Back to Journals » Journal of Inflammation Research » Volume 19
Circulating Plasma Proteins Influence the Risk of Aortic Dissection via Blood Pressure: A Network Mendelian Randomization and Multi-Omics Study
Authors Li Z ![]() , Liu J
, Liu J ![]() , Fan X, Shi L, Shi H, Li C, Duan C, Zhang C, Wu Q, Shao Y
, Fan X, Shi L, Shi H, Li C, Duan C, Zhang C, Wu Q, Shao Y
Received 24 February 2026
Accepted for publication 19 March 2026
Published 31 March 2026 Volume 2026:19 600596
DOI https://doi.org/10.2147/JIR.S600596
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Chengming Fan
Zhenghao Li,1,* Jian Liu,2,* Xiaoxi Fan,1 Liangtao Shi,1 Haoming Shi,1 Changying Li,1 Changzhu Duan,3 Cheng Zhang,1 Qingchen Wu,1 Yue Shao1
1Department of Cardiothoracic Surgery, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China; 2Department of Cardiology, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China; 3Department of Cell Biology and Genetics, Center for Molecular Medicine and Oncology Research, Chongqing Medical University, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yue Shao, Department of Cardiothoracic Surgery, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China, Email [email protected] Qingchen Wu, Department of Cardiothoracic Surgery, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China, Email [email protected]
Background: The causal link between plasma proteins and aortic dissection (AD), and the role of blood pressure (BP) in this relationship, is not well understood.
Methods: We applied an integrated network Mendelian randomization (MR) and multi-omics framework to investigate the causal effects of plasma proteins on AD and the mediating role of BP. Genetic instruments for plasma proteins were derived from deCODE genetics (n = 35,559), and genome-wide association studies (GWAS) data for BP and AD were sourced from UK Biobank (n = 458,855), Million Veteran Program (n = 449,042) and FinnGen (n = 464,256). We conducted proteome-wide MR, mediation MR, and colocalization analyses to pinpoint causal proteins and pathways. Robustness was checked through sensitivity and replication analyses. Validation was implemented by replication MR analyses and our proteomic sequencing of East Asian population. Downstream analyses, including single‑cell RNA sequencing analysis, drug target analysis and phenome-wide association study (PheWAS) analysis were performed.
Results: Genetically predicted hypertension and diastolic BP were linked to a higher risk of AD. Proteome-wide MR identified 9 plasma proteins affecting both BP and AD, notably CCN3, COL6A3, NPPB, and NQO1, with BP mediating 5.43– 22.74% of their effects on AD. Colocalization analysis showed shared genetic variants between CCN3 and DBP (PP.H4 = 92.10%), and COL6A3 and AD (PP.H4 = 71.60%). Subsequent analyses validated the mediating effects and expression tendency of candidate proteins, revealed their underlying molecular mechanisms, and identified potential targeted drugs.
Conclusion: By mapping plasma proteins that affect AD risk and quantifying the mediation effect of BP, our work nominates CCN3, COL6A3, NPPB, and NQO1 as promising biomarkers and therapeutic targets, aiming to provide novel strategies for the early identification and intervention of AD.
Keywords: plasma proteins, blood pressure, hypertension, aortic dissection, Mendelian randomization, multi-omics
Introduction
Aortic dissection (AD) is a critical cardiovascular condition involving a tear in the aorta’s inner layer, allowing blood to create a hematoma between layers,1 leading to potential complications like aortic rupture and shock, with a high early mortality rate of 30%.2 Hypertension (HTN) is the leading risk factor, significantly increasing the likelihood of AD compared to those with normal BP.3
Significant evidence suggests that HTN and AD share common lifestyle and genetic risk factors.3–5 However, the molecular links between HTN and AD are not fully understood. Proteomics provides a powerful approach to identify molecular targets involved in cellular processes and represents a major source of therapeutic targets for cardiovascular diseases (CVDs).6–8 Plasma proteins may act as mediators of HTN and AD or biomarkers linked to arterial pathophysiology. Therefore, investigating the proteomic landscape of HTN and AD could yield crucial insights into the molecular determinants and pathways. Mendelian randomization (MR) uses genetic variants to explore causal relationships between plasma proteins, BP, and AD, minimizing confounding issues.9–11 Recent large-scale proteomic and genomic data have facilitated the use of MR to systematically identify plasma proteins linked to these conditions.6 These proteins served as potential circulating biomarkers and promising therapeutic targets, providing a valuable strategy for the early prediction of AD and aortic aneurysm (AA).7
We hypothesize that there exists a portion of the plasma proteome is linked to both HTN and AD, and that these proteins influence the risk of AD by exerting their regulatory effects on BP. This study addresses the knowledge gap on the causal role of plasma proteins in AD by conducting an MR investigation. We aim to identify plasma proteins affecting AD and quantify BP’s mediating role. Using network MR, we seek to establish a framework for understanding the “plasma protein-BP-AD” axis and provide insights into the molecular pathophysiology of AD.
Methods
Study Design
As depicted in Figure 1, our network MR analysis encompasses several key steps. Initially, two-sample MR analysis was carried out to determine the causal relationships among BP traits [essential HTN, systolic BP (SBP), and diastolic BP (DBP)] and AD. Subsequently, we conducted a proteome-wide MR analysis utilizing cis protein quantitative trait loci (cis-pQTL) of plasma proteins to determine their causal effects on BP traits and AD. Following these analyses, we executed functional enrichment and colocalization analysis. Then a mediation MR analysis to estimate the causal effect of plasma proteins on AD mediated by BP traits was performed, along with the corresponding mediation proportion. To ensure the robustness of the MR results, we also conducted sensitivity and replication analyses. In the final phase, to prevent ethnic heterogeneity and enhance validation robustness, we recruited an independent clinical cohort. We engaged in single cell RNA sequencing (scRNA-seq) analysis to explore the cellular localization and interactions of genes encoding proteins and assessed druggability potential through data mining and phenome-wide association studies (PheWAS). The detailed methods are provided in the Supplementary Material.
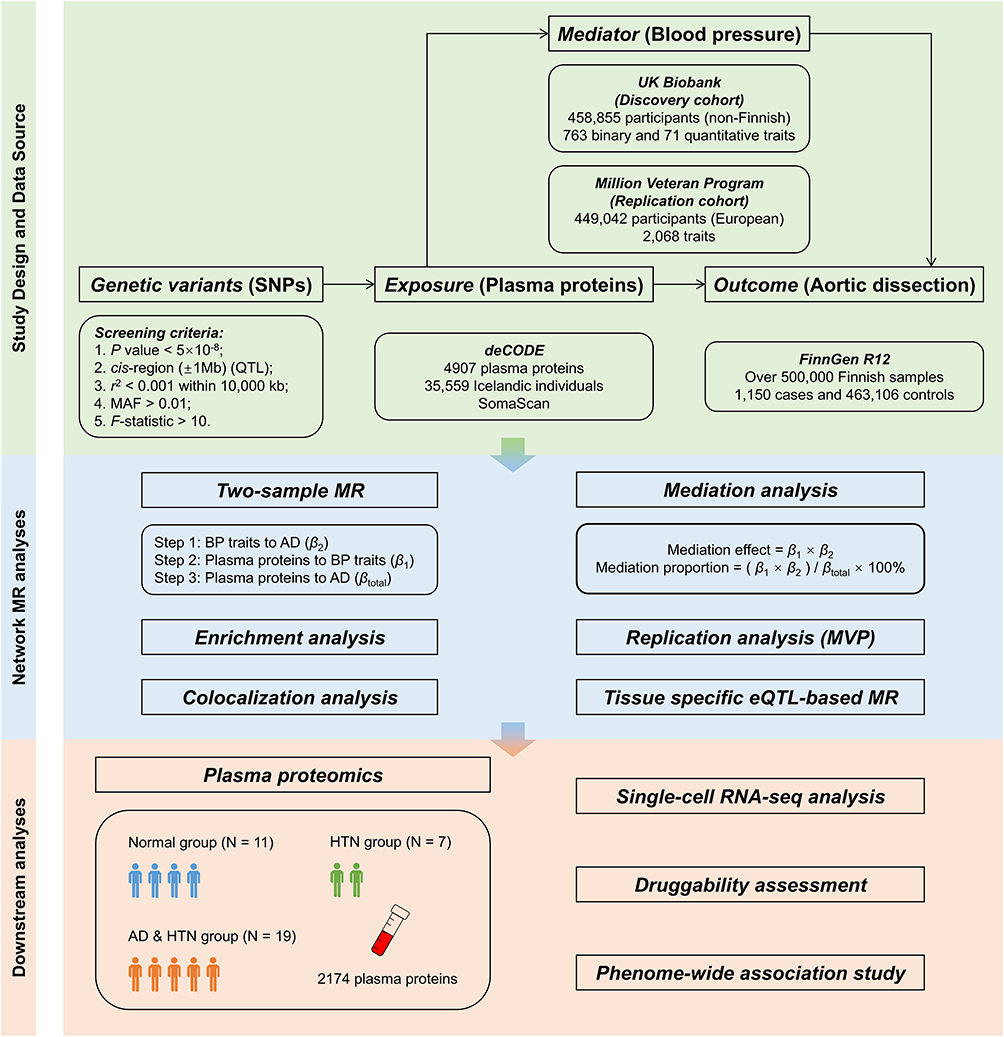 |
Figure 1 Overview of the study design. Abbreviations: SNP, single-nucleotide polymorphism; QTL, quantitative trait loci; MAF, minor allele frequency; MR, Mendelian randomization; BP, blood pressure; AD, aortic dissection; MVP, Million Veteran Program; HTN, hypertension; AD, aortic dissection. |
Data Sources
Table 1 provides details on the genome-wide association studies (GWAS) utilized in this research. The GWAS summary statistics for plasma proteins were obtained from deCODE. Using the SomaScan version 4 assay (SomaLogic), deCODE measured 4,907 plasma proteins in 35,559 Icelandic individuals from the Icelandic Cancer Project and various genetic programs at deCODE genetics, thereby identifying genetic variants associated with distinct plasma proteins.12 The GWAS summary statistics for BP traits were derived from the UK Biobank (UKB) (discovery cohort) and Million Veteran Program (MVP) (replication cohort) studies, which were stored in GWAS Catalog.13 The whole-genome sequencing of 490,640 participants from the UKB produced GWAS datasets encompassing 763 binary and 71 quantitative phenotypes. To avoid sample overlap, we only included 458,855 participants of non-Finnish European ancestry in our analysis.14 The Department of Veterans Affairs MVP conducted GWAS on 2,068 traits in 635,969 participants. To ensure consistency in population, we only selected 449,042 participants of European ancestry for analysis.15 The GWAS summary statistics for AD were obtained from the 12th release of the FinnGen study. FinnGen is a large-scale genomics initiative involving more than 500,000 samples from the Finnish biobank, which included 1,150 patients with AD and 463,106 healthy controls.16 The cis expression quantitative trait loci (cis-eQTL) data was obtained from eQTLGen Consortium and Genotype-Tissue Expression (GTEx) Portal.17,18
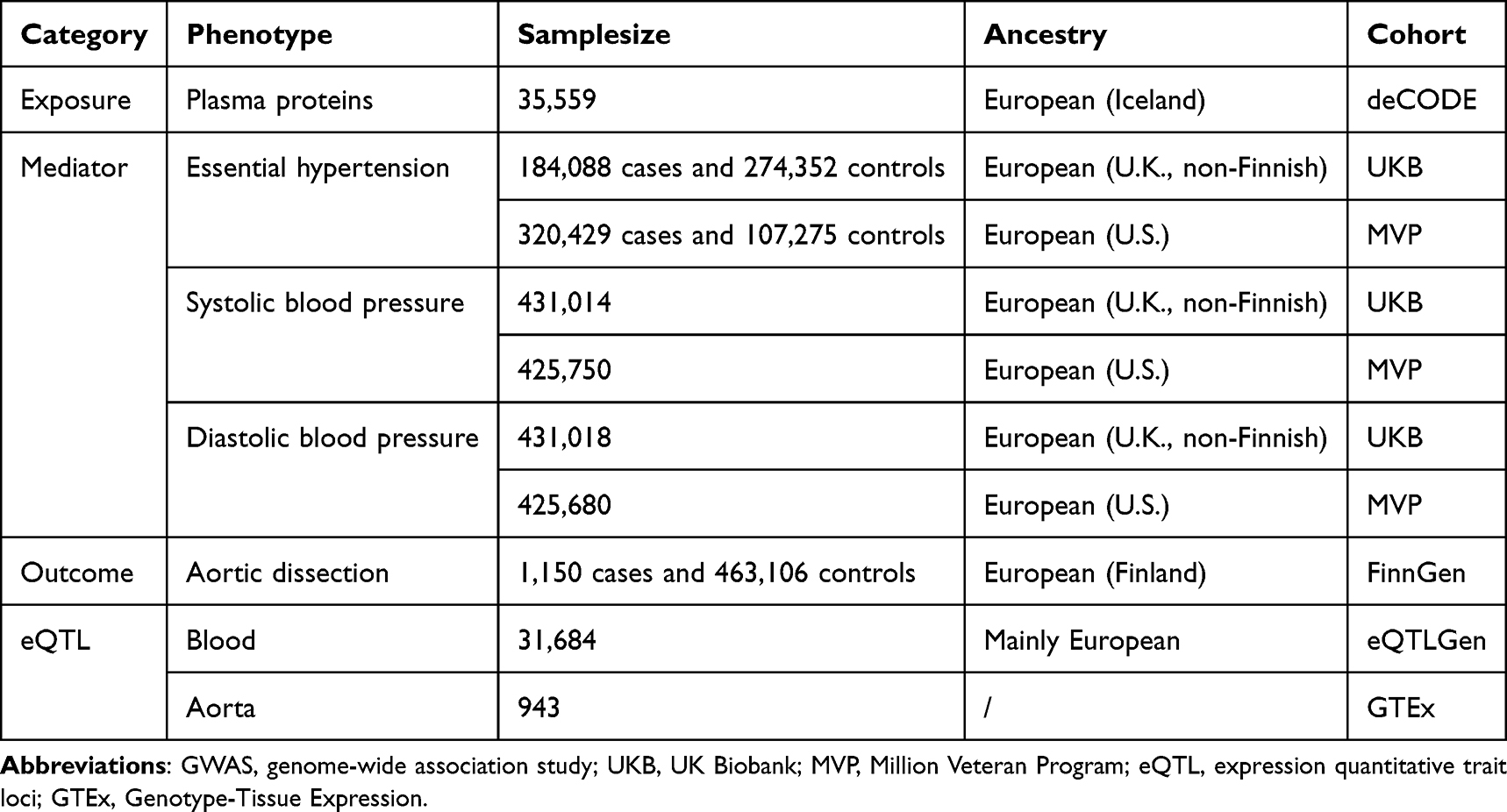 |
Table 1 Detailed Information of GWAS Datasets Used in This Study |
Plasma Proteomics
We collected blood samples from patients with acute Stanford type A AD complicated by a history of HTN who were admitted to the Yuanjiagang Branch of the First Affiliated Hospital of Chongqing Medical University between January 2024 and December 2025. The history of HTN was provided by the patients or their relatives, and all patients were diagnosis by aortic computed tomography angiography (CTA). Blood samples from healthy and hypertensive individuals were obtained from participants who underwent health examination at the Health Management Center of the Jingwei Branch of the First Affiliated Hospital of Chongqing Medical University during 2025. The collected blood samples were sent to plasma proteomic sequencing by Shanghai Personal Biotechnology Co., Ltd. This study was approved by the Ethics Committee of the First Affiliated Hospital of Chongqing Medical University, and written informed consent was obtained from all participants.
Statistical Analyses
All statistical analyses were performed using R software (version 4.3.3). The “TwoSampleMR” and “ieugwasr” packages were used for data acquisition, integration and analysis, the “tidyverse” and “xQTLbiolinks” packages were used for data organization and visualization. The “Seurat” and “CellChat” packages were used for scRNA-seq analyses. To identify more causal proteins, the Benjamini-Hochberg (BH) method was applied to correct for multiple testing of the false discovery rate (FDR) for the MR results excluding plasma protein to AD.
Results
Association Between BP Traits and AD
The results from two-sample MR analysis showed that genetically predicted HTN (OR = 1.52 [95% CI, 1.27–1.81], Adjusted P = 2.10×10−5) and DBP (OR = 2.97 [95% CI, 1.88–4.69], Adjusted P = 2.10×10−5) were associated with an increased risk of AD, and no significant causal association was observed between genetically predicted SBP and AD (OR = 1.15 [95% CI, 0.76–1.74], Adjusted P = 0.51) (Figure 2A and Table S1). However, heterogeneity and horizontal pleiotropy were detected in all the above results. After correcting for outlier SNPs that indicated potential horizontal pleiotropy, the associations remained (Table S1).
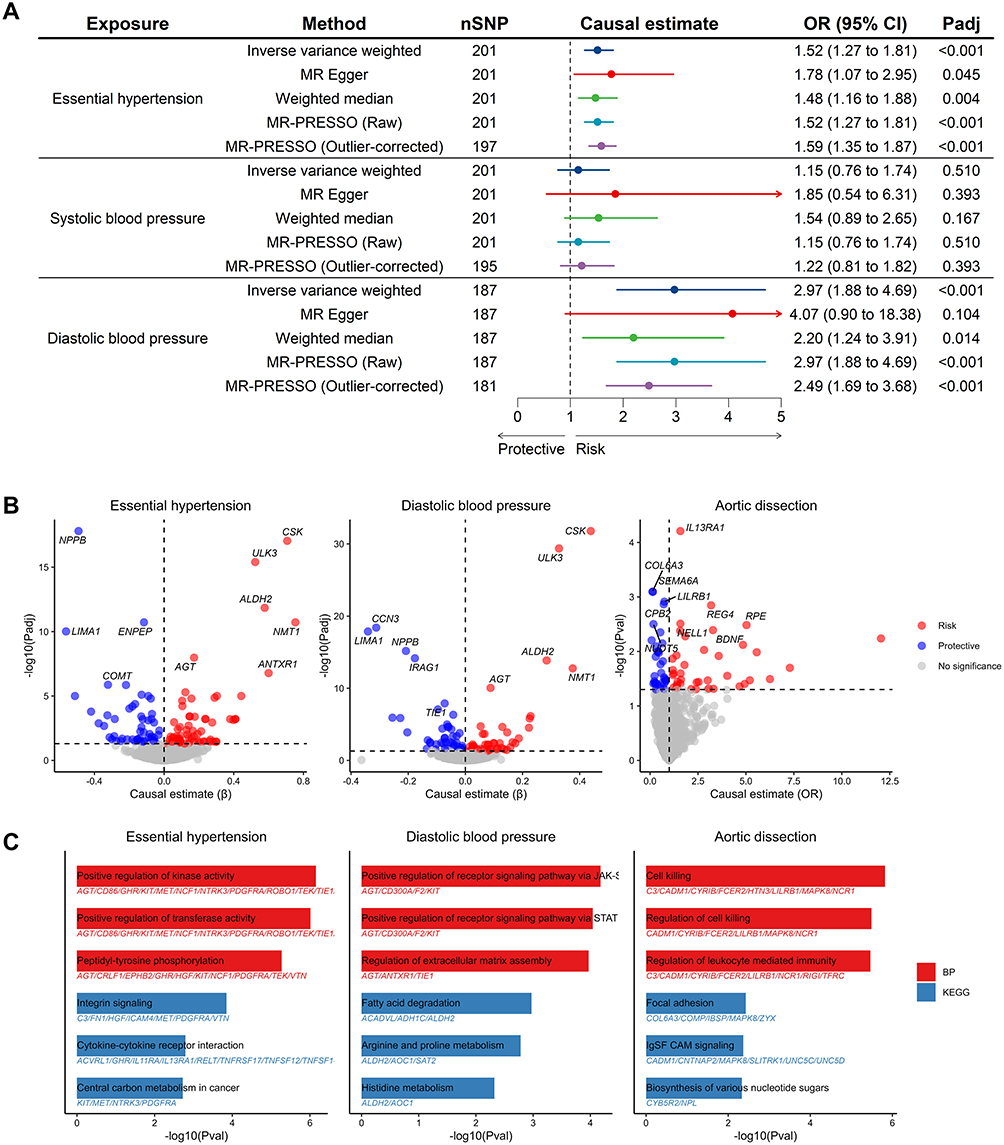 |
Figure 2 Causal association between plasma proteins, BP traits, and AD. (A) Causal association between BP traits and AD. (B) Causal association between plasma proteins and HTN, DBP, and AD, respectively. (C) Functional enrichment analysis of causal plasma proteins of HTN, DBP, and AD. Abbreviations: BP, blood pressure; HTN, hypertension; DBP, diastolic blood pressure; AD, aortic dissection. |
Genetically Predicted Proteins Associated with BP Traits and AD
The proteome-wide MR analysis identified 176 plasma proteins associated with HTN or DBP (118 with HTN, 90 with DBP, and 32 overlaps), as well as 73 plasma proteins associated with AD (Figure 2B and Tables S2–S4). Causal proteins were enriched in biological processes like regulation of cell junction assembly, leukocyte proliferation, angiogenesis and vasculature development, and pathways like cytokine-cytokine receptor interaction and biosynthesis of cofactors, suggesting that these proteins may exert their effects in HTN and AD through these pathophysiological progressions (Figure 2C and Tables S5–S7). Among these, 9 proteins exhibited causal relationships with both BP traits and AD (5 with HTN, 4 with DBP) (Figure 3A). Colocalization analysis showed that CCN3 and DBP (PPH4 = 92.10%), as well as COL6A3 and AD (PPH4 = 71.60%), shared a single causal variant (Figure 3B–D and Table S8). Multi-trait colocalization analysis did not identify genetic colocalization among plasma proteins, BP traits, and AD, but the strong colocalization between CCN3 and DBP was confirmed (PP = 92.08%, lending by rs56013326) (Tables S9–S10). Sensitivity analyses showed no horizontal pleiotropy, but heterogeneity was detected in NPPB to DBP, and MR results by different methods were consistent in causal direction except SHMT1 to AD (Table S11).
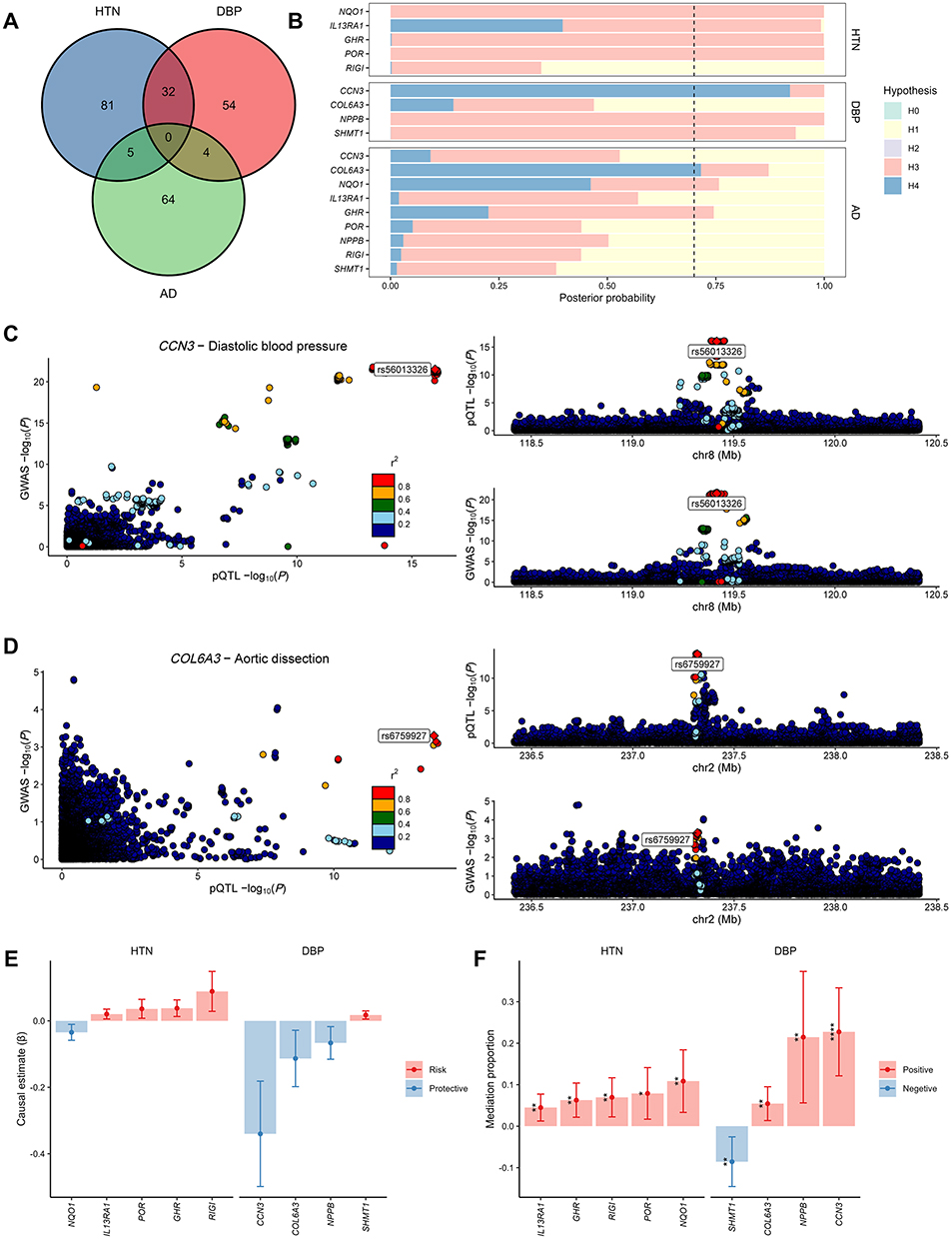 |
Figure 3 Colocalization and mediation analysis of causal proteins. (A) Venn diagram of the causal proteins between BP traits and AD. (B) Posterior probability of colocalization hypotheses on the overlapped causal proteins to BP traits and AD. The black dashed line represents the positive threshold (70%) for the posterior probability of hypothesis 4. (C) Combined comparing and visualizing regional signals plots between CCN3 cis-pQTL and DBP GWAS data. (D) Combined comparing and visualizing regional signals plots between COL6A3 cis-pQTL and AD GWAS data. (E) Mediation effects of overlapped causal proteins on AD mediated by BP traits. (F) Mediation proportion of overlapped causal proteins on AD mediated by BP traits (* P<0.05, ** P<0.01, *** P<0.001, **** P<0.0001). Abbreviations: BP, blood pressure; AD, aortic dissection; pQTL, protein quantitative trait loci; DBP, diastolic blood pressure; GWAS, genome-wide association study. |
Interestingly, previous studies have shown that increased levels of COL6A3 and its carboxyl terminus product are associated with a higher risk of coronary artery diseases, which is contrary to our results.19,20 Therefore, we conducted MR and colocalization analyses between COL6A3 and various CVDs. The results revealed that COL6A3 acts as a protective factor for aortic diseases with evidence of colocalization, while it is a risk factor for coronary artery diseases (Figure S1).
Mediation Effect of BP Traits on Protein-Associated AD
Among the 5 proteins associated with both HTN and AD, 5 mediation pathways were identified, in which the direction of mediating effect and total effect remained consistent. Specifically, 10.86% ([95% CI, 3.33–18.40%], P = 4.73×10−3) of the effect of NQO1 on AD may be mediated by HTN (Figure 3E–F and Table S12). Among the 4 proteins associated with both DBP and AD, 3 mediation pathways were identified. DBP explained 22.74% ([95% CI, 12.14–33.35%], P = 2.63×10−5), 5.43% ([95% CI, 1.36–9.51%], P = 8.93×10−3), and 21.47% ([95% CI, 5.62–37.31%], P = 7.94×10−3) of the effect of CCN3, COL6A3, and NPPB on AD, respectively (Figure 3E–F and Table S12).
Replication Analyses in MVP Cohort
We replicated MR analyses in MVP study and found that HTN and DBP also exhibited a risk effect on AD, while there was still no significant causal link found between SBP and AD (Figure 4A and Table S13). The proteome-wide MR identified 235 plasma proteins associated with HTN or DBP (150 with HTN, 147 with DBP, and 62 overlaps) (Figure 4B and Tables S14–S15). The biological processes and pathways causal proteins enriched in were almost consistent with the results of the discovery cohort, which emphasizes the importance of these progressions in the pathogenesis of BP traits and AD (Figure 4C and Tables S16–17 Integrating the MR results of the discovery and replication cohorts, 1 plasma protein (NQO1) was causally associated with AD via HTN, and 3 plasma proteins (CCN3, COL6A3, and NPPB) were causally associated with AD via DBP (Figure 4D–E), among which CCN3 (PPH4 = 61.76%) and COL6A3 (PPH4 = 69.53%) showed suggestive colocalization with DBP (Figure 4F and Table S18). Mediation analysis showed high consistency in both cohorts. Specifically, HTN could mediate the effects of NQO1 on AD, with mediation proportions of 12.04%. DBP could mediate the effects of CCN3, COL6A3, and NPPB on AD, with mediation proportions ranging from 10.22% to 26.24% (Figure 4G and Table S19).
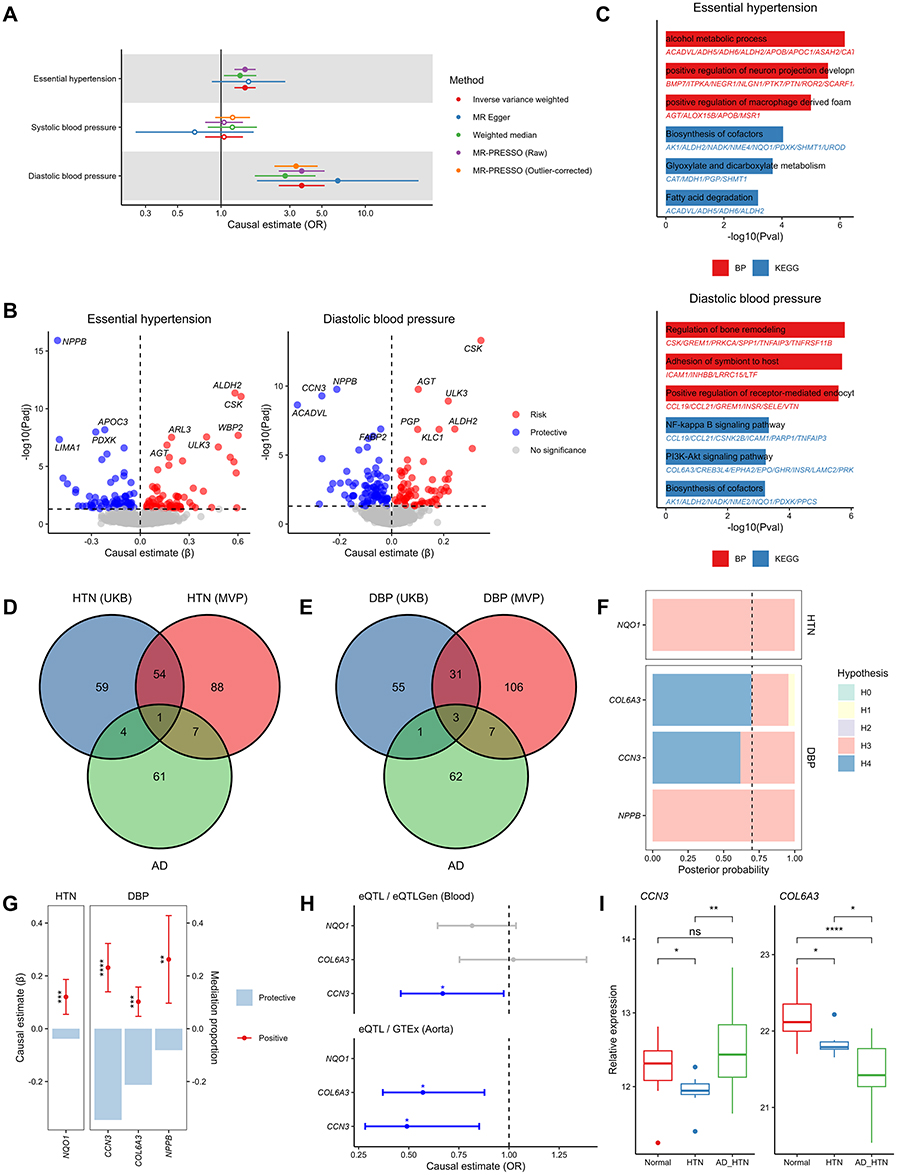 |
Figure 4 Replication analyses in MVP cohort and validation analyses. (A) Causal association between BP traits and AD. (B) Causal effect of genetically predicted plasma proteins on BP traits. (C) Functional enrichment analysis of causal plasma proteins of BP traits. (D) Venn diagram of the causal proteins between HTN in both cohorts and AD. (E) Venn diagram of the causal proteins between DBP in both cohorts and AD. (F) Posterior probability of colocalization hypotheses on the candidate causal proteins to BP traits. The black dashed line represents the positive threshold (70%) for the posterior probability of hypothesis 4. (G) Mediation effects (left y-axis) and proportion (right y-axis) of candidate causal proteins on AD mediated by BP traits. (H) Causal association between tissue specific genes encoding candidate causal proteins and AD (* P<0.05). (I) The relative expression differences of candidate causal proteins among Normal, HTN, and AD_HTN group in our plasma proteomic sequencing (*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). Abbreviations: MVP, Million Veteran Program; BP, blood pressure; AD, aortic dissection; HTN, hypertension; DBP, diastolic blood pressure; AD_HTN, aortic dissection with history of hypertension. |
Tissue Specific eQTL-Based MR
Summary-level cis-eQTL statistics of whole blood and aorta were respectively retrieved from the eQTLGen Consortium and GTEx Portal, aiming to investigate whether genetically predicted tissue-specific gene expression levels have a causal relationship with AD. Owing to discrepancies between transcriptomic and proteomic detection methodologies, NPPB was not found in the two eQTL cohorts. Two-sample MR results demonstrated that the expression levels of CCN3 in both whole blood and aorta, and COL6A3 in aorta, showed a protective effect against AD (Figure 4H and Table S20). These finding suggests that CCN3 and COL6A3 prevent AD at both transcriptional and translational levels, serving as potential robust therapeutic targets for AD.
Plasma Proteomics
We collected blood samples from 7 patients with HTN (HTN group), 19 patients with AD and history of HTN (AD_HTN group), and 11 healthy controls (Normal group), followed by plasma proteomic sequencing. After quality control, 2,174 plasma proteins were finally identified. Due to the diversity in sequencing platforms, methodologies, and sequencing depths between our study and the deCODE genetics, only CCN3 and COL6A3 were detected in our plasma proteome. Compared with Normal group, CCN3 in HTN group was significantly decreased. Whereas compared with HTN group, CCN3 in AD_HTN group was significantly increased. However, no significant difference was observed in CCN3 between Normal and AD_HTN group. The levels of COL6A3 showed a stepwise significant decrease across Normal, HTN, and AD_HTN group, indicating that the reduction of protective factor COL6A3 plays an important role in the progression from HTN to AD (Figure 4I).
Single-Cell RNA-Seq Analyses
After quality control of the scRNA-seq data of acute Stanford type A AD from GSE213740, we implemented a standard data process pipeline (Figure 5A and B). Based on the established aortic cell-specific markers reported in previous literature, we annotated 16 cell clusters into 8 distinct cell types, including smooth muscle cells (SMC), fibroblasts (FIB), endothelial cells (EC), Schwann cells, macrophages, T cells, B cells, and proliferating cells (Figure 5C and D). The top 5 differentially expressed genes of each cell type were consistent with the corresponding cell identity (Figure 5E). Cell-cell communication analysis revealed that COL6A3, which is expressed by SMC and FIB, engages in extensive cell-cell crosstalk with various cell types. Among these, the ligand-receptor pair consisting of COL6A3 derived from FIB and CD44 derived from macrophages exhibited the highest communication probability, suggesting that the cell-cell communication mediated by this pair may play a role in the pathological progression of AD (Figure 5F). CCN3 was predominantly expressed in SMC, NPPB showed low expression levels in aortic tissues, and NQO1 was mainly expressed in SMC, FIB, and EC (Figure 5G).
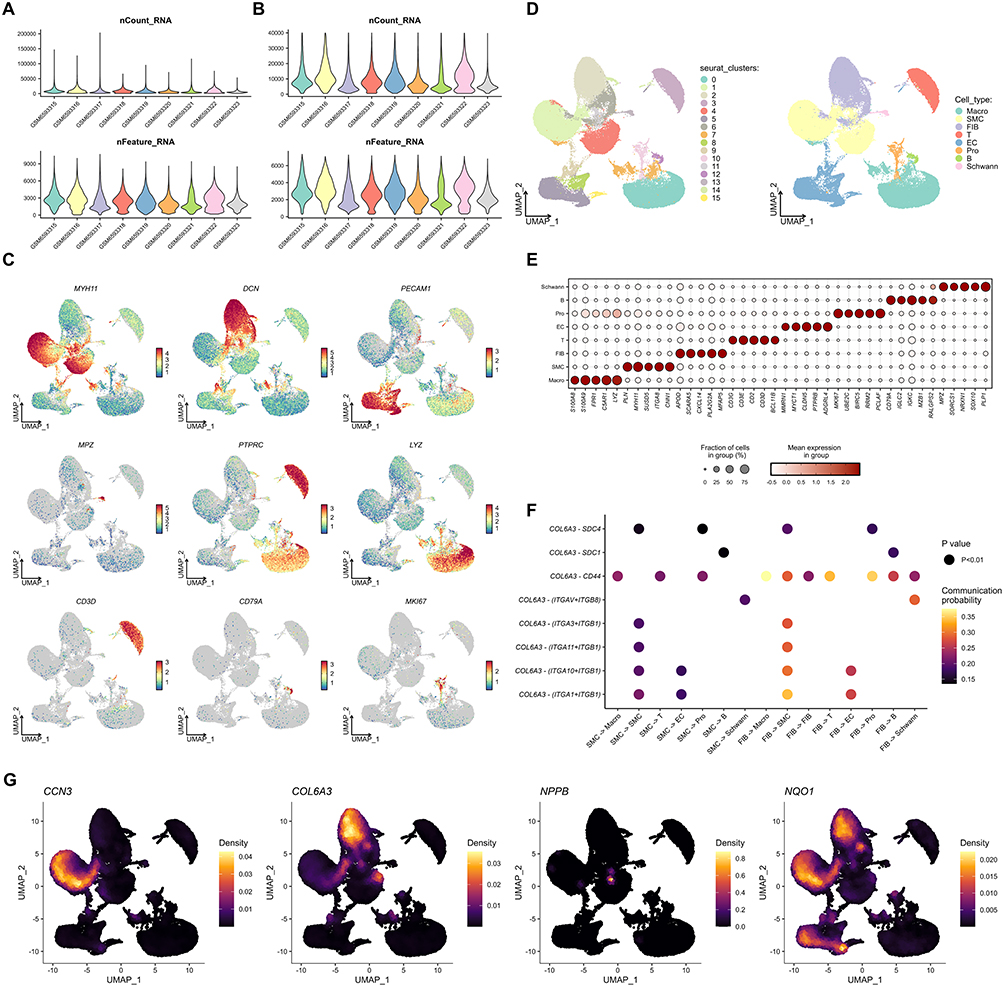 |
Figure 5 Single-cell RNA-seq analyses. (A) Number of counts and features of each sample before quality control. (B) Number of counts and features of each sample after quality control. (C) UMAP plots of expression distribution of the aortic cell-specific markers. (D) UMAP plots of 16 cell clusters and annotated 8 cell types. (E) Top 5 DEGs of each cell type. (F) Cell-cell communication in AD mediated by receptor-ligand pairs involving COL6A3. (G) Distribution density UMAP plots of genes encoding candidate causal proteins in AD cells. Abbreviations: UMAP, uniform manifold approximation and projection; DEG, differentially expressed genes; AD, aortic dissection. |
Drug Target Analysis
We searched and evaluated the druggability and drug development status of candidate proteins in the Therapeutic Target Database, Drug-Gene Interaction Database, DrugBank, and Open Targets Platform (Tables S21–S24). CCN3 is one of the targets of insulin human. Ocriplasmin, which is used to treat symptomatic vitreomacular adhesion, and collagenase Clostridium histolyticum, which is used to treat Dupuytren’s contracture and Peyronie’s disease, are drugs targeting COL6A3. Carvedilol, which targets NPPB, is widely used in the treatment of CVDs such as HTN, and the B-type natriuretic peptide analogue BD-NP is also under investigation. NQO1 is associated with beta-lapachone analogue, vitamin E analogue, and coenzyme Q10 analogue. Beta-lapachone analogue can promote NQO1-mediated programmed tumor cell necrosis, which is used in the treatment of solid tumors. Vitamin E analogue and coenzyme Q10 analogue act as effective cellular oxidative stress protectants. Drugs targeting these proteins hold significant potential for research in the treatment of aortic dissection.
Phenome-Wide Association Study Analysis
We conducted a PheWAS analysis on candidate proteins using the AstraZeneca PheWAS Portal. No gene-level results were available for CCN3 (also known as NOV). For the remaining proteins, none was significantly associated (P < 1×10−6/1×10−8) with other phenotypes as risk factor (OR > 1 or β > 0), indicating that no detrimental or adverse effects were examined in other binary or continuous phenotypes according to the gene-level results (Figure 6).
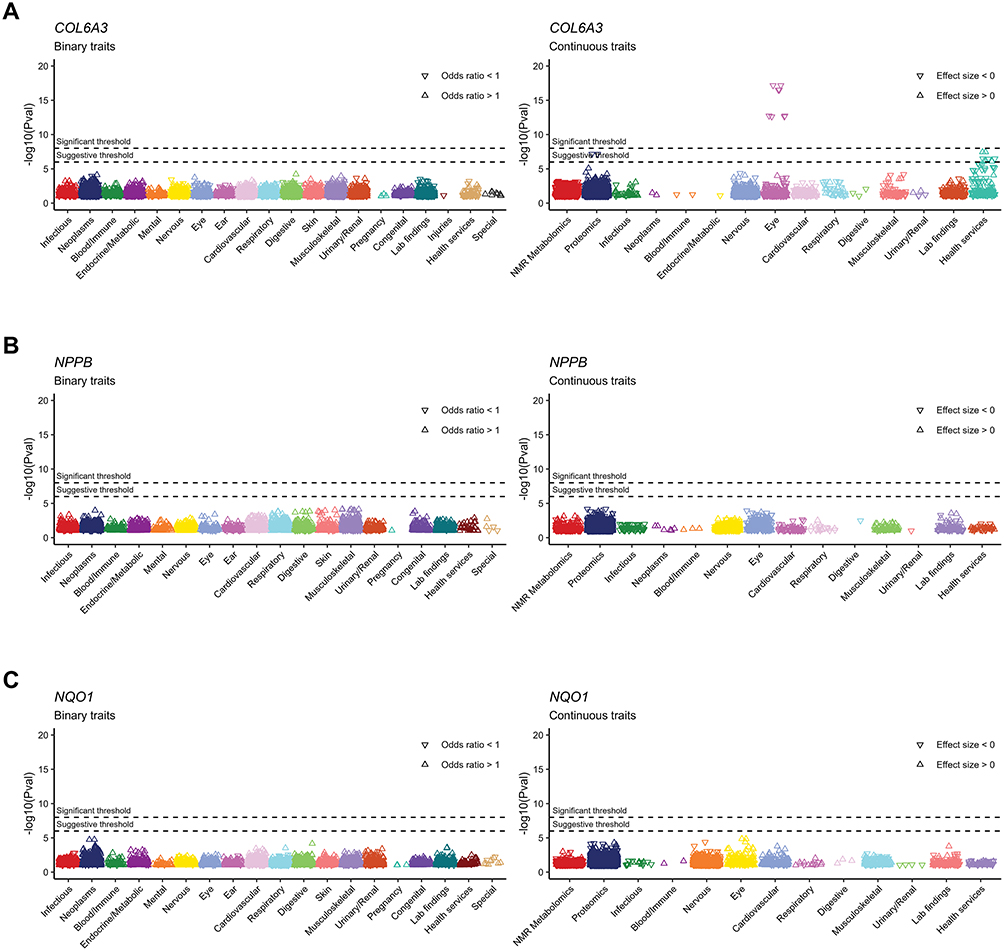 |
Figure 6 Phenome-wide association study analysis on candidate causal proteins and both binary and continuous traits. (A) COL6A3. (B) NPPB. (C) NQO1. |
Discussion
Our network MR identified CCN3, COL6A3, NPPB and NQO1 as key plasma proteins causally associated with both BP traits and AD. CCN3 exhibited protective effects against AD with DBP serving as its mediator (mediation proportion = 22.74%) and strong colocalization with DBP (PPH4 = 92.10% in discovery, 61.76% in replication). COL6A3 showed colocalization with AD (PPH4 = 71.60%) and suggestive colocalization with DBP (PPH4 = 69.54% in replication), and was mediated by DBP in its protective effect on AD (mediation proportion = 5.43%). NPPB and NQO1 prevent AD via DBP and HTN, respectively, with similar mediation effects observed across cohorts. Analyses of our plasma proteomic sequencing and scRNA-seq from public databases suggested that COL6A3 may serve as an important trigger in the development and progression of HTN and subsequent AD. Drug database mining revealed potential therapeutics targeting these proteins, and PheWAS analysis demonstrated that these proteins, which serve as protective factors, were not significantly associated with other phenotypes as risk factors, suggesting that therapeutic strategies targeting these proteins may possess favorable safety profiles.
HTN is a key risk factor for AD, as confirmed by research. A study of over 276,000 participants identified HTN as an independent risk factor for aortic-related deaths, with critical BP thresholds at 130/82 mmHg.21 Studies indicate DBP has a more direct impact on AD-related mortality, especially in individuals aged 50–75.21,22 Research by Rapsomaniki E et al found that DBP, rather than SBP, is a risk factor for AA in those over 60.23 A meta-analysis confirmed DBP ≥ 80 mmHg as a risk factor for AA, with better prognostic value than SBP.24 Our study provides evidence that HTN and DBP are causal risk factors for AD, whereas no causal association was identified between SBP and AD. These results suggest that DBP could play a more crucial part in preventing AD.
CCN3 is a cytokine that regulates the extracellular matrix and offers vascular protection by inhibiting vascular SMC migration and proliferation, reducing neointimal thickening, and protecting EC.25 It also has anti-inflammatory effects by downregulating NF-κB activity and suppressing inflammation-related molecules like MMP9 and MMP2.26,27 These actions are relevant to HTN-related changes such as endothelial inflammation and vascular thickening.28–30 CCN3 is significantly reduced in AD and AA, and its deficiency in animal studies leads to vascular issues, highlighting its protective role against AA.31,32 The study shows that over 20% of CCN3’s effect on AD is mediated by DBP, with strong colocalization observed in two independent cohorts, suggesting it may be a key molecule linking hemodynamic stress to vascular integrity. Our plasma proteomic sequencing analysis revealed decreased expression of CCN3 in patients with HTN, whereas an upward trend was observed in those with AD. This finding is not entirely consistent with previous reports indicating reduced CCN3 levels in AD. We speculate that the elevated CCN3 observed in AD patients may represent a compensatory or stress-induced response triggered following the acute onset of the disease. The specific underlying mechanisms and pathophysiological implications warrant further investigation.
COL6A3, a collagen family member, is vital for the extracellular matrix and cell interactions. It is associated with early-onset HTN in the Japanese population, suggesting a role in BP regulation.33 Reduced circulating COL6A3 levels were identified as a key risk factor for AA, likely due to their role in compromising vascular wall stability through disruptions in lipid metabolism, immune responses, and matrix remodeling.34 Hypertensive patients have lower serum endotrophin, a degradation product of COL6A3, compared to healthy individuals, indicating reduced aortic elasticity and increased stiffness, key features of HTN linked to negative cardiovascular outcomes.35 COL6A3 is also associated with coronary artery disease risk, and serum endotrophin correlates with higher cardiovascular event and mortality risk in atherosclerotic patients.19,20,36 COL6A3 consistently showed protective effects against aortic conditions like AD and AA, with confirmed genetic colocalization of COL6A3 with AA and AD. In contrast, it was associated with an increased risk of atherosclerotic diseases such as coronary artery disease. This indicates that COL6A3 may have different roles in various vascular areas, likely through distinct biological pathways related to each disease’s pathophysiology. Our plasma proteomic sequencing analysis validated the stepwise significant low expression of COL6A3 in the serum of patients with HTN and AD. Single-cell transcriptomic analysis revealed that COL6A3 is predominantly expressed in FIB. The potential COL6A3-CD44 receptor-ligand pair between FIB and macrophages indicates intercellular communication. CD44, a multifunctional receptor on macrophages, profoundly influences macrophage’s behavior in various physiological and pathological contexts. It mediates interactions between macrophages and the extracellular matrix as well as other cells, affecting macrophage migration, localization, polarization (particularly towards the M2 phenotype), initiation, and progression of inflammatory responses, and contributing to immunosuppression and chemotherapy resistance in the tumor microenvironment.37–39 These findings collectively suggest that COL6A3 serves as a pivotal molecular hub connecting mechanical integrity with the immune microenvironment.
NPPB is a prohormone secreted by ventricular cardiomyocytes, released in response to increased ventricular load (often associated with elevated blood volume). Its core functions include lowering BP, promoting diuresis, and inhibiting sodium and water reabsorption, making it a key substance in maintaining cardiovascular homeostasis.40 Consequently, the natriuretic peptide system may become a major therapeutic target for various related diseases and for reducing HTN and metabolism-related cardiovascular risks in the future.41 However, in pathological states, elevated levels of NPPB serve as a risk marker. In hypertensive patients, it reflects cardiac stress due to sustained high pressure, indicating early cardiac involvement;42 in acute AD, its elevation is associated with a sharp increase in right heart load, acting as a strong predictor of short-term mortality risk (with mortality potentially increasing up to fourfold in high-risk individuals).40 Furthermore, it correlates positively with aortic diameter and holds significant prognostic value.43,44 Interestingly, our study suggests that genetically determined higher levels of NPPB may be a protective factor against HTN, with approximately 20% of its effect on AD being mediated by DBP. This highlights the complexity of the system. While the natriuretic peptide pathway is inherently protective, its sustained elevation in disease essentially reflects the body’s challenging transition from compensation to decompensation, thereby transforming it into a signal of risk.
NQO1 exerts its antihypertensive effects through two core mechanisms. First, by elevating NAD+ levels, it activates SIRT1, leading to the deacetylation and activation of eNOS, which collectively improves endothelial function and promotes vasodilation.45,46 Second, as a key target gene of Nrf2, NQO1 constitutes a crucial antioxidant defense pathway, scavenging reactive oxygen species to mitigate oxidative damage in blood vessels and kidneys, and its downregulation in pathological conditions such as obesity and chronic kidney disease exacerbates oxidative stress and the progression of hypertension.47,48 Consistent with previous research, our study identifies NQO1 as a protective factor against both hypertension and aortic dissection.49,50
The intersections of biological processes and pathways enriched by the causal proteins for BP traits and AD highlight the roles of proteins in interconnected pathways. Key pathways include renal sodium excretion, crucial for fluid balance and BP regulation, whose dysfunction can trigger HTN and cardiovascular issues like AD.51,52 Cell junction assembly is vital for aortic wall integrity, and changes in adhesion genes and excessive calpain activation can reduce vascular SMC contractility and disrupt endothelial function.53,54 Angiogenesis regulation is crucial, as inhibitors of angiogenesis can raise AD risk.55,56 Inflammation and immune regulation are crucial, with phagocytosis preventing thoracic AD by clearing apoptotic cells.57 Leukocyte proliferation regulation plays a pivotal role in cardiovascular inflammation. Diseases such as HTN can lead to endothelial dysfunction in the bone marrow and increase inflammatory myeloid cells.58 In summary, these enriched biological processes and pathways may collectively constitute a dynamic pathological network that drives the development of HTN and AD.
Large samples from four major biobanks offer high statistical power, however, our study had some limitations. First, the GWAS datasets used in this study are predominantly from participants of European ancestry. While this reduces confounding from population stratification, differences in allele frequencies, linkage disequilibrium structure, and genetic diversity may limit the generalizability of our findings to other populations. Though we performed plasma proteomic validation in an East Asian cohort, this cohort was relatively small and did not involve whole-genome sequencing, failing to link phenotypes to genetic architecture. The validation would be strongly influenced by confounding factors such as diet, environment, and comorbidities, making it inadequate to fully validate our MR results. Future studies should conduct cross-ancestry validation in diverse large non-European cohorts to improve the applicability of our findings. Second, despite leveraging one of the most extensive proteome-wide GWAS resources, some proteins lacked significant instrumental variables (IVs), which restricted the exploration of their causal associations and potential therapeutic value. Additionally, heterogeneity and pleiotropy could not be effectively assessed. This requires GWAS data with larger scale and greater sequencing depth to identify more significant IVs. Third, although this study suggested causal associations and pathways linking plasma proteins, BP, and AD, further experimental researches are needed to elucidate the underlying molecular mechanisms. Future research could focus on constructing relevant models to explore the pathophysiological processes of specific proteins in maintaining vascular homeostasis and the development of AD, and evaluating the potential of targeting these proteins for the prevention or treatment of AD, with the aim of achieving early identification and stratified management of AD.
Conclusion
In summary, this study mapped how plasma proteins affect AD risk, highlighting BP’s key role. We identified new biomarkers and targets like CCN3, COL6A3, NPPB and NQO1. Subsequent investigations should seek to validate these findings among diverse populations and examine the molecular mechanisms of these proteins and pathways. These insights aim to advance precise prevention and targeted treatment of HTN and AD.
Abbreviations
AA, aortic aneurysm; AD, aortic dissection; BH, Benjamini-Hochberg; BP, blood pressure; CTA, computed tomography angiography; CVD, cardiovascular disease; DBP, diastolic blood pressure; EC, endothelial cell; eQTL, expression quantitative trait loci; FDR, false discovery rate; FIB, fibroblast; GTEx, Genotype-Tissue Expression; GWAS, genome-wide association study; HTN, hypertension; IV, instrumental variable; MR, Mendelian randomization; MVP, Million Veteran Program; PheWAS, phenome-wide association study; pQTL, protein quantitative trait loci; SBP, systolic blood pressure; scRNA-seq, single cell RNA sequencing; SMC, smooth muscle cell; UKB, UK Biobank.
Data Sharing Statement
The GWAS data used in this study are publicly available. The plasma proteomic sequencing data is available from the corresponding author on reasonable request.
Ethics Approval and Informed Consent
This study adhered to the STROBE-MR guidelines, and was approved by the Ethics Committee of the First Affiliated Hospital of Chongqing Medical University (No. 2024-397-01), and followed the guidelines stated in the Helsinki Declaration. Written informed consent was obtained from all participants.
Acknowledgments
The availability of GWAS summary statistics from the deCODE Genetics, UK Biobank, Million Veteran Program, FinnGen study, eQTLGen consortium, and GTEx Portal greatly facilitated this work. The authors thank all the participants and researchers who contributed to these publicly available resources.
Author Contributions
Zhenghao Li: Conceptualization, Data curation, Formal analysis, Visualization, Writing – original draft, Writing – review and editing. Jian Liu: Formal analysis, Visualization. Xiaoxi Fan: Data curation, Validation. Liangtao Shi: Data curation, Validation. Haoming Shi: Conceptualization, Writing – review and editing. Changying Li: Data curation, Validation. Changzhu Duan: Conceptualization, Resources, Supervision, Writing – review and editing. Cheng Zhang: Conceptualization, Resources, Supervision, Funding acquisition, Writing – review and editing. Qingchen Wu: Conceptualization, Resources, Supervision, Funding acquisition, Writing – review and editing. Yue Shao: Conceptualization, Writing – original draft, Writing – review and editing. All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from the National Natural Science Foundation of China (82270506).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Carrel T, Sundt TM, Von Kodolitsch Y, Czerny M. Acute aortic dissection. Lancet. 2023;401(10378):773–16. doi:10.1016/s0140-6736(22)01970-5
2. Evangelista A, Isselbacher EM, Bossone E, et al. Insights from the international registry of acute aortic dissection: a 20-year experience of collaborative clinical research. Circulation. 2018;137(17):1846–1860. doi:10.1161/circulationaha.117.031264
3. Hibino M, Otaki Y, Kobeissi E, et al. Blood pressure, hypertension, and the risk of aortic dissection incidence and mortality: results from the J-SCH study, the UK Biobank Study, and a meta-analysis of cohort studies. Circulation. 2022;145(9):633–644. doi:10.1161/circulationaha.121.056546
4. Bossone E, Eagle KA. Epidemiology and management of aortic disease: aortic aneurysms and acute aortic syndromes. Nat Rev Cardiol. 2021;18(5):331–348. doi:10.1038/s41569-020-00472-6
5. Gawinecka J, Schönrath F, von Eckardstein A. Acute aortic dissection: pathogenesis, risk factors and diagnosis. Swiss Med Wkly. 2017;
6. Wei Q, Li K, Su L, et al. Plasma proteomics implicate glutamic oxaloacetic transaminases as potential markers for acute myocardial infarction. J Proteomics. 2024;308:105286. doi:10.1016/j.jprot.2024.105286
7. Li M, He X, Gong W, et al. Plasma proteomics profiles predict the risk of future aortic aneurysm and aortic dissection. Int J Surg. 2025;111(10):6894–6904. doi:10.1097/js9.0000000000002845
8. Ho FK, Mark PB, Lees JS, et al. A proteomics-based approach for prediction of different cardiovascular diseases and dementia. Circulation. 2025;151(5):277–287. doi:10.1161/circulationaha.124.070454
9. Qiu S, Liu Z, Jiang WD, et al. Diabetes and aortic dissection: unraveling the role of 3-hydroxybutyrate through Mendelian randomization. Cardiovasc Diabetol. 2024;23(1):159. doi:10.1186/s12933-024-02266-3
10. Liu W, Yang C, Lei F, et al. Major lipids and lipoprotein levels and risk of blood pressure elevation: a Mendelian Randomisation Study. EBioMedicine. 2024;100:104964. doi:10.1016/j.ebiom.2023.104964
11. Liu Y, Wang H, Yu M, et al. Hypertriglyceridemia as a key contributor to abdominal aortic aneurysm development and rupture: insights from genetic and experimental models. Circulation. 2025;152(12):862–881. doi:10.1161/circulationaha.125.074737
12. Ferkingstad E, Sulem P, Atlason BA, et al. Large-scale integration of the plasma proteome with genetics and disease. Nat Genet. 2021;53(12):1712–1721. doi:10.1038/s41588-021-00978-w
13. Cerezo M, Sollis E, Ji Y, et al. The NHGRI-EBI GWAS Catalog: standards for reusability, sustainability and diversity. Nucleic Acids Res. 2025;53(D1):D998–d1005. doi:10.1093/nar/gkae1070
14. UK Biobank Whole-Genome Sequencing Consortium. Whole-genome sequencing of 490,640 UK Biobank participants. Nature. 2025;645(8081):692–701. doi:10.1038/s41586-025-09272-9
15. Verma A, Huffman JE, Rodriguez A, et al. Diversity and scale: genetic architecture of 2068 traits in the VA million veteran program. Science. 2024;385(6706):eadj1182. doi:10.1126/science.adj1182
16. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613(7944):508–518. doi:10.1038/s41586-022-05473-8
17. Võsa U, Claringbould A, Westra HJ, et al. Large-scale cis- and trans-eQTL analyses identify thousands of genetic loci and polygenic scores that regulate blood gene expression. Nat Genet. 2021;53(9):1300–1310. doi:10.1038/s41588-021-00913-z
18. Center BS. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science. 2020;369(6509):1318–1330. doi:10.1126/science.aaz1776
19. Yoshiji S, Lu T, Butler-Laporte G, et al. Integrative proteogenomic analysis identifies COL6A3-derived endotrophin as a mediator of the effect of obesity on coronary artery disease. Nat Genet. 2025;57(2):345–357. doi:10.1038/s41588-024-02052-7
20. Fan M, Li N, Huang L, Chen C, Dong X, Gao W. Exploring potential drug targets in multiple cardiovascular diseases: a study based on proteome-wide Mendelian randomization and colocalization analysis. Cardiovasc Ther. 2025;2025:5711316. doi:10.1155/cdr/5711316
21. Otaki Y, Watanabe T, Konta T, et al. Effect of hypertension on aortic artery disease-related mortality - 3.8-Year nationwide community-based prospective cohort study. Circ J. 2018;82(11):2776–2782. doi:10.1253/circj.CJ-18-0721
22. Otaki Y, Watanabe T, Konta T, et al. One-year change in diastolic blood pressure and aortic disease-related mortality in a Japanese general population aged 50-75 years. Circ J. 2021;85(12):2222–2231. doi:10.1253/circj.CJ-21-0514
23. Rapsomaniki E, Timmis A, George J, et al. Blood pressure and incidence of twelve cardiovascular diseases: lifetime risks, healthy life-years lost, and age-specific associations in 1·25 million people. Lancet. 2014;383(9932):1899–1911. doi:10.1016/s0140-6736(14)60685-1
24. Kobeissi E, Hibino M, Pan H, Aune D. Blood pressure, hypertension and the risk of abdominal aortic aneurysms: a systematic review and meta-analysis of cohort studies. Eur J Epidemiol. 2019;34(6):547–555. doi:10.1007/s10654-019-00510-9
25. Shimoyama T, Hiraoka S, Takemoto M, et al. CCN3 inhibits neointimal hyperplasia through modulation of smooth muscle cell growth and migration. Arterioscler Thromb Vasc Biol. 2010;30(4):675–682. doi:10.1161/atvbaha.110.203356
26. Lin Z, Natesan V, Shi H, et al. A novel role of CCN3 in regulating endothelial inflammation. J Cell Commun Signal. 2010;4(3):141–153. doi:10.1007/s12079-010-0095-x
27. Janune D, Kubota S, Nishida T, et al. Novel effects of CCN3 that may direct the differentiation of chondrocytes. FEBS Lett. 2011;585(19):3033–3040. doi:10.1016/j.febslet.2011.08.024
28. Dikalova A, Fehrenbach D, Mayorov V, et al. Mitochondrial CypD acetylation promotes endothelial dysfunction and hypertension. Circ Res. 2024;134(11):1451–1464. doi:10.1161/circresaha.123.323596
29. Naderi-Meshkin H, Setyaningsih WAW. Endothelial cell dysfunction: onset, progression, and consequences. Front Biosci. 2024;29(6):223. doi:10.31083/j.fbl2906223
30. Aboukhater D, Morad B, Nasrallah N, et al. Inflammation and hypertension: underlying mechanisms and emerging understandings. J Cell Physiol. 2023;238(6):1148–1159. doi:10.1002/jcp.31019
31. Ran H, Li C, Rizvi SMM, et al. Integrated analyses of Mendelian randomization, eQTL, and single-cell transcriptome identify CCN3 as a potential biomarker in aortic dissection. Sci Rep. 2024;14(1):32062. doi:10.1038/s41598-024-83611-0
32. Zhang C, van der Voort D, Shi H, et al. Matricellular protein CCN3 mitigates abdominal aortic aneurysm. J Clin Invest. 2016;126(4):1282–1299. doi:10.1172/jci82337
33. Yamada Y, Kato K, Oguri M, et al. Identification of 13 novel susceptibility loci for early-onset myocardial infarction, hypertension, or chronic kidney disease. Int J Mol Med. 2018;42(5):2415–2436. doi:10.3892/ijmm.2018.3852
34. Zhang K, Liu Y, Mao A, Li C, Geng L, Kan H. Proteome-wide Mendelian randomization identifies therapeutic targets for abdominal aortic aneurysm. J Am Heart Assoc. 2025;14(3):e038193. doi:10.1161/jaha.124.038193
35. Baykiz D, Demirtakan ZG, Govdeli EA, et al. Endotrophin and matrix metalloproteinase-2 levels in bicuspid aortic valve and hypertension associated aortopathy and their relationship with strain parameters of the ascending aorta. Eur Rev Med Pharmacol Sci. 2023;27(6):2358–2376. doi:10.26355/eurrev_202303_31771
36. Holm Nielsen S, Edsfeldt A, Tengryd C, et al. The novel collagen matrikine, endotrophin, is associated with mortality and cardiovascular events in patients with atherosclerosis. J Intern Med. 2021;290(1):179–189. doi:10.1111/joim.13253
37. Wang C, Li Y, Wang L, et al. SPP1 represents a therapeutic target that promotes the progression of oesophageal squamous cell carcinoma by driving M2 macrophage infiltration. Br J Cancer. 2024;130(11):1770–1782. doi:10.1038/s41416-024-02683-x
38. Dai Y, Ji Z, Liang H, et al. CD44v5 domain regulates crosstalk between TNBC cells and tumor-associated macrophages by enhancing the IL-4R/STAT3 axis. Cancer Sci. 2024;115(7):2235–2253. doi:10.1111/cas.16200
39. Kim SH, Kim CH, Lee CH, et al. Glycoengineered stem cell-derived extracellular vesicles for targeted therapy of acute kidney injury. Biomaterials. 2025;318:123165. doi:10.1016/j.biomaterials.2025.123165
40. Vrsalovic M, Vrsalovic Presecki A, Aboyans V. N-terminal pro-brain natriuretic peptide and short-term mortality in acute aortic dissection: a meta-analysis. Clin Cardiol. 2020;43(11):1255–1259. doi:10.1002/clc.23436
41. Sarzani R, Spannella F, Giulietti F, Balietti P, Cocci G, Bordicchia M. Cardiac natriuretic peptides, hypertension and cardiovascular risk. High Blood Press Cardiovasc Prev. 2017;24(2):115–126. doi:10.1007/s40292-017-0196-1
42. Rosół N, Procyk G, Kacperczyk-Bartnik J, Grabowski M, Gąsecka A. N-terminal prohormone of brain natriuretic peptide in gestational hypertension and preeclampsia - State of the art. Eur J Obstet Gynecol Reprod Biol. 2024;297:96–105. doi:10.1016/j.ejogrb.2024.04.007
43. Wen D, Jia P, Du X, Dong JZ, Ma CS. Value of N-terminal pro-brain natriuretic peptide and aortic diameter in predicting in-hospital mortality in acute aortic dissection. Cytokine. 2019;119:90–94. doi:10.1016/j.cyto.2019.03.004
44. Gutin LS, Bakalov VK, Rosing DR, Arai AE, Gharib AM, Bondy CA. N-terminal pro-brain natriuretic peptide levels and aortic diameters. Am Heart J. 2012;164(3):419–424. doi:10.1016/j.ahj.2012.06.021
45. Kim YH, Hwang JH, Noh JR, et al. Activation of NAD(P)H:quinone oxidoreductase ameliorates spontaneous hypertension in an animal model via modulation of eNOS activity. Cardiovasc Res. 2011;91(3):519–527. doi:10.1093/cvr/cvr110
46. Kim YH, Hwang JH, Kim KS, et al. NQO1 activation regulates angiotensin-converting enzyme shedding in spontaneously hypertensive rats. Cardiovasc Res. 2013;99(4):743–750. doi:10.1093/cvr/cvt147
47. Farooqui Z, Mohammad RS, Lokhandwala MF, Banday AA. Nrf2 inhibition induces oxidative stress, renal inflammation and hypertension in mice. Clin Exp Hypertens. 2021;43(2):175–180. doi:10.1080/10641963.2020.1836191
48. Sarkar P, Pecorelli A, Woodby B, et al. Evaluation of anti-oxinflammatory and ACE-Inhibitory properties of protein hydrolysates obtained from edible non-mulberry silkworm pupae (Antheraea assama and philosomia ricinii). Nutrients. 2023;15(4). doi:10.3390/nu15041035
49. Zou J, Zheng Z, Ye W, et al. Targeting the smooth muscle cell KEAP1-Nrf2-STING axis with pterostilbene attenuates abdominal aortic aneurysm. Phytomedicine. 2024;130:155696. doi:10.1016/j.phymed.2024.155696
50. Van Merrienboer TAR, Rombouts KB, Van Wijk A, et al. Proteome differences in smooth muscle cells from diabetic and non-diabetic abdominal aortic aneurysm patients reveal metformin-induced mechanisms. Med Sci. 2025;13(3). doi:10.3390/medsci13030184
51. Buryskova Salajova K, Malik J, Valerianova A. Cardiorenal syndromes and their role in water and sodium homeostasis. Physiol Res. 2024;73(2):173–187. doi:10.33549/physiolres.935110
52. Tsuruda T, Yamashita A, Otsu M, et al. Angiotensin II induces aortic rupture and dissection in osteoprotegerin-deficient mice. J Am Heart Assoc. 2022;11(8):e025336. doi:10.1161/jaha.122.025336
53. Li Y, Gao S, Han Y, et al. Variants of focal adhesion scaffold genes cause thoracic aortic aneurysm. Circ Res. 2021;128(1):8–23. doi:10.1161/circresaha.120.317361
54. Miyazaki T, Miyazaki A. Dysregulation of calpain proteolytic systems underlies degenerative vascular disorders. J Atheroscler Thromb. 2018;25(1):1–15. doi:10.5551/jat.RV17008
55. Tsujinaka K, Izawa-Ishizawa Y, Miyata K, et al. Angiogenesis inhibitor-specific hypertension increases the risk of developing aortic dissection. Biomed Pharmacother. 2023;167:115504. doi:10.1016/j.biopha.2023.115504
56. Yu Y, Su X, Qin Q, et al. Yes-associated protein and transcriptional coactivator with PDZ-binding motif as new targets in cardiovascular diseases. Pharmacol Res. 2020;159:105009. doi:10.1016/j.phrs.2020.105009
57. Yin Z, Zhang J, Zhao M, et al. EDIL3/Del-1 prevents aortic dissection through enhancing internalization and degradation of apoptotic vascular smooth muscle cells. Autophagy. 2024;20(11):2405–2425. doi:10.1080/15548627.2024.2367191
58. Rohde D, Vandoorne K, Lee IH, et al. Bone marrow endothelial dysfunction promotes myeloid cell expansion in cardiovascular disease. Nat Cardiovasc Res. 2022;1(1):28–44. doi:10.1038/s44161-021-00002-8
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Prevalence of Hyperuricemia and the Relationship Between Serum Uric Acid and Hypertension in New Onset Diabetic Patients: A Cross-Sectional Indian Study
Singh SK, Singh R, Singh SK, Iquebal MA, Jaiswal S, Singh A
Diabetes, Metabolic Syndrome and Obesity 2022, 15:1809-1817
Published Date: 14 June 2022
Prevalence of Hypertension and Its Associated Risk Factors Among Patients with Rheumatoid Arthritis in the Kingdom of Saudi Arabia
Al-Ahmari AK
International Journal of General Medicine 2022, 15:6507-6517
Published Date: 8 August 2022
Prevalence of Undiagnosed Hypertension and Its Predictors in Jordan: A Cross-Sectional Study
Alhawari H, AlShelleh S, Alhawari H, AlRyalat SA, Khanfar AN, Alzoubi O
International Journal of General Medicine 2022, 15:7919-7928
Published Date: 25 October 2022
Trends and Comparisons of Blood Pressure and Fasting Plasma Glucose in Patients with Hypertension, Diabetes, and Comorbidity: 4-Year Follow-Up Data
Xu L, Wen X, Yang Y, Cui D
Risk Management and Healthcare Policy 2022, 15:2221-2232
Published Date: 25 November 2022
Perseverative Cognition as a Mediator Between Personality Traits and Blood Pressure
Arroyave-Atehortua D, Cordoba-Sanchez V, Zambrano-Cruz R
Vascular Health and Risk Management 2023, 19:363-370
Published Date: 29 June 2023