramírez-Rodríguez GB, Garcés V, Delgado-López JM, Jiménez-Luna C, Perazzoli G, Melguizo C, Prados J, Ortíz R
International Journal of Nanomedicine 2023, 18:5075-5093
Published Date: 6 September 2023
Extracellular Vesicles in Idiopathic Pulmonary Fibrosis: Pathogenesis, Biomarkers and Innovative Therapeutic Strategies

Yang Y, Lv M, Xu Q, Wang X, Fang Z
International Journal of Nanomedicine 2024, 19:12593-12614
Published Date: 25 November 2024
Research and Application Prospect of Nanomedicine in Kidney Disease: A Bibliometric Analysis From 2003 to 2024

Meng Y, Sui L, Xu T, Zhao H, Yuan Q, Sun L
International Journal of Nanomedicine 2025, 20:3007-3030
Published Date: 12 March 2025