Back to Journals » International Journal of General Medicine » Volume 19
The Gut-Disc Axis: Mendelian Randomization Study Reveals Causal Effects of Gut Microbiota on Intervertebral Disc Degeneration Not Mediated by Systemic Inflammation
Authors Yan Z, Jiang Q, Lu Z, Cao S ![]() , Bai J, Ni Z, Ding Y
, Bai J, Ni Z, Ding Y
Received 14 April 2026
Accepted for publication 3 June 2026
Published 15 June 2026 Volume 2026:19 615488
DOI https://doi.org/10.2147/IJGM.S615488
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ke Huang
Ziqiao Yan,1,* Qiang Jiang,1,* Zhengcao Lu,1,* Shiqi Cao,1 Jie Bai,1 Zhexin Ni,2 Yu Ding1
1Department of Orthopaedics, Senior Department of Traditional Chinese Medicine, The Sixth Medical Center of PLA General Hospital, Beijing, People’s Republic of China; 2Beijing Institute of Radiation Medicine, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhexin Ni, Email [email protected] Yu Ding, Email [email protected]
Background: Intervertebral disc degeneration (IVDD) is a major cause of low back pain. Observational studies suggest a link with gut microbiota (GM), possibly mediated by systemic inflammation, but causality remains unclear.
Objective: To assess the suggestive associations of GM and inflammatory cytokines (ICs) on IVDD using Mendelian randomization (MR), and evaluate potential mediation by ICs.
Methods: We performed a two-sample MR analysis. Genetic instruments for GM came from MiBioGen (n=18,340), for 41 ICs from a meta-analysis (n=8337), and for IVDD from FinnGen (29,508 cases/227,388 controls). Inverse-variance weighted (IVW) was the primary method. Mediation was assessed via two-step MR, with sensitivity analyses (MR-Egger, MR-PRESSO).
Results: At the nominal significance level (P< 0.05), 11 microbial taxa showed suggestive causal associations with IVDD risk, of which only Eubacterium coprostanoligenes group approached the Bonferroni-corrected threshold (P=3.0× 10− 5). Seven inflammatory cytokines showed suggestive associations with IVDD; however, none remained statistically statistically significant after Bonferroni correction. Mediation analysis revealed no statistically statistically significant indirect effect of cytokines linking gut microbiota to IVDD.
Conclusion: This study provides genetic evidence supporting a potential gut-disc axis. The lack of evidence for cytokine mediation does not exclude alternative pathways but indicates insufficient support for systemic cytokine-mediated mechanisms among those examined; future mechanistic studies should explore microbial metabolites and local immune activity.
Keywords: intervertebral disc degeneration, gut microbiota, inflammatory cytokines, Mendelian randomization, mediation analysis, causality
Introduction
Intervertebral disc degeneration (IVDD) is a predominant worldwide cause of low back pain and disability, conferring a substantial socioeconomic burden on healthcare systems.1 While aging, mechanical stress, and genetic predisposition are well-established risk factors,2 emerging evidence suggests the gut microbiota (GM) as a potential contributor to IVDD pathogenesis.3 Observational studies have linked dysbiosis of the GM to an increased risk of IVDD, potentially through mechanisms involving systemic inflammation, metabolic dysfunction, or immune system modulation.4 Nonetheless, inherent limitations of observational designs—such as residual confounding and reverse causality—preclude definitive conclusions regarding a causal relationship between GM and IVDD.5
Concurrently, the role of inflammatory cytokines (ICs) in IVDD progression has gained increasing attention.6 Elevated levels of specific ICs are a hallmark of degenerated disc tissue, where they are believed to promote extracellular matrix degradation, neurovascular ingrowth, and pain sensitization.7 Given that GM composition is a known regulator of systemic host immune responses and circulating IC levels,8 it has been hypothesized that ICs may act as key mediators in the putative gut-disc axis.9 For instance, certain bacterial taxa can influence the production of prototypical cytokines and chemokines, which could subsequently disrupt the homeostasis of the disc microenvironment.10
To address these hypotheses and overcome the limitations of observational epidemiology, we employed a two-sample Mendelian randomization (MR) framework.11 This method utilizes genetic variants as instrumental variables to infer causality while minimizing confounding and bias from reverse causation. Leveraging summary-statistics from large-scale genome-wide association studies (GWAS) of GM, ICs, and IVDD, we aimed to: (i) assess the causal effect of GM on IVDD risk; (ii) evaluate the causal influence of ICs on IVDD; and (iii) investigate the mediating role of ICs in the pathway from GM to IVDD.
Although a limited number of recent MR studies have explored associations between gut microbiota and IVDD, none have systematically incorporated a comprehensive panel of circulating inflammatory cytokines as candidate mediators within a unified two-step MR framework. Whether systemic inflammation constitutes the principal biological bridge linking gut dysbiosis to IVDD remains unresolved. Our study fills this gap by simultaneously evaluating 211 microbial taxa, 41 inflammatory cytokines, and IVDD outcomes within a single causal-inference framework. This study provides genetic evidence supporting the gut-disc axis hypothesis.
Methods
Study Design
In brief, our analytical framework consisted of three sequential steps (Figure 1): (Step 1) two-sample MR to assess the total causal effect of each gut microbial taxon on IVDD; (Step 2) two-sample MR to assess the causal effect of each inflammatory cytokine on IVDD; and (Step 3) two-step mediation MR to test whether cytokines mediate the microbiota-IVDD relationship.12
 |
Figure 1 Study Design and Analytical Workflow. A schematic overview of the three-step Mendelian randomization (MR) analysis. Step 1: suggestive associations of GM on intervertebral disc degeneration (IVDD) were assessed. Step 2: suggestive associations of inflammatory cytokines (ICs) on IVDD were assessed. Step 3: A two-step MR mediation analysis was performed to investigate whether the identified ICs mediate the causal pathways from GM to IVDD. |
Data Sources
Genetic instruments for GM were obtained from the MiBioGen consortium GWAS, which included 18,340 individuals and provided summary statistics for 211 microbial taxa (spanning 9 phyla, 16 classes, 20 orders, 35 families, and 131 genera).13 Summary statistics for 41 inflammatory cytokines were sourced from a GWAS meta-analysis comprising 8337 individuals of European ancestry.14 Genetic association estimates for IVDD were obtained from the FinnGen consortium (R8 release)(https://www.finngen.fi/fi), including 29,508 cases and 227,388 controls. IVDD cases were identified using ICD-10 (M51), ICD-9 (722), and ICD-8 (725) codes. GWAS summary statistics were downloaded on 5th Nov 2025.
Selection of Genetic Instruments
The locus-wide significance threshold (P<1×10−5) for GM is widely adopted in microbiome MR studies because few microbial taxa reach genome-wide significance, and overly stringent thresholds would leave most taxa without analyzable instruments.15 For the included taxa, the number of SNPs per exposure ranged from 3 to 21 (mean ≈11), the minimum F-statistic was 19.5, and the mean F-statistic was 34.7, with cumulative R2 between 0.5% and 3.2% per exposure (Table S1), indicating sufficient instrument strength and minimal weak instrument bias.16 Data harmonization was conducted using the harmonise_data function in the TwoSampleMR R package. Effect alleles were aligned between exposure and outcome datasets based on the reference strand. Palindromic SNPs with intermediate allele frequencies (MAF > 0.42) were excluded to prevent strand-related ambiguity, while palindromic SNPs with MAF ≤ 0.42 were retained and aligned using allele frequency information. SNPs that could not be unambiguously harmonized were removed prior to MR analysis. To satisfy the second MR assumption (independence of instruments from confounders), all candidate SNPs were queried in the PhenoScanner V2 database to identify potential associations with known IVDD confounders (eg, body mass index, smoking, type 2 diabetes); SNPs showing genome-wide significant associations (P < 5×10−8) with these traits were excluded prior to downstream analysis.
Mendelian Randomization Analysis
Reverse MR analysis was conducted using IVDD as the exposure and the significant microbial taxa / cytokines as outcomes. SNPs associated with IVDD at genome-wide significance (P<5×10−8) were selected as instruments after clumping (r2<0.001, kb=10,000).17–20 The IVW method was used as the primary analysis; a P>0.05 in the reverse direction was interpreted as evidence against reverse causation.21 Bonferroni correction was applied separately at each analytical level: for the GM–IVDD analysis, the corrected threshold was P < 0.05/211 ≈ 2.37×10−4; for the IC–IVDD analysis, P < 0.05/41 ≈ 1.22×10−3; and for the mediation analysis, P < 0.05/77 ≈ 6.5×10−4. Findings with P < 0.05 but exceeding the corresponding Bonferroni threshold are reported as “suggestive” rather than statistically significant, consistent with established practice in microbiome MR studies where strict correction can be overly conservative given the correlated nature of microbial taxa.
Mediation Analysis
A two-step MR approach was used to quantify mediation effects. First, MR was performed to estimate the effect of GM on potential mediator ICs (Path A). Second, the effect of these ICs on IVDD was estimated (Path B). The 95% confidence intervals for the indirect effects (β_A × β_B) were estimated using the Delta method, which provides analytically derived standard errors via first-order Taylor expansion. The proportion mediated was calculated only when both the total effect and indirect effect were nominally significant; otherwise, the proportion mediated was considered statistically unstable and is not reported as a primary outcome.22
Bidirectional and Sensitivity Analyses
Reverse causation was assessed by testing the effects of genetic liability to IVDD on GM and ICs, using genome-wide statistically significant SNPs (P<5×10−8) as instruments. Sensitivity analyses included: (1) Cochran’s Q test to assess heterogeneity; (2) MR-Egger regression and MR-PRESSO to detect and correct for horizontal pleiotropy; (3) Leave-one-out analysis to evaluate the influence of individual variants. All analyses were conducted in R (v4.2.1) using the TwoSampleMR (v0.5.7) and MR-PRESSO (v1.0) packages.23–26
Ethical Approval
This study was exempt from institutional ethical review. Specifically, this exemption is strictly based on items 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (issued by the National Health Commission of China, dated February 18, 2023).
Result
Instrumental Variable Selection
Initially, we identified 224, 478, 1667, 280, and 125 SNPs associated with 210 GMs at the class, family, genus, order, and phylum levels, respectively, at a level of P < 1 × 10–5 (one gut microbiota was excluded due to no eligible SNPs). These 2774 SNPs were selected as IVs for the 210 GM taxa. Then, we identified 451 SNPs associated with 41 cytokines at a level of P < 5 × 10–6.
Suggestive Causal Associations Between GM and IVDD
Using the IVW method, we identified suggestive causal associations between 11 microbial taxa and IVDD risk at the nominal level (P < 0.05). None of these associations survived Bonferroni correction (P < 2.37×10−4); only the protective association of the Eubacterium coprostanoligenes group (P = 3.0×10−5) approached corrected significance. Therefore, with the exception of Eubacterium coprostanoligenes group, all microbiota–IVDD findings should be interpreted as suggestive rather than definitive (Figures 2 and 3, Table 1). The F-statistic for all SNPs exceeded 10, indicating no substantial weak instrument bias (Table S1).
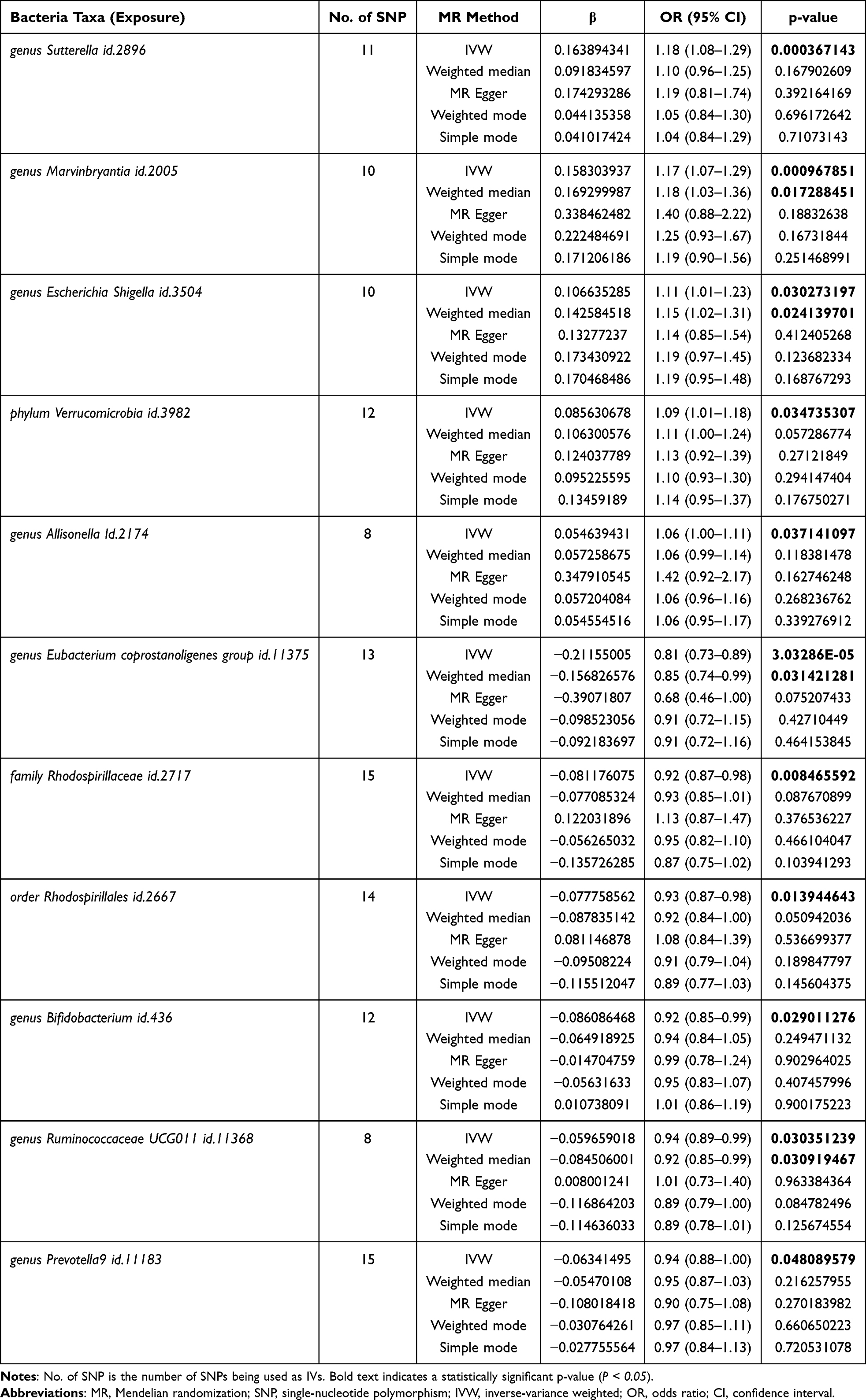 |
Table 1 Mendelian Randomization Analysis of the Causal Effects of Gut Microbiota on Intervertebral Disc Degeneration |
 |
Figure 2 Forest Plot of suggestive associations of GM on IVDD Risk. The suggestive associations of 11 gut microbial taxa statistically significantly associated with IVDD risk, as determined by the inverse-variance weighted (IVW) method, are displayed. Effect sizes are presented as odds ratios (ORs) with 95% confidence intervals (CIs). Taxa are categorized as risk factors (OR > 1) or protective factors (OR < 1). |
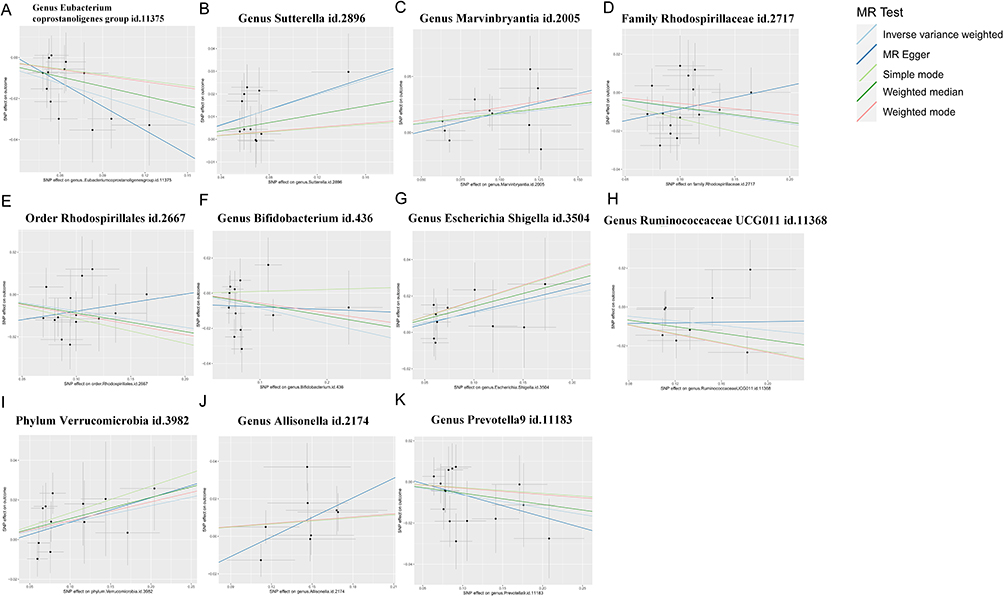 |
Figure 3 Scatter Plots of Genetic Associations for statistically significant GM and IVDD. Each point represents a single-nucleotide polymorphism (SNP). The slopes of the colored regression lines generated by different Mendelian randomization (MR) methods (including inverse-variance weighted, MR-Egger, simple mode, weighted median, and weighted mode) indicate the estimated causal effect. (A) Genus Eubacterium coprostanoligenes group. (B) Genus Sutterella. (C) Genus Marvinbryantia. (D) Family Rhodospirillaceae. (E) Order Rhodospirillales. (F) Genus Bifidobacterium. (G) Genus Escherichia-Shigella. (H) Genus Ruminococcaceae UCG011. (I) Phylum Verrucomicrobia. (J) Genus Allisonella. (K) Genus Prevotella9. |
Five taxa were identified as risk factors for IVDD: genus Sutterella (id.2896, OR = 1.18, 95% CI: 1.08–1.29, P = 0.0004), genus Marvinbryantia (id.2005, OR = 1.17, 95% CI: 1.07–1.29, P = 0.001), genus Escherichia-Shigella (id.3504, OR = 1.11, 95% CI: 1.01–1.23, P = 0.03), phylum Verrucomicrobia (id.3982, OR = 1.09, 95% CI: 1.01–1.18, P = 0.03), and genus Allisonella (id.2174, OR = 1.06, 95% CI: 1.00–1.11, P = 0.04).
Six taxa were identified as protective factors for IVDD: genus Eubacterium coprostanoligenes group (id.11375, OR = 0.81, 95% CI: 0.73–0.89, P = 3.0×10−5), family Rhodospirillaceae (id.2717, OR = 0.92, 95% CI: 0.87–0.98, P = 0.008), order Rhodospirillales (id.2667, OR = 0.93, 95% CI: 0.87–0.98, P = 0.014), genus Bifidobacterium (id.436, OR = 0.92, 95% CI: 0.85–0.99, P = 0.03), genus Ruminococcaceae UCG011 (id.11368, OR = 0.94, 95% CI: 0.89–0.99, P = 0.03), and genus Prevotella9 (id.11183, OR = 0.94, 95% CI: 0.88–1.00, P = 0.048).
Reverse MR analysis using IVDD-associated SNPs as instruments showed no significant reverse effect of IVDD on the identified gut microbial taxa (P>0.05), supporting the directional assumption from microbiota to IVDD rather than the reverse (Table S2), which helps to mitigate concerns about reverse causation.
Sensitivity analyses supported the robustness of the primary findings. No statistically significant heterogeneity was observed among the IVs for the gut microbiome traits (Table S3). We used the MR Pleiotropy RESidual Sum and Outlier (MR-PRESSO) method to assess horizontal pleiotropy, and the results indicated no evidence of pleiotropic bias (Table S4). Furthermore, leave-one-out analysis confirmed that no single SNP disproportionately drove the observed associations between GM and IVDD (Figure 4).
 |
Figure 4 Leave-One-Out Sensitivity Analysis for GM on IVDD. Each panel shows the inverse-variance weighted (IVW) estimate and 95% confidence interval when iteratively removing one single-nucleotide polymorphism (SNP) at a time, confirming that no single SNP was driving the observed causal associations. (A) Genus Eubacterium coprostanoligenes group. (B) Genus Sutterella. (C) Genus Marvinbryantia. (D) Family Rhodospirillaceae. (E) Order Rhodospirillales. (F) Genus Bifidobacterium. (G) Genus Escherichia-Shigella. (H) Genus Ruminococcaceae UCG011. (I) Phylum Verrucomicrobia. (J) Genus Allisonella. (K) Genus Prevotella9. |
Suggestive Associations of Inflammatory Cytokines on IVDD
We next assessed the suggestive associations of 41 inflammatory cytokines on IVDD. IVW analysis revealed suggestive associations for seven cytokines. Genetically predicted levels of RANTES (OR = 1.07, 95% CI: 1.01–1.12, P = 0.013) and SDF-1α (OR = 1.09, 95% CI: 1.00–1.19, P = 0.042) were associated with increased odds of IVDD. In contrast, genetically predicted levels of IFN-γ (OR = 0.89, 95% CI: 0.83–0.95, P = 0.0007), IL-1β (OR = 0.89, 95% CI: 0.83–0.97, P = 0.005), MIP-1β (OR = 0.97, 95% CI: 0.95–1.00, P = 0.02), MIG (OR = 0.95, 95% CI: 0.91–0.99, P = 0.025), and IL-18 (OR = 0.96, 95% CI: 0.93–1.00, P = 0.036) were associated with decreased odds of IVDD (Figures 5 and 6, Table 2). The F-statistic for all SNPs exceeded 10, indicating no substantial weak instrument bias (Table S5).
 |
Table 2 Mendelian Randomization Analysis of the Suggestive Causal Effects of Inflammatory Cytokines on Intervertebral Disc Degeneration |
 |
Figure 5 Forest Plot of Suggestive associations of Inflammatory Cytokines on IVDD. The suggestive associations of 7 inflammatory cytokines suggestively associated with IVDD risk (P < 0.05, pre-Bonferroni correction), as determined by the IVW method, are displayed. Effect sizes are presented as odds ratios (ORs) with 95% confidence intervals (CIs). |
 |
Figure 6 Scatter Plots of Genetic Associations for Suggestive Inflammatory Cytokines and IVDD. Each point represents a single-nucleotide polymorphism (SNP). The slopes of the colored regression lines generated by different Mendelian randomization (MR) methods illustrate the estimated causal direction. (A) IFN-γ. (B) IL-1β. (C) RANTES. (D) MIP-1β. (E) MIG. (F) IL-18. (G) SDF-1α. |
It should be noted that none of these associations remained statistically statistically significant after Bonferroni correction for multiple testing (corrected significance threshold P < 0.0012). Sensitivity analyses supported the robustness of these findings. No statistically significant heterogeneity was detected among the IVs for cytokine traits (Table S6). The MR-PRESSO test revealed no evidence of horizontal pleiotropy (Table S7). Leave-one-out analysis indicated that no single SNP disproportionately influenced the results (Figure 7). It must be emphasised that none of the cytokine–IVDD associations survived Bonferroni correction. The following findings are therefore exploratory in nature and should not be interpreted as validated suggestive associations.
 |
Figure 7 Leave-One-Out Sensitivity Analysis for Inflammatory Cytokines on IVDD. The plots suggests that the results were not disproportionately influenced by any individual single-nucleotide polymorphism (SNP). (A) IFN-γ. (B) IL-1β. (C) RANTES. (D) MIP-1β. (E) MIG. (F) IL-18. (G) SDF-1α. |
Mediation Analysis
We performed a two-step MR analysis to assess whether the suggestive associations of the 11 identified GM on IVDD were mediated by the 7 inflammatory cytokines suggestively associated with IVDD.
The overall findings from the mediation analysis were null. After applying a Bonferroni correction for the number of tested mediator-exposure-outcome combinations (corrected significance threshold P < [0.05/77] ≈ 0.00065), none of the indirect effects were statistically statistically significant. The 95% confidence intervals for all indirect effect estimates included the null value (Table 3), indicating a lack of robust evidence to support the mediating role of these systemic inflammatory cytokines in the pathways from GM to IVDD.
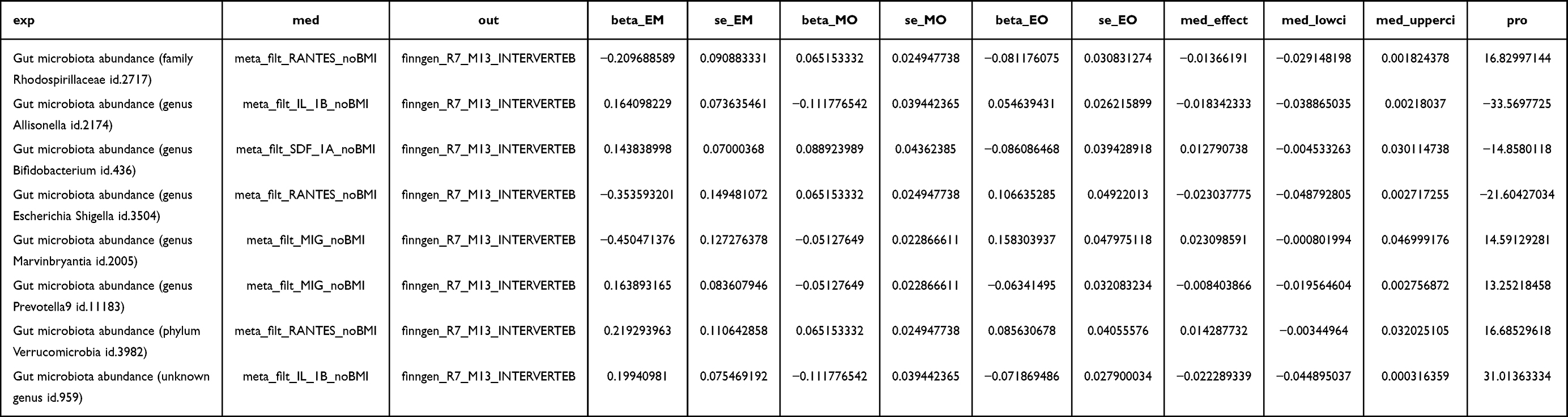 |
Table 3 Two-Step MR Mediation Analysis of Inflammatory Cytokines in the Gut Microbiota-IVDD Pathway |
Although none of the indirect effects reached statistical significance, point estimates and confidence intervals for all tested mediation pathways are presented in Table 3 for transparency. Overall, no robust evidence of mediation by the examined systemic cytokines was identified. The abundance of genus Marvinbryantia was associated with lower levels of MIG (β = −0.451, SE = 0.127), and lower MIG levels were in turn associated with a reduced risk of IVDD (β = −0.051, SE = 0.023). The resulting point estimate for the indirect effect was positive (β_indirect = 0.023), but its 95% confidence interval was wide and included zero (95% CI: −0.0008 to 0.0470), precluding a definitive conclusion.
Similarly, the observed suggestive association between phylum Verrucomicrobia, RANTES, and IVDD did not yield a statistically statistically significant mediation effect. Due to the non-statistically significant primary associations, the estimated proportion mediated for all pathways was highly unstable and should be interpreted with extreme caution.
Discussion
This two-sample Mendelian randomization study represents a comprehensive genetic investigation into the causal relationships between GM, inflammatory cytokines, and intervertebral disc degeneration. Our analysis yielded three principal findings. First, we identified several gut microbial taxa that exert suggestive associations on IVDD risk, providing genetic evidence supporting the hypothesis of a potential “gut-disc” axis.27 Second, we found suggestive evidence that genetically proxied levels of several inflammatory cytokines are associated with IVDD. Third, and most notably, our mediation analysis did not find statistically statistically significant evidence that the systemic inflammatory cytokines we investigated act as mediators between the implicated GM and IVDD after rigorous multiple testing correction.
The identification of specific GM, such as the protective Eubacterium coprostanoligenes group and the risk-associated Sutterella and Marvinbryantia, aligns with a growing body of literature linking GM to musculoskeletal health.28 Eubacterium species are known producers of short-chain fatty acids (SCFAs) such as butyrate, which could be hypothesised to contribute to the anti-inflammatory and anti-catabolic effects observed in various models, potentially partially explaining the protective signal observed here. However, as butyrate and other SCFA levels were not directly measured in this study, this mechanistic interpretation remains speculative and warrants experimental validation in future studies.29 Conversely, Sutterella has been associated with mucosal inflammation and could potentially promote systemic low-grade inflammation.30 Our use of MR strengthens the evidence for these relationships by minimizing the confounding and reverse causation that limit observational studies.
It must be emphasised that none of the cytokine–IVDD associations survived Bonferroni correction; the following biological interpretations are therefore exploratory and should not be regarded as validated causal effects. The suggestive associations between cytokines like RANTES (a risk factor) and IFN-γ (a protective factor) with IVDD are biologically plausible. RANTES (CCL5) is a chemokine that recruits monocytes and T-cells to sites of inflammation and has been implicated in matrix degradation processes relevant to IVDD.31 IFN-γ, typically considered a pro-inflammatory cytokine, may have complex, context-dependent roles, including immunoregulatory functions that could be protective in chronic degenerative settings.32 However, the fact that these associations did not survive Bonferroni correction necessitates caution in their interpretation and highlights the need for replication in larger GWAS.
The most critical finding of our mediation analysis is the lack of statistically significant evidence for cytokine mediation. Despite identifying several suggestive causal paths for both exposures and mediators, the formal two-step MR analysis did not identify any statistically statistically significant indirect effects after multiple testing correction. The confidence intervals for all mediation estimates included the null value. This suggests that the causal effect of these gut microbes on IVDD may operate through alternative pathways. Potential mechanisms not captured by our analysis include: (1) Local immune modulation: The GM may influence the immune cell populations within the disc microenvironment itself without altering systemic cytokine levels.33 (2) Metabolic products: Microbial metabolites (eg, SCFAs, bile acids, tryptophan derivatives) could directly influence disc cell homeostasis, apoptosis, or matrix synthesis.34 (3) Direct translocation: In cases of increased intestinal permeability (“leaky gut”), microbial components or whole bacteria may translocate, potentially eliciting a localized immune response in paravertebral tissues or even the disc itself.35 (4) Other unmeasured mediators: Pathways involving neurotransmitters, oxidative stress, or vascularization were not examined in this study.36
The major strength of this study is the application of MR, which provides a robust framework for assessing causality by leveraging genetic variants as instrumental variables.37 The use of large, publicly available GWAS consortia data ensured substantial statistical power. We also conducted extensive sensitivity analyses to check for pleiotropy and heterogeneity, strengthening the validity of our primary results.
However, several limitations must be acknowledged. First, the statistical power for the mediation analysis was a key constraint. The GWAS for ICs had a relatively smaller sample size (n≈8337) compared to those for GM and IVDD, which likely resulted in weaker genetic instruments and reduced power to detect true mediation effects (Type II error).38 The stringent Bonferroni correction, while conservative, further increased this risk. Second, the GWAS data for GM and ICs were derived from blood and fecal samples, respectively, and may not fully reflect the biological activity at the site of the intervertebral disc.39 Third, despite our efforts to control for horizontal pleiotropy, residual pleiotropy remains a potential caveat in any MR study.40 Fourth, Importantly, all GWAS datasets used in this study were derived from individuals of European ancestry. Therefore, the generalizability of our findings to other ethnic populations (eg, East Asian, African) remains uncertain, and replication in trans-ancestry MR studies is essential before any broader clinical extrapolation.41
Conclusion
In conclusion, our study provides novel genetic evidence that specific GM play a causal role in the development of IVDD, solidifying the concept of a gut-disc axis. However, it indicates no evidence of mediation by the examined systemic inflammatory cytokines was detected, rather than excluding the existence of such mediation. This important negative result shifts the focus of future research towards alternative mechanistic pathways, such as direct microbial metabolite actions, local immune regulation, or other biological processes. Future studies with larger GWAS for cytokines and more precise tissue-specific molecular data are needed to conclusively rule out or identify specific inflammatory mediators. Importantly, our findings do not provide direct evidence for clinical interventions, probiotic therapies, or biomarker applications. Rather, they highlight specific microbial taxa as priority candidates for future mechanistic and translational studies, particularly those focused on microbial metabolites and local immune microenvironments within the intervertebral disc. Ultimately, integrating multi-omics data (metagenomics, metabolomics) from well-phenotyped cohorts will be essential to unravel the precise mechanisms linking the gut microbiome to disc health and disease.
Data Access Statement
All data used in this present study are publicly available summary-level data from previously conducted genome-wide association studies (GWAS). Genetic instruments for gut microbiota were obtained from the MiBioGen consortium (https://mibiogen.gcc.rug.nl/). Genetic associations for inflammatory cytokines were sourced from the GWAS. Summary statistics for intervertebral disc degeneration were derived from FinnGen. Original datasets can be accessed and applied for via the links provided by the respective consortia or publications.
Ethics Statement
The authors confirm that the ethical policies of the journal, as noted on the journal’s author guidelines page, have been adhered to. The data used in this study were publicly available summary-level statistics obtained from large-scale genome-wide association studies (GWAS) consortia. All original studies included in this analysis had been approved by their respective institutional review boards or ethics committees, and informed consent was obtained from all participants in those primary studies. As our research involved secondary analysis of de-identified, publicly available data and did not involve any new collection of human biological samples or individual-level data, additional ethical approval was not required for this specific study. This study was exempt from institutional ethical review. Specifically, this exemption is strictly based on items 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (issued by the National Health Commission of China, dated February 18, 2023). Our research complied with the principles set forth in the Declaration of Helsinki.
Author Contributions
YD and ZN conceived the study. ZY, QJ and ZL analyzed data. SC, JB and ZN drafted the manuscript. QJ, JB, and SC provided guidance and checked the manuscript.
All authors made a statistically significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
References
1. Kuzu Ş, Jawad SR, Canli M, et al. Investigation of the efficacy of high and low intensity laser therapy in patients with lumbar disc herniation: a randomized controlled trial. J Med Biol Eng. 2025;45(5):738–14. doi:10.1007/s40846-025-00989-6
2. Xue Q, Li J, Qin R, et al. Nrf2 activation by pyrroloquinoline quinone inhibits natural aging-related intervertebral disk degeneration in mice. Aging Cell. 2024;23(8):e14202. doi:10.1111/acel.14202
3. Geng Z, Wang J, Chen G, et al. Gut microbiota and intervertebral disc degeneration: a bidirectional two-sample Mendelian randomization study. J Orthop Surg Res. 2023;18(1):601. doi:10.1186/s13018-023-04081-0
4. Ruiz-Fernández C, Francisco V, Pino J, et al. Molecular relationships among obesity, inflammation and intervertebral disc degeneration: are adipokines the common link? Int J Mol Sci. 2019;20(8):2030. doi:10.3390/ijms20082030
5. Liu C, Chu X, Biao Y, et al. Association between lipid-lowering agents with intervertebral disc degeneration, sciatica and low back pain: a drug-targeted mendelian randomized study and cross-sectional observation. Lipids Health Dis. 2024;23(1):327. doi:10.1186/s12944-024-02311-w
6. Chen S, Lei L, Li Z, et al. Grem1 accelerates nucleus pulposus cell apoptosis and intervertebral disc degeneration by inhibiting TGF-β-mediated Smad2/3 phosphorylation. Exp Mol Med. 2022;54:518–530.
7. Zhan J, Cui Y, Zhang P, et al. Cartilage endplate-targeted engineered exosome releasing and acid neutralizing hydrogel reverses intervertebral disc degeneration. Adv Healthc Mater. 2025;14(2):e2403315. doi:10.1002/adhm.202403315
8. Wang D, Zhang L, He D, et al. Systemic pharmacology reveal the mechanism by which the Qiangjin Zhuanggu Qufeng mixture inhibits LPS-induced pyroptosis of rat nucleus pulposus cells. Phytomedicine. 2023;119:154998. doi:10.1016/j.phymed.2023.154998
9. Sun Y, Li Z, Duan J, et al. Unveiling the Gut-Disc axis: how microbiome dysbiosis accelerates intervertebral disc degeneration. J Inflamm Res. 2024;17:8271–8280. doi:10.2147/JIR.S487936
10. Li W, Lai K, Chopra N, Zheng Z, Das A, Diwan AD. Gut-disc axis: a cause of intervertebral disc degeneration and low back pain? Eur Spine J. 2022;31(4):917–925. doi:10.1007/s00586-022-07152-8
11. Ji D, Chen WZ, Zhang L, Zhang ZH, Chen LJ. Gut microbiota, circulating cytokines and dementia: a Mendelian randomization study. J Neuroinflammation. 2024;21(1):2. doi:10.1186/s12974-023-02999-0
12. Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44(2):512–525. doi:10.1093/ije/dyv080
13. Kurilshikov A, Medina-Gomez C, Bacigalupe R, et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet. 2021;53(2):156–165. doi:10.1038/s41588-020-00763-1
14. Ahola-Olli AV, Würtz P, Havulinna AS, et al. Genome-wide association study identifies 27 loci influencing concentrations of circulating cytokines and growth factors. Am J Hum Genet. 2017;100(1):40–50. doi:10.1016/j.ajhg.2016.11.007
15. Myers TA, Chanock SJ, Machiela MJ. LDlinkR: an R package for rapidly calculating linkage disequilibrium statistics in diverse populations. Front Genet. 2020;11:157. doi:10.3389/fgene.2020.00157
16. Pierce BL, Ahsan H, Vanderweele TJ. Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int J Epidemiol. 2011;40(3):740–752. doi:10.1093/ije/dyq151
17. Choi KW, Chen CY, Stein MB, et al. Assessment of bidirectional relationships between physical activity and depression among adults: a 2-sample mendelian randomization study. JAMA Psychiatry. 2019;76(4):399–408. doi:10.1001/jamapsychiatry.2018.4175
18. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40:304–314.
19. Ho J, Mak CCH, Sharma V, To K, Khan W. Mendelian randomization studies of lifestyle-related risk factors for osteoarthritis: a PRISMA review and meta-analysis. Int J Mol Sci. 2022;23(19):11906. doi:10.3390/ijms231911906
20. Hartwig FP, Davey Smith G, Bowden J. Robust inference in summary data mendelian randomization via the zero modal pleiotropy assumption. Int J Epidemiol. 2017;46(6):1985–1998. doi:10.1093/ije/dyx102
21. Jiang P, Yu F, Zhou X, Shi H, He Q, Song X. Dissecting causal links between gut microbiota, inflammatory cytokines, and DLBCL: a Mendelian randomization study. Blood Adv. 2024;8(9):2268–2278. doi:10.1182/bloodadvances.2023012246
22. Caiyun M, Hebao W, Wenhao Y, Changqing L, Changqing L, Xiaojiang Z. Dissecting causal links between gut microbiota, inflammatory cytokines, and Parkinson’s disease: a Mendelian randomization study. Brain Behav. 2024;14(11):e70169. doi:10.1002/brb3.70169
23. Bowden J, Holmes MV. Meta-analysis and Mendelian randomization: a review. Res Synth Methods. 2019;10:486–496.
24. Burgess S, Bowden J, Fall T, Ingelsson E, Thompson SG. Sensitivity analyses for robust causal inference from Mendelian randomization analyses with multiple genetic variants. Epidemiology. 2017;28(1):30–42. doi:10.1097/EDE.0000000000000559
25. Morrison J, Knoblauch N, Marcus JH, Stephens M, He X. Mendelian randomization accounting for correlated and uncorrelated pleiotropic effects using genome-wide summary statistics. Nat Genet. 2020;52(7):740–747. doi:10.1038/s41588-020-0631-4
26. Cao J, Wang N, Luo Y, et al. A cause-effect relationship between Graves’ disease and the gut microbiome contributes to the thyroid-gut axis: a bidirectional two-sample Mendelian randomization study. Front Immunol. 2023;14:977587. doi:10.3389/fimmu.2023.977587
27. Zhang J, Wang B, Du P, Song H, Yang L, Zhou Y. Gut-disc axis: a Mendelian randomization study on the relationship between gut microbiota and cervical spondylosis. Medicine. 2025;104(7):e41536. doi:10.1097/MD.0000000000041536
28. Zhao T, Wei Y, Zhu Y, et al. Gut microbiota and rheumatoid arthritis: from pathogenesis to novel therapeutic opportunities. Front Immunol. 2022;13:1007165.
29. Chen WJ, Chen YT, Ko JL, et al. Butyrate modulates gut microbiota and anti-inflammatory response in attenuating cisplatin-induced kidney injury. Biomed Pharmacother. 2024;181:117689. doi:10.1016/j.biopha.2024.117689
30. Hiippala K, Kainulainen V, Kalliomäki M, Arkkila P, Satokari R. Mucosal prevalence and interactions with the epithelium indicate commensalism of Sutterella spp. Front Microbiol. 2016;7:1706.
31. Zhang Z, Qin F, Feng Y, et al. Icariin regulates stem cell migration for endogenous repair of intervertebral disc degeneration by increasing the expression of chemotactic cytokines. BMC Complement Med Ther. 2022;22(1):63. doi:10.1186/s12906-022-03544-x
32. Zhang C, Li H, Wang H, et al. Identifying myeloid-derived suppressor cells and lipocalin-2 as therapeutic targets for intervertebral disc degeneration. Adv Sci. 2025;12(34):e00505. doi:10.1002/advs.202500505
33. Ge Y, Yang H, Fu Y, et al. A Mendelian randomization study to reveal gut-disc axis: causal associations between gut microbiota with intervertebral disc diseases. Eur Spine J. 2025;34(6):2052–2065. doi:10.1007/s00586-025-08795-z
34. Yao B, Cai Y, Wang W, et al. The effect of gut microbiota on the progression of intervertebral disc degeneration. Orthop Surg. 2023;15:858–867.
35. Twardowska A, Makaro A, Binienda A, Fichna J, Salaga M. Preventing bacterial translocation in patients with leaky gut syndrome: nutrition and pharmacological treatment options. Int J Mol Sci. 2022;23(6):3204. doi:10.3390/ijms23063204
36. Luo H, Wang Z, He Z, et al. Injectable chondroitin sulfate-grafted self-antioxidant hydrogels ameliorate nucleus pulposus degeneration against overactive inflammation. Biomater Sci. 2023;11(10):3629–3644. doi:10.1039/D3BM00359K
37. Sekula P, Del Greco MF, Pattaro C, Köttgen A. Mendelian randomization as an approach to assess causality using observational data. J Am Soc Nephrol. 2016;27(11):3253–3265. doi:10.1681/ASN.2016010098
38. Jiang Y, Chen S, McGuire D, et al. Proper conditional analysis in the presence of missing data: application to large scale meta-analysis of tobacco use phenotypes. PLoS Genet. 2018;14(7):e1007452. doi:10.1371/journal.pgen.1007452
39. Tam V, Patel N, Turcotte M, Bossé Y, Paré G, Meyre D. Benefits and limitations of genome-wide association studies. Nat Rev Genet. 2019;20(8):467–484. doi:10.1038/s41576-019-0127-1
40. Sanderson E. Multivariable Mendelian randomization and mediation. Cold Spring Harb Perspect Med. 2021;11:a038984.
41. Liu C, Zeinomar N, Chung WK, et al. Generalizability of polygenic risk scores for breast cancer among women with European, African, and Latinx ancestry. JAMA Netw Open. 2021;4:e2119084.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Revealing the Impact of Gut Microbiota on Acne Through Mendelian Randomization Analysis

Ji X, Wu S, Zhao D, Bai Q, Wang Y, Gong K, Zheng H, Zhu M
Clinical, Cosmetic and Investigational Dermatology 2024, 17:383-393
Published Date: 8 February 2024
Genomics-Microbiome Based Assessment of Bidirectional Causality Between Gut Microbiota and Psoriasis
Gao Q, Liu JH, Ma WY, Cheng ZL, Hao PS, Luo NN
Clinical, Cosmetic and Investigational Dermatology 2024, 17:435-445
Published Date: 13 February 2024
Exploring the Causal Relationship Between Frailty and Chronic Obstructive Pulmonary Disease: Insights From Bidirectional Mendelian Randomization and Mediation Analysis
Cheng Z, Wu J, Xu C, Yan X
International Journal of Chronic Obstructive Pulmonary Disease 2025, 20:193-205
Published Date: 25 January 2025
The Mediation Role of Insulin Resistance and Chronic Systemic Inflammation in the Association Between Obesity and NAFLD: Two Cross-Sectional and a Mendelian Randomization Study
Huang X, Chen Q, Su Q, Gong J, Wu L, Xiang L, Li W, Chen J, Zhao H, Huang W, Du S, Ye W
Clinical Epidemiology 2025, 17:287-302
Published Date: 24 March 2025
Investigating the Gut Microbiota-Inflammatory Cytokine-Skin Axis in Inflammatory Skin Diseases: Evidence from Mendelian Randomization
Hu Y, Chen J, Zeng Q, Jiang L, Hu Y, Zhu S, Fu C, Lei L
Clinical, Cosmetic and Investigational Dermatology 2026, 19:611710
Published Date: 1 July 2026