Back to Journals » Clinical Interventions in Aging » Volume 20
The Protective Activity of Apigenin Against Bone and Cartilage Diseases
Received 19 March 2025
Accepted for publication 5 August 2025
Published 13 August 2025 Volume 2025:20 Pages 1235—1251
DOI https://doi.org/10.2147/CIA.S529148
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Zhi-Ying Wu
Jincai Chen,1 Xiaofei Liao,2 Longhuo Wu3
1Department of Orthopedics, First Affiliated Hospital of Gannan Medical University, Ganzhou, 341000, People’s Republic of China; 2Department of Pharmacy, Ganzhou People’s Hospital, Ganzhou, 341000, People’s Republic of China; 3School of Pharmacy, Gannan Medical University, Ganzhou, 341000, People’s Republic of China
Correspondence: Longhuo Wu, Email [email protected]
Abstract: Bone and cartilage diseases have become the leading causes of joint disability due to the destruction of bone and cartilage. No effective drugs are available to cure bone and cartilage diseases. Exploring natural compounds as therapeutic alternatives shows promise. Apigenin, a naturally occurring flavonoid, exhibits various pharmacological activities, such as anti-inflammation, antioxidant, and immune modulation. Apigenin acts as a phytoestrogen and mediates the balance of bone remodeling by inducing osteogenic differentiation, promoting osteoblast-associated bone formation, and inhibiting osteoclast-related bone resorption. In addition, apigenin exhibits protective effects against osteoporosis (OP), rheumatoid arthritis (RA), osteoarthritis (OA), gouty arthritis (GA), and intervertebral disc degeneration (IDD). Apigenin can be a promising agent in treating bone and cartilage. However, the application of apigenin is limited due to its low water solubility and bioavailability. More efforts are still needed.
Keywords: apigenin, osteogenic differentiation, osteoporosis, rheumatoid arthritis, osteoarthritis, gouty arthritis
Graphical Abstract:

Introduction
The skeletal system provides the framework for body structure and movement. Bones undergo continuous remodeling by replacing old bones with new bones, and bone remodeling is maintained by the balance between osteoblast-mediated bone formation and osteoclast-regulated bone resorption. The incidence of degenerative bone and cartilage diseases is increasingly high due to the aging population.1 Aging and estrogen deficiency may lead to the development of bone diseases, such as osteoporosis (OP),2 rheumatoid arthritis (RA),3 and osteoarthritis (OA).4 The prevalence of bone and cartilage diseases has caused economic and social stress worldwide. Currently, most therapeutic pharmaceuticals for OA, OP, and RA management are prescribed for symptom improvement.5,6 For example, non-steroidal anti-inflammatory drugs (NSAIDs) are commonly used for joint pain alleviation. Some disease-modifying anti-rheumatic drugs (DMARDs), such as tofacitinib, have been developed for treating RA.7 However, no effective drugs are available to cure them. This can be explained by the absence of a full understanding of the underlying molecular mechanisms. Thus, it is urgent to explore potential targets and effective drugs for managing bone and cartilage diseases.
The dynamic metabolism of bone is balanced by osteoblasts and osteoclasts. Osteoclast-mediated bone resorption is essential for the maintenance, repair, and remodeling of bone tissues. Osteoblasts activate the expression of receptor activator of nuclear factor (NF)-κB ligand (RANKL), which interacts with the RANKL receptor to mediate the differentiation, activation, and survival of osteoclasts.8 Osteoblasts secrete collagen and promote the deposition of calcium, phosphate, and other ions, leading to the formation of new bones. Osteoclasts digest proteins and minerals by upregulating the expression of tartrate-resistant acid phosphatase (TRAP).9 Chondrocytes, the unique cell type in the articular cartilage, maintain the structure and function of cartilage as well as extracellular matrix (ECM) metabolism.10 The pathological alterations of OA cartilage are associated with excessive chondrocyte cell death and ECM degradation.11 The main components of ECM include type II collagen and aggrecan. Particularly, type II collagen forms a network by interacting with proteoglycan aggregate, which is constituted by the large aggregating proteoglycan and aggrecan to a hyaluronan (HA) filament.12 The catabolic factor matrix metalloproteinases (MMPs), particularly MMP-13, are mainly responsible for the degradation of type II collagen, and a disintegrin and metalloproteinases with thrombospondin motifs (ADAMTSs) can degrade aggrecan.13
Flavonoids, the secondary metabolites in natural plants and nutraceuticals, have a common carbon skeleton of diphenylpropane, which is characterized by two benzene rings (A and B rings) connected by a three-carbon linking chain that forms a heterocyclic ring (C ring) containing oxygen. The B ring is commonly attached to position 2 in the C ring. However, position 3 or 4 can also be available for the B ring.14 Flavonoids exhibit protective effects on bone remodeling.15 Apigenin, also called 4’,5,7-trihydroxyflavone with the formula of C15H10O5 and a molecular weight of 270.05 g/mol, has been abundantly found in vegetables and fruits, such as celery, parsley, pistachio, onions, apple, oranges, and chamomile.16 In nature, apigenin is mainly present in O- and C-glycosidic forms, such as apigenin-8-C-glucoside (also known as vitexin), and apigenin-6-C-glucoside (also known as isovitexin)17–19 (Figure 1).
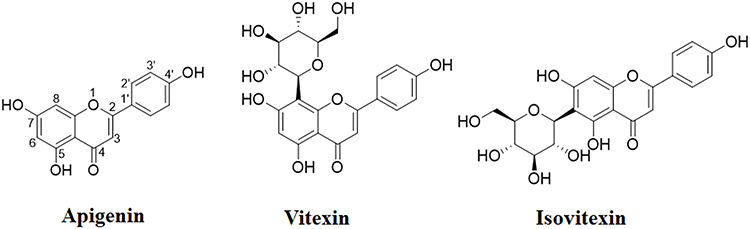 |
Figure 1 The chemical structures of apigenin, vitexin, and isovitexin. |
Botanical preparations have been proven to be safe and effective in treating bone and cartilage diseases.15,20 Supplemental natural flavonoids as nutraceuticals are increasingly recognized for treating many diseases. Ramesh et al have reviewed the functions and molecular mechanisms of flavonoids in mediating bone remodeling and suggested flavonoids as the promising agents in treating bone-related ailments in the future.15 Some flavonoids can function as phytoestrogens due to their structural similarity to estrogen and their ability to interact with estrogen receptor. The estrogen-like effects of flavonoids favor anabolism in the tissues of bone and cartilage.21 Apigenin is one of the important natural phytoestrogens derived from plants and foods.22 Phytoestrogens can interact with estrogen receptors due to their structural similarity with 17β-estradiol.23 Apigenin has various pharmacological activities, such as anti-inflammation,24,25 antioxidant,26 immune modulation,27 and bone protection effects.17 In this article, we mainly discuss the protective effects of apigenin and its glycosides against bone diseases.
The Protective Activities of Apigenin Against OP Development
OP is associated with decreased bone mass and increased bone fragility, and it is marked as a progressive systemic skeletal disease by the World Health Organization (WHO). Approximately 536,000 OP patients with fragility fractures develop annually. In the European Union, about 22,000,000 women and 5,500,000 men were affected by OP in 2010.28 It has been reported that the total medical expenses for OP increased by 51% from 2015 to 2019 (from USD 155 million to USD 234 million), while the medical expenses per patient increased by 3.7% annually, reaching USD 216.29 OP and the fragility fracture significantly challenge individuals and have become a public health concern worldwide.28
Estrogen deprivation in postmenopausal women induces high bone turnover and loss, resulting in primary OP.30 It is reported that hormone therapy shows efficacy in reduction or suppression of OP-associated fracture and most of the other symptoms related to menopause.30 However, long-term hormone therapy has been associated with a high risk of breast cancer and other diseases.31 Several medications in the clinic include anti-resorptive drugs, such as bisphosphonates, calcitonin, and denosumab, and anabolic agents, such as abaloparatide, romosozumab, and teriparatide. Healthy lifestyle, including nutrition (calcium, protein, vitamin C, and vitamin D) and suitable physical activity, exhibits relative small efficacy, but it may slow bone loss and prevent OP.32 Currently used therapy includes those that inhibit bone resorption, promote bone formation, and dual-action therapies.33 Combinational therapy of anabolic and anti-resorptive agents can produce synergistic effects and become the ideal strategy for OP treatment. However, this therapeutic strategy is not recommended due to the combined adverse effects.34 An alternative therapeutic strategy for OP management is essential.
In ovariectomized (OVX) mice, apigenin treatment can significantly reduce trabecular bone loss in the femurs, maintaining bone homeostasis.35 The protective activity of apigenin against bone loss has been confirmed in OVX rats, as shown by the increased mineral content and density of the trabecular bone and improved trabecular bone microarchitecture.36 Apigenin is one of the bioactive components of Cirsium setidens water extracts (CSEs), which has been shown to promote osteoblastic bone matrix mineralization and suppress osteoclastic bone resorption in OVX mice.37 (Table 1) It has been reported that the apigenin glycosides vitexin and isovitexin are the bioactive components of Acer palmatum hot water extract (APE), which promotes osteoblastogenesis and inhibits osteoclastogenesis in MC3T3-E1 cells.38
 |
Table 1 Summary of the Effects of Apigenin, Vitexin, and Isovitexin on Bone and Cartilage Diseases |
Apigenin Induces Osteogenic Differentiation
Mesenchymal stem cells (MSCs) have multiple directional differentiation potential. Under specific conditions, MSCs can differentiate into osteoblasts, adipocytes, and chondrocytes, and they can be used as important seed cells for gene therapy, regenerative medicine, and tissue engineering.62 Particularly, MSCs derived from bone marrow (BMSCs) can easily differentiate into osteoblasts, stimulate bone formation, and increase bone mass. The introduction of BMSCs into the therapeutic management of bone diseases has shown promising results.63 However, the potential of osteogenic differentiation of BMSCs in OP patients is reduced.64
Various signaling pathways have been involved in mediating osteogenic differentiation.65 For example, Wnt/β-catenin signaling is believed to be integral to osteoblastic differentiation by transcriptionally upregulating the gene expression of runt-related transcription factor 2 (Runx2) via binding to its promoter.66 Activation of β-catenin stimulates the formation of osteoblasts by inducing an increase in alkaline phosphatase (ALP) activity.67 Similarly, activation of the Hedgehog signaling pathway relieves the inhibitory effects of Ptch1 on Smo and activates the expression of nuclear transcription factor Gli1, promoting the osteogenic differentiation of BMSCs.65 BMP pathway is critical for promoting ossification by mediating osteogenic differentiation marker expression, such as ALP, osteopontin (OPN), and OCN.68 Another study shows that Sirt1 activation promotes angiogenesis and osteogenic differentiation in BMP9-induced MSCs.69
It has been reported that apigenin can significantly increase the activity of ALP and mineralization and upregulate the expression of OPN, osterix (OSX), and Runx2. Apigenin promotes osteogenic differentiation of human MSCs by activating the JNK and p38 MAPK signaling pathways.39 (Table 1) Consistently, apigenin can rescue the potential osteogenic differentiation of human BMSCs obtained from elderly patients with OP. Microarray-based gene expression profile in apigenin-treated human BMSCs has been explored. Apigenin promotes osteogenic differentiation by activating the focal adhesion kinase (FAK), ERK, and TGFβ/Smad2 signaling pathways. In addition, apigenin decreases senescence and oxidative stress in TBHP-treated human BMSCs.40 It is well known that Wnt/β-catenin signaling plays a critical role in cellular differentiation and ECM production during skeletal formation and development.70 One study reports that apigenin at the doses of 0.05 and 0.5 μM induces osteogenic differentiation of human MSCs by enhancing the Wnt/β-catenin signaling. Similar effects of apigenin (at the dose of 45 ng/kg for 6 weeks) on osteogenesis in a rat (male and 3-month old) femoral fracture model have been shown.71
Hypoxia plays a critical role in maintaining the pluripotency of stem cells. In human periodontal ligament (HPDL) cells, cobalt chloride (CoCl2) treatment upregulates the mRNA expression of stem-cell markers REX1 and OCT4, down-regulates the expression of osteogenesis-related factors, and suppresses the activities of ALP and mineralization. However, apigenin compromises the effects of CoCl2 on HPDL cells by inhibiting HIF1α activity.41 (Table 1) Increased inflammation regulated by the NLRP3 inflammasome and NF-κB signaling can suppress osteogenesis but promote adipogenesis. In LPS/palmitic acid (PA)-treated human MSCs, apigenin may significantly enhance the osteogenic activity by downregulating the expression of NLRP3, caspase-1, IL-1β, and NF-κB42 (Table 1).
Apigenin Promotes Osteoblast-Mediated Bone Formation
Quantitative structure-activity relationship (QSAR) has been explored to evaluate the bone-forming ability of flavonoids by activating the voltage-gated calcium (CaV) channel in osteoblasts. Apigenin has been predicted to be the bioactive compound in modulating the CaV channel and promoting osteoblast-mediated bone formation.72 Apigenin can enhance the mRNA expression of TGFβ1, TGFβ-R1, TGFβ-R2, TGFβ-R3, BMP-2, BMP-7, RUNX-2, ALP, COL-I, OSX, and OSC in MG63 osteoblasts. In addition, apigenin treatment also increases the mRNA expression of osteoprotegerin (OPG) and RANKL, compared to that in the negative control group. These indicate that apigenin treatment plays a role in maintaining the metabolism homeostasis of osteoblasts.43 (Table 1) Apigenin is one of the bioactive compounds in the leaf extract of Blainvillea acmella (L). Philipson, which shows bone anabolic effects in MC3T3-E1 cells by suppressing oxidative stress.73 Another study reports that an apigenin glycoside from Uraria crinita (L). Desv. ex DC. can promote osteoblast differentiation and stimulate new bone formation and regeneration by upregulating the expression of ALP, Runx2, OPN, and BMP-2 in HOb cells.74
Oxidative stress is associated with the imbalance between antioxidant defenses and ROS production. Oxidative stress has become a cause of various degenerative diseases, such as OP. Particularly, oxidative stress induces bone loss by inducing lipid peroxidation, inhibiting the expression of antioxidant enzymes, suppressing osteoblast differentiation, increasing osteoblast apoptosis, and enhancing osteoclast formation.75 In H2O2-treated MC3T3-E1 cells, apigenin prevents oxidative stress-induced cell damage and promotes osteoblast differentiation, as shown by increased expression of ALP, collagen, OPN, OPG, bone sialoprotein, OSX, osteocalcin (OC), and BMPs (BMP-2, BMP-4, and BMP-7) (Figure 2). Mechanically, the protective activity of apigenin might be associated with activation of the PI3K/AKT and ERK signaling pathways, which are inhibited by H2O2 treatment.44 (Table 1) Osteoblasts have been implicated in mineral formation by promoting ECM protein expression and releasing matrix vesicles. The processes of mineralization require the involvement of many factors, such as annexins (Anx) and tissue-nonspecific alkaline phosphatase (TNAP). Apigenin mediates the mineralization processes by regulating the expression of AnxA6 and TNAP.76
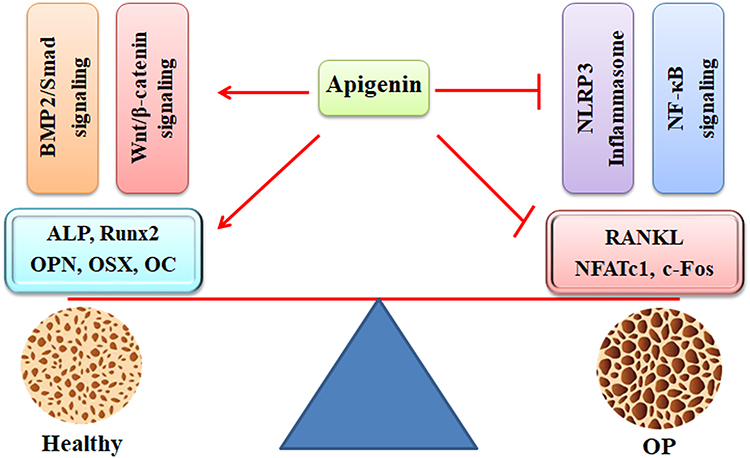 |
Figure 2 The protective activity of apigenin against OP development. Apigenin promotes bone formation by activating the BMP-2/Smad and Wnt/β-catenin signaling pathways, as shown by increased expression of ALP, Runx2, OPN, OSX, and OC. In contrast, apigenin inhibits bone resorption by suppressing the NLRP3 inflammasome and NF-κB signaling pathways, as indicated by decreased expression of RANKL, NFATc1, and c-Fos. Abbreviations: OP, osteoporosis; BMP-2, bone morphogenetic protein-2; ALP, alkaline phosphatase; Runx2, runt-related transcription factor 2; OPN, osteopontin; OSX, osterix; NLRP3, NOD-like receptor family pyrin domain containing 3; NF-κB, nuclear factor kappa-B; RANKL, receptor activator of nuclear factor (NF)-κB ligand; NFATc1, nuclear factor of activated T-cells cytoplasmic 1. |
It has been reported that isovitexin promotes bone formation at the osteotomy site and exhibits bone regenerative effects in a femur osteotomy mouse model. Isovitexin promotes osteoblast differentiation by stimulating mitochondrial biogenesis and respiration. In OVX osteopenic mice, isovitexin rescues bone mass and structure of trabecular bones and cortical bones, as demonstrated by the upregulation of the expression of Runx2, BMP-2, and type I collagen in bones, an increase in serum procollagen type I N-terminal propeptide (PINP), and a decrease in sclerostin and type I collagen cross-linked C-telopeptide (CTX-1).45 (Table 1) Consistently, vitexin promotes the osteoblast differentiation of MC3T3-E1 preosteoblasts, mouse calvarial primary cells, C3H10T1/2, and BMSCs, as shown by enhanced expression of Dlx5, Runx2, ALP, and Smad1/5/9.77
Apigenin Inhibits Osteoclast-Mediated Bone Resorption
Osteoclasts are derived from the hematopoietic bone marrow monocytic/macrophage (BMM) cell lineage and they are known to be capable of resorbing bone. Osteoblasts and osteocytes can produce and secrete RANKL, which can interact with the receptor RANK on BMM precursors, activating the bone-resorbing machinery and the downstream factors NFATc1 and c-Fos.78 (Figure 2) One study shows that inhibition of RANKL may lead to suppression of the NF-κB signaling pathway and NFATc1 expression, leading to attenuation of osteoclast-derived bone resorption.79 OP is a bone disorder with dysregulated inflammatory responses. Estrogen insufficiency may favor Th2 over Th1 in the immune profile, inducing the alteration of inflammatory cytokine ratio, the initiation of bone loss, and the promotion of OP pathogenesis.80 Inflammatory immune regulation can be a potential target for OP management.
Apigenin, one of the bioactive components from Cephalotaxus koreana Nakai, has been reported to inhibit osteoclast differentiation.81 In MC3T3-E1 cells, apigenin suppresses TNFα-induced expression of osteoclastogenic factors, such as IL-6, MCP-1, MCP-3, and RANTES, and inhibits IFNγ-induced secretion of osteoclastogenic monokines CXCL-9 and CXCL-10. In 3T3-L1 cells, apigenin downregulates the expression of IL-6, MCP-1, and leptin. In RAW 264.7 cells, apigenin inhibits RANKL-induced formation of multinucleated cells and suppresses osteoclastogenesis.46 (Table 1) It has been reported that vitexin (a glucoside of apigenin) can significantly inhibit RANKL-induced osteoclast formation and bone resorption by inhibiting the p38 MAPK and ERK signaling pathways and suppressing the expression of NFATc1 and c-Fos in LPS-treated BMM cells. Thus, apigenin and its glucoside may suppress RANKL-mediated osteoclastogenesis and bone resorption.82
The Protective Activities of Apigenin Against RA Development
RA, an autoimmune disease, is correlated with the deregulation of immune cells and synovial fibroblasts, which increase inflammatory responses, induce cartilage destruction, and promote osteoclast differentiation and bone erosion.83,84 Research indicates that the incidence of RA increases with age, peaking between 60 and 70 years. In 2020, an estimated 17.6 million people had RA worldwide, and it is forecasted that the number of RA patients will be 31.7 million by 2050.85 A significant upward trend in the global age-standardised incidence rate (ASIR) of RA is observed for both sexes from 1990 to 2019 (average annual percent change (AAPC): 0.214.86 One study reports that the global RA-related incidence rate increases from 11.66 to 13.48 per 100,000 people from 1990 to 2021. RA-related disability-adjusted life years (DALYs) rate increases from 26.37 to 30.71 per 100,000 people, with females bearing a higher burden.87 RA diagnosis (during the 5 years after diagnosis) has been associated with lower earnings, particularly for older individuals and those with lower education level, by about 5%.88 The annual average cost is $2337.73 per patient with RA in Morocco.89
Various potential biomolecular mechanisms have been proposed. However, the exact cause of RA remains unclear.90 Four classes of drugs, such as NSAIDs, DMARDs, glucocorticoids (GCs), and botanical preparations, have been used for treating RA. DMARDs are recommended by the American College of Rheumatology (ACR) to preserve joint functions.91 NSAIDs and GCs have become supplementary therapies to alleviate inflammatory responses in RA patients. Apigenin has been reported to inhibit the inflammatory responses in LPS-treated RAW 264.7 cells and the angiogenesis in TNFα-treated human umbilical vein endothelial cells (HUVECs).19
It has been reported that apigenin exhibits immunosuppressive activity against collagen-induced arthritis, as shown by reduced severity of pathological alterations and decreased production of pro-inflammatory cytokines in the serum and supernatants from the lymph nodes of the mice with collagen-induced arthritis. In lipopolysaccharide (LPS)-treated bone marrow-derived dendritic cells (DCs), apigenin restrains cell phenotype and maintains cell phagocytotic capability. Apigenin becomes a potent suppressor of DC maturation and migration.47 (Table 1) Consistently, apigenin can significantly reduce the infiltration of inflammation cells, edema formation, and paw thickness and decrease arthritis score in collagen antibody-induced arthritis-induced rat models. In addition, apigenin also decreases the neutrophil-to-lymphocyte ratio (NLR) in whole blood.92 Additionally, it is reported that apigenin can alleviate the arthritic symptoms by inhibiting synovial hyperplasia, angiogenesis, and osteoclast activation in the mice with collagen-induced arthritis. Mechanically, apigenin inhibits the expression of VEGF and VEGFR, suppresses the proliferation of RA-FLSs, promotes the apoptosis of RA-FLSs, and mediates the RANKL/RANK/OPG system in CIA mice48 (Table 1).
It has been reported that apigenin exhibits anti-inflammatory activity and inhibits LPS-activated collagenase activity by decreasing nitric oxide (NO)-mediated cyclooxygenase-2 (COX-2) expression and monocyte adherence.93 One study reports that apigenin can effectively inhibit cyclooxygenase 2 (COX-2) and nuclear factor κB (NF-κB) expression, induce lupus immune cell apoptosis, and suppress lupus development by inhibiting autoantigen presentation for expansion of autoreactive Th1 and Th17 cells. Apigenin may exhibit protective activities against inflammation- or immune-associated diseases, such as RA and OA.94 Apigenin is one of the bioactive compounds from the polar extract of Phoenix dactylifera L., which shows anti-inflammatory and anti-rheumatic effects in CFA-induced arthritis rat models by mediating the IL-23/IL-17/IL-22 axis and the NF-κB/IL-6/JAK1/STAT3 signaling pathway.49 (Figure 3) In addition, apigenin is the bioactive compound of Mansoa alliacea extract, which exhibits anti-inflammatory and anti-nociceptive activities in CFA-treated mice.95 Consistently, isovitexin-contained crude aqueous extract of Cecropia hololeuca Miq. also shows anti-inflammatory and anti-nociceptive effects in zymosan-induced mouse arthritis.96
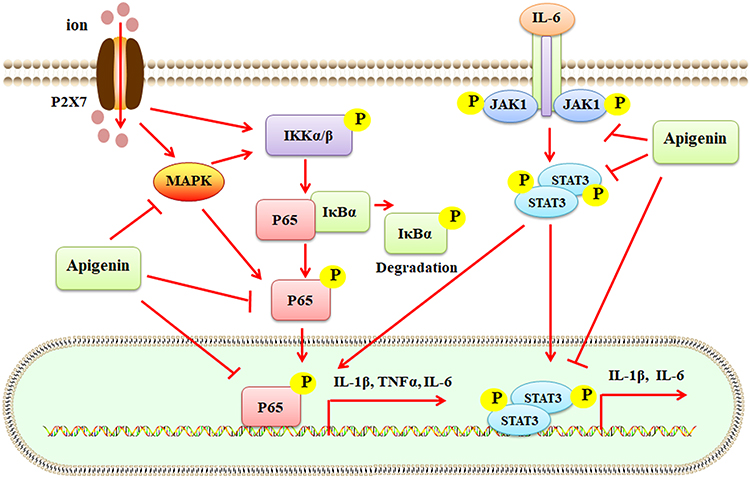 |
Figure 3 Apigenin suppresses inflammatory responses mediated by the P2X7/MAPK/NF-κB and IL-6/JAK1/STAT3 signaling pathways. Ion-activated P2X7 stimulates MAPK and IKKα/β, which induced the activation of p65. Phosphorylated p65 (p-p65) enters the nucleus for transcription regulation of IL-1β, TNFα, and IL-6 expression. IL-6 induces the phosphorylation of JAK1, which promotes the activation of STAT3. Phosphorylated STAT3 (p-STAT3) enters the nucleus for transcription regulation. Apigenin suppresses the activities of MAPK, p-p65, and p-STAT3, leading to inhibition of inflammatory responses. Abbreviations: MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor kappa-B; IL-1β, interleukin-1β; TNFα, tumor necrosis factor-α; JAK1, Janus kinase 1; STAT3, signal transducer and activator of transcription 3. |
P2X7 plays a role in the immune system and inflammatory diseases by mediating the efflux of Na+, Ca2+, and K+ and activating the MPAK/NF-κB signaling pathways (Figure 3). The P2X7/NF-κB signaling has been implicated in the pathological development of arthritis.50 Apigenin has been shown the protective activity against CFA-induced arthritis by suppressing the P2X7/NF-κB signaling and its downstream factors, such as IL-1β, TNFα, and IL-6 in rats.50 (Table 1) TNFα-triggered progressive inflammation has been implicated in the pathogenesis of RA. TNFα treatment increases the expression of IL-6, IL-1β, TTR, and receptors for advanced glycation end products (RAGE) by activating the NF-κB signaling pathway, compared to that in the negative control group in human synovial fibroblast SW982 cells. Apigenin can interact with TNFα and exhibit anti-inflammatory effects against TNFα-stimulated deregulation associated with RA pathogenesis51 (Table 1).
RA fibroblast-like synoviocytes (RA-FLSs) are different from normal FLSs due to their aggressive and invasive phenotype. RA-FLSs can grow in soft agarose in an anchorage-independent manner, and they can invade the articular cartilage in mouse RA models.97 RA-FLSs play a critical role in the pathological development of RA by promoting the synthesis of pro-inflammatory cytokines and upregulating the expression of matrix-degrading enzymes.98 Apigenin suppresses the aberrant proliferation of RA fibroblast-like synoviocytes (RA-FLSs) and decreases the production of NO and IL-6. Apigenin exhibits anti-RA effects by attenuating the activation of the NF-κB signaling and the expression of pro-inflammatory cytokines.52 (Table 1) TNF-related apoptosis-inducing ligand (TRAIL) can promote FLS proliferation by activating the MAPK and PI3K/AKT signaling pathways. However, TRAIL-induced apoptosis only accounts for a small amount of RA-FLSs. This might be associated with the death resistance of RA-FLSs, which exhibit high expression levels of Bcl-2 and Mcl-1. Apigenin can significantly suppress TRAIL-induced RA-FLS proliferation, restore the expression of p21 and p27, and inactivate the PI3K/AKT signaling pathway.99 Apigenin-4’-O-α-L- rhamnoside can decrease the expression of MMP-1, MMP-3, RANKL, and TNFα in RA-FLS cells and inhibit the MAPK signaling pathway.100
In human RA-FLSs, the effects of apigenin (at the dose of 10, 25, 50, and 100 μM) on cell apoptosis have been studied. Particularly, the activity of apigenin (at the dose of 100 μM) at the time-point of 1, 2, 4, and 8 h in promoting cell apoptosis is also investigated. Consequently, apigenin treatment may stimulate a large amount of ROS, stimulate the MAPK-ERK1/2 signaling pathway, and induce cell apoptosis.101 Apigenin, the effective compound of a Chinese herbal medicine Genkwa flos, can inhibit paw edema, decrease arthritis scores, and induce weight loss in complete Freund’s adjuvant (CFA)-treated rats by enhancing the levels of superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) and decreasing the production of NO and malondialdehyde (MDA). The protective effects of apigenin against CFA-induced RA might be associated its antioxidant activity53 (Table 1).
The Protective Activities of Apigenin Against OA Development
OA, characterized by chronic low-grade inflammation, degenerative cartilage erosion, osteophyte hyperplasia, and synovitis, is the leading cause of musculoskeletal pain, disability, and socioeconomic burden. Epidemiological data show that OA affects more than 300 million individuals worldwide, with higher incidence and prevalence in middle-aged and elderly populations.102 The number of years lived with disability (YLDs) for hip and knee OA has been ranked 11th highest in the 2010 Global Burden of Diseases (GBD) survey.103 In the USA, musculoskeletal diseases affect over 121 million people and account for the highest rate of disability among all disease groups.104 The traditional treatment for OA are mainly systemic administration of drugs (such as NSAIDs or opioid-strength painkillers) or end-stage total joint replacement-related surgery.105
Macrophage polarization plays a critical role in OA development.106 It has been reported that apigenin can inhibit macrophage M1 polarization and promote M2 polarization, as shown by reduced levels of IL-1, IL-6, TNFα, and IL-12 and enhanced levels of MG-L1, MG-L2, ARG-1, and IL-10 in cultured bovine articular cartilage explants. Mechanically, apigenin inhibits macrophage polarization-mediated inflammatory response and chondrocyte apoptosis by inhibiting the MAPK and TRPM7/mTOR signaling pathways.54 (Figure 4) One study reports the co-culture of bovine articular cartilage explants with cytokine combinations (including IL-1α, IL-1β, IL-6, IL-17, TNFα, and oncostatin M). The aggrecanase-mediated cleavage of aggrecan is promoted by all cytokine mixtures via MMPs. Particularly, oncostatin M can synergize with IL-1 and TNFα to induce a rapid cleavage of aggrecan and HA. Interestingly, apigenin acts as an inhibitor of hyaluronidase to suppress HA release. This suggests that hyaluronidase has been involved in cartilage matrix degradation, and HA release is associated with high levels of concomitant proteolysis.107
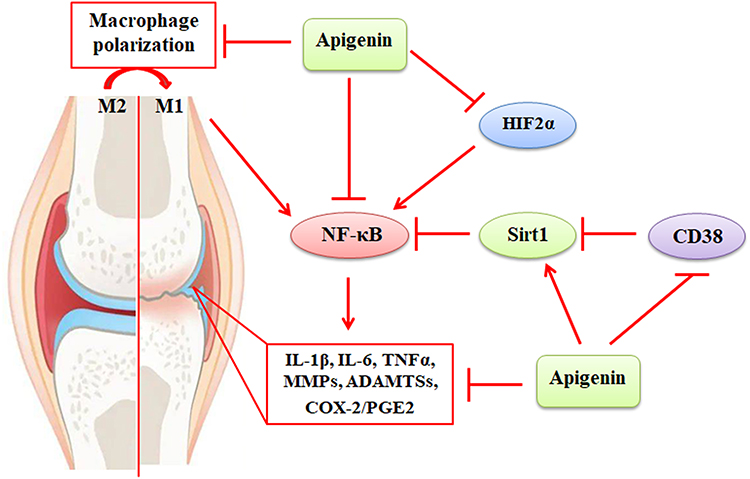 |
Figure 4 The protective activity of apigenin against inflammatory responses in OA. Activation of the NF-κB signaling enhances the expression of IL-1β, IL-6, TNFα, MMPs, ADAMTSs, COX-2, and PGE2, leading to destruction of joint cartilage. The NF-κB signaling can be activated by macrophage M1 polarization, HIF2α, and CD38. These can be compromised by apigenin, which exhibits protective activity against NF-κB-mediated inflammatory responses. Abbreviations: NF-κB, nuclear factor kappa-B; IL-1β, interleukin-1β; TNFα, tumor necrosis factor-α; MMPs, matrix metalloproteinases; ADAMTSs, a disintegrin and metalloproteinases with thrombospondin motifs; COX-2, cyclooxygenase-2; PGE2, prostaglandin E2. |
Cluster of differentiation 38 (CD38), the principal NADase, is ubiquitously expressed in immune cells. CD38-mediated NAD+ degradation is linked to reduced activity of Sirts, and the expression of CD38 is associated with increased inflammatory responses.108 The expression of CD38 is upregulated in human OA cartilage and IL-1β-treated chondrocytes. Overexpression of CD38 decreases cellular NAD+/NADH levels and promotes IL-1β-induced cartilage degradation, synovial inflammation, osteophyte formation, and subchondral bone sclerosis. Apigenin attenuates IL-1β-induced catabolic activities and exhibits chondroprotective effects by mediating the CD38 activity and the Sirt1/NF-κB signaling pathway.55 (Figure 4) (Table 1) Apigenin is one of the bioactive compounds in Vernonia amygdalina (VA), which has been shown to increase the expression of IL-10 and collagen 2a1 and decrease the expression of MMP-3, MMP-13, ADAMTS5, collagen type II degradation biomarker (CTX-II), NF-κB, COX-2, and PGE2.109 OA commonly co-exists with OP in the elderly. Apigenin glycosides can effectively decrease cartilage erosion, bone loss, serum levels of biomarkers, such as IL-1β and osteocalcin (OCN), and mRNA expression of NF-κB, IL-1β, cyclooxygenase-2 (COX-2), and matrix metalloproteinase-13 (MMP-13) in rats.18
The hypoxia‐inducible factor 2α (HIF2α) is a transcription factor for mastering OA pathogenesis by upregulating the expression of MMP-3, MMP-13, COX-2, and other catabolic factors via binding to hypoxia-responsive elements (-CGTG-) in their promoter specifically.110 Apigenin, one of the main bioactive compounds in Cirsium japonicum var. maackii (CJM), has been demonstrated to alleviate HIF2α-induced cartilage destruction and improve DMM-induced mouse OA progression. Apigenin downregulates HIF2α-mediated expression of MMP-3, MMP-13, COX-2, and ADAMTS4 by inhibiting the NF-κB and JNK signaling pathways.111 (Figure 4) Vitexin exhibits anti-inflammatory effects on IL-1β-treated human OA chondrocytes by reducing the levels of pro-inflammatory cytokines, such as IL-6, TNFα, HIF-1α, COX-2, PGE2, MMP-1, MMP-3, and MMP-13.56 (Table 1) In addition, vitexin suppresses IL-1β-induced rat chondrocyte apoptosis by inhibiting the expression of GRP78, PDI, and CHOP, which are the key factors in endoplasmic reticulum (ER) stress. Vitexin also inhibits ER stress-related activation of the NF-κB signaling, inflammation, and ECM degradation in rat chondrocytes. Thus, the protective activity of vitexin, a glucoside of apigenin, might be associated with the inhibition of ER stress in OA chondrocytes.112
The Protective Activities of Apigenin Against Gouty Arthritis (GA)
Gouty arthritis (GA) has been associated with the deregulation of purine metabolism and uric acid excretion. GA is characterized by the accumulation of uric acid crystals in the articular cavity, synovium, cartilage, and bone tissues, leading to joint deformation and stiffness.113 The development of GA might be associated with the interaction of genetic and environmental factors, such as obesity, purine-rich diets, and alcohol consumption. Monosodium urate (MSU) in the joints or urate in the serum may activate the inflammatory responses.114 NSAIDs, GCs, allopurinol, probenecid, and colchicine have been intensively used in the clinic for GA treatment. However, they are limited due to the adverse effects.115 Novel effective therapeutic strategies, such as anti-inflammation, urate reduction, and xanthine oxidase (XOD) inhibition, have been explored.
MSU-induced CD38 expression has been associated with NAD+ degradation and IL-1β and CXCL1 release in gouty mice. However, apigenin acts as an inhibitor of CD38 and reverses CD38-induced activation of the NLRP3 inflammasome and suppression of the Sirt3/SOD2 signaling pathway in bone marrow derived macrophages (BMDMs).57 (Table 1) It has been demonstrated that apigenin exhibits the strongest activity in inhibiting XOD functions (an IC50 value of 30 μM) among the isolated compounds. The aqueous extract of Selaginella moellendorffii Hieron containing apigenin has been reported to improve hyperuricemia, reduce gouty joint inflammatory responses, and suppress oxidative stress in potassium oxonate-treated mice.116 Another study reports that apigenin-7-O-glucoside reduces the serum levels of uric acid, creatinine, and MDA and enhances the activity of SOD in hyperuricemic mice. Mechanically, apigenin-7-O-glucoside inhibits the expression of XOD (an IC50 value of 0.24 μM) and promotes the excretion of uric acid by mediating the renal urate transporters URAT1, GLUT9, OAT1, and ABCG2.117 Isovitexin, a glucoside of apigenin, has been shown to alleviate the joint inflammatory responses in MSU-induced rat GA models by inhibiting the TLR4/MyD88/NF-κB signaling pathway. However, the mechanism of apigenin underlying the anti-inflammatory activity of independent pathways, such as JNK, MAPK, and other signaling pathway, still needs further investigation58 (Table 1).
The Protective Activities of Apigenin Against IDD Development
Intervertebral disc degeneration (IDD), an age-associated musculoskeletal disease, causes low-quality life and high clinical and socioeconomic burdens. The intervertebral disc (IVD), a cylindrical avascular connective tissue, is constituted by a central nucleus pulposus (NP), a peripheral annulus fibrosus (AF), and a cartilage endplate (CEP). The NP cells are responsible for the production of ECM metabolism by regulating the expression of proteoglycan and type II collagen. This indicates that the NP cells play a critical role in maintaining the function and flexibility of the discs.118 It has been reported that the dysfunction of NP and AF cells has been correlated with the pathological development of IDD by inducing abnormal ECM metabolism.119 A variety of factors, such as inflammation, oxidative stress, abnormal mechanical stress, and immune deregulation contribute to IVD cell apoptosis and ECM composition alterations.120
One study reports that the levels of the pro-inflammatory cytokines, such as TNFα, IL-1β, IL-2, IL-6, IL-8, and IL-17 are significantly enhanced in IDD rats. In addition, the catabolic factors, such as MMP-3, MMP-9, ADAMTS-4, syndecan-4, and COX-2/PGE2, are also upregulated. Apigenin has been reported to inhibit inflammatory responses and ECM degradation in the IDD rats and NP cells.59 (Table 1) Autophagy is a dynamic process for scavenging the senescent organelles and misfolded proteins. One study reports that impaired lysosomal biogenesis and autophagic flux promote IDD development.121 Tert-butyl hydroperoxide (TBHP) may induce autophagosome and lysosome fusion interruption and lysosomal dysfunction by decreasing the nuclear translocation of TFEB, the controller of autophagosome. Apigenin can reverse the decreased nuclear translocation of TFEB induced by TBHP. Particularly, apigenin can inhibit TBHP-induced NP cell apoptosis and senescence and ECM degradation by activating the AMPK/mTOR/TFBE signaling pathway and restoring autophagic flux.60 Thus, increasing autophagy has become a potential strategy for IDD treatment, and this suggests a new insight into understanding the potential of apigenin in the treatment of IDD (Table 1).
The Protective Activities of Apigenin Against Other Bone Diseases
Osteonecrosis of the femoral head (ONFH) is characterized by progressive alterations in the hip structure affected by various factors, such as GC use. The bioinformatics analysis shows that thirty-one hub genes are enriched in the AMPK pathway, TLR pathway, and immunity. Two ferroptosis-related factors NCF2 and SLC2A1 have been identified the potential diagnostic biomarkers. Vitexin, an apigenin glucoside, has been reported to promote osteoblast differentiation by inhibiting oxidative stress and the HIF-1 signaling pathway in dexamethasone (Dex)-treated MC3T3-E1 cells.122
Osteosarcoma (OS), the most common bone cancer, affects children and adolescents worldwide. The etiology of OS remains unclear, and the prognosis is poor due to the recurrence and metastasis.123,124 Apigenin has been shown to promote the mitochondrial apoptosis of OS U2OS cells and exhibit anticancer activity.125 The Wnt/β-catenin signaling plays a role in cell morphogenesis, differentiation, and proliferation. It has been reported that apigenin suppresses cell proliferation and reduces cell invasion by inhibiting the Wnt/β-catenin signaling in U2OS and MG63 cells.126 Consistently, apigenin suppresses the proliferation and epithelial-mesenchymal transition (EMT) of SOSP-9607 cells by inhibiting the PI3K/Akt/mTOR signaling-mediated Warburg effects61 (Table 1).
Clinical Perspectives
There are several clinical trials for apigenin reported on the website of clinicaltrials.gov. A single-center, single-blind, randomized, placebo-control pilot clinical trial (NCT05999682) in elderly septic patients has been performed to determine whether apigenin improves organ dysfunction scores. However, no results have been posted. Similarly, the clinical trial (NCT06560216) to study cryopreservation medium with apigenin in post-thaw human sperm has been conducted. The clinical trial on the protective effects of apigenin against neurodegenerative disorder (NCT05696665) has been documented. However, apigenin has low water solubility and poor oral bioavailability (less than 5% of the oral administered dose), which may significantly limit its clinical applications. Isovitexin has appropriately three-fold higher bioavailability than apigenin and has an osteogenic EC50 value of 620 nM in cultured osteoblasts.45,127 To improve the bioavailability of apigenin, several strategies have been explored. Nanoparticles, such as liposomes, lipid nanocapsules, and polymer-based nanocapsules have been prepared to improve the bioavailability of apigenin.16 For example, a novel formulation of apigenin loaded into nanoparticles has been developed for treating dry eye diseases.128 The self-nanoemulsifying drug delivery system (SNEDDS) has been employed to improve the dissolution and permeability of apigenin and promote its antioxidant activity.129 However, the new strategies for improving apigenin bioavailability in treating bone diseases are rarely reported.
The effects of apigenin on bone healing of critical-size defects in rat calvaria have been investigated. Apigenin treatment by gavage promotes the new formation of bone trabeculae, as shown by increased secretion of osteoid matrix and mineralization. Mechanically, apigenin enhances the expression of Runx-2, Smad5, Coll1, Coll4, and Coll5 in human dental pulp stem cells (hDPSCs).130 Bioactive mesoporous glasses (MBGs) have been used for treating bone defects. One study shows that the 3D scaffolds using MBGs loaded with extracts, rich in apigenin and kaempferol, promote bone metabolism, as indicated by improved cytocompatibility and enhanced expression of Runx-2, ALP, and biomineralization in MC3T3-E1 cells.131
MSC administration has been used as a cell-based therapeutic strategy for OA treatment, and it shows promising effects on cartilage regeneration. One study reports that a combination of apigenin and synovial membrane-derived MSC (SMMSCs) can enhance the therapeutic efficiency of cell therapy with a potential mechanism of anti-inflammation and antioxidant of apigenin. It shows that apigenin decreases the expression of IL-1β, TNFα, inducible nitric oxide synthase (iNOS), MMP-3, and MMP-13 and increases the expression of SOD, MDA, SOX-9, collagen 2a1, and aggrecan, compared to that in the negative control group.132 Co-treatment of MSC and apigenin in the rat knee by intra-articular injection can enhance the production of ECM components and improve histological changes by inhibiting inflammation and oxidative stress.133
One study shows that Sirt3- and SHMT2-mediated mitochondrial pathways have become the potential targets of apigenin in promoting caspase-3-dependent apoptosis in colorectal cancer cells.134 Apigenin is an anti-inflammatory agent that could induce the death of cancer cells using radiotherapy methods with X-ray. The presence of sensitizers and high atomic number nanoparticles coated with apigenin biomaterials can prevent DNA damage repair and increase the likelihood of cancer cell death. A biocompatible gold nanoparticle coated with apigenin (Api@AuNPs) as a sensitizer has been introduced.135
Conclusions
Apigenin, a naturally occurring flavonoid, has been shown the protective activities against bone diseases, such as OP, RA, OA, GA, and IDD, in cells and animals. Apigenin is a phytoestrogen that inhibits bone loss and improves bone microarchitecture. It is valuable that apigenin promotes osteogenic differentiation, induces osteoblast-associated bone formation, and suppresses osteoclast-related bone resorption. The anti-inflammation and antioxidant effects of apigenin facilitate its protection against OP, RA, OA, GA, and IDD. However, many issues still need to be addressed. For example, more potential targets and the related molecular mechanisms of apigenin should be explored. The clinical trials of apigenin in treating human bone diseases should be studied. The different chemical forms of apigenin, such as apigenin glycosides, might be linked to the distinct biological activities, although the sugar parts of these glycosides can be removed and converted into their aglycone moiety apigenin. The effects of apigenin on NAD+ elevation need more investigation, although apigenin acts as an inhibitor of CD38. The novel strategies for improving the bioavailability of apigenin should be further studied, particularly in the field of bone and cartilage diseases.
Data Sharing Statement
The data used to support the findings of this study are included within the article.
Funding
There is no funding to report.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Wu Z, Wang Y, Liu W, Lu M, Shi J. The role of neuropilin in bone/cartilage diseases. Life Sci. 2024;346:122630. doi:10.1016/j.lfs.2024.122630
2. Li J, Chen X, Lu L, Yu X. The relationship between bone marrow adipose tissue and bone metabolism in postmenopausal osteoporosis. Cytokine Growth Factor Rev. 2020;52:88–98. doi:10.1016/j.cytogfr.2020.02.003
3. Komatsu N, Takayanagi H. Mechanisms of joint destruction in rheumatoid arthritis - immune cell-fibroblast-bone interactions. Nat Rev Rheumatol. 2022;18(7):415–429. doi:10.1038/s41584-022-00793-5
4. Sacitharan PK. Ageing and Osteoarthritis. Subcell Biochem. 2019;91:123–159.
5. Gan J, Deng X, Le Y, Lai J, Liao X. The development of naringin for use against bone and cartilage disorders. Molecules. 2023;28(9):3716. doi:10.3390/molecules28093716
6. Ye Y, Zhou J. The protective activity of natural flavonoids against osteoarthritis by targeting NF-κB signaling pathway. Front Endocrinol. 2023;14:1117489. doi:10.3389/fendo.2023.1117489
7. Wollenhaupt J, Lee EB, Curtis JR, et al. Safety and efficacy of tofacitinib for up to 9.5 years in the treatment of rheumatoid arthritis: final results of a global, open-label, long-term extension study. Arthritis Res Ther. 2019;21(1):89. doi:10.1186/s13075-019-1866-2
8. Sobacchi C, Frattini A, Guerrini MM, et al. Osteoclast-poor human osteopetrosis due to mutations in the gene encoding RANKL. Nat Genet. 2007;39(8):960–962. doi:10.1038/ng2076
9. Blair HC, Robinson LJ, Huang CL, et al. Calcium and bone disease. Biofactors. 2011;37(3):159–167. doi:10.1002/biof.143
10. Li W, Shi Z, Jing H, et al. Streamlined metal-based hydrogel facilitates stem cell differentiation, extracellular matrix homeostasis and cartilage repair in male rats. Nat Commun. 2025;16(1):4344. doi:10.1038/s41467-025-59725-y
11. Peng Z, Sun H, Bunpetch V, et al. The regulation of cartilage extracellular matrix homeostasis in joint cartilage degeneration and regeneration. Biomaterials. 2021;268:120555. doi:10.1016/j.biomaterials.2020.120555
12. Fosang AJ, Hardingham TE. Isolation of the N-terminal globular protein domains from cartilage proteoglycans. Identification of G2 domain and its lack of interaction with hyaluronate and link protein. Biochem J. 1989;261(3):801–809. doi:10.1042/bj2610801
13. Mukherjee A, Das B. The role of inflammatory mediators and matrix metalloproteinases (MMPs) in the progression of osteoarthritis. Biomater Biosyst. 2024;13:100090. doi:10.1016/j.bbiosy.2024.100090
14. Fossatelli L, Maroccia Z, Fiorentini C, Bonucci M. Resources for human health from the plant kingdom: the potential role of the flavonoid apigenin in cancer counteraction. Int J Mol Sci. 2023;25(1):251. doi:10.3390/ijms25010251
15. Ramesh P, Jagadeesan R, Sekaran S, Dhanasekaran A, Vimalraj S. Flavonoids: classification, function, and molecular mechanisms involved in bone remodelling. Front Endocrinol. 2021;12:779638. doi:10.3389/fendo.2021.779638
16. Lotfi MS, Rassouli FB. Natural flavonoid apigenin, an effective agent against nervous system cancers. Mol Neurobiol. 2024;61(8):5572–5583. doi:10.1007/s12035-024-03917-y
17. Daneshvar S, Zamanian MY, Ivraghi MS, et al. A comprehensive view on the apigenin impact on colorectal cancer: focusing on cellular and molecular mechanisms. Food Sci Nutr. 2023;11(11):6789–6801. doi:10.1002/fsn3.3645
18. Tantowi N, Mohamed S, Lau SF, Hussin P. Comparison of diclofenac with apigenin-glycosides rich Clinacanthus nutans extract for amending inflammation and catabolic protease regulations in osteoporotic-osteoarthritis rat model. Daru. 2020;28(2):443–453. doi:10.1007/s40199-020-00343-y
19. Zhou DC, Zheng G, Jia LY, et al. Comprehensive evaluation on anti-inflammatory and anti-angiogenic activities in vitro of fourteen flavonoids from Daphne Genkwa based on the combination of efficacy coefficient method and principal component analysis. J Ethnopharmacol. 2021;268:113683. doi:10.1016/j.jep.2020.113683
20. Wu Z, Yang Z, Liu L, Xiao Y. Natural compounds protect against the pathogenesis of osteoarthritis by mediating the NRF2/ARE signaling. Front Pharmacol. 2023;14:1188215. doi:10.3389/fphar.2023.1188215
21. Guo J, Ma J, Cai K, et al. Isoflavones from semen sojae preparatum improve atherosclerosis and oxidative stress by modulating Nrf2 signaling pathway through estrogen-like effects. Evid Based Complement Alternat Med. 2022;2022:4242099. doi:10.1155/2022/4242099
22. Pavlopoulos DT, Myrtsi ED, Tryfinopoulou P, Iliopoulos V, Koulocheri SD, Haroutounian SA. Phytoestrogens as biomarkers of plant raw materials used for fish feed production. Molecules. 2023;28(8):3623. doi:10.3390/molecules28083623
23. Szukiewicz D. Insight into the potential mechanisms of endocrine disruption by dietary phytoestrogens in the context of the etiopathogenesis of endometriosis. Int J Mol Sci. 2023;24(15):12195.
24. Li K, Wu L, Jiang J. Apigenin accelerates wound healing in diabetic mice by promoting macrophage M2-type polarization via increasing miR-21 expression. Mol Cell Biochem. 2024;479(11):3119–31127.
25. Charrière K, Schneider V, Perrignon-Sommet M, et al. Exploring the role of apigenin in neuroinflammation: insights and implications. Int J Mol Sci. 2024;25(9):5041. doi:10.3390/ijms25095041
26. Almeer R, Alyami NM. The protective effect of apigenin against inorganic arsenic salt-induced toxicity in PC12 cells. Environ Sci Pollut Res Int. 2023;30(48):106625–106635. doi:10.1007/s11356-023-29884-w
27. Zhou XW, Wang J, Tan WF. Apigenin suppresses innate immune responses and ameliorates lipopolysaccharide-induced inflammation via inhibition of STING/IRF3 pathway. Am J Chin Med. 2024;52(2):471–492. doi:10.1142/S0192415X24500204
28. Gregson CL, Armstrong DJ, Bowden J, et al. UK clinical guideline for the prevention and treatment of osteoporosis. Arch Osteoporos. 2022;17(1):58. doi:10.1007/s11657-022-01061-5
29. Park JY, Chung YJ, Kim MR, Song JY. Five-year sales trends of osteoporosis medications in Korea: a market analysis based on IMS health sales audit data (2018-2023). Medicina. 2025;61(5):805. doi:10.3390/medicina61050805
30. Li L, Wang Z. Ovarian aging and osteoporosis. Adv Exp Med Biol. 2018;1086:199–215.
31. Levin VA, Jiang X, Kagan R. Estrogen therapy for osteoporosis in the modern era. Osteoporos Int. 2018;29(5):1049–1055. doi:10.1007/s00198-018-4414-z
32. Vázquez-Lorente H, García-Gavilán JF, Shyam S, et al. Mediterranean diet, physical activity, and bone health in older adults: a secondary analysis of a randomized clinical trial. JAMA Network Open. 2025;8(4):e253710. doi:10.1001/jamanetworkopen.2025.3710
33. Akkawi I, Zmerly H. Osteoporosis: current Concepts. Joints. 2018;6(2):122–127. doi:10.1055/s-0038-1660790
34. Kim B, Cho YJ, Lim W. Osteoporosis therapies and their mechanisms of action (Review). Exp Ther Med. 2021;22(6):1379. doi:10.3892/etm.2021.10815
35. Goto T, Hagiwara K, Shirai N, Yoshida K, Hagiwara H. Apigenin inhibits osteoblastogenesis and osteoclastogenesis and prevents bone loss in ovariectomized mice. Cytotechnology. 2015;67(2):357–365. doi:10.1007/s10616-014-9694-3
36. Park JA, Ha SK, Kang TH, et al. Protective effect of apigenin on ovariectomy-induced bone loss in rats. Life Sci. 2008;82(25–26):1217–1223. doi:10.1016/j.lfs.2008.03.021
37. Oh MS, Kim SI, Sim YE, et al. Cirsium setidens water extracts containing linarin block estrogen deprivation-induced bone loss in mice. Int J Mol Sci. 2023;24(2):1620. doi:10.3390/ijms24021620
38. Kuriya K, Nishio M, Wada S, et al. Antiosteoporotic effects of acer palmatum extract on osteoclastogenesis and osteoblastogenesis. J Med Food. 2019;22(4):365–373. doi:10.1089/jmf.2018.4243
39. Zhang X, Zhou C, Zha X, et al. Apigenin promotes osteogenic differentiation of human mesenchymal stem cells through JNK and p38 MAPK pathways. Mol Cell Biochem. 2015;407(1–2):41–50. doi:10.1007/s11010-015-2452-9
40. Ali D, Okla M, Abuelreich S, et al. Apigenin and Rutaecarpine reduce the burden of cellular senescence in bone marrow stromal stem cells. Front Endocrinol. 2024;15:1360054. doi:10.3389/fendo.2024.1360054
41. Osathanon T, Vivatbutsiri P, Sukarawan W, Sriarj W, Pavasant P, Sooampon S. Cobalt chloride supplementation induces stem-cell marker expression and inhibits osteoblastic differentiation in human periodontal ligament cells. Arch Oral Biol. 2015;60(1):29–36. doi:10.1016/j.archoralbio.2014.08.018
42. Asadi A, Goudarzi F, Ghanadian M, Mohammadalipour A. Evaluation of the osteogenic effect of apigenin on human mesenchymal stem cells by inhibiting inflammation through modulation of NF-κB/IκBα. Res Pharm Sci. 2022;17(6):697–706. doi:10.4103/1735-5362.359436
43. Melguizo-Rodríguez L, Manzano-Moreno FJ, Illescas-Montes R, et al. Bone protective effect of extra-virgin olive oil phenolic compounds by modulating osteoblast gene expression. Nutrients. 2019;11(8):1722. doi:10.3390/nu11081722
44. Jung WW. Protective effect of apigenin against oxidative stress-induced damage in osteoblastic cells. Int J Mol Med. 2014;33(5):1327–1334. doi:10.3892/ijmm.2014.1666
45. Pal S, Sharma S, Porwal K, et al. Oral administration of isovitexin, a naturally occurring apigenin derivative showed osteoanabolic effect in ovariectomized mice: a comparative study with teriparatide. Calcif Tissue Int. 2022;111(2):196–210. doi:10.1007/s00223-022-00979-9
46. Bandyopadhyay S, Lion JM, Mentaverri R, et al. Attenuation of osteoclastogenesis and osteoclast function by apigenin. Biochem Pharmacol. 2006;72(2):184–197. doi:10.1016/j.bcp.2006.04.018
47. Li X, Han Y, Zhou Q, et al. Apigenin, a potent suppressor of dendritic cell maturation and migration, protects against collagen-induced arthritis. J Cell Mol Med. 2016;20(1):170–180. doi:10.1111/jcmm.12717
48. Li Y, Yang B, Bai JY, et al. The roles of synovial hyperplasia, angiogenesis and osteoclastogenesis in the protective effect of apigenin on collagen-induced arthritis. Int Immunopharmacol. 2019;73:362–369. doi:10.1016/j.intimp.2019.05.024
49. El-Gendy ZA, Abdelazeem S, Abdel Jaleel GA, et al. Anti-inflammatory and anti-rheumatic effects of Phoenix dactylifera L. (date palm) seed by controlling cytokines and inhibiting JAK1/STAT3 pathway on CFA-induced arthritis rat and its phytochemical profiling. J Ethnopharmacol. 2024;329:118138. doi:10.1016/j.jep.2024.118138
50. Chang X, He H, Zhu L, et al. Protective effect of apigenin on Freund’s complete adjuvant-induced arthritis in rats via inhibiting P2X7/NF-κB pathway. Chem Biol Interact. 2015;236:41–46. doi:10.1016/j.cbi.2015.04.021
51. Monu AP, Saquib M, Biswas S, Biswas S. Targeting TNF-α-induced expression of TTR and RAGE in rheumatoid arthritis: apigenin’s mediated therapeutic approach. Cytokine. 2024;179:156616. doi:10.1016/j.cyto.2024.156616
52. Sun YW, Bao Y, Yu H, et al. Anti-rheumatoid arthritis effects of flavonoids from Daphne genkwa. Int Immunopharmacol. 2020;83:106384. doi:10.1016/j.intimp.2020.106384
53. Zhang CF, Zhang SL, He X, et al. Antioxidant effects of Genkwa flos flavonoids on Freund׳s adjuvant-induced rheumatoid arthritis in rats. J Ethnopharmacol. 2014;153(3):793–800. doi:10.1016/j.jep.2014.03.046
54. Ji X, Du W, Che W, Wang L, Zhao L. Apigenin inhibits the progression of osteoarthritis by mediating macrophage polarization. Molecules. 2023;28(7):2915. doi:10.3390/molecules28072915
55. Gil Alabarse P, Chen LY, Oliveira P, Qin H, Liu-Bryan R. Targeting CD38 to suppress osteoarthritis development and associated pain after joint injury in mice. Arthritis Rheumatol. 2023;75(3):364–374. doi:10.1002/art.42351
56. Yang H, Huang J, Mao Y, Wang L, Li R, Ha C. Vitexin alleviates interleukin-1β-induced inflammatory responses in chondrocytes from osteoarthritis patients: involvement of HIF-1α pathway. Scand J Immunol. 2019;90(2):e12773. doi:10.1111/sji.12773
57. Alabarse PG, Oliveira P, Qin H, et al. The NADase CD38 is a central regulator in gouty inflammation and a novel druggable therapeutic target. Inflamm Res. 2024;73(5):739–751. doi:10.1007/s00011-024-01863-y
58. Sun X, Li P, Qu X, Liu W. Isovitexin alleviates acute gouty arthritis in rats by inhibiting inflammation via the TLR4/MyD88/NF-κB pathway. Pharm Biol. 2021;59(1):1326–1333. doi:10.1080/13880209.2021.1979595
59. Ding F, Li X. Apigenin mitigates intervertebral disc degeneration through the amelioration of tumor necrosis factor α (TNF-α) signaling pathway. Med Sci Monit. 2020;26:e924587.
60. Xie C, Shi Y, Chen Z, et al. Apigenin alleviates intervertebral disc degeneration via restoring autophagy flux in nucleus pulposus cells. Front Cell Dev Biol. 2021;9:787278. doi:10.3389/fcell.2021.787278
61. Shi Y, Lian K, Jia J. Apigenin suppresses the Warburg effect and stem-like properties in SOSP-9607 cells by inactivating the PI3K/Akt/mTOR signaling pathway. Evid Based Complement Alternat Med. 2022;2022:3983637. doi:10.1155/2022/3983637
62. Kim N, Cho SG. Clinical applications of mesenchymal stem cells. Korean J Intern Med. 2013;28(4):387–402. doi:10.3904/kjim.2013.28.4.387
63. Gebraad A, Ohlsbom R, Miettinen JJ, et al. Growth response and differentiation of bone marrow-derived mesenchymal stem/stromal cells in the presence of novel multiple myeloma drug melflufen. Cells. 2022;11(9):1574. doi:10.3390/cells11091574
64. Benisch P, Schilling T, Klein-Hitpass L, et al. The transcriptional profile of mesenchymal stem cell populations in primary osteoporosis is distinct and shows overexpression of osteogenic inhibitors. PLoS One. 2012;7(9):e45142. doi:10.1371/journal.pone.0045142
65. Li N, Mu X, Zhang S, Wang H. Recent advances in the multifaceted mechanisms of catalpol in treating osteoporosis. Front Pharmacol. 2025;16:1560715. doi:10.3389/fphar.2025.1560715
66. Zhu Y, Wang Y, Jia Y, Xu J, Chai Y. Catalpol promotes the osteogenic differentiation of bone marrow mesenchymal stem cells via the Wnt/β-catenin pathway. Stem Cell Res Ther. 2019;10(1):37. doi:10.1186/s13287-019-1143-y
67. Jackson A, Vayssière B, Garcia T, et al. Gene array analysis of Wnt-regulated genes in C3H10T1/2 cells. Bone. 2005;36(4):585–598. doi:10.1016/j.bone.2005.01.007
68. Cheng A, Genever PG. SOX9 determines RUNX2 transactivity by directing intracellular degradation. J Bone Miner Res. 2010;25(12):2680–2689. doi:10.1002/jbmr.174
69. Lu Y, Ma ZX, Deng R, Jiang HT, Chu L, Deng ZL. The SIRT1 activator SRT2104 promotes BMP9-induced osteogenic and angiogenic differentiation in mesenchymal stem cells. Mech Ageing Dev. 2022;207:111724. doi:10.1016/j.mad.2022.111724
70. Duan P, Bonewald LF. The role of the wnt/β-catenin signaling pathway in formation and maintenance of bone and teeth. Int J Biochem Cell Biol. 2016;77(Pt A):23–29. doi:10.1016/j.biocel.2016.05.015
71. Pan FF, Shao J, Shi CJ, Li ZP, Fu WM, Zhang JF. Apigenin promotes osteogenic differentiation of mesenchymal stem cells and accelerates bone fracture healing via activating Wnt/β-catenin signaling. Am J Physiol Endocrinol Metab. 2021;320(4):E760–e771. doi:10.1152/ajpendo.00543.2019
72. Chan K, Leung HCM, Tsoi JK. Predictive QSAR model confirms flavonoids in Chinese medicine can activate voltage-gated calcium (CaV) channel in osteogenesis. Chin Med. 2020;15(1):31. doi:10.1186/s13020-020-00313-1
73. Abdul Rahim R, Jayusman PA, Lim V, et al. Phytochemical analysis, antioxidant and bone anabolic effects of blainvillea acmella (L.) Philipson. Front Pharmacol. 2021;12:796509. doi:10.3389/fphar.2021.796509
74. Mao YW, Lin RD, Hung HC, Lee MH. Stimulation of osteogenic activity in human osteoblast cells by edible Uraria crinita. J Agric Food Chem. 2014;62(24):5581–5588. doi:10.1021/jf5012177
75. Tao ZS, Wu XJ, Yang M, Shen CL. Astaxanthin prevents bone loss in osteoporotic rats with palmitic acid through suppressing oxidative stress. Redox Rep. 2024;29(1):2333096. doi:10.1080/13510002.2024.2333096
76. Mroczek J, Pikula S, Suski S, Weremiejczyk L, Biesaga M, Strzelecka-Kiliszek A. Apigenin modulates AnxA6- and TNAP-mediated osteoblast mineralization. Int J Mol Sci. 2022;23(21):13179. doi:10.3390/ijms232113179
77. Kim KM, Son HE, Min HY, Jang WG. Vitexin enhances osteoblast differentiation through phosphorylation of Smad and expression of Runx2 at in vitro and ex vivo. Mol Biol Rep. 2020;47(11):8809–8817. doi:10.1007/s11033-020-05929-y
78. Teitelbaum SL, Ross FP. Genetic regulation of osteoclast development and function. Nat Rev Genet. 2003;4(8):638–649. doi:10.1038/nrg1122
79. Meng J, Zhang W, Wang C, et al. Catalpol suppresses osteoclastogenesis and attenuates osteoclast-derived bone resorption by modulating PTEN activity. Biochem Pharmacol. 2020;171:113715. doi:10.1016/j.bcp.2019.113715
80. Fujii T, Kitaura H, Kimura K, Hakami ZW, Takano-Yamamoto T. IL-4 inhibits TNF-α-mediated osteoclast formation by inhibition of RANKL expression in TNF-α-activated stromal cells and direct inhibition of TNF-α-activated osteoclast precursors via a T-cell-independent mechanism in vivo. Bone. 2012;51(4):771–780. doi:10.1016/j.bone.2012.06.024
81. Yoon KD, Jeong DG, Hwang YH, Ryu JM, Kim J. Inhibitors of osteoclast differentiation from Cephalotaxus koreana. J Nat Prod. 2007;70(12):2029–2032. doi:10.1021/np070327e
82. Jiang J, Jia Y, Lu X, et al. Vitexin suppresses RANKL-induced osteoclastogenesis and prevents lipopolysaccharide (LPS)-induced osteolysis. J Cell Physiol. 2019;234(10):17549–17560. doi:10.1002/jcp.28378
83. Zhang Z, Liu Y, Liang X, et al. Advances in nanodelivery systems based on apoptosis strategies for enhanced rheumatoid arthritis therapy. Acta Biomater. 2025;197:87–103. doi:10.1016/j.actbio.2025.03.043
84. Kaur S, White S, Bartold M. Periodontal disease as a risk factor for rheumatoid arthritis: a systematic review. JBI Libr Syst Rev. 2012;10(42 Suppl):1–12. doi:10.11124/jbisrir-2012-288
85. Black RJ, Cross M, Haile LM; Collaborators GRA. Global, regional, and national burden of rheumatoid arthritis, 1990-2020, and projections to 2050: a systematic analysis of the global burden of disease study 2021. Lancet Rheumatol. 2023;5(10):e594–e610. doi:10.1016/S2665-9913(23)00211-4
86. Chen C, Shi Q, He W, Tian H, Ye T, Yang Y. Global trends in the burden of rheumatoid arthritis by sociodemographic index: a joinpoint and age-period-cohort analysis based on the global burden of disease study 2019. BMJ Open. 2024;14(11):e082966. doi:10.1136/bmjopen-2023-082966
87. Zhang Z, Gao X, Liu S, et al. Global, regional, and national epidemiology of rheumatoid arthritis among people aged 20-54 years from 1990 to 2021. Sci Rep. 2025;15(1):10736. doi:10.1038/s41598-025-92150-1
88. Miller H, Neovius M, Askling J, Bruze G. Impact of incident rheumatoid arthritis on earnings: a nationwide sibling comparison study. Rheumatology. 2025;64(6):3879–3883. doi:10.1093/rheumatology/keae535
89. Zarrik H, Hassani AC, Rkain H, Allali F, Bahiri R, Ahid S. Indirect costs assessment and intangible costs description of rheumatoid arthritis patients with biological therapy in Morocco: ECORAM Study. Int J Rheum Dis. 2024;27(10):e15367. doi:10.1111/1756-185X.15367
90. Wang Z, Wang S, Wang K, Wu X, Tu C, Gao C. Stimuli-sensitive nanotherapies for the treatment of osteoarthritis. Macromol Biosci. 2021;21(11):e2100280. doi:10.1002/mabi.202100280
91. England BR, Smith BJ, Baker NA, et al. 2022 American college of rheumatology guideline for exercise, rehabilitation, diet, and additional integrative interventions for rheumatoid arthritis. Arthritis Care Res. 2023;75(8):1603–1615. doi:10.1002/acr.25117
92. Jin CH, So Y, Nam B, Han SN, Kim JB. Isoegomaketone alleviates the development of collagen antibody-induced arthritis in male Balb/c mice. Molecules. 2017;22(7):1209. doi:10.3390/molecules22071209
93. Lee JH, Zhou HY, Cho SY, Kim YS, Lee YS, Jeong CS. Anti-inflammatory mechanisms of apigenin: inhibition of cyclooxygenase-2 expression, adhesion of monocytes to human umbilical vein endothelial cells, and expression of cellular adhesion molecules. Arch Pharm Res. 2007;30(10):1318–1327. doi:10.1007/BF02980273
94. Kang HK, Ecklund D, Liu M, Datta SK. Apigenin, a non-mutagenic dietary flavonoid, suppresses lupus by inhibiting autoantigen presentation for expansion of autoreactive Th1 and Th17 cells. Arthritis Res Ther. 2009;11(2):R59. doi:10.1186/ar2682
95. Hamann FR, Brusco I, de Campos Severo G, et al. Mansoa alliacea extract presents antinociceptive effect in a chronic inflammatory pain model in mice through opioid mechanisms. Neurochem Int. 2019;122:157–169. doi:10.1016/j.neuint.2018.11.017
96. Teixeira FM, Coelho MN, José-Chagas FDN, et al. Oral treatments with a flavonoid-enriched fraction from Cecropia hololeuca and with rutin reduce articular pain and inflammation in murine zymosan-induced arthritis. J Ethnopharmacol. 2020;260:112841. doi:10.1016/j.jep.2020.112841
97. Müller-Ladner U, Kriegsmann J, Franklin BN, et al. Synovial fibroblasts of patients with rheumatoid arthritis attach to and invade normal human cartilage when engrafted into SCID mice. Am J Pathol. 1996;149(5):1607–1615.
98. Pap T, Müller-Ladner U, Gay RE, Gay S. Fibroblast biology. Role of synovial fibroblasts in the pathogenesis of rheumatoid arthritis. Arthritis Res. 2000;2(5):361–367. doi:10.1186/ar113
99. Sun QW, Jiang SM, Yang K, Zheng JM, Zhang L, Xu WD. Apigenin enhances the cytotoxic effects of tumor necrosis factor-related apoptosis-inducing ligand in human rheumatoid arthritis fibroblast-like synoviocytes. Mol Biol Rep. 2012;39(5):5529–5535. doi:10.1007/s11033-011-1356-3
100. Cao D, Fan Q, Li Z, et al. Transcriptomic profiling revealed the role of apigenin-4’-O-α-L-rhamnoside in inhibiting the activation of rheumatoid arthritis fibroblast-like synoviocytes via MAPK signaling pathway. Phytomedicine. 2022;102:154201. doi:10.1016/j.phymed.2022.154201
101. Shin GC, Kim C, Lee JM, et al. Apigenin-induced apoptosis is mediated by reactive oxygen species and activation of ERK1/2 in rheumatoid fibroblast-like synoviocytes. Chem Biol Interact. 2009;182(1):29–36. doi:10.1016/j.cbi.2009.07.016
102. Safiri S, Kolahi AA, Smith E, et al. Global, regional and national burden of osteoarthritis 1990-2017: a systematic analysis of the global burden of disease study 2017. Ann Rheum Dis. 2020;79(6):819–828. doi:10.1136/annrheumdis-2019-216515
103. Cross M, Smith E, Hoy D, et al. The global burden of Hip and knee osteoarthritis: estimates from the global burden of disease 2010 study. Ann Rheum Dis. 2014;73(7):1323–1330. doi:10.1136/annrheumdis-2013-204763
104. Nguyen A, Lee P, Rodriguez EK, Chahal K, Freedman BR, Nazarian A. Addressing the growing burden of musculoskeletal diseases in the ageing US population: challenges and innovations. Lancet Healthy Longev. 2025;6(5):100707. doi:10.1016/j.lanhl.2025.100707
105. Deng W, Wang T, Li L, et al. A review of nanomaterials in osteoarthritis treatment and immune modulation. Regen Biomater. 2025;12:rbaf048.
106. Sun Y, Zuo Z, Kuang Y. An emerging target in the battle against osteoarthritis: macrophage polarization. Int J Mol Sci. 2020;21(22):8513. doi:10.3390/ijms21228513
107. Durigova M, Roughley PJ, Mort JS. Mechanism of proteoglycan aggregate degradation in cartilage stimulated with oncostatin M. Osteoarthritis Cartilage. 2008;16(1):98–104. doi:10.1016/j.joca.2007.05.002
108. Chini CCS, Peclat TR, Warner GM, et al. CD38 ecto-enzyme in immune cells is induced during aging and regulates NAD(+) and NMN levels. Nat Metab. 2020;2(11):1284–1304. doi:10.1038/s42255-020-00298-z
109. Madzuki IN, Lau SF, Abdullah R, Mohd Ishak NI, Mohamed S. Vernonia amygdalina inhibited osteoarthritis development by anti-inflammatory and anticollagenase pathways in cartilage explant and osteoarthritis-induced rat model. Phytother Res. 2019;33(7):1784–1793. doi:10.1002/ptr.6366
110. Yang S, Kim J, Ryu JH, et al. Hypoxia-inducible factor-2alpha is a catabolic regulator of osteoarthritic cartilage destruction. Nat Med. 2010;16(6):687–693. doi:10.1038/nm.2153
111. Cho C, Kang LJ, Jang D, et al. Cirsium japonicum var. maackii and apigenin block Hif-2α-induced osteoarthritic cartilage destruction. J Cell Mol Med. 2019;23(8):5369–5379. doi:10.1111/jcmm.14418
112. Xie CL, Li JL, Xue EX, et al. Vitexin alleviates ER-stress-activated apoptosis and the related inflammation in chondrocytes and inhibits the degeneration of cartilage in rats. Food Funct. 2018;9(11):5740–5749. doi:10.1039/C8FO01509K
113. Lyu S, Ding R, Liu P, et al. LC-MS analysis of serum for the metabolomic investigation of the effects of pulchinenoside b4 administration in monosodium urate crystal-induced gouty arthritis rat model. Molecules. 2019;24(17):3161. doi:10.3390/molecules24173161
114. Martin WJ, Herst PM, Chia EW, Harper JL. Sesquiterpene dialdehydes inhibit MSU crystal-induced superoxide production by infiltrating neutrophils in an in vivo model of gouty inflammation. Free Radic Biol Med. 2009;47(5):616–621. doi:10.1016/j.freeradbiomed.2009.05.035
115. Simon Taylor R. BET 1: prednisolone for the treatment of acute gouty arthritis. Emerg Med J. 2017;34(10):687–689. doi:10.1136/emermed-2017-207129.1
116. Zhao P, Chen KL, Zhang GL, Deng GR, Li J. Pharmacological basis for use of selaginella moellendorffii in gouty arthritis: antihyperuricemic, anti-inflammatory, and xanthine oxidase inhibition. Evid Based Complement Alternat Med. 2017;2017(1):2103254. doi:10.1155/2017/2103254
117. Zhang Y, Li Y, Li C, et al. Paeonia × suffruticosa Andrews leaf extract and its main component apigenin 7-O-glucoside ameliorate hyperuricemia by inhibiting xanthine oxidase activity and regulating renal urate transporters. Phytomedicine. 2023;118:154957. doi:10.1016/j.phymed.2023.154957
118. Huang J, Zhou Q, Ren Q, Luo L, Ji G, Zheng T. Endoplasmic reticulum stress associates with the development of intervertebral disc degeneration. Front Endocrinol. 2022;13:1094394. doi:10.3389/fendo.2022.1094394
119. Feng C, Yang M, Lan M, et al. ROS: crucial intermediators in the pathogenesis of intervertebral disc degeneration. Oxid Med Cell Longev. 2017;2017(1):5601593. doi:10.1155/2017/5601593
120. Zhang XB, Hu YC, Cheng P, et al. Targeted therapy for intervertebral disc degeneration: inhibiting apoptosis is a promising treatment strategy. Int J Med Sci. 2021;18(13):2799–2813. doi:10.7150/ijms.59171
121. Niu Z, Tang G, Wang X, et al. Trigonochinene E promotes lysosomal biogenesis and enhances autophagy via TFEB/TFE3 in human degenerative NP cells against oxidative stress. Phytomedicine. 2023;112:154720. doi:10.1016/j.phymed.2023.154720
122. Lu H, Fan Y, Yan Q, et al. Identification and validation of ferroptosis-related biomarkers in steroid-induced osteonecrosis of the femoral head. Int Immunopharmacol. 2023;124(Pt A):110906. doi:10.1016/j.intimp.2023.110906
123. Picci P. Osteosarcoma (osteogenic sarcoma). Orphanet J Rare Dis. 2007;2(1):6. doi:10.1186/1750-1172-2-6
124. Ferguson WS, Goorin AM. Current treatment of osteosarcoma. Cancer Invest. 2001;19(3):292–315. doi:10.1081/CNV-100102557
125. Lin CC, Chuang YJ, Yu CC, et al. Apigenin induces apoptosis through mitochondrial dysfunction in U-2 OS human osteosarcoma cells and inhibits osteosarcoma xenograft tumor growth in vivo. J Agric Food Chem. 2012;60(45):11395–11402. doi:10.1021/jf303446x
126. Liu X, Li L, Lv L, Chen D, Shen L, Xie Z. Apigenin inhibits the proliferation and invasion of osteosarcoma cells by suppressing the Wnt/β-catenin signaling pathway. Oncol Rep. 2015;34(2):1035–1041. doi:10.3892/or.2015.4022
127. Pal S, Singh M, Porwal K, et al. Adiponectin receptors by increasing mitochondrial biogenesis and respiration promote osteoblast differentiation: discovery of isovitexin as a new class of small molecule adiponectin receptor modulator with potential osteoanabolic function. Eur J Pharmacol. 2021;913:174634. doi:10.1016/j.ejphar.2021.174634
128. Bonilla-Vidal L, Espina M, García ML, et al. Novel nanostructured lipid carriers loading Apigenin for anterior segment ocular pathologies. Int J Pharm. 2024;658:124222. doi:10.1016/j.ijpharm.2024.124222
129. Morakul B, Teeranachaideekul V, Limwikrant W, Junyaprasert VB. Dissolution and antioxidant potential of apigenin self nanoemulsifying drug delivery system (SNEDDS) for oral delivery. Sci Rep. 2024;14(1):8851. doi:10.1038/s41598-024-59617-z
130. Lorusso F, Scarano A, Fulle S, Valbonetti L, Mancinelli R, Di Filippo ES. Effectiveness of apigenin, resveratrol, and curcumin as adjuvant nutraceuticals for calvarial bone defect healing: an in vitro and histological study on rats. Nutrients. 2023;15(5):1235. doi:10.3390/nu15051235
131. Arias-Rodríguez LI, Pablos JL, Vallet-Regí M, et al. Enhancing osteoblastic cell cultures with gelatin methacryloyl, bovine lactoferrin, and bioactive mesoporous glass scaffolds loaded with distinct parsley extracts. Biomolecules. 2023;13(12):1764. doi:10.3390/biom13121764
132. Estakhri F, Panjehshahin MR, Tanideh N, et al. The effect of kaempferol and apigenin on allogenic synovial membrane-derived stem cells therapy in knee osteoarthritic male rats. Knee. 2020;27(3):817–832. doi:10.1016/j.knee.2020.03.005
133. Estakhri F, Reza Panjehshahin M, Tanideh N, Gheisari R, Azarpira N, Gholijani N. Efficacy of combination therapy with apigenin and synovial membrane-derived mesenchymal stem cells on knee joint osteoarthritis in a rat model. Iran J Med Sci. 2021;46(5):383–394. doi:10.30476/IJMS.2020.83686.1301
134. Abdelmaksoud NM, Abulsoud AI, Abdelghany TM, et al. Uncovering SIRT3 and SHMT2-dependent pathways as novel targets for apigenin in modulating colorectal cancer: in vitro and in vivo studies. Exp Cell Res. 2024;441(1):114150. doi:10.1016/j.yexcr.2024.114150
135. Neshastehriz A, Hormozi-Moghaddam Z, Kichi ZA, Taheri SM, Amini SM, Aghaei A. Overcoming breast cancer cell treatment resistance by optimizing sonodynamic therapy and radiation sensitizers on lncRNA PVT1 and miR-1204 expression. Photodiagnosis Photodyn Ther. 2025;51:104433. doi:10.1016/j.pdpdt.2024.104433
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The m6A/m1A/m5C-Related Methylation Modification Patterns and Immune Landscapes in Rheumatoid Arthritis and Osteoarthritis Revealed by Microarray and Single-Cell Transcriptome
Zheng H, Aihaiti Y, Cai Y, Yuan Q, Yang M, Li Z, Xu K, Xu P
Journal of Inflammation Research 2023, 16:5001-5025
Published Date: 1 November 2023
The Hippo-YAP Signaling Pathway in Osteoarthritis and Rheumatoid Arthritis
Li M, Zhang FJ, Bai RJ
Journal of Inflammation Research 2024, 17:1105-1120
Published Date: 19 February 2024
Hospital-Treated Infections and 15-year Incidence of Musculoskeletal Disorders: A Large Population-Based Cohort Study
Gao Y, McGagh D, Ding L, Hong S, Ouyang Z, Wei J, Zeng C, Lei G, Xie J
Clinical Epidemiology 2025, 17:251-264
Published Date: 11 March 2025
Nanotechnology in Orthopedic Care: Advances in Drug Delivery, Implants, and Biocompatibility Considerations
Xia L, Zhou C, Li Q, Liu L, Jiang C, Dai H, Zhang H, Zhao J, Liang W
International Journal of Nanomedicine 2025, 20:9251-9274
Published Date: 21 July 2025
Bromodomain and Extraterminal Protein Inhibition: A Novel Therapeutic Strategy in Arthritis
Sheng W, Zhao J, Yu F, Wang D, Zeng H, Liu P
Journal of Inflammation Research 2025, 18:13503-13517
Published Date: 30 September 2025