Back to Journals » Journal of Pain Research » Volume 19
Causal Brain Network Alterations Across Disease Stages in Herpes Zoster and Postherpetic Neuralgia
Authors Zou W, Li Z, Gu L, Jiang X, Xiong J ![]() , Qin S, Lu J, Jiang J
, Qin S, Lu J, Jiang J
Received 14 August 2025
Accepted for publication 18 January 2026
Published 24 January 2026 Volume 2026:19 560620
DOI https://doi.org/10.2147/JPR.S560620
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor King Hei Stanley Lam
Wanqing Zou,1– 3,* Zihan Li,4,* Lili Gu,5 Xiaofeng Jiang,1– 3 Jiaxin Xiong,1– 3 Suhong Qin,1– 3 Jing Lu,1– 3 Jian Jiang1– 3
1Jiangxi Provincial Key Laboratory for Precision Pathology and Intelligent Diagnosis, Department of Radiology, the First Affiliated Hospital, Jiangxi Medical College, Nanchang University, Nanchang, Jiangxi, 330006, People’s Republic of China; 2Jiangxi Province Medical Imaging Research Institute, Nanchang, Jiangxi, 330006, People’s Republic of China; 3Clinical Research Center for Medical Imaging in Jiangxi Province, Nanchang, Jiangxi, 330006, People’s Republic of China; 4The First Radiology Department of Guangzhou Medical University, Guangzhou, Guangdong, 510163, People’s Republic of China; 5Department of Pain, The First Affiliated Hospital of Nanchang University, Nanchang, 330006, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jian Jiang, Jiangxi Provincial Key Laboratory for Precision Pathology and Intelligent Diagnosis, Department of Radiology, the First Affiliated Hospital, Jiangxi Medical College, Nanchang University, 17 Yongwaizheng Street, Nanchang, Jiangxi, 330006, People’s Republic of China, Tel +86 135 7612 9759, Email [email protected]
Background: Herpes zoster (HZ) and postherpetic neuralgia (PHN), which are common chronic pain disorders, can cause long-term pain and negative emotions accompanied by structural brain changes; however, their temporal dynamics and causal relationships remain unknown.
Purpose: This study employed causal structural covariance network (CaSCN) analysis was used to explore gray matter volume (GMV) alterations across disease stages and their causal relationships. CaSCN refers to directed causal influences between brain regions based on structural covariance.
Patients and Methods: This study employed a cross-sectional observational design. In this study, 157 treatment-naïve first-episode HZ and PHN patients (53 acute HZ, 53 subacute HZ and 52 chronic PHN) were enrolled, along with 85 sex- and age-matched healthy controls (HCs). Voxel-based morphometry (VBM) was applied to analyze high-resolution T1-weighted magnetic resonance images and measure GMV in each participant. On the basis of the results of the intergroup comparisons, the left pericalcarine cortex and bilateral thalamus (voxel-level p < 0.001, cluster-level p < 0.05) presented significant differences and were selected as seed regions for subsequent CaSCN analysis.
Results: Compared with healthy controls, patients with HZ and PHN presented stage-specific GMV changes, and areas such as the left pericalcarine cortex and bilateral thalamus presented GMV changes at the time of onset; causal structure analysis revealed that the left pericalcarine cortex and bilateral thalamus presented significant positive causal effects on the left middle occipital gyrus, left middle temporal gyrus, left angular gyrus, left cerebellum, right inferior temporal gyrus, left medial superior frontal gyrus, left cusai lobe and other brain regions.
Conclusion: This study revealed dynamic patterns of GMV changes over time in HZ and PHN patients by CaSCN analysis, providing new perspectives for understanding the neuroimaging mechanisms of HZ and PHN and clarifying the causal relationships of brain structural alterations during disease progression.
Keywords: causal structural covariance network, gray matter volume, herpes zoster, postherpetic neuralgia, structural covariance network
Introduction
Varicella-zoster virus (VZV) is a ubiquitous, double-stranded DNA alpha herpesvirus that establishes lifelong latency in sensory-ganglion neurons and causes two main human illnesses: primary varicella (chickenpox) and reactivation as herpes zoster (shingles).1 The three phases during HZ reactivation are defined as acute HZ-related pain, subacute HZ-related pain, and PHN.2 The annual incidence of HZ increases annually and increases with age.3 Approximately 5–30% of patients with HZ progress to postherpetic neuralgia (PHN),4 manifested by varying degrees of pain that last for 3 months after the rash heals.5 Patients with PHN are often present with persistent burning, intermittent shock-like pain, and triggering allodynia (hyperalgesia and allodynia).6 These pains often have a significant impact on the patient’s quality of life, sleep, emotional well-being, and may even cause anxiety and depression.7,8 The pathological mechanism of PHN involves a dual disturbance of the peripheral and central nervous systems. Varicella-zoster virus reactivation triggers neuroinflammation and axonal damage, leading to peripheral sensitization; Persistent peripheral nociceptive input induces altered neuronal plasticity in the dorsal horn of the spinal cord and passes through the thalamus-cortical circuits spread to higher brain regions, resulting in “central sensitization”.9,10 The transition to chronic pain involves key brain regions including the thalamus, anterior/posterior cingulate cortex, insula, and prefrontal cortex.11 Alterations in this network underpin persistent pain and associated negative affect. Structural magnetic resonance imaging studies using voxel-based morphometry (VBM) analysis have shown that the gray matter volume (GMV) of PHN patients is abnormally concentrated in pain-related areas (thalamus, anterior cingulate cortex, cerebellum, etc.),12–14 emotion regulation areas (prefrontal cortex, amygdala, etc.),14,15 visual processing and cognitive control areas (talar fissure, cuneiform, temporal lobe, angular gyrus, etc.).12,16 However, existing studies have not distinguished the causal relationship between structural changes in various brain regions, and it is difficult to analyze the temporal diffusion pattern of structural abnormalities. This limitation limits the understanding of the mechanism of PHN neuroimaging.
The structural covariance network (SCN) quantifies the cooperative changes in gray matter morphology indices in brain intervals,17 providing a new perspective for exploring disease-related brain network reorganization. SCN analysis revealed that the brain network of PHN patients showed global efficiency reduction, modular destruction, and dysfunction of pivot nodes (such as thalamus), suggesting that chronic pain may affect multidimensional clinical symptoms by reshaping the topological organization of the brain network.18 However, the traditional SCN can only describe the synchronicity of brain region changes on the basis of zero-time lag correlation and cannot reveal the causal association of these changes. Granger causality analysis provides methodological support to overcome this bottleneck. The core idea is that if the historical information of brain region A can significantly improve the ability to predict future changes in brain region B, then there is a “Granger causal effect” between A and B.19 In patients with PHN, the thalamus-to-anterior cingulate gyrus was found to be enhanced at rest, whereas the flow to descending pain modulation pathways (eg, the prefrontal cortex to the thalamus) was weakened, which may reflect imbalances in central pain modulation mechanisms.20 However, the relationship between brain structural changes and the course of the disease is still unclear, and the causal relationship among gray matter changes is still unclear.
Causal structural covariance networks apply Granger causality analysis to structural data arranged in a specific order (eg, disease progression, duration) and can reflect the causality of brain intersections well. This method, first applied in epilepsy research, reveals the causal relationship between progressive GMV damage between the hippocampus and other extrahippocampal structures in medial temporal lobe epilepsy.21 Subsequently, research using CaSCN analysis has made progress in the understanding of a variety of diseases, including epilepsy22,23 depression,24,25 Alzheimer’s disease26 schizophrenia,27 Parkinson’s disease,28 and so on. These studies demonstrate a causal relationship between the hippocampus, thalamus, and other brain regions, such as the amygdala, prefrontal cortex, and cerebellum, demonstrating the value of CaSCN in exploring disease progression mechanisms and identifying potential intervention targets.
In this study, CaSCN analysis is applied to patients with HZ and PHN, with the aim of determining the initial regions of brain structural changes in HZ and PHN and their diffusion patterns, providing a neuroimaging basis for early disease identification, progression prediction, and targeted intervention. In particular, the identification of key regions may contribute to more precise transcranial stimulation therapy, provide new treatment ideas for such refractory chronic painful diseases, and provide stronger insights into the brain structural networks involved in the development of HZ and PHN diseases.
Materials and Methods
Research Objects
This study employed a cross-sectional observational design. The patients selected for this study were treated for shingles from July 2019 to November 2024 in the pain department of the First Affiliated Hospital of Nanchang University in; these patients were hospitalized for HZ or postherpetic neuralgia PHN while in a treatment-naïve state. In addition, healthy volunteers were recruited from the community and surrounding areas as a control group. Some of these patients were part of a previous study group.20
Inclusion Criteria for Patients
- Right-handed;
- HZ in the acute phase was characterized by herpes, and the duration of herpes disease was longer than 1 month. In the subacute stage, HZ was characterized by herpes lesions, with a duration of longer than 1 month and less than 3 months. Chronic PHN was defined as a herpetic lesion that lasts for longer than 3 months.2
- Diagnosis was made by two attending physicians in the pain department according to the criteria of the International Association for the Study of Pain;29
Exclusion Criteria for Patients
- Having other types of chronic pain or neurological diseases;
- Already diagnosed with mental or psychological illness;
- T2 sequence found to have brain parenchymal lesions;
- Contraindications to MRI (including claustrophobia and ferromagnetic implants in the body).
Inclusion Criteria for Healthy Controls (HCs)
- Right-handed;
- Age- and sex-matched with the included HZ and PHN patients;
- No chronic pain or neurological disease;
- No parenchymal brain lesions found in conventional T2 sequences.
After excluding the subjects with poor image quality, a total of 53 acute HZ patients, 53 subacute HZ patients, 52 chronic PHN patients and 85 HCs were ultimately included.
Basic patient information, including sex, age, disease course on the first day of admission, visual analog scale score, site of onset, and skin lesion characteristics, was collected. The Hamilton Depression Scale30 and the Hamilton Anxiety Scale31 were used. The levels of depression and anxiety of the patients were assessed separately.
The study was conducted in accordance with the approved guidelines and the principles of the Declaration of Helsinki, and all procedures involved in the study were formally approved by the Ethics Committee of the First Affiliated Hospital of Nanchang University (ethics approval code: 20200145). All the participants were recruited from the pain department of the First Affiliated Hospital of Nanchang University, and before they participated in the study, they provided written informed consent.
MRI Data Acquisition
MRI data from all the patients were collected using a Siemens TIM Trio 3.0T (Erlangen, Bavaria, Germany) scanner equipped with the radiology department of our hospital. Three-dimensional, high-resolution T1-weighted imaging was obtained by magnetization preparation of fast gradient echo sequences with the following parameters: pulse repetition time = 1900 ms, pulse echo time = 2.26 ms, flip angle = 9°, field of view = 215 mm × 230 mm, and matrix = 240 × 256 sagittal scans of 176 layers with a thickness of 1.0 mm each. During the scan, the participants were asked to close their eyes, try not to think, keep their heads still, and avoid falling asleep. In addition, to check for substantial disease in the brain, each participant underwent a routine T2-weighted imaging scan. We also performed functional MR imaging sequences, but they were not used in this study.
Data Processing
Voxel-based morphometric analysis: All T1 images in SPM12 (https://www.fil.ion.ucl.ac.uk/spm/software/spm12/) were analyzed using CAT12 (http://dbm.neuro.unijena.de/cat12/) to obtain data to be used in MATLAB 2021b (MathWorks, Inc., Natick, MA, USA). The following preprocessing steps were applied: (1) The images were checked for artifacts, and they were repositioned to adjust the image origin at the commissure. (2) The T1-weighted image was normalized to the standard space, and the GM, white matter and cerebrospinal fluid regions were divided and resampled. (3) The segmented GM image was smoothed using an 8 mm half-peak full-width Gaussian core. The resulting smooth GM image was used as the GMV for subsequent group comparisons. The total intracranial volume (TIV) for each participant was calculated as a covariate for comparative analysis.
Voxel-Based Morphometric Analysis (VBM)
To map the progressive pattern of GMV atrophy in patients, all patients were divided into three subgroups (acute, subacute, chronic) according to the progressive stage of disease duration. The GMV images for each subgroup were subsequently compared with those of healthy controls using a two-sample t test (voxel-level p < 0.001, cluster-level p < 0.05). In these analyses, sex, age, and intracranial total volume were regressed as covariates.
CaSCN Analysis
GMV data for all patients were sorted in order of disease duration from low to high. The seed-based CaSCN was subsequently constructed using pseudotime series, similar to the method of GC analysis applied for functional MR imaging data analysis.21 The seed region was selected from the previously mentioned voxel-based morphometric analysis. The seed points were extracted using the toolkit RESTplus (http://www.restplus.net/forum/RESTplus). Granger causality values indicated that volume changes in brain regions lagged behind seed points, which may indicate that this volume change is driven by seeds. When performing the CaSCN analysis, sex, age, and total intracranial volume were regressed as covariates. To present statistical significance, the GC plot was converted to a z score plot, with the z score and Granger causality values used as thresholds for multiple comparative correction false discovery rate (FDR).21 To further investigate the causal relationship between regions of interest (ROIs), obtained from CaSCN analysis, we performed ROI-to-ROI GC analysis and signed path coefficient GC analysis and analyzed the causal relationship between ROIs. The software programs MRIcroGL (NITRC: MRIcroGL: Tool/Resource Info) and toolkitsBrainNet Viewer (NITRC: BrainNet Viewer: Tool/Resource Info) were used to present the results.
Statistical Analysis
Prospective power analysis (GPower) indicated that a total sample of 66 was required to detect a large effect (f = 0.40) in the primary one-way ANOVA (α = 0.05, power = 0.80). For pairwise comparisons, a total sample of 52 (26 per group) was needed to detect a large effect (d = 0.80). Our study sample (N = 157) and group sizes (n ≈ 53) substantially exceed these requirements, confirming adequate power for the primary study aims.
For all the measurement data, the Shapiro‒Wilk test was first used to test their normality. The normally distributed data are expressed as the means ± standard deviations and were compared using the independent sample t test. For metric data that did not conform to a normal distribution, the median (interquartile range) was calculated. For the analysis of the count data, ie, sex, the Pearson chi-square test was used for statistical analysis.
All of the above statistical analyses were performed with SPSS 27.0 (SPSS Inc., Chicago, IL, USA) software. A two-tailed test was used for all statistical inferences, and if the P value was less than 0.05, the results were considered statistically significant.
Results
Demographic and Clinical Characteristics
The clinical and demographic data are shown in Table 1.
 |
Table 1 Clinical Demographics of Patients with Different Course of Disease and Matched Healthy Controls |
Changes in Gray Matter Volume
To map the progressive pattern of GMV atrophy in patients, all patients were divided into three subgroups (acute, subacute, chronic) according to the progressive stage of disease duration. Compared with the healthy control group (voxel-level p < 0.001, cluster-level p < 0.05), patients in the acute phase of herpes zoster presented decreased GMV in the left talar fissure and bilateral lingual gyrus (MNI: −3, 88, −4; k = 1110; peak-t = 5.37; PFWE-corr= 0.015; Cohen’s d = 01.043), and no area of increased GMV was observed (Figure 1). Patients in the subacute stage of herpes zoster presented increased GMV in the bilateral thalamus, hippocampus, lingual gyrus, right parahippocampal gyrus (MNI: −18, −30, 4; k = 2721; peak-t = 4.54; PFWE-corr< 0.001; Cohen’s d = 0.886), and left anterior central gyrus and decreased GMV in the right inferior temporal gyrus, cerebellum, right fusiform gyrus, and right middle temporal gyrus (MNI: 46, −54, −24; k = 1916; peak-t = 4.39; PFWE-corr = 0.001; Cohen’s d = 0.857) (Figure 2). Patients with chronic postherpetic neuralgia presented reduced GMV in the left cerebellum, right temporal pole middle gyrus (MNI: 20, −40, −46; k = 130; peak-t = 3.65; PFWE-corr= 0.070; Cohen’s d = 0.779), right hippocampus, left superior temporal gyrus, left anterior and paracingulate gyrus, left medial and paracingulate gyrus, and left inferior parietal angular gyrus (Figure 3). The differences in gray matter volume (GMV) among the clinical stages are presented in Table 2.
 |
Table 2 Differences in Gray Matter Volume (GMV) Among the Clinical Stages |
 |
Figure 1 GMV changes in acute Herpes Zoster patients. Blue area: Compared with HC, patients with acute-phase Herpes Zoster exhibited decreased gray matter volume in the left pericalcarine cortex and bilateral lingual gyri. The left pericalcarine cortex, located in the occipital lobe, serves as the core region for primary visual information processing and facilitates the focusing of attention on specific locations in the visual field. The bilateral lingual gyri participate in higher-order visual functions (eg, visual feature processing). |
 |
Figure 2 GMV changes in patients with subacute Herpes Zoster. Blue area: Decreased GM was observed in the following regions compared to healthy controls (HC): bilateral thalamus, bilateral hippocampi, bilateral lingual gyri, right parahippocampal gyrus, left precentral gyrus, the right inferior temporal gyrus, cerebellum, right fusiform gyrus, right middle temporal gyrus. The thalamus serves as a key relay nucleus for sensory information transmission, and its structural changes may directly affect the transmission and integration efficiency of pain signals in the center. The involvement of the hippocampus and parahippocampal gyrus suggests that the limbic system emotional memory circuit is involved in the chronicity of pain, or provides a neural structural basis for the formation of painful memories and accompanying emotional disorders (such as anxiety and depression). |
 |
Figure 3 GMV changes in patients with chronic Postherpetic Neuralgia. Blue area: Patients with chronic phase Postherpetic Neuralgia have reduced GM in the right temporal pole middle gyrus, left cerebellum, right hippocampus, left superior temporal gyrus, left anterior and paracingulate gyrus, left medial and paracingulate gyrus, and left inferior parietal margin angular gyrus compared to HC. The anterior cingulate gyrus and medial prefrontal cortex are the core nodes of emotion regulation. Alterations in the parietal cortex (angular gyrus) may be associated with spatial attention integration and somatic representation dysfunction. |
CaSCN Analysis
CaSCN Analysis of Patients with Herpes Zoster in the Acute Phase
Taking the left thalamus as the seed point, there was a significant causal relationship (P = 0.05, FDR correction) in the right inferior temporal gyrus, the pericortex of the left talar fissure, the right middle temporal gyrus, the right thalamus, and the left middle temporal gyrus (areas with voxels greater than 10 were selected) (Table 3), and causal analysis was performed on the regions with significant causal relationships (suggesting that the thalamus may be the center of the directional network and that it may have a positive causal effect on other regions). These regions were extracted for ROI-to-ROI CaSCN analysis. The ROI-to-ROI results revealed a directional network that showed a causal relationship between regions and revealed that a decrease in left thalamic GMV affected the volume of the right inferior temporal gyrus, the left talar fissure pericortex, the right thalamus, and the left middle temporal gyrus (GC = 0.48, z = 3.80, p < 0.05, FDR-corrected) (Figure 4).
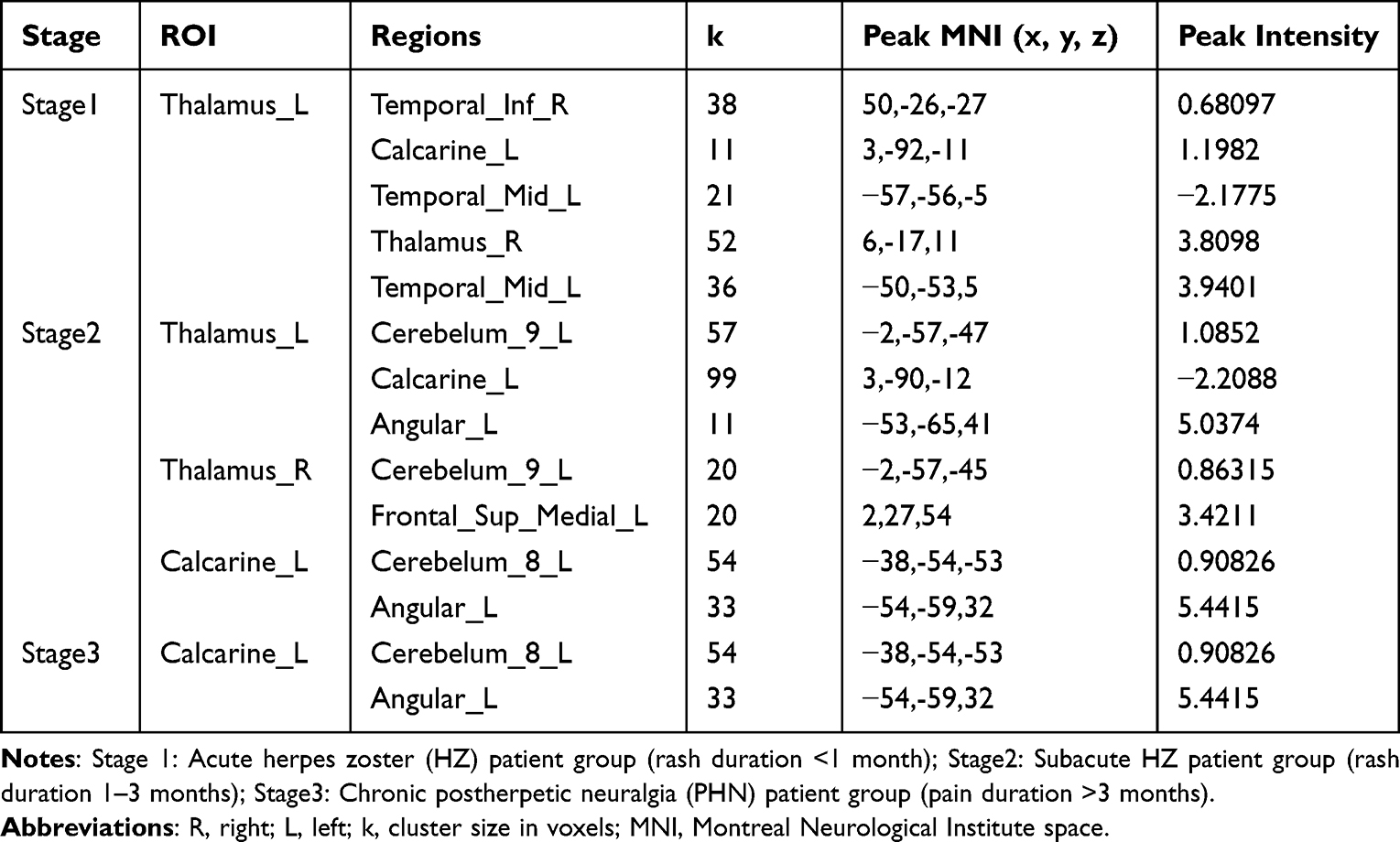 |
Table 3 Brain Regions with Significant Causal Relationships to the Calcarine_L and Thalamus_L/R |
 |
Figure 4 Causal Structural Covariance Network between brain regions in a group of patients with acute phase Herpes Zoster. The blue arrows indicate that decreased GM volume in the left thalamus affects the volume of the right inferior temporal gyrus, left pericalcarine cortex, right thalamus, and left middle temporal gyrus. The causal effect is dominated by the “thalamus→visual/temporal lobe network”. The significant causal flow of the left thalamus to the right inferior temporal gyrus, the left talar sulcus pericortex (including the V1 area), the right thalamus and the left middle temporal gyrus constitute the early information propagation network of the “thalamic–visual–temporal lobe”, suggesting the initial abnormality of pain-perception integration. |
CaSCN Analysis of Patients with Herpes Zoster in the Subacute Phase
Taking the left thalamus as the seed point, there was a significant causal relationship between the left cerebellum, the left talar fissure pericortex, and the left angular gyrus (P = 0.05, FDR correction) (Table 3), and causal analysis was carried out on the regions with significant causal relationships. The results revealed that the reduction in left thalamic GMV affected the volume of the left cerebellum and left angular gyrus. Changes in GMV in the left angular gyrus affected the volume of the left cerebellum (GC = 0.29, z = 3.98, p < 0.05, FDR-corrected).
Taking the right thalamus as the seed point, there was a significant causal relationship between the left cerebellum and the left medial superior frontal gyrus (P = 0.05, FDR correction) (Table 3), and causal analysis was carried out on the regions with significant causal relationships. The results showed that the reduction in right thalamic GMV affected the volume of the left cerebellum and the left medial superior frontal gyrus (GC = 0.34, z = 4.01, p < 0.05, FDR-corrected).
Taking the pericortex of the left talar fissure as the seed point, there was a significant causal relationship between the left cerebellum and the left angular gyrus (P = 0.05, FDR corrected) (Table 3), and causal analysis was carried out on the areas with significant causal relationships. The results revealed that the changes in GMV in the left talar fissure pericortex and left angular gyrus affected the volume of the left cerebellum (GC = 0.34, z = 4.10, p < 0.05, FDR-corrected) (Figure 5).
 |
Figure 5 Causal Structural Covariance Network between brain regions in a group of patients with Herpes Zoster in the subacute phase. The blue arrows indicate that the decreased GM volume in the left thalamus affects the volume of the left cerebellum and left angular gyrus; the GM alterations in the left angular gyrus affect the volume of the left cerebellum; the decreased GM volume in the right thalamus affects the volume of the left cerebellum and left superior medial frontal gyrus; the GM changes in both the left pericalcarine cortex and left angular gyrus affect the volume of the left cerebellum. Causal pathway diffusion of the thalamus → cerebellum/parietal lobe. Directional effects of the left thalamus on left cerebellar region VI and the left inferior parietal angular gyrus, and the right thalamus on the left cerebellar region VI and the left medial prefrontal cortex, indicating that the pain signal diffuses to the motor‒spatial coordination network. Notably, the causal effect of the left angular gyrus on the left cerebellum may reflect the compensatory regulatory mechanism of the parietal–cerebellar circuit. |
CaSCN Analysis of Patients with Herpes Zoster in the Chronic Phase
Taking the left talar fissure pericortex as the seed point, the left cuneiform cortex had a significant causal relationship (P = 0.05, FDR correction) (Table 3), and causal analysis was carried out on the areas with significant causal relationships. The results revealed that the decrease in GMV in the left-talar fissure cortex affected the volume of the left cuneiform cortex (GC = 0.55, z = 4.47, p < 0.05, FDR-corrected) (Figure 6).
 |
Figure 6 Causal Structural Covariance Network between brain regions in a group of patients with chronic Postherpetic Neuralgia. The blue arrow indicates that decreased GM volume in the left pericalcarine cortex affects the volume of the left cuneus. The causal pattern is simplified. The left talar sulcus cortex → the left cuneiform lobe (V2/V3 region) have significant positive causal flow, and network reorganization tends to be stable at this stage, but the dynamic regulation within the visual network is retained, which may suggest adaptive changes in neuroplasticity during the chronicity process. |
Discussion
In this study, CaSCN analysis was used to explore the development of HZ and PHN, as well as the changes in GMV, and the causal associations among these changes. The study design was based on disease duration ranking, and an SCN was constructed on the basis of changes in CMV,32 and the causal relationships among GMV changes in various brain regions were studied. The results revealed that the early stage of shingles was characterized by structural abnormalities in vision-related areas (the left pericalcarine cortex), which then spread to emotional, cognitive, and motor networks through the driving action of the thalamus and ultimately led to extensive brain structural reorganization. The main findings and their clinical significance are as follows:
Patterns of Brain Structure Changes During Disease Progression
In the group of patient in the acute stage of herpes zoster, a significant decrease in gray matter volume was observed in the left pericalcarine cortex (mainly including the primary visual cortex V1 region) and bilateral lingual gyri. The left pericalcarine corte is located in the occipital lobe and is the core area of primary visual information processing, visual stimuli focus and direct attention to specific locations in the field of vision,33 The bilateral lingual gyri participate in more advanced visual functions (eg, visual feature processing).34 Early structural alterations in these areas may indicate that acute HZ pain may trigger alterations at the primary cortical level through a visual–sensory integrated network. Previous studies have confirmed that visual input, particularly input that is received by the affected area, increases the perception of pain.35 This finding shows that patients in the acute stage of HZ not only experience nociceptive stimuli but also may experience sensitization across sensory channels due to the visual stimuli of skin lesions.
As the disease progresses to the subacute stage, the gray matter atrophy pattern reveals a diffusion trend that includes the bilateral thalamus, hippocampus, lingual gyrus, right parahippocampal gyrus, and left anterior central gyrus (primary motor cortex). Significant abnormalities in the thalamic structure at this stage provide key support for the “central sensitization” hypothesis; ie, continuous peripheral nociceptive input through the thalamus‒cortical circuits spreads pain signals to the broader central network, ultimately leading to dysfunction of the pain regulatory system.36 The thalamus serves as a key relay nucleus for sensory information transmission,37 and its structural changes may directly affect the transmission and integration efficiency of pain signals in the center.38 Moreover, the involvement of the hippocampus and parahippocampal gyrus suggests that the limbic system emotional memory circuit is involved in the chronicity of pain,39 or provides a neural structural basis for the formation of painful memories and accompanying emotional disorders (such as anxiety and depression).40
In the chronic PHN stage, gray matter atrophy extends to additional brain regions, such as the right temporal polar middle gyrus, left cerebellum, right hippocampus, and left superior temporal gyrus. This pattern of involving multiple networks suggests that chronic pain may trigger cognition-compensatory dysregulation of the emotion regulation network. The anterior cingulate gyrus and medial prefrontal cortex are the core nodes of emotion regulation, and their structural abnormalities may be the neuropathological basis for the high incidence of anxiety and depression in patients with PHN.41 Alterations in the parietal cortex (angular gyrus) may be associated with spatial attention integration and somatic representation dysfunction,42 providing an explanation for the common physical perception disorders of chronic pain. Persistent atrophy of the cerebellum is not only associated with motor coordination deficits but may also reflect pain-related abnormalities in atypical compensatory effects in the emotional circuit.43,44
Central Driving Role of the Thalamus in Network Reorganization
Structural neuroimaging studies have revealed that herpes zoster causes different structural changes in the brain at different stages. The thalamus is a key nerve center in pain signaling,45 and patients with chronic pain experience thalamocortical rhythm disturbances46 and motor control dysfunction.47 The thalamus is closely related to the mood and emotion of pain.48 The results of CaSCN analysis revealed that the thalamus plays a central driving role in the spread of disease. In the acute phase, the left thalamus showed significant causal effects on the right inferior temporal gyrus, the left talar fissure pericortex, the right thalamus, and the left middle temporal gyrus. This finding confirms the pivotal role of the thalamus in pain information relay,49 suggesting that structural alterations in the left thalamus may occur through transhemispheric connections (such as the corpus callosum) or the thalamus‒cortical loops that drive structural remodeling of a wider range of brain regions.50 The causal effect of the left thalamus on the left talar fissure cortex is particularly important, suggesting that although the talar fissure pericortex shows earlier structural changes in VBM analysis, its persistent structural remodeling may be regulated by thalamic dysfunction.
On the basis of causal structure covariance network (CaSCN) analysis, this study reveals the dynamic brain network reorganization pattern in the progression of herpes zoster neuralgia: 1) Acute phase (<1 month): The causal effect is dominated by the “thalamus→visual/temporal lobe network”. The significant causal flow of the left thalamus to the right inferior temporal gyrus, the left pericalcarine cortex (including the V1 area), the right thalamus and the left middle temporal gyrus constitute the early information propagation network of the “thalamic–visual–temporal lobe”, suggesting the initial abnormality of pain-perception integration. 2) Subacute phase (1–3 months): Causal pathway diffusion of the thalamus → cerebellum/parietal lobe. Directional effects of the left thalamus on left cerebellar region VI and the left inferior parietal angular gyrus, and the right thalamus on the left cerebellar region VI and the left medial prefrontal cortex, indicating that the pain signal diffuses to the motor‒spatial coordination network. Notably, the causal effect of the left angular gyrus on the left cerebellum may reflect the compensatory regulatory mechanism of the parietal–cerebellar circuit. 3) Chronic phase (>3 months): The causal pattern is simplified. The left talar sulcus cortex → the left cuneiform lobe (V2/V3 region) have significant positive causal flow, and network reorganization tends to be stable at this stage, but the dynamic regulation within the visual network is retained, which may suggest adaptive changes in neuroplasticity during the chronicity process.
Notably, we observed causal effects (eg, left angular gyrus → left cerebellum) in the subacute phase, which may reflect compensatory structural reorganization. In the early stages of chronic pain, the brain may attempt to regulate abnormal pain signals by reorganizing neural circuits, and the angular gyrus is involved in spatial attention and somatosensory integration.51 Its regulation of the cerebellum may be an attempt to restore sensation-compensatory mechanisms for movement coordination. However, as the course of the disease progresses to the chronic stage, this compensatory pattern gradually weakens and is replaced by a positive causal effect dominated by atrophy and diffusion. This dynamic may reflect the transition from compensated to decompensated, providing a new perspective on the neural mechanisms that make PHN pain chronic.
Methodological Value of Brain Structure Causal Network Analysis
In this study, causal structural covariance network (CaSCN) analysis was applied to patients with HZ and PHN, overcoming the limitation of the traditional structural covariance network (SCN), which can describe only synchronicity changes. The conventional SCN is based on zero-time lag correlation and cannot distinguish whether thalamic damage drives atrophy in other brain regions. Through Granger causal analysis, the CaSCN reveals the timing and directionality of structural changes in PHN, such as thalamus-driven visual cortex atrophy, providing a causal framework for analyzing disease transmission. This approach can help us to determine the “source” and “path” of disease transmission, not just static structural differences. This design provides an efficient way to explore the dynamic mechanisms of the disease compared with time-consuming true longitudinal studies. By stratifying patients by disease course, we are able to obtain a “snapshot” of the entire course of the disease in a single study, providing a more comprehensive perspective on the neural mechanisms of PHN. In conclusion, our study generates the first causal network hypothesis of brain alterations across HZ stages. We eagerly anticipate future research that will build upon this foundation to rigorously validate and refine these proposed causal pathways.
The initial atrophy regions and key driver nodes identified in this study provide potential targets for early intervention. The thalamus, as the central driver region of disease spread, may be an ideal target for neuromodulation (eg, deep brain stimulation and transcranial magnetic stimulation). Moreover, the structural changes in the perital cortex in the acute phase suggest that early cognitive behavioral interventions targeting the visual–sensory integration network may help prevent the development of PHN. Interventions targeting these early targets may interrupt the “waterfall effect” of disease spread, blocking the transition from acute HZ to chronic PHN. Clinical trials have shown that motor cortical stimulation relieves pain in some patients with chronic pain disorders.52,53 In particular, for patients whose pain has not resolved after standard treatment, imaging markers may become important auxiliary indicators for treatment decision-making and prognosis. Abnormal features of brain networks at different disease stages provide a neuroscientific basis for “stage-specific” treatment strategies, which may improve the accuracy and effectiveness of existing treatment methods. The progressive causal network alterations we mapped across HZ stages offer a potential objective framework for predicting disease trajectory. Future studies could investigate whether this causal network signature in the acute or subacute phase can serve as a neuroimaging biomarker to identify patients at high risk for chronification, thereby facilitating early and aggressive intervention to prevent the development of PHN.
Our study transcends mere descriptive observations to directly address core theoretical frameworks of chronic pain. First, in the context of chronic pain development, the progressive trajectory of causal network alterations we mapped provides a neural model for the transition from acute to chronic pain. This suggests that, much like what has been demonstrated in other chronic pain conditions such as neuropathic back pain,54,55 early identification of causal network features could potentially enable risk stratification in acute herpes zoster patients,56 thereby facilitating precision interventions to effectively prevent the progression to chronic pain. Second, these dynamic changes provide compelling evidence for maladaptive neuroplasticity as a central mechanism.57 The shift of causal influences from sensory-integrative hubs to those governing affective and self-referential processing likely reflects an initial failed compensatory response to persistent nociception that ultimately solidifies into a pathological architecture. Finally, our results underscore that the pathology of PHN is rooted in large-scale network reconfiguration. The central driving role of the thalamus in this reorganization demonstrates that the critical dysfunction lies not in isolated “pain centers” but in altered causal interactions within distributed networks.58
Research Limitations and Future Directions
(1) Although the design, which is based on the course of the disease, provides a new perspective on disease progression, it is necessary to carry out real longitudinal studies in the future to track the multitime point image trajectory of HZ to PHN transformation and verify the timing model with clinical and biomarker data. (2) Analyzing only gray matter volume failed to account for the multidimensional mechanism of pain conduction. In the future, DTI (white matter integrity) and fMRI (resting/task-state functional connectivity) should be integrated to construct a “structure–functional–clinical” model. The use of cross-modal coupling analysis, such as structural‒functional coupling, can improve early warning sensitivity. For instance, such integration could test whether the identified causal hub in the thalamus exerts aberrant control over functional networks involved in sensory and affective pain processing, thereby linking a structural cause to a functional consequence and ultimately to the clinical phenotype of PHN. (3) The documentation of educational levels and comorbidities was imperfectly documented for some patients, and no cognitive evaluation was performed; these limitations in clinical data collection will be systematically improved in future studies. (4) Unlike the network built on the basis of BOLD sequences and diffusion tensor imaging sequences, the network in this study is a group-level SCN, and individualized SCNs can be analyzed in the future when the sample size is sufficient.
Conclusion
In this study, causal structure covariance network analysis was applied to reveal the dynamic change pattern of gray matter volume in patients with HZ and PHN and the causal associations of these changes. The thalamus, which is a core driver node, has significant causal effects on multiple brain regions, supporting its key role in the chronicity of pain. The identified time-specific structural changes and key driver nodes provide a neuroimaging basis for early intervention, disease prediction, and personalized treatment strategies, which are expected to improve the clinical management of this refractory disease.
Acknowledgments
This work was supported by the National Key Research and Development Program of China (2022YFC3602202), the National Natural Science Foundation of China (grant no. 81960313), the Clinical Research Center For Medical Imaging In Jiangxi Province (No.20223BCG74001) and the Key research and development plans of Jiangxi Provincial Department of Science and Technology (grant no. S2020ZPYFB2343).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Hasan S, Ishrat Khan N, Nakeb AA, Tarranum F. Herpes zoster with oro-facial involvement – report of a case and detailed review of literature. Indian J Dentistry. 2012;3(2):94–15. doi:10.1016/j.ijd.2012.03.011
2. Adriaansen EJM, Jacobs JG, Vernooij LM, et al. 8. Herpes zoster and post herpetic neuralgia. Pain Pract. 2024;25(1). doi:10.1111/papr.13423
3. Patil A, Goldust M, Wollina U. Herpes zoster: a review of clinical manifestations and management. Viruses. 2022;14(2). doi:10.3390/v14020192
4. van Oorschot D, Vroling H, Bunge E, Diaz-Decaro J, Curran D, Yawn B. A systematic literature review of herpes zoster incidence worldwide. Human Vaccines Immunotherapeutics. 2021;17(6):1714–1732. doi:10.1080/21645515.2020.1847582
5. Giannelos N, Curran D, Nguyen C, Kagia C, Vroom N, Vroling H. The incidence of herpes zoster complications: a systematic literature review. Infect Dis Ther. 2024;13(7):1461–1486. doi:10.1007/s40121-024-01002-4
6. Hadley GR, Gayle JA, Ripoll J, et al. Post-herpetic neuralgia: a review. Curr Pain Headache Reports. 2016;20(3):17. doi:10.1007/s11916-016-0548-x
7. Mallick-Searle T, Snodgrass B, Brant JM. Postherpetic neuralgia: epidemiology, pathophysiology, and pain management pharmacology. J Multidiscip Healthc. 2016;9:447–454. doi:10.2147/jmdh.S106340
8. Curran D, Doherty TM, Lecrenier N, Breuer T. Healthy ageing: herpes zoster infection and the role of zoster vaccination. NPJ Vaccines. 2023;8(1):184. doi:10.1038/s41541-023-00757-0
9. Liu Q, Han J, Zhang X. Peripheral and central pathogenesis of postherpetic neuralgia. Skin Res Technol. 2024;30(8):e13867. doi:10.1111/srt.13867
10. Li X, Zhang H, Zhang X, et al. A central and peripheral dual neuromodulation strategy in pain management of zoster-associated pain. Scientific Reports. 2024;14(1):24672. doi:10.1038/s41598-024-75890-4
11. Kuner R, Kuner T. Cellular circuits in the brain and their modulation in acute and chronic pain. Physiological Rev. 2021;101(1):213–258. doi:10.1152/physrev.00040.2019
12. Cao S, Qin B, Zhang Y, et al. Herpes zoster chronification to postherpetic neuralgia induces brain activity and grey matter volume change. Am J Trans Res. 2018;10(1):184–199.
13. Liu J, Gu L, Huang Q, et al. Altered gray matter volume in patients with herpes zoster and postherpetic neuralgia. J Pain Res. 2019;12:605–616. doi:10.2147/jpr.S183561
14. Tang Y, Ren C, Wang M, et al. Altered gray matter volume and functional connectivity in patients with herpes zoster and postherpetic neuralgia. Brain Res. 2021;1769:147608. doi:10.1016/j.brainres.2021.147608
15. Jiang X, Kuang H, Lv H, et al. Aberrant functional and causal connectivity of the amygdala in herpes zoster and post-herpetic neuralgia patients. British J Radiol. 2023;96(1152):20230338. doi:10.1259/bjr.20230338
16. Ma T, Li ZY, Yu Y, et al. Gray matter abnormalities in patients with complex regional pain syndrome: a systematic review and meta-analysis of voxel-based morphometry studies. Brain Sci. 2022;12(8). doi:10.3390/brainsci12081115
17. Melie-Garcia L, Slater D, Ruef A, et al. Networks of myelin covariance. Human Brain Mapping. 2018;39(4):1532–1554. doi:10.1002/hbm.23929
18. Xiong J, Gu L, Jiang X, et al. Local structural indices changes during different periods of postherpetic neuralgia: a graphical study in structural covariance networks. J Pain Res. 2025;18:1175–1187. doi:10.2147/jpr.S515047
19. Bielczyk NZ, Uithol S, van Mourik T, Anderson P, Glennon JC, Buitelaar JK. Disentangling causal webs in the brain using functional magnetic resonance imaging: a review of current approaches. Network Neurosci. 2019;3(2):237–273. doi:10.1162/netn_a_00062
20. Lv H, Gu L, Xiong J, et al. Altered effective connectivity of the pain matrix in herpes zoster and postherpetic neuralgia patients: granger causality analysis of resting-state fMRI. J Integ Neurosci. 2024;23(8):147. doi:10.31083/j.jin2308147
21. Zhang Z, Liao W, Xu Q, et al. Hippocampus-associated causal network of structural covariance measuring structural damage progression in temporal lobe epilepsy. Human Brain Mapping. 2017;38(2):753–766. doi:10.1002/hbm.23415
22. Wan X, Wang W, Wu X, et al. Progressive structural damage in sleep-related hypermotor epilepsy. J Neurosci Res. 2023;101(9):1447–1456. doi:10.1002/jnr.25203
23. Hsieh H, Xu Q, Zhang Q, et al. Mapping progressive damage epicenters in epilepsy with generalized tonic-clonic seizures by causal structural covariance network density (CaSCNd). Brain Res. 2024;1828:148766. doi:10.1016/j.brainres.2024.148766
24. Han S, Zheng R, Li S, et al. Progressive brain structural abnormality in depression assessed with MR imaging by using causal network analysis. Psychol Med. 2023;53(5):2146–2155. doi:10.1017/s0033291721003986
25. Li Y, Wang C, Teng C, et al. Hippocampus-driving progressive structural alterations in medication-naïve major depressive disorder. J Affect Disord. 2019;256:148–155. doi:10.1016/j.jad.2019.05.053
26. Qing Z, Chen F, Lu J, et al. Causal structural covariance network revealing atrophy progression in Alzheimer’s disease continuum. Human Brain Mapping. 2021;42(12):3950–3962. doi:10.1002/hbm.25531
27. Zhou C, Zhang R, Ding M, et al. Progressive structural alterations associated with negative symptoms in schizophrenia: a causal structural covariance network analysis. Progress Neuro-Psychopharmacol Biol Psychiatry. 2025;136:111236. doi:10.1016/j.pnpbp.2024.111236
28. Mu R, Qin X, Zheng W, Yang P, Huang B, Zhu X. Progressive brain structural abnormality in cerebral small vessel disease assessed with MR imaging by using causal network analysis. NeuroImage Clin. 2024;44:103672. doi:10.1016/j.nicl.2024.103672
29. Ochoa JL. Neuropathic pain: redefinition and a grading system for clinical and research purposes. Neurology. 2009;72(14):1282–1283. doi:10.1212/01.wnl.0000346325.50431.5f
30. Hamilton M. A rating scale for depression. J Neurol Neurosurg Psychiatry. 1960;23(1):56–62. doi:10.1136/jnnp.23.1.56
31. Hamilton M. The assessment of anxiety states by rating. British J Med Psychol. 1959;32(1):50–55. doi:10.1111/j.2044-8341.1959.tb00467.x
32. Alexander-Bloch A, Giedd JN, Bullmore E. Imaging structural co-variance between human brain regions. Nat Rev Neurosci. 2013;14(5):322–336. doi:10.1038/nrn3465
33. Baroncelli L, Lunghi C. Neuroplasticity of the visual cortex: in sickness and in health. Experim neurol. 2021;335:113515. doi:10.1016/j.expneurol.2020.113515
34. Weiner KS, Grill-Spector K. Neural representations of faces and limbs neighbor in human high-level visual cortex: evidence for a new organization principle. Psychol Res. 2013;77(1):74–97. doi:10.1007/s00426-011-0392-x
35. Jordán-López J, Arguisuelas MD, Doménech J, et al. Modifying lumbar flexion pain thresholds in patients with chronic low back pain through visual-proprioceptive manipulation with virtual reality: a cross-sectional study. J Neuroengineer Rehab. 2025;22(1):138. doi:10.1186/s12984-025-01664-2
36. Yan Y, Zhu M, Cao X, et al. Thalamocortical circuit controls neuropathic pain via up-regulation of HCN2 in the ventral posterolateral thalamus. Neurosci Bulletin. 2023;39(5):774–792. doi:10.1007/s12264-022-00989-5
37. Moustafa AA, McMullan RD, Rostron B, Hewedi DH, Haladjian HH. The thalamus as a relay station and gatekeeper: relevance to brain disorders. Rev Neurosci. 2017;28(2):203–218. doi:10.1515/revneuro-2016-0067
38. Marcuse LV, Langan M, Hof PR, et al. The thalamus: structure, function, and neurotherapeutics. Neurotherapeutics. 2025;22(2):e00550. doi:10.1016/j.neurot.2025.e00550
39. Zhao W, Zhao L, Chang X, Lu X, Tu Y. Elevated dementia risk, cognitive decline, and hippocampal atrophy in multisite chronic pain. Proceed Nat Acad Sci USA. 2023;120(9):e2215192120. doi:10.1073/pnas.2215192120
40. Small SA, Schobel SA, Buxton RB, Witter MP, Barnes CA. A pathophysiological framework of hippocampal dysfunction in ageing and disease. Nat Rev Neurosci. 2011;12(10):585–601. doi:10.1038/nrn3085
41. Hein TP, Gong Z, Ivanova M, Fedele T, Nikulin V, Herrojo Ruiz M. Anterior cingulate and medial prefrontal cortex oscillations underlie learning alterations in trait anxiety in humans. Commun Biol. 2023;6(1):271. doi:10.1038/s42003-023-04628-1
42. Zhang B, Wang F, Zhang Q, Naya Y. Distinct networks coupled with parietal cortex for spatial representations inside and outside the visual field. NeuroImage. 2022;252:119041. doi:10.1016/j.neuroimage.2022.119041
43. Ji YW, Shen ZL, Zhang X, et al. Plasticity in ventral pallidal cholinergic neuron-derived circuits contributes to comorbid chronic pain-like and depression-like behaviour in male mice. Nat Commun. 2023;14(1):2182. doi:10.1038/s41467-023-37968-x
44. Ahmadian N, van Baarsen K, van Zandvoort M, Robe PA. The cerebellar cognitive affective syndrome-a meta-analysis. Cerebellum. 2019;18(5):941–950. doi:10.1007/s12311-019-01060-2
45. Coghill RC, Sang CN, Maisog JM, Iadarola MJ. Pain intensity processing within the human brain: a bilateral, distributed mechanism. J Neurophysiol. 1999;82(4):1934–1943. doi:10.1152/jn.1999.82.4.1934
46. Jones EG. Thalamocortical dysrhythmia and chronic pain. Pain. 2010;150(1):4–5. doi:10.1016/j.pain.2010.03.022
47. Knotkova H, Hamani C, Sivanesan E, et al. Neuromodulation for chronic pain. Lancet. 2021;397(10289):2111–2124. doi:10.1016/s0140-6736(21)00794-7
48. Grodd W, Kumar VJ, Schüz A, Lindig T, Scheffler K. The anterior and medial thalamic nuclei and the human limbic system: tracing the structural connectivity using diffusion-weighted imaging. Scientific Reports. 2020;10(1):10957. doi:10.1038/s41598-020-67770-4
49. Galdino G, Veras FP, Dos Anjos-Garcia T. The role of the thalamus in nociception: important but forgotten. Brain Sci. 2024;14(8). doi:10.3390/brainsci14080741
50. Lenz FA, Dougherty PM, Meeker TJ, Saffer MI, Oishi K. Neuroscience of the human thalamus related to acute pain and chronic “thalamic” pain. J Neurophysiol. 2024;132(6):1756–1778. doi:10.1152/jn.00065.2024
51. Saccani MS, Contemori G, Del Popolo Cristaldi F, Bonato M. Attentional load impacts multisensory integration, without leading to spatial processing asymmetries. Scientific Reports. 2025;15(1):16240. doi:10.1038/s41598-025-95717-0
52. Wang F, Tian ZC, Ding H, et al. A sensory-motor-sensory circuit underlies antinociception ignited by primary motor cortex in mice. Neuron. 2025;113(12):1947–1968.e7. doi:10.1016/j.neuron.2025.03.027
53. Kong Q, Li T, Reddy S, Hodges S, Kong J. Brain stimulation targets for chronic pain: insights from meta-analysis, functional connectivity and literature review. Neurotherapeutics. 2024;21(1):e00297. doi:10.1016/j.neurot.2023.10.007
54. Baliki MN, Petre B, Torbey S, et al. Corticostriatal functional connectivity predicts transition to chronic back pain. Nature Neurosci. 2012;15(8):1117–1119. doi:10.1038/nn.3153
55. Wager TD, Sutherland SP, Lindquist MA, Sluka KA. Accelerating discovery in pain science: the acute to chronic pain signatures program. Pain. 2025;166(11s):S95–s98. doi:10.1097/j.pain.0000000000003674
56. Wu Y, Wang C, Qian W, et al. Default mode network-basal ganglia network connectivity predicts the transition to postherpetic neuralgia. IBRO Neurosci Reports. 2025;18:135–141. doi:10.1016/j.ibneur.2025.01.009
57. Yang P, Chen H, Wang T, et al. Electroacupuncture promotes synaptic plasticity in rats with chronic inflammatory pain-related depression by upregulating BDNF/TrkB/CREB signaling pathway. Brain Behav. 2023;13(12):e3310. doi:10.1002/brb3.3310
58. Kucyi A, Davis KD. The dynamic pain connectome. Trends Neurosci. 2015;38(2):86–95. doi:10.1016/j.tins.2014.11.006
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Fire Needling Acupuncture for Adult Patients with Acute Herpes Zoster: Protocol of a Systematic Review and Meta-Analysis
Liu L, Chen Q, Yang J, Gang W, Zhao L, Lyu T, Jing X, Zhang CS, Li B
Journal of Pain Research 2022, 15:2161-2170
Published Date: 2 August 2022
Comparison of a One- versus Two-Week Treatment with Famciclovir Upon Reductions in Pain and Occurrence of Postherpetic Neuralgia in Herpes Zoster: A Randomized Open-Label Trial
Peng F, He H, Xia T, Lv S
Infection and Drug Resistance 2023, 16:721-726
Published Date: 2 February 2023
Effects of Intradermal Botulinum Toxin Injections on Herpes Zoster Related Neuralgia
Peng F, Xia TB
Infection and Drug Resistance 2023, 16:2159-2165
Published Date: 12 April 2023
The Effect of CT-Guided Pulsed Radiofrequency Combined with Ozone Injection on Zoster-Associated Pain: A Retrospective Study
Wang X, Yu J, Han CF, He JD, Yang WQ, Wang Q, Chen JP
Journal of Pain Research 2023, 16:1321-1332
Published Date: 20 April 2023
Local Structural Indices Changes During Different Periods of Postherpetic Neuralgia: A Graphical Study in Structural Covariance Networks
Xiong J, Gu L, Jiang X, Kuang H, Lv H, Li Z, Xie Y, Luo Q, Jiang J
Journal of Pain Research 2025, 18:1175-1187
Published Date: 11 March 2025