Back to Journals » Journal of Inflammation Research » Volume 18
The eNAMPT-Integrin α5β1 Axis Mediates Neutrophil-Endothelial Cell Interactions Driving Inflammation in Ulcerative Colitis
Authors Di Y, Ji W, Xiong W, Song W, Chen G, Li D, Qi F ![]()
Received 19 July 2025
Accepted for publication 8 November 2025
Published 30 December 2025 Volume 2025:18 Pages 18307—18321
DOI https://doi.org/10.2147/JIR.S554975
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Nadia Andrea Andreani
Yongcheng Di,1,2,* Wenbin Ji,1,2,* Wenhao Xiong,1,2,* Wenbin Song,1,2 Guoshan Chen,1,2 Danzhou Li,1,2 Feng Qi1,2
1Department of General Surgery, Tianjin Medical University General Hospital, Tianjin, People’s Republic of China; 2Tianjin Key Laboratory of Precise Vascular Reconstruction and Organ Function Repair, Tianjin, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Feng Qi, Email [email protected]
Background: Ulcerative colitis (UC), a chronic inflammatory bowel disease with rising global incidence, involves neutrophil-driven mucosal damage. The precise mechanisms remain elusive, hindering targeted therapies. Therefore, this study aims to integrate single-cell transcriptomics with in vivo experiments to reveal the key signaling axes driving pathogenic neutrophil activation in UC.
Methods: Single-cell transcriptomics characterized UC inflammatory microenvironments, focusing on neutrophil functional states and intercellular interactions. Based on key findings from bioinformatics analysis, we hypothesize that the eNAMPT-integrin α 5β 1 signaling axis drives abnormal neutrophil-endothelial cell communication and functionally validate this hypothesis in in vivo models.
Results: Neutrophils exhibited aberrant activation and significant NAMPT overexpression in UC. Extracellular eNAMPT functioned as a signaling molecule binding endothelial integrin α 5β 1, mediating pathological neutrophil-endothelial crosstalk. Pharmacological blockade of the eNAMPT/integrin α 5β 1 axis inhibited neutrophil mucosal infiltration, reducing inflammation and tissue damage in UC mouse models.
Conclusion: The eNAMPT-integrin α 5β 1-mediated neutrophil-endothelial communication axis represents a novel pathogenic pathway in UC, providing a foundation for precision therapies targeting this mechanism.
Keywords: ulcerative colitis, neutrophils, endothelial cells, NAMPT, bioinformatics
Introduction
Ulcerative colitis (UC) is a chronic inflammatory disease that affects the rectum and colon. In 2023, the global prevalence of UC was estimated to be 5 million cases, and the incidence rate continues to rise. Patients with ulcerative colitis may experience disease progression toward the proximal colon, eventually developing pancolitis. This is accompanied by structural and functional changes that lead to an impaired quality of life.1 Despite the increasing availability of clinical treatment options, controlling the disease remains challenging due to the complex underlying mechanisms, which are not yet fully understood.2 The prevailing view is that individuals with genetic susceptibility develop ulcerative colitis following environmental exposure. Intestinal epithelial barrier disruption, abnormal immune activation, and dysbiosis of the gut microbiota are the primary drivers of persistent inflammation.3
In ulcerative colitis, the role of immune factors in the pathogenesis of the disease is well established.4 The innate immune response serves as the first line of defense against any attack, mediated by various cell types, including classical immune cells such as neutrophils, monocytes, macrophages, and dendritic cells, as well as non-immune cells such as epithelial cells, endothelial cells, and mesenchymal cells.
In the pathological process of ulcerative colitis, neutrophils serve as core effector cells of acute mucosal inflammation and are among the first immune cells to be recruited during acute inflammatory responses in the intestinal mucosa. Their abnormal activation has been confirmed as a key driver of tissue damage.5 Studies have shown that neutrophils not only exacerbate mucosal barrier damage by releasing pro-inflammatory factors such as TNF-α and IL-1β upon early recruitment to the site of intestinal injury, but also promote the sustained deterioration of the inflammatory microenvironment by forming neutrophil extracellular traps (NETs).3,6 Notably, despite the well-established pro-inflammatory role of neutrophils in IBD, significant gaps remain in our understanding of their intercellular communication networks with intestinal mucosal endothelial cells and other stromal components, which limits the precise development of strategies targeting neutrophil regulation. Although strategies targeting neutrophil infiltration7,8 show promise, existing approaches still have limitations in terms of specificity, efficacy, or safety.
Based on this, the present study utilized single-cell transcriptomics to systematically analyze the inflammatory microenvironment of UC, identifying neutrophil-highly expressed nicotinamide phosphoribosyltransferase (NAMPT), a multifunctional molecule with both metabolic regulation and extracellular signaling functions. eNAMPT may establish a neutrophil-endothelial cell communication axis through its receptor integrin α5β1, thereby participating in the pathogenesis of IBD. Blocking the regulatory axis between neutrophils and endothelial cells can effectively alleviate intestinal inflammation in a UC mouse model. This study not only provides a new perspective on the pathogenesis of ulcerative colitis but also offers potential new therapeutic targets for the disease.
Materials and Methods
Data Sources
The gene expression matrices of normal human intestines and intestinal mucosal samples of UC patients were retrieved from the Gene Expression Omnibus (GEO) database (Home-GEO DataSets-NCBI (nih.gov)). Single cell RNA sequence files for UC and normal samples were obtained using the keyword “single cell ulcerative colitis”. GSE214695 was used to include colon cells from healthy controls (6), ulcerative colitis (6), and Crohn’s disease (6) patients selected from healthy controls and ulcerative colitis (6) patients. The healthy control and ulcerative colitis groups were selected for subsequent single-cell sequencing analysis. All these datasets were obtained in compliance with the ethical standards of their original institutions and have undergone complete anonymization. Pursuant to Article 32, Paragraphs 1 and 2 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (China, 2023), the use of such de-identified publicly available data does not require further ethical review by our institutional ethics review board.
scRNA-Seq Data Analysis
R (version 4.4.1) and the Seurat R package (version 4.3.0) were used for analysis. Using the Seurat R package, a Seurat object was created for each sample using CreateSeuratObject with a gene-by-gene count matrix (parameter: min.cells=5). Cells with high mitochondrial content (>15%) and low gene count assays (<200) were considered low quality cells and discarded. Single-cell sequencing data were normalized using the NormalizeData method, and the FindVariableFeatures function was used to find the top 2,000 high margin genes for subsequent analysis, which were then downscaled by PCA with T-SNE, and significant principal components (PCs) were identified based on the jackStraw function. When combining different samples, highly variable genes were identified, and batch correction was performed using the Harmony algorithm integrated in Seurat. Harmony removes batch-specific and technical variations across samples through iterative correction of principal components, while retaining true biological variability before clustering. Cells were clustered into 23 different cell types (clusters 0–22) at 1.0 resolution using the “FindClusters” function, and the “FindAllMarkers” function was used to identify differentially expressed genes in each cluster (clusters 0–22) differentially expressed genes (DEGs) in each cluster. In addition, some classical markers defined by cell subgroups were obtained from previous studies and manually annotated based on marker expression. The Benjamini–Hochberg false discovery rate (FDR) correction was applied to adjust p-values for multiple hypothesis testing, and genes with FDR < 0.05 were considered statistically significant. To further explore the biological significance of DEGs, Gene Ontology (GO) enrichment analysis was performed using the clusterProfiler R package. Terms with adjusted p-values (FDR < 0.05) were regarded as significantly enriched.
Augur Analysis
The single cell data was divided into ulcerative colitis samples and normal control samples according to the data in GEO, and the Augur package9 was used to determine the most sensitive cell types in the disease and normal groups. This analysis identified the cells that showed the most significant changes under the experimental conditions in a manner independent of the number of cells in each group.
Cell-Cell Communication Analysis
Cell-Cell communication analysis of single-cell transcriptome data was performed using the Cellchat.10 Based on the normalized expression matrix and known ligand-receptor (LR) databases, the probability of communication between each cell type was calculated and significant cell-communication pairs were screened by a permutation test (p < 0.05). Only LR pairs containing at least one receptor gene were retained, excluding receptor-receptor or undefined interactions. The Augur method was further combined to identify the most sensitive cell types in the disease state, which were used as signaling sources to deeply analyze the key ligand-receptor pairs and their associated signaling pathways.
Experimental Animals and Ethics Statement
This investigation utilized male BALB/C mice (aged 8 weeks, 20–21 g body weight) maintained under controlled environmental conditions (temperature: 22 ± 1 °C, humidity: 50 ± 10%) with ad libitum access to standard chow and water following a 7-day acclimation period. All experimental procedures were conducted in accordance with the National Institutes of Health Guidelines for Laboratory Animals and approved by the Institutional Animal Care and Use Committee of Tianjin Medical University General Hospital (Ethics Approval No. IRB2025-DW-50). This investigation adhered to the ARRIVE guidelines 2.0 for preclinical animal studies.
In addition, this study made use of publicly available human transcriptomic datasets obtained from the Gene Expression Omnibus (GEO) database. According to item 1 and item 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (issued on February 18, 2023, China), research involving the use of publicly accessible, de-identified human data is exempt from Institutional Review Board (IRB) approval. Therefore, ethical approval for the analysis of these publicly available human datasets was not required.
DSS-Induced Colitis in Mice
In this experiment, 30 adult BALB/C mice of 8 weeks of age were randomly divided into 5 groups of 6 mice each, of which 4 groups, ie, 24 mice, were administered with drinking water containing 3% Dextran Sulfate Sodium (DSS, Yeasen, China) for 7 days and returned to normal water intake for 3 days to induce acute colitis, and the other 6 mice were given normal water intake as Healthy control group, given normal drinking water. The groups of DSS-fed mice were: the UC model group; the anti-Ly6G group, which was injected intraperitoneally with anti-Ly6G antibody (1.25 mg/kg, Selleck, USA) every 2 days to deplete neutrophils;11 the Daporinad group, which was injected intraperitoneally with Daporinad (2.4 mg/kg, MedChemExpres, USA) every day to inhibit NAMPT activity and reduce eNAMPT levels;12 ATN-161 group, daily intraperitoneal injection of ATN-161 to block integrin α5β1 receptor function (1 mg/kg, MCE).13 Mice were weighed and recorded daily from the beginning of the laboratory and were executed by CO2 asphyxiation on day 10 and whole colon samples as well as serum were obtained for subsequent assays.
Flow Cytometric Analysis (FACS)
To assess the changes in systemic neutrophils, the proportion of whole blood neutrophils in each group of model mice was measured by flow cytometry. Zombie NIR™ stain (ACP-CY7, 423106, BioLegend, USA) was first incubated at room temperature for 10 min to differentiate live cells, followed by incubation of the cells with fluorescently labeled surface antibodies anti-Ly6G (APC, 127614, BioLegend) and anti-CD11b (PE, 101208, BioLegend) for 50 min at 4°C and then addition of PBS to terminate the staining, and finally resuspended in 200 μL PBS for on-line detection. Data were obtained and analyzed using FlowJo software.
Hematoxylin-Eosin Staining (H&E Staining)
Samples from the distal colon of mice were obtained and fixed in 4% paraformaldehyde for 24 hours. The samples were then dehydrated, paraffin-embedded, and cut into 5 μm sections for H&E staining using an ultramicrotome (LEICA, Germany). Each section was observed under a high magnification field of view (400×) of a light microscope and five fields of view were randomly selected, and then scoring criteria were used to quantify the severity of colonic epithelial damage and inflammatory cell infiltration in each group.14 Specifically, colonic epithelial injury was scored as follows: 0, normal; 1, hyperproliferation, loss of cup cells, and abnormal foci of crypts; 2, mild to moderate loss of crypt foci (10–50%); 3, extensive loss of crypt foci (50–90%); 4, complete loss of crypt foci, with epithelium intact; 5, small to medium-sized ulcers (crypt width <10); 6, large ulcers (crypt width ≥ 10). Inflammatory infiltration was categorized as mucosal (0, normal; 1, mild; 2, moderate; 3, severe), submucosal (0, normal; 1, mild to moderate; 2, severe), and muscular/plasma layer (0, regular; 1, moderate to severe), respectively. The total score ranged from 0 to 12 depending on epithelial damage and inflammatory infiltration.
Immunohistochemistry (IHC)
In order to clarify the differences in the expression of eNAMPT and integrin α5β1 in different experimental groups, IHC was used in this study for localization and semi-quantitative analysis. Paraffin-embedded tissue samples were taken and serial sections of 5 μm thickness were prepared, placed on anti-debonding slides and baked at 60°C for 2 hours to enhance tissue adhesion. Sections were deparaffinized to hydration by xylene and gradient ethanol and incubated with 3% H2O2 solution for 15 min at room temperature to eliminate endogenous peroxidase activity and reduce background staining. Sections were placed in EDTA antigen repair buffer (pH 9.0) and thermally induced antigen repair was performed using the microwave repair method (100°C, 15 min) to fully expose the target antigenic epitopes, and the sections were naturally cooled to room temperature after repair was completed. Sections were washed with PBS and closed for 30 min at room temperature using 10% normal goat serum to block non-specific binding. Then anti-NAMPT antibody (1:200, 11776-1-AP, Proteintech Group, USA) and anti-integrin α5β1 antibody (1:150, YT5589, ImmunoWay Biotechnology Company, USA) were added dropwise, respectively, and incubated in a wet box overnight at 4°C. The following day, the sections were rewarmed and incubated with HRP-labeled goat anti-rabbit IgG polymer (EnVision™ Detection System, DAB kit, ZSGB-BIO,China) for 20 min at room temperature, DAB color solution (3,3′-diaminobenzidine) for 3–5 min away from light, and the reaction was terminated by microscopic control. Hematoxylin was used to re-stain the nuclei for 4 min, ethanol hydrochloride was used to differentiate the nuclei, ammonia was used to return the blue color, and neutral dendrimer was used to seal the sections after gradient ethanol dehydration. Each section was observed under a high magnification field of view (400×) of a light microscope and 5 fields of view were randomly selected to count the number of NAMPT or integrin α5β1 positive cells.
Enzyme-Linked Immunosorbent Assay
The concentrations of inflammatory factors TNF-α, IL-6, IL-1β and chemokine CXCL1 in the serum of each group of mice were detected by ELISA kit (DAKEWE, China). Optical density (OD) values were measured with a microplate reader. Mouse serum was centrifuged at 12,000 × g for 15 min at 4°C, and the supernatant was removed after sediment removal and assayed in strict accordance with the manufacturer’s instructions.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism v.10.0 (GraphPad software). Results were expressed as mean ± standard error (SEM). Differences between multiple groups were analyzed using one-way ANOVA. Statistical significance was set at *P<0.05, **P<0.01, and ***P<0.001.
Workflow of the Study
A schematic overview of the overall experimental and analytical design is illustrated in Supplementary Figure 1. The workflow summarizes the entire process of this study, which included single-cell transcriptomic analysis, intercellular communication inference, and in vivo validation.
Result
Single-Cell Transcriptome Analysis of Ulcerative Colitis
In order to analyze the cell infiltration in ulcerative colitis tissues, we selected single-cell transcriptome data from ulcerative colitis (UC) tissues and healthy control (HC) tissues for analysis. A total of nine cell populations were identified by unsupervised clustering. Combining the classical cell marker genes and the expression characteristics of each clustering class, we manually annotated the major immune and intrinsic cell types including B cells, CD4⁺ T cells, NK cells, Fibroblasts, M1 macrophages, Neutrophils, Epithelial cells, Endothelial cells, and Mast cells (Figure 1A). Further comparison of cellular composition differences between the UC and HC groups showed that the proportion of neutrophils was significantly higher in the UC group (Figure 1B and C), suggesting that they may play a key role in the development of the disease.
 |
Figure 1 Single-cell transcriptome reveals differences in cellular composition and neutrophil characteristics between UC and healthy controls. (A) TSNE demonstrating a total of 9 major cell types identified. (B) Cellular composition of UC compared to healthy controls, with a significant increase in the proportion of neutrophils in the UC group. (C) Histogram of the proportion of the different cell types in each sample, further demonstrating a significant increase in neutrophils in the UC group. (D) NAMPT spatial expression distribution map of NAMPT across cell types, further demonstrating a significant increase in neutrophils in the UC group. (E) Violin plot of NAMPT expression in different cell types and different groups. (F) Differential gene distribution plot (Dot plot) of each cell type relative to the others, with the color indicating the average level of gene expression (logFC). (G) Differential expression volcano plot of neutrophils between UC and control, with up-regulated genes in red and down-regulated genes in blue. (H) Differential gene GO functional enrichment analysis. |
In addition, we further focused on the expression pattern of NAMPT gene (proposed by Cell Chat) in different cell types. The TSNE plot (Figure 1D) showed that NAMPT was mainly enriched in neutrophils of UC samples, suggesting that NAMPT may be an important factor mediating the pro-inflammatory function of neutrophils. Figure 1E also clearly showed that the expression of NAMPT in the neutrophils in the UC group had significantly elevated expression compared to the HC group, further emphasizing its specific activation in the disease state. In addition, we demonstrated the genes that were significantly over-expressed or under-expressed in each cell subpopulation relative to other cells (Figure 1F), and these differentially expressed genes helped to more clearly characterize the molecular features of various types of cells, for example, the genes that were significantly over-expressed in neutrophils, including S100A8, S100A9, etc, were closely related to inflammation and immune response. We then analyzed the differentially expressed genes (DEGs) between UC and HC groups (Figure 1G) and performed GO functional enrichment analysis to explore their functional significance (Figure 1H), which showed that these genes were significantly enriched in multiple inflammation-related biological processes such as positive regulation of cytokine production, immune response-activating signaling pathway, and leukocyte migration, and further supported that neutrophils play an important role in the pathogenesis of UC by regulating these pathways.
Augur Algorithm to Identify the Most Differentiated Cell Types Between the Ulcerative Colitis Group and the Normal Group
We used Augur to identify the most differentially responsive cell types in patients with ulcerative colitis versus healthy controls.15 We visualized individual cells based on the AUC difference calculated by the Augur algorithm (Figure 2A), and neutrophils had the highest area under the curve (AUC) for the difference between the two groups (0.712) (Figure 2B) This analysis identified the cells that showed the most pronounced change between the experimental conditions in a manner that was unaffected by the number of cells in each group as being neutrophils, suggesting that neutrophil granulocytes play an important role in the pathophysiologic process of ulcerative colitis.
 |
Figure 2 Cell type annotation based on AUC score. (A) TSNE plot showing the results of cell annotation based on AUC (area under the curve) score. The score is calculated from the set of classical marker genes for each cell. Colors in the graph indicate high or low AUC values. (B) Bar graph of maximum AUC values for each cell type. Neutrophils had the highest AUC (0.712). |
Complex Cellular Communication Networks in Ulcerative Colitis
To further explore the potential communication relationship between different cell types, we performed a systematic analysis of intercellular communication using the CellChat tool. The results showed that in the disease state, cellular communication was mainly mediated by Secreted Signaling, followed by ECM-Receptor and Cell-Cell Contact (Figure 3A), suggesting that in the inflammatory microenvironment of ulcerative colitis, multiple types of signaling pathways are involved. The further constructed cell-to-cell communication network (Figure 3B) reveals the complex interactions among the nine cell subpopulations. The size of the nodes in the figure represents the degree of activity of each cell type in the communication network, and the thickness of the connecting lines indicates the number or strength of interactions between cells. The results showed that B cells, CD4⁺ T cells, and neutrophils interacted most frequently with other cell populations, suggesting that they may play a key role in inflammatory regulation. Given that neutrophils showed significant differences between disease and control groups (confirmed by Augur analysis), we further focused on their associated signaling pathways. By analyzing ligand-receptor pairs between neutrophils and other cells (Supplementary Figure 2), we identified the NAMPT-ITGA5+ITGB1 (integrin α5β1) pathway as having the highest probability of communication between multiple cell types, especially between neutrophils and endothelial cells, suggesting that this pathway may be an important molecular axis for neutrophil-mediated inflammatory response, suggesting that this pathway may be an important molecular axis for neutrophil-mediated inflammatory response. Further pathway-specific network analysis showed that Visfatin (NAMPT) signaling was mainly emitted by neutrophils and acted on multiple cell subpopulations, including endothelial cells and fibroblasts (Figure 3C). Functional role analysis in Figure 3D further suggested that neutrophils played a major role in Visfatin signaling network. The functional role analysis of Figure 3D further suggests that neutrophils play a major role in the Visfatin signaling network as a “Sender”, while endothelial cells mainly act as a “Receiver”, which together constitute a pro-inflammatory communication network centered on neutrophils.
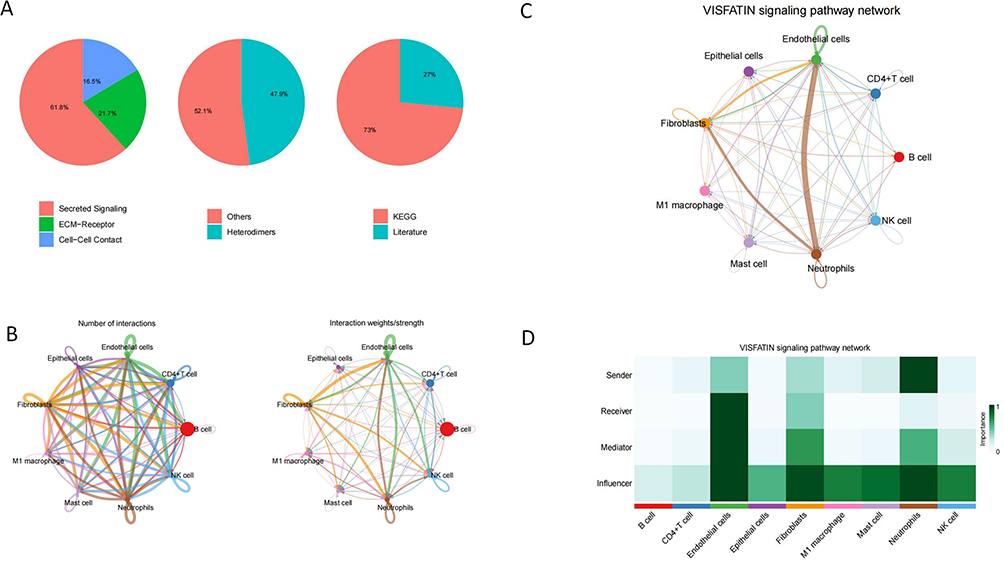 |
Figure 3 Overview of intercellular communication types and VISFATIN signaling pathway network. (A) Three pie charts summarize the types of ligand-receptor interactions, structural composition, and data sources. The majority of the interactions belonged to secretory-type signaling (61.8%), with the rest being ECM-receptor (21.7%) and cell-cell contacts (16.5%); 47.9% of the ligand-receptor pairs were heterodimers; 73% of the data were derived from the KEGG database, and 27% were from the literature support; and (B) Chordal graphs illustrating the overall intercellular communication network. The width of the connection represents the number or strength of interactions. (C) Diagram of the inter-cellular communication network of the Visfatin signaling pathway. Arrows indicate the direction of signaling, and thicker lines indicate higher communication strength. (D) Heatmap showing the roles of each cell type in the Visfatin pathway, including Sender, Receiver, Mediator, and Influencer. |
Neutrophil Depletion Ameliorates DSS-Induced Colitis in Mice by Attenuating the Inflammatory Response
To validate the role of neutrophils in colitis, we used a DSS-induced acute colitis mouse model. Compared with Healthy control group, UC model group mice lost weight (Figure 4A, p<0.001) and shortened colon length (Figure 4B, p<0.001), and HE staining showed localized depletion of cuprocytes, loss of focal crypts, and increase of inflammatory infiltration in the intestines (Figure 4C, p<0.001), and immunohistochemical staining showed a significant increase in neutrophils with localized infiltration in the intestine, suggesting that neutrophils are involved in the pathological process of ulcerative colitis (Figure 4D, p<0.001).
 |
Figure 4 Neutrophil depletion ameliorates DSS-induced colitis in mice by attenuating the inflammatory response. (A) Body weight of each group was monitored for 10 days (n = 6). (B) Representative images of colon and colon length in each group were measured on day 10 (n = 6). (C) Representative H&E-stained micrographs of colon from each group and quantitatively assessed for colon injury using histopathology scores (n = 6); black arrowheads indicate inflammatory cell infiltration (scale bar: 25 μm; original magnification ×400). (D) Representative immunohistochemical staining of Ly6G-positive cells in colonic mucosa (scale bar: 25 μm; original magnification ×400) and quantification of Ly6G-positive cells per high-power field (n = 6), with black arrows indicating positive cells. (E) Representative flow cytometry dot plots showing Ly6G+ cells in peripheral blood and quantification of Ly6G+ cells frequency (n = 6). (F) ELISA assay was performed to examine the serum concentrations of pro-inflammatory cytokines CXCL1, IL-6, TNF-α and IL-1β (n = 6). Statistical analysis was performed by one-way ANOVA. **: p<0.01; ***: p<0.001. |
Flow cytometry results showed a significant increase in the proportion of neutrophils in whole blood of mice in the UC model group compared with the Healthy control group (Figure 4E, p<0.001, for the gating strategy, see Supplementary Figure 3), and the anti-Ly6G antibody given by intraperitoneal injection effectively depleted neutrophils in whole blood of mice (Figure 4E, p<0.001), and immunohistochemical staining showed a significant reduction of localized neutrophil infiltration in the intestine (Figure 4D, p<0.001), as well as an increase in body weight (Figure 4D, p<0.001) compared with the UC model group. Compared with the UC model group, weight regain (Figure 4A, p<0.001), colorectal length lengthening, and histopathological examination showed a significant reduction in the degree of inflammatory damage (Figure 4A–D, p<0.001) (Figure 4B, p<0.001).
In addition elisa results suggested that compared with Healthy control group, the expression levels of pro-inflammatory cytokines TNF-α, IL-6, CXCL1, and IL-1β, which are mainly expressed by neutrophils, were significantly higher in serum of UC model group (Figure 4F, p<0.001), and after depletion of neutrophils The expression levels of pro-inflammatory cytokines TNF-α, IL-6, CXCL1, and IL-1β in serum were significantly decreased (Figure 4F, p<0.001).
The above results strongly confirmed that neutrophils have a crucial role in the development of ulcerative colitis, and the presence of neutrophils aggravated the response of ulcerative colitis, and the removal of neutrophils could attenuate the inflammatory response to ameliorate DSS-induced colitis in mice.
Inhibition of NAMPT Attenuates the Inflammatory Response in DSS-Induced Colitis in Mice
This study continued to investigate the role of eNAMPT released by neutrophils in colitis. The NAMPT inhibitor Daporinad was administered by intraperitoneal injection to inhibit its function. The results suggested that weight regain (Figure 5A, p<0.001) and colorectal length were increased (Figure 5B, p<0.001) after inhibition of NAMPT function in the Daporinad group compared to the UC model group, and histological assessment showed a significant reduction in inflammatory infiltration and damage (Figure 5C, p<0.001), which could be effective in investigating the role of eNAMPT released by neutrophils. 0.001), which could effectively improve intestinal inflammation in mice.
 |
Figure 5 Inhibition of NAMPT attenuates DSS-induced inflammatory response to colitis in mice. (A) Body weight of each group was monitored for 10 days (n = 6). (B) Representative images of colon and colon length in each group were measured on day 10 (n = 6). (C) Representative H&E stained micrographs of colon in each group and quantitative assessment of colon injury using histopathological scores (n = 6); black arrowheads indicate inflammatory cell infiltration (scale bar: 25 μm; original magnification ×400). (D) Representative immunohistochemical staining of NAMPT+ cells in colonic mucosa (scale bar: 25 μm; original magnification ×400) and quantification of NAMPT+ cells per high-power field (n = 6), with black arrows indicating positive cells. (E) ELISA assay was performed to examine the serum concentrations of pro-inflammatory cytokines CXCL1, IL-6, TNF-α and IL-1β (n = 6). Statistical analysis was performed by one-way ANOVA. ***: p<0.001. |
Immunohistochemical staining results suggested that the protein expression level of NAMPT was increased in the localized intestinal tract of the UC model group compared with that of the Healthy control group (Figure 5D, p<0.001), and the application of the NAMPT inhibitor effectively reduced the localized intestinal tract expression of NAMPT (Figure 5D, p<0.001), and the protein expression level of NAMPT was significantly reduced in the intestinal localization after neutrophil depletion (Figure 5D, p<0.001). In addition the expression levels of pro-inflammatory cytokines TNF-α, IL-6, CXCL1 and IL-1β were reduced in the Daporinad group compared to the UC model group (Figure 5E, p<0.001).
The above results suggest that the expression of neutrophil-derived eNAMPT is elevated in ulcerative colitis and inhibition of eNAMPT can effectively attenuate the inflammatory response of DSS-induced colitis in mice.
Inhibition of Integrin α5β1 Receptor Function Attenuates Inflammatory Injury in DSS-Induced Colitis in Mice
This study continues to investigate the role of integrin α5β1, the primary receptor for eNAMPT released by neutrophils, in colitis. ATN-161, an inhibitor of integrin α5β1, was given by intraperitoneal injection to inhibit its function. The results suggested that weight regain (Figure 6A, p<0.001) and colorectal length (Figure 6B, p<0.05) were increased in the ATN-161 group compared to the UC model group after inhibition of the function of integrin α5β1, and histological assessment showed a significant reduction in the inflammatory infiltration and damage (Figure 6C, p<0.05), which was effective in reducing the inflammatory infiltration and damage. p<0.05), effectively improving intestinal inflammation in mice.
 |
Figure 6 Inhibition of integrin α5β1 receptor function attenuates inflammatory injury in DSS-induced colitis in mice. (A) Body weight of each group was monitored for 10 days (n = 6). (B) Representative images of colon and colon length were measured in each group on day 10 (n = 6). (C) Representative H&E stained micrographs of colon in each group and quantitative assessment of colon injury using histopathology scores (n = 6); black arrowheads indicate inflammatory cell infiltration (scale bar: 25 μm; original. (D) Representative immunohistochemical staining of integrin α5β1+ cells in colonic mucosa (scale bar: 25 μm; original magnification ×400) and quantification of integrin α5β1+ cells per high-power field (n = 6), with black arrows indicating positive cells. (E) ELISA assay was performed to examine the serum concentrations of pro-inflammatory cytokines CXCL1, IL-6, TNF-α and IL-1β (n = 6). Statistical analysis was performed by one-way ANOVA. ns: not significant; *: p<0.05; ***: p<0.001. |
Immunohistochemical staining results suggested an increase in the number of integrin α5β1+ cells localized in the intestines of the UC model group compared with the Healthy control group (Figure 6D, p<0.001). There was no difference in the expression level of integrin α5β1 localized in the intestines of the ATN-161 group compared to the UC model group (Figure 6D, ns). In addition the expression levels of pro-inflammatory cytokines TNF-α, IL-6, CXCL1, and IL-1β were reduced in the ATN-161 group compared to the UC model group (Figure 6E, p<0.001).
The above results suggest that the expression of integrin α5β1 is elevated in ulcerative colitis and that inhibition of integrin α5β1 function can effectively attenuate the inflammatory injury in DSS-induced colitis in mice.
Discussion
The pathogenesis of ulcerative colitis is highly complex and has not been fully elucidated, but immune-mediated pathologic injury has been identified as a central pathogenetic mechanism. In this process, neutrophils, as the core effector cells of acute inflammation in the mucosa, are the first myeloid immune cells to be directionally recruited to the site of intestinal inflammation. Their aberrant activation drives tissue injury through pro-inflammatory mediator release, NETosis effects, and immune cell interactions. The infiltration density of neutrophils is significantly and positively correlated with the depth of mucosal ulceration and disease activity index in patients with UC, and is a key enforcer of tissue destruction in the acute phase.1–5
Indeed in this study we found neutrophils to be the most highly associated immune cells with a dominant role in UC by analyzing single-cell sequencing data. Combined with animal experiments we found that the proportion of neutrophils increased in modeled mice, as did the local infiltration of the intestinal tract, while depletion of neutrophils significantly improved the intestinal inflammatory response, reduced the expression levels of the pro-inflammatory cytokines TNF-α, IL-6, CXCL1, and IL-1β, and attenuated inflammatory injury while alleviating shortening of the intestinal tract length, which was favorable for weight regain, highlighting the role of the critical role of neutrophils in UC.
When analyzing the possible pathogenic mechanisms of neutrophils in depth, we found that neutrophils highly express Visfatin (NAMPT), which is the key rate-limiting enzyme for nicotinamide adenine dinucleotide (NAD) synthesis, and plays a key role in intracellular NAD metabolic homeostasis.12 In addition to functioning as an enzyme in the intracellular compartment to participate in the intracellular metabolic control, NAMPT can be secreted to the extracellular compartment to play a cytokine-like role.13 In addition to its intracellular role as an enzyme in the regulation of intracellular metabolism, NAMPT can also be secreted into the extracellular compartment to play cytokine-like roles in the regulation of inflammatory responses.13,16 Extracellular NAMPT (eNAMPT) is strongly up-regulated in a variety of immune disorders,17–20 including IBD, and plays an important role in the inflammatory process. These constitute an important mechanistic link between inflammatory, metabolic and transcriptional pathways and NAD metabolism.4
In fact, the present study found that the local expression level of NAMPT protein in the intestine of mice with experimental colitis was increased and significantly decreased after removal of neutrophils, whereas the application of inhibitors of NAMPT exerted a therapeutic effect by depleting intracellular NAMPT stores and thereby reducing the expression of eNAMPT to alleviate the intestinal inflammatory response, suggesting that UC neutrophils are the main source of eNAMPT and are the major source of eNAMPT.
Based on cellular communication and ligand-receptor pair analysis, we found that eNAMPT-integrin α5β1 ligand-receptor pairs play a key role in neutrophil-endothelial cell communication in ulcerative colitis. Indeed the current study concludes that integrin α5β1 acts as a major functional receptor for eNAMPT.21–24 Integrin family proteins act as adhesion molecules between cells and between cells and the extracellular matrix (ECM) and play a key role in immune cell migration and inflammatory responses,25 among which integrin α5β1 has been reported to potentially play an important role in several inflammatory and autoimmune diseases.26 Studies have shown that integrin α5β1 is significantly upregulated in intestinal inflammation and is involved in the migration of immune cells, especially neutrophils, to inflammatory sites.13 From a translational medicine perspective, targeting this axis holds promise for complementary or combined strategies with existing anti-inflammatory therapies (such as anti-TNF-α drugs), particularly offering new potential treatment options for patients with refractory ulcerative colitis.
Endothelial cells play an important role in the pathological process of ulcerative colitis27,28 and their dysfunction directly drives the inflammatory cascade response.29,30 Activated endothelial cells show barrier function disruption in response to stimulation by pro-inflammatory factors (eg, TNF-α, IL-1β), which is manifested by down-regulation of the expression of tight junction proteins (e.g.claudin-531) and an increase in vascular permeability, leading to plasma exudation and local tissue edema.32,33
Meanwhile, endothelial cells highly express integrins,34 especially integrin α5β1,35,36 in the inflammatory environment, driving the adhesion of immune cells, including neutrophils, and infiltration into the inflamed intestinal mucosa. In addition, cytokines such as IL-6 and IL-837 secreted by endothelial cells further activate mesenchymal stromal cells,38 forming a positive feedback amplification loop of pro-inflammatory signals. Targeted modulation of endothelial activation (e.g.blocking adhesion molecules) has emerged as a novel strategy for intervention in UC.39,40
In the present study, it was found that the expression of integrin α5β1 was increased locally in the intestine of mice with experimental colitis, and the application of an inhibitor of integrin α5β1 alleviated the intestinal inflammatory response by inhibiting the function of integrin α5β1. These results further suggest that neutrophils act on integrin α5β1 expressed by endothelial cells by releasing Visfatin/eNAMPT, thereby inducing inflammatory responses and intestinal injury.
In conclusion, the present study found that the abnormal activation of neutrophils in UC led to an increase in the expression of eNAMPT, and the increased eNAMPT induced the degree of inflammatory damage in ulcerative colitis by acting on the integrin α5β1 receptor of endothelial cells experimenting that neutrophils as well as endothelial cell communication together, which provides a new perspective for the pathogenesis of ulcerative colitis, and provides a new potential target for the treatment of ulcerative colitis, providing new potential targets. However, there are some limitations in this study. First, although we initially verified the relationship between eNAMPT and integrin α5β1 with the help of animal experiments, the specific downstream signaling pathways still need to be further explored and clarified. Second, this study mainly focused on the acute colitis model, and the mechanism of action in long-term chronic inflammation still needs more experimental verification.
This study has certain limitations. Bioinformatics analysis was based on a single cohort with a small sample size in vivo, and primarily utilized an acute colitis model, factors that limit the generalizability of the results. Furthermore, downstream signaling pathways of the eNAMPT-integrin α5β1 axis require further investigation. The lack of detailed clinical metadata in the databases used also represents a potential constraint. Therefore, future studies should validate this pathway in larger-scale studies and chronic disease models, while focusing on elucidating its downstream molecular mechanisms to assess its clinical translational value.
Conclusion
In this study, by integrating single-cell sequencing technology, bioinformatics analysis and in vivo experiments, we preliminarily revealed that aberrant activation of neutrophils in UC achieves cellular communication with endothelial cells and aggravates the inflammatory response in ulcerative colitis through the eNAMPT-integrin α5β1 pathway. This study provides a theoretical basis and experimental evidence to improve the understanding of the pathogenesis of ulcerative colitis and to develop new therapeutic strategies. At the same time, we recognize that these findings are preliminary, and their potential for clinical translation must be validated in larger-scale studies and more complex disease models.
Data Sharing Statement
The datasets generated during the current study are available in public databases. The dataset numbers were mentioned in Materials and Methods. The data that support the findings of this study are available from the corresponding author upon reasonable request.
Funding
The project was funded by National Natural Science Foundation of China (82170399) and The Natural Science Foundation of Tianjin (23JCZDJC00540).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Ordás I, Eckmann L, Talamini M. et al. Ulcerative colitis. Lancet. 2012;380(9853):1606–1619. doi:10.1016/S0140-6736(12)60150-0
2. de Souza HS, Fiocchi C. Immunopathogenesis of IBD: current state of the art. Nat Rev Gastroenterol Hepatol. 2016;13(1):13–27. doi:10.1038/nrgastro.2015.186
3. Brazil JC, Louis NA, Parkos CA. The role of polymorphonuclear leukocyte trafficking in the perpetuation of inflammation during inflammatory bowel disease. Inflamm Bowel Dis. 2013;19(7):1556–1565. doi:10.1097/MIB.0b013e318281f54e
4. Gerner RR, Klepsch V, Macheiner S, et al. NAD metabolism fuels human and mouse intestinal inflammation. Gut. 2018;67(10):1813–1823. doi:10.1136/gutjnl-2017-314241
5. Kim JS, HK Kim, M Kim, et al. Colon-Targeted eNAMPT-Specific Peptide Systems for Treatment of DSS-Induced Acute and Chronic Colitis in Mouse. Antioxidants. 2022;11(12):2376.
6. Dinallo V, Marafini I, Di Fusco D, et al. Neutrophil Extracellular Traps Sustain Inflammatory Signals in Ulcerative Colitis. J Crohns Colitis. 2019;13(6):772–784. doi:10.1093/ecco-jcc/jjy215
7. Wallace JL, Higa A, Webb McKnight G, et al. Prevention and reversal of experimental colitis by a monoclonal antibody which inhibits leukocyte adherence. Inflammation. 1992;16(4):343–354. doi:10.1007/BF00917626
8. Feagan BG, Rutgeerts P, Sands BE, et al. Vedolizumab as induction and maintenance therapy for ulcerative colitis. N Engl J Med. 2013;369(8):699–710. doi:10.1056/NEJMoa1215734
9. Skinnider MA, Squair JW, Kathe C, et al. Cell type prioritization in single-cell data. Nat Biotechnol. 2021;39(1):30–34. doi:10.1038/s41587-020-0605-1
10. Jin S, Guerrero-Juarez CF, Zhang L, et al. Inference and analysis of cell-cell communication using CellChat. Nat Commun. 2021;12(1):1088. doi:10.1038/s41467-021-21246-9
11. Boivin G, Faget J, Ancey P-B, et al. Durable and controlled depletion of neutrophils in mice. Nat Commun. 2020;11(1):2762. doi:10.1038/s41467-020-16596-9
12. Romacho T, Valencia I, Ramos-González M, et al. Visfatin/eNampt induces endothelial dysfunction in vivo: a role for Toll-Like Receptor 4 and NLRP3 inflammasome. Sci Rep. 2020;10(1):5386. doi:10.1038/s41598-020-62190-w
13. Danese S, Sans M, Spencer DM, et al. Angiogenesis blockade as a new therapeutic approach to experimental colitis. Gut. 2007;56(6):855–862. doi:10.1136/gut.2006.114314
14. Lee Y, Sugihara K, Gillilland MG, et al. Hyaluronic acid-bilirubin nanomedicine for targeted modulation of dysregulated intestinal barrier, microbiome and immune responses in colitis. Nat Mater. 2020;19(1):118–126. doi:10.1038/s41563-019-0462-9
15. Squair JW, Skinnider MA, Gautier M, et al. Prioritization of cell types responsive to biological perturbations in single-cell data with Augur. Nat Protoc. 2021;16(8):3836–3873. doi:10.1038/s41596-021-00561-x
16. Travelli C, Colombo G, Aliotta M, et al. Extracellular nicotinamide phosphoribosyltransferase (eNAMPT) neutralization counteracts T cell immune evasion in breast cancer. J Immunother Cancer. 2023;11(10):e007010. doi:10.1136/jitc-2023-007010
17. Kumari B, Yadav UCS. Adipokine Visfatin’s Role in Pathogenesis of Diabesity and Related Metabolic Derangements. Curr Mol Med. 2018;18(2):116–125. doi:10.2174/1566524018666180705114131
18. Garcia AN, Casanova NG, Kempf CL, et al. eNAMPT Is a Novel Damage-associated Molecular Pattern Protein That Contributes to the Severity of Radiation-induced Lung Fibrosis. Am J Respir Cell Mol Biol. 2022;66(5):497–509. doi:10.1165/rcmb.2021-0357OC
19. Moschen AR, Kaser A, Enrich B, et al. Visfatin, an adipocytokine with proinflammatory and immunomodulating properties. J Immunol. 2007;178(3):1748–1758. doi:10.4049/jimmunol.178.3.1748
20. Romacho T, Sánchez-Ferrer CF, Peiró C. Visfatin/Nampt: an adipokine with cardiovascular impact. Mediators Inflamm. 2013;2013:946427. doi:10.1155/2013/946427
21. Liao Z, Jin Y, Chu Y, et al. Single-cell transcriptome analysis reveals aberrant stromal cells and heterogeneous endothelial cells in alcohol-induced osteonecrosis of the femoral head. Commun Biol. 2022;5(1):324. doi:10.1038/s42003-022-03271-6
22. Shi J, Q Li, J Li, et al. Single-Cell RNA Sequencing Reveals the Spatial Heterogeneity and Functional Alteration of Endothelial Cells in Chronic Hepatitis B Infection. Int J Mol Sci. 2024;25(13).
23. Yang W, Wang P, Luo M, et al. DeepCCI: a deep learning framework for identifying cell-cell interactions from single-cell RNA sequencing data. Bioinformatics. 2023;39(10). doi:10.1093/bioinformatics/btad596
24. Peng Z, Ren Z, Tong Z, et al. Interactions between MFAP5 + fibroblasts and tumor-infiltrating myeloid cells shape the malignant microenvironment of colorectal cancer. J Transl Med. 2023;21(1):405. doi:10.1186/s12967-023-04281-6
25. Pang X, He X, Qiu Z, et al. Targeting integrin pathways: mechanisms and advances in therapy. Signal Transduct Target Ther. 2023;8(1):1. doi:10.1038/s41392-022-01259-6
26. Morshed A, Abbas AB, Hu J, et al. Shedding New Light on The Role of ανβ3 and α5β1 Integrins in Rheumatoid Arthritis. Molecules. 2019;24(8):1537. doi:10.3390/molecules24081537
27. Tolstanova G, Deng X, French SW, et al. Early endothelial damage and increased colonic vascular permeability in the development of experimental ulcerative colitis in rats and mice. Lab Invest. 2012;92(1):9–21. doi:10.1038/labinvest.2011.122
28. Britzen-Laurent N, Weidinger C, Stürzl M. Contribution of Blood Vessel Activation, Remodeling and Barrier Function to Inflammatory Bowel Diseases. Int J Mol Sci. 2023;24(6). doi:10.3390/ijms24065517
29. Cromer WE, JM Mathis, DN Granger, et al. Role of the endothelium in inflammatory bowel diseases. World J Gastroenterol. 2011;17(5):578–593. doi:10.3748/wjg.v17.i5.578
30. Gravina AG, Dallio M, Masarone M, et al. Vascular Endothelial Dysfunction in Inflammatory Bowel Diseases: pharmacological and Nonpharmacological Targets. Oxid Med Cell Longev. 2018;2018(1):2568569. doi:10.1155/2018/2568569
31. Kakogiannos N, Ferrari L, Giampietro C, et al. JAM-A Acts via C/EBP-α to Promote Claudin-5 Expression and Enhance Endothelial Barrier Function. Circ Res. 2020;127(8):1056–1073. doi:10.1161/CIRCRESAHA.120.316742
32. Park JS, Cresci GAM. Dysfunctional intestinal microvascular endothelial cells: insights and therapeutic implications in gastrointestinal inflammation. Immunometabolis. 2024;6(2):e00043. doi:10.1097/IN9.0000000000000043
33. Naschberger E, Flierl C, Huang J, et al. Analysis of the interferon-γ-induced secretome of intestinal endothelial cells: putative impact on epithelial barrier dysfunction in IBD. Front Cell Dev Biol. 2023;11:1213383. doi:10.3389/fcell.2023.1213383
34. Aman J, Margadant C. Integrin-Dependent Cell-Matrix Adhesion in Endothelial Health and Disease. Circ Res. 2023;132(3):355–378. doi:10.1161/CIRCRESAHA.122.322332
35. Li L, Welser-Alves J, van der Flier A, et al. An angiogenic role for the α5β1 integrin in promoting endothelial cell proliferation during cerebral hypoxia. Exp Neurol. 2012;237(1):46–54. doi:10.1016/j.expneurol.2012.06.005
36. Valdembri D, Caswell PT, Anderson KI, et al. Neuropilin-1/GIPC1 signaling regulates alpha5beta1 integrin traffic and function in endothelial cells. PLoS Biol. 2009;7(1):e25. doi:10.1371/journal.pbio.1000025
37. Romani de Wit T, De Leeuw HPJC, Rondaij MG, et al. Von Willebrand factor targets IL-8 to Weibel-Palade bodies in an endothelial cell line. Exp Cell Res. 2003;286(1):67–74. doi:10.1016/S0014-4827(03)00094-6
38. Sierra-Parraga JM, Merino A, Eijken M, et al. Reparative effect of mesenchymal stromal cells on endothelial cells after hypoxic and inflammatory injury. Stem Cell Res Ther. 2020;11(1):352. doi:10.1186/s13287-020-01869-3
39. Soriano A, Salas A, Salas A, et al. VCAM-1, but not ICAM-1 or MAdCAM-1, immunoblockade ameliorates DSS-induced colitis in mice. Lab Invest. 2000;80(10):1541–1551. doi:10.1038/labinvest.3780164
40. Ghosh S, Panaccione R. Anti-adhesion molecule therapy for inflammatory bowel disease. Therap Adv Gastroenterol. 2010;3(4):239–258. doi:10.1177/1756283X10373176
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification and Validation of Hub Genes Related to Neutrophil Extracellular Traps-Mediated Cell Damage During Myocardial Infarction

Ke D, Ni J, Yuan Y, Cao M, Chen S, Zhou H
Journal of Inflammation Research 2024, 17:617-637
Published Date: 1 February 2024
Network Analysis of Osteoarthritis Progression Using a Steiner Minimal Tree Algorithm
Xie Y, Shao F, Ji Y, Feng D, Wang L, Huang Z, Wu S, Sun F, Jiang H, Miyamoto A, Wang H, Zhang C
Journal of Inflammation Research 2024, 17:3201-3209
Published Date: 18 May 2024
Differential Expression of IFI16, IL-33 and CD55 Link Potential Common Pathogenic Mechanisms for COVID-19 and Ulcerative Colitis
Zhang F, Di Q, Li Y, Ye J, Wang B, Ding Y
Journal of Multidisciplinary Healthcare 2025, 18:8027-8041
Published Date: 17 December 2025
Identification and Validation of Key Purine Metabolism-Related Genes in Ulcerative Colitis Using Bioinformatics and Machine Learning
Zhang S, Zhang Y, Du D, Zeng Y, Zhang S, Wang Q, Xue W, Wen X, Lan Y, Hu W
Journal of Inflammation Research 2026, 19:557806
Published Date: 8 January 2026