Back to Journals » Nature and Science of Sleep » Volume 17

Gut Microbiome Composition Changes During Insomnia Treatment with Lemborexant
Authors Miyata S ![]() , Iwamoto K
, Iwamoto K ![]() , Ito M, Okada I, Matsuyama N, Fujimoto A, Kogo Y, Nishiwaki H, Ueyama J, Ohno K, Ozaki N
, Ito M, Okada I, Matsuyama N, Fujimoto A, Kogo Y, Nishiwaki H, Ueyama J, Ohno K, Ozaki N
Received 5 March 2025
Accepted for publication 24 June 2025
Published 25 July 2025 Volume 2025:17 Pages 1709—1726
DOI https://doi.org/10.2147/NSS.S526037
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah L Appleton
Seiko Miyata,1,* Kunihiro Iwamoto,1,* Mikako Ito,2 Ippei Okada,3 Nao Matsuyama,1,3 Akihiro Fujimoto,4 Yuki Kogo,4 Hiroshi Nishiwaki,2,5 Jun Ueyama,6 Kinji Ohno,2,7 Norio Ozaki1,8
1Department of Psychiatry, Nagoya University Graduate School of Medicine, Nagoya, Aichi, Japan; 2Division of Neurogenetics, Center for Neurological Diseases and Cancer, Nagoya University Graduate School of Medicine, Nagoya, Aichi, Japan; 3PharmaLex Japan, Inc., Shibuya, Tokyo, Japan; 4Medical Headquarters, Eisai Co., Ltd., Bunkyo, Tokyo, Japan; 5Department of Genetics, Stanford University School of Medicine, Stanford, CA, USA; 6Department of Biomolecular Sciences, Field of Omics Health Sciences, Nagoya University Graduate School of Medicine, Nagoya, Aichi, Japan; 7Graduate School of Nutritional Sciences, Nagoya University of Arts and Sciences, Nisshin, Aichi, Japan; 8Pathophysiology of Mental Disorders, Nagoya University Graduate School of Medicine, Nagoya, Aichi, Japan
*These authors contributed equally to this work
Correspondence: Seiko Miyata, Department of Psychiatry, Nagoya University Graduate School of Medicine 65 Tsurumai-Cho, Showa-Ku, Nagoya, 466-8550, Japan, Email [email protected]
Purpose: Insomnia is a common disorder worldwide. Growing evidence has revealed that the microbiota–gut–brain axis contributes to the regulation of sleep continuity and duration, both directly and indirectly. Although lemborexant is effective in treating insomnia, its effect on the gut microbiota remains unclear. Here, we investigated the relationship between the gut microbiota and hypnotic use in insomnia.
Participants and Methods: We enrolled 29 adults with insomnia and performed sleep electroencephalography and stool analyses at baseline and after 4 and 12 weeks of open-label lemborexant treatment. Changes in gut microbiota were analyzed using 16S rRNA sequencing and metabolite analysis was performed to assess short-chain fatty acids (SCFAs).
Results: Beta diversity (Jaccard dissimilarity) and Firmicutes/Bacteroidetes ratio significantly increased after administration of lemborexant for 12 weeks (p < 0.05). Seven genera were significantly different (p < 0.05). Among these, Tannerellaceae Parabacteroides decreased significantly after 12 weeks of lemborexant treatment (p = 0.013), even after correcting for false discovery rates. Akkermansia was strongly negatively correlated with sleep efficiency (r = − 0.754, p = 0.0003). Allisonella showed opposite correlations with latency to persistent sleep and sleep efficiency after 12 weeks of lemborexant treatment (r = 0.523, p = 0.018, r = − 0.516, p = 0.020, respectively). There were no significant differences in SCFAs during the treatment period.
Conclusion: Our findings suggest that prolonged lemborexant treatment in individuals with insomnia may induce notable shifts in gut microbiota composition, including a significant reduction in Parabacteroides underscoring the potential interaction between hypnotic use and gut microbial balance.
Keywords: insomnia intervention, gut microbiota, lemborexant, short-chain fatty acids, elderly, home sleep monitoring
Introduction
Insomnia is a widespread disorder in many countries, and its prevalence has increased in recent decades.1,2 Insufficient sleep affects both physical and mental health and contributes to the pathogenesis of various neuropsychiatric disorders.3–5 Therefore, exploring the role of sleep in these disorders is important for the optimal diagnosis and treatment. There is considerable evidence that the gut microbiota regulates host sleep and mental states through the microbiota–gut–brain axis.6–8 A better understanding of gut microbiota in relation to diseases may be advantageous in elucidating the relationship between insomnia and psychiatric disorders.
The microbiota–gut–brain axis directly and indirectly contributes to regulating sleep behavior, such as sleep continuity and duration, and may play a critical role in the etiology and pathogenesis of sleep disorders.9 Both acute and chronic insomnias are associated with reduced microbial richness, decreased diversity, and depleted short-chain fatty acid (SCFA)-producing bacteria compared to those in healthy controls.10 A canonical correspondence analysis of 72 older patients with insomnia revealed that sleep efficiency and cognitive performance explained 7.5–7.9% of the variation7 in gut microbiota composition. Gut microbiota-targeted interventions using different strategies, such as probiotics, prebiotics, and fecal microbiota transplantation, have been shown to exert therapeutic effects under different conditions.8 However, to the best of our knowledge, the effect of hypnotic insomnia treatment on the gut microbiota has not been studied.
According to two Phase III trials using polysomnography and/or sleep diaries, lemborexant, a competitive dual orexin receptor antagonist, improves sleep onset and sleep maintenance in patients with insomnia without inducing rebound insomnia or withdrawal.11,12 In our home monitoring of subjective and objective sleep (FLUID study), all subjective sleep parameters (sleep onset latency, wake after sleep onset [WASO], total sleep time [TST], and sleep efficiency) improved significantly with treatment, and all objective sleep parameters, including measurements of TST and WASO, also improved when considering baseline values.13,14
Given the growing body of evidence that the gut microbiota is intricately linked to sleep regulation and mental health, it is crucial to explore how pharmacological interventions, such as lemborexant, might influence this relationship. However, the effects of sleep medications on the gut microbiome, particularly in the context of insomnia, remain unclear. We hypothesized that lemborexant treatment would affect the diversity and balance of the gut microbiota. Thus, the purpose of this study was to investigate the impact of lemborexant on the gut microbiome in patients with insomnia and to evaluate changes in microbial diversity, the abundance of key bacterial populations, and the production of SCFAs, which are thought to influence sleep and brain health.
Materials and Methods
Participants
The inclusion criteria of the FLUID study were as follows; (1) ability to provide written informed consent before the study commenced, (2) age ≥50 years, (3) diagnosis of insomnia disorder based on the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition, and (4) a score of ≥24 on the Japanese version of the Mini-Mental State Examination. The exclusion and discontinuation criteria are described in the protocol elsewhere.13 Additionally, participants who claimed to have taken antibiotics in the past month were excluded from gut microbiome analysis. This FLUID study, open-label, single-arm, single-center, was conducted at Nagoya University Hospital from June 1, 2021, to September 30, 2022.The study protocol was approved by the Nagoya University Certified Review Board (2021–0079) and registered in the Japanese Registry of Clinical Trials (jRCT s041210024). The study was conducted in compliance with the principles of the Declaration of Helsinki and the Ministry of Health, Labour, and Welfare. All participants agreed to the purpose and procedures of this study, and informed consent was obtained from all participants.
Study Procedure
The study design has been previously described in detail.13 Briefly, after the initial screening period, eligible participants were treated with lemborexant (5 mg; LEM5) for 12 weeks, ensued by a follow-up period of approximately 4 weeks. If the participants did not feel that their sleep or other symptoms had improved, the dose was increased to 10 mg (LEM10) after consultation with a study physician based on the participants’ complaints. Fecal samples were collected by the participants in their homes within one week of each visit (baseline, week 4, and week 12), and samples were delivered to the hospital at 0 °C within 24 h. These samples were stored at −80 °C until analysis. The medication doses prescribed before the screening period did not change until the last observation period. Medications other than prohibited ones were permitted for temporary use. Lifestyle habits including diet were assessed at each visit; however, no special controls were used.
Sleep Analysis
Home-based objective sleep measurements were conducted using a single-channel EEG device called Zmachine Insight+ (General Sleep Corporation, Cleveland, OH), which monitors sleep patterns and provides an algorithmic sleep-stage classification. This device has undergone validation studies involving healthy individuals, patients with insomnia, and those with psychiatric conditions, with methodological details available in the published literature.15,16 The device was designed for easy self-application by participants before bedtime. Four key metrics were assessed: the time until consistent sleep onset (latency to persistent sleep or LPS, defined as the minutes elapsed before reaching a period where 10 out of 12 min was classified as sleep), representing objective sleep onset latency, WASO, TST, and sleep efficiency (SE, calculated as the proportion of TST to total recording duration). For subjective sleep assessment, electronic sleep diaries were utilized to gather data on subjective sleep onset latency (sSOL), subjective WASO (sWASO), subjective TST (sTST), and subjective SE (sSE, calculated as the proportion of TST to time in bed). Sleep diary data were averaged over the previous seven days, including the visit date, and used for analysis.
Questionnaires
Sleep quality and disturbances during the previous month were evaluated using the Pittsburgh Sleep Quality Index (PSQI).17 This tool consists of 19 items that evaluate seven different aspects of sleep, including how participants subjectively rate their sleep quality, how long it takes them to fall asleep, and whether they use sleep medications. Sleep disturbance was indicated when a participant scored 6 or higher on the PSQI. The Epworth Sleepiness Scale (ESS)18 was employed to assess daytime drowsiness. Participants who scored 11 or higher on this scale were considered to have experienced excessive daytime sleepiness.
Sample Collection, DNA Isolation, and V3–V4 16S rRNA sequencing
The gut microbiota analysis was performed according to the procedure described previously.19 All fecal samples were collected in screw-top fecal containers (height: 54 mm; diameter: 28 mm; polypropylene) with brown screw caps and an integrated spoon (high-density polyethylene; SARSTEDT Inc., Nümbrecht, Germany) by the study participants at home. The samples were transported to the laboratory at 0 °C and stored at −80 °C. Freezing was performed using a freeze-dryer (FDU-2110) connected to a drying chamber (DRC-1100; EYELA, Tokyo, Japan). After drying, the fecal samples were moved from the fecal containers to a disposable grinding chamber (MT 40, IKA, Staufen, Germany). The samples were then ground (IKA tube mill control), and the fine-grain dried fecal samples were stored at −30 °C.
A QIAamp PowerFecal DNA Kit (QIAGEN, Hilden, Germany) was used to extract DNA from 20 mg freeze-dried samples. To ensure efficient bacterial DNA extraction, FastPrep-24 5G (MP Biomedicals, Qbiogene, Montreal, Canada) was used instead of a vortex. The samples were homogenized in PowerBead Solution and Solution C1 of the QIAamp PowerFecal DNA Kit with beads (Lysing Matrix E; MP Biomedicals, Santa Ana, CA) using FastPrep-24 5G at 6.0 m/s for 60s and three cycles.
The V3–V4 hypervariable region of the bacterial 16S rRNA gene was amplified using a pair of primers (341F, 5’-CCTACGGGNGGCWGCAG-3 and 805R, 5’-GACTACHVGGGTATCTAATCC-3’). The KAPA HiFi HotStart Ready-mix PCR kit (Kapa Biosystems, Wilmington, MA) and Nextera XT index kit (Illumina, San Diego, CA) were used to prepare the metagenomic sequencing library. Nucleotide fragments were sequenced using the MiSeq System (Illumina) and MiSeq Reagent Kit V3 (600-cycle). QIIME2 was used for taxonomic analysis. FASTQ files were quality-filtered and DADA2 was used to generate amplicon sequence variants. The samples were not discarded during filtration. For taxonomic analysis, a pre-trained naïve Bayes classifier and q2-feature-classifier plugin from QIIME2 were used. The q2-feature-classifier was used to produce a trained reference from the SILVA taxonomic database that released 138 references for taxonomic identification.
The taxa were filtered at the genus and family levels under the following conditions: taxa with average relative abundances > 0.01% were used for the analysis.
Metabolite Analysis in Fecal samples
Short-chain fatty acids (acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, and isovaleric acid) were quantified using a standardized derivatization-gas chromatography-mass spectrometry protocol.20 Lyophilized fecal samples (20 mg) were extracted with 5 mmol/L NaOH. After centrifugation, the supernatant was chemically derivatized using 2-methyl-1-propanol, pyridine, and isobutyl chloroformate with hexanoic acid 6,6,6-d3 as the internal standard. The derivatized samples were extracted with hexane and analyzed using an Agilent 7890A gas chromatograph coupled to an Agilent 5975 inert mass spectrometer (Agilent Technologies, Santa Clara, CA) for metabolite quantification.
Statistical Analysis
The basic statistics were calculated for each score at each evaluation point. The alpha diversity was evaluated using Shannon’s index and Simpson’s diversity, and the beta diversity was evaluated by Bray–Curtis analysis,21 Jaccard dissimilarity,22 and Chao23 at each visit. For beta diversity, the difference between weeks 4 and 12 for each participant was calculated, using the baseline visit as a reference. Next, principal coordinate analysis (PCoA) was used to analyze the effects of lemborexant administration using permutational multivariate analysis of variance (PERMANOVA).24 The effects of each feature were evaluated using three distance metrics: Bray–Curtis, Jaccard dissimilarity, and Chao. The values were calculated using the vegan R package. The ratio of Firmicutes/Bacteroidetes (F/B) was calculated. All the genera were included in the analysis. A comparison of these indices at baseline and at weeks 4 and 12 was performed using the Wilcoxon signed-sum rank test. Statistical significance was set at p < 0.05.
The taxa were filtered at the genus and family levels using the following two conditions. First, for each taxon, the number of samples was counted, and the relative abundance of the taxon of interest was greater than 1E-4. The number of such samples should be >20% of all samples. Second, taxa with an average relative abundance of >0.001 were selected.
Then, gut microbiota candidates associated with insomnia disorders and sleep quality (sleep efficiency, PSQI, and Insomnia Severity Index) were extracted from previous papers analyzing them in human studies.6,7,10,25–27 We identified one gut microbial order and 18 genera (Supplementary Table 1).
Analysis of variance (ANOVA) with Friedman test and Wilcoxon signed-rank sum test were used to compare the differential abundance of taxa at baseline and at weeks 4 and 12. The p-values were corrected for false discovery rate (FDR) using the Benjamini–Hochberg method for all detected enteric gut microbiomes, and a q value < 0.05 was considered significant. If Friedman test results were significant, the Jonckheere–Terpstra test was used to determine whether the increase or decrease in the gut microbiome was a constant trend throughout the study period. The associations between all bacterial families or genera, fecal metabolites and sleep parameters (LPS, WASO, TST, SE, sSOL, sWASO, sTST, and sSE) as well as PSQI and ESS scores were analyzed using Pearson’s correlation coefficient. Statistical significance was defined as p<0.05. Changes in fecal metabolite levels at each visit were compared using the Wilcoxon signed-rank sum test.
All analyses were performed using R ver 4.4.1 (R Foundation for Statistical Computing, Vienna, Austria), EZR (Saitama Medical Center, Jichi Medical University, Saitama, Japan),28 and JMP pro ver.17 (SAS Institute, Cary, NC). The study was exploratory, conducted as an adjunct to the FLUID study, and did not independently calculate the sample size before the study. Therefore, the effect size (Cohen’s d) was calculated using post-hoc analysis.
Results
Participants’ Background
Twenty-nine participants provided consent and completed the sample collection at baseline and week 4. Two participants discontinued the study before the week 12-visit, and fecal samples were collected from 27 participants at week 12. All participants were initially administered 5 mg lemborexant, which was subsequently titrated as needed to 10 mg at week 4 based on subjective symptomatology and clinical assessment. Of the 27 participants who completed the study, 10 received a 10-mg dose of lemborexant (Figure 1). The participants’ backgrounds are listed in Table 1. Overall, the mean age was 58.8 years (range: 50–72 years). Among them, 14 (48%) were male, and 10 were titrated to LEM10 after week 4. The most common symptoms were difficulty maintaining sleep (n = 24), waking up earlier than desired (n = 19), and difficulty initiating sleep (n = 17); insomnia duration was 6.3 ± 7.2 years.
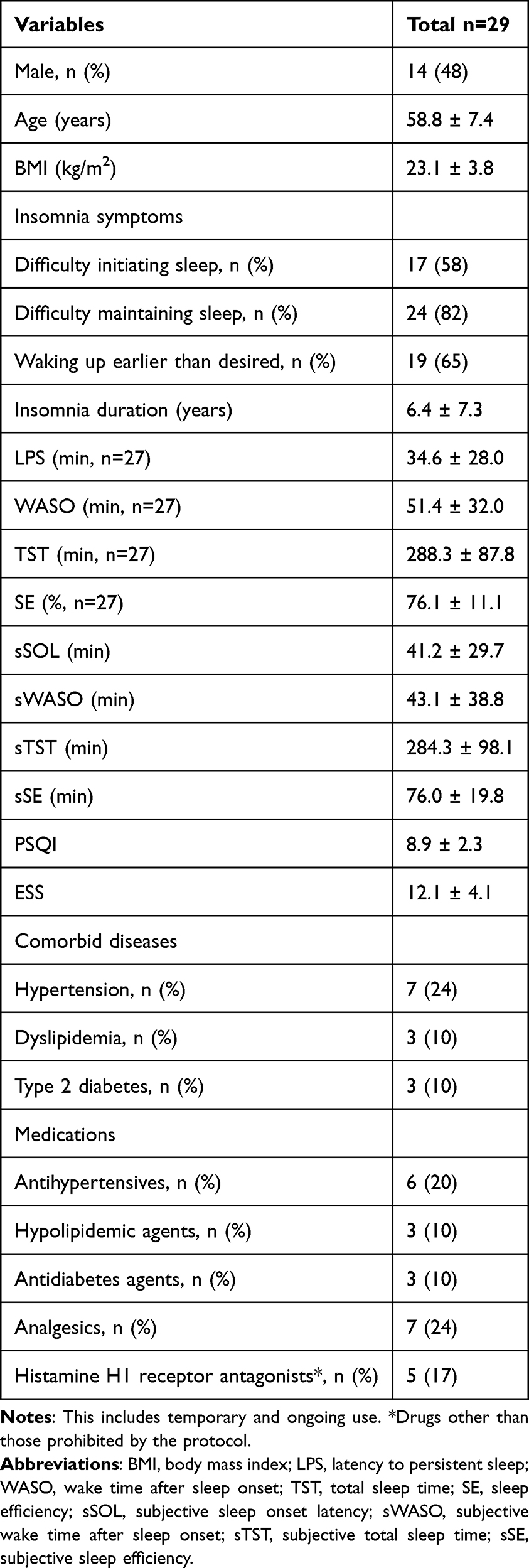 |
Table 1 Participants’ Backgrounds |
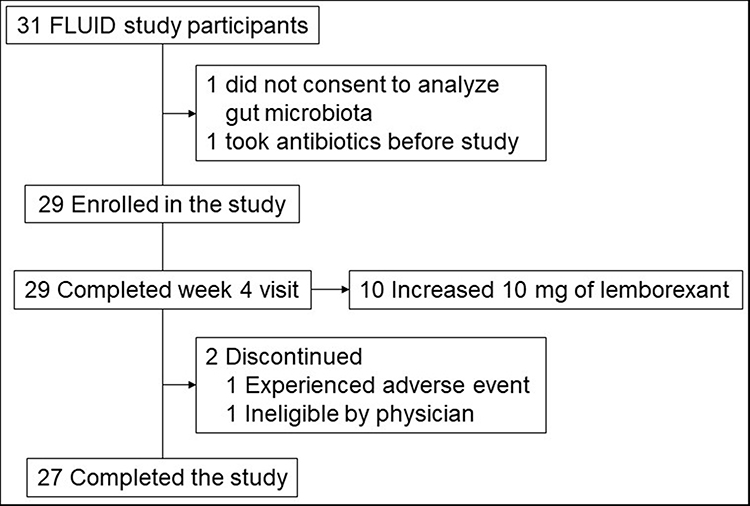 |
Figure 1 Participants through the study. |
Changes of the Gut Microbiome Diversity with Lemborexant Administration
Alpha diversity, calculated as Shannon’s index and Simpson’s diversity, did not show any significant differences between the baseline and weeks 4 and 12 (Figure 2a). However, the beta diversity calculated using Jaccard dissimilarity significantly increased at week 12 compared to week 4. The Chao index at week 12 slightly increased compared to that at week 4 but did not reach a significant level. Bray–Curtis analysis showed no significant differences between weeks 4 and 12 (Figure 2b). In all diversity estimations by Bray–Curtis analysis, Jaccard dissimilarity, and Chao index, the centers of gravity at baseline and weeks 4 and 12 overlapped in the PCoA of the bacterial species. The overall bacterial composition by PERMANOVA also showed no significant differences (Figure 2c). The F/B ratio was significantly higher at week 12 than that at baseline. However, there was no significant difference in the F/B ratio between the baseline and week 4 (Figure 2d).
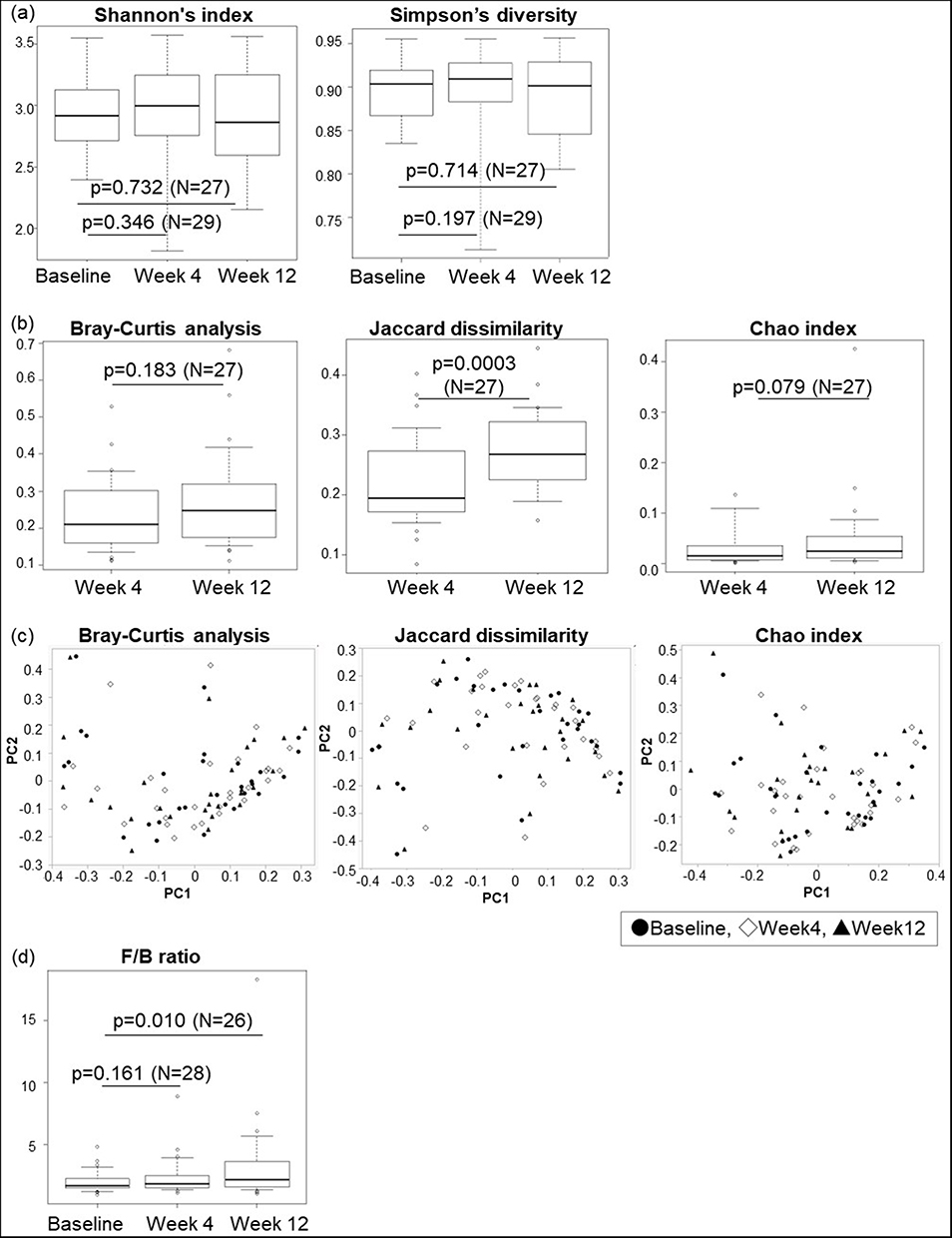 |
Figure 2 Gut microbiome diversity at each visit. (a) Alpha diversity was based on Shannon’s index and Simpson’s diversity. (b) Beta diversity by Bray–Curtis analysis, Jaccard dissimilarity, and Chao index. The number of cases used in the analysis was 27 cases paired in weeks 4 and 12. (c) Principal coordinate analysis using Bray–Curtis analysis, Jaccard dissimilarity, and Chao index. (d) Firmicutes/Bacteroidetes ratio. In one case, data was missing. Therefore, baseline to week 4 comparisons were performed in 28 cases and baseline to week 12 comparisons in 26. The p-values obtained by Wilcoxon’s signed-rank test were all < 0.05. |
Changes of on the Composition of the Gut Microbiome with Lemborexant Administration
Firmicutes, Bacteroidetes, and Actinobacteria were the three most abundant phyla detected during the three visits. At the family level, the relative abundance of two families was significantly increased, and that of one family was significantly decreased at week 4 compared to baseline, but these were not significantly different at week 12 (Supplementary Figure 1). Seven families were significantly different at week 12 compared with baseline, and Tannerellaceae remained significant at week 12 after FDR correction (q value = 0.004, Supplementary Figure 2).
At the genus level, significant differences were observed among the three visits. Six bacterial taxa showed distinct relative abundances between baseline and week 4 (Figure 3 and Supplementary Figure 3). Anaerovoracaceae Family XIII AD3011 group increased at week 4 compared to baseline (0.0007±0.0007 vs 0.0010±0.0008, n = 29, p = 0.045, q = 0.738) and remained at week 12 (0.0007±0.0008 vs 0.0018±0.0036, n = 27, p = 0.003, q = 0.185), although it did not reach significance after FDR correction (Figure 3). At week 12, seven bacterial taxa showed distinct relative abundances compared to those at baseline. Among them, Tannerellaceae Parabacteroides was still significantly lower at week 12 than at baseline after adjustment for FDR (0.0179±0.0098 vs 0.0111±0.0063, n = 27, p = 0.0001, q < 0.0001, Cohen’s d = 0.53) (Figure 3) and tended to decrease at week 4. Trend analysis revealed that Parabacteroides significantly decreased (p = 0.001) during the study period. Decreases in Bacteroidaceae Bacteroides (p = 0.014, q = 0.387) and Erysipelatoclostridiaceae Erysipelotrichaceae UCG-003 (p = 0.017, q = 0.387) and increases in Oscillospiraceae uncultured (p = 0.017, q = 0.387), Streptococcaceae Streptococcus (p = 0.022, q = 0.387), and Akkermansiaceae Akkermansia (p = 0.021, q = 0.387) did not reach significance after adjustment for FDR.
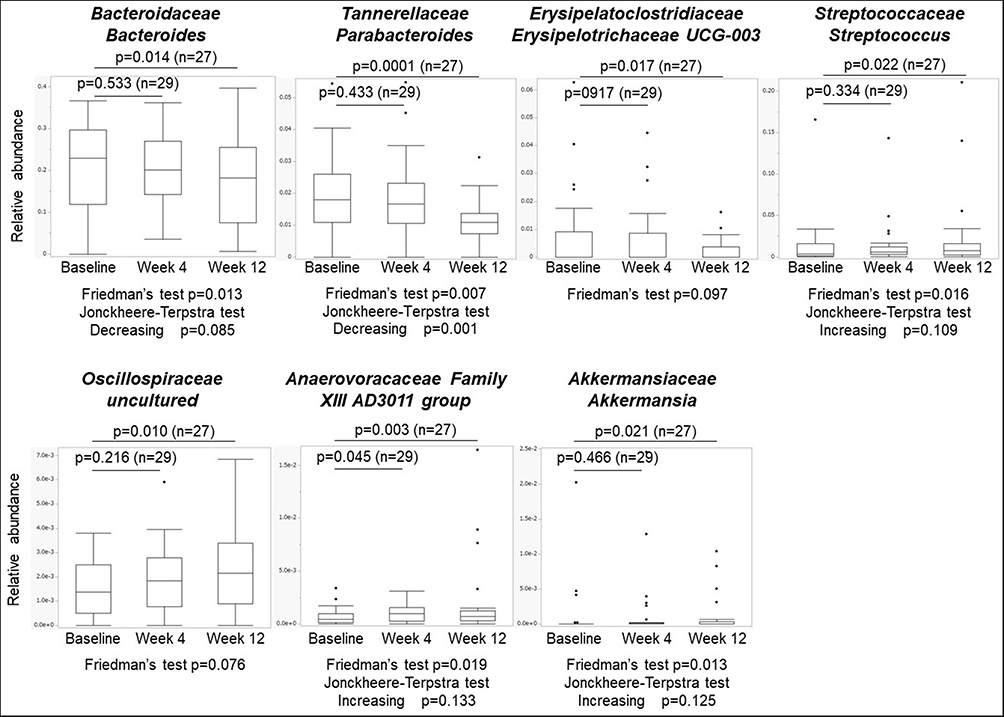 |
Figure 3 Composition of the gut microbiome at the genus level. P-values were calculated using the Wilcoxon signed-rank test. Q values were calculated using the Benjamini–Hochberg method. |
Changes in Gut Microbiome Associated with Insomnia/Sleep Disturbances Reported in the Previous studies
Of the 18 bacterial genera previously associated with insomnia, Parabacteroides was also significantly reduced. The order Lactobacillales appeared to be increased by the administration of lemborexant but did not reach significance levels (week 4, p = 0.188; week 12, p = 0.052). Barnesiellaceae Coprobacter increased significantly from the baseline at week 4 (p = 0.041), but there was no significant difference at week 12 (p = 0.052). The other 16 bacterial genera did not show any significant changes after lemborexant administration in our study population (Supplementary Table 1).6,7,10,25–27
Correlation Analysis of Sleep Measures and Questionnaire Scores with Gut Microbiome
Several gut microbiota were significantly correlated with both objective and subjective sleep parameters, with some taxa showing consistent directional associations.
Akkermansia showed a strong negative correlation with SE (r = −0.754, p = 0.0003) and sSOL (r = −0.612, p = 0.001 at week 12). Similarly, Megasphaera was consistently negatively correlated with both sTST and sSE, whereas the Eubacterium ruminantium group showed a strong negative correlation with sSOL (r = −0.713, p < 0.0001). At week 12, Christensenellaceae and Subdoligranulum were negatively correlated with LPS (r = −0.691 and r = −0.606, respectively; p < 0.01) (Tables 2 and 3).
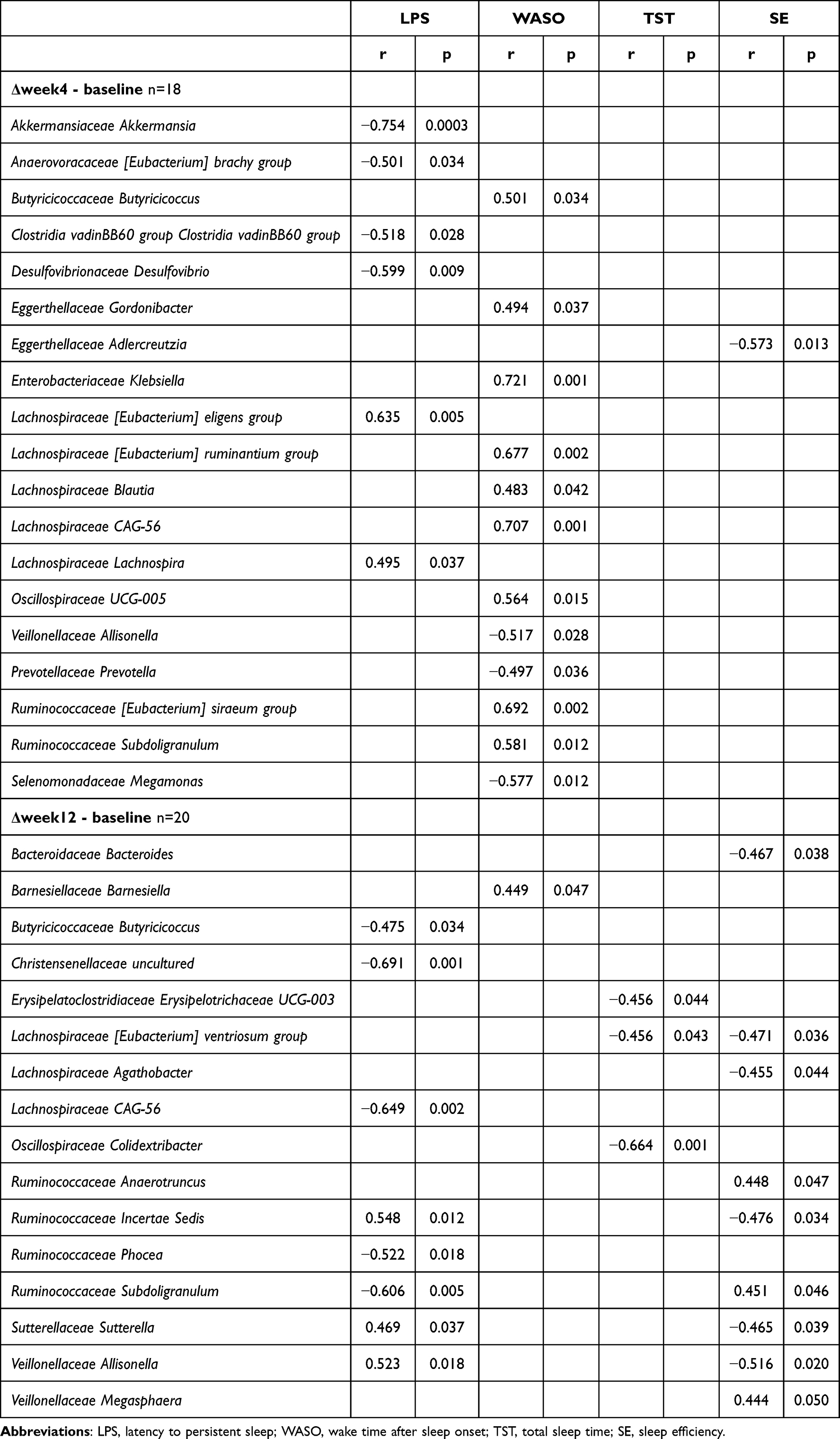 |
Table 2 Correlation Between Objective Sleep Indices and Gut Microbiomes |
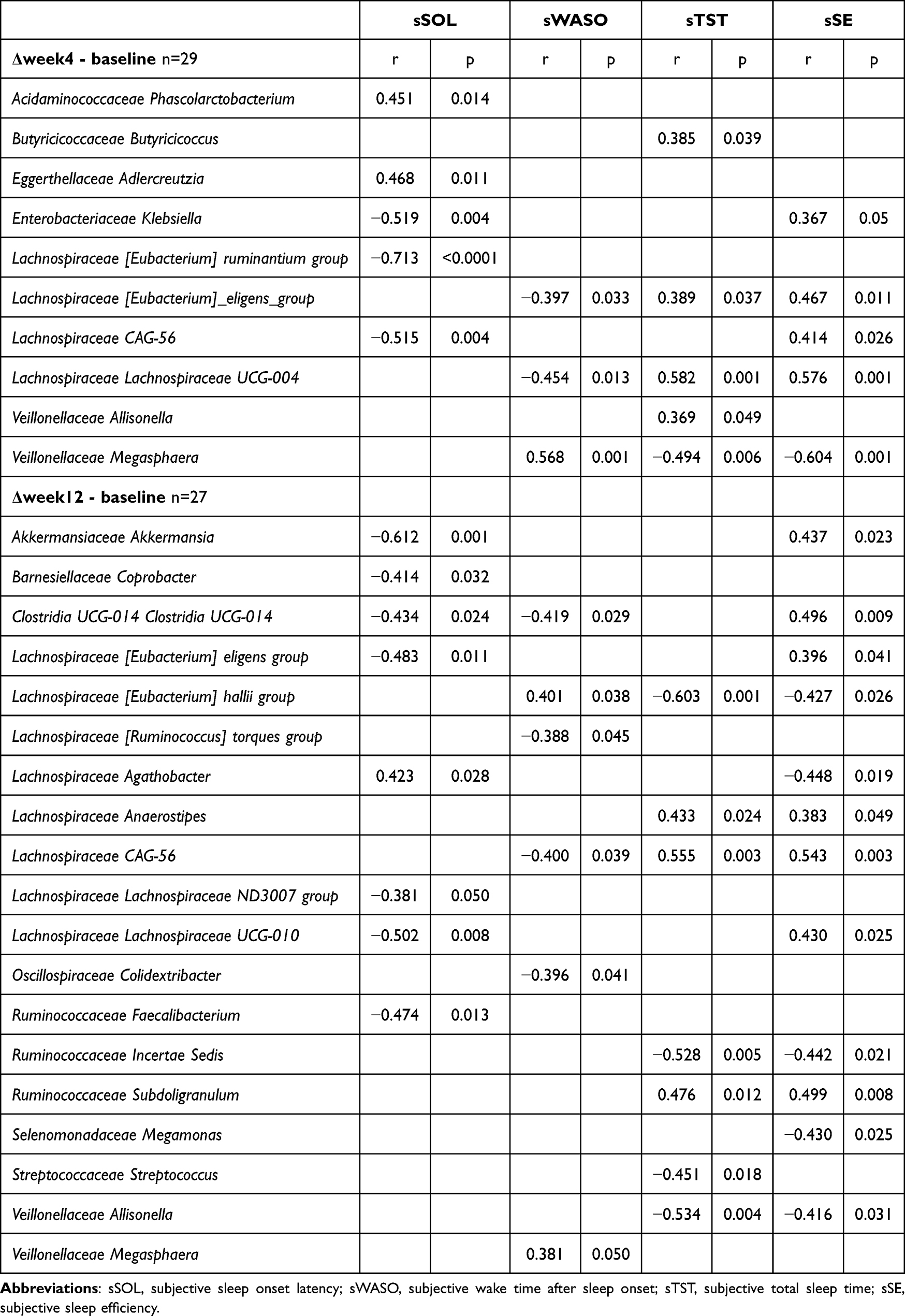 |
Table 3 Correlation Between Subjective Sleep Indices and Gut Microbiomes |
Klebsiella and CAG-56 were positively associated with TST (r = 0.721 and r = 0.707, respectively; p < 0.01) (Table 2). CAG-56 was also positively correlated with both sTST and sSE at week 12 (r = 0.555, p = 0.003 and r = 0.543, p = 0.003, respectively) (Table 3).
Allisonella demonstrated a significant positive correlation with LPS (r = 0.523, p = 0.018), and a significant negative correlation with SE (r = −0.516, p = 0.020) (Table 2).
Gut microbiota was also associated with self-reported sleep quality measures. At week 4, Phascolarctobacterium was positively correlated with the ESS (r = 0.591, p = 0.001), and Anaerotruncus was positively associated with the PSQI (r = 0.555, p = 0.002). At week 12, the Collinsella, Eubacterium, and Lachnospiraceae hallii groups showed strong positive correlations with the ESS (r = 0.516, 0.514, and 0.521, respectively; p < 0.01). In contrast, Desulfovibrio and Prevotella were negatively associated with the ESS (r = −0.507 and −0.509, respectively; p = 0.007), and Lachnospiraceae UCG-004 was inversely correlated with the PSQI (r = −0.522, p = 0.005) (Table 4).
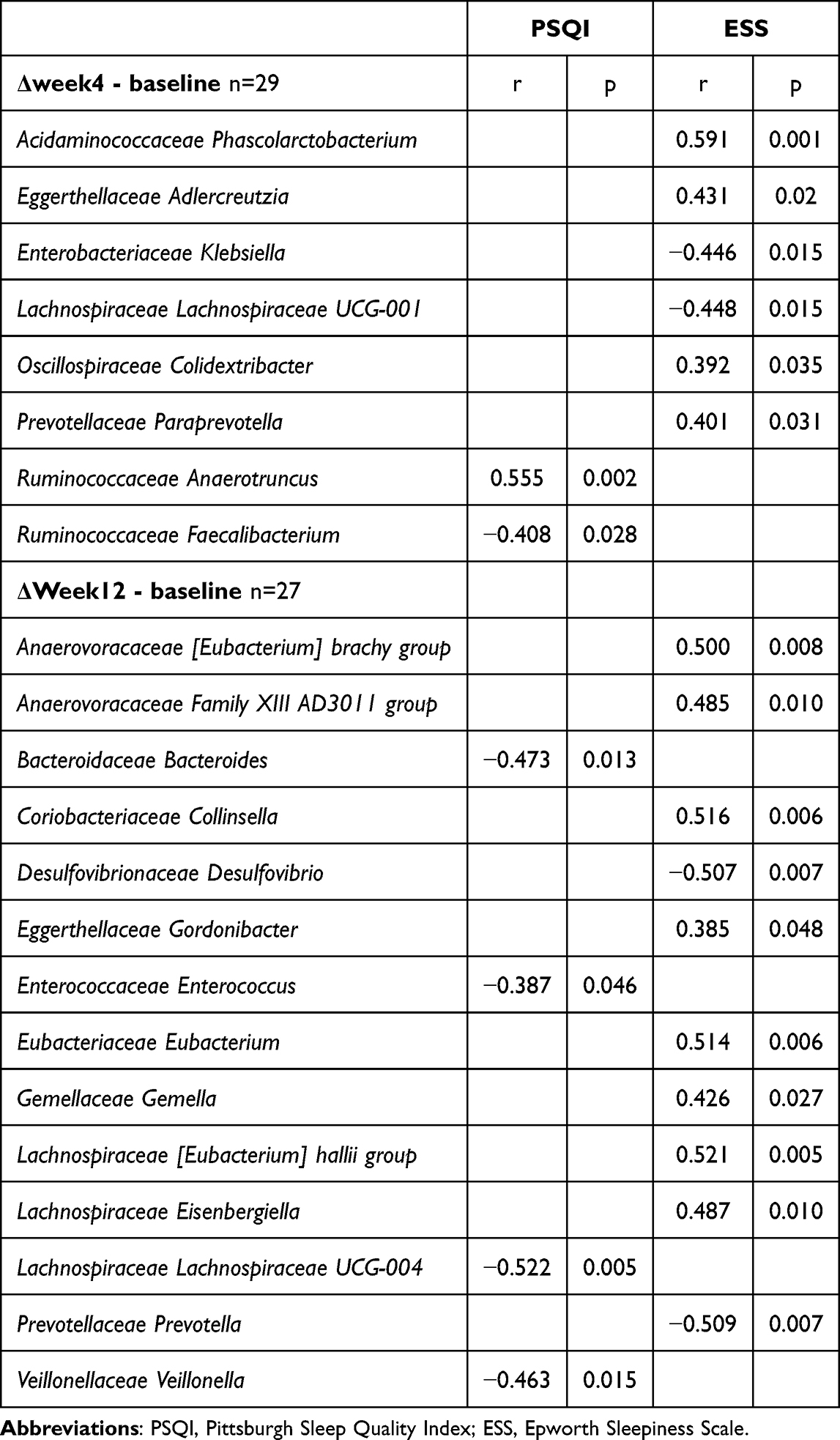 |
Table 4 Correlation Between Questionnaire Scores and Gut Microbiomes |
Changes of the Fecal Metabolites with Lemborexant Administration
The concentrations of SCFAs in the feces are summarized in Table 5. Acetic acid levels tended to increase during the study period; however, this increase was not statistically significant. Other SCFAs, such as propionic acid, butyric acid, isobutyric acid, valeric acid, and isovaleric acid, did not differ significantly among the three visits.
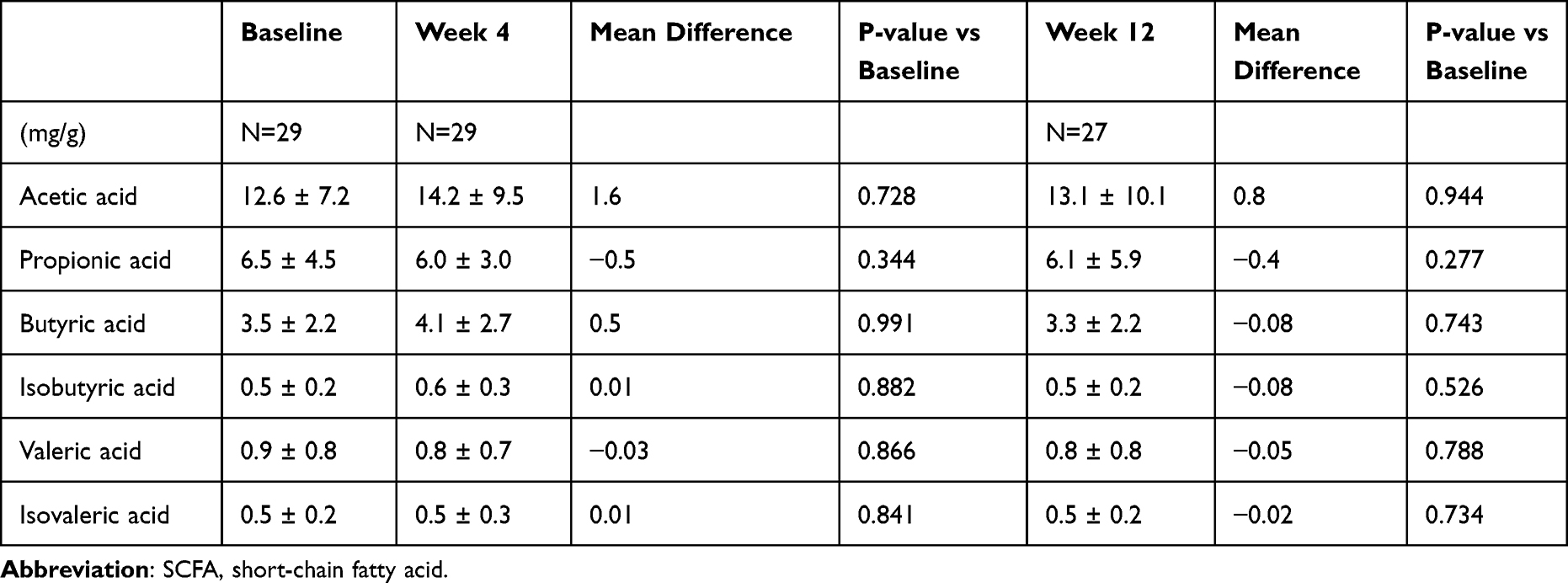 |
Table 5 Concentration of Fecal Metabolites of SCFAs |
Butyric acid showed significant correlations with multiple objective sleep parameters, including longer LPS (r = 0.580, p = 0.012), shorter TST (r = −0.480, p = 0.044), and reduced WASO (r = −0.482, p = 0.031). None of the SCFAs showed a strong correlation (r ≥ 0.5) with the subjective sleep index at weeks 4 or 12. No statistically significant or substantially stronger correlations were found for all SCFAs with PSQI and ESS (Supplementary Tables 2–4).
Discussion
To our knowledge, this is the first study to investigate the effects of hypnotics on the gut microbiome and its metabolites. This study provides evidence that lemborexant administration in patients with insomnia leads to the following significant alterations in the gut microbiota: 1) an increase in beta-diversity by the Jaccard dissimilarity, 2) an increase in the F/B ratio, 3) a reduction in the genus T. Parabacteroides, and 4) some gut microbiota showing significant correlation with sleep measures, thereby highlighting a potential microbiota–gut–brain axis modulation through hypnotic use.
We showed that beta diversity and F/B ratio increased after 12 weeks of lemborexant administration. Firmicutes and Bacteroidetes are the two dominant phyla accounting for approximately 90% of the total gut microbiome.29 The association between F/B ratio and various diseases has been repeatedly reported. Both changes in beta diversity and F/B ratio have been reported in patients with insomnia.26 In healthy controls, self-reported sleep quality was positively associated with microbial diversity and F/B ratio.30 Sleep duration was positively correlated, and waking after sleep onset was negatively correlated with microbiome diversity.31 The importance of balanced gut microbiota for host health has also been demonstrated. The gut microbiome, the largest reservoir of microorganisms in the human body, plays an important role in neurodevelopment, aging, and brain diseases.32 Our results provide evidence that hypnotic administration affects the gut microbiota composition associated with sleep quality.
In our study, after 12 weeks of treatment with lemborexant, the composition of the gut microbiome changed significantly compared with that at baseline. Among them, Parabacteroides remained significantly decreased, even after multiple corrections with FDR. A study in sleep-deprived mice showed that Tannerellaceae was significantly increased in sleep-deprived mice and was positively correlated with TNF-α, suggesting that pro-inflammatory cytokine responses may be caused by the disorder of the microbiota.33 T. Parabacteroides and Pyy, which regulates gastrointestinal functions and satiety, increased in response to a high-fat diet and affected the homeostasis of the intestinal epithelium.34 To the best of our knowledge, there have been no previous reports of the association between insomnia and Tannerellaceae. However, we observed a significant decrease in Parabacteroides following lemborexant treatment. Our observation of significantly reduced Parabacteroides levels after lemborexant administration provides evidence of this microbiome shift in insomnia treatment, potentially opening new avenues for understanding sleep-gut microbiome interactions.
Six other bacterial genera (A. Family XIII AD3011 group, B. Bacteroides, A. Akkermansia, E. Erysipelotrichaceae UCG-003, S. Streptococcus, Oscillospiraceae uncultured), in addition to T. Parabacteroides, were significantly altered by insomnia treatment, although they did not show any significance after FDR correction. A. Family XIII AD3011 group has been reported to be associated with elevated levels of atherosclerosis biomarkers35 and an increased risk of neurodevelopmental disorders due to dysbiosis in the maternal lineage of mice fed a high-fat diet.36 Bacteroides is known as a distinguishable biomarker in insomnia from healthy controls.10,26 A. Akkermansia is adversely affected by chronic intermittent hypoxia and sleep fragmentation in mice.37 E. Erysipelotrichaceae UCG-003 increases in Alzheimer’s disease and mild cognitive impairment.38 Streptococcus spp. is decreased in patients with a PSQI score ≥16 after traumatic brain injury.39 A decrease in Oscillospiracea is related to inflammatory diseases40 and depression.41 Changes in the bacterial genera after 12 weeks of lemborexant treatment were favorable, although the changes were not significant after correction. Our findings support previously reported associations between sleep disorders and the gut microbiota.
Several studies have reported changes in the gut microbiota of patients with insomnia disorders compared with healthy controls. Therefore, we investigated whether the treatment altered these effects. B. Bacteroides, which has been repeatedly reported in insomnia disorders,10,26 tended to decrease continuously during the study period. Moreover, the order Lactobacillales, to which the lactic acid-producing bacteria belong, and which are related to brain function in insomnia disorders,6 changed significantly after lemborexant administration. The changes observed in insomnia-related gut bacteria following treatment may shed light on the mechanisms underlying the relationship between insomnia and the gut microbiome.
Our study identified significant relationships between the gut microbiome and various sleep parameters, including objective and subjective indices, sleep quality, and daytime sleepiness. Building on prior work,11 which linked Bacteroides and PSQI scores in acute insomnia, we found similar but weaker correlations. Our comprehensive sleep indices identified multiple gut bacterial genera associated with specific sleep parameters. Interestingly, some bacterial genera differed from those previously linked to changes in gut microbiota during sleep interventions. Our earlier research showed that while subjective symptoms improved after medication, objective improvement varied based on pre-medication values,14 suggesting a subject-objective measurement discrepancy that may explain the inconsistent results. Larger validation studies with diverse populations are required to confirm the association between insomnia symptoms, sleep indices, and gut microbiota changes.
When comparing the beta diversity between baseline and week 4 or 12 using the three different methods, only Jaccard dissimilarity showed a significant difference between week 4 and 12. Several indicators have been proposed for assessing beta diversity. Jaccard dissimilarity, Bray–Curtis analysis, and Chao index were used in this study. Jaccard dissimilarity is qualitative, based on the number of species observed, and is not affected by the amount of each species present.22 Bray–Curtis analysis is also quantitative, based on observed species abundance, and is therefore susceptible to the distribution of key bacterial species.21 Finally, the Chao index is a versatile similarity index considering rare species.23 Differences in how the beta diversity was calculated may have affected the detection of significant differences.
No significant changes were observed in any SCFAs during the treatment period, except for a slight increase in acetic acid. SCFAs, the main metabolites produced by bacterial fermentation of dietary fiber in the gastrointestinal tract, are speculated to play a key role in the microbiota–gut–brain crosstalk.42 Higher concentrations of acetic acid, butyric acid, and propionic acid are associated with lower sleep efficiency and longer sleep-onset latency in older adults with insomnia symptoms.43 As previously reported,14 sleep improvement after the intervention varied among individuals; therefore, the relationship between sleep index improvement and changes in SCFAs should be investigated in further studies.
To the best of our knowledge, no studies have examined the changes in gut microbiota due to sleep drug interventions. From previous studies,6–8,10,25–27 dual orexin receptor antagonists might indirectly affect the gut microbiota via changes in sleep patterns. Orexin neurons project to multiple brain regions and regulate stress responses and energy and glucose metabolism via the autonomic and endocrine systems.44 Orexin is a chemical mediator in the gut–brain axis, and its effects on the gut microbiota have been reported.45 Therefore, direct or indirect effects of lemborexant on gut microbiota are expected, though no studies have investigated this relationship. Further studies are needed to elucidate the mechanism underlying the relationship between sleeping pills and changes in the gut microbiota.
Regarding the relationship between improvement in ESS and changes in gut microbiomes, six genera were identified, with r > 0.5, indicating a relatively strong relationship. Patients with narcolepsy have been shown to have different gut microbiota profiles than healthy controls, and changes in the gut microbiota affect microbiota–gut–brain communication, which may be associated with the development of narcolepsy and may also induce changes in the gut microbiota.46
In relation to PSQI, Lachnospiraceae showed a strong correlation and Bacteroides a moderate correlation. Both these bacteria have been associated with sleep problems, suggesting that this relationship may be mediated by microbial metabolites and sleep-related neurochemicals linked to these bacteria.47,48
Many internal and external factors affect gut microbiota. Routinely used non-antibiotics, including proton pump inhibitors and statins, also provide diversity in the gut microbiota.49,50 Nutrition induces diet-derived signals at the host–microbiota interface to maintain homeostasis or enhance disease susceptibility.51 Physical activity has the potential to modulate gut microbiota diversity and composition, which may influence depressive symptoms via immune, hormonal, and neural mechanisms within the gut–brain axis.52 This study did not control for previously prescribed medications or daily routines. Therefore, these factors must be considered in addition to the effects of lemborexant. However, the fact that this was an intervention study rather than a cross-sectional analysis and that medications and lifestyle habits continued to be monitored suggests that these factors were minimized.
From the perspective of the brain–gut–microbiota axis, both mental and sleep disorders have been reported to be related to gut bacteria.9,32 Insomnia plays an important role as a risk factor, comorbid condition, and transdiagnostic symptom of many mental disorders, including mood and anxiety disorders and schizophrenia.53,54 An analysis focusing on this axis may help us better understand the relationship between sleep and mental disorders. Studies on the gut microbiota and their metabolites may provide new insights into the relationship between sleep and mental disorders.
This exploratory study has several limitations, including the absence of a placebo control group, small and age-limited sample size, and non-standardized fecal sample collection timing.55 The study had a low success rate for sleep EEG recordings and a limited amount of objective sleep data. These factors may have contributed to the absence of significant gut biome changes. Although the effect size of a significantly altered bacterial genus was significant (Cohen’s d = 0.53), the number of participants was not optimized and required revalidation. Menopause is known to affect the composition of the gut microbiota,56 and all the women in this study were postmenopausal. Although the participants’ daily lives were not strictly controlled, significant changes were reviewed by physicians. Medication use was controlled; one participant using high-impact antibiotics was excluded, and other drugs remained unchanged. Therefore, monitoring diet, exercise, and medications during the study period may have minimized confounding effects. Future studies should incorporate a placebo-controlled design to more rigorously evaluate the causal relationship between the intervention and the observed outcomes, thereby enhancing the internal validity of the findings.
Conclusion
The composition of the gut microbiome at week 12 of lemborexant treatment differed from that at baseline, including an increase in beta diversity and B/F ratio. Particularly, T. Parabacteroides, which is related to sleep deprivation and inflammatory responses, significantly decreased after 12 weeks of insomnia treatment. In addition, several gut microbiome genera were associated with objective and subjective sleep indices, and subjective symptom indices. Our findings may help us understand the complex relationship between sleep and the diversity of the gut microbiota because of the deep relationship between sleep and the brain–gut axis.
Abbreviations
SCFA, short-chain fatty acid; TST, total sleep time; WASO, wake after sleep onset; LEM5, lemborexant 5 mg; LEM10, lemborexant 10 mg; PCoA, principal coordinate analysis; PERMANOVA, permutational multivariate analysis of variance; F/B, Firmicutes/Bacteroidetes; FDR, false discovery rate.
Data Sharing Statement
The datasets generated during and/or analyzed during the current study are available in the DDBJ repository (https://www.ddbj.nig.ac.jp/index-e.html). BioSample metadata are available in the DDBJ BioSample database under accession number SAMD00848081-SAMD00848165. Please contact the corresponding author for further information if necessary.
Ethics Approval and Informed Consent
The study protocol was approved by the Nagoya University Certified Review Board (2021–0079) and registered in the Japan Registry of Clinical Trials (jRCT s041210024, https://jrct.niph.go.jp/en-latest-detail/jRCTs041210024). The study was conducted in compliance with the principles of the Declaration of Helsinki and the Ministry of Health, Labour, and Welfare. All participants agreed to the purpose and procedures of the study and provided written informed consent before starting the study.
Acknowledgments
We thank Tomoko Wada, Keita Kawai, and Minami Kinouchi for their help with sample and data collection and Tomomi Yamada for the analysis of the samples. The MMSE-J was purchased from Nippon Bunka Sha (https://www.nichibun.co.jp/english/products/), printing the authorized Japanese edition of the MMSE, which was prepared under contract with the original publisher (Psychological Assessment Resources). We have registered the Japanese version of ESS with Qualitest Co., Ltd., which holds the rights to the Japanese version of ESS.
Author Contributions
SM: Methodology, Formal analysis, Investigation, Data Curation, Writing-Original Draft, Visualization, Funding Acquisition; KI: Methodology, Investigation, Writing-Review & Editing; MI: Investigation, Writing-Review & Editing; IO: Investigation, Writing-Review & Editing; NM: Investigation, Writing-Review & Editing; AF: Interpretation, Writing-Review & Editing; YK: Interpretation, Writing-Review & Editing; HN: Formal analysis, Writing-Review & Editing; JU: Investigation, Writing-Review & Editing; KO: Investigation, Supervision, Writing - Review & Editing, NO: Conceptualization, Writing - Review & Editing, Supervision, Funding acquisition.
All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was financially supported by Eisai Co., Ltd., the owner and manufacturer of lemborexant, and grants from AMED (No. JP24zf0127011) and JSPS KAKENHI (Grant Number 23K10934).
Disclosure
AF and YK are employees of Eisai Co., Ltd. KI received personal fees from Eisai, Lundbeck Japan, MSD, Otsuka, Sumitomo Pharma, Takeda, Viatris, Yoshitomi, Meiji Seika Pharma, DAIICHI SANKYO and Kyowa outside of the submitted work. NO has received research support or speaker honoraria from, or has served as a joint researcher with, or a consultant to, Otsuka Pharmaceutical Co., Ltd., Sumitomo Pharma Co., Ltd., Viatris Inc., Eisai Co., Ltd., The KAITEKI Institute, Inc., Ricoh Co., Ltd., Mitsubishi Tanabe Pharma Co.; reports personal fees/grants from Kyowa Pharmaceutical Industry, Medical Review, Eli Lilly, DAIICHI SANKYO, TSUMURA, Takeda, Mochida, Meiji Seika Pharma, EA Pharma, Viatris, Ricoh, Nippon Boehringer Ingelheim, Lundbeck Japan, Nihon Medi-Physics, Nippon Chemiphar, outside the submitted work. The other authors declare no conflicts of interest.
References
1. Ahn E, Baek Y, Park J-E, Lee S, Jin H-J. Elevated prevalence and treatment of sleep disorders from 2011 to 2020: a nationwide population-based retrospective cohort study in Korea. BMJ Open. 2024;14(2):e075809. doi:10.1136/bmjopen-2023-075809
2. Kronholm E, Partonen T, Härmä M, et al. Prevalence of insomnia-related symptoms continues to increase in the Finnish working-age population. J Sleep Res. 2016;25(4):454–457. doi:10.1111/jsr.12398
3. Palagini L, Bastien CH, Marazziti D, Ellis JG, Riemann D. The key role of insomnia and sleep loss in the dysregulation of multiple systems involved in mood disorders: a proposed model. J Sleep Res. 2019;28(6):e12841. doi:10.1111/jsr.12841
4. Hertenstein E, Feige B, Gmeiner T, et al. Insomnia as a predictor of mental disorders: a systematic review and meta-analysis. Sleep Med Rev. 2019;43:96–105. doi:10.1016/j.smrv.2018.10.006
5. Chellappa SL, Aeschbach D. Sleep and anxiety: from mechanisms to interventions. Sleep Med Rev. 2022;61:101583. doi:10.1016/j.smrv.2021.101583
6. Feng Y, Fu S, Li C, et al. Interaction of gut microbiota and brain function in patients with chronic insomnia: a regional homogeneity study. Front Neurosci. 2022;15:804843. doi:10.3389/fnins.2021.804843
7. Haimov I, Magzal F, Tamir S, et al. Variation in gut microbiota composition is associated with sleep quality and cognitive performance in older adults with insomnia. Nat Sci Sleep. 2022;14:1753–1767. doi:10.2147/NSS.S377114
8. Neroni B, Evangelisti M, Radocchia G, et al. Relationship between sleep disorders and gut dysbiosis: what affects what? Sleep Med. 2021;87:1–7. doi:10.1016/j.sleep.2021.08.003
9. Wang Z, Lu T, Lu T, et al. The microbiota-gut-brain axis in sleep disorders. Sleep Med Rev. 2022;65:101691. doi:10.1016/j.smrv.2022.101691
10. Li Y, Zhang B, Zhou Y, et al. Gut microbiota changes and their relationship with inflammation in patients with acute and chronic insomnia. Nat Sci Sleep. 2020;12:895–905. doi:10.2147/NSS.S271927
11. Rosenberg R, Murphy P, Zammit G, et al. Comparison of lemborexant with placebo and zolpidem tartrate extended release for the treatment of older adults with insomnia disorder: a Phase 3 randomized clinical trial. JAMA Network Open. 2019;2:e1918254–e1918254. doi:10.1001/jamanetworkopen.2019.18254
12. Yardley J, Kärppä M, Inoue Y, et al. Long-term effectiveness and safety of lemborexant in adults with insomnia disorder: results from a phase 3 randomized clinical trial. Sleep Med. 2021;80:333–342. doi:10.1016/j.sleep.2021.01.048
13. Okada I, Iwamoto K, Miyata S, et al. FLUID study: study protocol for an open-label, single-centre pilot study to investigate the effect of lemborexant on sleep management in Japanese subjects aged 50 years and older with insomnia disorder. BMJ Open. 2021;11(11):e054885. doi:10.1136/bmjopen-2021-054885
14. Miyata S, Iwamoto K, Okada I, et al. Assessing the real-world, long-term impact of lemborexant on sleep quality in a home-based clinical study. Nat Sci Sleep. 2024;16:291–303. doi:10.2147/NSS.S448871
15. Wang Y, Loparo KA, Kelly MR, et al. Evaluation of an automated single-channel sleep staging algorithm. Nat Sci Sleep. 2015;7:101–111. doi:10.2147/NSS.S77888
16. Miyata S, Iwamoto K, Banno M, et al. Performance of an ambulatory electroencephalogram sleep monitor in patients with psychiatric disorders. J Sleep Res. 2021;30:e13273.
17. Buysse DJ, Reynolds CF 3rd, Monk TH, et al. The Pittsburgh sleep quality index: a new instrument for psychiatric practice and research. Psychiatry Res. 1989;28(2):193–213. doi:10.1016/0165-1781(89)90047-4
18. Johns MW. A new method for measuring daytime sleepiness: the Epworth sleepiness scale. Sleep. 1991;14(6):540–545. doi:10.1093/sleep/14.6.540
19. Nishiwaki H, Ueyama J, Kashihara K, et al. Gut microbiota in dementia with Lewy bodies. NPJ Parkinson’s Dis. 2022;8(1):169. doi:10.1038/s41531-022-00428-2
20. Ueyama J, Hayashi M, Hirayama M, et al. Effects of pesticide intake on gut microbiota and metabolites in healthy adults. Int J Envir Res Public Health. 2023;20:213. doi:10.3390/ijerph20010213
21. Bray JR, Curtis JT. An ordination of the upland forest communities of southern Wisconsin. Ecol Monogr. 1957;27:325–349. doi:10.2307/1942268
22. Ludwig JA, Reynolds JF. Statistical Ecology: A Primer in Methods and Computing. Vol 1. John Wiley & Sons; 1988.
23. Chao A, Chazdon RL, Colwell RK, Shen T-J. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 2005;8:148–159. doi:10.1111/j.1461-0248.2004.00707.x
24. Anderson MJ. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001;26:32–46.
25. Fang H, Yao T, Li W, et al. Efficacy and safety of fecal microbiota transplantation for chronic insomnia in adults: a real world study. Front Microbiol. 2023;14:1299816. doi:10.3389/fmicb.2023.1299816
26. Liu B, Lin W, Chen S, et al. Gut microbiota as an objective measurement for auxiliary diagnosis of insomnia disorder. Front Microbiol. 2019;10:01770. doi:10.3389/fmicb.2019.01770
27. Zhang Q, Yun Y, An H, et al. Gut microbiome composition associated with major depressive disorder and sleep quality. Front Psychiat. 2021;12:645045. doi:10.3389/fpsyt.2021.645045
28. Kanda Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013;48:452–458. doi:10.1038/bmt.2012.244
29. Huttenhower C, Gevers D, Knight R, et al. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486:207–214.
30. Grosicki GJ, Riemann BL, Flatt AA, Valentino T, Lustgarten MS. Self-reported sleep quality is associated with gut microbiome composition in young, healthy individuals: a pilot study. Sleep Med. 2020;73:76–81. doi:10.1016/j.sleep.2020.04.013
31. Smith RP, Easson C, Lyle SM, et al. Gut microbiome diversity is associated with sleep physiology in humans. PLoS One. 2019;14(10):e0222394. doi:10.1371/journal.pone.0222394
32. Basiji K, Sendani AA, Ghavami SB, et al. The critical role of gut-brain axis microbiome in mental disorders. Metab Brain Dis. 2023;38:2547–2561. doi:10.1007/s11011-023-01248-w
33. Zhang M, Zhang M, Kou G, Li Y. The relationship between gut microbiota and inflammatory response, learning and memory in mice by sleep deprivation. Front Cell Infect Microbiol. 2023;13:1159771. doi:10.3389/fcimb.2023.1159771
34. Farzi A, Ip CK, Reed F, et al. Lack of peptide YY signaling in mice disturbs gut microbiome composition in response to high-fat diet. FASEB J. 2021;35:e21435. doi:10.1096/fj.202002215R
35. Burton KJ, Krüger R, Scherz V, et al. Trimethylamine-N-Oxide postprandial response in plasma and urine is lower after fermented compared to non-fermented dairy consumption in healthy adults. Nutrients. 2020;12:234. doi:10.3390/nu12010234
36. Gesù CM D, Matz LM, Bolding IJ, et al. Maternal gut microbiota mediate intergenerational effects of high-fat diet on descendant social behavior. Cell Rep. 2022;41:111461. doi:10.1016/j.celrep.2022.111461
37. Wang F, Zou J, Xu H, et al. Effects of chronic intermittent hypoxia and chronic sleep fragmentation on gut microbiome, serum metabolome, liver and adipose tissue morphology. Front Endocrinol. 2022;13:i820939. doi:10.3389/fendo.2022.820939
38. Zhu Z, Ma X, Wu J, et al. Altered Gut microbiota and its clinical relevance in mild cognitive impairment and Alzheimer’s disease: shanghai aging study and Shanghai memory study. Nutrients. 2022;14:3959. doi:10.3390/nu14193959
39. Zhanfeng N, Liang W, Jing K, Jinbo B, Yanjun C, Hechun X. Regulation of sleep disorders in patients with traumatic brain injury by intestinal flora based on the background of brain-gut axis. Front Neuroscience. 2022;16:934822. doi:10.3389/fnins.2022.934822
40. Gophna U, Konikoff T, Nielsen HB. Oscillospira and related bacteria – from metagenomic species to metabolic features. Environ Microbiol. 2017;19:835–841. doi:10.1111/1462-2920.13658
41. Jiang H, Ling Z, Zhang Y, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behavior Immunity. 2015;48:186–194. doi:10.1016/j.bbi.2015.03.016
42. Dalile B, Van Oudenhove L, Vervliet B, Verbeke K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nature Rev Gastroenterol Hepatol. 2019;16:461–478.
43. Magzal F, Even C, Haimov I, et al. Associations between fecal short-chain fatty acids and sleep continuity in older adults with insomnia symptoms. Sci Rep. 2021;11:4052. doi:10.1038/s41598-021-83389-5
44. Wang Q, Wang X, Yang C, et al. The role of sleep disorders in cardiovascular diseases: culprit or accomplice? Life Sci. 2021;283:119851. doi:10.1016/j.lfs.2021.119851
45. Mediavilla C. Bidirectional gut-brain communication: a role for orexin-A. Neurochem Int. 2020;141:104882. doi:10.1016/j.neuint.2020.104882
46. Wu J, Li Y. Relationship between type 1 narcolepsy and gut microbiota: sleepy people, sleepy microbiota? Sleep. 2024;47(4):zsae050. doi:10.1093/sleep/zsae050
47. Tanaka A, Sanada K, Miyaho K, et al. The relationship between sleep, gut microbiota, and metabolome in patients with depression and anxiety: a secondary analysis of the observational study. PLoS One. 2023;18(12):e0296047. doi:10.1371/journal.pone.0296047
48. Wang Y, van de Wouw M, Drogos L, et al. Sleep and the gut microbiota in preschool-aged children. Sleep. 2022;45(6):zsac020. doi:10.1093/sleep/zsac020
49. Nagata N, Nishijima S, Miyoshi-Akiyama T, et al. Population-level metagenomics uncovers distinct effects of multiple medications on the human gut microbiome. Gastroenterology. 2022;163:1038–1052. doi:10.1053/j.gastro.2022.06.070
50. Weersma RK, Zhernakova A, Fu J. Interaction between drugs and the gut microbiome. Gut. 2020;69(8):1510–1519. doi:10.1136/gutjnl-2019-320204
51. Zmora N, Suez J, Elinav E. You are what you eat: diet, health and the gut microbiota. Nat Rev Gastroenterol Hepatol. 2019;16(1):35–56. doi:10.1038/s41575-018-0061-2
52. Yao M, Qu Y, Zheng Y, Guo H. The effect of exercise on depression and gut microbiota: possible mechanisms. Brain Res Bull. 2025;220:111130. doi:10.1016/j.brainresbull.2024.111130
53. Ford DE, Kamerow DB. Epidemiologic study of sleep disturbances and psychiatric disorders: an opportunity for prevention? JAMA. 1989;262:1479–1484. doi:10.1001/jama.1989.03430110069030
54. Seow LSE, Verma SK, Mok YM, et al. Evaluating DSM-5 insomnia disorder and the treatment of sleep problems in a psychiatric population. J Clin Sleep Med. 2018;14:237–244. doi:10.5664/jcsm.6942
55. Matenchuk BA, Mandhane PJ, Kozyrskyj AL. Sleep, circadian rhythm, and gut microbiota. Sleep Med Rev. 2020;53:101340. doi:10.1016/j.smrv.2020.101340
56. Santos-Marcos JA, Rangel-Zuñiga OA, Jimenez-Lucena R, et al. Influence of gender and menopausal status on gut microbiota. Maturitas. 2018;116:43–53. doi:10.1016/j.maturitas.2018.07.008
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Effects of Empagliflozin on Gut Microbiota in Heart Failure with a Preserved Ejection Fraction: The Design of a Pragmatic Randomized, Open-Label Controlled Trial (EMPAGUM)
Guan XQ, Wang CH, Cheng P, Fu LY, Wu QJ, Cheng G, Guan L, Sun ZJ
Drug Design, Development and Therapy 2023, 17:1495-1502
Published Date: 18 May 2023
Alterations in Gut Microbiota and Serum Metabolites in Children with Mycoplasma pneumoniae Pneumonia
Wang S, Liu C, Ding R, Wang S, Ye Y, He M
Infection and Drug Resistance 2024, 17:5097-5110
Published Date: 20 November 2024
Disruption of Gut Microbiota and Associated Fecal Metabolites in Collagen-Induced Arthritis Mice During the Early Stage

Wu D, Yin M, Cao D, Zhang X, Zhu Y, Wei Y, Li Y, Wen C, Zhou J
Journal of Inflammation Research 2025, 18:1703-1717
Published Date: 4 February 2025
Synergistic Effect of Acupuncture and Traditional Chinese Medicine on Cerebral Infarction in Rats: Roles of Short-Chain Fatty Acids and Interleukin-17
Kang Z, Lin P, Chen Z, Ye H, Fang L, Zhang P
International Journal of General Medicine 2025, 18:7231-7242
Published Date: 3 December 2025