Back to Journals » International Journal of Nanomedicine » Volume 21
Targeting Lipid Metabolic Reprogramming to Overcome Immunotherapy Resistance: Systemic Nutritional Modulation and Precision Nanomedicine
Authors Li Q, Yang J, Chen D, Chen Y ![]() , Jiang S
, Jiang S
Received 20 May 2026
Accepted for publication 8 July 2026
Published 15 July 2026 Volume 2026:21 626371
DOI https://doi.org/10.2147/IJN.S626371
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. RDK Misra
Qi Li,1,2,* Jingyi Yang,1,2,* Daozhen Chen,1– 3 Yu Chen,1,2 Sunmin Jiang1,2
1Affiliated Women’s Hospital of Jiangnan University, Wuxi School of Medicine, Jiangnan University, Wuxi, Jiangsu, People’s Republic of China; 2Wuxi School of Medicine, Jiangnan University, Wuxi, Jiangsu, People’s Republic of China; 3Wuxi Higher Health Vocational Technology School, Wuxi, Jiangsu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Sunmin Jiang, Email [email protected] Yu Chen, Email [email protected]
Abstract: Despite the clinical success of immune checkpoint inhibitors (ICIs), resistance driven by a metabolically hostile tumor microenvironment (TME), particularly lipid metabolic reprogramming, remains a formidable challenge. Tumors actively exploit lipid mediators, notably prostaglandin E2 (PGE2), to suppress CD8+ T cells and promote immunosuppressive macrophage polarization. Although targeted nanotherapeutics aim to locally reverse these defects, their in vivo efficacy is frequently abrogated by the host’s nutritional baseline. Specifically, high dietary intake of Omega-6 polyunsaturated fatty acids fuels systemic PGE2 biosynthesis, creating a metabolic “sink” that localized nanotherapies struggle to neutralize. To address this limitation, this review proposes a dual-compartment therapeutic framework combining systemic dietary modulation with precision nanotherapy. As a distinctive contribution, we bridge these biological mechanisms with pharmaceutical design by integrating critical translational barriers, advanced formulation strategies, and emerging technologies for lipid reprogramming. Building upon this, we critically evaluate the most promising formulations, including stimuli-responsive and lipid-targeted nanocarriers, to elucidate their synergistic potential with nutritional interventions. Ultimately, pairing systemic Omega-3 dietary preconditioning with localized nanotherapeutics provides a biologically rational strategy to dismantle lipid-driven immune evasion. Maximizing patient outcomes and advancing future clinical prospects will depend on the rigorous translation of these combinatorial regimens to effectively overcome ICI resistance.
Keywords: metabolic immunotherapy, immunotherapy resistance, lipid metabolic reprogramming, nanotherapeutics, omega-3 fatty acids
Introduction
Immune checkpoint inhibitors (ICIs) achieve durable remissions in a limited fraction of patients with advanced solid tumors.1,2 Frequent intrinsic and acquired resistance severely restricts broader clinical success, demanding new strategies to reverse immune evasion.3 Aberrant lipid metabolism within the tumor microenvironment (TME) represents a highly actionable target to overcome this therapeutic barrier.4,5 Lipid metabolic reprogramming in both malignant cells and tumor-infiltrating immune populations reshapes the intratumoral immune landscape through multiple mechanisms, including the polarization of tumor-associated macrophages (TAMs) toward immunosuppressive M2 phenotypes, functional exhaustion of cytotoxic T lymphocytes, and metabolic exclusion of antitumor effector cells from nutrient-depleted niches.6 Indeed, the targeted reversal of such M2 macrophage polarization is increasingly recognized as a vital strategy to restore robust antitumor immunity and improve overall therapeutic efficacy.7 Crucially, these immunometabolic changes are not solely dictated by cell-intrinsic oncogenic programs. The host systemic nutritional status substantially influences the lipid composition within the tumor and thereby governs the functional states of resident immune populations.8
Dietary polyunsaturated fatty acids (PUFAs) serve as physiologically accessible and mechanistically significant modulators of the immunometabolic axis. The ratio of systemic omega-6 to omega-3 PUFA intake directly influences the equilibrium between proinflammatory and anti-inflammatory eicosanoid biosynthesis within the TME.9 The increased availability of dietary omega-6 PUFA enhances the production of prostaglandin E2 (PGE2) via cyclooxygenase-2 (COX-2)/microsomal prostaglandin E synthase-1 (mPGES-1) pathway. This process suppresses antitumor immunity by inhibiting effector T cell function through EP2/EP4 receptors and promoting the recruitment of myeloid-derived suppressor cells (MDSCs).10 Conversely, dietary supplementation with omega-3 PUFA competitively reduces eicosanoid synthesis from arachidonic acid (AA), decreases the availability of substrates necessary for the biosynthesis of immunosuppressive prostaglandins, and facilitates the resolution of chronic intratumoral inflammation.11,12 Independent of these systemic effects, the intratumoral lipid microenvironment directly influence TAMs polarization through transcriptional and epigenetic mechanisms. This includes the upregulation of the fatty acid scavenger receptor CD36, which enhances the internalization of exogenous lipids and supports fatty acid oxidation (FAO)-dependent M2 metabolic programming.13,14 These observations suggest that the baseline systemic lipid environment serves as an upstream determinant of TME immunosuppression, functioning in parallel with and potentially modulating the efficacy of targeted intratumoral interventions.
Nanomedicine platforms engineered to reprogram intra-tumoral lipid metabolism present precisely targeted mechanisms to disrupt localized immunosuppressive circuits. However, their translation encounters a fundamental challenge that has not received adequate conceptual attention. Actively targeted lipid nanoparticles, biomimetic exosomes, and polymeric nanocarriers are capable of delivering FAO inhibitors, cholesterol metabolism modulators, and PGE2 pathway antagonists with high spatial selectivity to TAMs and other TME populations.15,16 Nonetheless, the pharmacological efficacy of these localized interventions is contingent upon the upstream systemic lipid environment. An abundant systemic supply of pro-inflammatory omega-6-derived lipid mediators can perpetually regenerate the intra-tumoral immunosuppressive milieu that nanomedicine aims to rectify, potentially limiting the durability of the therapeutic effect. This systemic-local interaction has not been explicitly considered as a design factor in the development of nanomedicine for cancer immunotherapy, representing a conceptual gap with direct translational relevance. Addressing such complex interactions demands the development of an integrated mechanistic taxonomy and a clear translational roadmap, analogous to frameworks proposed for other precision immunotherapies.17
The integration of systemic dietary PUFA modulation with precision nanomedicine serves as a critical strategy to overcome lipid-driven immune evasion in solid tumors. The present review will delve into the mechanisms by which dietary PUFA profiles shape the intratumoral immune landscape and influence ICIs responsiveness, and then will detail the design of nanocarrier systems tailored for TME penetration and lipid immunometabolic reprogramming. Ultimately, the review outlines a translational framework that combines systemic nutritional interventions with targeted nanotechnology. This combined approach aims to establish a reproducible and clinically viable strategy for enhancing ICIs efficacy in patients with lipid-metabolically hostile solid tumors (Figure 1).
 |
Figure 1 Mechanistic rationale for synergistic nutritional immunotherapy. (A) PGE2-mediated immune evasion. Circulating AA drives intratumoral PGE2 synthesis via COX-2 in TAMs. This metabolic PGE2 sink suppresses CD8+ T cells, recruits Tregs, and locks TAMs in an immunosuppressive M2-like state. (B) Monotherapy failure. Conventional treatments face two barriers: the desmoplastic stroma (①) physically blocks nanomedicine penetration, while the PGE2 metabolic overload (②) sustains localized immunosuppression and therapy resistance. (C) Synergistic intervention. Dietary omega-3 (EPA/DHA) competitively displaces AA in cellular membranes, restricting PGE2 synthesis and generating SPMs. The reduced PGE2 environment enhances the efficacy of lipid-targeting nanoplatforms. The synergistic blockade forces M2-to-M1 TAM repolarization, triggers ICD, and promotes DC-mediated CD8+ T cell activation for robust tumor eradication. Abbreviations: AA, arachidonic acid; CRT, calreticulin; DAMPs, damage-associated molecular patterns; DC, dendritic cell; DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid; ER, endoplasmic reticulum; HMGB1, high-mobility group box 1; ICD, immunogenic cell death; LNP, lipid nanoparticle; MDSC, myeloid-derived suppressor cell; PGE2, prostaglandin E2; SPM, specialized pro-resolving mediator; TAM, tumor-associated macrophage; TME, tumor microenvironment; TNF-α, tumor necrosis factor-alpha; Treg, regulatory T cell. |
Literature Search Strategy
To ensure a comprehensive and objective overview of the current landscape, a literature search was conducted using primary scientific databases, including PubMed, Web of Science, and Scopus. The search included peer-reviewed articles published in English up to May 2026. We utilized combinations of the following core search terms: “metabolic immunotherapy”, “immunotherapy resistance”, “tumor microenvironment”, “lipid metabolic reprogramming”, “prostaglandin E2”, “omega-3 fatty acids”, “dietary modulation”, “combination therapy”, and “nanotherapeutics”. Articles were manually screened for relevance, with a primary focus on studies elucidating the mechanisms of lipid-driven immune evasion, the application of targeted nanotherapeutics, and the combinatorial strategies integrating systemic nutritional interventions with nanomedicine in cancer immunotherapy.
Dietary PUFAs and Metabolic Remodeling of the Tumor Microenvironment
Mammals lack the δ-12 and δ-15 desaturases necessary for de novo synthesis of essential PUFAs, making systemic PUFA profiles entirely dependent on dietary intake.18–20 Under physiological conditions, exogenous lipids integrate into cellular membranes to regulate bilayer fluidity and receptor signaling.21,22 During oncogenesis, structurally abnormal tumor neovasculature, characterized by fenestrated endothelium and deficient pericyte coverage, facilitates disproportionate extravasation of circulating lipoproteins into the TME.23,24 This localized lipid accumulation is further amplified by transcriptional upregulation of lipid-scavenging receptors, notably CD36 and fatty acid-binding proteins, in malignant cells and tumor-infiltrating immune populations.25,26 As a result, the intratumoral lipid pool reflects the habitual dietary composition of the host and provides the substrates that drive immunometabolic reprogramming across multiple immune cell compartments.27,28
Lipid Accumulation and Transporter-Mediated Immunosuppression
Aberrant lipid partitioning within the TME establishes a metabolically hostile niche that selectively impairs infiltrating cytotoxic populations, including CD8+ T lymphocytes and natural killer (NK) cells.29 CD36-mediated internalization of excess oxidized lipids in CD8+ tumor-infiltrating lymphocytes triggers intracellular lipid peroxidation and a ferroptosis program that abrogates cytotoxic capacity, and elevated CD36 expression correlates with progressive T cell dysfunction and poor clinical outcomes in multiple tumor types.30–33 Within TAMs, CD36-mediated uptake of oxidized low-density lipoprotein activates the IRE1α and ATF6 branches of the unfolded protein response rather than simply accumulating as inert lipid droplets, promoting the formation of lipid-laden immunosuppressive macrophages that reinforce pro-tumoral polarization.34,35 Importantly, the functional consequences of CD36-driven lipid loading differ across immune cell subsets: in CD8+ T cells it drives ferroptotic elimination, while in regulatory T cells it enhances immunosuppressive function through effects on mitochondrial metabolism, indicating that the lipid-enriched TME exploits a shared receptor to generate divergent immunosuppressive outputs across the immune cell repertoire.36,37
Enzymatic Substrate Competition and Eicosanoid-Mediated Immune Evasion
Once incorporated into cellular phospholipids, PUFAs are released as free precursors of bioactive lipid mediators via cytosolic phospholipase A2 (cPLA2).38 The immunological outcome of this cascade depends on the competition between omega-6 and omega-3 PUFAs for the active site of the inducible COX-2, which predominates in tumor cells and infiltrating immune populations, as well as lipoxygenase isoforms.39 In dietary patterns weighted toward high omega-6 intake, AA predominates at the COX-2 catalytic site, sustaining overproduction of PGE2, the principal immunosuppressive eicosanoid in the TME.40,41 PGE2 suppresses CD8+ T cell proliferative expansion and effector differentiation principally through EP2 and EP4-mediated cAMP-PKA signaling, which attenuates TCR-proximal signal transduction and reduces autocrine IL-2 availability, thereby impairing mTORC1-dependent metabolic reprogramming required for effector function.42 Through the same EP2/EP4-cAMP axis, PGE2 abrogates conventional type 1 dendritic cell (cDC1) cytokine and chemokine production and promotes MDSC accumulation and regulatory T cell suppressive activity.43,44 The detailed cellular mechanisms downstream of PGE2 in each immune subset are discussed in Dietary PUFAs and Metabolic Remodeling of the Tumor Microenvironment. Increasing systemic omega-3 availability enables EPA and DHA to competitively occupy COX-2 catalytic sites, reducing PGE2 synthesis and redirecting enzymatic output toward specialized pro-resolving mediators including resolvins, protectins, and maresins.45,46 These mediators suppress chronic intratumoral inflammation and restore conditions permissive for antitumor immunosurveillance primarily by promoting apoptotic cell clearance, antagonizing EP receptor signaling, and restoring dendritic cells (DCs) and NK cells functional activity. It should be noted that the pharmacology of this switch is not binary: certain protectins and D-series resolvins can act as positive allosteric modulators of EP4 at low concentrations, indicating that the immunological outcome of altering the omega-3 to omega-6 ratio reflects a dynamic receptor pharmacology rather than a simple on-off transition between immunosuppressive and immunoprotective states.
Dietary-Microbiota-Epigenetic Axis
Beyond direct cellular effects, systemic PUFA profiles shape gut microbial ecosystem composition. Omega-3-enriched diets selectively promote the proliferation of fermentatively active commensal taxa,47 which metabolize complex dietary carbohydrates into short-chain fatty acids (SCFAs), primarily butyrate, propionate, and acetate, that enter systemic circulation and modulate immunity at sites distant from the gut.48,49 Circulating SCFAs influence antitumor immunity through direct activation of GPR43 and GPR109A on immune and epithelial cells, and through epigenetic reprogramming via selective inhibition of class I histone deacetylases (HDACs).50 By antagonizing class I HDACs activity, butyrate and related SCFAs prevent histone deacetylation in CD8+ T cells, promoting transcriptionally permissive chromatin marks at key effector gene loci including IFNG and TNF.51,52 This chromatin remodeling enhances IFN-γ, TNF-α, and granzyme B (GZMB) secretion, while concurrent mTOR activation, driven in part by increased intracellular acetyl-CoA availability, sustains the metabolic fitness and clonal expansion of effector T cell populations.53,54 Importantly, butyrate also increases histone acetylation at the promoter regions of PDCD1 and CD28 in CD8+ T cells, upregulating PD-1 and CD28 surface expression and thereby enhancing responsiveness to anti-PD-1 checkpoint blockade, a mechanism that offers a direct molecular explanation for the observed correlation between gut microbiota diversity, dietary fiber intake, and clinical response to immunotherapy (Figure 2).55 The net effect of this dietary-microbiome-epigenome axis is systemic host conditioning that lowers the threshold for CD8+ T cell intratumoral infiltration and functionally sensitizes established malignancies to immune checkpoint blockade.
 |
Figure 2 The Omega-3-Microbiota-Epigenetic Axis in CD8+ T Cell Activation. Dietary Omega-3 PUFAs modulate intestinal microbial ecology to favor the expansion of SCFA-producing commensals. Systemically absorbed SCFAs penetrate the TME via hematogenous routes to modulate tumor-infiltrating lymphocytes. Within CD8+ T cells, SCFAs act as potent HDAC inhibitors. This inhibition promotes permissive histone acetylation at the IFNG locus, such epigenetic remodeling primes CD8+ T cells for robust IFN-γ secretion upon cognate antigen recognition, ultimately amplifying antitumor immunosurveillance. Abbreviations: HDAC, histone deacetylase; IFNγ, interferon-gamma; PUFA, polyunsaturated fatty acid; SCFA, short-chain fatty acid. |
Lipid-Driven Metabolic Reprogramming in the TME
Dietary lipid composition directly shapes the intratumoral metabolic landscape and governs the functional states of three critical immune populations, namely TAMs, CD8+ T cells, and cDC1s. Lipid-derived signals suppress each population through mechanistically distinct but interconnected pathways, collectively establishing a self-reinforcing immunosuppressive network.
TAM Metabolic Rewiring and Its Consequences for Tumor Cell Fate
TAMs represent the predominant stromal immune population in solid tumors and exhibit considerable phenotypic plasticity closely linked to intracellular metabolite availability.56–58 In pro-inflammatory macrophages, aerobic glycolysis and the pentose phosphate pathway form the metabolic foundation for antitumor effector functions.59,60 The pentose phosphate pathway maintains a cytosolic NADPH pool that supplies electrons to the NOX2 complex,61,62 whose ROS generation supports the oxidative burst and production of pro-inflammatory mediators including TNF-α.63,64
In a TME enriched with omega-6 dietary lipids, elevated PGE2 concentrations disrupt this program. PGE2 engages EP2 and EP4 receptors, activating adenylate cyclase and PKA, which downregulates c-Myc and its downstream glycolytic transcriptional targets.65–67 Concurrently, upregulation of CD36, FABP4, and CPT1A shifts macrophage bioenergetics toward fatty acid oxidation and mitochondrial OXPHOS.26,68,69 The resulting FAO-derived acetyl-CoA fuels selective histone acetylation at promoters of pro-tumoral genes including Arg1, Il10, and Tgfb1, epigenetically consolidating an immunosuppressive macrophage state.70,71 Reduced flux through the pentose phosphate pathway simultaneously depletes the NADPH pool available to NOX2, impairing oxidative burst capacity (Figure 3A).72–74
 |
Figure 3 Lipid-Driven Metabolic Reprogramming in the TME. (A) PGE2 signaling dictates M2 macrophage metabolic rewiring. PGE2 binds EP2 and EP4 receptors to elevate cAMP. PKA activation suppresses HIF-1α and glycolysis while enhancing Acetyl-CoA production. Nuclear Acetyl-CoA drives HAT-dependent H3K27 acetylation to secure Arg1 and Il10 transcription. Concurrently, membrane CD36 internalizes extracellular fatty acids. CPT1A transports these lipids into mitochondria to fuel FAO. The generated NADPH pool suppresses ROS via NOX2. These integrated metabolic shifts firmly establish the immunosuppressive M2 phenotype. (B) Lipid dysregulation drives CD8+ T cell ferroptosis. CD36 internalizes oxLDL. ACSL4 integrates these lipids to form vulnerable PE-PUFA membrane species. Simultaneously, tumor-driven cystine depletion collapses the intracellular GSH pool. This inactivates GPX4 and unleashes lipid peroxidation. Additionally, PD-1 signaling activates the Akt/GATA1 axis to suppress PLPP1 expression. The resulting phospholipid deficiency physically sensitizes the membrane. Unchecked lipid peroxide accumulation ultimately executes T cell ferroptosis. (C) cDC1 Dysfunction and Systemic Priming Disruption. PGE2 signals through cDC1 EP2 and EP4 to activate the nuclear cAMP/PKA/CREM cascade. This signaling thoroughly represses IRF8 and CCR7 transcription. The loss of CCR7 traps antigen-loaded cDC1s within the tumor stroma, completely severing the link to systemic immune priming. Intracellular aberrant lipid bodies further blockade cross-presentation. Persistent cAMP signaling also induces IDO1 expression, whose paracrine kynurenine product propagates tolerogenic reprogramming across neighboring cDC2s. The concurrent elimination of spatial localization and functional identity enforce systemic immune tolerance across the tumor-draining lymph node axis. In this figure, solid arrows indicate direct activation, molecular conversion, or signaling pathways; dashed arrows represent indirect processes, paracrine signaling, or multi-step metabolic conversions; T-bars indicate negative regulation or direct inhibition; red downward arrows denote downregulation or reduced levels; and crosses denote blocked pathways, suppression, or loss of function. Abbreviations: AA, arachidonic acid; ACSL4, acyl CoA synthetase long chain family member 4; Akt, protein kinase B; cAMP, cyclic adenosine monophosphate; CPT1A, carnitine palmitoyltransferase 1A; EP2/EP4, prostaglandin E2 receptors 2/4; FAO, fatty acid oxidation; GATA1, GATA binding protein 1; GPX4, glutathione peroxidase 4; GSH, reduced glutathione; HAT, histone acetyltransferase; HIF 1α, hypoxia inducible factor 1-α; IDO1, indoleamine 2,3 dioxygenase 1; IRF8, interferon regulatory factor 8; NADPH, reduced nicotinamide adenine dinucleotide phosphate; NOX2, NADPH oxidase 2; oxLDL, oxidized low density lipoprotein; PD-1, programmed cell death protein 1; PE-PUFA, polyunsaturated phosphatidylethanolamine; PGE2, prostaglandin E2; PKA, protein kinase A; PLPP1, phospholipid phosphatase 1; PUFA, polyunsaturated fatty acid; ROS, reactive oxygen species; TAM, tumor associated macrophage; tdLN, tumor draining lymph node. |
The attenuation of macrophage oxidative output carries meaningful consequences for neighboring tumor cells. Under physiological conditions, paracrine ROS and TNF-α from pro-inflammatory macrophages engage all three canonical UPR sensors in tumor cells.75 When proteotoxic load is high and sustained, PERK-dependent phosphorylation of eIF2α drives CHOP induction, committing cells to apoptosis accompanied by calreticulin surface translocation, ATP release, and HMGB1 secretion, the hallmark DAMP triad of ICD.76–78 PGE2-mediated TAM reprogramming lowers this oxidative paracrine output, reducing the probability that tumor cell ER stress reaches the pro-apoptotic threshold. Under diminished stress, tumor cells preferentially engage adaptive UPR arms, upregulating GRP78 to restore protein-folding capacity and transcriptionally inducing PD-L1 on the cell surface.79–81 Beyond reducing the upstream oxidative trigger, PGE2 functions as an inhibitory DAMP that neutralizes downstream immunostimulatory signals even when canonical DAMPs are generated, dominantly skewing the net immunological outcome toward tolerogenic rather than immunogenic cell death.82,83
CD8+ T Cell Ferroptosis
The aberrant lipid composition of the TME exerts direct cytotoxic pressure on infiltrating CD8+ T lymphocytes through a multi-input ferroptosis sensitization mechanism. Through upregulated CD36, oxidized lipids and PUFAs accumulating in the TME are excessively internalized.30,31 ACSL4 converts free PUFAs into PUFA-PE species incorporated into membranes, directly sensitizing cells to iron-catalyzed lipid peroxidation.84,85
Two independent pathways amplify this susceptibility. First, tumor cells competitively monopolize extracellular cystine through high SLC7A11 expression, depriving adjacent T cells of the principal glutathione precursor.86 This cystine deprivation impairs GPX4 activity and independently upregulates CD36-mediated lipid uptake in T cells, creating a feed-forward amplification circuit in which antioxidant depletion and lipid overloading reinforce each other.87 Second, PD-1 signaling suppresses PLPP1 expression through the Akt-GATA1 axis, reducing total membrane phospholipid pools and lowering the threshold at which PUFA-PE species accumulate to ferroptosis-executing concentrations (Figure 3B).32
An important contextual paradox warrants acknowledgment. The IFN-γ that CD8+ T cells deploy to drive tumor cell ferroptosis simultaneously creates conditions that render T cells themselves more susceptible to ferroptotic elimination.30 Furthermore, tumor cells sustain GPX4-independent ferroptosis resistance through the FSP1-CoQ10 axis, which regenerates CoQ10 to quench lipid radical propagation without requiring glutathione.88 This asymmetric resistance profile means the lipid-enriched TME functions as a selective elimination pressure that preferentially depletes effector immune cells while sparing malignant cells, representing a fundamental barrier to ICIs efficacy in solid tumors.
cDC1 Dysfunction and Disruption of Systemic Immune Priming
Effective anti-tumor immunity requires coupling intratumoral cytotoxicity with systemic adaptive priming in tumor-draining lymph nodes (tdLNs), a process physically bridged by cDC1s.89,90 Aberrant lipid accumulation impairs cDC1 function at two mechanistically distinct levels. At the intracellular level, oxidized lipids generated as byproducts of omega-6 PUFA metabolism are taken up by cDC1s via scavenger receptors.91 The resulting oxidatively truncated lipid bodies covalently bind HSP70, preventing peptide-MHC class I complex translocation to the cell surface and abolishing cross-presentation capacity.92
At the receptor signaling level, tumor-derived PGE2 activates EP2 and EP4 on cDC1s, engaging the downstream cAMP-PKA-CREM pathway.93 Critically, combined EP2 and EP4 signaling rather than either receptor alone drives cDC1 transcriptional reprogramming, and dual but not single-receptor blockade completely rescues IRF8 expression. IRF8 loss effectively dedifferentiates cDC1s toward a tolerogenic phenotype and reduces CCR7 expression required for migration toward tdLNs, physically confining antigen-loaded cDC1s within the tumor stroma.93 Persistent cAMP signaling additionally induces IDO1, whose paracrine kynurenine product propagates tolerogenic reprogramming across neighboring cDC2s (Figure 3C).94
PGE2 thus operates as a dual inhibitor of cDC1-mediated immunity, simultaneously dismantling cDC1 functional identity through the EP2/EP4-cAMP-IRF8 axis and neutralizing extracellular DAMP-mediated DCs activation. Together with the TAM and CD8+ T cell suppression mechanisms described above, these defects prevent naive CD8+ T cell clonal expansion and enforce systemic immune tolerance, completing the self-reinforcing immunosuppressive network across all three cellular axes.
Synergistic Nutritional Immunotherapy: Integrating Dietary Lipids with Targeted Nanomedicine
Reversing the multi-layered immunosuppressive network described above requires interventions that address both systemic substrate availability and local intratumoral signaling simultaneously. Neither dietary modulation nor pharmacological blockade alone achieves this dual objective, which motivates a combinatorial paradigm termed synergistic nutritional immunotherapy.
Pharmacokinetic Constraints of Monotherapies
Systemic enrichment of the host lipid pool with omega-3 PUFAs reduces baseline pro-inflammatory eicosanoid production and shifts the systemic lipid environment toward an immunologically permissive state.95,96 However, reliance on dietary modulation alone faces structural barriers imposed by the tumor. Cancer-associated fibroblasts (CAFs) within desmoplastic stroma continuously cross-link collagen and hyaluronan, physically compressing local vasculature and generating interstitial fluid pressures of 5 to 40 mmHg compared to near-zero baseline levels.97,98 This convective barrier traps circulating lipid metabolites at the tumor periphery, preventing dietary interventions from reaching concentrations sufficient to interrupt TAM-tumor cell metabolic crosstalk in the hypoxic core.44
Targeted pharmacological blockade of the PGE2-EP2/EP4 axis offers a direct therapeutic strategy, but systemic administration of free COX-2 inhibitors or EP4 antagonists carries significant clinical liabilities. Broad prostaglandin suppression disrupts physiological lipid homeostasis, producing gastrointestinal mucosal injury and elevated thromboembolic risk.99,100 Free therapeutic agents also lack active targeting capacity, undergo rapid systemic clearance, and cannot overcome elevated interstitial fluid pressure that restricts passive diffusion into dense myeloid infiltrates, resulting in subtherapeutic local concentrations.101,102 These delivery and efficacy deficits establish the pharmacological rationale for coupling systemic dietary lipid recalibration with the spatiotemporal precision of engineered nanomedicines to address both systemic and localized immunosuppressive barriers.
Nanomedicine-Driven Precision Lipid Metabolic Reprogramming
Precision immunometabolic therapy requires selective intratumoral drug delivery. As a prominent strategy in macrophage-targeted nanomedicine, coating nanocarriers with mannose or M2-targeting peptides directs payload delivery specifically to immunosuppressive TAMs, which overexpress CD206 and CD163 receptors relative to surrounding stromal cells. Ligand binding triggers receptor-mediated endocytosis, depositing therapeutic cargo directly into the macrophage cytoplasm.103–105 To preserve payload integrity against lysosomal degradation, formulations incorporating ionizable lipids or pH-responsive polymers exploit endosomal acidification to release active cargo into the cytosol.106,107
Cytosolic delivery initiates TAM metabolic rewiring through three convergent mechanisms. First, CPT1 inhibitors such as etomoxir have been used as proof-of-concept tools to block mitochondrial fatty acid import in preclinical models, forcing a metabolic shift from OXPHOS toward aerobic glycolysis that promotes pro-inflammatory macrophage repolarization, CD8+ T cell recruitment, and elevated GZMB expression.108,109 The clinical translation of etomoxir itself is limited by off-target mitochondrial toxicity at effective concentrations, and more selective CPT1-targeting strategies are an active area of development. Second, SOAT1 inhibitors such as avasimibe block aberrant cholesterol esterification. In CD8+ T cells, SOAT1 inhibition increases free membrane cholesterol, enhancing T cell receptor clustering and immunological synapse formation to amplify cytotoxic capacity.110,111 Third, nanomedicine-driven CD36 blockade eliminates the exogenous lipid supply to TAMs. Without this substrate, the elevated FAO that drives mitochondrial ROS production, JAK1 phosphorylation, SHP1 dephosphorylation, and downstream STAT6 activation to maintain immunosuppressive macrophage function is extinguished.26,112
Targeting the COX-2-PGE2 paracrine axis restores cDC1 function through two complementary approaches. Nanoparticle-delivered COX-2 siRNA reduces PGE2 synthesis at the transcriptional level, preventing EP2/EP4-mediated IRF8 downregulation in cDC1s and restoring their capacity to coordinate CD8+ T cell responses.113 Alternatively, PLGA nanoparticles encapsulating EP2/EP4 antagonists block PGE2 signaling at the receptor level, enabling DCs to recover co-stimulatory molecule expression including CD80 and CD86, and reactivating CD8+ T cell priming.114 Single-cell transcriptomic analyses demonstrate that EP2/EP4 antagonism restores c-Myc target gene expression and OXPHOS capacity in both T cells and macrophages, reversing the PGE2-mediated metabolic impairment of effector immunity.115,116 Representative nanocarrier platforms implementing these strategies are detailed in Table 1.
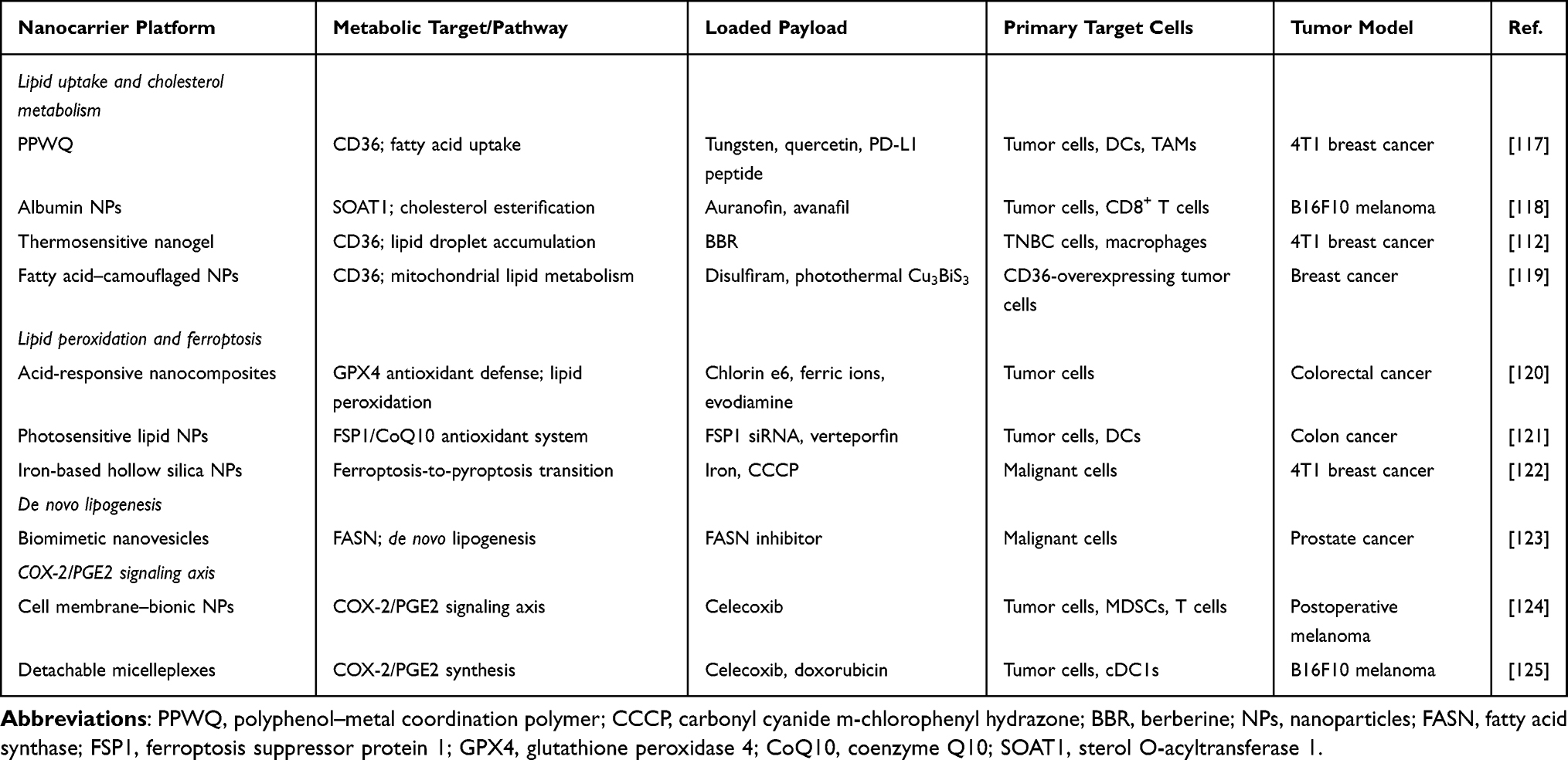 |
Table 1 Representative Nanomedicines Targeting Lipid Metabolism in the TME |
Dietary Conditioning as a Pharmacological Complement to Nanomedicine
The efficacy of targeted nanomedicines is directly conditioned by the dietary lipid environment. High omega-6 intake drives systemic AA accumulation, providing abundant COX-2 substrate that elevates intratumoral PGE2 to concentrations substantially exceeding the physiological range.126 This ligand excess creates a fundamental pharmacological challenge for receptor-level interventions. Preclinical evidence demonstrates that dual EP2/EP4 blockade is required for optimal immune activation, as EP2 and EP4 share overlapping cAMP-mediated downstream signaling and single-node antagonism is insufficient to restore immune cell function when endogenous PGE2 concentrations are high.127 Endogenous PGE2 therefore competes with delivered antagonists for receptor occupancy under conditions of high ligand availability, and because solid tumor barriers inherently limit intratumoral nanoparticle accumulation, antagonist concentrations may remain below the endogenous PGE2 threshold.128
Omega-3 supplementation addresses this constraint at the substrate level. EPA and DHA compete with AA for COX-2 processing, generating the substantially less immunosuppressive PGE2 and thereby reducing baseline pro-inflammatory eicosanoid output.129 Clinical evidence from a randomized crossover trial confirms that supplementation with 2.4 g EPA and 1.2 g DHA daily for six months significantly reduces plasma PGE2 concentrations at one, three, and six months, demonstrating that dietary omega-3 enrichment reliably lowers the systemic endogenous ligand burden.130 In murine cancer models, dietary omega-3 enrichment reduces intratumoral immunosuppressive cytokines, decreases regulatory T cell proportions, enhances cytotoxic T cell activity, and synergizes with anti-PD-1 blockade to suppress tumor growth.131 A parallel line of evidence demonstrates that dietary omega-3 supplementation stabilizes endogenous epoxy fatty acids in the TME and enhances ICIs efficacy in multiple murine tumor models.132 These independent lines of evidence support the concept that dietary lipid modification lowers the endogenous PGE2 burden and may therefore reduce the competitive threshold that currently limits receptor antagonist efficacy under conditions of restricted intratumoral nanoparticle penetration. Whether dietary omega-3 preconditioning meaningfully augments the activity of targeted EP2/EP4 antagonist nanomedicines in a controlled experimental model has not yet been directly tested, representing a defined research gap that warrants prospective investigation. The evidence summarized in Table 2 presents representative nanocarrier platforms achieving TME immunomodulation, which provide the pharmacological foundation against which such dietary combination strategies should be evaluated.
 |
Table 2 Dietary Restriction Strategies Combined with Precision Nanocarrier Delivery for TME Reprogramming |
Tumor Heterogeneity and the Limits of Uniform Intervention
The immunometabolic mechanisms and therapeutic strategies discussed in preceding sections represent generalizable principles, but their practical relevance depends critically on the physical and metabolic context of individual tumor types. Solid tumors differ substantially in stromal architecture and lipid niche composition, and these differences impose constraints that uniform dietary or nanomedicine strategies cannot adequately address.137
The Desmoplastic Barrier in Pancreatic Cancer
Pancreatic ductal adenocarcinoma (PDAC) exemplifies the immunologically cold tumor, defined by a dense desmoplastic stroma in which CAFs continuously deposit fibrillar type I collagen and high-molecular-weight hyaluronan.138 The resulting structural network elevates interstitial fluid pressure, compresses intratumoral vasculature, and generates chronic hypoxia that severely restricts both CD8+ T cell infiltration and the deep-tissue penetration of systemically administered omega-3 PUFAs.139 Single-cell RNA sequencing data from PDAC models and patient samples reveal that eicosanoid synthesis is distributed across multiple cell compartments. Analysis of scRNA-seq datasets identifies PGE2 and prostacyclins as products of CAFs, PGD2 and thromboxanes as predominantly myeloid-derived, and a transition from PGD2 to PGE2 production in the tumor epithelium during malignant progression.140 This multicellular eicosanoid network introduces spatial heterogeneity that bulk-tissue lipidomic measurements cannot fully resolve, with implications for the specificity of systemic PGE2-lowering strategies.
Within nutrient-deprived zones, TAMs sustain FAO and OXPHOS through PPAR pathway activation, a state further stabilized by reduced RIPK3 expression that prevents caspase-1-mediated cleavage of PPAR-γ.141 This bioenergetic adaptation preserves an immunosuppressive phenotype that is partially insulated from circulating lipid composition, presenting a fundamental limitation for systemic nutritional interventions alone. Accessing these compartments requires nanocarriers engineered for active stromal penetration, including size-shrinkable architectures that exploit MMP activity in the stroma and surface functionalization with tumor-penetrating peptides such as iRGD to activate transcytosis-dependent deep tissue transport.142–145
The Adipocyte-Rich Niche in Ovarian and Breast Cancer
Tumors developing within adipocyte-rich environments face a distinct metabolic landscape. In omental metastases of high-grade serous ovarian cancer and breast cancer invading adjacent adipose depots, malignant cells drive adipocyte lipolysis through paracrine IL-6, IL-8, and PTHrP signaling, upregulating ATGL and HSL to release free fatty acids that are transferred to tumor cells and macrophages via CD36-mediated uptake and FABP4-facilitated redistribution.146–149 The sustained local abundance of adipocyte-derived fatty acids maintains CD36-mediated lipid internalization in TAMs, continuously supplying substrates for both FAO and COX-2-dependent PGE2 synthesis.150 This locally amplified immunosuppressive lipid cycle operates at concentrations that standard-dose systemic omega-3 supplementation is unlikely to overcome. Locally targeted co-delivery of CD36 siRNA and EP2/EP4 antagonists offers a more direct approach to simultaneously interrupt exogenous lipid influx and downstream immunosuppressive signaling in these niches, representing precisely the kind of tailored intervention required to sensitize the highly immunosuppressive ovarian cancer microenvironment.151–153
Together, these tumor type-specific contexts make clear that lipid-targeted immunotherapy cannot be uniformly applied across solid malignancies. The dominant lipid metabolic program of each tumor niche must guide the choice of nanocarrier architecture, targeting strategy, and dietary adjunct dosing. This principle directly informs how patients are selected and stratified in clinical trials evaluating lipid-targeted approaches.
Clinical Translation from Biomarker Evaluation to Precision Trials
Quantitative Lipidomic Evaluation and Patient Stratification
Tumor heterogeneity requires precise patient selection for lipid targeted therapies. Observational studies demonstrate that systemic lipidomic profiles predict immune checkpoint inhibitor efficacy. Advanced NSCLC patients with elevated baseline serum EPA to AA ratios achieve better overall survival. This survival benefit operates independently of tumor mutational burden and PD-L1 expression.154–157 Pro-inflammatory lipid profiles drive primary immune resistance. Sustained PGE2 signaling impairs IL-2 responsiveness and mitochondrial function in TCF1+ CD8+ T cells.158 This lipid environment also downregulates IRF8 in cDC1 cells. This specific downregulation impairs cDC1 effector functions including IL-12 and CXCL9 production, disrupting their ability to orchestrate effective CD8+ T cell responses within tumors and tdLNs.93 These combined defects cause the accumulation of exhausted T cells expressing high levels of TOX, PD-1, TIM-3, and LAG-3.159 Omega 3 supplementation requires careful application based on the microenvironment. While DHA exhibits anti-inflammatory effects on DCs by inhibiting maturation and pro-inflammatory cytokine production, the optimal dosing and timing relative to immunotherapy remains to be established in clinical trials.160
Advanced liquid chromatography tandem mass spectrometry combined with multi-omics pipelines provides quantitative measurements of plasma eicosanoids and structural phospholipids.161,162 A six lipid plasma panel successfully distinguished disease control from progressive disease in NSCLC patients receiving chemoimmunotherapy.163 Profiling circulating tumor derived extracellular vesicles offers superior diagnostic specificity. These vesicles carry lipid signatures directly from parent tumor cells. Tumor secreted vesicle associated PGE2 actively promotes MDSC accumulation and macrophage immunosuppression.164–166 Table 3 summarizes these predictive peripheral lipidomic signatures across different solid tumors.
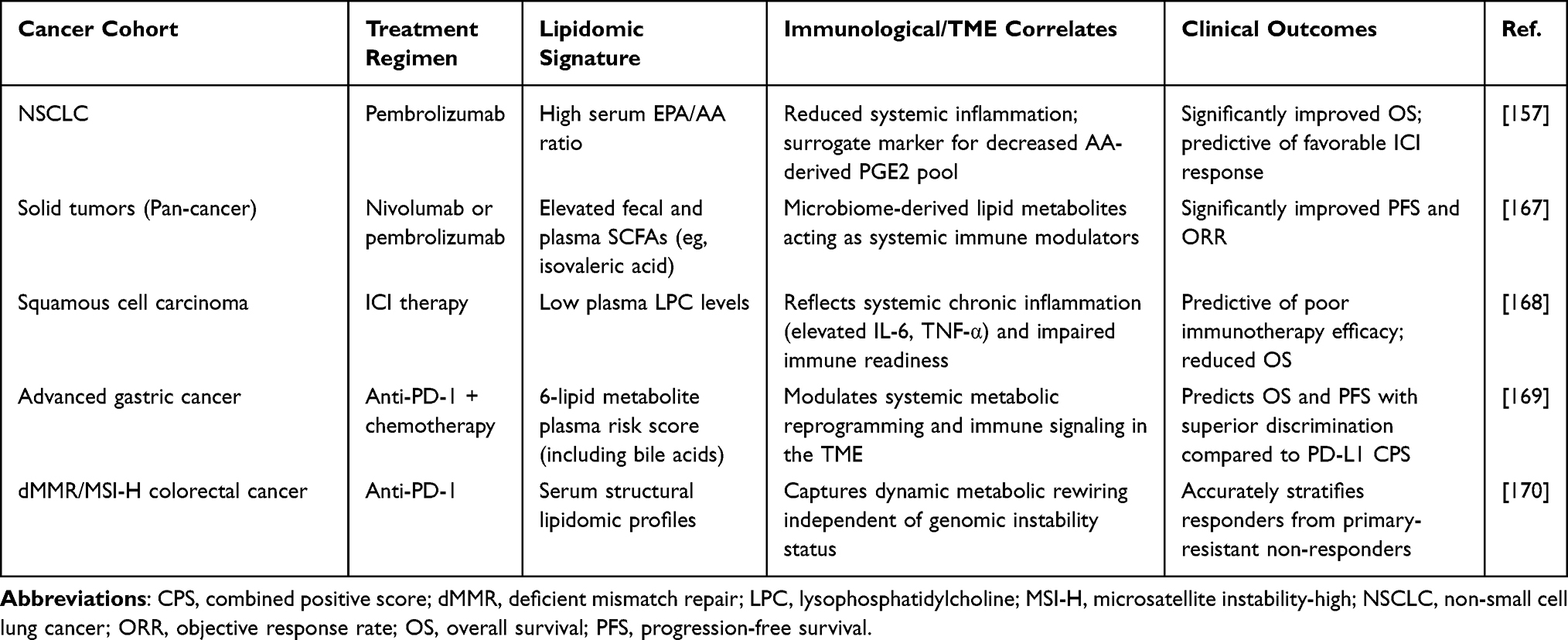 |
Table 3 Clinical Evidence of Peripheral Lipidomic Signatures as Predictive Biomarkers for ICI Efficacy |
Prospective Clinical Trials of Nutritional Immunotherapy
Current prospective trials offer early pharmacodynamic support for combining Omega-3 supplementation with ICIs. Peripheral blood analyses from supplemented cohorts show a consistent decrease in polymorphonuclear MDSCs alongside stable regulatory T cell populations.171 Meta-analyses further confirm that EPA and DHA co-administration lowers systemic serum levels of IL-6, TNF-α, and C-reactive protein.172 These favorable shifts in peripheral immune markers provide proof-of-concept for systemic eicosanoid remodeling. However, they cannot confirm actual immune reprogramming within the TME. Several ongoing trials are evaluating dietary lipid modulation, caloric restriction, and gut microbiome interventions alongside immunotherapy (Table 4). Methodological limitations in their design restrict their ability to validate the combined mechanisms proposed in this review. Most studies fail to quantify intratumoral PGE2 concentrations or local tissue PUFA composition, leaving our core hypothesis without direct histological validation. Habitual Omega-6 intake also remains largely unstandardized at baseline. This inter-patient variability in AA supply inevitably dilutes the resulting eicosanoid-related immune signals. Most importantly, current protocols lack an Omega-3 preconditioning phase before ICIs initiation. Early nutritional intervention is necessary to suppress baseline intratumoral PGE2 levels. Lowering this baseline breaks the pharmacodynamic ceiling of immune tolerance and creates the optimal microenvironment required for subsequent targeted nanocarriers to achieve maximum efficacy.
 |
Table 4 Prospective Clinical Trials Combining Systemic Metabolic Interventions with Cancer Immunotherapy |
Translational Limitations and Future Directions
Pharmacokinetic and Biological Delivery Barriers
Several physiological barriers restrict the clinical application of synergistic nutritional immunotherapy. Individual variations in gut microbiome composition and bile acid metabolism dictate the gastrointestinal absorption of oral lipids.173,174 Free fatty acids in the systemic circulation bind extensively to albumin or accumulate in the liver and adipose tissues. This systemic distribution severely reduces the active lipid fraction reaching the malignant tissue.175 The high interstitial fluid pressure of desmoplastic stroma further restricts lipid penetration. Intratumoral accumulation often remains too low to overcome local PGE2 driven immunosuppression.101 Oral supplementation modulates the peripheral immune system but fails to independently remodel the dense tumor core. Current trial designs reflect these limitations. Most studies do not quantify intratumoral PGE2 concentrations or local tissue PUFA composition. Unstandardized baseline Omega 6 intake causes high inter-patient variability and dilutes the observable immune signals.
Nanomedicine platforms designed to cross these barriers face distinct biological challenges. Serum proteins rapidly coat intravenously administered nanoparticles. This dynamic protein corona hides engineered targeting ligands such as mannose residues and reduces targeting precision.176–178 Complement proteins deposit C3b fragments on the carrier surface. The mononuclear phagocyte system recognizes these fragments and clears the nanoparticles into the liver and spleen.179,180 PEGylation strategies often fail due to pre-existing anti-PEG antibodies in the general population. These antibodies enrich in the protein corona and accelerate macrophage uptake.181,182 Biomimetic cell membrane cloaking offers a physiological alternative. Coating nanocarriers with cell membranes introduces CD47 to engage inhibitory SIRPalpha receptors on circulating phagocytes. This cloaking incorporates CD55 and CD59 to block opsonin deposition and extend systemic circulation time.183,184 Manufacturing these membrane coated nanocarriers at a clinical scale remains a major engineering challenge.
Biomarker Limitations and Precision Oncology Design
Current biomarker platforms require significant technical refinement. Bulk plasma profiling captures systemic eicosanoid states but cannot reliably isolate tumor specific metabolic signals. Tumor derived extracellular vesicle lipidomics offers better specificity. Standardized isolation protocols must resolve the low proportion of tumor derived vesicles in the blood before reliable clinical application.185 Clinical patient selection must also exclude lipid independent resistance mechanisms. Intrinsic Wnt β catenin activation suppresses chemokine production and prevents cDC1 recruitment independently of eicosanoid biology.186 The loss of HLA class I expression through genomic deletion or NLRC5 silencing blocks tumor antigen recognition and lipid targeted therapies cannot reverse these specific genetic and epigenetic resistance programs.25,187,188
Future adaptive clinical trials must incorporate Omega 3 preconditioning phases before initiating immunotherapy or nanomedicine delivery. This early intervention lowers the baseline intratumoral PGE2 burden prior to targeted receptor blockade. Trial designs should require paired biopsies to quantify local tissue PUFA composition and validate intratumoral eicosanoid suppression. Enrolling patients based on elevated circulating PGE2 metabolites and unfavorable Omega 6 to Omega 3 ratios will concentrate trials on the most responsive populations.171,189 Preclinical research should evaluate the direct combination of dietary preconditioning and targeted nanomedicines in desmoplastic and adipocyte rich tumor models. Standardizing vesicle isolation protocols will eventually enable clinicians to pair metabolic vulnerabilities with targeted nanomedicine delivery. This integrated approach represents the biological foundation of precision immunometabolic oncology (Figure 4).
 |
Figure 4 Proposed clinical translational roadmap for synergistic nutritional immunotherapy. Left panel: Patient stratification. Integrates liquid biopsies (plasma lipidomics, PGE2 metabolites, tEVs), tumor biopsies (baseline TME lipid profiles), and exploratory fecal microbiome profiling to guide precise patient selection. Center panel: Two-phase intervention. Phase I: Omega-3 dietary pre-conditioning to deplete the intratumoral PGE2 sink and restore nanomedicine sensitivity. Phase II: Co-administration of lipid-targeting nanocarriers (eg, EP4 antagonists) and anti-PD-1 immune checkpoint inhibitors (ICI). Right panel: Outcome monitoring. Assesses primary clinical endpoints (ORR, PFS, OS), evaluates TME immune infiltration (eg, CD8+ T cells and M1 macrophages), and tracks safety through irAEs and systemic inflammatory markers (eg, IL-6, CRP, transaminases). Abbreviations: CRP, C-reactive protein; EP4, prostaglandin E2 receptor subtype 4; ICI, immune checkpoint inhibitor; IL-6, interleukin-6; irAE, immune-related adverse event; ORR, objective response rate; OS, overall survival; PFS, progression-free survival; PGE2, prostaglandin E2; tEV, tumor-derived extracellular vesicle; TME, tumor microenvironment. |
Conclusion
Malignant tissues actively orchestrate local lipid partitioning to enforce immune exclusion. Within the TME, dietary omega 6 fatty acids are funneled into the AA cascade, fueling COX-2 mediated PGE2 synthesis. This localized eicosanoid pool locks TAMs into a suppressive state dependent on fatty acid oxidation, precipitates lipid peroxidation driven ferroptosis in infiltrating CD8+ T cells, and paralyzes cDC1 lymph node trafficking. Consequently, systemic immune surveillance collapses at multiple spatial checkpoints.
Monotherapeutic attempts to dismantle this lipid shield remain fundamentally constrained by spatial and biochemical barriers. Systemic dietary shifts toward omega 3 fatty acids effectively deplete circulating AA but fail to breach the dense stromal architecture of established tumors. Conversely, nanomedicines engineered to block PGE2 signaling are frequently outcompeted by the overwhelming local concentration of TME derived eicosanoids.
To resolve these isolated vulnerabilities, we propose a sequential intervention paradigm. Systemic preconditioning with dietary omega 3 fundamentally erodes the baseline AA supply, depressing the intratumoral PGE2 reservoir. This metabolically sensitized environment opens a critical therapeutic window, allowing EP4 antagonist loaded nanocarriers to achieve functional TAM reprogramming and rescue cDC1 migration. Liquid biopsy platforms quantifying circulating omega 6 to omega 3 ratios, PGE2 metabolites, and extracellular vesicle lipidomes will enable the precise prospective stratification of patients harboring this lipid dependent phenotype.
Realizing this clinical translation necessitates overcoming inherent nanomedicine delivery barriers and validating robust lipidomic biomarkers. Future Phase I trial designs must map longitudinal metabolic responses, establish precise dosing kinetics, and verify intratumoral immune remodeling. Ultimately, neutralizing this lipid driven evasion network provides a mechanistic rationale to rescue systemic antigen presentation, laying the essential groundwork for durable responses to ICIs.
Data Sharing Statement
No new data were created.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the National Natural Science Foundation of China (No. 82473350, China); Wuxi Municipal Science and Technology Development Fund Project (No. K20253012, China), General scientific research project of Wuxi Municipal Health Commission (No. M202430, China), and Youth Scientific Research Project of Wuxi Municipal Health Commission (No. Q202530, China).
Disclosure
The authors declare no competing interests in this work.
References
1. Cao J, Ding X, Ji J, Zhang L, Luo C. Efficacy and safety of immune checkpoint inhibitors rechallenge in advanced solid tumors: a systematic review and meta-analysis. Front Oncol. 2024;14:1475502. doi:10.3389/fonc.2024.1475502
2. Poorkhani A, Tajaldini M, Ahmadi F, et al. Emerging new immune checkpoint inhibitors in solid tumor immunotherapy. Naunyn Schmiedebergs Arch Pharmacol. 2025;398(9):11733–21. doi:10.1007/s00210-025-04131-w
3. Zhang C, Zhang C, Wang H. Immune-checkpoint inhibitor resistance in cancer treatment: current progress and future directions. Cancer Lett. 2023;562:216182. doi:10.1016/j.canlet.2023.216182
4. Zhang H, Li S, Wang D, et al. Metabolic reprogramming and immune evasion: the interplay in the tumor microenvironment. Biomark Res. 2024;12(1):96. doi:10.1186/s40364-024-00646-1
5. Gao B, Lu Y, Lai X, et al. Metabolic reprogramming in hepatocellular carcinoma: mechanisms of immune evasion and therapeutic implications. Front Immunol. 2025;16:1592837. doi:10.3389/fimmu.2025.1592837
6. Wang J, He Y, Hu F, et al. Metabolic reprogramming of immune cells in the tumor microenvironment. Int J Mol Sci. 2024;25(22):12223. doi:10.3390/ijms252212223
7. Liu K, Kong L, Cui H, et al. Thymosin α1 reverses oncolytic adenovirus-induced M2 polarization of macrophages to improve antitumor immunity and therapeutic efficacy. Cell Rep Med. 2024;5(10):101751. doi:10.1016/j.xcrm.2024.101751
8. Goswami S, Zhang Q, Celik CE, Reich EM, Yilmaz ÖH. Dietary fat and lipid metabolism in the tumor microenvironment. Biochim Biophys Acta BBA - Rev Cancer. 2023;1878(6):188984. doi:10.1016/j.bbcan.2023.188984
9. Liu R, Wang C, Tao Z, Hu G. Lipid metabolism reprogramming in cancer: insights into tumor cells and immune cells within the tumor microenvironment. Biomedicines. 2025;13(8):1895. doi:10.3390/biomedicines13081895
10. Fadaee M, Lahouty M, Abdi M, Asghari Ozma M, Lotfollahi L, Kazemi T. The effects of n-3 PUFAs on colorectal cancer carcinogenesis: comprehensive review. Food Agric Immunol. 2025;36(1):2501967. doi:10.1080/09540105.2025.2501967
11. Zou B, Zhao D, Zhou S, Kang JX, Wang B. Insight into the effects of Omega-3 fatty acids on gut microbiota: impact of a balanced tissue Omega-6/Omega-3 ratio. Front Nutr. 2025;12:1575323. doi:10.3389/fnut.2025.1575323
12. Yang YC, Chang SC, Hung CS, Shen MH, Lai CL, Huang CJ. Gut-microbiota-derived metabolites and probiotic strategies in colorectal cancer: implications for disease modulation and precision therapy. Nutrients. 2025;17(15):2501. doi:10.3390/nu17152501
13. Wang J, Zhang W, Liu C, et al. Reprogramming of lipid metabolism mediates crosstalk, remodeling, and intervention of microenvironment components in breast cancer. Int J Biol Sci. 2024;20(5):1884–1904. doi:10.7150/ijbs.92125
14. Ma Y, Huang Y, Hu F, Shu K. Lipid metabolic rewiring in glioma‑associated microglia/macrophages (review). Int J Mol Med. 2024;54(5):102. doi:10.3892/ijmm.2024.5426
15. Kang X, Huang Y, Wang H, et al. Tumor-associated macrophage targeting of nanomedicines in cancer therapy. Pharmaceutics. 2023;16(1):61. doi:10.3390/pharmaceutics16010061
16. Shi X, Askari Rizvi SF, Yang Y, Liu G. Emerging nanomedicines for macrophage-mediated cancer therapy. Biomaterials. 2025;316:123028. doi:10.1016/j.biomaterials.2024.123028
17. Al-Azzani H, Arthur Vithran DT, Aliouat H, Zhou W, Mao X. Precision bacterial immunotherapy: an integrated mechanistic taxonomy and translational roadmap against antimicrobial resistance. Front Immunol. 2025;16:1675682. doi:10.3389/fimmu.2025.1675682
18. Nakamura MT, Nara TY. Structure, function, and dietary regulation of Δ6, Δ5, and Δ9 desaturases. Annu Rev Nutr. 2004;24(1):345–376. doi:10.1146/annurev.nutr.24.121803.063211
19. Koundouros N, Poulogiannis G. Reprogramming of fatty acid metabolism in cancer. Br J Cancer. 2020;122(1):4–22. doi:10.1038/s41416-019-0650-z
20. Ringel AE, Drijvers JM, Baker GJ, et al. Obesity shapes metabolism in the tumor microenvironment to suppress anti-tumor immunity. Cell. 2020;183(7):1848–1866.e26. doi:10.1016/j.cell.2020.11.009
21. Levental KR, Malmberg E, Symons JL, et al. Lipidomic and biophysical homeostasis of mammalian membranes counteracts dietary lipid perturbations to maintain cellular fitness. Nat Commun. 2020;11(1):1339. doi:10.1038/s41467-020-15203-1
22. Ali O, Szabó A. Review of eukaryote cellular membrane lipid composition, with special attention to the fatty acids. Int J Mol Sci. 2023;24(21):15693. doi:10.3390/ijms242115693
23. Schaaf MB, Garg AD, Agostinis P. Defining the role of the tumor vasculature in antitumor immunity and immunotherapy. Cell Death Dis. 2018;9(2):115. doi:10.1038/s41419-017-0061-0
24. Lim SA, Wei J, Nguyen TLM, et al. Lipid signalling enforces functional specialization of Treg cells in tumours. Nature. 2021;591(7849):306–311. doi:10.1038/s41586-021-03235-6
25. Wang D, Ye Q, Gu H, Chen Z. The role of lipid metabolism in tumor immune microenvironment and potential therapeutic strategies. Front Oncol. 2022;12:984560. doi:10.3389/fonc.2022.984560
26. Su P, Wang Q, Bi E, et al. Enhanced lipid accumulation and metabolism are required for the differentiation and activation of tumor-associated macrophages. Cancer Res. 2020;80(7):1438–1450. doi:10.1158/0008-5472.CAN-19-2994
27. Zhao S, Peralta RM, Avina-Ochoa N, Delgoffe GM, Kaech SM. Metabolic regulation of T cells in the tumor microenvironment by nutrient availability and diet. Semin Immunol. 2021;52:101485. doi:10.1016/j.smim.2021.101485
28. Prendeville H, Lynch L. Diet, lipids, and antitumor immunity. Cell Mol Immunol. 2022;19(3):432–444. doi:10.1038/s41423-021-00781-x
29. Zhang Y, Kurupati R, Liu L, et al. Enhancing CD8+ T cell fatty acid catabolism within a metabolically challenging tumor microenvironment increases the efficacy of melanoma immunotherapy. Cancer Cell. 2017;32(3):377–391.e9. doi:10.1016/j.ccell.2017.08.004
30. Ma X, Xiao L, Liu L, et al. CD36-mediated ferroptosis dampens intratumoral CD8+ T cell effector function and impairs their antitumor ability. Cell Metab. 2021;33(5):1001–1012.e5. doi:10.1016/j.cmet.2021.02.015
31. Xu S, Chaudhary O, Rodríguez-Morales P, et al. Uptake of oxidized lipids by the scavenger receptor CD36 promotes lipid peroxidation and dysfunction in CD8+ T cells in tumors. Immunity. 2021;54(7):1561–1577.e7. doi:10.1016/j.immuni.2021.05.003
32. Ping Y, Shan J, Qin H, et al. PD-1 signaling limits expression of phospholipid phosphatase 1 and promotes intratumoral CD8+ T cell ferroptosis. Immunity. 2024;57(9):2122–2139.e9. doi:10.1016/j.immuni.2024.08.003
33. Kado T, Nawaz A, Takikawa A, Usui I, Tobe K. Linkage of CD8+ T cell exhaustion with high-fat diet-induced tumourigenesis. Sci Rep. 2019;9(1):12284. doi:10.1038/s41598-019-48678-0
34. Di Conza G, Tsai CH, Gallart-Ayala H, et al. Tumor-induced reshuffling of lipid composition on the endoplasmic reticulum membrane sustains macrophage survival and pro-tumorigenic activity. Nat Immunol. 2021;22(11):1403–1415. doi:10.1038/s41590-021-01047-4
35. Goossens P, Rodriguez-Vita J, Etzerodt A, et al. Membrane cholesterol efflux drives tumor-associated macrophage reprogramming and tumor progression. Cell Metab. 2019;29(6):1376–1389.e4. doi:10.1016/j.cmet.2019.02.016
36. Michelet X, Dyck L, Hogan A, et al. Metabolic reprogramming of natural killer cells in obesity limits antitumor responses. Nat Immunol. 2018;19(12):1330–1340. doi:10.1038/s41590-018-0251-7
37. Bosteels V, Maréchal S, De Nolf C, et al. LXR signaling controls homeostatic dendritic cell maturation. Sci Immunol. 2023;8(83):eadd3955. doi:10.1126/sciimmunol.add3955
38. Dennis EA, Norris PC. Eicosanoid storm in infection and inflammation. Nat Rev Immunol. 2015;15(8):511–523. doi:10.1038/nri3859
39. Zelenay S, van der Veen AG, Böttcher JP, et al. Cyclooxygenase-dependent tumor growth through evasion of immunity. Cell. 2015;162(6):1257–1270. doi:10.1016/j.cell.2015.08.015
40. Chen JH, Perry CJ, Tsui YC, et al. Prostaglandin E2 and programmed cell death 1 signaling coordinately impair CTL function and survival during chronic viral infection. Nat Med. 2015;21(4):327–334. doi:10.1038/nm.3831
41. Böttcher JP, Bonavita E, Chakravarty P, et al. NK cells stimulate recruitment of cDC1 into the tumor microenvironment promoting cancer immune control. Cell. 2018;172(5):1022–1037.e14. doi:10.1016/j.cell.2018.01.004
42. Miao J, Lu X, Hu Y, et al. Prostaglandin E2 and PD-1 mediated inhibition of antitumor CTL responses in the human tumor microenvironment. Oncotarget. 2017;8(52):89802–89810. doi:10.18632/oncotarget.21155
43. Matsuura R, Punyawatthananukool S, Kawakami R, Mikami N, Sakaguchi S, Narumiya S. Prostaglandin E2 -EP2/EP4 signaling induces the tumor-infiltrating Treg phenotype for tumor growth. Proc Natl Acad Sci. 2025;122(49):e2424251122. doi:10.1073/pnas.2424251122
44. Bonavita E, Bromley CP, Jonsson G, et al. Antagonistic inflammatory phenotypes dictate tumor fate and response to immune checkpoint blockade. Immunity. 2020;53(6):1215–1229. doi:10.1016/j.immuni.2020.10.020
45. Serhan CN, Levy BD. Resolvins in inflammation: emergence of the pro-resolving superfamily of mediators. J Clin Invest. 2018;128(7):2657–2669. doi:10.1172/JCI97943
46. Panigrahy D, Edin ML, Lee CR, et al. Epoxyeicosanoids stimulate multiorgan metastasis and tumor dormancy escape in mice. J Clin Invest. 2012;122(1):178–191. doi:10.1172/JCI58128
47. Spencer CN, McQuade JL, Gopalakrishnan V, et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science. 2021;374(6575):1632–1640. doi:10.1126/science.aaz7015
48. Makki K, Deehan EC, Walter J, Bäckhed F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe. 2018;23(6):705–715. doi:10.1016/j.chom.2018.05.012
49. He Y, Fu L, Li Y, et al. Gut microbial metabolites facilitate anticancer therapy efficacy by modulating cytotoxic CD8+ T cell immunity. Cell Metab. 2021;33(5):988–1000.e7. doi:10.1016/j.cmet.2021.03.002
50. Trompette A, Gollwitzer ES, Pattaroni C, et al. Dietary fiber confers protection against flu by shaping Ly6c− patrolling monocyte hematopoiesis and CD8+ T cell metabolism. Immunity. 2018;48(5):992–1005.e8. doi:10.1016/j.immuni.2018.04.022
51. Luu M, Riester Z, Baldrich A, et al. Microbial short-chain fatty acids modulate CD8+ T cell responses and improve adoptive immunotherapy for cancer. Nat Commun. 2021;12(1):4077. doi:10.1038/s41467-021-24331-1
52. Bachem A, Makhlouf C, Binger KJ, et al. Microbiota-derived short-chain fatty acids promote the memory potential of antigen-activated CD8+ T cells. Immunity. 2019;51(2):285–297.e5. doi:10.1016/j.immuni.2019.06.002
53. Kaymak I, Watson MJ, Oswald BM, et al. ACLY and ACSS2 link nutrient-dependent chromatin accessibility to CD8 T cell effector responses. J Exp Med. 2024;221(9):e20231820. doi:10.1084/jem.20231820
54. Zhang H, Tang K, Ma J, et al. Ketogenesis-generated β-hydroxybutyrate is an epigenetic regulator of CD8+ T-cell memory development. Nat Cell Biol. 2020;22(1):18–25. doi:10.1038/s41556-019-0440-0
55. Zhu X, Li K, Liu G, et al. Microbial metabolite butyrate promotes anti-PD-1 antitumor efficacy by modulating T cell receptor signaling of cytotoxic CD8 T cell. Gut Microbes. 2023;15(2):2249143. doi:10.1080/19490976.2023.2249143
56. Pittet MJ, Michielin O, Migliorini D. Clinical relevance of tumour-associated macrophages. Nat Rev Clin Oncol. 2022;19(6):402–421. doi:10.1038/s41571-022-00620-6
57. Viola A, Munari F, Sánchez-Rodríguez R, Scolaro T, Castegna A. The metabolic signature of macrophage responses. Front Immunol. 2019;10:1462. doi:10.3389/fimmu.2019.01462
58. Vitale I, Manic G, Coussens LM, Kroemer G, Galluzzi L. Macrophages and metabolism in the tumor microenvironment. Cell Metab. 2019;30(1):36–50. doi:10.1016/j.cmet.2019.06.001
59. Makowski L, Chaib M, Rathmell JC. Immunometabolism: from basic mechanisms to translation. Immunol Rev. 2020;295(1):5–14. doi:10.1111/imr.12858
60. Liu J, Gao M, Yang Z, et al. Macrophages and metabolic reprograming in the tumor microenvironment. Front Oncol. 2022;12:795159. doi:10.3389/fonc.2022.795159
61. Williams NC, O’Neill LAJ. A role for the krebs cycle intermediate citrate in metabolic reprogramming in innate immunity and inflammation. Front Immunol. 2018;9:141. doi:10.3389/fimmu.2018.00141
62. Canton M, Sánchez-Rodríguez R, Spera I, et al. Reactive oxygen species in macrophages: sources and targets. Front Immunol. 2021;12:734229. doi:10.3389/fimmu.2021.734229
63. Cameron AM, Castoldi A, Sanin DE, et al. Inflammatory macrophage dependence on NAD+ salvage is a consequence of reactive oxygen species–mediated DNA damage. Nat Immunol. 2019;20(4):420–432. doi:10.1038/s41590-019-0336-y
64. Herb M, Schramm M. Functions of ROS in macrophages and antimicrobial immunity. Antioxidants. 2021;10(2):313. doi:10.3390/antiox10020313
65. Jin K, Qian C, Lin J, Liu B. Cyclooxygenase-2-Prostaglandin E2 pathway: a key player in tumor-associated immune cells. Front Oncol. 2023;13:1099811. doi:10.3389/fonc.2023.1099811
66. Ching MM, Reader J, Fulton AM. Eicosanoids in cancer: prostaglandin E2 receptor 4 in cancer therapeutics and immunotherapy. Front Pharmacol. 2020;11:819. doi:10.3389/fphar.2020.00819
67. Sokhi UK, Yuan R, Mishra B, et al. Opposing regulation of TNF responses by IFN-γ and a PGE2-cAMP axis that is apparent in rheumatoid and immune checkpoint inhibitor-induced arthritis human IL-1β+ macrophages. eLife. 2025;14:RP104367. doi:10.7554/eLife.104367
68. Zingg J, Hasan ST, Nakagawa K, et al. Modulation of cAMP levels by high-fat diet and curcumin and regulatory effects on CD36/FAT scavenger receptor/fatty acids transporter gene expression. BioFactors. 2017;43(1):42–53. doi:10.1002/biof.1307
69. Di Biase S, Lee C, Brandhorst S, et al. Fasting-mimicking diet reduces HO-1 to promote T cell-mediated tumor cytotoxicity. Cancer Cell. 2016;30(1):136–146. doi:10.1016/j.ccell.2016.06.005
70. McDonnell E, Crown SB, Fox DB, et al. Lipids reprogram metabolism to become a major carbon source for histone acetylation. Cell Rep. 2016;17(6):1463–1472. doi:10.1016/j.celrep.2016.10.012
71. Etzerodt A, Tsalkitzi K, Maniecki M, et al. Specific targeting of CD163+ TAMs mobilizes inflammatory monocytes and promotes T cell–mediated tumor regression. J Exp Med. 2019;216(10):2394–2411. doi:10.1084/jem.20182124
72. Muri J, Kopf M. Redox regulation of immunometabolism. Nat Rev Immunol. 2021;21(6):363–381. doi:10.1038/s41577-020-00478-8
73. Savina A, Jancic C, Hugues S, et al. NOX2 controls phagosomal pH to regulate antigen processing during crosspresentation by dendritic cells. Cell. 2006;126(1):205–218. doi:10.1016/j.cell.2006.05.035
74. Zhang T, Yin W, Zhao Y, et al. NOX2 enzyme mimicking nano-networks regulate tumor-associated macrophages to initiate both innate and adaptive immune effects. Adv Healthc Mater. 2024;13(20):2302387. doi:10.1002/adhm.202302387
75. Galluzzi L, Vitale I, Warren S, et al. Consensus guidelines for the definition, detection and interpretation of immunogenic cell death. J Immunother Cancer. 2020;8(1):e000337. doi:10.1136/jitc-2019-000337
76. Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, Sancho D. Dendritic cells in cancer immunology and immunotherapy. Nat Rev Immunol. 2020;20(1):7–24. doi:10.1038/s41577-019-0210-z
77. Choi M, Shin J, Lee CE, et al. Immunogenic cell death in cancer immunotherapy. BMB Rep. 2023;56(5):275–286. doi:10.5483/BMBRep.2023-0024
78. Krysko DV, Garg AD, Kaczmarek A, Krysko O, Agostinis P, Vandenabeele P. Immunogenic cell death and DAMPs in cancer therapy. Nat Rev Cancer. 2012;12(12):860–875. doi:10.1038/nrc3380
79. Gawlak-Socka S, Kowalczyk E, Wiktorowska-Owczarek A. Unfolded protein response at the crossroads: integrating endoplasmic reticulum stress with cellular stress networks. Int J Mol Sci. 2026;27(4):1986. doi:10.3390/ijms27041986
80. Lee AS. Glucose-regulated proteins in cancer: molecular mechanisms and therapeutic potential. Nat Rev Cancer. 2014;14(4):263–276. doi:10.1038/nrc3701
81. Fu Z, Li M, Zhou H, et al. Endoplasmic reticulum stress orchestrates tumor metabolism and immunity: new insights into immunometabolic therapeutics. Front Immunol. 2025;16:1674163. doi:10.3389/fimmu.2025.1674163
82. Hangai S, Ao T, Kimura Y, et al. PGE2 induced in and released by dying cells functions as an inhibitory DAMP. Proc Natl Acad Sci. 2016;113(14):3844–3849. doi:10.1073/pnas.1602023113
83. Hayashi K, Nikolos F, Lee YC, et al. Tipping the immunostimulatory and inhibitory DAMP balance to harness immunogenic cell death. Nat Commun. 2020;11(1):6299. doi:10.1038/s41467-020-19970-9
84. Liao P, Wang W, Wang W, et al. CD8+ T cells and fatty acids orchestrate tumor ferroptosis and immunity via ACSL4. Cancer Cell. 2022;40(4):365–378.e6. doi:10.1016/j.ccell.2022.02.003
85. Doll S, Proneth B, Tyurina YY, et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat Chem Biol. 2017;13(1):91–98. doi:10.1038/nchembio.2239
86. Kagan VE, Mao G, Qu F, et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat Chem Biol. 2017;13(1):81–90. doi:10.1038/nchembio.2238
87. Han C, Ge M, Xing P, et al. Cystine deprivation triggers CD36-mediated ferroptosis and dysfunction of tumor infiltrating CD8+ T cells. Cell Death Dis. 2024;15(2):145. doi:10.1038/s41419-024-06503-1
88. Bersuker K, Hendricks JM, Li Z, et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature. 2019;575(7784):688–692. doi:10.1038/s41586-019-1705-2
89. Reticker-Flynn NE, Zhang W, Belk JA, et al. Lymph node colonization induces tumor-immune tolerance to promote distant metastasis. Cell. 2022;185(11):1924–1942.e23. doi:10.1016/j.cell.2022.04.019
90. You Z, Chi H. Lipid metabolism in dendritic cell biology. Immunol Rev. 2023;317(1):137–151. doi:10.1111/imr.13215
91. Veglia F, Tyurin VA, Mohammadyani D, et al. Lipid bodies containing oxidatively truncated lipids block antigen cross-presentation by dendritic cells in cancer. Nat Commun. 2017;8(1):2122. doi:10.1038/s41467-017-02186-9
92. Herber DL, Cao W, Nefedova Y, et al. Lipid accumulation and dendritic cell dysfunction in cancer. Nat Med. 2010;16(8):880–886. doi:10.1038/nm.2172
93. Bayerl F, Meiser P, Donakonda S, et al. Tumor-derived prostaglandin E2 programs cDC1 dysfunction to impair intratumoral orchestration of anti-cancer T cell responses. Immunity. 2023;56(6):1341–1358.e11. doi:10.1016/j.immuni.2023.05.011
94. Braun D, Longman RS, Albert ML. A two-step induction of indoleamine 2,3 dioxygenase (IDO) activity during dendritic-cell maturation. Blood. 2005;106(7):2375–2381. doi:10.1182/blood-2005-03-0979
95. Plata-Gómez AB, Ho PC. Age- and diet-instructed metabolic rewiring of the tumor–immune microenvironment. J Exp Med. 2025;222(6):e20241102. doi:10.1084/jem.20241102
96. Golonko A, Pienkowski T, Swislocka R, et al. Dietary factors and their influence on immunotherapy strategies in oncology: a comprehensive review. Cell Death Dis. 2024;15(4):254. doi:10.1038/s41419-024-06641-6
97. Liu J, Zhang J, Gao Y, et al. Barrier permeation and improved nanomedicine delivery in tumor microenvironments. Cancer Lett. 2023;562:216166. doi:10.1016/j.canlet.2023.216166
98. Jain RK. Vascular and interstitial barriers to delivery of therapeutic agents in tumors. Cancer Metastasis Rev. 1990;9(3):253–266. doi:10.1007/BF00046364
99. Bresalier RS, Sandler RS, Quan H, et al. Cardiovascular events associated with rofecoxib in a colorectal adenoma chemoprevention trial. N Engl J Med. 2005;352(11):1092–1102. doi:10.1056/NEJMoa050493
100. Bindu S, Mazumder S, Bandyopadhyay U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: a current perspective. Biochem Pharmacol. 2020;180:114147. doi:10.1016/j.bcp.2020.114147
101. Minchinton AI, Tannock IF. Drug penetration in solid tumours. Nat Rev Cancer. 2006;6(8):583–592. doi:10.1038/nrc1893
102. Kuznetsova AB, Kolesova EP, Parodi A, Zamyatnin AA, Egorova VS. Reprogramming tumor-associated macrophage using nanocarriers: new perspectives to halt cancer progression. Pharmaceutics. 2024;16(5):636. doi:10.3390/pharmaceutics16050636
103. Song Y, Huang Y, Zhou F, Ding J, Zhou W. Macrophage-targeted nanomedicine for chronic diseases immunotherapy. Chin Chem Lett. 2022;33(2):597–612. doi:10.1016/j.cclet.2021.08.090
104. Zhou Y, Yuan J, Xu K, Li S, Liu Y. Nanotechnology reprogramming metabolism for enhanced tumor immunotherapy. ACS Nano. 2024;18(3):1846–1864. doi:10.1021/acsnano.3c11260
105. Ren M, Zheng X, Gao H, Jiang A, Yao Y, He W. Nanomedicines targeting metabolism in the tumor microenvironment. Front Bioeng Biotechnol. 2022;10:943906. doi:10.3389/fbioe.2022.943906
106. Mrksich K, Padilla MS, Mitchell MJ. Breaking the final barrier: evolution of cationic and ionizable lipid structure in lipid nanoparticles to escape the endosome. Adv Drug Deliv Rev. 2024;214:115446. doi:10.1016/j.addr.2024.115446
107. Wang Y, Xu Y, Song J, et al. Tumor cell-targeting and tumor microenvironment–responsive nanoplatforms for the multimodal imaging-guided photodynamic/photothermal/chemodynamic treatment of cervical cancer. Int J Nanomed. 2024;19:5837–5858. doi:10.2147/IJN.S466042
108. Hossain F, Al-Khami AA, Wyczechowska D, et al. Inhibition of fatty acid oxidation modulates immunosuppressive functions of myeloid-derived suppressor cells and enhances cancer therapies. Cancer Immunol Res. 2015;3(11):1236–1247. doi:10.1158/2326-6066.CIR-15-0036
109. Zeng W, Li F, Jin S, Ho PC, Liu PS, Xie X. Functional polarization of tumor-associated macrophages dictated by metabolic reprogramming. J Exp Clin Cancer Res. 2023;42(1):245. doi:10.1186/s13046-023-02832-9
110. Yang W, Bai Y, Xiong Y, et al. Potentiating the antitumour response of CD8+ T cells by modulating cholesterol metabolism. Nature. 2016;531(7596):651–655. doi:10.1038/nature17412
111. Lee SSY, Li J, Tai JN, Ratliff TL, Park K, Cheng JX. Avasimibe encapsulated in human serum albumin blocks cholesterol esterification for selective cancer treatment. ACS Nano. 2015;9(3):2420–2432. doi:10.1021/nn504025a
112. Jin W, Xu S, Zhang H, et al. A CD36-targeting thermosensitive berberine nanogel blocks tumor lipid hijacking and potentiates anti-PD-L1 immunotherapy in triple-negative breast cancer. Mol Pharm. 2026;23(2):958–974. doi:10.1021/acs.molpharmaceut.5c01378
113. Kim W, Ye Z, Simonenko V, et al. Codelivery of TGFβ and Cox2 siRNA inhibits HCC by promoting T-cell penetration into the tumor and improves response to immune checkpoint inhibitors. NAR Cancer. 2024;6(1):zcad059. doi:10.1093/narcan/zcad059
114. Bödder J, Kok LM, Fauerbach JA, Flórez-Grau G, De Vries IJM. Tailored PGE2 immunomodulation of moDCs by nano-encapsulated EP2/EP4 antagonists. Int J Mol Sci. 2023;24(2):1392. doi:10.3390/ijms24021392
115. Sinha P, Clements VK, Fulton AM, Ostrand-Rosenberg S. Prostaglandin E2 promotes tumor progression by inducing myeloid-derived suppressor cells. Cancer Res. 2007;67(9):4507–4513. doi:10.1158/0008-5472.CAN-06-4174
116. Luan B, Yoon YS, Le Lay J, Kaestner KH, Hedrick S, Montminy M. CREB pathway links PGE2 signaling with macrophage polarization. Proc Natl Acad Sci. 2015;112(51):15642–15647. doi:10.1073/pnas.1519644112
117. Wang G, Wang D, Xia L, et al. Metal–phenolic nanomedicines targeting fatty acid metabolic reprogramming to overcome immunosuppression in radiometabolic cancer therapy. ACS Appl Mater Interf. 2025;17(5):7478–7488. doi:10.1021/acsami.4c21028
118. Han D, Ding B, Zheng P, et al. Albumin nanoparticles co-loaded with dual drugs for enhanced ferroptosis-based cancer therapy through modulating cholesterol metabolism. ACS Nano. 2026;20(2):2225–2238. doi:10.1021/acsnano.5c17428
119. Wang S, Shen Y, Hu B, et al. Light-activated CD36-targeted cuproptosi triggers metabolic-immune synergy for precision breast cancer therapy. ACS Appl Mater Interf. 2025;17(36):50317–50336. doi:10.1021/acsami.5c11318
120. Luo J, Xu L, Feng J, et al. Tumor microenvironment-activated and ROS-augmented nanoplatform amplified PDT against colorectal cancer through impairing GPX4 to induce ferroptosis. ACS Appl Mater Interf. 2025;17(29):41586–41596. doi:10.1021/acsami.5c05523
121. Liu J, Wu M, Yang C, et al. Synergizing ferroptosis suppressor protein 1 gene silencing and photodynamic therapy based on photosensitive lipid nanoparticles for colon cancer immunotherapy. ACS Nano. 2025;19(32):29341–29359. doi:10.1021/acsnano.5c06115
122. Feng Q, Qi F, Fang W, Hu P, Shi J. Ferroptosis to pyroptosis regulation by iron-based nanocatalysts for enhanced tumor immunotherapy. J Am Chem Soc. 2024;146(47):32403–32414. doi:10.1021/jacs.4c08304
123. Cheng X, Xu J, Cui Y, et al. Nanovesicles for lipid metabolism reprogram-enhanced ferroptosis and magnetotherapy of refractory tumors and inhibiting metastasis with activated innate immunity. ACS Nano. 2025;19(7):7213–7230. doi:10.1021/acsnano.4c16981
124. Liu Y, He J, Li M, Ren K, Zhao Z. Inflammation-driven nanohitchhiker enhances postoperative immunotherapy by alleviating prostaglandin E2-mediated immunosuppression. ACS Appl Mater Interf. 2024;16(6):6879–6893. doi:10.1021/acsami.3c17357
125. Wang L, Ding K, Zheng C, et al. Detachable nanoparticle-enhanced chemoimmunotherapy based on precise killing of tumor seeds and normalizing the growing soil strategy. Nano Lett. 2020;20(9):6272–6280. doi:10.1021/acs.nanolett.0c01415
126. Montecillo-Aguado M, Tirado-Rodriguez B, Antonio-Andres G, et al. Omega-6 polyunsaturated fatty acids enhance tumor aggressiveness in experimental lung cancer model: important role of oxylipins. Int J Mol Sci. 2022;23(11):6179. doi:10.3390/ijms23116179
127. Panigrahy D, Wang D, Chen A, et al. Dual blockade of EP2 and EP4 signaling is required for optimal immune activation and antitumor activity against prostaglandin-expressing tumors. Cancer Res Commun. 2023;3(8):1486–1500. doi:10.1158/2767-9764.CRC-23-0249
128. Boumelha J, De Castro A, Bah N, et al. CRISPR–Cas9 screening identifies KRAS-induced COX2 as a driver of immunotherapy resistance in lung cancer. Cancer Res. 2024;84(14):2231–2246. doi:10.1158/0008-5472.CAN-23-2627
129. Tiwary S, Hsu KS, Goldfarbmuren KC, Xia Z, Berzofsky JA. High levels of endogenous omega-3 fatty acids promote dendritic cell antigen presentation and improve dendritic cell–based cancer vaccine efficacy in mice. Cancer Immunol Res. 2025;13(10):1609–1622. doi:10.1158/2326-6066.CIR-24-0927
130. Elisia I, Yeung M, Kowalski S, et al. Omega 3 supplementation reduces C-reactive protein, prostaglandin E2 and the granulocyte/lymphocyte ratio in heavy smokers: an open-label randomized crossover trial. Front Nutr. 2022;9:1051418. doi:10.3389/fnut.2022.1051418
131. Harre P, Hohenberger K, Krammer S, et al. Dietary ω−3 polyunsaturated fatty acids (PUFAs) reduce cholesterol-driven non-small cell lung cancer (NSCLC) progression in mouse models of disease. Commun Med. 2025;5(1):432. doi:10.1038/s43856-025-01193-y
132. Kelly AG, Wang W, Rothenberger E, et al. Enhancing cancer immunotherapy via inhibition of soluble epoxide hydrolase. Proc Natl Acad Sci. 2024;121(7):e2314085121. doi:10.1073/pnas.2314085121
133. Qian R, Cao G, Su W, et al. Enhanced sensitivity of tumor cells to autophagy inhibitors using fasting-mimicking diet and targeted lysosomal delivery nanoplatform. Nano Lett. 2022;22(22):9154–9162. doi:10.1021/acs.nanolett.2c03890
134. Licha D, Vidali S, Aminzadeh-Gohari S, et al. Untargeted metabolomics reveals molecular effects of ketogenic diet on healthy and tumor xenograft mouse models. Int J Mol Sci. 2019;20(16):3873. doi:10.3390/ijms20163873
135. Becker S, Momoh J, Biancacci I, et al. Intermittent fasting primes the tumor microenvironment and improves nanomedicine delivery in hepatocellular carcinoma. Small. 2023;19(43):2208042. doi:10.1002/smll.202208042
136. Chiang ST, Chen Q, Han T, et al. Biomimetic nanovesicles synergize with short-term fasting for enhanced chemotherapy of triple-negative breast cancer. ACS Nano. 2024;18(50):33875–33889. doi:10.1021/acsnano.4c07074
137. Larson CR, Mandloi A, Acharyya S, Carstens JL. The tumor microenvironment across four dimensions: assessing space and time in cancer biology. Front Immunol. 2025;16:1554114. doi:10.3389/fimmu.2025.1554114
138. Huang Z, Chen J, Zhu T, et al. Cancer-associated fibroblasts in the tumor microenvironment: heterogeneity, crosstalk mechanisms, and therapeutic implications. Mol Cancer. 2025;25(1):19. doi:10.1186/s12943-025-02533-1
139. Heldin CH, Rubin K, Pietras K, Östman A. High interstitial fluid pressure—an obstacle in cancer therapy. Nat Rev Cancer. 2004;4(10):806–813. doi:10.1038/nrc1456
140. Gubbala VB, Jytosana N, Trinh VQ, et al. Eicosanoids in the pancreatic tumor microenvironment—a multicellular, multifaceted progression. Gastro Hep Adv. 2022;1(4):682–697. doi:10.1016/j.gastha.2022.02.007
141. Wu L, Zhang X, Zheng L, et al. RIPK3 orchestrates fatty acid metabolism in tumor-associated macrophages and hepatocarcinogenesis. Cancer Immunol Res. 2020;8(5):710–721. doi:10.1158/2326-6066.CIR-19-0261
142. Wong C, Stylianopoulos T, Cui J, et al. Multistage nanoparticle delivery system for deep penetration into tumor tissue. Proc Natl Acad Sci. 2011;108(6):2426–2431. doi:10.1073/pnas.1018382108
143. Ruoslahti E. Tumor penetrating peptides for improved drug delivery. Adv Drug Deliv Rev. 2017;110–111:3–12. doi:10.1016/j.addr.2016.03.008
144. Kang S, Lee S, Park S. iRGD peptide as a tumor-penetrating enhancer for tumor-targeted drug delivery. Polymers. 2020;12(9):1906. doi:10.3390/polym12091906
145. Zhao X, Yang X, Wang X, et al. Penetration cascade of size switchable nanosystem in desmoplastic stroma for improved pancreatic cancer therapy. ACS Nano. 2021;15(9):14149–14161. doi:10.1021/acsnano.0c08860
146. Nieman KM, Kenny HA, Penicka CV, et al. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat Med. 2011;17(11):1498–1503. doi:10.1038/nm.2492
147. Kir S, White JP, Kleiner S, et al. Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nature. 2014;513(7516):100–104. doi:10.1038/nature13528
148. Zhou C, He X, Tong C, et al. Cancer-associated adipocytes promote the invasion and metastasis in breast cancer through LIF/CXCLs positive feedback loop. Int J Biol Sci. 2022;18(4):1363–1380. doi:10.7150/ijbs.65227
149. Mukherjee A, Chiang CY, Daifotis HA, et al. Adipocyte-induced FABP4 expression in ovarian cancer cells promotes metastasis and mediates carboplatin resistance. Cancer Res. 2020;80(8):1748–1761. doi:10.1158/0008-5472.CAN-19-1999
150. Wu H, Han Y, Rodriguez Sillke Y, et al. Lipid droplet-dependent fatty acid metabolism controls the immune suppressive phenotype of tumor-associated macrophages. EMBO Mol Med. 2019;11(11):e10698. doi:10.15252/emmm.201910698
151. Wang Z, Zhang X, Fang C, et al. Immunotherapy and the ovarian cancer microenvironment: exploring potential strategies for enhanced treatment efficacy. Immunology. 2024;173(1):14–32. doi:10.1111/imm.13793
152. Qian Y, Qiao S, Dai Y, et al. Molecular-targeted immunotherapeutic strategy for melanoma via dual-targeting nanoparticles delivering small interfering RNA to tumor-associated macrophages. ACS Nano. 2017;11(9):9536–9549. doi:10.1021/acsnano.7b05465
153. Rodell CB, Arlauckas SP, Cuccarese MF, et al. TLR7/8-agonist-loaded nanoparticles promote the polarization of tumour-associated macrophages to enhance cancer immunotherapy. Nat Biomed Eng. 2018;2(8):578–588. doi:10.1038/s41551-018-0236-8
154. Bolte LA, Lee KA, Björk JR, et al. Association of a mediterranean diet with outcomes for patients treated with immune checkpoint blockade for advanced melanoma. JAMA Oncol. 2023;9(5):705. doi:10.1001/jamaoncol.2022.7753
155. Hatae R, Chamoto K, Kim YH, et al. Combination of host immune metabolic biomarkers for the PD-1 blockade cancer immunotherapy. JCI Insight. 2020;5(2):e133501. doi:10.1172/jci.insight.133501
156. Pecci F, Cantini L, Cognigni V, et al. Prognostic impact of blood lipid profile in patients with advanced solid tumors treated with immune checkpoint inhibitors: a multicenter cohort study. Oncologist. 2024;29(3):e372–e381. doi:10.1093/oncolo/oyad273
157. Tanaka I, Yano Y, Mori M, Manabe S, Fukuo K. Impact of serum eicosapentaenoic acid/arachidonic acid ratio on overall survival in lung cancer patients treated with pembrolizumab: a pilot study. Sci Rep. 2024;14(1):1384. doi:10.1038/s41598-024-51967-y
158. Fumet JD, Latour C, Nuttin L, et al. Tumor-associated macrophages produce PGE2 to promote CD8+ T-cell exhaustion and drive resistance to PD-L1 blockade in microsatellite-stable colorectal cancer. Cancer Res. 2026;86(3):785–801. doi:10.1158/0008-5472.CAN-25-0079
159. Lacher SB, Dörr J, De Almeida GP, et al. PGE2 limits effector expansion of tumour-infiltrating stem-like CD8+ T cells. Nature. 2024;629(8011):417–425. doi:10.1038/s41586-024-07254-x
160. Kong W, Yen JH, Vassiliou E, Adhikary S, Toscano MG, Ganea D. Docosahexaenoic acid prevents dendritic cell maturation and in vitro and in vivo expression of the IL-12 cytokine family. Lipids Health Dis. 2010;9(1):12. doi:10.1186/1476-511X-9-12
161. Holzlechner M, Eugenin E, Prideaux B. Mass spectrometry imaging to detect lipid biomarkers and disease signatures in cancer. Cancer Rep. 2019;2(6):e1229. doi:10.1002/cnr2.1229
162. Irún P, Carrera-Lasfuentes P, Sánchez-Luengo M, et al. Pharmacokinetics and changes in lipid mediator profiling after consumption of specialized pro-resolving lipid-mediator-enriched marine oil in healthy subjects. Int J Mol Sci. 2023;24(22):16143. doi:10.3390/ijms242216143
163. Jiang H, Li XS, Yang Y, Qi RX. Plasma lipidomics profiling in predicting the chemo-immunotherapy response in advanced non-small cell lung cancer. Front Oncol. 2024;14:1348164. doi:10.3389/fonc.2024.1348164
164. Ghadami S, Dellinger K. The lipid composition of extracellular vesicles: applications in diagnostics and therapeutic delivery. Front Mol Biosci. 2023;10:1198044. doi:10.3389/fmolb.2023.1198044
165. Greening DW, Xu R, Rai A, Suwakulsiri W, Chen M, Simpson RJ. Clinical relevance of extracellular vesicles in cancer — therapeutic and diagnostic potential. Nat Rev Clin Oncol. 2025;22(12):924–952. doi:10.1038/s41571-025-01074-2
166. Espiau-Romera P, Gordo-Ortiz A, Ortiz-de-Solórzano I, Sancho P. Metabolic features of tumor-derived extracellular vesicles: challenges and opportunities. Extracell Vesicles Circ Nucleic Acids. 2024;5(3):555–570. doi:10.20517/evcna.2024.12
167. Nomura M, Nagatomo R, Doi K, et al. Association of short-chain fatty acids in the gut microbiome with clinical response to treatment with Nivolumab or Pembrolizumab in patients with solid cancer tumors. JAMA Network Open. 2020;3(4):e202895. doi:10.1001/jamanetworkopen.2020.2895
168. Iwasaki T, Shirota H, Hishinuma E, et al. Plasma lysophosphatidylcholine levels correlate with prognosis and immunotherapy response in squamous cell carcinoma. Int J Mol Sci. 2025;26(15):7528. doi:10.3390/ijms26157528
169. Zhan Z, Wang X, He M, et al. Identification of plasma lipidomic biomarkers for prognostic stratification in advanced gastric cancer treated with PD-1 inhibitor plus chemotherapy. Front Immunol. 2026;17:1714472. doi:10.3389/fimmu.2026.1714472
170. Tou Y, Sun W, Liu P, Sun J, Ma T, Zhang YH. Structural lipidomics uncovers C═C location-specific lipid signatures and the response to immune checkpoint inhibitor in dMMR/MSI-H colorectal cancer. ACS Meas Sci Au. 2026;
171. Westheim AJF, Stoffels LM, Dubois LJ, et al. Fatty acids as a tool to boost cancer immunotherapy efficacy. Front Nutr. 2022;9:868436. doi:10.3389/fnut.2022.868436
172. Bodur M, Yilmaz B, Ağagündüz D, Ozogul Y. Immunomodulatory effects of omega-3 fatty acids: mechanistic insights and health implications. Mol Nutr Food Res. 2025;69(10):e202400752. doi:10.1002/mnfr.202400752
173. Alijani S, Hahn A, Harris WS, Schuchardt JP. Bioavailability of EPA and DHA in humans – a comprehensive review. Prog Lipid Res. 2025;97:101318. doi:10.1016/j.plipres.2024.101318
174. Fu Y, Wang Y, Gao H, et al. Associations among dietary omega-3 polyunsaturated fatty acids, the gut microbiota, and intestinal immunity. Mediators Inflamm. 2021;2021(1):8879227. doi:10.1155/2021/8879227
175. Kazantzis M, Stahl A. Fatty acid transport proteins, implications in physiology and disease. Biochim Biophys Acta BBA. 2012;1821(5):852–857. doi:10.1016/j.bbalip.2011.09.010
176. Monopoli MP, Åberg C, Salvati A, Dawson KA. Biomolecular coronas provide the biological identity of nanosized materials. Nat Nanotechnol. 2012;7(12):779–786. doi:10.1038/nnano.2012.207
177. Cheng YH, He C, Riviere JE, Monteiro-Riviere NA, Lin Z. Meta-analysis of nanoparticle delivery to tumors using a physiologically based pharmacokinetic modeling and simulation approach. ACS Nano. 2020;14(3):3075–3095. doi:10.1021/acsnano.9b08142
178. Sterin EH, Weinstein LA, Tiwari A, et al. Influence of the protein corona on hematopoietic stem and progenitor cell uptake and macrophage clearance of membrane-wrapped nanoparticles. Proc Natl Acad Sci. 2025;122(40):e2507922122. doi:10.1073/pnas.2507922122
179. Walkey CD, Chan WCW. Understanding and controlling the interaction of nanomaterials with proteins in a physiological environment. Chem Soc Rev. 2012;41(7):2780–2799. doi:10.1039/C1CS15233E
180. Tsoi KM, MacParland SA, Ma XZ, et al. Mechanism of hard-nanomaterial clearance by the liver. Nat Mater. 2016;15(11):1212–1221. doi:10.1038/nmat4718
181. Kozma GT, Shimizu T, Ishida T, Szebeni J. Anti-PEG antibodies: properties, formation, testing and role in adverse immune reactions to PEGylated nano-biopharmaceuticals. Adv Drug Deliv Rev. 2020;154–155:163–175. doi:10.1016/j.addr.2020.07.024
182. Yang Q, Lai SK. Anti-
183. Jalil AR, Tobin MP, Discher DE. Suppressing or enhancing macrophage engulfment through the use of CD47 and related peptides. Bioconjug Chem. 2022;33(11):1989–1995. doi:10.1021/acs.bioconjchem.2c00019
184. Liu H, Su YY, Jiang XC, Gao JQ. Cell membrane-coated nanoparticles: a novel multifunctional biomimetic drug delivery system. Drug Deliv Transl Res. 2023;13(3):716–737. doi:10.1007/s13346-022-01252-0
185. Imanbekova M, Sharma M, Wachsmann-Hogiu S. On the dilemma of using single EV analysis for liquid biopsy: the challenge of low abundance of tumor EVs in blood. Theranostics. 2025;15(16):8031–8048. doi:10.7150/thno.115131
186. Luke JJ, Bao R, Sweis RF, Spranger S, Gajewski TF. WNT/β-catenin pathway activation correlates with immune exclusion across human cancers. Clin Cancer Res. 2019;25(10):3074–3083. doi:10.1158/1078-0432.CCR-18-1942
187. Gettinger S, Choi J, Hastings K, et al. Impaired HLA class I antigen processing and presentation as a mechanism of acquired resistance to immune checkpoint inhibitors in lung cancer. Cancer Discov. 2017;7(12):1420–1435. doi:10.1158/2159-8290.CD-17-0593
188. Chowell D, Morris LGT, Grigg CM, et al. Patient HLA class I genotype influences cancer response to checkpoint blockade immunotherapy. Science. 2018;359(6375):582–587. doi:10.1126/science.aao4572
189. Serini S, Cassano R, Trombino S, Calviello G. Nanomedicine-based formulations containing ω-3 polyunsaturated fatty acids: potential application in cardiovascular and neoplastic diseases. Int J Nanomed. 2019;14:2809–2828. doi:10.2147/IJN.S197499
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Stimulus Responsive Nanocarrier for Enhanced Antitumor Responses Against Hepatocellular Carcinoma
Zhang D, Song J, Jing Z, Qin H, Wu Y, Zhou J, Zang X
International Journal of Nanomedicine 2024, 19:13339-13355
Published Date: 10 December 2024
Immunotherapy Resistance and Therapeutic Strategies in PD-L1 High Expression Non-Small Cell Lung Cancer
Liu J, Cai Y, Liu J, Chen D, Wu X
OncoTargets and Therapy 2025, 18:953-966
Published Date: 29 August 2025
Key Clinical Frontiers of mRNA Loaded Lipid Nanoparticles in Cancer Vaccines
Cao L, Min J, Yu M, Zhang Z, Yuan D, Chen D
International Journal of Nanomedicine 2025, 20:14935-14953
Published Date: 12 December 2025
The Role of Bacterial Outer Membrane Vesicles in Tumor Development: Facts, Potential Applications and Future Perspectives
Gu Z, Zhang M, Shen F, Chen J, Gu Y, Gong Y, Xue Q, Zhou H, Liu Y, Shao S, Zhang Y, Bai J, Hua K, Zhou G, Ding J
International Journal of Nanomedicine 2026, 21:582600
Published Date: 11 March 2026
Nanodynamic Therapy in Colorectal Cancer: Engineering Precision Immunotherapy and Multimodal Synergy
Wang F, Zhang Z, Chen J, Qian L, Chen B, Zhai L, Zhao J, Chen T, Zhou J, Gou K, Zhao Z, Zhu X, Xiao Z
International Journal of Nanomedicine 2026, 21:598076
Published Date: 23 March 2026