Back to Journals » Journal of Inflammation Research » Volume 18
The Neuroimmune Axis in Atopic Dermatitis: From Pathogenic Mechanisms to Targeted Neuroimmunotherapy
Authors Zhang Z ![]() , Chang C, Xiao L, Su H, Lyu Y, Zhao J
, Chang C, Xiao L, Su H, Lyu Y, Zhao J ![]() , Chen J
, Chen J ![]() , Gou K, Zhou J
, Gou K, Zhou J ![]() , Wang C
, Wang C ![]() , Zhu X, Fu S, Lu M
, Zhu X, Fu S, Lu M ![]() , Shi L
, Shi L
Received 2 November 2025
Accepted for publication 3 December 2025
Published 25 December 2025 Volume 2025:18 Pages 18079—18113
DOI https://doi.org/10.2147/JIR.S578036
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Anish R. Maskey
Zhongsong Zhang,1,2,* Chao Chang,1,2,* Lifan Xiao,1,2,* Hang Su,1,2,* Yaping Lyu,3,* Junxian Zhao,4 Junhao Chen,5 Keyi Gou,2 Jingfeng Zhou,2 Chengjie Wang,6 Xingcheng Zhu,7 Shi Fu,5 Mao Lu,1 Lei Shi8
1Department of Dermatovenereology, Clinical Medical College and The First Affiliated Hospital of Chengdu Medical College, Chengdu, Sichuan, People’s Republic of China; 2School of Clinical Medicine, Chengdu Medical College, Chengdu, 610550, People’s Republic of China; 3Department of Ophthalmology, Clinical Medical College and The First Affiliated Hospital of Chengdu Medical College, Chengdu, Sichuan, People’s Republic of China; 4Department of Urology, 920th Hospital of Joint Logistics Support Force of Chinese People’s Liberation Army, Kunming, Yunnan, People’s Republic of China; 5Department of Urology, The Second Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China; 6School of Stomatology, Xinjiang Second Medical College, Karamay, People’s Republic of China; 7Department of Clinical Laboratory, The Second People’s Hospital of Qujing City, Qujing, Yunnan, People’s Republic of China; 8Department of Clinical Laboratory, Shuguang Hospital Affiliated to Shanghai University of Chinese Traditional Medicine, Shanghai, 201203, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Mao Lu, Email [email protected] Lei Shi, Email [email protected]
Abstract: Atopic dermatitis (AD) is a chronic, relapsing inflammatory skin disease in which intense pruritus and eczematous lesions arise from tightly coupled disturbances of the nervous and immune systems. Emerging evidence indicates that a dysregulated neuroimmune axis—as reflected by peripheral neuronal sensitization, epidermal barrier dysfunction, and central itch processing—plays a central role in disease onset, persistence, and clinical heterogeneity. In this review, we synthesize current knowledge on the neuroimmune mechanisms that drive AD, focusing on how cytokines such as IL-31, IL-4/IL-13, TSLP and IL-33, together with neuropeptides including substance P, CGRP and VIP, establish self-reinforcing itch–scratch and inflammation loops at the level of the skin and the central nervous system. We then highlight recent multi-omics and systems biology approaches, including single-cell and spatial transcriptomics, neuroimaging, and microbiome profiling, that have reshaped the understanding of neuroimmune “neighbourhoods” and the gut–skin–brain axis in AD. Building on these mechanistic insights, we summarize key neuroimmune biomarkers—such as NGF, IL-31, TARC/CCL17, S100 proteins, barrier-related lipids, neurofunctional readouts from fMRI, and microbial signatures—and discuss their potential for improving diagnosis, patient stratification, and treatment monitoring within a precision medicine framework. Finally, we review established and emerging neuroimmune-targeted therapies, including IL-4/IL-13 and IL-31 pathway inhibitors, JAK inhibitors, OX40/OX40L-directed biologics, TRP and NK1R antagonists, phototherapy, and microbiome-based interventions, with a particular emphasis on biomarker-guided sequencing and combination strategies. Overall, we propose that positioning the neuroimmune axis at the core of AD pathogenesis provides a conceptual basis for developing stratified, durable, and patient-centred neuroimmunotherapy, while also outlining the remaining challenges regarding clinical validation of biomarkers, long-term safety, accessibility, and implementation across diverse patient populations.
Keywords: atopic dermatitis, neuroimmune, neuropeptides, skin barrier, precision medicine
Introduction
Atopic dermatitis (AD) is a common chronic, relapsing inflammatory skin disease that affects hundreds of millions of people worldwide and imposes a substantial burden on patients’ quality of life.1,2 Although AD is often colloquially referred to as “eczema”, it represents a specific, well-defined subtype within the broader spectrum of eczematous disorders. Patients typically present with xerosis, intense pruritus, and eczematous lesions that may show erythema, edema, lichenification, excoriations, fissures, and occasional exudation.2,3 Epidemiological studies report a prevalence of approximately 2–10% in the general population, with even higher rates in children (up to 15–30%), underscoring the public health impact of the disease.4–6 The pathogenesis of AD is multifactorial and involves a complex interplay between genetic predisposition, environmental exposures, skin barrier dysfunction, immune dysregulation, and neural alterations.7,8 Genetic variants affecting epidermal barrier proteins, such as filaggrin, increase susceptibility to barrier impairment and allergen penetration, thereby facilitating type 2–skewed immune responses. Environmental triggers—including allergens, microbial exposures, climate factors, irritants, and psychosocial stress—can provoke disease flares or aggravate existing lesions in genetically susceptible individuals.9–11 AD is also closely associated with other atopic comorbidities, such as asthma, allergic rhinitis, and food allergy, a progression often referred to as the “atopic march”, reflecting the presence of a broader systemic immune imbalance.7,12 In addition, it has been reported that an important but overlooked aspect of AD is the bidirectional communication between the nervous and immune systems—known as neuroimmune crosstalk—which can further promote inflammation, reinforce itching, and induce the development of chronic diseases.13–15
In recent years, an important but historically underappreciated dimension of AD pathophysiology has come into focus: the neuroimmune axis.16,17 The skin is a highly innervated and immunologically active organ in which sensory neurons, keratinocytes, and immune cells engage in bidirectional communication to regulate itch and inflammation.18 Patients with AD frequently exhibit increased cutaneous nerve fiber density, elevated levels of neuropeptides such as substance P and calcitonin gene–related peptide, and upregulated nerve growth factor, all of which contribute to peripheral neuronal sensitization.19,20 Conversely, immune-derived mediators—including IL-31 and other type 2 cytokines—can directly activate or sensitize pruriceptive neurons, lowering their activation threshold and promoting chronic pruritus.16 These reciprocal interactions drive a self-sustaining itch–scratch–inflammation cycle that perpetuates barrier damage and disease chronicity.21 Beyond local skin events, psychosocial stress is now recognized as a key exacerbating factor that links the central nervous system to cutaneous immune dysregulation.22 Stress can activate the hypothalamic–pituitary–adrenal axis and the sympathetic nervous system, altering the production of neuroendocrine mediators such as cortisol and catecholamines.21 Dysregulated stress responses may disturb the homeostatic “brain–skin” connection, enhance mast cell activation and cytokine release, and thereby amplify pruritus, inflammation, and disease flares in AD.23 This neuroimmunologic perspective helps explain the tight association between AD severity, sleep disturbance, anxiety, depression, and reduced quality of life.
Despite substantial progress, there is still a lack of an integrated synthesis that brings together molecular and cellular neuroimmune mechanisms, stress-related pathways, clinical phenotypes, biomarker candidates, and emerging targeted treatments within a unified framework.17,24–26 Therefore, this review aims to summarize current knowledge of neuroimmune interactions in AD, including their roles in pathogenesis, itch perception, and stress-related exacerbation, highlight recent methodological advances and neuroimmune biomarkers with potential relevance for disease stratification and monitoring; and discuss existing and emerging therapeutic strategies that target the neuroimmune axis. By doing so, we seek to provide a conceptual and translational basis for future precision medicine approaches in AD, ultimately improving long-term disease control and patients’ quality of life.
Neuroimmune Mechanisms in the Pathogenesis of AD
Understanding the diverse mediators involved in neuroimmune communication is essential for appreciating how these pathways collectively drive the chronicity and clinical heterogeneity of AD. The interactions among sensory neurons, immune cells, keratinocytes, and stress-related neuroendocrine circuits are orchestrated through a wide range of cytokines, neuropeptides, growth factors, and pruritogenic signals. These molecules not only initiate and amplify inflammation and pruritus but also establish self-reinforcing feedback loops that link peripheral barrier dysfunction to systemic immune activation and central nervous system modulation. To provide a clearer and more integrated overview of these mechanisms, the key neuroimmune mediators discussed in this section—including their principal cellular sources and functional roles in AD—are summarized in Table 1 below.
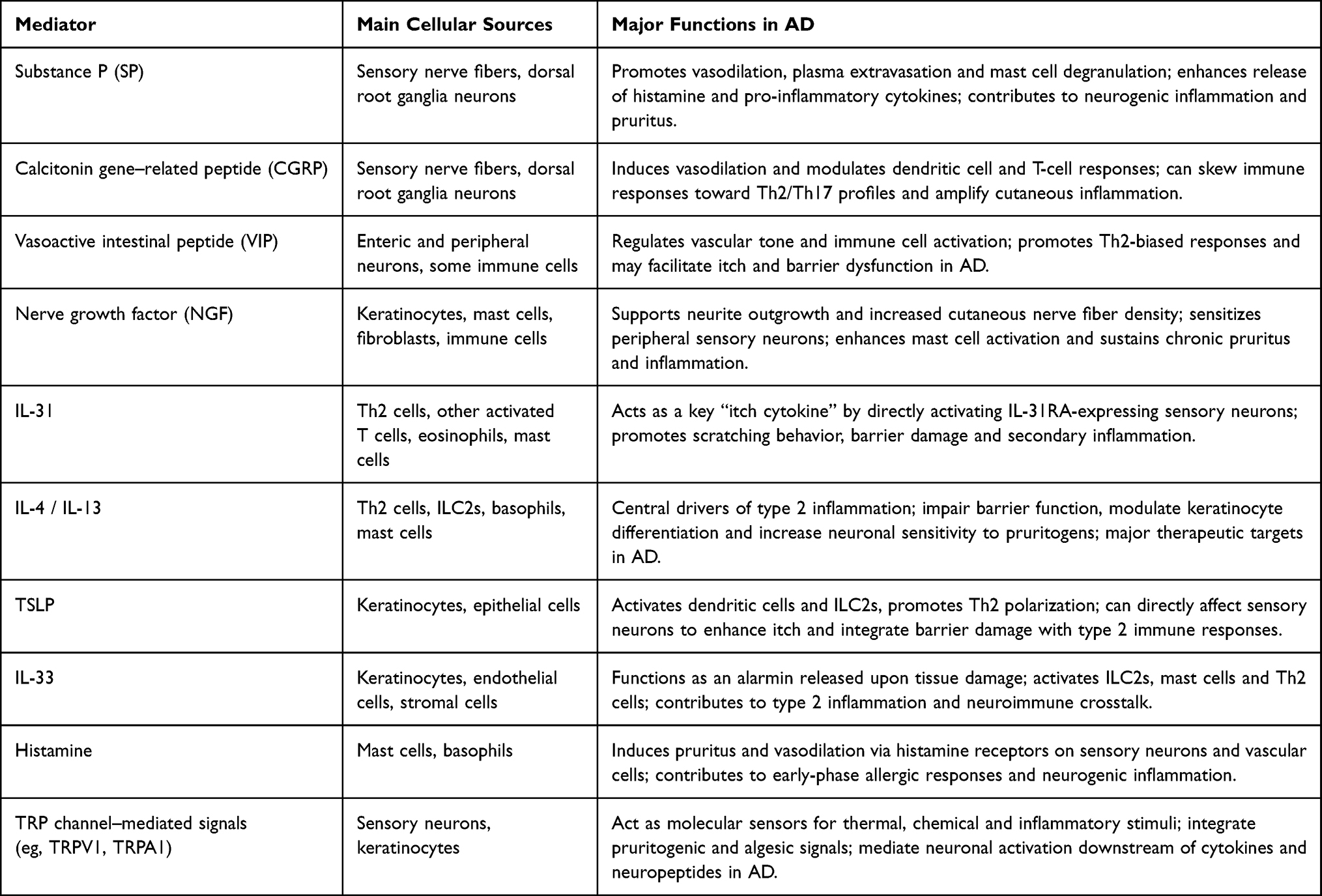 |
Table 1 Key Neuroimmune Mediators in AD: Principal Sources and Functions |
Neuronal Sensitization and Itch Pathway
In AD, the interaction between the nervous and immune systems plays a crucial role in causing itching. The inflammatory chemicals released by immune cells directly affect sensory neurons, causing the itch scratch cycle to continue and worsening the protective barrier of the skin (Figure 1).27 IL-31 is mainly produced by active Th2 cells and binds to receptor complexes on sensory neurons, which are composed of IL-31RA and tumor suppressor M receptor (OSMR). This activates a signaling pathway called JAK/STAT, which increases neuronal activity and triggers itching by altering the way certain proteins work and increasing the production of itch related genes.28 Additionally, we found many clinical studies support this process. For example, in AD patients, higher levels of IL-31 are closely associated with more severe itching. Compared to non-itchy skin areas, the level of IL-31 in itchy skin areas is much higher. This increase leads to the release of more brain natriuretic peptide (BNP) from neurons in the dorsal root ganglion, which in turn causes skin cells to release more cytokines, exacerbating inflammation.29 This IL-31 driven cycle helps cause chronic itching by increasing nerve growth in the skin and making itch fibers more sensitive. The success of nemolizumab, a drug targeting IL-31RA, in clinical trials demonstrates the importance of this pathway. It reduces itching by 50% in patients with moderate to severe AD.30 Another molecule, thymic stromal lymphopoietin (TSLP), is produced by skin cells when the skin barrier is damaged or allergens are present. TSLP activates immune cells and promotes Th2 cell growth by binding to receptors on dendritic cells and T cells. Interestingly, TSLP can also directly activate sensory neurons related to itching, even without immune cells.31 Recent studies have shown that TSLP levels are higher in the skin of AD and increase neuropeptide like substance P (SP) in the nerves. In a mouse model, injection of TSLP causes more scratching, but blocking TSLP receptors prevents this behavior.32 In clinical practice, these findings indicate the potential of the anti TSLP drug tezepelumab, which significantly reduces the severity of eczema and itching in clinical trials. This indicates that it has the ability to disrupt the neuroimmune circuits associated with AD.33
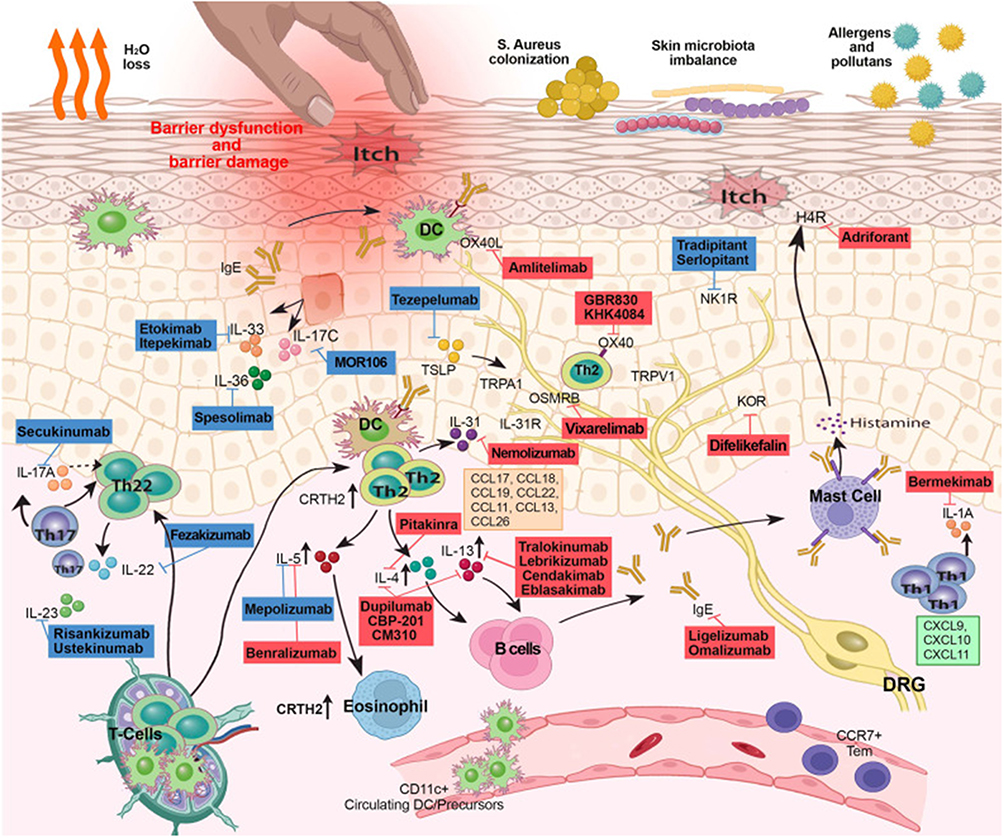 |
Figure 1 This figure illustrates the complex mechanisms of skin inflammation and itching, highlighting barrier dysfunction, immune response, and microbial imbalance. It displays various immune cells (such as T cells, B cells, mast cells, dendritic cells) and cytokines (such as IL-17, IL-31) involved in the pathogenesis of diseases such as AD. The image also details the effects of therapies targeting these immune pathways, including monoclonal antibodies such as dupilumab, mepolizumab, etc. This figure emphasizes the interactions between immune cells, skin barriers, and external factors such as allergens and microorganisms, which can lead to itching and skin damage. Copyright ©2023. Reproduced from Facheris P, Jeffery J, Del Duca E, Guttman-Yassky E. The translational revolution in atopic dermatitis: the paradigm shift from pathogenesis to treatment. Cell Mol Immunol. 2023 May;20(5):448–474. doi: 10.1038/s41423-023-00992-4.34 |
In addition, transient receptor potential (TRP) channels, particularly TRPV1 and TRPA1, play a crucial role in itch conduction, serving as molecular sensors for sensory neurons. These channels are activated by itch inducing agents such as IL-31 and TSLP, triggering calcium influx, membrane depolarization, and the generation of action potentials in itch specific C-fibers.35 For example, in AD, sensitization of TRPV1 by histamine or protease activated receptor 2 (PAR2) agonists can exacerbate itching, as evidenced by upregulation of TRPV1 expression in lesion skin biopsies, which is associated with the severity of itching during disease onset.36 Similarly, TRPA1 channels integrate microbial and environmental signals, including ligands derived from Staphylococcus aureus, to enhance neurogenic inflammation and itching. Consistent with this, TRPA1 knockout mice showed significantly reduced scratching behavior and inflammation in the AD model.29 Moreover, these TRP mediated mechanisms are closely related to IL-31 and TSLP driven pathways, forming a self-reinforcing loop in which cytokine induced neuronal hypersensitivity reduces itch threshold, promotes scratch induced barrier disruption, and maintains chronic inflammation. Longitudinal clinical studies on AD patients have shown that TRP antagonists (such as local capsaicin analogs) can alleviate itching by desensitizing overactive channels, highlighting their transformative potential.37 As for current clinical practice, this comprehensive framework supports the development of novel TRPV1 antagonists. In preclinical AD models, this antagonist significantly reduces itching behavior and epidermal hyperplasia, paving the way for new local formulations targeting the neuroimmune axis to improve patients’ quality of life.38 In addition, protease activated receptor 2 (PAR2) expressed on sensory neurons interacts with proteases derived from mast cells, synergizes with TRP channels, and establishes a feedforward loop of neurogenic inflammation and chronic itching in AD.26 Therefore, from a current perspective, this interconnected network - including IL-31 mediated neuronal overexcitation, TSLP driven immune neuron crosstalk, and TRP dependent signal transduction - highlights the therapeutic potential of neuroimmune targeted interventions aimed at breaking the itch scratch cycle and improving clinical outcomes of AD.26
Immune Cell Neuron Interaction Axis
At present, it is widely believed that the interaction between immune cells and neurons constitutes the central nervous immune axis of AD, amplifying inflammation, itching, and barrier dysfunction through reciprocal signaling between resident immune cells and sensory neurons39 (Figure 2 and Table 2). Specifically, mast cells located near sensory nerve endings undergo degranulation when neurons release SP. SP is a neuropeptide that can activate Mas related G protein coupled receptor X2 (MRGPRX2) on human mast cells. This activation triggers the rapid release of pro-inflammatory mediators such as histamine, TNF - α, and chemokines, which in turn sensitizes sensory neurons and perpetuates the itch scratch cycle.40 In many experimental AD models, SP induced degranulation of mast cells through MRGPRX2 has been shown to enhance vascular permeability and neurogenic inflammation. This was confirmed in the knockout of MRGPRX2 in mice, where intradermal injection of SP significantly increased mast cell activation and vascular leakage, highlighting a feedforward loop that exacerbated erythema and edema in diseased skin.41 In addition, this mechanism links neuroimmune activation with stress-induced disease exacerbation, as epithelial stress factors stimulate nerve endings to release SP, stimulate mast cell degranulation and mediator secretion, thereby enhancing itch perception. These findings have translational significance, indicating that therapeutic inhibition of MRGPRX2 can alleviate rapid allergic reactions and itching episodes in AD patients.42 Consistent with this, clinical studies have shown that MRGPRX2 antagonists may disrupt the interaction between mast cells and nerves, thereby reducing neurogenic inflammation and scratching behavior, with potential benefits for nodular prurigo like variants of AD and overall improvement in patients’ quality of life.40 In addition, eosinophils abundant in AD lesions secrete NGF, which binds to tropomyosin receptor kinase A (TrkA) and p75 neurotrophic factor receptor (p75NTR) on sensory neurons. This special interaction promotes the growth, branching, and hypersensitivity reactions of neurons, thereby exacerbating itching and epidermal hyperplasia.43 For example, in AD patients, elevated levels of NGF derived from eosinophils are associated with disease severity, with increased expression of TrkA and p75NTR on eosinophils compared to allergic rhinitis or non allergic individuals. These changes can inhibit cell apoptosis, enhance chemotaxis, and maintain neuroimmune crosstalk, collectively driving the chronic inflammatory state characteristics of AD.44
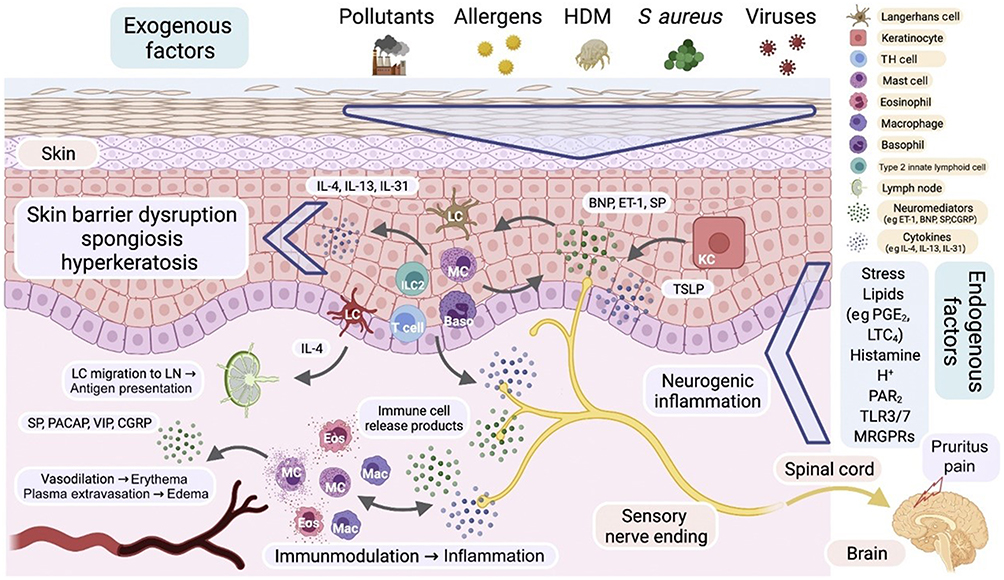 |
Figure 2 Role of neuroimmune interactions in AD. The exogenous factors in the figure (such as pollutants, allergens, dust mites, Staphylococcus aureus, etc). activate local immune responses by disrupting the skin barrier (such as skin barrier damage, sponge like transformation, and hyperkeratosis in the figure), further triggering inflammation and itching. Neuropeptides (such as substance P, calcitonin gene-related peptide [CGRP], and intestinal active peptide [VIP]) and neurotransmitters (such as histamine) are released through sensory nerve endings in the skin, acting on immune cells (such as dendritic cells, mast cells, eosinophils, T cells, and B cells), regulating immune responses, promoting cytokine release (such as IL-4, IL-13), and exacerbating skin inflammation and itching. In addition, neuropeptides enhance local immune responses by activating neural receptors (such as NK1R receptors) and immune cell receptors, forming a neuroimmune feedback loop. Neuroinflammation exacerbates the activation and infiltration of immune cells, leading to the chronicity of AD. Ultimately, the neural immune interaction not only exacerbates skin inflammation and itching symptoms, but may also affect the long-term management and treatment outcomes of the disease. In addition, endogenous factors such as stress, lipids, histamine, and neurotransmitters further enhance neuroimmune interactions, keeping the immune system in AD in a continuously activated state, leading to chronic inflammation and abnormal immune cell activity. Neuropeptides and neurotransmitters promote cytokine release by binding to receptors on immune cells, forming a vicious cycle that further exacerbates inflammation and itching, becoming a key mechanism in the long-term and chronic pathological state of AD. Copyright ©2022. Reproduced from Steinhoff M, Ahmad F, Pandey A et al. Neuroimmune communication regulating pruritus in atopic dermatitis. J Allergy Clin Immunol. 2022 Jun;149(6):1875–1898. doi: 10.1016/j.jaci.2022.03.010.26 |
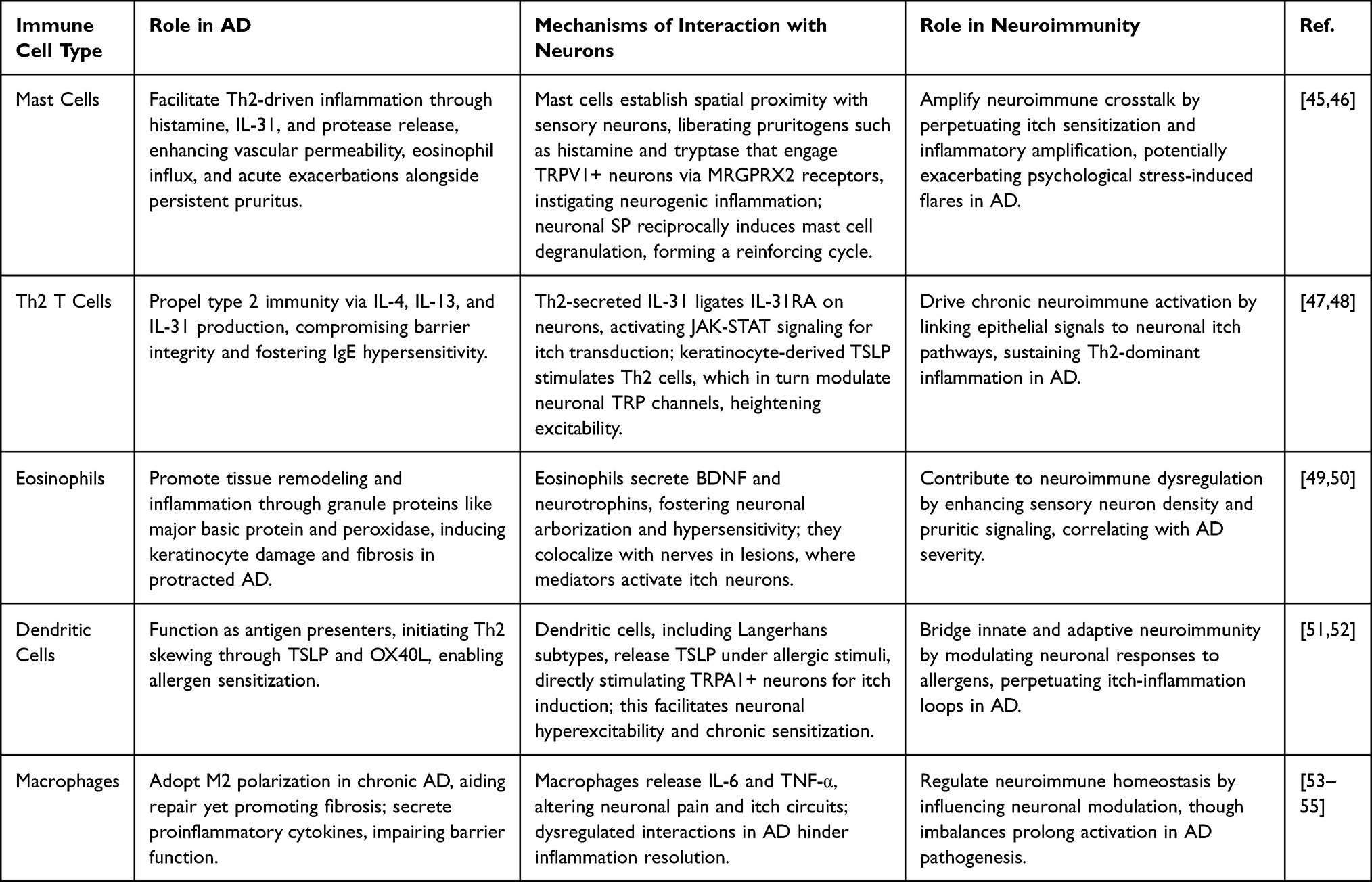 |
Table 2 Summary of Immune Cell-Neuron Interactions in the Neuroimmune Axis of AD |
In addition, this NGF mediated signaling regulates the function of eosinophils, while brain-derived neurotrophic factor (BDNF) - a related neurotrophic factor - further enhances the survival and activation of eosinophils in AD. Experimental evidence suggests that BDNF derived from AD eosinophils promotes the growth of dorsal root ganglion (DRG) neurons, leading to persistent chronic itching and neural remodeling.43 Therefore, targeting the NGF pathway is highly meaningful for clinical translation, as anti NGF antibodies have been shown to reduce skin inflammation and scratching behavior in preclinical AD models, paving the way for biologic therapies to alleviate neuroimmune drive symptoms in refractory AD cases.56 In addition, Th2 cytokines such as IL-4, IL-13, and IL-31 bind to receptors on itch specific sensory neurons and can enhance TRP channels, particularly TRPV1 and TRPA1, directly regulating neuronal excitability, thereby reducing itch threshold and increasing neuronal responsiveness to itch.56 For example, in AD, IL-31 released by Th2 cells binds to IL-31RA on TRPV1 ⁺/TRPA1 ⁺ neurons, activating STAT3 dependent signaling, promoting neuronal proliferation and itch propagation, as shown in a mouse model, where IL-31 overexpression reproduces AD like itch and hair loss.26 In addition, IL-13 exacerbates this process by downregulating epidermal differentiation genes and increasing neuronal sensitivity, which is associated with increased Th2 infiltration and cytokine levels in human AD biopsies, thereby promoting a self-sustaining cycle of inflammation and neuropathic itching.34 Therefore, the interaction between Th2 neurons is the basis for central sensitization and therapeutic resistance, which can be demonstrated by the clinical efficacy of Dupuzumab (an IL-4R α blocker). In Phase III trials of moderate to severe AD, Dupuzumab significantly reduced itch scores by simultaneously targeting immune and neuronal pathways.26 Therefore, these interrelated mechanisms highlight the therapeutic potential of neuroimmunomodulators such as JAK inhibitors and anti cytokine biologics, which can disrupt immune neuron crosstalk and achieve sustained relief and improved patient outcomes by reducing itching and inflammation in AD.57
Neuropeptides - Innate Immune Microcircuit
Neuropeptides, as key regulatory factors of nervous system activity, play a crucial mediating role in the neural control of AD. Currently, an increasing number of evidence suggests that the interaction between neuropeptides and the innate immune microenvironment establishes a dynamic regulatory network that amplifies type 2 inflammation and itching through local neuroimmune signaling centers in the skin57 (Figure 2). Specifically, the neuroregulatory protein U (NMU) released by sensory neurons binds to the NMUR1 receptor on the second group of innate lymphocytes (ILC2s), forming a specialized “neuro ILC2 unit” that rapidly promotes ILC2 proliferation and secretion of cytokines such as IL-5 and IL-13, thereby driving eosinophil recruitment and Th2 polarization in inflamed skin tissue.58 In addition, the NMU-NMUR1 axis can enhance the activity of ILC2 effectors in response to epithelial stress signals such as IL-33 and TSLP. For instance, a mouse AD model suggests that NMU deficiency reduces the accumulation of ILC2 and alleviates skin inflammation, suggesting that targeting NMUR1 may be a promising strategy for disrupting the early neuroimmune inflammatory cascade.59 In addition, CGRP, Vasoactive intestinal peptide (VIP) and SP exert strong immunomodulatory effects on dendritic cells (DCs), mast cells, and vascular endothelial cells by binding to their respective receptors - CGRP’s RAMP1 and SP’s NK1R. This activation promotes degranulation of mast cells, histamine release, and endothelial vasodilation, promoting immune cell infiltration and neurogenic edema in AD lesions.60 For example, in human AD skin biopsy, elevated SP levels are associated with increased mast cell activation and DC maturation, leading to enhanced antigen presentation and persistent allergic inflammation. In clinical practice, SP antagonists have shown efficacy in reducing itching and erythema in Phase II trials of pruritus, emphasizing their therapeutic potential in AD.61 In addition, CGRP affects endothelial function by inducing nitric oxide dependent permeability, synergistically inhibits regulatory T cell (Treg) activity with VIP, and enhances ILC2 and basophil responses. Studies using organoid models derived from AD patients have shown that CGRP blockade can reduce excessive cytokine production, restore barrier integrity, and further highlight the pathogenic role of neuropeptide immune crosstalk in AD.62 Therefore, these neuropeptides form a complex microcircuit in which neuronal signals fine tune innate immune reactivity, exacerbating barrier dysfunction, maintaining chronic inflammation, and promoting the persistence of diseases in genetically or immunologically susceptible individuals.63
In addition, tissue-specific programming of the group 2 innate lymphoid cells (ILC2s) endows the skin with unique activation features of resident ILC2s, characterized by increased expression of dual regulatory proteins and IL-13 in response to skin alarm gradients. This distinguishes them from lung or intestinal ILC2s, which primarily produce IL-5 under similar stimuli.64 For example, in AD, skin ILC2s exhibit epigenetic modifications driven by mediators derived from keratinocytes, leading to preferential amplification during disease onset. A longitudinal clinical study showed that biologics targeting ILC2, such as anti-IL-13 antibodies, can significantly reduce the severity score and itching intensity of eczema in patients with refractory diseases.65 In addition, this organ specific activation bias is combined with neuropeptide signaling, where neuroregulatory protein U (NMU) preferentially amplifies the skin ILC2 response rather than other tissue subgroups, leading to local hyperresponsiveness. This provides a translational opportunity for precise treatment, such as local NMUR1 inhibitors, which can selectively alleviate skin inflammation without inducing systemic immune suppression.66 At the same time, other components of the neuroimmune microcircuit include neuronal derived acetylcholine, which affects the polarization of macrophages towards the M2 phenotype, thereby maintaining fibrosis remodeling in chronic AD. Recent proteomic analysis of diseased skin suggests that acetylcholine receptor antagonists can alleviate extracellular matrix deposition, highlighting a potential therapeutic pathway for reversing fibrosis related remodeling.67 Elucidating these immune neural mechanisms has accelerated the development of targeted biologics for AD therapy.34,68 For example, dupilumab - a monoclonal antibody that blocks IL-4R α - effectively inhibits the IL-4 and IL-13 signaling pathways, thereby suppressing Th2 driven inflammation.69 However, in addition to immunomodulatory therapies, the neuroimmune interface remains a core determinant of the pathogenesis of AD, such as the secretion of IL-31 by Th2 cells, which directly induces itching by activating receptors on sensory nerve fibers.70,71
The Role of Central Nervous System in the Pathological Mechanism of AD
So far, an increasing amount of new evidence emphasizes the complex regulatory role of the central nervous system (CNS) in the pathogenesis of AD, where neural circuits not only amplify peripheral inflammation but also integrate environmental and psychological stress factors to maintain chronic disease status.72 Specifically, the brain skin axis mediates stress-induced AD exacerbation by activating the hypothalamic pituitary adrenal (HPA) axis. Under psychological pressure, the hypothalamus releases corticotropin releasing hormone (CRH), which stimulates the pituitary gland to secrete adrenocorticotropic hormone (ACTH), leading to an increase in cortisol levels in the adrenal cortex. This cascade reaction disrupts the epidermal barrier function, promotes the dominance of Th2 cytokines, and thus links neuroendocrine stress response with immune dysfunction.73 For example, in AD patients who experience acute stress, increased HPA activity is associated with increased epidermal permeability and decreased expression of filaggrin, as demonstrated by longitudinal cohort studies where transient cortisol surges preceded inflammatory episodes. These changes can promote a pro-inflammatory environment, impair keratinocyte differentiation and barrier repair.74 In addition, the HPA axis functionally also intersects with sympathetic nervous system (SNS) activity, leading to catecholamine driven recruitment of immune cells and degranulation of mast cells in the dermis. For example, clinical evidence supports this association, as in randomized controlled trials, mindfulness based interventions that normalize HPA response reduced AD severity scores by 30%.75 Therefore, the treatment of HPA dysregulation has clinical potential, as glucocorticoid receptor modulators have been shown in preclinical models to alleviate stress-induced lesions, paving the way for a comprehensive approach that combines psychopharmacology and dermatology interventions to improve relief outcomes.76 In addition, neuroimaging studies on itch perception have revealed abnormal central nervous system activation patterns in AD, with functional magnetic resonance imaging (fMRI) showing overactivation of the anterior cingulate cortex (ACC) and insula regions during itch stimulation, reflecting significant changes in network involvement and exacerbating subjective perception of itch.77 For example, in AD patients, compared to healthy controls, histamine induced itching can cause significant bilateral activation of the thalamus and prefrontal cortex. Positron emission tomography (PET) studies have shown that reduced binding of μ-opioid receptors in these areas is associated with persistent chronic itching.78 Furthermore, these altered neural activation patterns reflect central sensitivity, as diffusion tensor imaging (DTI) reveals abnormalities in the white matter tracts within the corpus callosum, linking chronic itching to emotional disorders. These findings support neurofeedback training as a transformative approach to regulate maladaptive itch circuits and improve the quality of life for patients with refractory AD.79 In addition, the mechanism associated with placebo amplifies the itch perception of AD patients through the central expectation network, as fMRI shows enhanced activation of the amygdala and orbitofrontal cortex during placebo allergen exposure, mimicking true allergic reactions. This phenomenon suggests that cognitive-behavioral therapy (CBT) may help reconnect maladaptive neural pathways and reduce symptom burden.80 When it comes to the interaction between psychological nervous system and immune system, emotional stress exacerbates AD through bidirectional central nervous system immune communication, where anxiety induced changes in vagus nerve tone inhibit the anti-inflammatory cholinergic pathway, leading to increased release of IL-31 and TSLP derived from keratinocytes and intensified neuronal hypersensitivity reactions.81 For example, in a longitudinal AD cohort, it was shown that an increase in perceived stress score is associated with upregulation of NGF expression in the skin, which may be driven by signals mediated by the central nervous system that promote eosinophil infiltration and barrier damage. Proteomic analysis confirmed an increase in NGF before clinical onset,82 but there is currently not much research to prove a strong correlation between the two. In addition, serotonin dysregulation associated with depression in the central nervous system can worsen AD by impairing peripheral immune tolerance, and selective serotonin reuptake inhibitors (SSRIs) have shown adjuvant efficacy in reducing itch intensity and eczema area severity index (EASI) scores in comorbid patients in phase II clinical trials.83 At the same time, dysfunction of the autonomic nervous system (ANS), particularly insufficient parasympathetic activity, plays a supplementary role, impairs sweating function, and promotes microbial dysregulation in AD skin. Based on Sudoscan’s evaluation, the electrochemical skin conductance of AD patients is reduced, providing diagnostic insights and supporting biofeedback therapy aimed at restoring autonomic balance and enhancing disease resistance.84
Emerging research emphasizes the crucial role of the nervous system in driving the chronicity and recurrence of AD as well.16,71 Some viewpoints suggest that abnormal neuronal activation not only maintains dysfunction of the epidermal barrier, but also amplifies immune dysfunction, leading to permanent inflammation and itching.85 While persistent itching can cause central sensitization, lower the threshold for itch perception, and lead to excessive reactions from patients to other harmless stimuli.86 It is worth noting that Guseva et al’s study showed that brain-derived neurotrophic factor (BDNF) derived from eosinophils promotes the sprouting of sensory neurons, increases neuronal excitability, and thus enhances itch sensitivity.87 Repeated scratching can further damage the epidermal barrier, release additional inflammatory mediators, and strengthen the self-sustained neuroimmune circuits that drive the progression of chronic diseases.88 Therefore, these findings emphasize the multifaceted involvement of the nervous system in AD pathophysiology and establish it as a promising therapeutic target for disrupting the itch inflammation cycle and improving disease management.68
The Role of Neuroinflammation in AD
The rapid understanding of neuroinflammation as a dynamic interaction between neural and immune components is fundamentally reshaping our understanding of chronic skin diseases, particularly in AD, where it manifests as increased neuronal activation and cytokine mediated crosstalk, leading to permanent itching and barrier dysfunction.39 Neuroinflammation, broadly defined as an inflammatory response within nerve tissue, is characterized by the release of pro-inflammatory mediators, activation of peripheral glial like cells, and regulation of neuronal signaling. Its scope extends beyond the central nervous system to peripheral organs such as the skin, bridging local and systemic pathological processes26 (Figure 3). In AD, neuroinflammation leads to disease chronicity by affecting the activity of adaptive immune cells, particularly T and B lymphocytes.26,89 There is evidence to suggest that the nervous system regulates immune function through the release of neurotransmitters, as these signaling molecules bind to receptors on immune cells, regulating their transport, differentiation, and effector responses, thereby maintaining immune activation and promoting skin infiltration.89,90 The accumulation of activated immune cells further promotes the release of inflammatory cytokines and chemokines, amplifies local immune responses, and ultimately leads to the chronic and recurrent processes of AD.26,91 Moreover, neuroinflammation induced epidermal barrier damage exacerbates the persistence of AD, as neuropeptides such as CGRP and SP can disrupt the function of keratinocytes and damage the integrity of the barrier. The destruction of this barrier enhances the penetration of allergens and microbial products, thereby triggering sustained immune activation, establishing a self-reinforcing inflammatory cycle, and maintaining disease progression.26,92
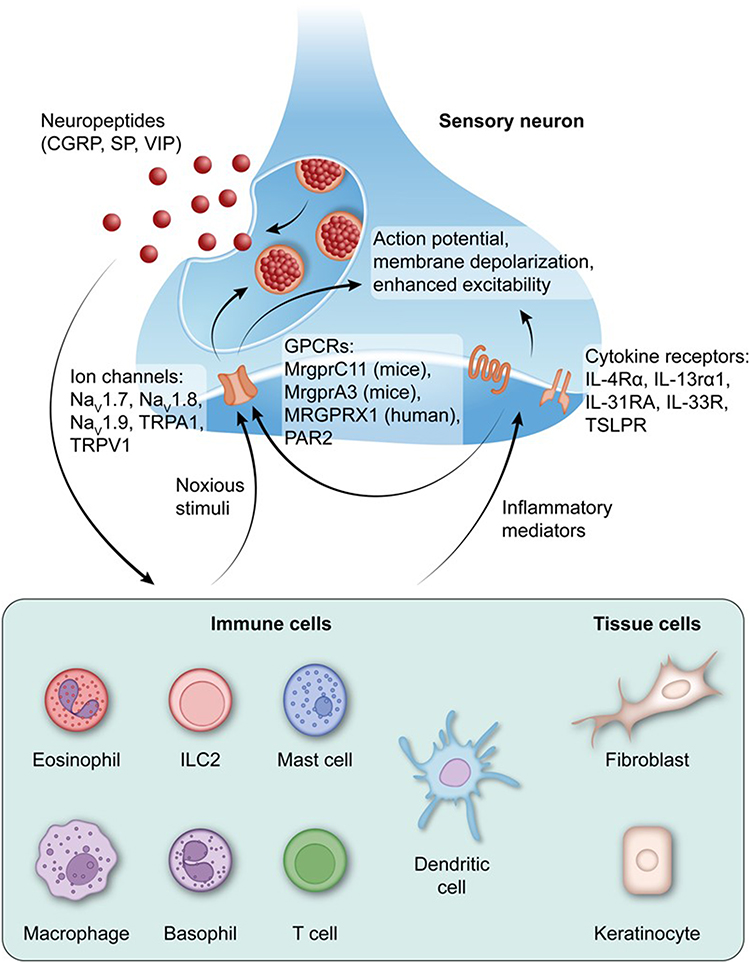 |
Figure 3 Neuroimmune interactions drive neuroinflammation in AD. The schematic diagram depicts the bidirectional crosstalk between sensory neurons, immune cells, and tissue cells that maintain neuroinflammation in AD. Harmful stimuli and inflammatory mediators activate neuronal ion channels and G protein coupled receptors (GPCRs; MrgprC11, MrgprA3, MRGPRX1, PAR2), leading to neuronal depolarization, enhanced excitability, and release of neuropeptides such as calcitonin gene-related peptide (CGRP), substance P (SP), and vasoactive enteropeptide (VIP). These neuropeptides regulate immune and tissue cell functions, induce the production of cytokines and chemokines, degranulate mast cells, and further recruit immune cells, including eosinophils, basophils, macrophages, dendritic cells, and T cells. Type 2 cytokines (IL-4, IL-13, IL-31, IL-33, TSLP) act on neuronal cytokine receptors (IL-4R α, IL-13R α 1, IL-31RA, IL-33R, TSLPR), enhancing neuronal excitability and neuropeptide release, forming a feedforward loop for neuroinflammation. This sustained neuronal immune epidermal axis promotes itching, epidermal barrier disruption, and chronic inflammation unique to AD. Copyright ©2024. Reproduced from Kim B, Rothenberg ME, Sun X et al. Neuroimmune interplay during type 2 inflammation: Symptoms, mechanisms, and therapeutic targets in atopic diseases. J Allergy Clin Immunol. 2024 Apr;153(4):879–893. doi: 10.1016/j.jaci.2023.08.017.39 |
Interestingly, recent studies seem to reshape the research landscape, indicating that neuroinflammation in AD is not only a secondary phenomenon, but also a key pathogenic driving factor. Advanced mouse models and human biopsy analyses have shown that neuroinflammatory activity intensifies during disease onset and is closely related to the severity of symptoms.93 For example, in AD, type 2 cytokines such as IL-4, IL-13, and IL-31 coordinate neuroinflammatory signaling by binding to receptors on sensory neurons, thereby enhancing neuronal excitability through the JAK/STAT pathway and promoting the release of neuropeptides such as SP. These neuropeptides activate mast cells through MRGPRX2/MgprB2 receptors, triggering degranulation of mast cells and amplification of local inflammatory cascades.39 In addition, this bidirectional skin nerve immune axis is driven by nociceptors, where scratching stimulates sensory neurons expressing Mrgpra3 to release SP, which in turn induces the production of mast cell-derived tumor necrosis factor - α (TNF - α) and neutrophil infiltration, thereby exacerbating vascular permeability and epidermal thickening. This effect has been confirmed in Fc ε RI dependent AD mouse models.93 In addition, epithelial derived scaremongs such as thymic stromal lymphopoietin (TSLP) and interleukin-33 (IL-33) exacerbate neuroinflammation through transient receptor potential (TRP) channels on sensitized sensory neurons, leading to chronic itch sensitization and establishing a feedforward loop to maintain barrier dysfunction by downregulating filaggrin expression and increasing keratinocyte protease activity.26 Therefore, we found that clinical analysis of AD patients showed elevated concentrations of neurotransmitters (including SP and brain natriuretic peptide (BNP)) in the affected skin, which were associated with increased severity of itching and sleep disorders, as demonstrated by proteomic studies linking these mediators to disease activity scores.94 Meanwhile, psychological stress exacerbates this neuroinflammatory cascade by activating the hypothalamic pituitary adrenal (HPA) axis, leading to the release of glucocorticoids and indirectly regulating neuronal hypersensitivity and immune cell transport. Queue studies have shown that compared to the control group, the levels of neuroinflammatory biomarkers in patients with stress-induced AD are significantly elevated.95 Therefore, these mechanisms highlight key translation opportunities. Biological products, such as dupilumab, targeting IL-4R alpha to block IL-4/IL-13 signaling, have been shown in phase III trials to reduce itch severity by over 50%, improve quality of life, and disrupt cytokine neuron crosstalk, thereby reducing neuroimmune activation.39 At the same time, newly emerging JAK inhibitors, such as abrocitinib, have broad inhibitory effects on neuroinflammatory signals, providing rapid symptom relief in moderate to severe AD and paving the way for precise treatment of the interwoven skin nerve immune axis.94 In the future, therapeutic regulation of neuroinflammation through the skin nerve immune interface may be a promising strategy that can affect the chronicity and recurrence of diseases. There is enormous therapeutic potential for this neuroimmune relationship, and we hope that future research may develop more effective and personalized treatment methods, especially for patients with refractory or recurrent chronic inflammatory skin diseases (such as AD).34,96,97
Current Research Frontiers and Hot Topics on Neuroimmune Axis in AD
Multi Omics and Systems Biology Methods
Single Cell Sequencing and Systems Biology Methods
As for the rapidly developing field of AD research, studying the neuroimmune axis has become a central pathway for exploring the intersection of neuronal sensitization and immune dysregulation to maintain chronic itching and inflammation. This process is increasingly being elucidated through advanced multi omics and systems biology methods, which can provide high-resolution analysis of molecular interactions.98 At present, these innovative methods, including scRNA seq, proteomics, and epigenetic analysis, are widely used in the study of the pathogenesis of AD and neuroimmune regulation, helping to identify cellular heterogeneity, mediator networks, and regulatory modifications that are the basis of sensory hypersensitivity and barrier dysfunction.57,99 It is worth noting that scRNA seq completely changed our understanding of the neuroimmune interface by mapping the transcriptional status of individual cells in AD lesion skin, revealing different clusters of itch sensing neurons and Th2 skewed immune cells that collectively express itch inducing cytokines such as IL-13 and IL-31. This interaction initiates a cascade reaction, in which IL-31 binds to neuronal OSMR/IL-31RA receptors, enhancing TRPV1 mediated calcium signaling, thereby amplifying neurogenic inflammation and promoting epidermal proliferation, which is associated with elevated SCORAD scores in clinical cohorts.100 For example, the recent scRNA seq study of peripheral blood mononuclear cells in patients with severe AD by Seon Pil Jin et al found that dysregulated congenital lymphocytes (ILCs) showed lineage infidelity, ILC2s overexpressed IL-13, and activated neurons through osteocutaneous protein dependent pathways. This mechanism promotes sustained itching and barrier disruption, providing a promising conversion pathway for IL-13-targeted biologics such as trastuzumab, which have been shown to effectively alleviate the severity of itching in refractory adult patients.101 As a supplement, Zhou and his colleagues combined scRNA seq with spatial transcriptome analysis of AD skin, revealing that abnormal differentiation of keratinocytes is associated with increased inflammatory innate lymphocytes (ILCs) and neuronal proximity. In this case, IL-22 upregulated by ILC3s inhibits the expression of filaggrin through STAT3 phosphorylation, thereby exacerbating transepidermal dehydration and enhancing allergen penetration. These findings suggest that JAK inhibitors have therapeutic potential in restoring barrier integrity and reducing seizure frequency, particularly in pediatric AD.102 Proteomics, as an effective biological detection method, can identify new mediators of the neuroimmune cascade by quantifying protein abundance in AD skin and serum samples. Analysis revealed an increase in SP and CGRP levels, which activate mast cells through NK1R signaling, leading to the release of histamine and IL-4. This process sensitizes the peripheral nerves and maintains a neurogenic feedback loop, which is associated with increased itching intensity and persistent chronic disease.103 In addition, Yu Ri Woo et al proteomic study of systemic biomarkers of AD revealed upregulation of CCL17 and IL-33, which serve as molecular bridges between immune activation and neuronal overexcitation. IL-33 induces TRPA1 mediated itching through degranulation of mast cells, which has clinical applicability as an indicator of disease severity and potential therapeutic targets. These findings highlight the prospects of biologics such as dupilumab, which can alleviate inflammation and sensory hypersensitivity reactions, thereby improving the quality of life of long-term or refractory AD patients (Figure 4).104
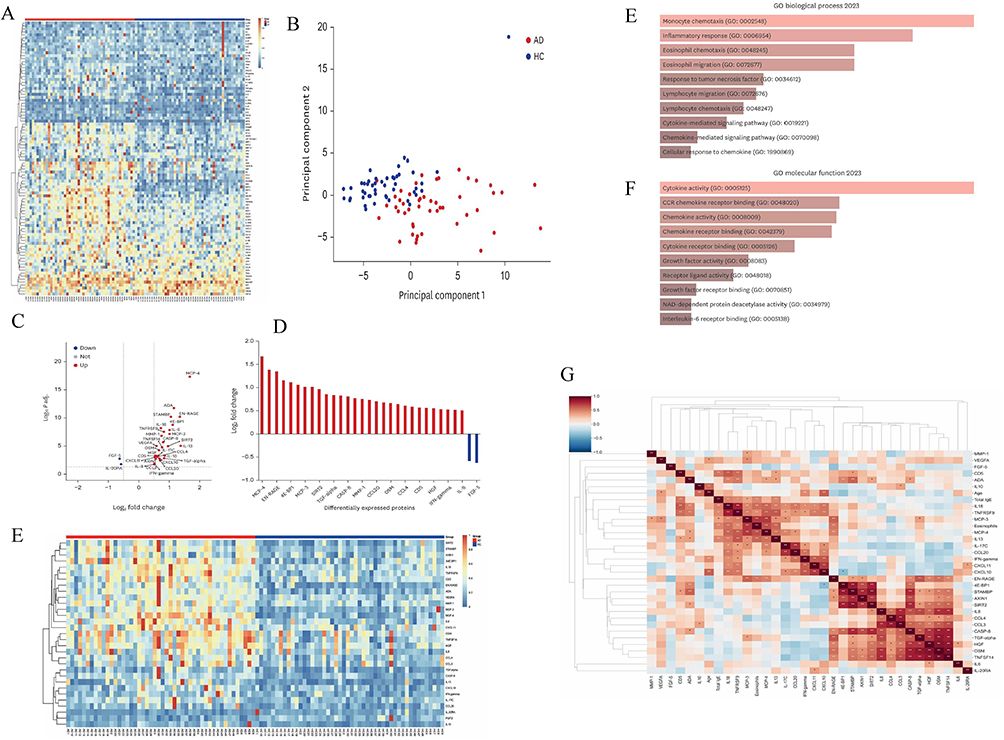 |
Figure 4 Serum inflammation proteome distinguishes AD from healthy controls. (A and B) Global serum differences. (A) Heatmap of 92 inflammation-panel proteins in AD and HCs. (B) Principal component analysis (PCA) showing separation between AD and HCs. (C–E) Differential expression. (C) Volcano plot of the 92 proteins highlighting those significantly altered in AD vs HC. (D) Log2_2 fold changes for DEPs meeting ||Log2_2FC|| ≥ 0.5 and P < 0.05. (E) Heatmap of DEPs across samples. (F and G) GO enrichment of DEPs. (F) Top enriched Biological Process terms. (G) Top enriched Molecular Function terms. (H) Clinical correlations. Correlation matrix of 31 DEPs with age, eosinophil counts, and total IgE in AD. Copyright ©2024. Reproduced from Woo YR, Moon JH, Shin HY et al. Systemic Inflammatory Proteomic Biomarkers in Atopic Dermatitis: Exploring Potential Indicators for Disease Severity. J Korean Med Sci. 2024 Aug 12;39(31): e223. doi: 10.3346/jkms.2024.39.e223.104 Abbreviations: AD, atopic dermatitis; HC(s), healthy control(s); DEP, differentially expressed protein; GO, Gene Ontology; IgE, immunoglobulin E. |
In addition, Michael Koch et al used quantitative proteomics to reveal a decrease in NRF2 activity in the epidermis of AD patients, linking mitochondrial dysfunction to oxidative stress, which disrupts neuronal homeostasis and amplifies Th2 response through IL-4/IL-13 signaling. These findings suggest that antioxidant interventions have therapeutic potential, aimed at enhancing mitochondrial elasticity and preventing the deterioration of neuroimmune drive, especially in elderly AD populations, which may provide new ideas for the treatment of AD patients.105 In addition, epigenetic modifications coordinate the regulation of neuroimmune genes in AD, and abnormal DNA methylation at the NGF and IL4RA promoters was observed in diseased keratinocytes, leading to increased expression, promoting neuronal growth and Th2 polarization, thereby exacerbating the itch scratch cycle and worsening barrier dysfunction.106 Specifically, in a study by Alberto Jos é da Silva Duarte et al, they investigated epigenetic control in the inflammatory pathway of AD. Histone deacetylation at the IL17A site was associated with sustained Th17 activation, promoting cytokine crosstalk, and upregulation of TRPV4 induced by IL-17 made nociceptors sensitive and associated with greater disease persistence. These findings support the potential of HDAC inhibitors as adjunctive therapies to regulate epigenetic markers and promote remission of refractory AD.107 In addition, microRNA analysis revealed overexpression of miR-155 in AD immune cells, which inhibits SOCS1 to enhance JAK-STAT signaling and IL-31 production, thereby linking epigenetic dysregulation with neuroimmune amplification, and indicating that miRNA antagonists have therapeutic potential in relieving itching and restoring skin homeostasis in different AD populations.108
Neuroimmune Axis From a Multi Omics Perspective
In the continuous development of AD neuroimmune research, integrating spatial and cellular lineage perspectives through multi omics methods has many unique advantages, which changes our understanding of dynamic cellular ecosystems and revealing how spatial proximity and lineage specific gene expression coordinate inflammatory cascades and itch responses.109 Specifically, we have carefully examined the single-cell profile of AD lesion skin, which depicts the neuroimmune “neighborhood” where sensory neurons expressing receptors such as MRGPRX2 cluster near type 2 innate lymphocytes (ILC2s) and mast cells. This organization promotes ligand receptor networks, such as the SP-NK1R axis, inducing calcium dependent degranulation of mast cells and subsequent IL-13 release, thereby amplifying epidermal hyperproliferation and barrier dysfunction in chronic lesions.101 For example, He et al used scRNA seq in AD biopsy and identified enriched ligand receptor pairs, including IL-31 from keratinocytes binding to OSMR on neurons, which enhanced neuronal excitability and continued the feedforward itch scratch cycle. At the same time, clinical observations further indicate that elevated levels of IL-31 in severe childhood AD are partially associated with the reactivity of anti-IL-31 biologics such as nemolizumab, highlighting the transformative potential of disrupting this neuroimmune network to reduce disease onset.110 In addition, this spatial framework emphasizes the importance of cell lineages in maintaining chronic inflammation. Th2 polarized CD4 ⁺ originates from immature precursor T cells infiltrating these neuroimmune regions and interacting with fibroblasts through CXCL12-CR4 signaling, thereby promoting a pro fibrotic microenvironment associated with increased skin thickness in refractory AD. These insights provide therapeutic opportunities for lineage targeting strategies such as CCR4 antagonists to regulate T cell migration and improve long-term response outcomes.99 For the application of spatial transcriptomics in neural immune epithelial units, these techniques map cell neighborhoods while preserving tissue structure, enabling the identification of pathway hotspots, such as upregulated JAK-STAT signaling in the perivascular region at the junction of neurons with epithelial cells and macrophages. This interaction inhibits the expression of filaggrin through IL-4/IL-13 signaling, leading to increased transepidermal dehydration, increased allergen penetration, and exacerbation of dryness and inflammation in AD.111 In a noteworthy study, spatial transcriptomics of AD lesions revealed concentrated hotspots of TNF - α and IFN - γ activity within neuroimmune epithelial clusters, with epithelial derived chemokines such as CCL20 recruiting CCR6 ⁺ neurons and T cells. This triggers a cascade reaction of keratinocyte apoptosis and immune amplification, consistent with histopathological observations of sponge like lesions during acute exacerbations, and provides clinical insights for spatially informed interventions, such as local JAK inhibitors, which can target immune suppression while minimizing systemic side effects.112 In addition, these analyses elucidate how cellular neighborhoods promote bidirectional signaling, such as neuropeptide Y released by neurons stimulating the production of IL-6 in adjacent epithelial units. Conversely, this in turn recruits eosinophils through IL-5, thereby perpetuating the observed eosinophilic sponge like changes in biopsy confirmed AD and emphasizing the therapeutic value of IL-5 inhibitors such as pembrolizumab for AD subtypes dominated by eosinophils, which can alleviate itching and restore barrier integrity.17 Advancing to the multimodal integration of scRNA seq, spatial genomics, and proteomics, these methods generate comprehensive molecular profiles by overlaying transcriptome profiles with spatial distribution and protein abundance, prioritizing potential drug targets. For example, elevated periostitis proteins found in the neuroimmune center have been shown to link neuronal activation with extracellular matrix remodeling and Th2 polarization, highlighting their critical role in AD progression.113 A recent multi omics study conducted by Chen et al integrated these patterns and identified RRM2 as a central protein within ILC2 and neuronal spatial clusters. Overexpression of RRM2 drives proliferation response by enhancing nucleotide synthesis, which is associated with increased epidermal renewal in diseased skin. RRM2 inhibitors have been proposed as potential combination partners with dupilumab to improve therapeutic efficacy. This is supported by preclinical data indicating a decrease in SCORAD score in treatment resistant adult AD models.114 In addition, this comprehensive framework promotes target prioritization through network-based analysis, revealing the rich CXCL10-CXCR3 interactions at the epithelial neural immune interface. Proteomic validation confirmed their role in recruiting sensitized T cells, which exacerbate neurogenic inflammation. This suggests that CXCR3 antagonists may prevent itch sensitization and promote skin homeostasis, as demonstrated by longitudinal cohort studies of AD patients.115
As we enter the next paradigm of multi omics in the neuroimmune field of AD, the fusion of spatial transcriptomics and single-cell lineage tracing is expected to depict individualized molecular features, thereby fundamentally changing the classification of subtypes and promoting tailored immune regulatory interventions.116 In addition, combining proteomics with spatial frameworks is expected to reveal dynamic ligand receptor interactions within the neuroimmune center, which may accelerate the identification of new biomarkers and predict the therapeutic response of refractory itch patients to biologics such as dupilumab.117 Meanwhile, future multi omics studies combining real-time epigenetic analysis can elucidate temporal changes in pathway hotspots, providing information for adaptive clinical trial design to optimize JAK inhibitor regimens, restore epidermal barrier integrity, and alleviate atopic progression in pediatric cohorts.118 In addition, in the future, utilizing this holistic and comprehensive paradigm may become a bridge between basic experiments and clinical applications, while machine learning assisted omics data synthesis can guide precision medicine strategies. These methods may ultimately reduce the comorbidity burden of genetically susceptible AD patients, including asthma, through early neuroimmune targeted prevention.119
Microbial Nervous Immune Axis
Nowadays, emerging evidence underscores the microbial-neuroimmune axis as a pivotal modulator in AD, highlighting how disruptions in both the cutaneous and enteric microbiota contribute to neural sensitization and inflammatory cascades. This growing research provides new avenues for exploring the gut skin brain interactions behind the persistence of diseases120 (Figure 5). The dysbiosis of the skin microbiome is characterized by the dominance of Staphylococcus aureus and the loss of symbiotic diversity, especially Staphylococcus epidermidis, which initiates axis activation by triggering toll like receptor 2 (TLR2) - mediated release of thymic stromal lymphopoietin (TSLP) from keratinocytes. This signal cascade subsequently sensitizes nociceptive C fibers through TSLP receptors on sensory neurons, thereby amplifying itching and neurogenic inflammation through SP induced degranulation of mast cells.121 At the same time, the gut microbiota of AD patients is imbalanced, characterized by a decrease in the ratio of Firmicutes to Bacteroidetes, which spreads systemic effects through short chain fatty acid (SCFA) deficiency (especially butyrate deficiency), impairing the differentiation of regulatory T cells (Tregs). This imbalance increases the levels of IL-4 and IL-13 in the circulation, promotes a Th2 dominated immune environment, enhances vagus nerve signaling, exacerbates skin nerve hypersensitivity and barrier dysfunction.122 Longitudinal cohort studies further support the regulation of cutaneous neuroinflammation by the gut, indicating that early exposure to antibiotics depletes the species of bifidobacteria, increases sympathetic outflow, and leads to increased expression of IL-31 neurons in pediatric AD. These processes maintain the feedforward loop of itch scratch behavior and microbial translocation.123 In addition to microbial factors, environmental confounding factors such as pollution, especially fine particulate matter (PM2.5), further exacerbate this axis by promoting the transformation of skin microbiota into pro-inflammatory Proteobacteria. Activation of TRPV1 channels on afferent nerves mediated by lipopolysaccharide (LPS) induces the release of calcitonin gene-related peptide (CGRP), thereby enhancing eosinophil recruitment and increasing epidermal nerve fiber density. Epidemiological studies have confirmed these findings by linking poorer air quality indices with higher SCORAD scores in urban AD cohorts.124 Similarly, dietary triggers such as high glycemic index foods can impair the integrity of the intestinal barrier, allowing microbial metabolites and histamine precursors to enter the circulation and activate the histamine itch pathway in the dorsal root ganglia. This mechanism links Western dietary patterns with accelerated onset of AD through the neural immune activation of genetically susceptible individuals.125 In this case, dysbiosis of the skin microbiome directly promotes neural sensitization, as superantigens from Staphylococcus aureus crosslink IgE on Langerhans cells, thereby initiating neuropeptide Y-mediated sympathetic nervous activation. This cascade reaction leads to upregulation of TRPA1 expression in sensory terminals, resulting in chronic hypersensitivity reactions in the affected skin. These findings were confirmed by scRNA seq analysis of human AD biopsies, which revealed rich microbial neuronal gene features.126 To counteract these interferences, probiotic interventions targeting neuroimmune balance have shown promising translational potential. For example, oral administration of Lactobacillus rhamnosus GG can restore SCFA production and weaken vagus nerve mediated release of IL-33 from intestinal glial cells, thereby reducing excessive excitability of peripheral nerves. In a randomized controlled trial involving adult AD patients, this method significantly reduced the normalization of itch visual analog score and epidermal filaggrin expression.127 In addition, local application of the symbiotic probiotic Rosa rugosa mucosa can repair the fungal community in the skin, inhibit the quorum sensing signals of pathogenic fungi, otherwise these signals would amplify the neurogenic vasodilation mediated by protease activated receptor 2 (PAR2). Evidence from Phase II clinical studies suggests that reduced transepidermal dehydration and decreased neurofibrillary light chain markers of axonal stress enhance the therapeutic potential of microbiome regulation in AD.128 On the basis of these findings, it has been demonstrated that synbiotic preparations containing phytic acid rich diets can enhance the biosynthesis of indole-3-propionic acid in skin associated clostridia, thereby regulating the signaling of aryl hydrocarbon receptors in neurons. This pathway inhibits Th17 driven inflammation, promotes sensory axonal pruning, and facilitates itch resolution. The preclinical model has translated these results into an improvement in EASI scores in subsequent dietary intervention trials, emphasizing the systemic potential of gut skin metabolic crosstalk.129
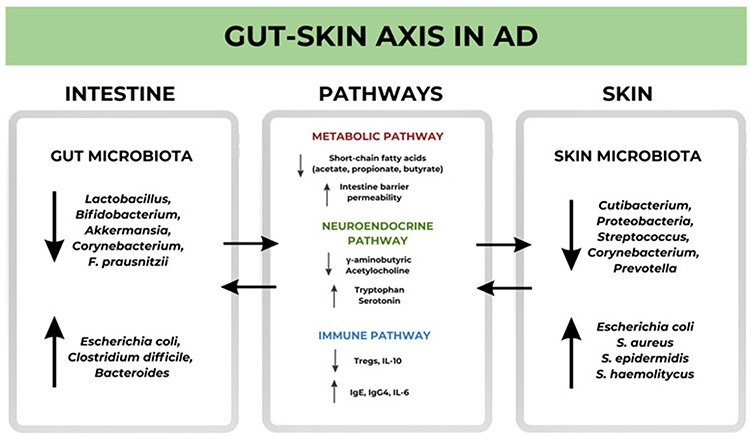 |
Figure 5 Microbial–neuroimmune gut–skin axis in AD. Schematic diagram showing bidirectional communication between gut and skin microbiota, providing fuel for neuroinflammation in AD. Gut dysbiosis—characterized by depletion of beneficial commensals (eg, Lactobacillus, Bifidobacterium, Akkermansia, Faecalibacterium prausnitzii) and enrichment of pathobionts (eg, Escherichia coli, Clostridium difficile, Bacteroides)—reduces short-chain fatty acids (acetate, propionate, butyrate), increases intestinal permeability, alters neurotransmitter/tryptophan metabolism, and skews immunity (↓Tregs/IL-10; ↑IgE/IgG4/IL-6). Skin dysbiosis with overgrowth of Staphylococcus aureus and shifts in resident taxa (eg, Cutibacterium, Proteobacteria, Streptococcus, Corynebacterium, Prevotella) triggers TLR2-driven keratinocyte TSLP, sensitizes sensory neurons/TRP channels, and promotes neuropeptide (SP/CGRP) release and mast-cell activation, amplifying itch and inflammation. Arrows represent gut skin signals that are transmitted through metabolic, neuroendocrine, and immune pathways, leading to barrier dysfunction and permanent chronic diseases. Copyright ©2024. Reproduced from Wrześniewska M, Wołoszczak J, Świrkosz G, Szyller H, Gomułka K. The Role of the Microbiota in the Pathogenesis and Treatment of Atopic Dermatitis-A Literature Review. Int J Mol Sci. 2024 Jun 13;25(12):6539. doi: 10.3390/ijms25126539.123 |
In addition, studies have found that extracellular vesicles from Lactobacillus fermentum regulate the serotonin pathway through the reconfiguration of the gut brain axis, leading to a decrease in 5-HT3 receptor density on sensory fibers and alleviation of AD symptoms in mouse models. This strategy demonstrates synergistic potential with anti-IL-4 biologics in restoring microbial neural homeostasis and provides promising directions for clinical progress.130 Overall, these comprehensive interventions highlight the therapeutic extensibility of the microbial neuroimmune axis, paving the way for personalized microbiome engineering strategies to disrupt neuroinflammatory circuits driven by ecological dysbiosis and promote persistent disease control.131
Looking ahead, the microbial neuroimmune axis in AD marks a paradigm shift towards comprehensive, multi omics research that elucidates the dynamic interactions between skin dysbiosis, gut metabolites, and neural sensitization. This approach is expected to establish a resilient, patient-centered therapeutic ecosystem that goes beyond traditional symptom management and targets potential neuroimmune networks for disease persistence.17,132,133 In this constantly evolving framework, emerging strategies emphasize the importance of developing standardized, AI enhanced fecal microbiota transplantation (FMT) protocols tailored specifically for AD patients. In addition, through these innovations, we can envision a future of precise monitoring, where metabolomics driven dashboards continuously evaluate SCFA and neuropeptide flux to guide the optimization of adjuvant therapy. These advances collectively aim to promote lasting neuroimmune balance and comprehensive disease management throughout the entire lifecycle of patients.134,135
Neuroimmune Biomarkers in the Diagnosis and Treatment of AD
At present, the traditional diagnosis of AD still mainly depends on clinical manifestations and patient medical history, lacking objective and quantifiable biomarkers that can reflect potential neuroimmune disorders. However, based on current insights into new developments, research on neuroimmune biomarkers for AD is becoming increasingly widespread and mature, and the future of research increasingly relies on translating neuroimmune findings into measurable clinical indicators, thereby adapting current limited diagnostic approaches to precision medicine. Meanwhile, advances in multi omics technologies such as transcriptomics, proteomics, and immunohistochemistry have catalyzed the identification of novel biomarkers associated with the neuroimmune pathways of AD. These biomarkers not only deepen our understanding of disease etiology, but also provide promising avenues for improving diagnostic accuracy, treatment monitoring, and patient stratification in clinical practice.
Neuroimmune Biomarkers in AD
In the paradigm shift towards precision dermatology, AD is considered a multifactorial disease caused by the interaction of barrier dysfunction, immune dysregulation, and environmental triggers. Novel biomarkers have become indispensable tools for improving diagnostic accuracy and guiding treatment decisions. However, due to challenges in validation, patient differences, and technical standardization, their integration with routine clinical practice is still limited136,137 (Figure 6 and Table 3). For example, serum thymus and activated regulatory chemokines (TARC/CCL17), a Th2 related cytokine, have been identified as reliable indicators of AD disease severity and treatment responsiveness. Mechanistic studies have shown that elevated levels of TARC contribute to eosinophil recruitment and amplify type 2 inflammation through CCR4 receptor signaling on T cells, thereby perpetuating the itch scratch cycle. Meanwhile, in clinical practice, a phase III trial conducted in 2024 showed that baseline TARC concentrations exceeding 1000 pg/mL were associated with reduced itching in patients receiving dupilumab treatment, emphasizing its translational relevance among stratified responders. However, given its overlap with asthma phenotype, its limited specificity remains a key limiting factor.138 In addition, recent studies on ceramides as skin barrier biomarkers have further elucidated their pathogenic significance in AD. Changes in ceramide chain length and sphingolipid metabolism can impair the integrity of the epidermis, leading to increased transepidermal dehydration and enhanced allergen penetration. For example, Yong et al reported a pediatric AD cohort study based on lipidomics, which showed that a ceramide ratio (such as CER [NS]/CER [AS]) below 1.5 can predict early-onset disease and poor response to local corticosteroids, providing valuable prognostic insights for preventive interventions. However, the clinical application of ceramide analysis is hindered by the need for invasive biopsy and the lack of standardized methods for different ethnic populations.139 Furthermore, multiple omics methods have identified spatial transcriptomic features, such as upregulation of IL-31 and OX40 ligands in diseased skin, which lead to neuronal sensitization and chronic Th2 polarization through the JAK-STAT and NF - κ B pathways. A famous study combining spatial transcriptomics with scRNA seq was able to predict recurrence after JAK inhibitor treatment, indicating that patients with high IL-31 expression only achieved sustained remission in 60% of cases. These findings highlight the potential of such biomarkers in upgrading personalized therapy to biologics such as pembrolizumab, despite the current high cost and complex bioinformatics requirements hindering their widespread clinical application.140 Similarly, emerging immune biomarkers such as S100A8/A9 (calprotectin) have attracted attention for their role in reflecting innate immune activation in AD. S100A8/A9 heterodimer promotes excessive proliferation of keratinocytes and cytokine release through TLR4 binding, exacerbating barrier dysfunction. Real world data from European registry shows that elevated serum S100A8/A9 levels (>500 ng/mL) distinguish moderate to severe AD from mild AD with an accuracy rate of 85%, supporting its application in early diagnosis and treatment monitoring, particularly in tracking the efficacy of Atrotinib. However, its elevation in other inflammatory diseases can impair disease specificity, highlighting the necessity of combining biomarker panels to improve diagnostic accuracy.141 For the functional evaluation of biomarkers, neuroimaging derived biomarkers have become valuable tools for elucidating the central mechanism of AD itching so far. The changes in functional connectivity between the insula and somatosensory cortex have been shown to reflect neural processing abnormalities associated with chronic itching. fMRI studies have shown that the reduction in gray matter volume leads to persistent itching through maladaptive neural plasticity changes. A recent clinical trial further combined these imaging biomarkers with serum neurokinin B measurement results to predict treatment response to trastuzumab, revealing that recovery of functional connectivity after treatment is associated with improvement in EASI score. These findings advance the neuropsychiatric management of AD by connecting central and peripheral biomarkers, although implementation is still limited by high imaging costs, accessibility issues in resource limited environments, and inherent subjectivity in itch assessment.142 In addition, the latest research indicates that the emergence of microbiome related biomarkers highlights the role of skin dysbiosis in the pathogenesis of AD. For example, the decreased abundance of Staphylococcus aureus alpha toxin inhibitors in the skin metagenome has been identified as a key indicator of dysregulation driven inflammation, where toxin mediated pore formation in keratinocytes triggers the release of IL-1 β and biases adaptive immunity towards a pro-inflammatory phenotype. Recent studies on Asian AD populations have shown that low abundance of inhibitors indicates susceptibility to infection and poor responsiveness to moisturizer treatment, supporting probiotics targeting the microbiome as a potential adjuvant intervention. However, inter batch differences in sequencing results and limited longitudinal datasets continue to hinder their clinical translation.136
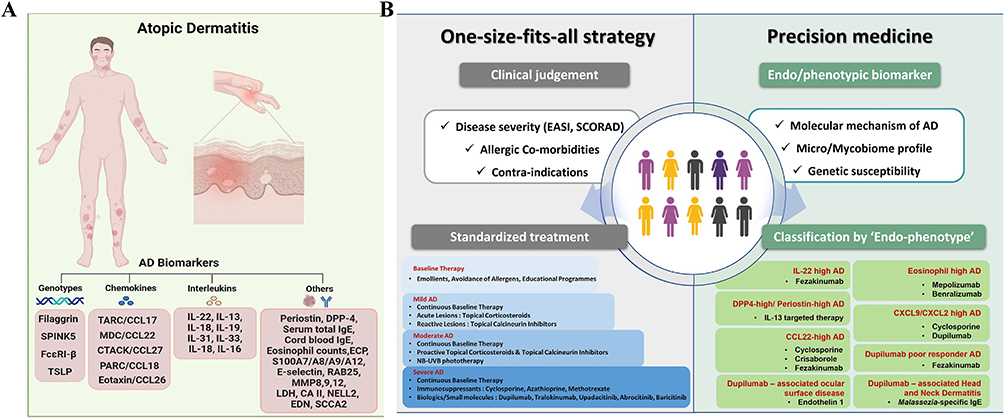 |
Figure 6 (A) Overview of Biomarkers in AD. This panel illustrates the typical distribution of skin lesions (red plaques) in AD patients and categorizes key biomarkers linked to AD pathogenesis. These include genotypes (eg, filaggrin, SPINK5, FcεRI-β, TSLP), chemokines (eg, TARC/CCL17, MDC/CCL22, CTACK/CCL27, PARC/CCL18, Eotaxin/CCL26), interleukins (eg, IL-22, IL-13, IL-31, IL-33, IL-16), and other markers (eg, periostin, DPP-4, serum IgE, eosinophil count, ECP, S100A7/A8/A9/A12, LDH, CA II, NELL2, EDN, SCCA2). These biomarkers demonstrate the complex neuroimmune interactions, inflammatory processes, and skin barrier damage underlying AD. (B) Comparison of AD treatment strategies: traditional medicine and precision medicine. The traditional “one size fits all” approach based on clinical judgment (left) and precision medicine stratified by intrinsic/phenotypic biomarkers (right) highlight personalized treatment for AD subtypes to address the complex heterogeneity of neuroimmunity. Copyright ©2024. Reproduced from Park CO, Kim SM, Lee KH, Bieber T. Biomarkers for phenotype-endotype relationship in atopic dermatitis: a critical review. EBioMedicine. 2024 May;103:105121. doi: 10.1016/j.ebiom.2024.105121.143 |
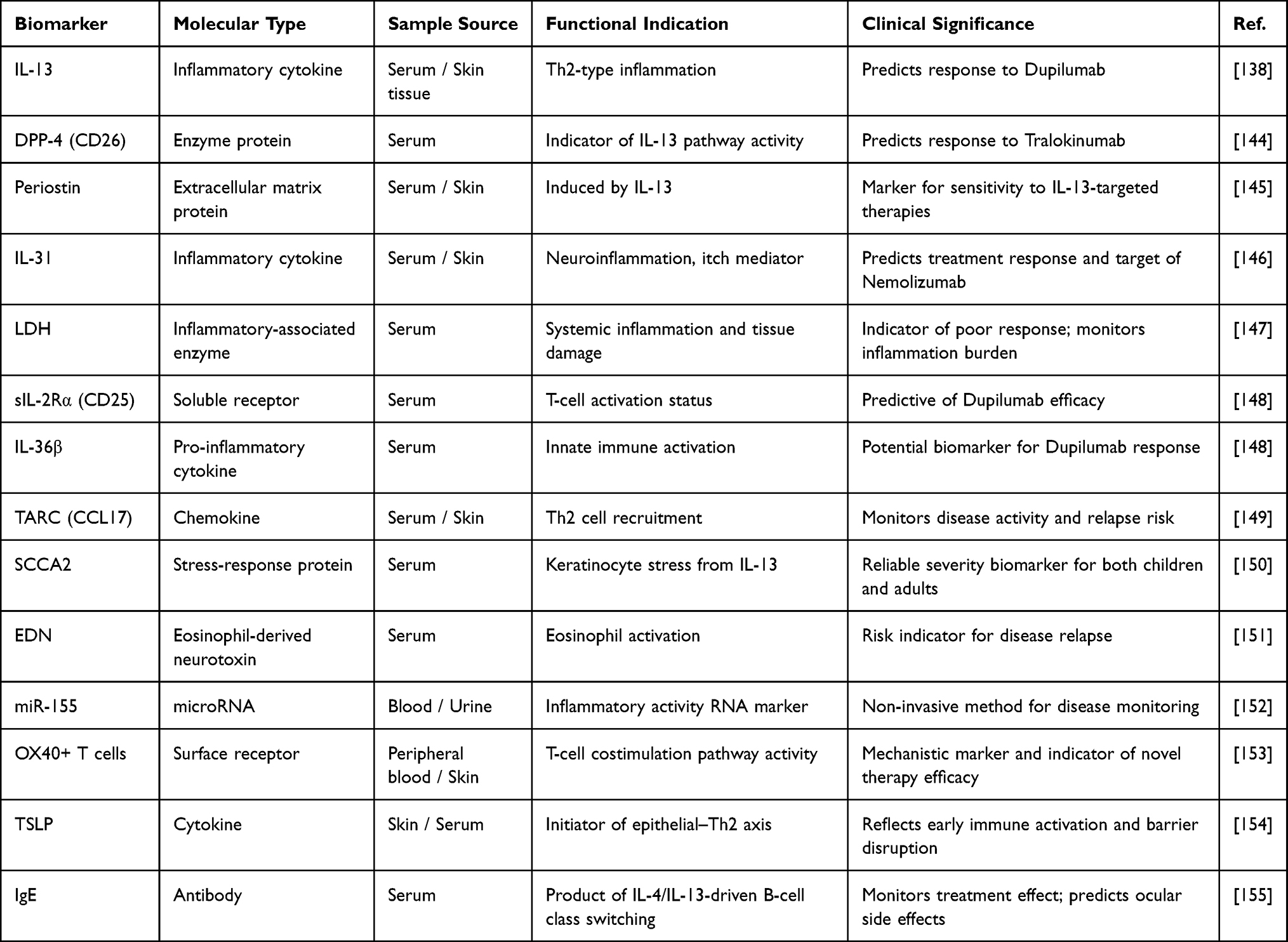 |
Table 3 Biomarkers in the Neuroimmune Mechanism of AD |
In addition, a recent review by Lyubchenko et al found a positive correlation between thymic stromal lymphopoietin (TSLP) levels, SCORAD score, and transepidermal dehydration (TEWL), highlighting its critical role in AD neuroimmune signaling.156 TSLP is an epithelial derived cytokine closely related to disease severity and barrier dysfunction, which promotes neuroimmune crosstalk by activating dendritic cells and mast cells. Similarly, S100 family proteins (S100A7, S100A8, and S100A9) are upregulated in acute and chronic AD lesions, interacting with IL-22 and IL-17-mediated inflammatory pathways, and may affect neuroimmune dynamics by regulating epidermal differentiation.34,157,158 A meta-analysis conducted in 2025 further confirmed that elevated levels of S100 protein are associated with the inflammatory burden and severity of itching in AD, emphasizing its potential diagnostic and prognostic utility.157 Based on these molecular insights, a recent bioinformatics study integrating machine learning methods identified candidate genes such as S100A9, SH3BGRL2, RAB27B, TMEM158, DAB2, FSTL1, CALD1, and XK, which may contribute to the neuroimmune mechanisms of AD.159
Stratified Medicine and Precision Medicine in AD Neuroimmune Biomarkers
In the dynamic field of precision dermatology, stratified medicine has become a key method for decoding disease molecules and cellular heterogeneity using biomarkers, as AD is characterized by complex interactions between immune dysregulation, epidermal barrier damage, and chronic itching. From this perspective, we can use biomarker guided stratification to align tailored treatment strategies with different disease subtypes, optimize clinical outcomes, and minimize adverse reactions136,160 (Figure 6). By combining high-resolution biomarker analysis with clinical phenotypes, precision medicine for AD not only improves diagnostic accuracy and prognostic capabilities, but also lays the foundation for a transformative, patient-centered treatment paradigm.138,161–163 Among the most notable neuroimmune biomarkers, serum NGF—a neurotrophin that supports sensory neuron survival and mediates neuronal hypersensitivity—is significantly elevated in AD patients compared with healthy controls. Mechanism studies have shown that NGF stimulates the sprouting of epidermal nerve fibers and induces keratinocytes to secrete pro-inflammatory cytokines such as IL-4 and IL-13, thereby amplifying itching and inflammation in a self-sustaining feedback loop.164 In addition, cytokines such as IL-31 play a central role in itch signaling by activating the IL-31 axis. IL-31 binds to heterodimeric receptors (IL-31RA/OSMR) on dorsal root ganglion neurons, triggering the JAK-STAT signaling cascade and inducing calcium influx and subsequent transmission of itching sensation. Clinical studies have shown that an increase in serum IL-31 concentration is closely related to the severity score of itching, especially in the pediatric AD cohort, highlighting its potential as a neuroimmune driven biomarker for itching.165 In addition, neuropeptides such as SP and CGRP, which released from high-density cutaneous nerve endings in AD affected skin, can further amplify the Th2 twisted immune response. They achieve this goal by promoting degranulation of mast cells and cytokine release, which enhances neurogenic inflammation and itching. Immunohistochemical analysis showed that the density of nerve endings in atopic lesions doubled, which is associated with chronic scratching behavior and increased disease activity.166 Multiple omics integrated studies further confirmed this hyperactivity and persistent inflammation, with spatial transcriptomics combined with scRNA seq revealing key ligand receptor interactions in the fibroblast leukocyte microenvironment, such as COL6A5-CCL19. These spatially resolved inflammatory niches drive sustained neuroimmune activation and serve as predictive molecular features for AD disease progression, linking basic biology with clinical translation.167 Advanced diagnostic methods, such as neuroimaging techniques, particularly fMRI, have revealed characteristic changes in brain connectivity in AD patients. In the itch stimulation task, the reduced activity in the caudate nucleus and posterior cingulate cortex indicates the presence of central sensitization mechanisms that lead to sleep disorders and cognitive dysfunction. A recent study further suggests that AD patients who respond to treatment exhibit normal neural activation patterns after treatment, emphasizing the potential of neuroimaging biomarkers in monitoring treatment efficacy.168 Similarly, resting state fMRI analysis showed that itching caused by eczema enhances the activation of the somatosensory cortex and insular cortex, providing functional evidence for neural plasticity remodeling. These findings are closely related to the intensity of itching reported by patients and can serve as objective endpoints for evaluating treatment interventions in clinical trials, thereby linking subjective symptoms with quantifiable neurophysiological data.169 Integrating these neurofunctional biomarkers into the precision medicine framework can improve treatment stratification and prediction accuracy. For example, JAK inhibitors such as upadatinib exhibit rapid anti itch effects by blocking IL-31 mediated JAK1 signaling, significantly reducing itching within a few weeks after initiation. Phase III clinical data indicate that baseline serum IL-31 levels below 50 pg/mL can predict better EASI-75 response at week 16, highlighting the translational value of biomarker based patient selection in optimizing treatment outcomes.170 On the contrary, IL-31 inhibitors such as nemolizumab maintain long-term control of itching by directly antagonizing IL-31RA receptors and blocking downstream neuronal excitation, thereby maintaining remission of moderate to severe AD. At the same time, real world evidence from the Japanese cohort further supports the use of multi omics analysis to assess skin IL-31 expression before treatment and reduce recurrence rates, strengthening the role of informed monitoring of biomarkers in personalized treatment.171
Secondly, OX40 targeted therapies, such as amlitelimab, have slower but more profound immunomodulatory effects. By disrupting the OX40-OX40L interaction on memory T cells, these therapies inhibit chronic Th2 polarization and reduce susceptibility to recurrence. Spatial transcriptome analysis has identified ligand receptor features that predict delayed but persistent clinical responses in patients with refractory AD, which can help design tailored treatment plans that balance efficacy and safety through biomarker guided precision care and minimize overtreatment.172 In addition, a study conducted by Ishiuji et al used fMRI to monitor cortical activity in AD patients during itch stimulation, revealing that central sensitization may be a potential diagnostic biomarker for the disease.173 These neurofunctional biomarkers provide valuable tools for enhancing disease diagnosis and patient stratification, bridging the gap between subjective symptom perception and objective neurophysiological evidence.
At the same time, non-invasive diagnostic techniques such as transepidermal dehydration (TEWL) and measurement of skin pH provide complementary insights into the integrity of the epidermal barrier and neuroimmune activation.137 A prospective study in 2023 further confirmed that the combination of familial susceptibility, elevated levels of type 2 cytokines, and abnormal lipid metabolism in infants significantly predicted the future development of AD (OR 54.0, 95% CI 9.2–317.5). Despite these promising findings, such predictive biomarkers have not yet been included in standard clinical workflows due to limited validation of early screening and practical challenges.174 Looking ahead, the integration of multiple omics platforms is expected to completely change the discovery of biomarkers and greatly improve the diagnostic accuracy of AD.137 With the accumulation of large-scale validation studies, these biomarkers are expected to transition from research frameworks to clinical implementation, thereby improving diagnostic accuracy, guiding personalized treatment, and enhancing overall patient outcomes.175 With the accumulation of large-scale validation studies, these biomarkers are expected to transition from research frameworks to clinical implementation, thereby improving diagnostic accuracy, guiding personalized treatment, and improving overall patient prognosis.
Therapeutic Strategies Targeting the Neuroimmune Axis in AD
Key Nodes and Potential Drug Targets in the Neuroimmune Circuit
In the intricate neuroimmune axis behind AD, bidirectional communication between sensory neurons and immune effectors maintains a self-reinforcing cycle of itching and inflammation. Therefore, treatment strategies targeting key regulatory nodes in this cycle can change disease management by selectively disrupting pathological crosstalk.17 The center of this paradigm is the so-called “itch axis”, controlled by IL-31 and its receptor IL-31R α. IL-31 is mainly secreted by Th2 cells and binds to IL-31R α/OSMR heterodimers on dorsal root ganglion neurons, thereby activating the JAK-STAT signaling cascade, inducing clinical manifestations of calcium influx, neuronal overexcitement, and refractory itching.176,177 Recent mechanistic studies further suggest that this axis exerts immunomodulatory effects by promoting neuronal release of CGRP, which in turn amplifies type 2 inflammation through mast cell activation. It is worth noting that a study by Marlys S. Fassett et al showed that IL-31 dependent neurogenic inflammation paradoxically suppressed the skin Th2 response in a mouse model, emphasizing the dual role of IL-31 signaling in itch transmission and immune homeostasis.178 From a clinical perspective, monoclonal antibodies such as nemolizumab that selectively target IL-31R α have achieved rapid and sustained itch relief in phase III clinical trials. The transformed evidence emphasizes the significant reduction in EASI scores and improvement in sleep quality in patients with moderate to severe AD, highlighting the potential of biomarker guided therapy to interrupt the chronic itch inflammation cycle and prevent disease recurrence.178 In addition to IL-31 signaling, JAK1 has become a key “common node” in the neuroimmune circuit. JAK1 mediates downstream responses to cytokines including IL-4, IL-13, and IL-31, driving epidermal proliferation and neuronal sensitization through STAT6 phosphorylation and itch inducing gene expression.179,180 Studies on JAK1 functional variants have shown that type 2 cytokine driven allergic inflammation is exacerbated in AD, and sensory neurons regulate lung immune homeostasis through JAK1 dependent pathways, indicating that atopic progression has broader systemic relevance.181 So, for AD patients, selective JAK1 inhibitors such as upadatinib show a rapid onset in relieving symptoms of neuroimmune drive. Evidence from the real world suggests that activation of the baseline JAK1 pathway can predict better itch relief and lasting relief, promote precise dosing strategies, minimize off target effects, and improve long-term treatment compliance.181
T cell reprogramming is a new technology for AD, and the OX40/OX40L signaling axis plays a key role in maintaining chronic Th2 polarization by mediating co stimulatory interactions on memory T cells, thereby driving sustained cytokine production and amplifying the inflammatory environment, enhancing neural sensitivity. The mechanism study conducted in 2024 elucidated that the involvement of OX40L-OX40 maintains the immune activation cycle of AD, promotes sustained T cell proliferation and effector function, thereby exacerbating the severity of the disease.182 It is worth noting that anti-OX40 biologics, such as Rocatinib monoclonal antibody and Amlitinib monoclonal antibody, have shown long-lasting efficacy in moderate to severe AD. A narrative review by Asaad Abdelhalim et al emphasized their potential to reduce recurrence rates through targeted pathway blockade, providing translational value for refractory AD cases resistant to traditional therapies and paving the way for biomarker guided combination regimens in personalized medicine.183 In addition, as a supplement to T cell guided methods, sensory nerve endings and mast cells represent key neuroimmune interfaces regulated by TRP channels (especially TRPV1 and TRPA1) and MRGPRX2 receptors. Activation of TRPV1 by endogenous ligands such as protons or heat triggers cation influx and neuronal depolarization, leading to SP release and subsequent degranulation of mast cells, thereby exacerbating cytokine driven inflammation. Jessica W. Hui Beckman’s review emphasizes the therapeutic potential of TRPV1 antagonists in alleviating temperature mediated itching and barrier dysfunction in AD, highlighting their potential utility in local formulations aimed at reducing environmental trigger sensitivity.184 Similarly, TRPA1 responds to oxidative stress and environmental stimuli, synergistically enhancing neuropeptide mediated inflammation in this cascade reaction. Hu et al demonstrated that mechanical scratching of the epidermis induces reactive oxygen species (ROS) activation of TRPA1 on mast cells, thereby exacerbating Th2 bias and inflammatory progression in AD models.185 At the same time, MRGPRX2 on mast cells acts as a non IgE receptor for itch, mediating the release of trypsin and activating PAR2 on sensory neurons, further amplifying the itch scratch cycle.186 Recent preclinical studies in 2024 have shown that MRGPRX2 antagonist GE1111 effectively improves DNFB induced AD in mouse models by inhibiting the production of inflammatory cytokines and restoring barrier integrity. This suggests that treating pruritus dominant AD phenotypes through feasible oral or local administration routes has therapeutic potential, aimed at improving patients’ quality of life.185 On the contrary, other neuroimmune pathways, such as the NK-1R axis (targeted by SP antagonists), have limited or inconsistent efficacy in AD. For example, in a retrospective analysis, Apriconazole showed moderate relief of refractory itching in patients with bullous epidermolysis and AD, while early clinical trials using Celostane failed to achieve the primary endpoint of itching relief, limiting its adjuvant role in selected subgroups of neurogenic itching rather than as a broad-spectrum treatment option.187 Another significant advancement is focused on the NGF signaling pathway.188 Livery’s research has shown that NGF enhances the excitability of sensory neurons through TrkA receptor activation, while stimulating keratinocytes to secrete IL-4 and IL-13, further prolonging Th2 inflammation.189 Based on these insights, a study by Nunomura et al identified periostitis proteins as a novel mediator of neuroimmune crosstalk. The periosteum secreted by keratinocytes activates ILC2 and sensory neurons, thereby exacerbating inflammation and itching through neuronal immune cell signaling.190
Although the complex pathways of the neuroimmune axis are crucial for advancing treatment strategies for AD - enabling the development of targeted interventions to break the vicious cycle of itching and inflammation at the molecular level - several key limitations continue to hinder their widespread clinical translation.17 These challenges include limited specificity of biomarkers such as IL-31 or JAK1 activity in inflammatory diseases, significant patient to patient variability in treatment response, and high costs of biologics such as nemolizumab and rocatinlimab, all of which limit accessibility and scalability.178,183 In addition, in resource constrained environments, the lack of standardized frameworks for integrating multiple omics datasets and the limited availability of advanced diagnostics such as functional MRI further hinder the practical implementation of precise dermatology.191,192 Looking ahead to the future, the integration of single-cell multi omics, machine learning driven biomarker panels, and innovative drug delivery technologies, such as TRP channel modulators encapsulated in nanoparticles and next-generation biologics with enhanced tissue specificity, brings great hope for overcoming these obstacles. Overall, these advances can design tailored and cost-effective treatment plans, achieve lasting relief for different types of AD, and significantly improve patients’ quality of life.
The Current Application and Clinical Strategies of Neuroimmune Regulation in AD
The treatment strategies targeting the neuroimmune axis are rapidly developing. For example, nemolizumab is an anti-IL-31 receptor monoclonal antibody that has shown significant efficacy in reducing itching in AD patients.193 Clinical trials have shown that pembrolizumab reduced itching by approximately 42.8% within 16 weeks, compared to 21.4% in the placebo group.193 In addition, JAK inhibitors such as baricitinib and upadatinib have been shown to alleviate inflammation and itching by inhibiting the JAK-STAT signaling pathway.194 TRPV1 channel antagonists, such as PAC-14028, have also shown promise in clinical trials, particularly in reducing itching.195 These emerging therapeutics offer new options for patients with moderate to severe AD.68 New therapeutic strategies based on neuroimmune mechanisms are rapidly developing. Dupilumab is an anti-IL-4R α monoclonal antibody that reduces inflammation and itching in AD by blocking IL-4 and IL-13 signaling pathways.196 Nimozumab targeting the IL-31 receptor specifically targets itching.68 Nemolizumab, which targets IL-31 receptors, specifically addresses pruritis.197 Currently, increasing evidence suggests that the clinical response rate of JAK inhibitors in patients with moderate to severe AD is 60% −80%.198,199 Neuropeptide receptor antagonists have also shown therapeutic potential. A study by Yosipovitch et al showed that NK1R antagonists, such as selenopitan, can alleviate chronic itching and improve quality of life.200 In addition, TRPV1 antagonists such as asivatrap have been shown to be effective in relieving itching in early clinical trials.201 These therapies have successfully translated neuroimmune research into clinical practice.
To date, multiple clinical trials have provided strong evidence for neuroimmune targeted therapy (Table 4).202,203 In the phase III trial of dupilumab, adult patients with moderate to severe AD showed significant improvements in eczema area and severity index (EASI) and itch visual analogue scale (VAS).202 The phase II trial of Nimozumab showed that it was superior to placebo in reducing itching, although the overall improvement in EASI score was not significant.203 Monoclonal antibodies targeting IL-13, including trastuzumab and lebrikizumab, have been approved by regulatory authorities in Europe and have been proven effective in improving AD symptoms.204–206 Vixarelimab (anti-OSMRβ) improved pruritus scores in clinical trials, though its impact on overall AD severity was limited.207 NK1 receptor antagonists (including selenopitan and traditional drugs) and TRPV1 antagonists (including PAC-14028) are still in clinical trials, and efficacy evaluation is ongoing.208–210 These findings highlight the therapeutic potential of neuroimmune targeted interventions in AD management, while emphasizing the necessity for further optimization to achieve comprehensive symptom control. Recent clinical trials have further confirmed the importance of neuroimmune mechanisms in the pathogenesis of AD. The dupilumab test (anti-IL-4R α monoclonal antibody) showed that over 80% of moderate to severe AD patients experienced a 50% reduction in itching within 16 weeks.202 The Phase III trial of nemolizumab showed a 60% reduction in itch score and a 40% reduction in lesion area within 12 weeks.211 The trial of JAK inhibitor baricitinib showed that 70% of patients in the 4mg dose group experienced relief of itching.212 In addition, a recent study using scRNA seq revealed heterogeneity in neuronal and immune cell populations within AD lesions, suggesting that variability in IL-31 signaling pathway activation may be a contributing factor to different treatment responses.213 These findings not only demonstrate the therapeutic potential of neuroimmune targeted interventions but also emphasize the importance of personalized medical approaches in AD management.
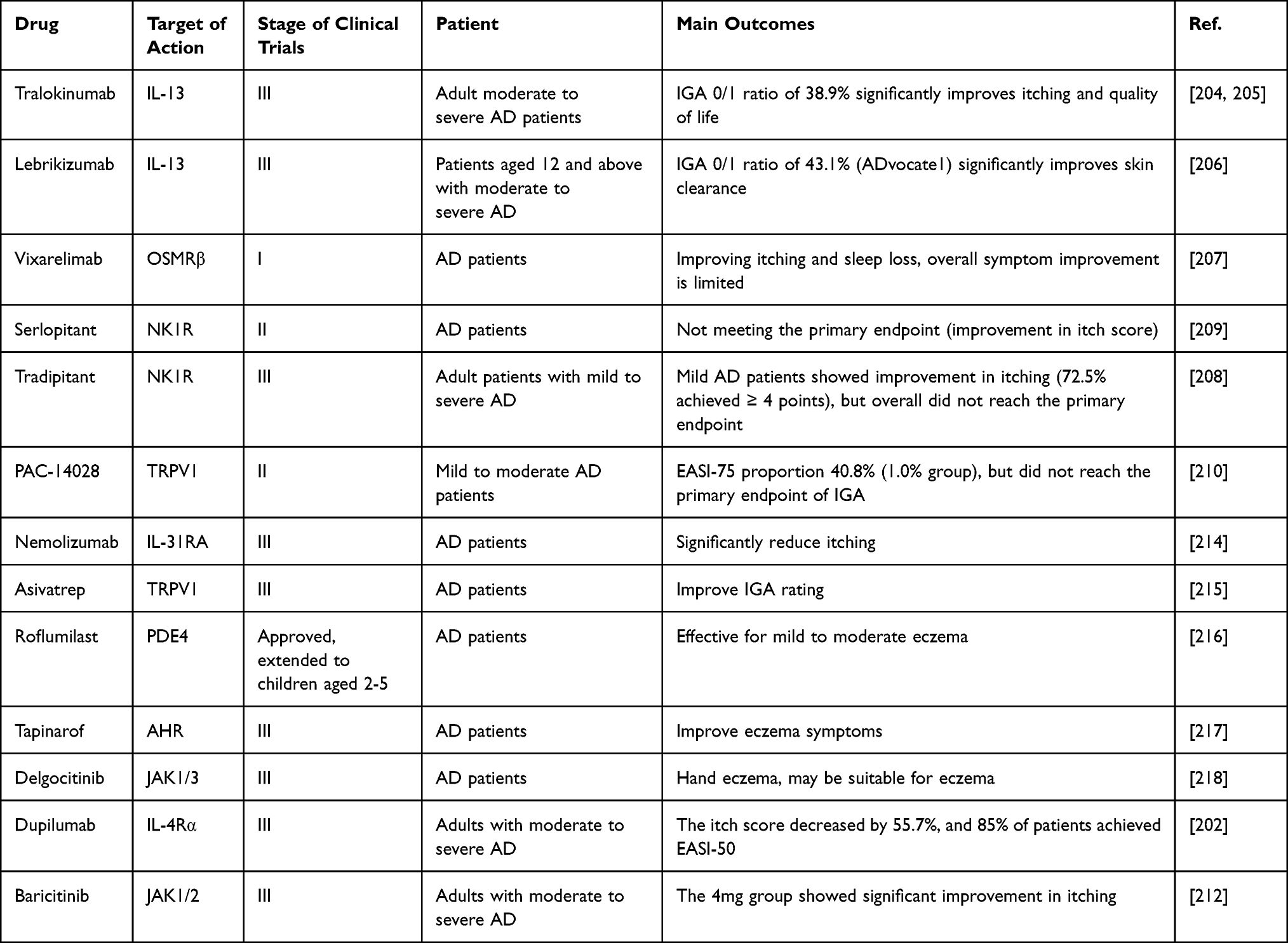 |
Table 4 Summary of Clinical Trials on AD |
Recent studies have shown that narrowband ultraviolet B (NBUVB) phototherapy remains a multifunctional adjuvant strategy, particularly effective in bridging the therapeutic gap between systemic biologics and local drugs by regulating the interrelated processes of neural sensitization and immune dysregulation that drive chronic AD attacks.219 In clinical practice, a multicenter observational study conducted in 2022 in adults with moderate to severe AD showed that the combination of three weekly sessions of NBUVB treatment (median cumulative dose of 21.5 J/cm 2 over 12 weeks) and dupilumab resulted in a 68% reduction in EASI score and a 75% improvement in pruritus NRS. Evidence from the 2b prospective cohort further supports that when NBUVB is used as a steroid backup adjuvant, remission can last up to six months, although non responders (over 20% of cases) emphasize the importance of selecting patients with baseline SCORAD>50 for optimal outcomes.220 In the pediatric population, neuroimmune hypersensitivity reactions usually manifest earlier, and NBUVB phototherapy has shown promising efficacy, although treatment regimens need to be adjusted. In 2018, a randomized controlled trial was conducted on children aged 5–12 years (n=30) using a low-dose regimen (0.5–1.5 MED increments, twice a week, for 10 weeks). Compared with placebo, PO-SCORD improved by 52%, providing level 1b evidence to support its safety and tolerability. Less than 10% of participants developed erythema, and no long-term photo induced cancer risk was observed during the two-year follow-up. Importantly, this pattern enhances the restoration of the epidermal barrier through upregulation of ceramides, reducing dupilumab activation rate by 40% in high load cases, demonstrating translation relevance.221 However, multiple studies have emphasized the limitations and potential risks of NBUVB phototherapy, including variable response rates, cumulative dose issues, and inconsistent long-term response persistence.222,223 Therefore, its widespread clinical application requires further confirmation through high-quality, large-scale evidence-based investigations to establish standardized protocols, optimize dosing strategies, and clarify long-term safety outcomes.
Emerging Neuroimmune-Targeted Therapies for AD: Mechanisms and Translational Horizons
Pioneering the forefront of neuroimmune regulation in AD - sustained peripheral neural circuits and chronic tissue remodeling permanently reinforce the self reinforcing itch inflammation cycle - emerging therapies targeting new molecular axes, including OX40/OX40L, TRP channels, MRGPRX2, and IL-31/OSMR β pathways, have transformative potential. These methods have addressed previously underserved neuronal hypersensitivity reactions and immune memory mechanisms, thereby reshaping the treatment landscape and achieving lasting remission and long-term disease improvement.224 Rocatinlimab is an OX40 inhibitor that can disrupt the OX40-OX40L interaction on activated T cells, leading to selective depletion of pathogenic Th2 and Th22 subsets while retaining regulatory T cells. This immune regulatory mechanism weakens cytokine driven sensitization by reducing IL-4 and IL-13 signaling, otherwise it enhances the expression of TRPV1 and SP release in sensory neurons. In the Phase III Rocket Program of 2025, the monotherapy of Rocatinib achieved EASI-75 response in 45% of moderate to severe AD patients by week 24, with sustained improvement in head and neck lesions translated into better quality of life and lower recurrence rates. These findings emphasize its potential for long-term disease modification in refractory AD cases.225 I At the same time, the OX40L antagonist amlitelimab blocks ligand mediated co stimulation to inhibit effector T cell proliferation and alleviate cytokine driven inflammation, thereby indirectly suppressing neuroimmune crosstalk by reducing the production of IL-31, which stimulates the discharge of itch neurons. A recent phase IIb trial showed that at week 52, molecular biomarkers and clinical severity continued to decrease, with 60% of patients maintaining an Investigator Global Assessment (IGA) score of 0 or 1, enhancing its translational value in preventing chronic neuroremodeling and positioning it as a differentiated therapy for inflammation dominant AD.226 Transitioning to a sensory regulation strategy, the local TRPV1 antagonist asivatrap works by selectively desensitizing capsaicin sensitive C fibers, thereby blocking proton or heat induced cation influx. This interrupts the release of neuropeptides, intensifies the activation of mast cells, and damages the epidermal barrier. The Phase III clinical data from 2021 confirms that it has significant efficacy in relieving itching and eczema severity in adolescents and adults, and has the potential to enhance local control when used in combination with systemic biologics such as dupilumab. In clinical practice, asivatrap provides a non immunosuppressive option that can reduce scratch induced flares and improve skin barrier function.227 Meanwhile, TRPA1 modulators - currently undergoing early human pharmacology evaluation - target oxidative stress-induced channel activation in mast cells and sensory neurons. In this pathway, ROS promotes the release of trypsin like enzymes and macrophage infiltration, leading to Th2 polarization and permanent amplification of itching. Hu et al demonstrated in an AD model that mechanical scratching activates TRPA1, which exacerbates inflammation through macrophage recruitment, paving the way for AD specific clinical trials targeting environmentally induced flares. Although human data is still preliminary, TRPA1 targeted drugs represent a promising adjuvant strategy for controlling oxidative stress-induced neuroimmune activation.228
In addition, recent studies have emphasized the therapeutic potential of MRGPRX2 antagonists such as EVO756, which dual targets mast cells and sensory neurons by inhibiting non IgE mediated degranulation. This can prevent the release of trypsin and histamine, thereby activating PAR2 on incoming neurons and disrupting the self sustained itch scratch cycle. In a preclinical study, Wong et al demonstrated that the relevant antagonist GE1111 alleviated DNFB induced AD in mice by reducing pro-inflammatory cytokines and restoring epidermal barrier integrity. EVO756 is currently undergoing phase IIb trials of AD and is being explored in combination with IL-31R α or JAK1 inhibitors to achieve synergistic neuroimmune blockade and redefine the treatment outcomes of pruritus type AD phenotype.229 In addition to this method, vixarelimab - a dual IL-31/OSMR β inhibitor - blocks the signaling of tumor suppressor M and IL-31, thereby alleviating itching and inflammation by inhibiting JAK-STAT activation in keratinocytes and sensory neurons. This mechanism has dual benefits of relieving itching and restoring the epidermal barrier. Although initially validated in nodular prurigo trials, a recent phase IIa study on AD showed that by week 8, approximately one-third of patients experienced rapid relief of itching and complete or near complete clearance of the skin, indicating that AD subtypes have different therapeutic effects compared to patients with concurrent pruritus dominant disease. However, further clinical trials are needed to clarify its comparative advantage over blocking the single IL-31 pathway.230 By combining clinical phenotypes with molecular mechanisms, stratified therapy algorithms now divide AD patients into different subgroups. Pruritus dominant phenotype, usually characterized by severe sleep disorders or nodular prurigo, may benefit most from initial IL-31R α targeted therapy, which may be supplemented with local or short-term oral JAK1 inhibitors to rapidly alleviate symptoms. In contrast, inflammation dominant endometrium associated with recurrence tendency or steroid dependent characteristics is favorable for IL-13 antibodies or OX40 (L) targeted biologics. The US Food and Drug Administration reviewed data for 2023 and further emphasized the long-term maintenance role of OX40 (L) inhibition in maintaining EASI score improvement and preventing disease recurrence.231 Sequential treatment strategies are also emerging to optimize disease control and long-term compliance. For example, oral JAK1 inhibitors were initially used to rapidly interrupt the itch inflammation feedback loop through extensive cytokine blockade, followed by IL-31R α antagonists for itch maintenance or IL-13/OX40 (L) inhibitors for long-term inflammation remodeling. This step-by-step accuracy based approach, supported by the SpringerLink guided algorithm, provides a dynamic treatment framework tailored to the patient’s specific disease trajectory.232 In addition, the rational integration of systemic and local patterns further enhances the synergistic effect of treatment. Local use of rosoliztinib cream can be used to treat focal lesions and reduce systemic treatment burden, while NBUVB phototherapy can be used as a staged adjuvant therapy for patients with extensive or refractory diseases. Incyte’s research has shown that when used in combination with systemic drugs, neuroimmune balance and barrier function are improved.233 In the near future, integrating these advances into the neuroimmune spectrum of AD will require multifaceted, data-driven research efforts. Future directions include multi omics guided clinical trials to improve combination regimens, explore biologics that regulate the microbiome, and develop novel delivery systems that enhance tissue specificity and minimize systemic toxicity. These innovations are expected to deepen our understanding of the mechanisms and advance personalized and durable treatment strategies for patients with different subtypes of AD.234
Future Research Directions and Challenges
In the current research landscape of AD, the integration of dermatology, immunology, and neuroscience has highlighted the profound interactions between neuronal circuits and immune effectors, driving a multidisciplinary revival aimed at decoding chronic itching and barrier dysfunction that signify this common disease. However, despite significant progress in understanding AD and its neuroimmune mechanisms, there are still some key challenges. AD exhibits high clinical heterogeneity, with patients presenting different symptoms, triggers, and treatment responses, typically associated with different neuroimmune features and comorbidities such as depression, anxiety, and sleep disorders.34,157,235,236 This means that treatment strategies must be tailored to each patient’s specific phenotype and immune system. Therefore, more research is needed on potential mechanisms to better understand the different forms of AD and develop more targeted treatment methods.237,238 Secondly, high treatment costs and long-term safety issues are obstacles to the widespread use of new therapies. Safety is particularly important when treating children and pregnant women.239,240 Although some new biological treatment methods, such as dupilumab and nemolizumab, have shown good clinical efficacy, their high cost and long-term effects still need further evaluation.241
To address these challenges, we believe that future research should focus on several key areas. Firstly, precision medicine methods should be used to identify AD subtypes through multi omics data (including genomics, transcriptomics, and proteomics) to treat patients. This strategy will develop personalized treatment plans based on the molecular characteristics of each patient, thereby improving treatment effectiveness while minimizing side effects.242,243 Secondly, new neuroimmune targets such as TSLP (thymic stromal lymphopoietin), IL-33, and TRPV1 channels are worth further exploration. Therapeutic interventions targeting these pathways may lead to more effective treatment methods, especially considering the critical roles of TRPV1 channels and MrgPRs in non-histamine dependent itching, which deserves further research and clinical translation.244–246 Thirdly, optimizing clinical trials through biomarker based patient stratification is a key priority as this approach can achieve more accurate participant recruitment and outcome prediction.140,247,248 For example, layering the trial through the high IL-31 phenotype of pruritus dominant AD, the high spectrum of osteocutaneous protein/DPP-4 driven by IL-13 pathway, or the high TARC group with an increased risk of recurrence can enhance treatment matching and reduce trial failure.140,247 This stratification strategy will facilitate targeted interventions, such as prioritizing anti-IL-31 therapy for itch focused cohorts or providing IL-13 inhibitors for subgroups with impaired barriers, thereby improving clinical trial efficiency and real-world applicability. Looking ahead, future research directions may utilize advanced multi omics platforms to identify personalized neuroimmune biomarkers. By analyzing machine learning algorithms from a comprehensive dataset of immunology, neuroscience, and bioinformatics, it is possible to establish predictive models for treatment response.249 In addition, exploring neuroregulatory therapies such as optogenetics or targeted neuroregulatory devices for pain management can provide innovative assistance to existing biologics by directly intervening in neural circuits to address residual itching in stubborn cases.39 Beyond cutaneous barrier disruption, accumulating evidence suggests that impairment of the intestinal barrier is a critical component of the gut–skin–brain axis in atopic dermatitis.250 In patients with AD, dysbiosis of the gut microbiota, together with reduced expression or altered distribution of tight junction proteins (such as claudins, occludin and zonula occludens proteins), can increase intestinal permeability (“leaky gut”).251,252 This facilitates the translocation of microbial products and dietary antigens into the systemic circulation, leading to low-grade systemic inflammation and skewing of immune responses toward type 2 and type 17 profiles. These circulating mediators may, in turn, influence cutaneous immune cells and sensory neurons, exacerbate pruritus and inflammation, and modulate central stress and emotional circuits that are closely linked to AD severity.253 Thus, impaired intestinal barrier function represents an important upstream driver of gut–microbiota–derived signals within the gut–skin–brain axis and may offer novel opportunities for preventive and therapeutic interventions targeting both the microbiome and barrier integrity.26
In addition, for the advancement of clinical applications, the development of combination therapies that focus on repairing the skin barrier, regulating the immune system, and controlling itching is an important area of future research. For example, combining skin barrier repair with immunotherapy can more comprehensively address the complex etiology of AD, effectively control symptoms, and reduce episodes.254,255 In addition, elucidating the role of skin microbiota in neuroimmune interactions provides promising therapeutic opportunities. Emerging evidence suggests that dysbiosis of the skin microbiome may play a key role in the pathogenesis of AD.256,257 Therefore, studying the interaction between microbial communities and immune responses can reveal new therapeutic targets.256,258 In addition, incorporating objective itch measurement tools into clinical studies will improve the accuracy of neuroimmune outcome assessment and comorbidity assessment. Beyond subjective scales such as the Peak Itching Rating Scale (PP-NRS), researchers can now use neuroimaging techniques such as wearable scratch monitoring devices, sleep trackers, and fMRI to assess central sensitivity.259,260 These techniques can quantify nighttime scratching, sleep fragmentation, and changes in the central nervous system network, providing strong endpoints for evaluating the impact of therapeutic interventions on the itch sleep depression cycle in real-world environments.21 In addition, these measures facilitate correlation analysis with health economic outcomes, enabling a comprehensive evaluation of the cost-effectiveness of treatment in reducing the burden of comorbidities.261 Ultimately, these evolving strategies are expected to bridge the current gap in understanding and pave the way for holistic interventions targeting the multifaceted neuroimmune landscape of AD.
Conclusion
In summary, accumulating evidence indicates that atopic dermatitis is not merely a barrier or immune disorder, but a prototypical disease of the neuroimmune axis, in which peripheral sensory neurons, immune cells, keratinocytes, and central stress pathways form tightly interconnected circuits that drive chronic pruritus, inflammation, and disease heterogeneity. By synthesizing current data on neuroimmune mediators and pathways—such as type 2 cytokines, IL-31, neuropeptides, nerve growth factor, and stress-related neuroendocrine signals—this review highlights how their reciprocal interactions sustain the itch–scratch–inflammation cycle and link cutaneous events to systemic and psychoneurological comorbidities. Advances in multi-omics, spatial profiling, neuroimaging, and microbiome research are beginning to identify candidate neuroimmune biomarkers that may support patient stratification, disease monitoring, and prediction of therapeutic response. At the same time, the rapid development of biologics and small molecules targeting key neuroimmune nodes, including IL-4/IL-13, IL-31, JAK–STAT signaling, and co-stimulatory pathways, is transforming the therapeutic landscape and providing proof-of-concept for neuroimmune-directed interventions. Together, these insights support a more integrated, mechanism-based view of atopic dermatitis and point toward future precision medicine strategies that combine neuroimmune profiling, biomarker-guided treatment selection, and comprehensive management of stress and psychosocial burden to achieve durable disease control and improved quality of life for patients.
Abbreviation
AD, Atopic Dermatitis; Th2, Type 2 Helper T Cells; IL, Interleukin; IL-4Rα, Interleukin-4 Receptor Alpha; JAK/STAT, Janus Kinase/Signal Transducer and Activator of Transcription Pathway; TRPV1, Receptor Potential Vanilloid 1; TRPA1, Transient Receptor Potential Ankyrin 1; SP, Substance P; CGRP, Calcitonin Gene-Related Peptide; VIP, Vasoactive Intestinal Peptide; NK1R, Neurokinin-1 Receptor; Treg, Regulatory T Cells; ILC2, Type 2 Innate Lymphoid Cells; NMU, Neuromedin U; NMUR1, Neuromedin U Receptor 1; NGF, Nerve Growth Factor; BDNF, Brain-Derived Neurotrophic Factor; DRG, Dorsal Root Ganglion; PAR2, Protease-Activated Receptor 2; MRGPRX2, Mas-related G Protein-Coupled Receptor X2; NRF2, Nuclear Factor E2-Related Factor 2; TSLP, Thymic Stromal Lymphopoietin; OSMR, Oncostatin M Receptor; OSMRβ, Oncostatin M Receptor Beta; IL-31RA, Interleukin-31 Receptor Alpha; OX40/OX40L, OX40/OX40 Ligand; TEWL, Transepidermal Water Loss; SCORAD, Atopic Dermatitis Severity Indices; EASI, Eczema Area and Severity Index; HC, Healthy Control; DEP, Differentially Expressed Protein; GO, Gene Ontology; PCA, Principal Component Analysis; IgE, Immunoglobulin E; DPP-4, Dipeptidyl Peptidase-4; TARC/CCL17, Thymus and Activation-Regulated Chemokine; MDC/CCL22, Macrophage-Derived Chemokine; CTACK/CCL27, Cutaneous T-Cell Attracting Chemokine; PARC/CCL18, Pulmonary and Activation-Regulated Chemokine; ECP, Eosinophil Cationic Protein; NBUVB, Narrowband Ultraviolet B; DNFB, 2,4-Dinitrofluorobenzene; ROS, Reactive Oxygen Species; FMT, Fecal Microbiota Transplantation.
Data Sharing Statement
Data availability is not applicable as no new data were generated or analyzed for this study.
Author Contributions
Zhongsong Zhang: Writing – review & editing, Writing – original draft, Validation, Methodology, Conceptualization, Data curation, Investigation, Software, Project Administration
Chao Chang: Writing – review & editing, Writing – original draft, Visualization, Data curation, Resources, Software, Supervision,
Lifan Xiao: Writing – review & editing, Writing – original draft, Validation, Data curation, Resources, Software, Methodology
Hang Su: Writing – review & editing, Writing – original draft, Conceptualization, Data curation, Resources, Validation, Visualization
Yaping Lyu: Writing – review & editing, Writing – original draft, Validation, Visualization, Formal analysis, Conceptualization, Software.
Junxian Zhao: Writing – review & editing, Resources, Formal Analysis, Funding acquisition, Project administration, Conceptualization.
Junhao Chen: Writing – review & editing, Resources, Formal Analysis, Funding acquisition, Conceptualization
Keyi Gou: Writing – review & editing, Resources, Formal Analysis, Data curation, Methodology
Jingfeng Zhou: Writing – review & editing, Resources, Data curation, Investigation
Chengjie Wang: Writing – review & editing, Resources, Supervision, Methodology
Xingcheng Zhu: Writing – review & editing, Resources, Methodology,
Shi Fu: Writing – review & editing, Data curation, Investigation, Software, Conceptualization, Supervision
Mao Lu: Writing – original draft, Visualization, Methodology, Conceptualization, Data curation, Investigation, Software, Supervision, Validation, Funding acquisition, Resources, Project administration, Formal analysis
Lei Shi: Writing – review & editing, Resources, Formal Analysis, Funding acquisition, Project administration, Writing – original draft, Visualization, Methodology, Conceptualization, Data curation, Investigation
All authors gave final approval of the version to be published, agreed on the journal to which the paper was submitted, and agree to be responsible for all the contents of this paper.
Funding
This research is supported by the Graduate Innovation Fund project of Kunming Medical University: 2025S249.
Disclosure
The authors declare that there are no competing interests associated with the paper.
References
1. Luo CH, Lai AC-Y, Tsai CC, et al. Staphylococcus aureus exacerbates dermal IL-33/ILC2 axis activation through evoking RIPK3/MLKL-mediated necroptosis of dry skin. JCI Insight. 2024;9:e185878. doi:10.1172/jci.insight.185878
2. Volke A, Toompere K, Laisaar K-T, et al. 12-month prevalence of atopic dermatitis in resource-rich countries: a systematic review and meta-analysis. Sci Rep. 2022;12:15125. doi:10.1038/s41598-022-19508-7
3. Huang -C-C, Lo Y-H, Hsu Y-J, et al. Anti-atopic dermatitis activity of epi-oxyzoanthamine isolated from zoanthid. Mar Drugs. 2023;21:447. doi:10.3390/md21080447
4. Hu Y, Zhao J, Shen Q, et al. Early-life quarantine increases the development of infant atopic dermatitis: a birth cohort study in China. Allergy. 2025;80:1526–1529. doi:10.1111/all.16561
5. Ou Y, Shao X, Zhang J, Chen J. Global, regional, and national burden of older adult atopic dermatitis in 204 countries and territories worldwide. Front Public Health. 2025;13:1569119. doi:10.3389/fpubh.2025.1569119
6. Chuang -C-C, Braham-Chaouche L, Thomas R, Mnif T. Epidemiology and burden of pediatric atopic dermatitis in China. Dermatol Ther. 2025;15:1319–1329. doi:10.1007/s13555-025-01403-4
7. Tsuge M, Ikeda M, Matsumoto N, Yorifuji T, Tsukahara H. Current insights into atopic march. Children. 2021;8:1067. doi:10.3390/children8111067
8. Kader HA, Azeem M, Jwayed SA, et al. Current insights into immunology and novel therapeutics of atopic dermatitis. Cells. 2021;10:1392. doi:10.3390/cells10061392
9. Narla S, Silverberg JI. The role of environmental exposures in atopic dermatitis. Curr Allergy Asthma Rep. 2020;20:74. doi:10.1007/s11882-020-00971-z
10. Kantor R, Silverberg JI. Environmental risk factors and their role in the management of atopic dermatitis. Expert Rev Clin Immunol. 2017;13:15–26. doi:10.1080/1744666X.2016.1212660
11. Davidson WF, Leung DYM, Beck LA, et al. Report from the national institute of allergy and infectious diseases workshop on “atopic dermatitis and the atopic march: mechanisms and interventions. J Allergy Clin Immunol. 2019;143:894–913. doi:10.1016/j.jaci.2019.01.003
12. Bradshaw LE, Wyatt LA, Brown SJ, et al. Emollient application from birth to prevent eczema in high-risk children: the BEEP RCT. Health Technol Assess. 2024;28:1–116. doi:10.3310/RHDN9613
13. Chu DK, Chu AWL, Rayner DG, et al. Topical treatments for atopic dermatitis (eczema): systematic review and network meta-analysis of randomized trials. J Allergy Clin Immunol. 2023;152:1493–1519. doi:10.1016/j.jaci.2023.08.030
14. Murota H, Nakahara T, Wang X, et al. Systematic literature review and network meta-analysis of clinical efficacy and safety of topical treatments for patients with atopic dermatitis. Dermatol Ther. 2025;15:1045–1062. doi:10.1007/s13555-025-01390-6
15. Fan J, Mishra SK. The emerging role of neuroimmune interactions in atopic dermatitis and itch. FEBS J. 2022;289:2723–2735. doi:10.1111/febs.15860
16. Liu AW, Gillis JE, Sumpter TL, Kaplan DH. Neuroimmune interactions in atopic and allergic contact dermatitis. J Allergy Clin Immunol. 2023;151:1169–1177. doi:10.1016/j.jaci.2023.03.013
17. Brabenec L, Gupta S, Eichwald T, Rafei M, Talbot S. Decoding the neuroimmune axis in the atopic march: mechanisms and implications. Curr Opin Immunol. 2024;91:102507. doi:10.1016/j.coi.2024.102507
18. Sun P-Y, Li H-G, Xu Q-Y, et al. Lidocaine alleviates inflammation and pruritus in atopic dermatitis by blocking different population of sensory neurons. Br J Pharmacol. 2023;180:1339–1361. doi:10.1111/bph.16012
19. Choi JE, Di Nardo A. Skin neurogenic inflammation. Semin Immunopathol. 2018;40:249–259. doi:10.1007/s00281-018-0675-z
20. Kim JH, Samra MS. Moderate to severe atopic dermatitis in children: focus on systemic Th2 cytokine receptor antagonists and Janus kinase inhibitors. Clin Exp Pediatr. 2024;67:64–79. doi:10.3345/cep.2022.00346
21. Cameron S, Donnelly A, Broderick C, et al. Mind and skin: exploring the links between inflammation, sleep disturbance and neurocognitive function in patients with atopic dermatitis. Allergy. 2024;79:26–36. doi:10.1111/all.15818
22. Ahn J, Shin S, Lee GC, et al. Unraveling the link between atopic dermatitis and autoimmune diseases in children: insights from a large-scale cohort study with 15-year follow-up and shared gene ontology analysis. Allergol Int. 2024;73:243–254. doi:10.1016/j.alit.2023.12.005
23. Sidbury R, Alpizar S, Laquer V, et al. Pharmacokinetics, safety, efficacy, and biomarker profiles during nemolizumab treatment of atopic dermatitis in adolescents. Dermatol Ther. 2022;12:631–642. doi:10.1007/s13555-021-00678-7
24. Lu Z, Xiao S, Chen W, et al. IL-20 promotes cutaneous inflammation and peripheral itch sensation in atopic dermatitis. FASEB J. 2022;36:e22334. doi:10.1096/fj.202101800R
25. Hashimoto T, Okuno S. The big four in the pathogenesis and pathophysiology of prurigo nodularis: interplay among type 2 inflammation, epidermal hyperplasia, dermal fibrosis, and itch from neuroimmune dysregulation. Clin Dermatol. 2025;43:S0738–081X(25)00093–8. doi:10.1016/j.clindermatol.2025.03.010
26. Steinhoff M, Ahmad F, Pandey A, et al. Neuroimmune communication regulating pruritus in atopic dermatitis. J Allergy Clin Immunol. 2022;149:1875–1898. doi:10.1016/j.jaci.2022.03.010
27. Meng J, Moriyama M, Feld M, et al. New mechanism underlying IL-31-induced atopic dermatitis. J Allergy Clin Immunol. 2018;141:1677–1689.e8. doi:10.1016/j.jaci.2017.12.1002
28. Datsi A, Steinhoff M, Ahmad F, Alam M, Buddenkotte J. Interleukin-31: the “itchy” cytokine in inflammation and therapy. Allergy. 2021;76:2982–2997. doi:10.1111/all.14791
29. Kwatra SG, Misery L, Clibborn C, Steinhoff M. Molecular and cellular mechanisms of itch and pain in atopic dermatitis and implications for novel therapeutics. Clin Transl Immunol. 2022;11:e1390. doi:10.1002/cti2.1390
30. Orfali RL, Aoki V. Blockage of the IL-31 pathway as a potential target therapy for atopic dermatitis. Pharmaceutics. 2023;15:577. doi:10.3390/pharmaceutics15020577
31. Wilson SR, Thé L, Batia LM, et al. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell. 2013;155:285–295. doi:10.1016/j.cell.2013.08.057
32. Luo J, Zhu Z, Zhai Y, et al. The role of tslp in atopic dermatitis: from pathogenetic molecule to therapeutical target. Mediators Inflamm. 2023;2023:7697699. doi:10.1155/2023/7697699
33. Ruppenstein A, Limberg MM, Loser K, Kremer AE, Homey B, Raap U. Involvement of neuro-immune interactions in pruritus with special focus on receptor expressions. Front Med. 2021;8:627985. doi:10.3389/fmed.2021.627985
34. Facheris P, Jeffery J, Del Duca E, Guttman-Yassky E. The translational revolution in atopic dermatitis: the paradigm shift from pathogenesis to treatment. Cell Mol Immunol. 2023;20:448–474. doi:10.1038/s41423-023-00992-4
35. Mahmoud O, Soares GB, Yosipovitch G. Transient receptor potential channels and itch. Int J Mol Sci. 2022;24:420. doi:10.3390/ijms24010420
36. Tsagareli MG, Follansbee T, Iodi Carstens M, Carstens E. Targeting Transient Receptor Potential (TRP) channels, Mas-Related G-Protein-Coupled Receptors (Mrgprs), and Protease-Activated Receptors (PARs) to relieve itch. Pharmaceuticals. 2023;16:1707. doi:10.3390/ph16121707
37. Yosipovitch G, Kim B, Luger T, et al. Similarities and differences in peripheral itch and pain pathways in atopic dermatitis. J Allergy Clin Immunol. 2024;153:904–912. doi:10.1016/j.jaci.2023.10.034
38. Go EJ, Lee JY, Kim YH, Park C-K. Site-specific transient receptor potential channel mechanisms and their characteristics for targeted chronic itch treatment. Biomolecules. 2024;14:107. doi:10.3390/biom14010107
39. Kim B, Rothenberg ME, Sun X, et al. Neuroimmune interplay during type 2 inflammation: symptoms, mechanisms, and therapeutic targets in atopic diseases. J Allergy Clin Immunol. 2024;153:879–893. doi:10.1016/j.jaci.2023.08.017
40. Thapaliya M, Chompunud Na Ayudhya C, Amponnawarat A, Roy S, Ali H. Mast cell-specific MRGPRX2: a key modulator of neuro-immune interaction in allergic diseases. Curr Allergy Asthma Rep. 2021;21:3. doi:10.1007/s11882-020-00979-5
41. Nagamine M, Kaitani A, Izawa K, et al. Neuronal substance P-driven MRGPRX2-dependent mast cell degranulation products differentially promote vascular permeability. Front Immunol. 2024;15:1477072. doi:10.3389/fimmu.2024.1477072
42. Aitella E, Romano C, Ginaldi L, Cozzolino D. Mast cells at the crossroads of hypersensitivity reactions and neurogenic inflammation. Int J Mol Sci. 2025;26:927. doi:10.3390/ijms26030927
43. Weihrauch T, Limberg MM, Gray N, Schmelz M, Raap U. Neurotrophins: neuroimmune interactions in human atopic diseases. Int J Mol Sci. 2023;24:6105. doi:10.3390/ijms24076105
44. Raap U, Deneka N, Bruder M, Kapp A, Wedi B. Differential up-regulation of neurotrophin receptors and functional activity of neurotrophins on peripheral blood eosinophils of patients with allergic rhinitis, atopic dermatitis and nonatopic subjects. Clin Exp Allergy. 2008;38:1493–1498. doi:10.1111/j.1365-2222.2008.03035.x
45. Xiao Z, Zhuo Y, Li R, Tan Y. The recent advances of mast cells in the pathogenesis of atopic dermatitis. Front Allergy. 2025;6:1668742. doi:10.3389/falgy.2025.1668742
46. Siiskonen H, Harvima I. Mast cells and sensory nerves contribute to neurogenic inflammation and pruritus in chronic skin inflammation. Front Cell Neurosci. 2019;13:422. doi:10.3389/fncel.2019.00422
47. Meng J, Li Y, Fischer MJM, Steinhoff M, Chen W, Wang J. Th2 modulation of transient receptor potential channels: an unmet therapeutic intervention for atopic dermatitis. Front Immunol. 2021;12:696784. doi:10.3389/fimmu.2021.696784
48. Cevikbas F, Wang X, Akiyama T, et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: involvement of TRPV1 and TRPA1. J Allergy Clin Immunol. 2014;133:448–460. doi:10.1016/j.jaci.2013.10.048
49. Guseva D, Rüdrich U, Kotnik N, et al. Neuronal branching of sensory neurons is associated with BDNF-positive eosinophils in atopic dermatitis. Clin Exp Allergy. 2020;50:577–584. doi:10.1111/cea.13560
50. Lee JJ, Protheroe CA, Luo H, et al. Eosinophil-dependent skin innervation and itching following contact toxicant exposure in mice. J Allergy Clin Immunol. 2015;135:477–487. doi:10.1016/j.jaci.2014.07.003
51. Singto T, Filor V, Vidak J, Klopfleisch R, Bäumer W. Dendritic cells under allergic condition enhance the activation of pruritogen-responsive neurons via inducing itch receptors in a co-culture study. BMC Immunol. 2024;25:17. doi:10.1186/s12865-024-00604-4
52. Wilcox NC, Taheri G, Halievski K, Talbot S, Silva JR, Ghasemlou N. Interactions between skin-resident dendritic and Langerhans cells and pain-sensing neurons. J Allergy Clin Immunol. 2024;154:11–19. doi:10.1016/j.jaci.2024.03.006
53. Kasraie S, Werfel T. Role of macrophages in the pathogenesis of atopic dermatitis. Mediators Inflamm. 2013;2013:942375. doi:10.1155/2013/942375
54. Gu S, Xu L, Huang B, Xiong K, Yang X, Ye J. Decoding macrophage dynamics: a pathway to understanding and treating inflammatory skin diseases. Int J Mol Sci. 2025;26:4287. doi:10.3390/ijms26094287
55. Lee SH, Sacks DL. Resilience of dermis resident macrophages to inflammatory challenges. Exp Mol Med. 2024;56:2105–2112. doi:10.1038/s12276-024-01313-z
56. Meng J, Li Y, Fischer MJM, Steinhoff M, Chen W, Wang J. Th2 modulation of transient receptor potential channels: an unmet therapeutic intervention for atopic dermatitis. Front Immunol. 2021;12. doi: 10.3389/fimmu.2021.696784
57. Liu AW, Gillis JE, Sumpter TL, Kaplan DH. Neuroimmune interactions in atopic and allergic contact dermatitis. J Allergy Clin Immunol. 2023;151:1169–1177. doi:10.1016/j.jaci.2023.03.013
58. Ye Y, Luo J, Zeng N, et al. Neuromedin U promotes human type 2 immune responses. Mucosal Immunol. 2022;15:990–999. doi:10.1038/s41385-022-00543-6
59. Ju X, Nagashima A, Dvorkin-Gheva A, et al. Neuromedin-U mediates rapid activation of airway group 2 innate lymphoid cells in mild asthma. Am J Respir Crit Care Med. 2024;210:755–765. doi:10.1164/rccm.202311-2164OC
60. Marek-Jozefowicz L, Nedoszytko B, Grochocka M, et al. Molecular Mechanisms of Neurogenic Inflammation of the Skin. Int J Mol Sci. 2023;24:5001. doi:10.3390/ijms24055001
61. Oetjen LK, Kim BS. Interactions of the immune and sensory nervous systems in atopy. FEBS J. 2018;285:3138–3151. doi:10.1111/febs.14465
62. Kim YJ, Granstein RD. Roles of calcitonin gene-related peptide in the skin, and other physiological and pathophysiological functions. Brain Behav Immun. 2021;18:100361. doi:10.1016/j.bbih.2021.100361
63. Yosipovitch G, Berger T, Fassett MS. Neuroimmune interactions in chronic itch of atopic dermatitis. J Eur Acad Dermatol Venereol. 2020;34:239–250. doi:10.1111/jdv.15973
64. Kobayashi T, Moro K. Tissue-specific diversity of group 2 innate lymphoid cells in the skin. Front Immunol. 2022;13:885642. doi:10.3389/fimmu.2022.885642
65. Imai Y. ILC2s in skin disorders. Allergol Int. 2023;72:201–206. doi:10.1016/j.alit.2023.01.002
66. Jia H, Wan H, Zhang D. Innate lymphoid cells: a new key player in atopic dermatitis. Front Immunol. 2023;14:1277120. doi:10.3389/fimmu.2023.1277120
67. Huang X, Li F, Wang F. Neural regulation of innate immunity in inflammatory skin diseases. Pharmaceuticals. 2023;16:246. doi:10.3390/ph16020246
68. Bieber T. Atopic dermatitis: an expanding therapeutic pipeline for a complex disease. Nat Rev Drug Discov. 2022;21:21–40. doi:10.1038/s41573-021-00266-6
69. Ferreira S, Torres T. Dupilumab for the treatment of atopic dermatitis. Actas Dermosifiliogr. 2018;109:230–240. doi:10.1016/j.ad.2017.10.012
70. Oetjen LK, Kim BS. Interactions of the immune and sensory nervous systems in atopy. FEBS J. 2018;285:3138–3151. doi:10.1111/febs.14465
71. Hall JM, Cruser D, Podawiltz A, Mummert DI, Jones H, Mummert ME. Psychological stress and the cutaneous immune response: roles of the HPA axis and the sympathetic nervous system in atopic dermatitis and psoriasis. Dermatol Res Pract. 2012;2012:403908. doi:10.1155/2012/403908
72. Escalante A, Serra-Baldrich E. Pathogenic mechanisms underlying itch in atopic dermatitis: the emerging role of neuroimmune interactions. Eur J Dermatol. 2023;33:343–349. doi:10.1684/ejd.2023.4514
73. Zhang H, Wang M, Zhao X, Wang Y, Chen X, Su J. Role of stress in skin diseases: a neuroendocrine-immune interaction view. Brain Behav Immun. 2024;116:286–302. doi:10.1016/j.bbi.2023.12.005
74. Mar K, Rivers JK. The mind body connection in dermatologic conditions: a literature review. J Cutan Med Surg. 2023;27:628–640. doi:10.1177/12034754231204295
75. Alexopoulos A, Chrousos GP. Stress-related skin disorders. Rev Endocr Metab Disord. 2016;17:295–304. doi:10.1007/s11154-016-9367-y
76. Arndt J, Smith N, Tausk F. Stress and atopic dermatitis. Curr Allergy Asthma Rep. 2008;8:312–317. doi:10.1007/s11882-008-0050-6
77. Mochizuki H, Schut C, Nattkemper LA, Yosipovitch G. Brain mechanism of itch in atopic dermatitis and its possible alteration through non-invasive treatments. Allergol Int. 2017;66:14–21. doi:10.1016/j.alit.2016.08.013
78. Ishiuji Y, Coghill RC, Patel TS, Oshiro Y, Kraft RA, Yosipovitch G. Distinct patterns of brain activity evoked by histamine-induced itch reveal an association with itch intensity and disease severity in atopic dermatitis. Br J Dermatol. 2009;161:1072–1080. doi:10.1111/j.1365-2133.2009.09308.x
79. Schut C, Mochizuki H, Grossman SK, et al. Brain processing of contagious itch in patients with atopic dermatitis. Front Psychol. 2017;8:1267. doi:10.3389/fpsyg.2017.01267
80. Napadow V, Li A, Loggia ML, et al. The imagined itch: brain circuitry supporting nocebo-induced itch in atopic dermatitis patients. Allergy. 2015;70:1485–1492. doi:10.1111/all.12727
81. Pondeljak N, Lugović-Mihić L. Stress-induced interaction of skin immune cells, hormones, and neurotransmitters. Clin Ther. 2020;42:757–770. doi:10.1016/j.clinthera.2020.03.008
82. Peters EM, Liezmann C, Spatz K, et al. Nerve growth factor partially recovers inflamed skin from stress-induced worsening in allergic inflammation. J Invest Dermatol. 2011;131. doi: 10.1038/jid.2010.317
83. Suárez AL, Feramisco JD, Koo J, Steinhoff M. Psychoneuroimmunology of psychological stress and atopic dermatitis: pathophysiologic and therapeutic updates. Acta Derm Venereol. 2012;92:7–15. doi:10.2340/00015555-1188
84. Tran BW, Papoiu ADP, Russoniello CV, et al. Effect of itch, scratching and mental stress on autonomic nervous system function in atopic dermatitis. Acta Derm Venereol. 2010;90:354–361. doi:10.2340/00015555-0890
85. Huang X, Li F, Wang F. Neural regulation of innate immunity in inflammatory skin diseases. Pharmaceuticals. 2023;16:246. doi:10.3390/ph16020246
86. Li C, Kim HJ, Back SK, Na HS. Common and discrete mechanisms underlying chronic pain and itch: peripheral and central sensitization. Pflugers Arch. 2021;473:1603–1615. doi:10.1007/s00424-021-02599-y
87. Guseva D, Rüdrich U, Kotnik N, et al. Neuronal branching of sensory neurons is associated with BDNF-positive eosinophils in atopic dermatitis. Clin Exp Allergy. 2020;50:577–584. doi:10.1111/cea.13560
88. Murota H, Katayama I. Exacerbating factors of itch in atopic dermatitis. Allergol Int. 2017;66:8–13. doi:10.1016/j.alit.2016.10.005
89. Siiskonen H, Harvima I. Mast cells and sensory nerves contribute to neurogenic inflammation and pruritus in chronic skin inflammation. Front Cell Neurosci. 2019;13. doi: 10.3389/fncel.2019.00422
90. Zhang Y, Zhang H, Jiang B, Tong X, Yan S, Lu J. Current views on neuropeptides in atopic dermatitis. Exp Dermatol. 2021;30:1588–1597. doi:10.1111/exd.14382
91. Biedermann T, Skabytska Y, Kaesler S, Volz T. Regulation of T cell immunity in atopic dermatitis by microbes: the yin and yang of cutaneous inflammation. Front Immunol. 2015;6:353. doi:10.3389/fimmu.2015.00353
92. Kim BE, Leung DYM. Significance of skin barrier dysfunction in atopic dermatitis. Allergy Asthma Immunol Res. 2018;10:207–215. doi:10.4168/aair.2018.10.3.207
93. Liu AW, Zhang YR, Chen C-S, et al. Scratching promotes allergic inflammation and host defense via neurogenic mast cell activation. Science. 2025;387:eadn9390. doi:10.1126/science.adn9390
94. Trier AM, Kim BS. Insights into atopic dermatitis pathogenesis lead to newly approved systemic therapies. Br J Dermatol. 2023;188:698–708. doi:10.1093/bjd/ljac016
95. Conway AE, Verdi M, Kartha N, et al. Allergic diseases and mental health. J Allergy Clin Immunol Pract. 2024;12:2298–2309. doi:10.1016/j.jaip.2024.05.049
96. Ständer S, Yosipovitch G. Substance P and neurokinin 1 receptor are new targets for the treatment of chronic pruritus. Br J Dermatol. 2019;181:932–938. doi:10.1111/bjd.18025
97. Alam M, Buddenkotte J, Ahmad F, Steinhoff M. Neurokinin 1 receptor antagonists for pruritus. Drugs. 2021;81:621–634. doi:10.1007/s40265-021-01478-1
98. Yosipovitch G, Berger T, Fassett MS. Neuroimmune interactions in chronic itch of atopic dermatitis. J Eur Acad Dermatol Venereol. 2020;34:239–250. doi:10.1111/jdv.15973
99. Liu AW, Gillis JE, Sumpter TL, Kaplan DH. Neuroimmune interactions in atopic and allergic contact dermatitis. J Allergy Clin Immunol. 2023;151:1169. doi:10.1016/j.jaci.2023.03.013
100. He H, Suryawanshi H, Morozov P, et al. Single-cell transcriptome analysis of human skin identifies novel fibroblast subpopulation and enrichment of immune subsets in atopic dermatitis. J Allergy Clin Immunol. 2020;145:1615–1628. doi:10.1016/j.jaci.2020.01.042
101. Jin S-P, Lee K, Bang YJ, et al. Mapping the immune cell landscape of severe atopic dermatitis by single-cell RNA-seq. Allergy. 2024;79:1584–1597. doi:10.1111/all.16121
102. Zhou J, Liang G, Liu L, et al. Single-cell RNA-seq reveals abnormal differentiation of keratinocytes and increased inflammatory differentiated keratinocytes in atopic dermatitis. J Eur Acad Dermatol Venereol. 2023;37:2336–2348. doi:10.1111/jdv.19256
103. Teresiak-Mikołajczak E, Czarnecka-Operacz M, Jenerowicz D, Silny W. Neurogenic markers of the inflammatory process in atopic dermatitis: relation to the severity and pruritus. Postepy Dermatol Alergol. 2013;30:286–292. doi:10.5114/pdia.2013.38357
104. Woo YR, Moon JH, Shin HY, et al. Systemic inflammatory proteomic biomarkers in atopic dermatitis: exploring potential indicators for disease severity. J Korean Med Sci. 2024;39:e223. doi:10.3346/jkms.2024.39.e223
105. Koch M, Kockmann T, Rodriguez E, et al. Quantitative proteomics identifies reduced nrf2 activity and mitochondrial dysfunction in atopic dermatitis. J Invest Dermatol. 2023;143:220–231.e7. doi:10.1016/j.jid.2022.08.048
106. Schmidt AD, de Guzman Strong C. Current understanding of epigenetics in atopic dermatitis. Exp Dermatol. 2021;30:1150–1155. doi:10.1111/exd.14392
107. da Silva Duarte AJ, Sanabani SS. Deciphering epigenetic regulations in the inflammatory pathways of atopic dermatitis. Life Sci. 2024;348:122713. doi:10.1016/j.lfs.2024.122713
108. Chen C, Zeng J, Lu J. Critical role of epigenetic modification in the pathogenesis of atopic dermatitis. Indian J Dermatol Venereol Leprol. 2023;89:700–709. doi:10.25259/IJDVL_298_2021
109. Mitamura Y, Reiger M, Kim J, et al. Spatial transcriptomics combined with single-cell RNA-sequencing unravels the complex inflammatory cell network in atopic dermatitis. Allergy. 2023;78:2215–2231. doi:10.1111/all.15781
110. He H, Suryawanshi H, Morozov P, et al. Single-cell transcriptome analysis of human skin identifies novel fibroblast subpopulation and enrichment of immune subsets in atopic dermatitis. J Allergy Clin Immunol. 2020;145:1615–1628. doi:10.1016/j.jaci.2020.01.042
111. Schäbitz A, Hillig C, Mubarak M, et al. Spatial transcriptomics landscape of lesions from non-communicable inflammatory skin diseases. Nat Commun. 2022;13:7729. doi:10.1038/s41467-022-35319-w
112. Hu T, Todberg T, Ewald DA, et al. Assessment of spatial and temporal variation in the skin transcriptome of atopic dermatitis by use of 1.5 mm minipunch biopsies. J Invest Dermatol. 2023;143:612–620.e6. doi:10.1016/j.jid.2022.10.004
113. Fiskin E, Eraslan G, Alora-Palli MB, et al. Multi-modal skin atlas identifies a multicellular immune-stromal community associated with altered cornification and specific T cell expansion in atopic dermatitis. bioRxiv. 2023:
114. Chen X, Cao B, Tan Z, et al. Integrated multi-omics analyses reveal innovative diagnostic and therapeutic targets associated with atopic dermatitis. J Inflamm Res. 2025;18:7951–7972. doi:10.2147/JIR.S526983
115. Nakajima S, Nakamizo S, Nomura T, Ishida Y, Sawada Y, Kabashima K. Integrating multi-omics approaches in deciphering atopic dermatitis pathogenesis and future therapeutic directions. Allergy. 2024;79:2366–2379. doi:10.1111/all.16183
116. Rusiñol L, Puig L. Multi-omics approach to improved diagnosis and treatment of atopic dermatitis and psoriasis. Int J Mol Sci. 2024;25:1042. doi:10.3390/ijms25021042
117. Gryak J, Georgievska A, Zhang J, et al. Prediction of pediatric peanut oral food challenge outcomes using machine learning. J Allergy Clin Immunol Glob. 2024;3:100252. doi:10.1016/j.jacig.2024.100252
118. Mercadié A, Gravier É, Josse G, et al. NMFProfiler: a multi-omics integration method for samples stratified in groups. Bioinformatics. 2025;41:btaf066. doi:10.1093/bioinformatics/btaf066
119. Sasaki A, Sargen M, Maskey AR, Li X-M. Scratching the surface: biomarkers and neurobiomarkers for improved allergic contact dermatitis management. Front Allergy. 2025;6:1564528. doi:10.3389/falgy.2025.1564528
120. Rios-Carlos M, Cervantes-García D, Córdova-Dávalos LE, Bermúdez-Humarán LG, Salinas E. Unraveling the gut-skin axis in atopic dermatitis: exploiting insights for therapeutic strategies. Gut Microbes. 2024;16:2430420. doi:10.1080/19490976.2024.2430420
121. Kim HB, Alexander H, Um JY, et al. Skin microbiome dynamics in atopic dermatitis: understanding host-microbiome interactions. Allergy Asthma Immunol Res. 2025;17:165–180. doi:10.4168/aair.2025.17.2.165
122. Díez-Madueño K, de la Cueva Dobao P, Torres-Rojas I, Fernández-Gosende M, Hidalgo-Cantabrana C, Coto-Segura P. Gut dysbiosis and adult atopic dermatitis: a systematic review. J Clin Med. 2024;14:19. doi:10.3390/jcm14010019
123. Wrześniewska M, Wołoszczak J, Świrkosz G, Szyller H, Gomułka K. The role of the microbiota in the pathogenesis and treatment of atopic dermatitis-a literature review. Int J Mol Sci. 2024;25:6539. doi:10.3390/ijms25126539
124. Minić-Pantić D, Abela B, Lehtimäki J, et al. Examining atopic dermatitis through the one health concept lens. Allergy. 2025. doi:10.1111/all.70080
125. Borrego-Ruiz A, Borrego JJ. Nutritional and microbial strategies for treating acne, alopecia, and atopic dermatitis. Nutrients. 2024;16:3559. doi:10.3390/nu16203559
126. Kim HB, Alexander H, Um JY, et al. Skin microbiome dynamics in atopic dermatitis: understanding host-microbiome interactions. Allergy Asthma Immunol Res. 2025;17:165–180. doi:10.4168/aair.2025.17.2.165
127. Giorgio CM, Licata G, Sorbo R, et al. Transforming atopic dermatitis management: probiotics as a game-changer in immune modulation-a double-blind, placebo-controlled clinical trial. Dermatitis. 2025. doi:10.1177/17103568251367725
128. Song H, Mun S-H, Han D-W, et al. Probiotics ameliorate atopic dermatitis by modulating the dysbiosis of the gut microbiota in dogs. BMC Microbiol. 2025;25:228. doi:10.1186/s12866-025-03924-6
129. Wang P, Yang L, Chen L, et al. Phytic acid improves the skin barrier function in murine models of atopic dermatitis through skin microbiota-derived indole-3-propionic acid. Allergy. 2025;80:2258–2270. doi:10.1111/all.16644
130. Choi H, Kwak M-J, Choi Y, et al. Extracellular vesicles of Limosilactobacillus fermentum SLAM216 ameliorate skin symptoms of atopic dermatitis by regulating gut microbiome on serotonin metabolism. Gut Microbes. 2025;17:2474256. doi:10.1080/19490976.2025.2474256
131. Thomas J, Sachdeva M, Dhar S, et al. Delphi consensus statement on the role of probiotics in the treatment of atopic dermatitis. Cureus. 2024;16:e64583. doi:10.7759/cureus.64583
132. Zhang Z, Wang R, Li M, Lu M. Current insights and trends in atopic dermatitis and microbiota interactions: a systematic review and bibliometric analysis. Front Microbiol. 2025;16:1613315. doi:10.3389/fmicb.2025.1613315
133. Qu B, Zhang X-E, Feng H, et al. Microbial perspective on the skin-gut axis and atopic dermatitis. Open Life Sci. 2024;19:20220782. doi:10.1515/biol-2022-0782
134. Qu B, Zhang X-E, Feng H, et al. Microbial perspective on the skin-gut axis and atopic dermatitis. Open Life Sci. 2024;19:20220782. doi:10.1515/biol-2022-0782
135. Chen M, Wang R, Wang T. Gut microbiota and skin pathologies: mechanism of the gut-skin axis in atopic dermatitis and psoriasis. Int Immunopharmacol. 2024;141:112658. doi:10.1016/j.intimp.2024.112658
136. Bakker D, de Bruin-Weller M, Drylewicz J, van Wijk F, Thijs J. Biomarkers in atopic dermatitis. J Allergy Clin Immunol. 2023;151:1163–1168. doi:10.1016/j.jaci.2023.01.019
137. Renert-Yuval Y, Thyssen JP, Bissonnette R, et al. Biomarkers in atopic dermatitis-a review on behalf of the International Eczema Council. J Allergy Clin Immunol. 2021;147:1174–1190.e1. doi:10.1016/j.jaci.2021.01.013
138. Gorelick J, Nguyen A, Schneider SKR, Martel BC, Madsen DE, Armstrong AW. Biomarkers in atopic dermatitis: a review of the role of IL-13 and the impact of tralokinumab treatment. Am J Clin Dermatol. 2025;26:199–211. doi:10.1007/s40257-024-00913-9
139. Yong TL, Zaman R, Rehman N, Tan CK. Ceramides and skin health: new insights. Exp Dermatol. 2025;34:e70042. doi:10.1111/exd.70042
140. Indolfi C, Grella C, Klain A, et al. Biomarkers in atopic dermatitis in children: a comprehensive review. Life. 2025;15:375. doi:10.3390/life15030375
141. Mortlock RD, Ma EC, Cohen JM, Damsky W. Assessment of treatment-relevant immune biomarkers in psoriasis and atopic dermatitis: toward personalized medicine in dermatology. J Invest Dermatol. 2023;143:1412–1422. doi:10.1016/j.jid.2023.04.005
142. Bai R, Zheng Y, Dai X. Atopic dermatitis: diagnosis, molecular pathogenesis, and therapeutics. Mol Biomed. 2025;6:71. doi:10.1186/s43556-025-00313-3
143. Park CO, Kim SM, Lee KH, Bieber T. Biomarkers for phenotype-endotype relationship in atopic dermatitis: a critical review. eBioMedicine. 2024;103:105121. doi:10.1016/j.ebiom.2024.105121
144. Maintz L, Welchowski T, Herrmann N, et al. IL-13, periostin and dipeptidyl-peptidase-4 reveal endotype-phenotype associations in atopic dermatitis. Allergy. 2023;78(6):1554–1569. doi:10.1111/all.15647
145. Kido-Nakahara M, Onozuka D, Izuhara K, et al. Biomarkers and patient-related factors associated with clinical outcomes in dupilumab-treated atopic dermatitis. J Allergy Clin Immunol Glob. 2024;3:100317. doi:10.1016/j.jacig.2024.100317
146. Wu Y, Gu C, Wang S, et al. Serum biomarker-based endotypes of atopic dermatitis in China and prediction for efficacy of dupilumab. Br J Dermatol. 2023;188:649–660. doi:10.1093/bjd/ljad032
147. Kido-Nakahara M, Onozuka D, Izuhara K, et al. Biomarkers and patient-related factors associated with clinical outcomes in dupilumab-treated atopic dermatitis. J Allergy Clin Immunol Glob. 2024;3:100317. doi:10.1016/j.jacig.2024.100317
148. Wu Y, Gu C, Wang S, et al. Serum biomarker-based endotypes of atopic dermatitis in China and prediction for efficacy of dupilumab. Br J Dermatol. 2023;188:649–660. doi:10.1093/bjd/ljad032
149. Kataoka Y. Thymus and activation-regulated chemokine (CCL17) as a clinical biomarker in atopic dermatitis: significance and limitations in the new treatment era. Front Allergy. 2024;5:1473902. doi:10.3389/falgy.2024.1473902
150. Izuhara K, Nunomura S, Nakahara T, Onozuka D. The search for blood biomarkers useful in treating atopic dermatitis patients. Expert Rev Clin Pharmacol. 2024;17:1163–1172. doi:10.1080/17512433.2024.2438192
151. Callaway Z, Kim C-K. Eosinophil-derived neurotoxin levels can predict allergic disease development and atopic march in children. Clin Exp Pediatr. 2025;68:398–405. doi:10.3345/cep.2024.01382
152. El-Korashi LA, Nafea OE, Nafea AE, Elkholy BM, Elhawy LL, Abdelhadi AA. MicroRNA-155 is a potential predictive tool for atopic dermatitis severity in children: a preliminary study. Egypt J Immunol. 2024;31:131–139. doi:10.55133/eji.310313
153. Weidinger S, Blauvelt A, Papp KA, et al. Phase 2b randomized clinical trial of amlitelimab, an anti-OX40 ligand antibody, in patients with moderate-to-severe atopic dermatitis. J Allergy Clin Immunol. 2025;155:1264–1275. doi:10.1016/j.jaci.2024.10.031
154. Sikorska-Szaflik H, Dębińska A, Połomska J, Drabik-Chamerska A, Sozańska B. The role of TSLP and IL-1 β and their genetic variants in the pathogenesis of single and multiple atopic diseases in children. J Clin Med. 2025;14:598. doi:10.3390/jcm14020598
155. Scala E, Madonna S, Abeni D, et al. A microarray-based IgE-molecular mimicry index (IgE-MMI): a biomarker for disease severity, clinical phenotypes, and therapeutic response in atopic dermatitis? Allergy. 2024;79:3415–3429. doi:10.1111/all.16377
156. Lyubchenko T, Collins HK, Goleva E, Leung DYM. Skin tape sampling technique identifies proinflammatory cytokines in atopic dermatitis skin. Ann Allergy Asthma Immunol. 2021;126:46–53.e2. doi:10.1016/j.anai.2020.08.397
157. Abdel-Mageed HM. Atopic dermatitis: a comprehensive updated review of this intriguing disease with futuristic insights. Inflammopharmacology. 2025;33:1161–1187. doi:10.1007/s10787-025-01642-z
158. Hammad H, Lambrecht BN. Barrier epithelial cells and the control of type 2 immunity. Immunity. 2015;43:29–40. doi:10.1016/j.immuni.2015.07.007
159. Jiang H, Gong B, Yan Z, Wang P, Hong J. Identification of novel biomarkers associated with immune infiltration in major depression disorder and atopic dermatitis. Arch Dermatol Res. 2025;317:417. doi:10.1007/s00403-025-03907-7
160. Hartmann D, Retamal C, Valenzuela F. Precision medicine and treat-to-target approach in atopic dermatitis: enhancing personalized care and outcomes. An Bras Dermatol. 2025;100:501135. doi:10.1016/j.abd.2025.501135
161. Mandlik DS, Mandlik SK. Atopic dermatitis: new insight into the etiology, pathogenesis, diagnosis and novel treatment strategies. Immunopharmacol Immunotoxicol. 2021;43:105–125. doi:10.1080/08923973.2021.1889583
162. Mitamura Y, Reiger M, Kim J, et al. Spatial transcriptomics combined with single-cell RNA-sequencing unravels the complex inflammatory cell network in atopic dermatitis. Allergy. 2023;78:2215–2231. doi:10.1111/all.15781
163. Lo Y, Cheng -T-T, Huang C-J, Cheng Y-C, Chyuan I-T. Advancing therapeutic strategies in atopic dermatitis: emerging targets and personalized approaches. Biomolecules. 2025;15:838. doi:10.3390/biom15060838
164. Hodeib A, El-Samad ZA, Hanafy H, El-Latief AA, El-bendary A, Abu-Raya A. Nerve growth factor, neuropeptides and cutaneous nerves in atopic dermatitis. Indian J Dermatol. 2010;55:135–139. doi:10.4103/0019-5154.62735
165. Ozceker D, Bulut M, Ozbay AC, et al. Assessment of IL-31 levels and disease severity in children with atopic dermatitis. Allergologia et Immunopathologia. 2018;46:322–325. doi:10.1016/j.aller.2017.10.005
166. Tobin D, Nabarro G, de la Faute HB, van Vloten WA, van der Putte SCJ, Schuurman H-J. Increased number of immunoreactive nerve fibers in atopic dermatitis. J Allergy Clin Immunol. 1992;90:613–622. doi:10.1016/0091-6749(92)90134-N
167. Mitamura Y, Reiger M, Kim J, et al. Spatial transcriptomics combined with single-cell RNA-sequencing unravels the complex inflammatory cell network in atopic dermatitis. Allergy. 2023;78:2215–2231. doi:10.1111/all.15781
168. Lee I-S, Yoon D-E, Lee S, et al. Neural biomarkers for identifying atopic dermatitis and assessing acupuncture treatment response using resting-state fMRI. J Asthma Allergy. 2024;17:383–389. doi:10.2147/JAA.S454807
169. Kim T, Li J, Tao L, Tao J, Wei X. Neuroimaging characteristics of pruritus induced by eczema: an fMRI study. Brain Behav. 2025;15:e70415. doi:10.1002/brb3.70415
170. Nitulescu G, Olaru OT, Andrei C, Nitulescu GM, Zanfirescu A. Targeting intracellular pathways in atopic dermatitis with small molecule therapeutics. Curr Issues Mol Biol. 2025;47:659. doi:10.3390/cimb47080659
171. Łacwik J, Kraik K, Laska J, Tota M, Sędek Ł, Gomułka K. IL-31/33 axis in atopic dermatitis. Int J Mol Sci. 2025;26:10162. doi:10.3390/ijms262010162
172. Yuan M, Lee J, Taylor M, Cho RJ, Cheng JB. Advancing precision medicine in inflammatory skin disease. Am J Clin Dermatol. 2025;26:853–861. doi:10.1007/s40257-025-00963-7
173. Ishiuji Y, Coghill RC, Patel TS, Oshiro Y, Kraft RA, Yosipovitch G. Distinct patterns of brain activity evoked by histamine-induced itch reveal an association with itch intensity and disease severity in atopic dermatitis. Br J Dermatol. 2009;161:1072–1080. doi:10.1111/j.1365-2133.2009.09308.x
174. Berdyshev E, Kim J, Kim BE, et al. Stratum corneum lipid and cytokine biomarkers at age 2 months predict the future onset of atopic dermatitis. J Allergy Clin Immunol. 2023;151:1307–1316. doi:10.1016/j.jaci.2023.02.013
175. Yu L, Li L. Potential biomarkers of atopic dermatitis. Front Med. 2022;9:1028694. doi:10.3389/fmed.2022.1028694
176. Cevikbas F, Wang X, Akiyama T, et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: involvement of TRPV1 and TRPA1. J Allergy Clin Immunol. 2014;133:448–460. doi:10.1016/j.jaci.2013.10.048
177. Hashimoto T, Yokozeki H, Karasuyama H, Satoh T. IL-31-generating network in atopic dermatitis comprising macrophages, basophils, thymic stromal lymphopoietin, and periostin. J Allergy Clin Immunol. 2023;151:737–746.e6. doi:10.1016/j.jaci.2022.11.009
178. Fassett MS, Braz JM, Castellanos CA, et al. IL-31-dependent neurogenic inflammation restrains cutaneous type 2 immune response in allergic dermatitis. Sci Immunol. 2023;8:eabi6887. doi:10.1126/sciimmunol.abi6887
179. Shim W-S, Tak M-H, Lee M-H, et al. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J Neurosci. 2007;27:2331–2337. doi:10.1523/JNEUROSCI.4643-06.2007
180. Bonchak JG, Swerlick RA. Emerging therapies for atopic dermatitis: TRPV1 antagonists. J Am Acad Dermatol. 2018;78:S63–S66. doi:10.1016/j.jaad.2017.12.023
181. Tamari M, Del Bel KL, Ver Heul AM, et al. Sensory neurons promote immune homeostasis in the lung. Cell. 2024;187:44–61.e17. doi:10.1016/j.cell.2023.11.027
182. Guttman-Yassky E, Croft M, Geng B, et al. The role of OX40 ligand/OX40 axis signalling in atopic dermatitis. Br J Dermatol. 2024;191:488–496. doi:10.1093/bjd/ljae230
183. Abdelhalim A, Yilmaz O, Elshaikh Berair M, Torres T. A narrative review of the OX40-OX40L pathway as a potential therapeutic target in atopic dermatitis: focus on rocatinlimab and amlitelimab. Dermatol Ther. 2024;14:3197–3210. doi:10.1007/s13555-024-01308-8
184. Hui-Beckman JW, Goleva E, Leung DYM, Kim BE. The impact of temperature on the skin barrier and atopic dermatitis. Ann Allergy Asthma Immunol. 2023;131:713–719. doi:10.1016/j.anai.2023.08.007
185. Hu J, Zhao Q, Che D, et al. Epidermal mechanical scratching-induced ROS exacerbates the itch-scratch cycle through TRPA1 activation on mast cells in atopic dermatitis. J Invest Dermatol. 2025;145:2034–2048.e7. doi:10.1016/j.jid.2024.12.026
186. Meixiong J, Anderson M, Limjunyawong N, et al. Activation of mast-cell-expressed mas-related G-protein-coupled receptors drives non-histaminergic itch. Immunity. 2019;50:1163–1171.e5. doi:10.1016/j.immuni.2019.03.013
187. Hartzell CJ, Szabova A, Collins AB, Morel AN, Mijatovic D, Goldschneider KR. Efficacy of aprepitant for refractory pruritus in patients with epidermolysis bullosa and atopic dermatitis: a retrospective study. Pediatr Dermatol. 2024;41:819–821. doi:10.1111/pde.15704
188. Deng J, Parthasarathy V, Marani M, et al. Extracellular matrix and dermal nerve growth factor dysregulation in prurigo nodularis compared to atopic dermatitis. Front Med. 2022;9:1022889. doi:10.3389/fmed.2022.1022889
189. Olivry T, Mayhew D, Paps JS, et al. Early activation of Th2/Th22 inflammatory and pruritogenic pathways in acute canine atopic dermatitis skin lesions. J Invest Dermatol. 2016;136:1961–1969. doi:10.1016/j.jid.2016.05.117
190. Nunomura S, Uta D, Kitajima I, et al. Periostin activates distinct modules of inflammation and itching downstream of the type 2 inflammation pathway. Cell Rep. 2023;42:111933. doi:10.1016/j.celrep.2022.111933
191. Mitamura Y, Reiger M, Kim J, et al. Spatial transcriptomics combined with single-cell RNA-sequencing unravels the complex inflammatory cell network in atopic dermatitis. Allergy. 2023;78:2215–2231. doi:10.1111/all.15781
192. Hui-Beckman JW, Goleva E, Leung DYM, Kim BE. The impact of temperature on the skin barrier and atopic dermatitis. Ann Allergy Asthma Immunol. 2023;131:713–719. doi:10.1016/j.anai.2023.08.007
193. Kabashima K, Matsumura T, Komazaki H, Kawashima M, Nemolizumab-JP01 Study Group. Trial of nemolizumab and topical agents for atopic dermatitis with pruritus. N Engl J Med. 2020;383:141–150. doi: 10.1056/NEJMoa1917006
194. Huang I-H, Chung W-H, Wu P-C, Chen C-B. JAK-STAT signaling pathway in the pathogenesis of atopic dermatitis: an updated review. Front Immunol. 2022;13:1068260. doi:10.3389/fimmu.2022.1068260
195. Yun J-W, Seo JA, Jeong YS, et al. TRPV1 antagonist can suppress the atopic dermatitis-like symptoms by accelerating skin barrier recovery. J Dermatol Sci. 2011;62:8–15. doi:10.1016/j.jdermsci.2010.10.014
196. Blauvelt A, de Bruin-Weller M, Gooderham M, et al. Long-term management of moderate-to-severe atopic dermatitis with dupilumab and concomitant topical corticosteroids (LIBERTY AD CHRONOS): a 1-year, randomised, double-blinded, placebo-controlled, Phase 3 trial. Lancet. 2017;389:2287–2303. doi:10.1016/S0140-6736(17)31191-1
197. Silverberg JI, Wollenberg A, Reich A, et al. Nemolizumab with concomitant topical therapy in adolescents and adults with moderate-to-severe atopic dermatitis (ARCADIA 1 and ARCADIA 2): results from two replicate, double-blind, randomised controlled phase 3 trials. Lancet. 2024;404:445–460. doi:10.1016/S0140-6736(24)01203-0
198. Le M, Berman-Rosa M, Ghazawi FM, et al. Systematic review on the efficacy and safety of oral janus kinase inhibitors for the treatment of atopic dermatitis. Front Med. 2021;8. doi:10.3389/fmed.2021.682547
199. Shih P-Y, Li C-J, Yong S-B. Emerging trends in clinical research on janus kinase inhibitors for atopic dermatitis treatment. Int Immunopharmacol. 2023;124:111029. doi:10.1016/j.intimp.2023.111029
200. Yosipovitch G, Ständer S, Kerby MB, et al. Serlopitant for the treatment of chronic pruritus: results of a randomized, multicenter, placebo-controlled phase 2 clinical trial. J Am Acad Dermatol. 2018;78:882–891.e10. doi:10.1016/j.jaad.2018.02.030
201. Park CW, Kim BJ, Lee YW, et al. Asivatrep, a TRPV1 antagonist, for the topical treatment of atopic dermatitis: phase 3, randomized, vehicle-controlled study (CAPTAIN-AD). J Allergy Clin Immunol. 2022;149:1340–1347.e4. doi:10.1016/j.jaci.2021.09.024
202. Two phase 3 trials of dupilumab versus placebo in atopic dermatitis. N Engl J Med. 2016;375:2235–48.doi:10.1056/NEJMoa1610020
203. Tyring SK, Rich P, Tada Y, et al. Risankizumab in patients with moderate-to-severe atopic dermatitis: a phase 2, randomized, double-blind, placebo-controlled study. Dermatol Ther. 2023;13:595–608. doi:10.1007/s13555-022-00876-x
204. Reichert J. Tralokinumab granted first approval in the European Union. The Antibody Society 2021. Available from: https://www.antibodysociety.org/antibody-therapeutic/tralokinumab-granted-first-approval-in-the-european-union/.
205. Tralokinumab gets nod for atopic dermatitis in Europe. Medscape. Available from: https://www.medscape.com/viewarticle/949912.
206. Two phase 3 trials of lebrikizumab for moderate-to-severe atopic dermatitis. N Engl J Med. 2023;388:1080–91.doi:10.1056/NEJMoa2206714
207. Sofen H, Bissonnette R, Yosipovitch G, et al. Efficacy and safety of vixarelimab, a human monoclonal oncostatin M receptor β antibody, in moderate-to-severe prurigo nodularis: a randomised, double-blind, placebo-controlled, phase 2a study. EClinicalMedicine. 2023;57:101826. doi:10.1016/j.eclinm.2023.101826
208. Welsh SE, Xiao C, Kaden AR, et al. Neurokinin-1 receptor antagonist tradipitant has mixed effects on itch in atopic dermatitis: results from EPIONE, a randomized clinical trial. J Eur Acad Dermatol Venereol. 2021;35:e338–e340. doi:10.1111/jdv.17090
209. Pojawa-Gołąb M, Jaworecka K, Reich A. NK-1 receptor antagonists and pruritus: review of current literature. Dermatol Ther. 2019;9:391–405. doi:10.1007/s13555-019-0305-2
210. Lee YW, Won C-H, Jung K, et al. Efficacy and safety of PAC-14028 cream - a novel, topical, nonsteroidal, selective TRPV1 antagonist in patients with mild-to-moderate atopic dermatitis: a phase IIb randomized trial. Br J Dermatol. 2019;180:1030–1038. doi:10.1111/bjd.17455
211. Narla S, Silverberg JI. Efficacy and risk stratification of janus kinase inhibitors in the treatment of moderate-to-severe atopic dermatitis. Dermatitis. 2024;35:S24–S38. doi:10.1089/derm.2023.0058
212. Torrelo A, Rewerska B, Galimberti M, et al. Efficacy and safety of baricitinib in combination with topical corticosteroids in paediatric patients with moderate-to-severe atopic dermatitis with an inadequate response to topical corticosteroids: results from a Phase III, randomized, double-blind, placebo-controlled study (BREEZE-AD PEDS). Br J Dermatol. 2023;189:23–32. doi:10.1093/bjd/ljad096
213. Jin S-P, Lee K, Bang YJ, et al. Mapping the immune cell landscape of severe atopic dermatitis by single-cell RNA-seq. Allergy. 2024;79:1584–1597. doi:10.1111/all.16121
214. Silverberg JI, Wollenberg A, Reich A, et al. Nemolizumab with concomitant topical therapy in adolescents and adults with moderate-to-severe atopic dermatitis (ARCADIA 1 and ARCADIA 2): results from two replicate, double-blind, randomised controlled phase 3 trials. Lancet. 2024;404:445–460. doi:10.1016/S0140-6736(24)01203-0
215. Park CW, Kim BJ, Lee YW, et al. Asivatrep, a TRPV1 antagonist, for the topical treatment of atopic dermatitis: phase 3, randomized, vehicle-controlled study (CAPTAIN-AD). J Allergy Clin Immunol. 2022;149:1340–1347.e4. doi:10.1016/j.jaci.2021.09.024
216. Simpson EL, Eichenfield LF, Alonso-Llamazares J, et al. Roflumilast cream, 0.15%, for atopic dermatitis in adults and children: INTEGUMENT-1 and INTEGUMENT-2 randomized clinical trials. JAMA Dermatol. 2024;160:1161–1170. doi:10.1001/jamadermatol.2024.3121
217. Silverberg JI, Eichenfield LF, Hebert AA, et al. Tapinarof cream 1% once daily: significant efficacy in the treatment of moderate to severe atopic dermatitis in adults and children down to 2 years of age in the pivotal phase 3 ADORING trials. J Am Acad Dermatol. 2024;91:457–465. doi:10.1016/j.jaad.2024.05.023
218. Bissonnette R, Warren RB, Pinter A, et al. Efficacy and safety of delgocitinib cream in adults with moderate to severe chronic hand eczema (DELTA 1 and DELTA 2): results from multicentre, randomised, controlled, double-blind, phase 3 trials. Lancet. 2024;404:461–473. doi:10.1016/S0140-6736(24)01027-4
219. Borgia F, Li Pomi F, Vaccaro M, Alessandrello C, Papa V, Gangemi S. Oxidative stress and phototherapy in atopic dermatitis: mechanisms, role, and future perspectives. Biomolecules. 2022;12:1904. doi:10.3390/biom12121904
220. Väkevä L, Niemelä S, Lauha M, et al. Narrowband ultraviolet B phototherapy improves quality of life of psoriasis and atopic dermatitis patients up to 3 months: results from an observational multicenter study. Photodermatol Photoimmunol Photomed. 2019;35:332–338. doi:10.1111/phpp.12479
221. Dayal S, Pathak K, Sahu P, Jain VK. Narrowband UV-B phototherapy in childhood atopic dermatitis: efficacy and safety. An Bras Dermatol. 2017;92:801–806. doi:10.1590/abd1806-4841.20175958
222. Choi JY, Owusu-Ayim M, Dawe R, Ibbotson S, Fleming C, Foerster J. Narrowband ultraviolet B phototherapy is associated with a reduction in topical corticosteroid and clinical improvement in atopic dermatitis: a historical inception cohort study. Clin Exp Dermatol. 2021;46:1067–1074. doi:10.1111/ced.14676
223. Musters AH, Mashayekhi S, Harvey J, et al. Phototherapy for atopic eczema. Cochrane Database Syst Rev. 2021;10:CD013870. doi:10.1002/14651858.CD013870.pub2
224. Biliński K, Rakoczy K, Karwowska A, et al. Anti-inflammatory therapies for atopic dermatitis: a new era in targeted treatment. J Clin Med. 2025;14:5053. doi:10.3390/jcm14145053
225. Guttman-Yassky E, Simpson E, Bissonnette R, et al. ROCKET: a phase 3 program evaluating the efficacy and safety of rocatinlimab in moderate-to-severe atopic dermatitis. Immunotherapy. 2025;17:83–94. doi:10.1080/1750743X.2025.2464528
226. Weidinger S, Blauvelt A, Papp KA, et al. Phase 2b randomized clinical trial of amlitelimab, an anti-OX40 ligand antibody, in patients with moderate-to-severe atopic dermatitis. J Allergy Clin Immunol. 2025;155:1264–1275. doi:10.1016/j.jaci.2024.10.031
227. Park CW, Kim BJ, Lee YW, et al. Asivatrep, a TRPV1 antagonist, for the topical treatment of atopic dermatitis: phase 3, randomized, vehicle-controlled study (CAPTAIN-AD). J Allergy Clin Immunol. 2022;149:1340–1347.e4. doi:10.1016/j.jaci.2021.09.024
228. Hu J, Zhao Q, Che D, et al. Epidermal mechanical scratching-induced ROS exacerbates the itch-scratch cycle through TRPA1 activation on mast cells in atopic dermatitis. J Invest Dermatol. 2025;145:2034–2048.e7. doi:10.1016/j.jid.2024.12.026
229. Wong TK, Choi YG, Li PH, Chow BKC, Kumar M. MRGPRX2 antagonist GE1111 attenuated DNFB-induced atopic dermatitis in mice by reducing inflammatory cytokines and restoring skin integrity. Front Immunol. 2024;15:1406438. doi:10.3389/fimmu.2024.1406438
230. Sofen H, Bissonnette R, Yosipovitch G, et al. Efficacy and safety of vixarelimab, a human monoclonal oncostatin M receptor β antibody, in moderate-to-severe prurigo nodularis: a randomised, double-blind, placebo-controlled, phase 2a study. EClinicalMedicine. 2023;57:101826. doi:10.1016/j.eclinm.2023.101826
231. Hartmann D, Retamal C, Valenzuela F. Precision medicine and treat-to-target approach in atopic dermatitis: enhancing personalized care and outcomes. An Bras Dermatol. 2025;100:501135. doi:10.1016/j.abd.2025.501135
232. Hawkins K, David E, Glickman JW, Del Duca E, Guttman-Yassky E, Krueger JG. Atopic dermatitis stratification: current and future perspective on skin and blood transcriptomic and proteomic profiling. Expert Rev Clin Immunol. 2024;20:1083–1088. doi:10.1080/1744666X.2024.2323964
233. Carrascosa-Carrillo JM, Aterido A, Li T, et al. Toward precision medicine in atopic dermatitis using molecular-based approaches. Actas Dermosifiliogr. 2024;115:66–75. doi:10.1016/j.ad.2023.08.003
234. Gallagher K, Halperin-Goldstein S, Paller AS. New treatments in atopic dermatitis update. Ann Allergy Asthma Immunol. 2025;S1081-1206(25):00312. doi:10.1016/j.anai.2025.06.020
235. Gatmaitan JG, Lee JH. Challenges and future trends in atopic dermatitis. Int J Mol Sci. 2023;24:11380. doi:10.3390/ijms241411380
236. Zhou Z, Yang J, Liu Q, Gao J, Ji W. Patho-immunological mechanisms of atopic dermatitis: the role of the three major human microbiomes. Scand J Immunol. 2024;100:e13403. doi:10.1111/sji.13403
237. Hu T, Todberg T, Ewald DA, et al. Assessment of spatial and temporal variation in the skin transcriptome of atopic dermatitis by use of 1.5 mm minipunch biopsies. J Invest Dermatol. 2023;143:612–620.e6. doi:10.1016/j.jid.2022.10.004
238. Kobyletzki von LV, Ballardini N, Henrohn D, et al. Care pathways in atopic dermatitis: a retrospective population‐based cohort study. J Eur Acad Dermatol Venereol. 2022;36:1456–1466. doi:10.1111/jdv.18185
239. Eichenfield LF, Stripling S, Fung S, Cha A, O’Brien A, Schachner LA. recent developments and advances in atopic dermatitis: a focus on epidemiology, pathophysiology, and treatment in the pediatric setting. Paediatr Drugs. 2022;24:293–305. doi:10.1007/s40272-022-00499-x
240. Jia T, Che D, Zheng Y, et al. Mast cells initiate type 2 inflammation via tryptase released by MRGPRX2/MrgprB2 activation in atopic dermatitis. J Invest Dermatol. 2023;144:53–62.e2. doi:10.1016/j.jid.2023.06.201
241. Long-term safety of dupilumab in adults with moderate to severe atopic dermatitis. Available from: https://www.dermatologytimes.com/view/long-term-safety-of-dupilumab-in-adults-with-moderate-to-severe-atopic-dermatitis.
242. Yamamura Y, Nakashima C, Otsuka A. Interplay of cytokines in the pathophysiology of atopic dermatitis: insights from Murin models and human. Front Med. 2024;11. doi: 10.3389/fmed.2024.1342176
243. Maintz L, Welchowski T, Herrmann N, et al. IL‐13, periostin and dipeptidyl‐peptidase‐4 reveal endotype‐phenotype associations in atopic dermatitis. Allergy. 2023;78:1554–1569. doi:10.1111/all.15647
244. Ahn J, Kim S-E, Kim D-Y, et al. Cav3.2 T-type calcium channel mediates acute itch and contributes to chronic itch and inflammation in experimental atopic dermatitis. J Invest Dermatol. 2023;144:612–620.e6. doi:10.1016/j.jid.2023.07.029
245. Dai X, Muto J, Shiraishi K, et al. TSLP impairs epidermal barrier integrity by stimulating the formation of nuclear IL-33/phosphorylated STAT3 complex in human keratinocytes. J Invest Dermatol. 2022;142:2100–2108.e5. doi:10.1016/j.jid.2022.01.005
246. Li H, Liang J, Li P, et al. Schizonepeta tenuifolia Briq-Saposhnikovia divaricata decoction alleviates atopic dermatitis via downregulating macrophage TRPV1. Front Pharmacol. 2024;15. doi: 10.3389/fphar.2024.1413513
247. Biliński K, Rakoczy K, Karwowska A, et al. Anti-inflammatory therapies for atopic dermatitis: a new era in targeted treatment. J Clin Med. 2025;14:5053. doi:10.3390/jcm14145053
248. Li H, Zhang Z, Zhang H, Guo Y, Yao Z. Update on the pathogenesis and therapy of atopic dermatitis. Clin Rev Allergy Immunol. 2021;61:324–338. doi:10.1007/s12016-021-08880-3
249. Escalante A, Serra-Baldrich E. Pathogenic mechanisms underlying itch in atopic dermatitis: the emerging role of neuroimmune interactions. Eur J Dermatol. 2023;33:343–349. doi:10.1684/ejd.2023.4514
250. Moniaga CS, Tominaga M, Takamori K. An altered skin and gut microbiota are involved in the modulation of itch in atopic dermatitis. cells. 2022;11:3930. doi:10.3390/cells11233930
251. Mohammad S, Karim MR, Iqbal S, et al. Atopic dermatitis: pathophysiology, microbiota, and metabolome - A comprehensive review. Microbiol Res. 2024;281:127595. doi:10.1016/j.micres.2023.127595
252. Zhang Z, Tan R, Xiong Z, Feng Y, Chen L. Dysregulation of autophagy during photoaging reduce oxidative stress and inflammatory damage caused by UV. Front Pharmacol. 2025;16:1562845. doi:10.3389/fphar.2025.1562845
253. Microbiota, mind, and skin: bridging the Gut-Brain-Skin axis in psychodermatology. Archives of Dermatological Res. 2025;317:875.doi:10.1007/s00403-025-04385-7
254. Liu X, Qin Y, Dong L, et al. Living symbiotic bacteria-involved skin dressing to combat indigenous pathogens for microbiome-based biotherapy toward atopic dermatitis. Bioact Mater. 2022;21:253–266. doi:10.1016/j.bioactmat.2022.08.019
255. Çetinarslan T, Kümper L, Fölster-Holst R. The immunological and structural epidermal barrier dysfunction and skin microbiome in atopic dermatitis-an update. Front Mol Biosci. 2023;10. doi: 10.3389/fmolb.2023.1159404
256. Hammond M, Gamal A, Mukherjee P, et al. Cutaneous dysbiosis may amplify barrier dysfunction in patients with atopic dermatitis. Front Microbiol. 2022;13. doi: 10.3389/fmicb.2022.944365
257. Kim GI, Jeong HY, Kim I, et al. Interconnection of the gut-skin axis in NC/Nga mouse with atopic dermatitis: effects of the three types of bifidobacterium bifidum CBT-BF3 (probiotics, postbiotics, and cytosine-phosphate-guanine oligodeoxynucleotide) on T cell differentiation and gut microbiota. Food Sci Anim Resour. 2024;44:1417–1439. doi:10.5851/kosfa.2024.e100
258. Demessant-Flavigny A-L, Connétable S, Kerob D, Moreau M, Aguilar L, Wollenberg A. Skin microbiome dysbiosis and the role of Staphylococcus aureus in atopic dermatitis in adults and children: a narrative review. J Eur Acad Dermatol Venereol. 2023;37(Suppl 5):3–17. doi:10.1111/jdv.19125
259. Au C-Y, Manazir N, Kang H, Saleem Bhagat AA. Development of objective measurements of scratching as a proxy of atopic dermatitis—a review. Sensors. 2025;25:4316. doi:10.3390/s25144316
260. Yasuda K-I, Ishiuji Y, Ebata T. Monitoring sleep and scratch improves quality of life in patients with atopic dermatitis. Acta Derm Venereol. 2023;103:11922. doi:10.2340/actadv.v103.11922
261. Kamata M, Sun DI, Paller AS. Deciding which patients with atopic dermatitis to prioritize for biologics and janus kinase inhibitors. J Allergy Clin Immunol Pract. 2025;13:1901–1910.e1. doi:10.1016/j.jaip.2025.04.042
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Awareness and Predictors of the Use of Bioinformatics in Genome Research in Saudi Arabia
Alomair L, Abolfotouh MA
International Journal of General Medicine 2023, 16:3413-3425
Published Date: 11 August 2023
Skin Barrier Repair and Nursing Care in Patients with Atopic Dermatitis: A Narrative Review
Chen X
International Journal of General Medicine 2025, 18:6803-6819
Published Date: 7 November 2025
The Immune Architecture of Eosinophilic Esophagitis: Mechanisms, Therapeutic Targets, and Precision Management
Bertin L, Caldart F, Barchi A, Seregni N, Pasta A, Calabrese F, Marabotto E, Mari A, Farah A, Sirinic E, Sorge A, Ghisa M, Chahuán J, Savarino V, Savarino EV
ImmunoTargets and Therapy 2026, 15:510865
Published Date: 27 May 2026