Back to Journals » Journal of Inflammation Research » Volume 19
Targeting the Gut Microbiota with Herbal Compounds from Traditional Chinese Medicine: A Mechanistic Synthesis of a Novel Therapeutic Approach for Ulcerative Colitis
Authors Chen J ![]() , Chen J
, Chen J ![]() , Wang Z
, Wang Z ![]()
Received 22 December 2025
Accepted for publication 26 May 2026
Published 29 June 2026 Volume 2026:19 590456
DOI https://doi.org/10.2147/JIR.S590456
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Syed Adeel Hassan
Junyi Chen,1,* Jiaxu Chen,2,* Zhenyi Wang1
1Department of Coloproctology, Yueyang Hospital of Integrated Traditional Chinese and Western Medicine, Shanghai, 200080, People’s Republic of China; Shanghai University of Traditional Chinese Medicine, Shanghai, 201203, People’s Republic of China; 2Department of Rheumatology and Immunology, Yueyang Hospital of Integrated Traditional Chinese and Western Medicine, Shanghai, 200080, People’s Republic of China; Shanghai University of Traditional Chinese Medicine, Shanghai, 201203, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhenyi Wang, Email [email protected]
Abstract: Ulcerative colitis (UC) represents a chronic relapsing inflammatory bowel disease characterized by substantial unmet clinical needs and limited curative modalities. Accumulating evidence implicates gut microbiota dysbiosis as a pivotal pathogenic driver, positioning microbiota-targeted interventions as promising therapeutic strategies. This review systematically delineates the mechanisms by which herbal compounds from Traditional Chinese Medicine ameliorate UC through the restoration of microbial and metabolic homeostasis—including the modulation of beneficial commensals and their bioactive metabolites—thereby reinforcing intestinal barrier integrity and dampening mucosal inflammation. Although translational bottlenecks persist, integrative multi-omics frameworks coupled with advanced pharmaceutical engineering offer viable pathways to bridge preclinical findings and clinical application. Taken together, deciphering the bidirectional crosstalk between herbal compounds and the gut microbiome paves the way for mechanism-based, personalized botanical therapeutics in UC management.
Keywords: ulcerative colitis, herbal compounds, gut microbiota, intestinal barrier, translational challenges
Introduction
Ulcerative colitis (UC), a chronic, relapsing inflammatory bowel disease (IBD) characterized by mucosal inflammation of the colon and rectum,1 has emerged as a major global health challenge due to its increasing incidence and prevalence worldwide,2,3 debilitating clinical outcomes, and the inadequacy of current therapeutic strategies. A systematic review demonstrated that UC incidence has increased by 2 to 5 fold over the past three decades in newly industrialized regions, including East Asia and Latin America, while prevalence in Western countries remains elevated at 50–200 cases per 100,000 population.4 Clinically, UC imposes a significant burden due to its chronic relapsing course, which often leads to complications such as severe diarrhea, rectal bleeding, and malnutrition,5,6 and confers a 1.7-fold increased lifetime risk of colorectal cancer in long-standing cases.7,8 Economically, UC imposes a substantial global healthcare burden, with high direct costs arising from hospitalization for disease flares and long-term medication use, as well as significant indirect costs due to productivity losses.9 Despite the availability of conventional therapeutic classes including 5-aminosalicylic acid compounds, corticosteroids, immunosuppressants, and biologic agents such as anti-tumor necrosis factor monoclonal antibodies,10 a substantial proportion of patients (up to 30%) fail to achieve sustained remission, underscoring a critical efficacy gap in current management strategies. Furthermore, long-term use of these agents is associated with serious adverse effects, including opportunistic infections, osteoporosis, and an elevated risk of lymphoma, which collectively contribute to the significant limitations of existing pharmacologic approaches.11 These efficacy and safety gaps underscore the urgent imperative for novel therapies that target the core pathogenic pathways of UC.12
Intestinal microbiota dysbiosis is a well-established central driver in the pathogenesis of ulcerative colitis.13 Clinical and animal studies consistently identify hallmark features of UC-associated dysbiosis, including a decreased relative abundance of Firmicutes alongside an expansion of Proteobacteria.14,15 Expanded Proteobacteria, particularly Enterobacteriaceae, elevate luminal lipopolysaccharide (LPS) levels,16 activates Toll-like receptor 4 (TLR4) on epithelial and immune cells, triggering a nuclear transcription factor-κB (NF-κB)-mediated production of pro-inflammatory cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α).17,18 This process directly links microbial dysbiosis to the inflammatory cascades characteristic of ulcerative colitis flares. Concurrently, the depletion of butyrate-producing commensals results in diminished luminal butyrate levels, which is a short-chain fatty acid (SCFA) that serves as the primary energy source for colonocytes and critically regulates the expression and function of tight junction proteins.19 This impairment of butyrate-mediated epithelial homeostasis directly compromises intestinal barrier integrity, leading to increased permeability and mucosal damage that characterizes active ulcerative colitis.20 Evidence from germ-free mouse models demonstrates that colonization with microbiota derived from UC patients can induce severe colitis, confirming the pathogenic potential of dysbiotic communities,21 while clinical studies show that fecal microbiota transplantation (FMT) from healthy donors achieves symptom alleviation and clinical remission in a significant proportion of refractory UC cases, further substantiating the causal role of microbiota imbalance in disease pathogenesis.22,23
Traditional Chinese Medicine (TCM) has been utilized for millennia to treat gastrointestinal disorders, and contemporary preclinical and clinical research now substantiates its efficacy in promoting gut health.24 Ancient TCM texts, such as the Huangdi Neijing and Shennong Bencao Jing, document the use of herbal formulas like Scutellaria Decoction (Huangqin Tang) for treating symptoms analogous to UC, with efficacy in ameliorating intestinal inflammation and restoring gut barrier function demonstrated in murine colitis models.25 In dextran sulfate sodium (DSS)-induced mice models of colitis, herbals like Scutellaria baicalensis (Huangqin) significantly reduce intestinal inflammation and improve barrier integrity,26,27 while Astragalus membranaceus (Huangqi) modulate key inflammatory pathways such as NF-κB and NOD-like Receptor Pyrin domain containing 3 (NLRP3) inflammasome activation.28,29 Meta-analyses of randomized controlled trials have established that TCM interventions, particularly herbal medicines, significantly improve clinical remission rates in UC patients when combined with conventional therapy,30 thereby providing robust clinical validation for TCM’s efficacy in gut health management.
Despite the well-established association of gut microbiota dysbiosis in UC pathogenesis13 and the historical application of herbal remedies for gastrointestinal disorders,24 the mechanistic connections between herbal compounds, gut microbiota modulation, and UC resolution remain incompletely elucidated. To address this gap, this review synthesizes mechanistic evidence to clarify the relationship between the microbiota and UC pathogenesis, links herbal compounds to gut microbiota modulation, and ultimately elucidates how herbal compounds improve UC by regulating the microbiota. Furthermore, we critically examine the translational bottlenecks between preclinical findings and clinical implementation of herbal therapeutics. Integrating contemporary technological advances, we propose that multi-omics frameworks coupled with advanced pharmaceutical delivery systems represent pivotal strategies to bridge this translational divide and drive future breakthroughs in microbiota-targeted UC management.
The Gut Microbiota in Health and UC
Normal Gut Microbiota: Composition, Functions, and Host-Microbe Symbiosis
The normal gut microbiota is a complex and dynamic community, predominantly composed of trillions of microorganisms from the phyla Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, and Verrucomicrobia (Figure 1). These microorganisms engage in a mutually beneficial symbiotic relationship with the host, performing critical metabolic, immune-modulatory, and barrier-supporting functions. Firmicutes (eg., Lactobacillus, Faecalibacterium prausnitzii, Roseburia spp.) and Bacteroidetes (eg., Bacteroides thetaiotaomicron, Prevotella copri) collectively account for approximately 90% of the adult gut microbiota. Actinobacteria (eg., Bifidobacterium spp)., Proteobacteria (eg., Escherichia coli), and Verrucomicrobia (Akkermansia muciniphila) represent the remaining minority.31
 |
Figure 1 The core gut microbiota in health and its symbiotic influence on the host. |
This microbial community establishes a mutualistic symbiosis with the human host, representing an integral component of intestinal anatomy and physiology. The human gastrointestinal tract provides a unique ecological niche for these microorganisms, characterized by a strictly anaerobic environment, stable temperature, controlled pH gradients, and a continuous supply of nutrients derived from the diet (indigestible fibers, resistant starches) and host secretions (mucins, sloughed epithelial cells). In return, these commensal microbes actively shape intestinal function through their metabolic output, immunomodulatory signals, and competitive exclusion of pathogens, thereby forming a functional meta-organism essential for host homeostasis.32
A primary function of the gut microbiota is the metabolic biotransformation of indigestible dietary components and host-derived substrates into a diverse array of bioactive metabolites that regulate host physiology. Central to this metabolic capacity is the fermentation of dietary fibers and complex carbohydrates, which evade host enzymatic digestion in the small intestine and reach the distal gut intact. Microbial carbohydrate-active enzymes, including polysaccharidases and glycosidases, hydrolyze these complex glycans into five- or six-carbon monosaccharides that undergo catabolism via the pentose phosphate pathway or Embden–Meyerhof–Parnas glycolysis to phosphoenolpyruvate and pyruvate. These intermediates subsequently fuel multiple anaerobic fermentation pathways to produce short-chain fatty acids (SCFAs; predominantly acetate, propionate, and butyrate) as terminal end products. Concurrently, the microbiota engages in extensive protein fermentation, wherein microbial proteases hydrolyze dietary and host-derived proteins into amino acids that undergo dissimilatory catabolic reactions, generating not only SCFAs and branched-chain fatty acids (BCFAs) but also ammonia, phenolic compounds, and indole derivatives. Furthermore, microbial bile salt hydrolases deconjugate primary bile acids, enabling subsequent 7α/β-dehydroxylation, epimerization, and oxidation reactions that yield diverse secondary bile acid pools, while tryptophanase activity directly transforms tryptophan into indole and its derivatives.33
Beyond metabolism, the normal gut microbiota plays a vital role in educating the host immune system by promoting immunoregulatory responses. For example, non-toxigenic Bacteroides fragilis produces polysaccharide A (PSA), which induces Foxp3⁺ regulatory T cells (Tregs) and enhances interleukin-10 (IL-10) secretion, thereby suppressing excessive pro-inflammatory responses and supporting intestinal immune homeostasis.34 Similarly, Bacteroides uniformis modulates intestinal bile acid metabolism35 and restores the expression of intestinal immune barrier proteins by suppressing T-helper 17 (Th17) cell differentiation in colonic epithelial cells and inhibiting downstream NF-κB and MAPK inflammatory signaling pathways.36
In addition to immune modulation, beneficial commensals from minority phyla are pivotal in maintaining physical barrier integrity. Beneficial commensals from minority phyla, including Actinobacteria and Verrucomicrobia, reinforce the intestinal barrier through distinct but complementary mechanisms. Bifidobacterium spp. (Actinobacteria) produce acetate and lactate to lower luminal pH and inhibit pathogen growth, while generating exopolysaccharides that strengthen the mucus layer and competing with detrimental microorganisms for nutrients and adhesion sites.37–40 Concurrently, A. muciniphila (Verrucomicrobia) enhances barrier integrity by degrading mucin to stimulate goblet cell production and upregulating tight junction proteins (zonula occludens-1 (ZO-1), occludin, and claudin-1), thereby maintaining epithelial tightness.41
Finally, the symbiotic equilibrium depends on the proper management of even low-abundance phyla, such as Proteobacteria (eg., E. coli). At baseline levels, these bacteria contribute to metabolic functions and microbial balance; however, their overgrowth transforms them into opportunistic pathogens that drive inflammatory pathology.42
Gut Dysbiosis in UC: Altered Diversity, Metabolism, and Immune Interactions
Gut dysbiosis in UC represents a profound disruption in the composition and balance of the gut microbiota, which alters microbial metabolism, undermines epithelial function, and perpetuates chronic intestinal inflammation.13,43 One of the most notable changes in UC is the depletion of beneficial microbes, particularly those belonging to the Firmicutes phylum, which are key producers of SCFAs, especially butyrate.44 The depletion of these key SCFA-producing bacteria, initiates a cascade of detrimental effects. It deprives colonocytes of their primary metabolic fuel, leading to an energy crisis that impairs cellular function and survival. Crucially, the loss of SCFA signaling undermines multiple protective mechanisms: it compromises epithelial barrier integrity by reducing the expression of tight junction proteins (eg., claudin-1) and mucin (MUC2), weakens the colonic hypoxic barrier favorable to commensals, and diminishes the production of antimicrobial peptides (AMPs). Furthermore, the absence of SCFAs, notably butyrate and propionate, disrupts immunomodulation by failing to inhibit histone deacetylases (HDAC) and activate receptors like G-protein-coupled receptor 43 (GPR43) and GPR109a on immune cells. This results in diminished anti-inflammatory cytokine production (eg., IL-10, TGF-β1), impaired differentiation of Tregs, and a consequent loss of braking mechanisms on the mucosal immune system, thereby directly fueling and perpetuating chronic inflammation45 Beyond SCFA production, F. prausnitzii also plays a crucial role in suppressing NF-κB activation, which regulates the release of pro-inflammatory cytokines; its depletion thus worsens both inflammation and impedes mucosal healing.46 Similarly, Roseburia species help reinforce the mucus barrier and promote Treg differentiation, vital for maintaining immune tolerance.47
Conversely, UC is marked by the expansion of pathogenic bacteria, particularly within the Proteobacteria phylum.48 Pathobionts such as adherent-invasive Escherichia coli (AIEC) thrive in the inflammatory milieu of the UC colon.49 These bacteria exploit nitrate and oxygen radicals, byproducts of the host’s inflammatory response, to fuel their growth.50 The overgrowth of these pathogens leads to epithelial invasion and triggers the activation of TLR4 via LPS binding. This activation subsequently stimulates the NLRP3 inflammasome, leading to the production of potent pro-inflammatory cytokines like IL-1β and TNF-α. This initiates a self-perpetuating inflammatory loop, intensifying mucosal immune activation and driving UC progression.51
Beyond these dominant phyla, multifaceted taxonomic perturbations further compromise mucosal homeostasis. Bacteroidetes exhibits strain-level heterogeneity with divergent pathogenic potentials: enterotoxigenic B. fragilis strains which highly secrete the highly pro-inflammatory B. fragilis toxin expand in the UC microenvironment to drive epithelial cleavage and exacerbate mucosal inflammation52, whereas beneficial mucin-degrading species such as B. thetaiotaomicron, critical for mucus production and SCFA generation, are significantly depleted, weakening epithelial barrier integrity.53 Concurrently, Akkermansia muciniphila (Verrucomicrobia), a specialized mucin-degrading bacterium that produces acetate and propionate essential for colonic mucus maintenance, is reduced in UC, resulting in diminished sulfated mucins and increased susceptibility to bacterial translocation.54 Similarly, immunoregulatory Actinobacteria, particularly Bifidobacterium species (eg., B. longum, B. adolescentis) that modulate Th1/Th2 and Th17/Treg balance, are depleted, impairing immune tolerance and Treg mitochondrial fitness.55–57 Lastly, sulfate-reducing Desulfovibrionaceae (Proteobacteria) perpetuate epithelial damage through hydrogen sulfide-mediated cytotoxicity, compromising colonocyte viability and mucosal integrity.58
In conclusion, UC-associated dysbiosis is characterized by a complex shift in the gut microbiota. This involves the depletion of beneficial microbes, such as SCFA-producing Firmicutes and immunoregulatory Actinobacteria; the expansion of pro-inflammatory pathogens from the Proteobacteria and specific Bacteroides strains; and the loss of mucin-degrading Verrucomicrobia. These profound microbial imbalances disrupt critical metabolic processes and immune regulation, ultimately leading to a breakdown in epithelial energy homeostasis, severe barrier dysfunction, and chronic intestinal inflammation.
The Pathogenic Role of the Gut Microbiota in Ulcerative Colitis
Compelling evidence from preclinical models,59 human interventional trials,60 and observational studies collectively validates the gut microbiota as a primary driver of ulcerative colitis pathogenesis, thereby underscoring its potential as a viable therapeutic target.61 Germ-free (GF) mice, which entirely lack a gut microbiota, consistently demonstrate significantly attenuated colitis severity-exhibiting up to 60% lower disease activity indices, reduced epithelial damage, and diminished pro-inflammatory cytokine levels-when challenged with UC-inducing agents such as DSS or trinitrobenzene sulfonic acid (TNBS),62,63 and this attenuated phenotype is reversed upon reconstitution with a complex microbiota.
Beyond compositional shifts, specific strains have been causally implicated in UC pathogenesis. Proteus mirabilis, detected at significantly higher frequencies in UC patients (65.9% vs. controls), exacerbates DSS-induced colitis in mice by downregulating IL-18-mediated mucin production.64 Similarly, specific strains of Streptococcus mutans, isolated more frequently from UC patients, aggravate colitis in murine models through hepatic interferon-γ activation.65 Additionally, Enterococcus faecalis strain OG1RF worsens experimental colitis through a glucosamine phosphotransferase system that enhances metabolic fitness.66
Conversely, FMT from UC patients into GF or antibiotic-treated mice has been shown to recapitulate UC-like phenotypes, including epithelial damage, immune infiltration, and elevated pro-inflammatory cytokine expression,21,67 directly implicating dysbiotic microbiota in the induction of ulcerative colitis. In human interventional trials, FMT achieves clinical remission in approximately 30–50% of patients with refractory UC, with responders exhibiting restored microbial diversity and enrichment of anti-inflammatory taxa such as F. prausnitzii.68,69
Specifically, a recent randomised, double-blind, placebo-controlled trial showed that oral lyophilised FMT following antibiotic conditioning induced corticosteroid-free clinical remission with endoscopic response in 53% of active UC patients at week 8, compared with 15% in the placebo group (difference 38.3%, 95% CI 8.6–68.0; p=0.027), with sustained benefits observed at week 56 in patients continuing maintenance therapy.70
Furthermore, longitudinal cohort studies substantiate this relationship, demonstrating that individuals with pre-existing dysbiosis which is characterized by a reduced abundance of butyrate-producing bacteria exhibit a significantly elevated risk of developing UC, while cross-sectional analyses reveal that disease severity correlates with the degree of microbial disruption.71
Collectively, these findings unequivocally establish the gut microbiota as a central driver in UC pathogenesis, establishing microbial modulation as a viable therapeutic target. This mechanistic foundation necessitates a detailed examination of how TCM herbal compounds engage with and modulate the gut microbial ecosystem to exert therapeutic effects.
Interaction Between Herbal Compounds and Gut Microbiota
Herbal compounds derived from TCM can be systematically classified according to their distinct structural frameworks, which fundamentally determine their bioavailability, metabolic fate, and interactions with gut microbiota. These interactions are critical in mediating the pharmacological activities of TCM, as many compounds undergo microbial biotransformation prior to exerting systemic effects72 (Table 1).
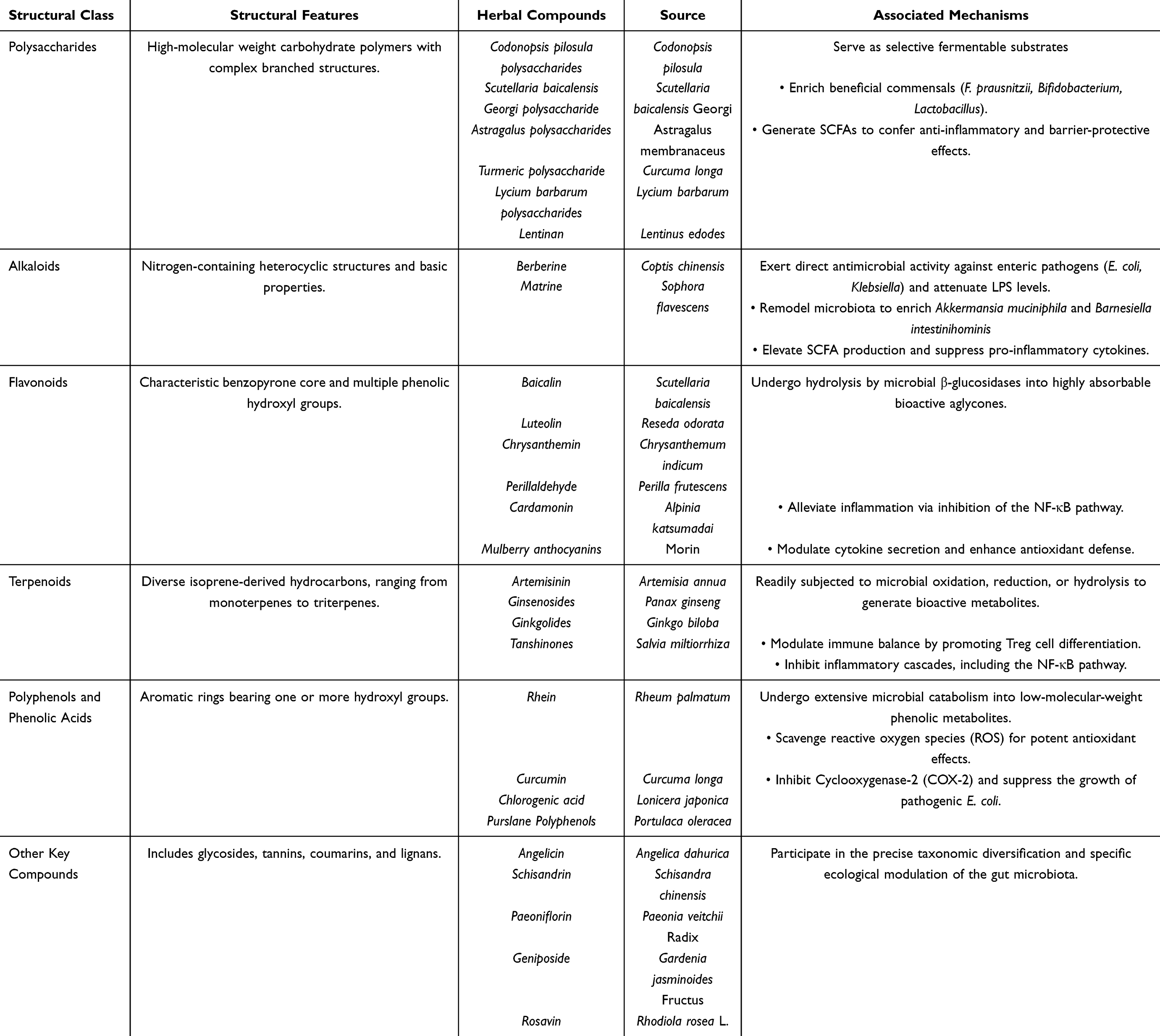 |
Table 1 The Interaction Between Herbal Compounds and UC |
Flavonoids, defined by their characteristic benzopyrone core and multiple phenolic hydroxyl groups,73 represent one of the most abundant classes of bioactive molecules in medicinal plants. Representative compounds include baicalin from Scutellaria baicalensis (Huangqin), luteolin from Reseda odorata (Muxicao), chrysanthemin from Chrysanthemum indicum (Yejuhua), perillaldehyde from Perilla frutescens (Zisu), and cardamonin from Alpinia katsumadai (Caodoukou).74 These compounds primarily exist as glycosides, which must be hydrolyzed by microbial β-glucosidases in the colon to yield absorbable aglycones.75,76 Such enzymatic activation underscores the indispensable role of gut microbiota in flavonoid bioavailability.77 These aglycones have been shown to alleviate UC through inhibition of the NF-κB pathway, modulation of cytokine secretion, and enhancement of antioxidant defense.75
Polysaccharides, characterized as high-molecular-weight carbohydrate polymers with complex branched structures, constitute major bioactive constituents of numerous TCM herbals,78 including Astragalus polysaccharides (APS) from Astragalus membranaceus (Huangqi), ginseng polysaccharides from Panax ginseng (Renshen), lycium barbarum polysaccharides (LBP) from Lycium barbarum (Gouqi), and lentinan from Lentinus edodes (Xianggu).79 Due to their large molecular size, these compounds resist upper gastrointestinal absorption and reach the colon intact, where they serve as selective fermentable substrates for beneficial commensals (eg., Bifidobacterium, Faecalibacterium).80 Specifically, APS increases F. prausnitzii abundance 2.5-fold in fecal samples from UC patients, whereas ginseng and LBP promote proliferation of Bifidobacterium and Lactobacillus species. Microbial fermentation of these substrates generates SCFAs such as butyrate, which confer anti-inflammatory and barrier-protective effects.81
Alkaloids, characterized by nitrogen-containing heterocyclic structures and basic properties, are predominantly isolated from TCM herbals such as Coptis chinensis (Huanglian) and Sophora flavescens (Kushen).82–84 Berberine and matrine represent prototypical examples whose lipophilic nature and relatively small molecular size enable direct antimicrobial activity against enteric pathogens, including E. coli and Klebsiella species frequently overrepresented in dysbiotic UC microbiota.85–87 Beyond direct bactericidal effects, these alkaloids remodel microbial composition, enriching beneficial taxa such as Akkermansia muciniphila and Barnesiella intestinihominis while reducing Helicobacter colonization.88,89 This ecological shift elevates fecal concentrations of SCFAs, including butyrate, acetic acid, and propionic acid, while attenuating harmful LPS levels.90 Consequently, these compounds suppress pro-inflammatory cytokine production, enhance intestinal barrier integrity, and restore mucosal homeostasis.
Terpenoids, comprising diverse isoprene-derived hydrocarbons ranging from monoterpenes to triterpenes, form another major class of TCM metabolites. Typical representatives include artemisinin from Artemisia annua (Qinghao), ginsenosides from Panax ginseng (Renshen), ginkgolides from Ginkgo biloba (Yinxing), and tanshinones from Salvia miltiorrhiza (Danshen).91,92 Owing to their lipophilicity, terpenoids exhibit high intestinal permeability and are readily subjected to microbial oxidation, reduction, or hydrolysis, generating metabolites with enhanced bioavailability and bioactivity.93 These compounds have been shown to modulate immune balance (eg., promoting Treg differentiation) and inhibit inflammatory pathways such as NF-κB, largely through microbially mediated transformation.94,95
Polyphenols and phenolic acids, characterized by aromatic rings bearing one or more hydroxyl groups, encompass a structurally heterogeneous group of metabolites ranging from simple phenolic acids to polymerized tannins. Representative examples include rhein from Rheum palmatum (Dahuang), curcumin from Curcuma longa (Jianghuang), chlorogenic acid from Lonicera japonica (Jinyinhua), and polyphenolic extracts from Portulaca oleracea (Machixian) and Origanum vulgare (Niuzhicao).96 Their partial hydrophilicity allows prolonged retention in the colon, where they undergo extensive microbial catabolism into low-molecular weight phenolic metabolites.97 These biotransformation processes are associated with antioxidant (eg., reactive oxygen species scavenging),98 anti-inflammatory (eg., Cyclooxygenase-2 inhibition),99 and antimicrobial effects (eg., suppression of E. coli growth).100 Consequently, the gut microbiota serves as a pivotal intermediary that converts polyphenols into bioactive forms with systemic health benefits.
Furthermore, other crucial compounds, such as glycosides (eg., salidroside),101 tannins (eg., dimeric proanthocyanidins),102 coumarins (eg., angelicin),103 and lignans (eg., schisandrin),104 also participate in the diversification and precise modulation of gut microbiota.
Collectively, the structural diversity of herbal compounds governs their microbial accessibility, metabolic conversion, and downstream physiological effects. The dynamic interplay between these phytochemicals and the gut microbiome represents a key mechanistic basis for the therapeutic efficacy of TCM in inflammatory and metabolic disorders.
Restoring Multi-Tiered Intestinal Barrier Integrity: The Role of Herbal Compounds and Microbial Metabolites in UC Therapy
A healthy intestinal mucosal barrier is essential to maintain the physiological processes. There are four parts of the complete intestinal mucosal barrier: mechanical, chemical, immune, and biological barriers.105 The pathogenesis of UC is characterized by the progressive disruption of the four-tiered intestinal mucosal barrier, a structural breach that perpetuates chronic inflammation106 (Figure 2 and Supplementary Table S1).
 |
Figure 2 Multidimensional restoration of the intestinal barrier mediated by gut microbiota and their metabolites. Herbal compounds restore intestinal barrier integrity through four hierarchical layers: (i) Biological barrier: Restructuring gut microbiota to enrich beneficial bacteria and suppress pathogens, thereby increasing production of SCFAs, secondary bile acids, and indole derivatives. (ii) Chemical barrier: Metabolites fortify the mucus layer by upregulating MUC2 and trefoil factor 3 (TFF3), enhance secretory IgA (sIgA) production, and stimulate Paneth cells to release antimicrobial proteins via STAT3 signaling. (iii) Mechanical barrier: Metabolites drive assembly of apical junctional complexes, preserving tight junctions (occludin, claudin, ZO-1) and adherens junctions (E-cadherin) to seal paracellular spaces. (iv) Immune barrier: In the lamina propria, compounds and metabolites reprogram immune networks—suppressing STAT3/NF-κB to shift macrophages from M1 to M2 phenotypes, modulating Th17/Treg balance toward Foxp3+ Treg differentiation, stimulating IgA production, and activating AhR on ILC3 cells to induce IL-22 release. Collectively, this microbiome-dependent metabolic network resolves colonic inflammation and restores mucosal homeostasis. |
Targeting the Biological Barrier: Reversing Dysbiosis and Reinstating Colonization Resistance
The intestinal biological barrier comprises a dynamic commensal microbial ecosystem that excludes pathogens to maintain mucosal homeostasis.107 In UC, this microecological equilibrium collapses, manifesting as reduced microbial diversity, depletion of beneficial commensals, and expansion of opportunistic pathogens.108 Herbal compounds intervene within this microenvironment through antimicrobial, prebiotic, and ecological restoration mechanisms to reinstate colonization resistance.17
Alkaloids, characterized by lipophilic nitrogen-containing heterocycles, exhibit potent antimicrobial activity against gut pathobionts while preserving commensal networks.109 In vitro and in vivo studies demonstrate that Berberine effectively suppresses proliferation of UC-associated pathogens E. coli that are aberrantly enriched in dysbiotic microbiota, thereby limiting luminal LPS accumulation.110 Similarly, matrine reduces Helicobacter colonization while enriching beneficial taxa such as Barnesiella intestinihominis, attenuating microbial triggers of mucosal inflammation.88
In contrast to alkaloid bactericidal activity, high-molecular-weight polysaccharides resist upper gastrointestinal digestion, reaching the colon intact to serve as selective fermentable substrates. For instance, polysaccharides derived from both Codonopsis pilosula and Astragalus membranaceus synergistically stimulate the proliferation of Faecalibacterium prausnitzii, a major butyrate-producing commensal that is characteristically depleted in the inflamed mucosa of UC patients.111 Similarly, LBP drive the expansion of keystone genera like Bifidobacterium and Lactobacillus, which selectively ferment these substrates to yield elevated SCFAs.112 These SCFAs serve as obligate bioenergetic substrates for colonocyte proliferation and epithelial repair.113
Beyond targeted taxonomic modulation, herbal interventions restore the macroscopic architecture of the gut microbiome, which in UC is universally compromised by collapsed α-diversity and skewed Firmicutes/Bacteroidetes (F/B) ratios.114 Terpenoids, such as ginsenoside Rg1, restore this macroscopic composition in murine models, increasing α-diversity and normalizing dysregulated F/B ratios.115 Importantly, this architectural restoration is not limited to isolated monomers but is a shared characteristic of broad-spectrum herbal therapies. Recent studies demonstrate that the classic herbal pair Polygonum hydropiper L. and Coptis chinensis comprehensively reverses DSS-induced ecological collapse, restoring α-diversity to near-healthy baseline levels and stabilizing the microbial network116 Similarly, polysaccharides derived from Poria cocos promote microbial richness and diversity, correlating directly with intestinal barrier repair and mucosal inflammation suppression.117
Fortifying Intestinal Chemical Barrier: Enhancing Mucin Networks and Antimicrobial Defenses
The intestinal chemical barrier constitutes a biochemical interface comprising mucins, trefoil factors, secretory IgA (sIgA), AMPs, and electrolytes that segregates the epithelium from luminal microbiota.118 In UC, this defensive framework undergoes multifaceted compromise, manifesting as goblet cell depletion, mucin hyposulfation, and diminished antimicrobial secretions.119 Herbal compounds exert therapeutic effects through distinct yet convergent mechanisms targeting secreted mucin networks, antimicrobial factor production, and transmembrane glycocalyx stability.120
As principal gel-forming constituents, MUC2 and trefoil factor 3 (TFF3) are critical for mucus viscosity and epithelial repair.121,122 Diverse herbal compounds restore mucus barrier integrity by fundamentally reprogramming microbiota-host metabolic interactions. Multi-omics studies demonstrate that Polyphenols such as quercetin remodel gut microbiota to enrich protective metabolites (eg., isovanillic acid, SCFAs), which reduce the expression of MUC2, thereby restoring mature mucin synthesis and secretion.123 Concurrently, these interventions re-establish the mucin-foraging feedback loop: berberine enrich Akkermansia muciniphila, which catabolizes mucin glycans to generate propionate SCFAs and upregulate MUC2 expression in colonic epithelium.124 In addition to SCFA-mediated mechanisms, multi-omics analyses reveal that berberine actively upregulates the colonic expression of TFF3 to accelerate mucosal repair. This upregulation is intricately linked to berberine-induced microbiota remodeling; specifically, berberine significantly enriches beneficial genera such as Megasphaera, whose abundance exhibits a strong positive correlation with enhanced TFF3 transcription and overall barrier fortification.125
Neutralization of luminal pathobionts depends upon abundant sIgA and AMPs secretion.107 Longan pulp polysaccharide promotes the synthesis and secretion of intestinal SIgA through regulating IgA class switch recombination-related factors, facilitating gut homing of IgA+ plasma cells and upregulating polymeric immunoglobulin receptor (pIgR) and secretory component expression, thereby reinforcing the intestinal mucosal barrier function.126 In a distinct regulatory pathway, total glucosides of peony (TGP) and its primary active component, paeoniflorin (PF), reduce the accumulation of microbial indole-3-lactate (ILA) to mitigate UC. As ILA serves as a negative regulator of intestinal epithelial autophagy, its suppression by TGP/PF effectively unleashes the protective autophagic machinery, thereby enhancing epithelial barrier robustness and suppressing inflammatory progression.127
Distinct from secreted mucins, transmembrane mucins (MUC1, MUC13) form the apical glycocalyx and transduce survival signals to underlying epithelium.128 In UC, however, these structures are downregulated; specific herbal interventions effectively rescue this structural deficit. For instance, indigo naturalis-derived components function as potent AhR ligands to stimulate IL-22 production, driving STAT3-dependent transcriptional upregulation of MUC1 and MUC13 in colonic epithelium.129 Concurrently, to prevent the inflammatory shedding of these mucin-anchored epithelial cells, flavonoids like baicalin actively suppress the NF-κB/Caspase-3-mediated apoptotic cascade, thereby preserving the physical cellular anchor required for transmembrane mucin stability.130 Furthermore, specific polyphenols like resveratrol enhance the abundance of A. muciniphila, which in turn augments transmembrane mucin expression,131 collectively stabilizing the epithelial-mucus interface and restoring chemical barrier homeostasis.
Repairing Intestinal Mechanical Barrier: Stabilizing Apical Junctions and Driving Stem Cell-Mediated Restitution
The intestinal mechanical barrier comprises a polarized epithelial monolayer sealed by apical junctional complexes (AJCs), including tight junctions (TJs) containing claudins, occludin, and ZO-1, supplemented by adherens junctions (eg., E-cadherin) and desmosomes.132 In UC, inflammatory mediators drive downregulation of junctional proteins and accelerate epithelial apoptosis, resulting in a “leaky gut” architecture characterized by endotoxemia and mucosal ulceration.133 Herbal compounds counteract this pathology through three convergent mechanisms: upregulating tight junction proteins, attenuating excessive epithelial shedding, and promoting stem cell-mediated restitution.108
TJs undergo dynamic assembly and disassembly to regulate paracellular permeability.132 Herbal compounds fortify this physical barrier by modulating gut microbiota composition and enriching beneficial metabolites that upregulate TJ protein expression. For instance, mulberry anthocyanins ameliorate DSS-induced dysbiosis by reducing the abundance of Escherichia-Shigella and increasing beneficial taxa such as Akkermansia, Muribaculaceae, and Allobaculum.134 These positive microbial shifts lead to the transcriptional upregulation of key tight junction proteins, including occludin, claudin-1, and ZO-1, thereby reducing pathological intestinal permeability. Similarly, polyphenol-rich Portulaca oleracea L. enriches Butyricoccus, Dorea, and Bifidobacterium, which elevate acetate and lactate production.135,136 These metabolites serve as substrates for butyrate-producing bacteria like F. prausnitzii via cross-feeding interactions, resulting in increased colonic butyrate that activates GPR41 on intestinal epithelial cells. This triggers signaling pathways that transcriptionally upregulate occludin and claudin-1 and facilitate their functional assembly at the epithelial membrane, stabilizing junctional networks and restoring barrier integrity.137,138
Beyond reinforcing AJCs, herbal compounds attenuate pathological epithelial loss. In UC, pathobiont overgrowth elevates LPS, triggering epithelial shedding and mucosal denudation.139 For instance, andrographolide minimizes epithelial loss by suppressing LPS-producing Enterobacteriaceae while deactivating NLRP3-mediated pyroptosis in intestinal epithelial cells (IECs).140 Similarly, cardamonin inhibits the RIPK1/RIPK3/MLKL-dependent necroptotic pathway, a destructive form of inflammatory cell death that promotes barrier collapse. By disrupting necrosome formation, it helps maintain epithelial monolayer continuity.141
While halting excessive cell shedding is essential for damage control,142 the ultimate restoration of barrier integrity necessitates active epithelial restitution driven by intestinal stem cells (ISCs).143 Herbal compounds play a pivotal role in re-awakening this regenerative niche. For instance, Rosavin effectively repairs intestinal epithelial injury and restores barrier function by inhibiting inflammatory responses and apoptosis, significantly increasing the populations of Lgr5+ intestinal stem cells, Lyz1+ Paneth cells, and Muc2+ goblet cells. Intestinal organoid studies further demonstrate that Rosavin directly facilitates epithelial cell differentiation and protects against TNF-α-induced damage, with the underlying mechanism involving the upregulation of gene expression associated with cell proliferation and defensin secretion.144 Additionally, notoginsenoside R1 has been shown to upregulates the Wnt/β-catenin cascade, amplifying the Lgr5+ stem cell pool to accelerate mucosal wound healing145 Furthermore, epithelial repair is significantly regulated by IL-22, primarily produced by Innate Lymphoid Cells-Type 3 (ILC3s) in the colon.146 In DSS-induced colitis mice, berberine restores gut microbial homeostasis by increasing probiotics (eg., Lactobacillus/Lactococcus) and decreasing pathobionts such as mouse intestinal Bacteroides, segmented filamentous bacteria, and Enterobacteriaceae. This immunomodulation is coupled with a suppression of ILC1 and an enhancement of ILC3s frequency and IL-22 production in colonic lamina propria lymphocytes, collectively promoting epithelial restitution.147
Reprogramming Intestinal Immune Barrier: Modulating Macrophage Polarization and Th17/Treg Homeostasis
The intestinal immune barrier comprises tissue-resident macrophages, T lymphocytes, B lymphocytes, and innate lymphoid cells (ILCs) distributed across intraepithelial compartments, lamina propria, and Peyer’s patches.148 In UC, chronic mucosal damage is driven by dysregulated macrophage polarization and disruption of the Th17/Treg balance.149 Herbal compounds orchestrate mucosal healing by leveraging microbial biotransformation to reprogram innate macrophage polarization and recalibrate adaptive T-cell responses.
During active UC, pro-inflammatory M1 macrophages expand significantly, releasing cytokines (IL-6, TNF-α) that activate NF-κB signaling, thereby intensifying inflammatory infiltration and tissue damage.150 This immune dysregulation is counteracted via a dual modulatory strategy. For instance, curcumin formulations suppress the TLR4/NF-κB inflammatory cascade while concurrently enriching SCFA-producing microbiota (eg., Lactobacillus).151 The resulting elevation in microbial SCFAs, particularly butyrate, activates GPR43 on macrophages and functions as a HDAC inhibitor, driving polarization toward the tissue-repairing M2 phenotype.152 Crucially, the immunomodulatory effects of herbal compounds are also highly dependent on the specific microbial context. While Akkermansia muciniphila is generally barrier-protective in homeostasis, its over-enrichment in the severely compromised mucosal niche of acute DSS colitis can pathologically trigger M1-like macrophage polarization via its secreted microbial protein Amuc_2172. In this specific pathological context, the administration of puerarin acts as a targeted immunomodulator.153 It significantly attenuates the DSS-mediated over-enrichment of A. muciniphila, effectively inhibiting Amuc_2172-triggered M1 macrophage activation and ultimately halting disease progression. Similarly, Codonopsis pilosula polysaccharide enriches beneficial bacteria (eg., Ligilactobacillus, Akkermansia) to elevate cecal acetic and butyric acid levels, which bind GPR receptors to suppress NLRP3 inflammasome activation, thereby attenuating intestinal inflammation.154
Herbal compounds leverage complex host-microbe co-metabolism to directly intervene in T cell differentiation. Phytochemicals orchestrate mucosal healing by leveraging microbial biotransformation to reprogram innate macrophage polarization and recalibrate adaptive T-cell responses. UC is further characterized by pathological skewing toward RORγt-driven Th17 inflammatory phenotypes alongside functional impairment of FOXP3+ Treg networks.155 Lycium barbarum polysaccharides stimulate Clostridium clusters (eg., C. butyricum) to produce propionate and butyrate, which epigenetically modify naïve CD4+ T cells by promoting acetylation of the Foxp3 promoter locus, thereby stabilizing Treg lineages.156 Additionally, Astragalus polysaccharides restore SCFA-producing microbiota in a microbiota-dependent manner (validated by FMT from treated donors), with elevated SCFAs inhibiting NF-κB activation via TLR4 and HDAC3 pathways to improve Th17/Treg balance.157 Furthermore, the Th17/Treg equilibrium is maintained through cross-talk with ILC3s.158 Indirubin specifically activates AhR signaling within intestinal ILC3s, stimulating barrier-protective IL-22 release while restoring ILC3-dependent suppression of Th17 polarization, thereby shifting mucosal immunity toward Treg-mediated tolerance.159
Translational Challenges and Future Actionable Directions
Translational Challenges: Phytochemical Heterogeneity, Preclinical Model Limitations, and Therapeutic Safety Risks
The clinical translation of microbiota-targeted herbal compounds for UC is constrained by phytochemical heterogeneity and complex host metabolism. Botanical raw materials exhibit inherent compositional variability determined by geographic origin, cultivation conditions, and post-harvest processing160–163 (Figure 3). For example, baicalin concentrations in Scutellaria baicalensis demonstrate significant inter-batch variation contingent on cultivation geography and harvest timing.160,163 Expanding beyond flavonoids, this heterogeneity universally affects other structural classes. The molecular weight and monosaccharide composition of Astragalus membranaceus polysaccharides fluctuate extensively depending on specific extraction protocols, which directly alters their prebiotic consistency and efficacy.164 Similarly, the precise ratios of bioactive ginsenosides in Panax ginseng are profoundly dictated by the plant’s age (eg., 3-year versus 6-year roots) and processing techniques (eg., fresh versus steamed roots).165 This variability is compounded by pharmacokinetic constraints: numerous TCM compounds exhibit limited oral bioavailability yet demonstrate clinical efficacy through microbiome-mediated biotransformation. Consequently, therapeutic outcomes depend on host-specific microbial enzymatic capacity, establishing a bidirectional relationship wherein baseline microbiota composition dictates drug bioactivation while the therapeutic agent concurrently reshapes microbial ecology.166–168 This interplay between variable phytochemical inputs and individualized microbial metabolism renders therapeutic responses unpredictable without rigorous chemical standardization coupled with patient stratification based on microbial profiling.
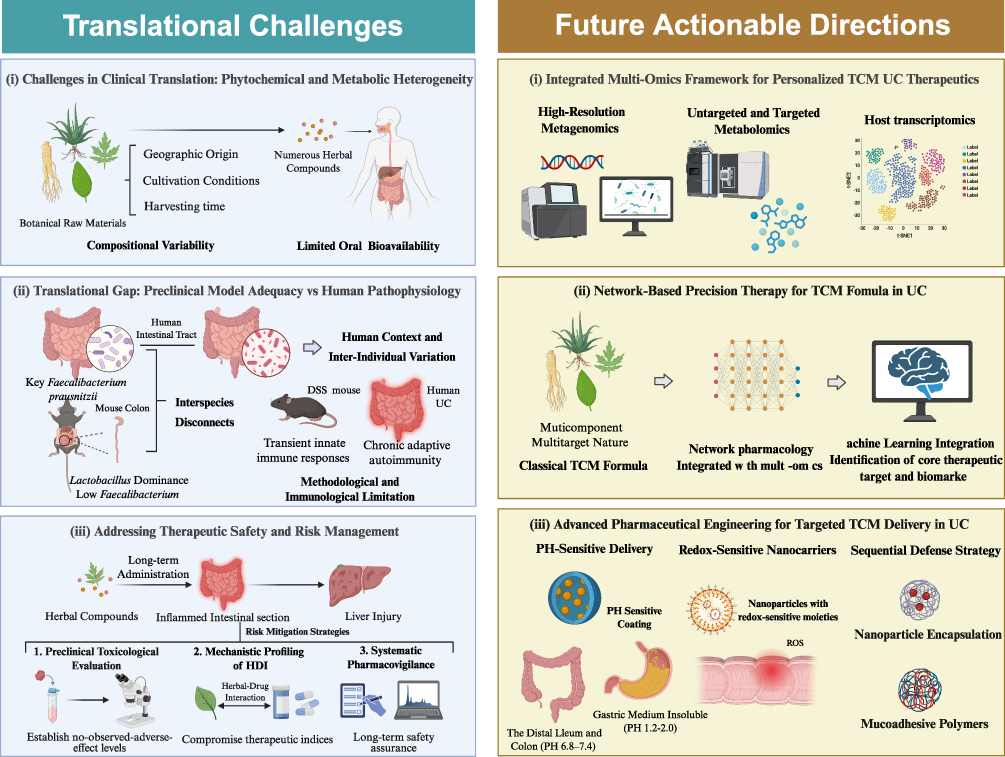 |
Figure 3 Translational challenges and future directions for TCM therapeutics in UC. (Left) Current translational bottlenecks: (i) phytochemical heterogeneity and limited bioavailability; (ii) discrepancies between preclinical models and human pathophysiology, including inter-individual microbiota variation; and (iii) safety concerns regarding long-term hepatotoxicity. (Right) Strategic solutions: (i) multi-omics frameworks for personalized therapeutics; (ii) network pharmacology integrated with machine learning to decode multi-component formulas; and (iii) stimuli-responsive nanocarriers (pH/redox-sensitive) for targeted colonic delivery. |
Despite rapid advances in microbiome-targeted therapeutics for gastrointestinal disease, clinical translation remains substantially impeded by fundamental biological disconnects between preclinical models and human pathophysiology. Marked taxonomic incongruence characterizes these interspecies differences: keystone human butyrate-producers such as Faecalibacterium prausnitzii are essentially absent in murine systems, whereas Lactobacillus species dominate rodent microbiota yet constitute negligible proportions in human gut ecosystems.169,170 These discrepancies are amplified by anatomical and physiological divergence, including differential mucus barrier architecture, distinct gastrointestinal transit kinetics, and divergent anatomical segregation of microbial fermentation—cecum-dominant in rodents versus distributed colonic fermentation in humans.169,171 Methodologically, acute chemical injury models (eg., DSS) induce transient innate immune responses that fail to recapitulate the chronic, relapsing-remitting adaptive autoimmunity characteristic of human UC.172 This translational gap is further exacerbated by the “hygiene problem” inherent to Specific Pathogen-Free housing, wherein artificially restricted microbial exposure generates immunologically naïve animals incapable of mimicking the antigen-experienced human immune system, potentially yielding artifactual efficacy signals reflecting toxicity resolution rather than true pharmacological modulation of chronic dysbiosis.173,174 Transitioning to clinical contexts, these biological limitations converge with profound inter-individual variation in baseline gut microbiome composition and enzymatic capacity—specifically heterogeneous β-glucuronidase activity generating distinct patient metabotypes—which emerges as the primary determinant of divergent therapeutic outcomes, rendering universal treatment paradigms inherently inadequate.175–177 This interplay between preclinical physiological inadequacy and patient-specific microbial enzymatic variation constitutes a pervasive bottleneck constraining clinical translation across microbiome-based therapeutics, demanding rigorous model standardization coupled with multi-omics-based stratification and mechanism-driven experimental frameworks to ensure therapeutic predictability.
Therapeutic safety requires rigorous scrutiny, particularly given documented contraindications and hepatotoxic risks associated with specific botanical compounds. Indigo naturalis, despite efficacy in refractory UC mucosal healing, induces dose-dependent hepatic dysfunction and pulmonary arterial hypertension upon chronic administration.178,179 Mechanistically, macrophage-mediated translocation of insoluble indirubin from inflamed intestine to hepatic tissue precipitates oxidative stress-driven liver injury.178 Similarly, baicalin-containing Scutellaria baicalensis formulations have been implicated in idiosyncratic drug-induced liver injury.180 Mitigating these risks necessitates standardized preclinical toxicological evaluation following to establish no-observed-adverse-effect levels.181 Additionally, mechanistic profiling of pharmacokinetic herb–drug interactions (HDI) is imperative given the polypharmacy prevalent in UC management; for instance, curcumin-mediated inhibition of intestinal BCRP/ABCG2 efflux transporters can substantially alter systemic exposure to conventional substrates such as sulfasalazine, potentially compromising therapeutic indices.182 Long-term safety assurance further requires structured pharmacovigilance, such as employing validated hepatotoxicity assessment algorithms for objective causality adjudication of suspected herb-induced liver injury.183 Integration of predictive toxicology, transporter-level HDI characterization, and systematic pharmacovigilance thus enables evidence-based risk stratification, ensuring that personalized therapeutic strategies are balanced against individualized safety profiles within precision medicine frameworks for UC.
Future Actionable Directions: Towards Precision Microbiome Therapies
To address the complexity of gut microbiota and phytochemical constituents constraining empirical TCM approaches, future UC therapeutics must adopt integrated multi-omics frameworks (Figure 3). Moving beyond descriptive microbiome surveys, research must systematically resolve the “active ingredient-microbiome-receptor” axis. High-resolution metagenomics enables precise delineation of patient- or syndrome-specific dysbiotic signatures establishing baseline microbial architectures.25 Layered upon this, untargeted and targeted metabolomics capture dynamic functional outputs of microbial communities, specifically tracking TCM-microbe cometabolites. For instance, metabolomic profiling has traced microbial biotransformation of botanical compounds into SCFAs and tryptophan derivatives such as xanthurenic acid, which subsequently activate anti-inflammatory host receptors including the AhR.184,185 Host transcriptomics completes this framework by providing phenotypic readouts mapping microbial cometabolite modulation of mucosal healing pathways and immune gene networks within intestinal epithelium.186 Mathematical integration of metagenomic, metabolomic, and transcriptomic datasets enables predictive mechanistic modeling beyond associative correlations, facilitating truly personalized TCM interventions based on patient-specific multi-omic signatures.
The multicomponent, multitarget nature of TCM challenges conventional “one drug, one target” paradigms while aligning with the complex pathogenesis of UC. Network pharmacology integrated with multi-omics enables a shift toward network-based precision therapy.187,188 Systems-level analyses reveal that classical formulas orchestrate synergistic modulation across multiple pathological pathways rather than isolated targets. Furthermore, machine learning integration refines core therapeutic target and biomarker identification, facilitating alignment of herbal compounds with distinct UC endotypes.189,190 This data-driven framework transitions TCM from empirical practice to quantifiable precision medicine, advancing personalized therapeutic strategies.
Poor aqueous solubility and extensive first-pass metabolism of bioactive compounds (eg., curcumin, berberine), coupled with complex drug-microbiota metabolic networks, constrain therapeutic development and clinical application. To circumvent these limitations, advanced pharmaceutical engineering has developed targeted delivery systems that bypass the harsh upper gastrointestinal environment to achieve localized, high-concentration release at sites of colonic inflammation. These platforms predominantly employ stimuli-responsive mechanisms tailored to the physiological hallmarks of UC. pH-sensitive coatings (eg., methacrylic acid copolymers) remain insoluble in gastric medium (pH 1.2–2.0) while dissolving in the neutral-to-alkaline environment of the distal ileum and colon (pH 6.8–7.4), ensuring intestinal targeting.191,192 Furthermore, nanoparticles functionalized with redox-sensitive moieties (eg., diselenide or thioketal linkers) exploit elevated reactive oxygen species levels in inflamed mucosa to enable site-specific payload release, maximizing bioavailability at sites of acute oxidative stress.193,194 The integration of stimuli-responsive nanocarriers with other materials, such as cell membrane nanomaterials and enzymatically degradable polysaccharide-based hydrogels (eg., pectin or chitosan), establishes a synergistic, multi-level protective strategy. This sequential defense system first shields the active compounds from enzymatic degradation in the upper gastrointestinal tract; it then utilizes the specific enzymes produced by the colonic microbiome to precisely degrade the hydrogel matrix, thereby ensuring targeted payload release within the microbial-rich inflammatory microenvironment of the colon.195–197 Collectively, these engineering strategies transform unstable phytochemicals into precision-targeted nanomedicines, fundamentally addressing bioavailability and site-specificity challenges in UC management.
Conclusion
This review systematically elucidates the multifaceted mechanisms by which herbal compounds alleviate UC symptoms by modulating the gut microbiota—promoting beneficial commensals, suppressing opportunistic pathogens, and restoring microbial diversity—thereby orchestrating improvements across mechanical, chemical, immune, and biological barrier functions. However, it is crucial to acknowledge that the current evidence base derives predominantly from preclinical models, necessitating cautious interpretation of causality until validated by rigorous human trials. To bridge this translational gap, future research must prioritize chemical standardization of raw materials, advanced pharmaceutical engineering to enhance bioavailability and colonic targeting, and the integration of multi-omics profiling to stratify patients according to baseline microbiome signatures and metabolic capacity, ultimately transforming empirical TCM application into precision microbiota-targeted therapeutics for UC.
Abbreviations
AhR, Aryl hydrocarbon receptor; AIEC, Adherent-invasive Escherichia coli; AJCs, Apical junctional complexes; AMPs, Antimicrobial peptides; APS, Astragalus polysaccharides; BCFAs, Branched-chain fatty acids; DSS, Dextran sulfate sodium; ER, Endoplasmic reticulum; F/B, Firmicutes/Bacteroidetes; FMT, Fecal microbiota transplantation; GF, Germ-free; GI, Gastrointestinal; GM, Gut microbiota; GPR43, G-protein-coupled receptor 43; HDAC, Histone deacetylase; HDI, Herb-drug interactions; IBD, Inflammatory bowel disease; IECs, Intestinal epithelial cells; IL-6, Interleukin-6; IL-10, Interleukin-10; ILA, indole-3-lactate; ILCs, Innate lymphoid cells; ILC3s, Group 3 innate lymphoid cells; IPA, Indole-3-propionic acid; ISCs, Intestinal stem cells; LBP, Lycium barbarum polysaccharides; LPS, Lipopolysaccharide; NF-κB, Nuclear transcription factor-κB; NLRP3, NOD-like Receptor Pyrin domain containing 3; pIgR, Polymeric immunoglobulin receptor; PSA, Polysaccharide A; SCFAs, Short-chain fatty acids; sIgA, Secretory IgA; TCM, Traditional Chinese Medicine; TFF3, Trefoil factor 3; Th17, T-helper 17; TJs, Tight junctions; TLR4, Toll-like receptor 4; TNBS, Trinitrobenzene sulfonic acid; TNF-α, Tumor necrosis factor-α; Tregs, Regulatory T cells; UC, Ulcerative colitis; ZO-1, Zonula occludens-1.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this study.
Author Contributions
Junyi Chen: Conceptualization, Literature review, Investigation, Writing – original draft, Writing – review and editing. Jiaxv Chen: Conceptualization, Literature review, Investigation, Writing – original draft, Writing – review and editing. Zhenyi Wang.: Supervision, Validation, Writing – review and editing, Funding acquisition. All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agreed to be accountable for all aspects of the work.
Funding
This work was fully supported by the National Natural Science Foundation of China (No. 82274531) and Shanghai Municipal Health Commission (No. 202540035).
Disclosure
The authors declare that they have no competing interests.
References
1. Kobayashi T, Siegmund B, Le Berre C, et al. Ulcerative colitis. Nat Rev Dis Primers. 2020;6(1):74. doi:10.1038/s41572-020-0205-x
2. Ananthakrishnan AN. Epidemiology and risk factors for IBD. Nat Rev Gastroenterol Hepatol. 2015;12(4):205–23. doi:10.1038/nrgastro.2015.34
3. Dharni K, Singh A, Sharma S, et al. Trends of inflammatory bowel disease from the Global Burden of Disease Study (1990-2019). Indian J Gastroenterol. 2024;43(1):188–198. doi:10.1007/s12664-023-01430-z
4. Ng SC, Shi HY, Hamidi N, et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet. 2017;390(10114):2769–2778. doi:10.1016/S0140-6736(17)32448-0
5. Jabłońska B, Mrowiec S. Nutritional Status and Its Detection in Patients with Inflammatory Bowel Diseases. Nutrients. 2023;15(8):1991. doi:10.3390/nu15081991
6. Gros B, Kaplan GG. Ulcerative Colitis in Adults: a Review. JAMA. 2023;330(10):951–965. doi:10.1001/jama.2023.15389
7. Abu-Freha N, Cohen B, Gordon M, et al. Colorectal cancer among inflammatory bowel disease patients: risk factors and prevalence compared to the general population. Front Med Lausanne. 2023;10:1225616. doi:10.3389/fmed.2023.1225616
8. Olén O, Erichsen R, Sachs MC, et al. Colorectal cancer in ulcerative colitis: a Scandinavian population-based cohort study. Lancet. 2020;395(10218):123–131. doi:10.1016/S0140-6736(19)32545-0
9. Kato S, Teixeira BC, Laurent T, et al. Treatment Patterns and Economic Burden of Ulcerative Colitis in Japan: a Retrospective Claims Analysis. Adv Ther. 2025;42(3):1435–1447. doi:10.1007/s12325-024-03096-1
10. Le Berre C, Honap S, Peyrin-Biroulet L. Ulcerative colitis. Lancet. 2023;402(10401):571–584. doi:10.1016/S0140-6736(23)00966-2
11. Abbas A, Di Fonzo DMP, Wetwittayakhlang P, Al-Jabri R, Lakatos PL, Bessissow T. Management of ulcerative colitis: where are we at and where are we heading? Expert Rev Gastroenterol Hepatol. 2024;18(10):567–574. doi:10.1080/17474124.2024.2422370
12. Guo M, Wang X. Pathological mechanism and targeted drugs of ulcerative colitis: a review. Medicine. 2023;102(37):e35020. doi:10.1097/MD.0000000000035020
13. Foppa C, Rizkala T, Repici A, Hassan C, Spinelli A. Microbiota and IBD: current knowledge and future perspectives. Dig Liver Dis. 2024;56(6):911–922. doi:10.1016/j.dld.2023.11.015
14. Danilova NA, Abdulkhakov SR, Grigoryeva TV, et al. Markers of dysbiosis in patients with ulcerative colitis and Crohn’s disease. Ter Arkh. 2019;91(4):17–24. doi:10.26442/00403660.2019.04.000211
15. Do KH, Ko SH, Kim KB, Seo K, Lee WK. Comparative Study of Intestinal Microbiome in Patients with Ulcerative Colitis and Healthy Controls in Korea. Microorganisms. 2023;11(11):2750. doi:10.3390/microorganisms11112750
16. Khorsand B, Asadzadeh Aghdaei H, Nazemalhosseini-Mojarad E, Nadalian B, Nadalian B, Houri H. Overrepresentation of Enterobacteriaceae and Escherichia coli is the major gut microbiome signature in Crohn’s disease and ulcerative colitis; a comprehensive metagenomic analysis of IBDMDB datasets. Front Cell Infect Microbiol. 2022;12:1015890. doi:10.3389/fcimb.2022.1015890
17. Xiong T, Zheng X, Zhang K, et al. Ganluyin ameliorates DSS-induced ulcerative colitis by inhibiting the enteric-origin LPS/TLR4/NF-κB pathway. J Ethnopharmacol. 2022;289:115001. doi:10.1016/j.jep.2022.115001
18. Huang X, Lin R, Liu H, et al. Resatorvid (TAK-242) Ameliorates Ulcerative Colitis by Modulating Macrophage Polarization and T Helper Cell Balance via TLR4/JAK2/STAT3 Signaling Pathway. Inflammation. 2024;47(6):2108–2128. doi:10.1007/s10753-024-02028-z
19. Ratajczak W, Rył A, Mizerski A, Walczakiewicz K, Sipak O, Laszczyńska M. Immunomodulatory potential of gut microbiome-derived short-chain fatty acids (SCFAs). Acta Biochim Pol. 2019;66(1):1–12. doi:10.18388/abp.2018_2648
20. Peña-Rodríguez M, Vega-Magaña N, García-Benavides L, et al. Butyrate administration strengthens the intestinal epithelium and improves intestinal dysbiosis in a cholestasis fibrosis model. J Appl Microbiol. 2022;132(1):571–583. doi:10.1111/jam.15135
21. Knudsen LA, Zachariassen LS, Strube ML, et al. Assessment of the Inflammatory Effects of Gut Microbiota from Human Twins Discordant for Ulcerative Colitis on Germ-free Mice. Comp Med. 2024;74(2):55–69. doi:10.30802/AALAS-CM-23-000065
22. Lima SF, Gogokhia L, Viladomiu M, et al. Transferable Immunoglobulin A-Coated Odoribacter splanchnicus in Responders to Fecal Microbiota Transplantation for Ulcerative Colitis Limits Colonic Inflammation. Gastroenterology. 2022;162(1):166–178. doi:10.1053/j.gastro.2021.09.061
23. Uygun A, Ozturk K, Demirci H, et al. Fecal microbiota transplantation is a rescue treatment modality for refractory ulcerative colitis. Medicine. 2017;96(16):e6479. doi:10.1097/MD.0000000000006479
24. Wang M, Fu R, Xu D, et al. Traditional Chinese Medicine: a promising strategy to regulate the imbalance of bacterial flora, impaired intestinal barrier and immune function attributed to ulcerative colitis through intestinal microecology. J Ethnopharmacol. 2024;318(Pt A):116879. doi:10.1016/j.jep.2023.116879
25. Li MX, Li MY, Lei JX, et al. Huangqin decoction ameliorates DSS-induced ulcerative colitis: role of gut microbiota and amino acid metabolism, mTOR pathway and intestinal epithelial barrier. Phytomedicine. 2022;100:154052. doi:10.1016/j.phymed.2022.154052
26. Tang X, Wang X, Feng B, Cui Y. Simultaneous Determination of Eight Bioactive Components of Huangqin Decoction in Rat Plasma by Ultra-high-performance Liquid Chromatography-tandem Mass Spectrometry and Its Application to Comparative Pharmacokinetic Study of Huangqin Decoction in Ulcerative Colitis and Normal Rats. J Sep Sci. 2025;48(4):e70140. doi:10.1002/jssc.70140
27. Li R, Chi H, Liao X, Cen S, Zou Y. The Glabridin from Huangqin Decoction Prevents the Development of Ulcerative Colitis into Colitis-Associated Colorectal Cancer by Modulating MMP1/MMP3 Activity. Int Immunopharmacol. 2024;135:112262. doi:10.1016/j.intimp.2024.112262
28. Dai Y, Lu Q, Li P, et al. Xianglian Pill attenuates ulcerative colitis through TLR4/MyD88/NF-κB signaling pathway. J Ethnopharmacol. 2023;300:115690. doi:10.1016/j.jep.2022.115690
29. Wei YY, Fan YM, Ga Y, Zhang YN, Han JC, Hao ZH. Shaoyao decoction attenuates DSS-induced ulcerative colitis, macrophage and NLRP3 inflammasome activation through the MKP1/NF-κB pathway. Phytomedicine. 2021;92:153743. doi:10.1016/j.phymed.2021.153743
30. Zeng L, Yang T, Yang K, et al. Curcumin and Curcuma longa Extract in the Treatment of 10 Types of Autoimmune Diseases: a Systematic Review and Meta-Analysis of 31 Randomized Controlled Trials. Front Immunol. 2022;13:896476. doi:10.3389/fimmu.2022.896476
31. Jandhyala SM, Talukdar R, Subramanyam C, Vuyyuru H, Sasikala M, Nageshwar Reddy D. Role of the normal gut microbiota. World J Gastroenterol. 2015;21(29):8787–8803. doi:10.3748/wjg.v21.i29.8787
32. Chen Y, Xiao L, Zhou M, Zhang H. The microbiota: a crucial mediator in gut homeostasis and colonization resistance. Front Microbiol. 2024;15:1417864. doi:10.3389/fmicb.2024.1417864
33. Krautkramer KA, Fan J, Bäckhed F. Gut microbial metabolites as multi-kingdom intermediates. Nat Rev Microbiol. 2021;19(2):77–94. doi:10.1038/s41579-020-0438-4
34. Zhang Y, Sun D, Zhao X, et al. Bacteroides fragilis prevents aging-related atrial fibrillation in rats via regulatory T cells-mediated regulation of inflammation. Pharmacol Res. 2022;177:106141. doi:10.1016/j.phrs.2022.106141
35. Wolf PG, Devendran S, Doden HL, et al. Berberine alters gut microbial function through modulation of bile acids. BMC Microbiol. 2021;21(1):24. doi:10.1186/s12866-020-02020-1
36. Tang Y, Fu A, Wang L, Ge Q. Microbiota-dependent metabolites - New engine for T cell warriors. Gut Microbes. 2025;17(1):2523815. doi:10.1080/19490976.2025.2523815
37. Meng D, Sommella E, Salviati E, et al. Indole-3-lactic acid, a metabolite of tryptophan, secreted by Bifidobacterium longum subspecies infantis is anti-inflammatory in the immature intestine. Pediatr Res. 2020;88(2):209–217. doi:10.1038/s41390-019-0740-x
38. Nie X, Li Q, Ji H, et al. Bifidobacterium longum NSP001-derived extracellular vesicles ameliorate ulcerative colitis by modulating T cell responses in gut microbiota-(in)dependent manners. NPJ Biofilms Microbiomes. 2025;11(1):27. doi:10.1038/s41522-025-00663-4
39. Yang J, Yang H. Antibacterial Activity of Bifidobacterium breve Against Clostridioides difficile. Front Cell Infect Microbiol. 2019;9:288. doi:10.3389/fcimb.2019.00288
40. Hu Y, Jin X, Gao F, et al. Selenium-enriched Bifidobacterium longum DD98 effectively ameliorates dextran sulfate sodium-induced ulcerative colitis in mice. Front Microbiol. 2022;13:955112. doi:10.3389/fmicb.2022.955112
41. Wang J, Zhao X, Li X, Jin X. Akkermansia muciniphila: a deworming partner independent of type 2 immunity. Gut Microbes. 2024;16(1):2338947. doi:10.1080/19490976.2024.2338947
42. Mukhopadhya I, Hansen R, El-Omar EM, Hold GL. IBD-what role do Proteobacteria play? Nat Rev Gastroenterol Hepatol. 2012;9(4):219–230. doi:10.1038/nrgastro.2012.14
43. Ramos GP, Papadakis KA. Mechanisms of Disease: inflammatory Bowel Diseases. Mayo Clin Proc. 2019;94(1):155–165. doi:10.1016/j.mayocp.2018.09.013
44. Deleu S, Machiels K, Raes J, Verbeke K, Vermeire S. Short chain fatty acids and its producing organisms: an overlooked therapy for IBD? EBioMedicine. 2021;66:103293. doi:10.1016/j.ebiom.2021.103293
45. Martin-Gallausiaux C, Marinelli L, Blottière HM, Larraufie P, Lapaque N. SCFA: mechanisms and functional importance in the gut. Proc Nutr Soc. 2021;80(1):37–49. doi:10.1017/S0029665120006916
46. Otte ML, Lama Tamang R, Papapanagiotou J, Ahmad R, Dhawan P, Singh AB. Mucosal healing and inflammatory bowel disease: therapeutic implications and new targets. World J Gastroenterol. 2023;29(7):1157–1172. doi:10.3748/wjg.v29.i7.1157
47. Nie K, Ma K, Luo W, et al. Roseburia intestinalis: a Beneficial Gut Organism From the Discoveries in Genus and Species. Front Cell Infect Microbiol. 2021;11:757718. doi:10.3389/fcimb.2021.757718
48. Wang H, Huang W, Pan X, et al. Quzhou Aurantii Fructus Flavonoids Ameliorate Inflammatory Responses, Intestinal Barrier Dysfunction in DSS-Induced Colitis by Modulating PI3K/AKT Signaling Pathway and Gut Microbiome. J Inflamm Res. 2025;18:1855–1874. doi:10.2147/JIR.S500014
49. Nadalian B, Nadalian B, Zali MR, Yadegar A. Outer Membrane Vesicles Derived from Adherent-Invasive Escherichia coli Induce Inflammatory Response and Alter the Gene Expression of Junction-Associated Proteins in Human Intestinal Epithelial Cells. Can J Infect Dis Med Microbiol. 2024;2024:2701675. doi:10.1155/2024/2701675
50. Phan J, Jain S, Nijkamp JF, et al. Gut health predictive indices linking gut microbiota dysbiosis with healthy state, mild gut discomfort, and inflammatory bowel disease phenotypes using gut microbiome profiling. Microbiol Spectr. 2025;13(8):e0027125. doi:10.1128/spectrum.00271-25
51. Alqudah A, Qnais E, Gammoh O, et al. Exploring Scopoletin’s Therapeutic Efficacy in DSS-Induced Ulcerative Colitis: insights into Inflammatory Pathways, Immune Modulation, and Microbial Dynamics. Inflammation. 2025;48(2):575–589. doi:10.1007/s10753-024-02048-9
52. Jasemi S, Molicotti P, Fais M, Cossu I, Simula ER, Sechi LA. Biological Mechanisms of Enterotoxigenic Bacteroides fragilis Toxin: linking Inflammation, Colorectal Cancer, and Clinical Implications. Toxins. 2025;17(6):305. doi:10.3390/toxins17060305
53. Liu J, Lin H, Cao M, et al. Shifts and importance of viable bacteria in treatment of DSS-induced ulcerative colitis mice with FMT. Front Cell Infect Microbiol. 2023;13:1124256. doi:10.3389/fcimb.2023.1124256
54. Zheng M, Han R, Yuan Y, et al. The role of Akkermansia muciniphila in inflammatory bowel disease: current knowledge and perspectives. Front Immunol. 2022;13:1089600. doi:10.3389/fimmu.2022.1089600
55. Li XH, Liu L, Wu WZ. Trans-Anethole Alleviates DSS-Induced Ulcerative Colitis by Remodeling the Intestinal Flora to Regulate Immunity and Bile Acid Metabolism. Mediators Inflamm. 2023;2023:4188510. doi:10.1155/2023/4188510
56. Mirsepasi-Lauridsen HC. Therapy Used to Promote Disease Remission Targeting Gut Dysbiosis, in UC Patients with Active Disease. J Clin Med. 2022;11(24):7472. doi:10.3390/jcm11247472
57. Dong J, Ping L, Cao T, et al. Immunomodulatory effects of the Bifidobacterium longum BL-10 on lipopolysaccharide-induced intestinal mucosal immune injury. Front Immunol. 2022;13:947755. doi:10.3389/fimmu.2022.947755
58. Leonardi I, Gerstgrasser A, Schmidt TSB, et al. Preventive Trichuris suis ova (TSO) treatment protects immunocompetent rabbits from DSS colitis but may be detrimental under conditions of immunosuppression. Sci Rep. 2017;7(1):16500. doi:10.1038/s41598-017-16287-4
59. Galipeau HJ, Caminero A, Turpin W, et al. Novel Fecal Biomarkers That Precede Clinical Diagnosis of Ulcerative Colitis. Gastroenterology. 2021;160(5):1532–1545. doi:10.1053/j.gastro.2020.12.004
60. Wang X, Peng J, Cai P, et al. The emerging role of the gut microbiota and its application in inflammatory bowel disease. Biomed Pharmacother. 2024;179:117302. doi:10.1016/j.biopha.2024.117302
61. Santana PT, Rosas SLB, Ribeiro BE, Marinho Y, de Souza HSP. Dysbiosis in Inflammatory Bowel Disease: pathogenic Role and Potential Therapeutic Targets. Int J Mol Sci. 2022;23(7):3464. doi:10.3390/ijms23073464
62. Maslowski KM, Vieira AT, Ng A, et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009;461(7268):1282–1286. doi:10.1038/nature08530
63. Camba-Gómez M, Gualillo O, Conde-Aranda J. New Perspectives in the Study of Intestinal Inflammation: focus on the Resolution of Inflammation. Int J Mol Sci. 2021;22(5):2605. doi:10.3390/ijms22052605
64. Jiang Z, Li P, Qiu K, et al. Proteus mirabilis exacerbates ulcerative colitis by inhibiting mucin production. Front Microbiol. 2025;16:1556953. doi:10.3389/fmicb.2025.1556953
65. Kojima A, Nakano K, Wada K, et al. Infection of specific strains of Streptococcus mutans, oral bacteria, confers a risk of ulcerative colitis. Sci Rep. 2012;2:332. doi:10.1038/srep00332
66. Fan TJ, Goeser L, Lu K, Faith JJ, Hansen JJ. Enterococcus faecalis Glucosamine Metabolism Exacerbates Experimental Colitis. Cell Mol Gastroenterol Hepatol. 2021;12(4):1373–1389. doi:10.1016/j.jcmgh.2021.06.017
67. Yang Y, Zheng X, Wang Y, et al. Human Fecal Microbiota Transplantation Reduces the Susceptibility to Dextran Sulfate Sodium-Induced Germ-Free Mouse Colitis. Front Immunol. 2022;13:836542. doi:10.3389/fimmu.2022.836542
68. Paramsothy S, Kamm MA, Kaakoush NO, et al. Multidonor intensive faecal microbiota transplantation for active ulcerative colitis: a randomised placebo-controlled trial. Lancet. 2017;389(10075):1218–1228. doi:10.1016/S0140-6736(17)30182-4
69. Imdad A, Pandit NG, Zaman M, et al. Fecal transplantation for treatment of inflammatory bowel disease. Cochrane Database Syst Rev. 2023;4(4):CD012774. doi:10.1002/14651858.CD012774.pub3
70. Haifer C, Paramsothy S, Kaakoush NO, et al. Lyophilised oral faecal microbiota transplantation for ulcerative colitis (LOTUS): a randomised, double-blind, placebo-controlled trial. Lancet Gastroenterol Hepatol. 2022;7(2):141–151. doi:10.1016/S2468-1253(21)00400-3
71. Lemay JA, Yamamoto M, Kroezen Z, et al. Lyophilized fecal short-chain fatty acid and electrolyte determination by capillary electrophoresis with indirect UV detection for assessment of pediatric inflammatory bowel disease. J Pharm Biomed Anal. 2021;192:113658. doi:10.1016/j.jpba.2020.113658
72. Cao SY, Ye SJ, Wang WW, Wang B, Zhang T, Pu YQ. Progress in active compounds effective on ulcerative colitis from Chinese medicines. Chin J Nat Med. 2019;17(2):81–102. doi:10.1016/S1875-5364(19)30012-3
73. Chanu NR, Gogoi P, Barbhuiya PA, Dutta PP, Pathak MP, Sen S. Natural Flavonoids as Potential Therapeutics in the Management of Diabetic Wound: a Review. Curr Top Med Chem. 2023;23(8):690–710. doi:10.2174/1568026623666230419102140
74. Tsou LK, Lara-Tejero M, RoseFigura J, et al. Antibacterial Flavonoids from Medicinal Plants Covalently Inactivate Type III Protein Secretion Substrates. J Am Chem Soc. 2016;138(7):2209–2218. doi:10.1021/jacs.5b11575
75. Gade A, Kumar MS. Gut microbial metabolites of dietary polyphenols and their potential role in human health and diseases. J Physiol Biochem. 2023;79(4):695–718. doi:10.1007/s13105-023-00981-1
76. Murota K, Nakamura Y, Uehara M. Flavonoid metabolism: the interaction of metabolites and gut microbiota. Biosci Biotechnol Biochem. 2018;82(4):600–610. doi:10.1080/09168451.2018.1444467
77. Amaretti A, Raimondi S, Leonardi A, Quartieri A, Rossi M. Hydrolysis of the rutinose-conjugates flavonoids rutin and hesperidin by the gut microbiota and bifidobacteria. Nutrients. 2015;7(4):2788–2800. doi:10.3390/nu7042788
78. Chen N, Jiang T, Xu J, et al. The relationship between polysaccharide structure and its antioxidant activity needs to be systematically elucidated. Int J Biol Macromol. 2024;270(Pt 2):132391. doi:10.1016/j.ijbiomac.2024.132391
79. Xie JH, Jin ML, Morris GA, et al. Advances on Bioactive Polysaccharides from Medicinal Plants. Crit Rev Food Sci Nutr. 2016;56(1):S60–84. doi:10.1080/10408398.2015.1069255
80. PubMed. Lycium barbarum polysaccharide alleviates DSS-induced chronic ulcerative colitis by restoring intestinal barrier function and modulating gut microbiota. Available from: https://pubmed.ncbi.nlm.nih.gov/38061697/.
81. Liang J, Zhang M, Wang X, et al. Edible fungal polysaccharides, the gut microbiota, and host health. Carbohydr Polym. 2021;273:118558. doi:10.1016/j.carbpol.2021.118558
82. Qu Z, Wang W, Adelson DL. Chromosomal level genome assembly of medicinal plant Sophora flavescens. Sci Data. 2023;10(1):572. doi:10.1038/s41597-023-02490-8
83. Zheng Z, Hu J, Sun D, et al. Structural and functional properties of common natural organic cations. Phytomedicine. 2025;141:156662. doi:10.1016/j.phymed.2025.156662
84. Liu Y, Wang B, Shu S, et al. Analysis of the Coptis chinensis genome reveals the diversification of protoberberine-type alkaloids. Nat Commun. 2021;12(1):3276. doi:10.1038/s41467-021-23611-0
85. Li J, Feng S, Liu X, et al. Effects of Traditional Chinese Medicine and its Active Ingredients on Drug-Resistant Bacteria. Front Pharmacol. 2022;13:837907. doi:10.3389/fphar.2022.837907
86. He K, Hu Y, Ma H, et al. Rhizoma Coptidis alkaloids alleviate hyperlipidemia in B6 mice by modulating gut microbiota and bile acid pathways. Biochim Biophys Acta. 2016;1862(9):1696–1709. doi:10.1016/j.bbadis.2016.06.006
87. Gasmi A, Asghar F, Zafar S, et al. Berberine: pharmacological Features in Health, Disease and Aging. Curr Med Chem. 2024;31(10):1214–1234. doi:10.2174/0929867330666230207112539
88. Yao H, Shi Y, Yuan J, Sa R, Chen W, Wan X. Matrine protects against DSS-induced murine colitis by improving gut barrier integrity, inhibiting the PPAR-α signaling pathway, and modulating gut microbiota. Int Immunopharmacol. 2021;100:108091. doi:10.1016/j.intimp.2021.108091
89. Huang J, Xu H. Matrine: bioactivities and Structural Modifications. Curr Top Med Chem. 2016;16(28):3365–3378. doi:10.2174/1568026616666160506131012
90. Yan S, Chang J, Hao X, et al. Berberine regulates short-chain fatty acid metabolism and alleviates the colitis-associated colorectal tumorigenesis through remodeling intestinal flora. Phytomedicine. 2022;102:154217. doi:10.1016/j.phymed.2022.154217
91. Wang Q, Zhao X, Jiang Y, Jin B, Wang L. Functions of Representative Terpenoids and Their Biosynthesis Mechanisms in Medicinal Plants. Biomolecules. 2023;13(12):1725. doi:10.3390/biom13121725
92. Vardhan PV, Shukla LI. Gamma irradiation of medicinally important plants and the enhancement of secondary metabolite production. Int J Radiat Biol. 2017;93(9):967–979. doi:10.1080/09553002.2017.1344788
93. Prabhu S, Poorniammal R, Dufossé L. Microbial Metabolites: a Sustainable Approach to Combat Plant Pests. Metabolites. 2025;15(6):418. doi:10.3390/metabo15060418
94. Chen Y, Lei Z, Gu D, et al. Gut microbiota: a potential enhancing factor for the therapeutic efficacy of bioactive compounds in herbal medicines. Fitoterapia. 2025;183:106570. doi:10.1016/j.fitote.2025.106570
95. Lu Y, Yu X, Wang Z, et al. Microbiota-gut-brain axis: natural antidepressants molecular mechanism. Phytomedicine. 2024;134:156012. doi:10.1016/j.phymed.2024.156012
96. Myrtsi ED, Koulocheri SD, Iliopoulos V, Haroutounian SA. High-Throughput Quantification of 32 Bioactive Antioxidant Phenolic Compounds in Grapes, Wines and Vinification Byproducts by LC-MS/MS. Antioxidants. 2021;10(8):1174. doi:10.3390/antiox10081174
97. Das T, Chatterjee N, Capanoglu E, Lorenzo JM, Das AK, Dhar P. The synergistic ramification of insoluble dietary fiber and associated non-extractable polyphenols on gut microbial population escorting alleviation of lifestyle diseases. Food Chem X. 2023;18:100697. doi:10.1016/j.fochx.2023.100697
98. Yan Z, Zhong Y, Duan Y, Chen Q, Li F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim Nutr. 2020;6(2):115–123. doi:10.1016/j.aninu.2020.01.001
99. Oliviero F, Scanu A, Zamudio-Cuevas Y, Punzi L, Spinella P. Anti-inflammatory effects of polyphenols in arthritis. J Sci Food Agric. 2018;98(5):1653–1659. doi:10.1002/jsfa.8664
100. Bouarab-Chibane L, Forquet V, Lantéri P, et al. Antibacterial Properties of Polyphenols: characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front Microbiol. 2019;10:829. doi:10.3389/fmicb.2019.00829
101. Niu Y, Zhang J, Shi D, Zang W, Niu J. Glycosides as Potential Medicinal Components for Ulcerative Colitis: a Review. Molecules. 2023;28(13):5210. doi:10.3390/molecules28135210
102. Lv M, Wan X, Wang Y, et al. Combined gut microbiome and metabolomics to reveal the mechanism of proanthocyanidins from the roots of Ephedra sinica Stapf on the treatment of ulcerative colitis. J Pharm Biomed Anal. 2024;249:116351. doi:10.1016/j.jpba.2024.116351
103. Wang K, Guo J, Chang X, Gui S. Painong-San extract alleviates dextran sulfate sodium-induced colitis in mice by modulating gut microbiota, restoring intestinal barrier function and attenuating TLR4/NF-κB signaling cascades. J Pharm Biomed Anal. 2022;209:114529. doi:10.1016/j.jpba.2021.114529
104. Li J, Lu Y, Wang D, et al. Schisandrin B prevents ulcerative colitis and colitis-associated-cancer by activating focal adhesion kinase and influence on gut microbiota in an in vivo and in vitro model. Eur J Pharmacol. 2019;854:9–21. doi:10.1016/j.ejphar.2019.03.059
105. Li Y. Gut microbiota in the pathogenesis and treatment of inflammatory bowel disease: a critical review of mechanisms and therapeutic advances. Front Med Lausanne. 2026;13:1738292. doi:10.3389/fmed.2026.1738292
106. Chen K, Wang H, Yang Y, et al. Common mechanisms of Gut microbe-based strategies for the treatment of intestine-related diseases: based on multi-target interactions with the intestinal barrier. Cell Commun Signal. 2025;23(1):288. doi:10.1186/s12964-025-02299-5
107. Caballero-Flores G, Pickard JM, Núñez G. Microbiota-mediated colonization resistance: mechanisms and regulation. Nat Rev Microbiol. 2023;21(6):347–360. doi:10.1038/s41579-022-00833-7
108. Zong Y, Meng J, Mao T, Han Q, Zhang P, Shi L. Repairing the intestinal mucosal barrier of traditional Chinese medicine for ulcerative colitis: a review. Front Pharmacol. 2023;14:1273407. doi:10.3389/fphar.2023.1273407
109. Zhu C, Li K, Peng -X-X, et al. Berberine a traditional Chinese drug repurposing: its actions in inflammation-associated ulcerative colitis and cancer therapy. Front Immunol. 2022;13:1083788. doi:10.3389/fimmu.2022.1083788
110. Du M, Liu X, Ji X, et al. Berberine alleviates enterotoxigenic Escherichia coli-induced intestinal mucosal barrier function damage in a piglet model by modulation of the intestinal microbiome. Front Nutr. 2025;11:1494348. doi:10.3389/fnut.2024.1494348
111. Tang S, Liu W, Zhao Q, et al. Combination of polysaccharides from Astragalus membranaceus and Codonopsis pilosula ameliorated mice colitis and underlying mechanisms. J Ethnopharmacol. 2021;264:113280. doi:10.1016/j.jep.2020.113280
112. Li Z-Y, Lin L-H, Liang H-J, et al. Lycium barbarum polysaccharide alleviates DSS-induced chronic ulcerative colitis by restoring intestinal barrier function and modulating gut microbiota. Ann Med. 2023;55(2):2290213. doi:10.1080/07853890.2023.2290213
113. Martínez-Ruiz M, Robeson MS, Piccolo BD. Fueling the fire: colonocyte metabolism and its effect on the colonic epithelia. Crit Rev Food Sci Nutr. 2025;65(33):8737–8756. doi:10.1080/10408398.2025.2507701
114. Zhang Y, Zhu M, Dai Y, Gao L, Cheng L. Research Progress in Ulcerative Colitis: the Role of Traditional Chinese Medicine on Gut Microbiota and Signaling Pathways. Am J Chin Med. 2024;52(8):2277–2336. doi:10.1142/S0192415X24500885
115. Zhong Y, Xiao Q, Huang J, et al. Ginsenoside Rg1 Alleviates Ulcerative Colitis in Obese Mice by Regulating the Gut Microbiota–Lipid Metabolism–Th1/Th2/Th17 Cells Axis. J Agric Food Chem. 2023;71(50):20073–20091. doi:10.1021/acs.jafc.3c04811
116. Zhu F, Ren C, Zhou K, et al. The Herbal PairPolygonum hydropiperL-Coptis chinensis Franch Attenuate DSS -Induced Ulcerative Colitis by Modulating Metabolism and Intestinal Flora. Food Sci Nutr. 2025;13(7):e70584. doi:10.1002/fsn3.70584
117. Song X, Wang W, Liu L, et al. Poria cocos Attenuated DSS-Induced Ulcerative Colitis via NF-κB Signaling Pathway and Regulating Gut Microbiota. Molecules. 2024;29(9):2154. doi:10.3390/molecules29092154
118. Yang S, Yu M. Role of Goblet Cells in Intestinal Barrier and Mucosal Immunity. J Inflamm Res. 2021;14:3171–3183. doi:10.2147/JIR.S318327
119. van der Post S, Jabbar KS, Birchenough G, et al. Structural weakening of the colonic mucus barrier is an early event in ulcerative colitis pathogenesis. Gut. 2019;68(12):2142–2151. doi:10.1136/gutjnl-2018-317571
120. Zhou Y, Zhang D, Cheng H, et al. Repairing gut barrier by traditional Chinese medicine: roles of gut microbiota. Front Cell Infect Microbiol. 2024;14:1389925. doi:10.3389/fcimb.2024.1389925
121. Mishra A, Vishwa R, Alqahtani MS, et al. The role of MUC2 in cancer - Biological significance and therapeutic implications: a review. Int J Biol Macromol. 2025;333(Pt 2):148480. doi:10.1016/j.ijbiomac.2025.148480
122. Hoffmann W. Trefoil Factor Family (TFF) Peptides and Their Links to Inflammation: a Re-evaluation and New Medical Perspectives. Int J Mol Sci. 2021;22(9):4909. doi:10.3390/ijms22094909
123. Lei L, Wang J, Wang J, et al. Quercetin boosts gut microbiota-driven production of isovanillic acid to alleviate colitis via enhancing intestinal barrier function. Curr Res Food Sci. 2025;11:101183. doi:10.1016/j.crfs.2025.101183
124. Xiao Y, Li X, Fang Y, et al. Berberine suppresses colon inflammation via integrated modulation of host metabolism, microbial ecology, and innate immune signaling. Theranostics. 2026;16(4):2019–2036. doi:10.7150/thno.116546
125. Nie X, Lu Q, Yin Y, He Z, Bai Y, Zhu C. Microbiome and metabolome analyses reveal significant alterations of gut microbiota and bile acid metabolism in ETEC-challenged weaned piglets by dietary berberine supplementation. Front Microbiol. 2024;15:1428287. doi:10.3389/fmicb.2024.1428287
126. Bai Y, Huang F, Zhang R, et al. Longan pulp polysaccharide protects against cyclophosphamide-induced immunosuppression in mice by promoting intestinal secretory IgA synthesis. Food Funct. 2020;11(3):2738–2748. doi:10.1039/c9fo02780g
127. Fan Q, Guan X, Hou Y, et al. Paeoniflorin modulates gut microbial production of indole-3-lactate and epithelial autophagy to alleviate colitis in mice. Phytomedicine. 2020;79:153345. doi:10.1016/j.phymed.2020.153345
128. Pelaseyed T, Bergström JH, Gustafsson JK, et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol Rev. 2014;260(1):8–20. doi:10.1111/imr.12182
129. Mizoguchi A, Yano A, Himuro H, Ezaki Y, Sadanaga T, Mizoguchi E. Clinical importance of IL-22 cascade in IBD. J Gastroenterol. 2018;53(4):465–474. doi:10.1007/s00535-017-1401-7
130. Shen J, Cheng J, Zhu S, et al. Regulating effect of baicalin on IKK/IKB/NF-kB signaling pathway and apoptosis-related proteins in rats with ulcerative colitis. Int Immunopharmacol. 2019;73:193–200. doi:10.1016/j.intimp.2019.04.052
131. Alrafas HR, Busbee PB, Nagarkatti M, Nagarkatti PS. Resveratrol modulates the gut microbiota to prevent murine colitis development through induction of Tregs and suppression of Th17 cells. J Leukoc Biol. 2019;106(2):467–480. doi:10.1002/JLB.3A1218-476RR
132. Mehandru S, Colombel JF. The intestinal barrier, an arbitrator turned provocateur in IBD. Nat Rev Gastroenterol Hepatol. 2021;18(2):83–84. doi:10.1038/s41575-020-00399-w
133. Braga-Neto MB, Ivanov AI. A Century of Progress Toward Gut Barrier Targeting Therapies in Inflammatory Diseases. Am J Pathol. 2025;195(11):1964–1966. doi:10.1016/j.ajpath.2025.04.021
134. Mo J, Ni J, Zhang M, et al. Mulberry Anthocyanins Ameliorate DSS-Induced Ulcerative Colitis by Improving Intestinal Barrier Function and Modulating Gut Microbiota. Antioxidants. 2022;11(9):1674. doi:10.3390/antiox11091674
135. Li Z, Chu T, Sun X, et al. Polyphenols-rich Portulaca oleracea L. (purslane) alleviates ulcerative colitis through restiring the intestinal barrier, gut microbiota and metabolites. Food Chem. 2025;468:142391. doi:10.1016/j.foodchem.2024.142391
136. Rios-Covian D, Gueimonde M, Duncan SH, Flint HJ, de Los Reyes-Gavilan CG. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol Lett. 2015;362(21):fnv176. doi:10.1093/femsle/fnv176
137. Rivière A, Selak M, Lantin D, Leroy F, De Vuyst L. Bifidobacteria and Butyrate-Producing Colon Bacteria: importance and Strategies for Their Stimulation in the Human Gut. Front Microbiol. 2016;7:979. doi:10.3389/fmicb.2016.00979
138. Fakharian F, Thirugnanam S, Welsh DA, et al. The Role of Gut Dysbiosis in the Loss of Intestinal Immune Cell Functions and Viral Pathogenesis. Microorganisms. 2023;11(7):1849. doi:10.3390/microorganisms11071849
139. Camilleri M. Leaky gut: mechanisms, measurement and clinical implications in humans. Gut. 2019;68(8):1516–1526. doi:10.1136/gutjnl-2019-318427
140. Zhang HL, Chang J, Sun CP, et al. Andrographolide and its sulfated metabolite alleviated DSS-induced inflammatory bowel disease through regulating inflammation and immune via MAPK/NLRP3 pathways and the balance of Th17/Treg cells. Mol Immunol. 2025;183:313–320. doi:10.1016/j.molimm.2025.05.015
141. Shen X, Chen H, Wen T, et al. A natural chalcone cardamonin inhibits necroptosis and ameliorates dextran sulfate sodium (DSS)-induced colitis by targeting RIPK1/3 kinases. Eur J Pharmacol. 2023;954:175840. doi:10.1016/j.ejphar.2023.175840
142. Patankar JV, Becker C. Cell death in the gut epithelium and implications for chronic inflammation. Nat Rev Gastroenterol Hepatol. 2020;17(9):543–556. doi:10.1038/s41575-020-0326-4
143. PubMed. Tales from the crypt: new insights into intestinal stem cells. Available from: https://pubmed.ncbi.nlm.nih.gov/30429586/.
144. Luo H, Guo M, Li M, et al. Protective Effect of Rosavin Against Intestinal Epithelial Injury in Colitis Mice and Intestinal Organoids. J Inflamm Res. 2024;17:6023–6038. doi:10.2147/JIR.S474368
145. Yu ZL, Gao RY, Lv C, et al. Notoginsenoside R1 promotes Lgr5+ stem cell and epithelium renovation in colitis mice via activating Wnt/β-Catenin signaling. Acta Pharmacol Sin. 2024;45(7):1451–1465. doi:10.1038/s41401-024-01250-7
146. Schmitz H, Barmeyer C, Fromm M, et al. Altered tight junction structure contributes to the impaired epithelial barrier function in ulcerative colitis. Gastroenterology. 1999;116(2):301–309. doi:10.1016/s0016-5085(99)70126-5
147. Dong Y, Fan H, Zhang Z, et al. Berberine ameliorates DSS-induced intestinal mucosal barrier dysfunction through microbiota-dependence and Wnt/β-catenin pathway. Int J Biol Sci. 2022;18(4):1381–1397. doi:10.7150/ijbs.65476
148. Wang Q, Meng Q, Chen Y, et al. Interaction between gut microbiota and immunity in health and intestinal disease. Front Immunol. 2025;16:1673852. doi:10.3389/fimmu.2025.1673852
149. Neurath MF. Targeting immune cell circuits and trafficking in inflammatory bowel disease. Nat Immunol. 2019;20(8):970–979. doi:10.1038/s41590-019-0415-0
150. Moraitis I, Taelman J, Arozamena B, et al. Mucosal Macrophages Govern Intestinal Regeneration in Response to Injury. Gastroenterology. 2025;169(1):119–135.e26. doi:10.1053/j.gastro.2025.01.252
151. Li S, Lu S, Xiong J, et al. Hydroxyethyl starch-curcumin nanoparticles ameliorate DSS-induced ulcerative colitis in mice via synergistic TLR4/NF-κB suppression, Nrf2 activation, intestinal barrier restoration, and gut microbiota modulation. Front Pharmacol. 2025;16:1610711. doi:10.3389/fphar.2025.1610711
152. Schulthess J, Pandey S, Capitani M, et al. The Short Chain Fatty Acid Butyrate Imprints an Antimicrobial Program in Macrophages. Immunity. 2019;50(2):432–445.e7. doi:10.1016/j.immuni.2018.12.018
153. Tao Q, Liang Q, Fu Y, et al. Puerarin ameliorates colitis by direct suppression of macrophage M1 polarization in DSS mice. Phytomedicine. 2024;135:156048. doi:10.1016/j.phymed.2024.156048
154. Zhou J, Yang Q, Wei W, Huo J, Wang W. Codonopsis pilosula polysaccharide alleviates ulcerative colitis by modulating gut microbiota and SCFA/GPR/NLRP3 pathway. J Ethnopharmacol. 2025;337(Pt 2):118928. doi:10.1016/j.jep.2024.118928
155. Gomez-Bris R, Saez A, Herrero-Fernandez B, Rius C, Sanchez-Martinez H, Gonzalez-Granado JM. CD4 T-Cell Subsets and the Pathophysiology of Inflammatory Bowel Disease. Int J Mol Sci. 2023;24(3):2696. doi:10.3390/ijms24032696
156. Li Q, Li L, Li Q, Wang J, Nie S, Xie M. Influence of Natural Polysaccharides on Intestinal Microbiota in Inflammatory Bowel Diseases: an Overview. Foods. 2022;11(8):1084. doi:10.3390/foods11081084
157. Zhang Y, Ji W, Qin H, et al. Astragalus polysaccharides alleviate DSS-induced ulcerative colitis in mice by restoring SCFA production and regulating Th17/Treg cell homeostasis in a microbiota-dependent manner. Carbohydr Polym. 2025;349(Pt A):122829. doi:10.1016/j.carbpol.2024.122829
158. Ruder B, Becker C. At the Forefront of the Mucosal Barrier: the Role of Macrophages in the Intestine. Cells. 2020;9(10):2162. doi:10.3390/cells9102162
159. Hu Y, Chen LL, Ye Z, et al. Indigo naturalis as a potential drug in the treatment of ulcerative colitis: a comprehensive review of current evidence. Pharm Biol. 2024;62(1):818–832. doi:10.1080/13880209.2024.2415652
160. Wu X, Wang S, Lu J, et al. Seeing the unseen of Chinese herbal medicine processing (Paozhi): advances in new perspectives. Chin Med. 2018;13:4. doi:10.1186/s13020-018-0163-3
161. Heinrich M, Appendino G, Efferth T, et al. Best practice in research - Overcoming common challenges in phytopharmacological research. J Ethnopharmacol. 2020;246:112230. doi:10.1016/j.jep.2019.112230
162. Liu CL, Jiang Y, Li HJ. Quality Consistency Evaluation of Traditional Chinese Medicines: current Status and Future Perspectives. Crit Rev Anal Chem. 2025;55(4):684–701. doi:10.1080/10408347.2024.2305267
163. Zhao Q, Chen XY, Martin C. Scutellaria baicalensis, the golden herb from the garden of Chinese medicinal plants. Sci Bull. 2016;61(18):1391–1398. doi:10.1007/s11434-016-1136-5
164. Dong M, Li J, Yang D, Li M, Wei J. Biosynthesis and Pharmacological Activities of Flavonoids, Triterpene Saponins and Polysaccharides Derived from Astragalus membranaceus. Molecules. 2023;28(13):5018. doi:10.3390/molecules28135018
165. Wang L, Shao L, Chen MY, et al. Panax notoginseng Alleviates Colitis via the Regulation of Gut Microbiota. Am J Chin Med. 2023;51(1):107–127. doi:10.1142/S0192415X23500076
166. Cheng H, Zhang D, Wu J, et al. Interactions between gut microbiota and polyphenols: a mechanistic and metabolomic review. Phytomedicine. 2023;119:154979. doi:10.1016/j.phymed.2023.154979
167. Dey P. Gut microbiota in phytopharmacology: a comprehensive overview of concepts, reciprocal interactions, biotransformations and mode of actions. Pharmacol Res. 2019;147:104367. doi:10.1016/j.phrs.2019.104367
168. Wahlström A, Sayin SI, Marschall HU, Bäckhed F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016;24(1):41–50. doi:10.1016/j.cmet.2016.05.005
169. Hugenholtz F, de Vos WM. Mouse models for human intestinal microbiota research: a critical evaluation. Cell Mol Life Sci. 2018;75(1):149–160. doi:10.1007/s00018-017-2693-8
170. Nguyen TLA, Vieira-Silva S, Liston A, Raes J. How informative is the mouse for human gut microbiota research? Dis Model Mech. 2015;8(1):1–16. doi:10.1242/dmm.017400
171. Johansson MEV, Phillipson M, Petersson J, Velcich A, Holm L, Hansson GC. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc Natl Acad Sci U S A. 2008;105(39):15064–15069. doi:10.1073/pnas.0803124105
172. Katsandegwaza B, Horsnell W, Smith K. Inflammatory Bowel Disease: a Review of Pre-Clinical Murine Models of Human Disease. Int J Mol Sci. 2022;23(16):9344. doi:10.3390/ijms23169344
173. Runge S, von Zedtwitz S, Maucher AM, et al. Laboratory mice engrafted with natural gut microbiota possess a wildling-like phenotype. Nat Commun. 2025;16(1):5301. doi:10.1038/s41467-025-60554-2
174. Ma J, Urgard E, Runge S, et al. Laboratory mice with a wild microbiota generate strong allergic immune responses. Sci Immunol. 2023;8(87):eadf7702. doi:10.1126/sciimmunol.adf7702
175. Espín JC, Jarrín-Orozco MP, Osuna-Galisteo L, Ávila-Gálvez MÁ, Romo-Vaquero M, Selma MV. Perspective on the Coevolutionary Role of Host and Gut Microbiota in Polyphenol Health Effects: metabotypes and Precision Health. Mol Nutr Food Res. 2024;68(22):e2400526. doi:10.1002/mnfr.202400526
176. Lee JWJ, Plichta D, Hogstrom L, et al. Multi-omics reveal microbial determinants impacting responses to biologic therapies in inflammatory bowel disease. Cell Host Microbe. 2021;29(8):1294–1304.e4. doi:10.1016/j.chom.2021.06.019
177. Alam Y, Hakopian S, Ortiz de Ora L, et al. Variation in human gut microbiota impacts tamoxifen pharmacokinetics. mBio. 2025;16(1):e0167924. doi:10.1128/mbio.01679-24
178. Xu Y, Shi J, Mok HL, et al. Macrophage-Mediated Transport of Insoluble Indirubin Induces Hepatic Injury During Intestinal Inflammation. Adv Sci. 2025;12(36):e02993. doi:10.1002/advs.202502993
179. Naganuma M, Sugimoto S, Mitsuyama K, et al. Efficacy of Indigo Naturalis in a Multicenter Randomized Controlled Trial of Patients With Ulcerative Colitis. Gastroenterology. 2018;154(4):935–947. doi:10.1053/j.gastro.2017.11.024
180. Chalasani N, Vuppalanchi R, Navarro V, et al. Acute liver injury due to flavocoxid (Limbrel), a medical food for osteoarthritis: a case series. Ann Intern Med. 2012;156(12):
181. Bemidinezhad A, Zojaji SA, Taraz jamshidi S, Mohammadi M, Alavi MS, Ghorbani A. Evaluation of acute, subacute, and subchronic toxicity of a hepatoprotective herbal formulation. Toxicol Rep. 2023;11:452–459. doi:10.1016/j.toxrep.2023.11.002
182. Kusuhara H, Furuie H, Inano A, et al. Pharmacokinetic interaction study of sulphasalazine in healthy subjects and the impact of curcumin as an in vivo inhibitor of BCRP. Br J Pharmacol. 2012;166(6):1793–1803. doi:10.1111/j.1476-5381.2012.01887.x
183. Teschke R, Danan G. Idiosyncratic Drug-Induced Liver Injury (Dili) and Herb-Induced Liver Injury (HILI): diagnostic Algorithm Based on the Quantitative Roussel Uclaf Causality Assessment Method (RUCAM). Diagnostics. 2021;11(3):458. doi:10.3390/diagnostics11030458
184. Wu X, Wei J, Ran W, et al. The Gut Microbiota-Xanthurenic Acid-Aromatic Hydrocarbon Receptor Axis Mediates the Anticolitic Effects of Trilobatin. Adv Sci. 2025;12(10):e2412234. doi:10.1002/advs.202412234
185. Yu Z, Li D, Sun H. Herba Origani alleviated DSS-induced ulcerative colitis in mice through remolding gut microbiota to regulate bile acid and short-chain fatty acid metabolisms. Biomed Pharmacother. 2023;161:114409. doi:10.1016/j.biopha.2023.114409
186. Jiang Z, Yan M, Qin Y, et al. Quercetin alleviates ulcerative colitis through inhibiting CXCL8-CXCR1/2 axis: a network and transcriptome analysis. Front Pharmacol. 2024;15:1485255. doi:10.3389/fphar.2024.1485255
187. Tang X, Huang Y, Zhu Y, Xu Y. Immune dysregulation in ulcerative colitis: pathogenic mechanisms and therapeutic strategies of traditional Chinese medicine. Front Cell Dev Biol. 2025;13:1610435. doi:10.3389/fcell.2025.1610435
188. Zheng S, Xue T, Wang B, Guo H, Liu Q. Application of network pharmacology in the study of mechanism of Chinese medicine in the treatment of ulcerative colitis: a review. Front Bioinform. 2022;2:928116. doi:10.3389/fbinf.2022.928116
189. Huang Y, Lin X, Wu Q, et al. Integrated serum pharmacochemistry and network pharmacology to reveal the kernel material basis and underlying mechanisms of the fuzi-lizhong pill for ulcerative colitis. J Tradit Complement Med. 2025;15(3):307–318. doi:10.1016/j.jtcme.2024.06.003
190. Lu X, Sun Y, Gong M, et al. Unveiling the Therapeutic Potential of Jiawei Jianpi Huoyu Formula in Ulcerative Colitis: a Multi-Strategies and Experimental Study Integrating Network Pharmacology, Machine Learning, and Mendelian Randomization. J Inflamm Res. 2025;18:10885–10903. doi:10.2147/JIR.S527482
191. Ma H, Li Y, Shi H, et al. Berberine‑calcium alginate-coated macrophage membrane-derived nanovesicles for the oral treatment of ulcerative colitis. Int J Biol Macromol. 2025;294:139114. doi:10.1016/j.ijbiomac.2024.139114
192. Zeng Z, Ouyang J, Sun L, Zeng F, Wu S. A Biomarker-Responsive Nanosystem with Colon-Targeted Delivery for Ulcerative Colitis’s Detection and Treatment with Optoacoustic/NIR-II Fluorescence Imaging. Adv Healthc Mater. 2022;11(22):e2201544. doi:10.1002/adhm.202201544
193. Cai WQ, Liang W, Li D, et al. Reactive Oxygen Species-Responsive Polymer Drug Delivery System Targeted Oxidative Stressed Colon Cells to Ameliorate Colitis. ACS Nano. 2025;19(18):17287–17308. doi:10.1021/acsnano.4c14373
194. Wan X, Zhang C, Lei P, et al. Precision therapeutics for inflammatory bowel disease: advancing ROS-responsive nanoparticles for targeted and multifunctional drug delivery. J Mater Chem B. 2025;13(10):3245–3269. doi:10.1039/d4tb02868f
195. Lei P, Yu H, Ma J, et al. Cell membrane nanomaterials composed of phospholipids and glycoproteins for drug delivery in inflammatory bowel disease: a review. Int J Biol Macromol. 2023;249:126000. doi:10.1016/j.ijbiomac.2023.126000
196. Pandey M, Choudhury H, Singh SK, et al. Budesonide-Loaded Pectin/Polyacrylamide Hydrogel for Sustained Delivery: fabrication, Characterization and In Vitro Release Kinetics. Molecules. 2021;26(9):2704. doi:10.3390/molecules26092704
197. Yang R, Chen M, Song Y, et al. A pH-sensitive dual-encapsulating delivery system: chitosan-alginate hydrogels trapping glycyrrhizic acid/curcumin micelles for synergistic ulcerative colitis. Int J Biol Macromol. 2026;339(Pt 1):149975. doi:10.1016/j.ijbiomac.2025.149975
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Taurine Regulates the Expression of Interleukin −17/10 and Intestinal Flora and Protects the Liver and Intestinal Mucosa in a Nonalcoholic Fatty Liver Disease Rat Model
Zhu FL, Huang T, Lv ZL, Liang G, Yao Z, Lan LC, Qadir A, Chen XQ, Shan QW
Diabetes, Metabolic Syndrome and Obesity 2024, 17:675-689
Published Date: 9 February 2024
Portulaca Oleracea L. as a Potential Therapeutic Drug Intervention in Ulcerative Colitis: Mechanisms of Action and Clinical Studies
Liu XY, Liu ZX, Tan WW, Zhang WB, Zhang YL, Zheng L, Que RY, Wen HZ, Dai YC
Drug Design, Development and Therapy 2024, 18:5931-5946
Published Date: 11 December 2024
Coix Seed Oil Alleviates DSS-Induced Ulcerative Colitis via Intestinal Barrier Repair and Ferroptosis Regulation
Zeng YX, Li NR, Deng BY, Gu YF, Lu SF, Liu Y
Journal of Inflammation Research 2025, 18:2557-2581
Published Date: 20 February 2025
Si-Ni Decoction as a Potential Treatment for Ulcerative Colitis: Modulation of Gut Microbiota and AKT1 Inhibition Through Network Pharmacology and in vivo Validation
Shi L, Chen L, Jin G, Yang Y, Zhu F, Zhou G
Journal of Inflammation Research 2025, 18:6263-6280
Published Date: 14 May 2025
Pneumococcal Endopeptidase O Attenuates Colitis by Inhibiting the Macrophage–CCL2 Axis and Reshaping Gut Microbiota
Wang H, Shao S, Peng Y, Xu W, Zhang J, Liu X, Bian X, Huang K, Zhang X
Journal of Inflammation Research 2026, 19:601434
Published Date: 27 May 2026