Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Selective Modulation of Cutibacterium acnes Biofilms in Acne: Limitations of Conventional Therapies and Emerging Anti-Virulence Strategies
Authors Kusuma SAF ![]()
Received 2 May 2026
Accepted for publication 23 June 2026
Published 30 June 2026 Volume 2026:19 621646
DOI https://doi.org/10.2147/CCID.S621646
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Sri Agung Fitri Kusuma
Department of Pharmaceutical Biology, Faculty of Pharmacy, Padjadjaran University, Sumedang, 45363, Indonesia
Correspondence: Sri Agung Fitri Kusuma, Department of Pharmaceutical Biology, Faculty of Pharmacy, Padjadjaran University, Sumedang, Jl. Raya Bandung-Sumedang Km. 21, Bandung, West Java, 45363, Indonesia, Email [email protected]
Abstract: Acne vulgaris is a multifactorial inflammatory skin disorder in which Cutibacterium acnes biofilm formation contributes to disease persistence, antimicrobial tolerance, and treatment failure. Conventional therapies primarily target bacterial viability but often fail to address biofilm-associated resilience and strain-specific virulence. Emerging strategies have therefore shifted toward the selective modulation of bacterial behavior rather than broad-spectrum eradication, with the aim of attenuating pathogenicity while minimizing disruption of the skin microbiome. This review critically evaluates selective modulation approaches targeting biofilm integrity, virulence pathways, and microbial ecology, including anti-virulence therapy, quorum-sensing inhibition, biofilm disruption, nanocarrier-based delivery systems, and microbiome-informed interventions. Preclinical studies suggest that these strategies may disrupt biofilm architecture, attenuate virulence factor expression, and potentially reduce selective pressure associated with conventional antimicrobial therapies. Approaches such as antimicrobial peptides, quorum-sensing inhibitors, and advanced delivery systems have demonstrated promising in vitro, ex vivo, and early preclinical outcomes; however, clinical evidence remains limited. Significant challenges remain, including insufficient in vivo validation, formulation instability, biofilm-associated delivery barriers, regulatory considerations, and limited long-term safety data. Overall, selective modulation represents a promising emerging framework for acne management, although its successful clinical translation will require robust clinical validation, improved disease-relevant models, and the integration of personalized strategies based on microbiome profiling and advanced delivery technologies. The infographic is divided into three sections: The Problem, Conceptual Biofilm Barrier and The Solution. The Problem highlights how biofilm formation leads to persistence, reduced antimicrobial effectiveness and treatment failure, using C. acnes biofilm as an example. The Conceptual Biofilm Barrier explains studies showing how a dense EPS matrix limits diffusion, with subpopulations that are metabolically active or tolerant, hindering antibiotic penetration. The Solution presents strategies for selective modulation: 1. Anti-virulence therapy to decrease virulence and inflammation. 2. Quorum sensing inhibition to disrupt bacterial communication. 3. Biofilm disruption to break down EPS and destabilize biofilms. 4. Advanced nanocarrier delivery for improved targeting and controlled release. 5. Microbiome modulation to target harmful strains while preserving the microbiome. The aim is to shift from broad antimicrobial use to targeted modulation for enhanced acne management.An infographic on biofilm formation, its barriers and solutions for acne management.
Keywords: anti-virulence, biofilm, Cutibacterium acnes, microbiome modulation, nanocarrier systems, quorum sensing
Introduction
Acne vulgaris is a common chronic inflammatory disorder of the pilosebaceous unit that affects up to 85% of adolescents and approximately 9.4% of the global population, making it one of the most prevalent dermatological conditions worldwide.1–6 The disease imposes a substantial physical and psychological burden, often leading to scarring, post-inflammatory hyperpigmentation, reduced quality of life, and the need for prolonged therapeutic management.7–9
Acne pathogenesis is multifactorial and involves sebum overproduction, follicular hyperkeratinization, inflammation, microbial colonization, genetic predisposition, dietary factors, hormonal influences, and immune dysregulation.10–12 In addition, disruption of the skin microbiome has emerged as a critical contributor to disease persistence and progression.11,13,14 The skin microbiome plays an essential role in maintaining cutaneous homeostasis by regulating immune responses, barrier integrity, lipid metabolism, and colonization resistance. Consequently, microbial dysbiosis has increasingly been recognized as both a pathogenic driver and a potential therapeutic target in acne vulgaris.11,13,14
Current therapeutic approaches, including topical and systemic antibiotics, retinoids, benzoyl peroxide, and adjunctive physical therapies, remain widely used and are supported by clinical guidelines.15 However, many patients experience incomplete responses, treatment relapse, adverse effects, and poor long-term adherence.15,16 Of particular concern is the rising prevalence of antibiotic-resistant Cutibacterium acnes strains due to prolonged antimicrobial exposure.17–20 Moreover, conventional therapies often act in a non-selective manner and may further disrupt the skin microbiota, potentially exacerbating dysbiosis.16–18
Emerging evidence suggests that acne severity is not primarily driven by an increased abundance of C. acnes, but rather by strain-level dysbiosis characterized by reduced phylotype diversity and the predominance of virulent strains such as IA1.11,14 These strains exhibit enhanced inflammatory potential, increased biofilm-forming capacity, and greater tolerance to antimicrobial therapy.11,14,21 Consequently, acne persistence is increasingly understood as a consequence of virulence-associated mechanisms and biofilm-mediated resilience rather than simple bacterial overgrowth.
Beyond antimicrobial resistance, bacterial persistence is strongly supported by adaptive mechanisms, including biofilm formation, quorum sensing, and virulence-associated gene expression.20,22 A central yet often underestimated driver of therapeutic failure is C. acnes’s ability to form biofilms within the pilosebaceous unit. Biofilm-associated cells exhibit markedly increased tolerance to antimicrobial agents, reduced metabolic activity, enhanced stress adaptation, and protection from host immune responses.21,23,24 These observations highlight a fundamental mismatch between conventional antimicrobial strategies and the biofilm-driven, virulence-mediated nature of acne pathogenesis.
Building on these limitations, recent research has shifted toward selective modulation strategies that target virulence pathways, biofilm integrity, and microbial ecology rather than broad-spectrum eradication of bacteria.14,25–30 Such approaches aim to attenuate pathogenic behavior while minimizing disruption of the resident microbiota. Accordingly, this review critically examines the limitations of conventional acne therapies through the lens of C. acnes biofilm biology and virulence mechanisms, and highlights selective modulation strategies as emerging therapeutic approaches to overcome treatment resistance and improve long-term acne management.
Literature Search Strategy and Selection Criteria
A comprehensive literature search was conducted across PubMed, Scopus, and ScienceDirect databases to identify relevant studies concerning Cutibacterium acnes biofilm dynamics and emerging therapeutic strategies in acne management, spanning from 2010 to 2026. Search terms included combinations of “Cutibacterium acnes,” “biofilm,” “anti-virulence,” “quorum sensing,” “microbiome,” and “targeted therapy.”
Studies were included if they were peer-reviewed English-language articles that provided mechanistic, microbiological, translational, or clinical insights relevant to C. acnes biofilms and emerging therapeutic approaches. Articles focusing on biofilm biology, virulence regulation, quorum sensing, microbiome modulation, biofilm-disrupting strategies, and advanced drug delivery systems were prioritized.
Exclusion criteria included conference abstracts, editorials, commentaries, non-English publications, duplicate records, studies lacking sufficient methodological detail, and articles not directly relevant to C. acnes biofilms or acne pathogenesis. To enhance transparency, the inclusion and exclusion criteria used in the literature selection are summarized in Table 1.
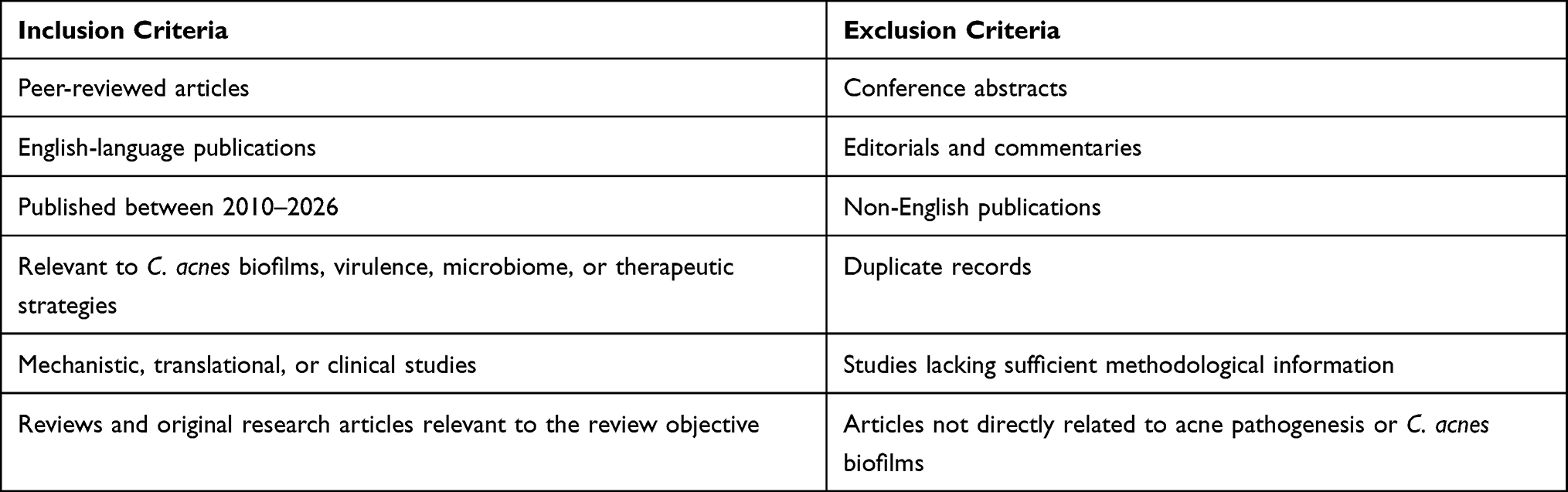 |
Table 1 Inclusion and Exclusion Criteria Used in Literature Selection |
This work is presented as a narrative critical review, focusing on the synthesis of mechanistic insights rather than a formal systematic review protocol, to provide a focused analysis of current and future therapeutic paradigms. Evidence was interpreted according to its translational relevance: direct clinical and human data were prioritized for clinical implications, C. acnes-specific experimental studies were used to support mechanistic statements, and evidence extrapolated from broader biofilm biology or non-follicular models was described cautiously as preliminary or hypothesis-generating.
The Biofilm Landscape in Acne
Biofilm formation by C. acnes represents a critical yet underrecognized factor in acne pathogenesis, contributing to persistence, immune evasion, and antimicrobial tolerance.21,24 Notably, bacterial density within a single follicular unit can reach up to 108 colony-forming units, creating a highly concentrated microenvironment that may favor quorum-sensing activation and biofilm stabilization.31 Biofilms are structured microbial communities embedded within a self-produced extracellular polymeric substance (EPS) matrix. In experimental C. acnes biofilm models, this matrix has been reported to contain polysaccharides, proteins, lipids, extracellular DNA, and biofilm-associated metabolites.32 Although follicular C. acnes macrocolonies and biofilm-like aggregates have been reported in acne lesions, the precise biochemical composition of the EPS matrix within the human pilosebaceous unit has not been directly characterized. Therefore, extrapolation from experimental biofilm models to the in vivo follicular environment should be interpreted with caution.
Importantly, biofilm formation has been proposed to contribute to the early stages of acne development. C. acnes biofilms have been described as a biological glue that may enhance cohesion between keratinocytes, thereby promoting retention of keratin and sebum within the follicular duct and facilitating comedone formation.31,33 This adhesive property may interfere with normal desquamation and contribute to follicular obstruction, which is considered a key initiating event in acne pathogenesis.
Biofilm development generally occurs through sequential stages, beginning with reversible bacterial adhesion to the follicular surface, followed by irreversible attachment, microcolony formation, EPS matrix production, maturation, and eventual dispersion. Early-stage biofilms are characterized by relatively active bacterial growth and dependence on adhesion processes, whereas mature biofilms possess a more established EPS matrix and increased structural complexity. These differences are clinically relevant because early biofilms may be more susceptible to adhesion-blocking or quorum-sensing-targeted interventions, while mature biofilms often require matrix-disrupting agents, advanced drug delivery systems, or combination therapies to overcome biofilm-associated tolerance.21,23,28
Beyond structural support, biofilms significantly enhance the pathogenic potential of C. acnes. Although C. acnes is a commensal organism present in both healthy and acne-affected skin, it can behave as an opportunistic pathogen under specific host and microenvironmental conditions. This transition is influenced by host susceptibility, changes in the follicular microenvironment, and strain-level dysbiosis, particularly the enrichment of acne-associated phylotypes such as IA1.14,34,35 These conditions promote biofilm formation and enhance the expression of virulence-associated factors, including lipases, adhesion proteins, porphyrins, and pro-inflammatory mediators, thereby contributing to follicular colonization, inflammation, and disease progression.33,34 In particular, lipase-mediated hydrolysis of sebum triglycerides generates free fatty acids that are both comedogenic and pro-inflammatory.34
At the molecular level, experimental analyses of C. acnes biofilm matrices have shown that the extracellular matrix is complex and functionally dynamic. In these model systems, polysaccharides can account for more than 60% of the matrix, alongside proteins, extracellular DNA, and metabolites such as porphyrins.32 The matrix not only provides structural integrity but also functions as a biochemical interface that facilitates nutrient retention, intercellular signaling, and protection against environmental stress. The presence of enzymes, including hydrolases and metabolic proteins, further suggests that the matrix may participate in metabolic adaptation and virulence regulation rather than serving as a passive scaffold.32 However, because these observations are primarily derived from experimental biofilm systems, their direct applicability to native follicular C. acnes biofilms should be interpreted cautiously.
Moreover, the biofilm matrix functions as a protective barrier that limits antimicrobial penetration and shields bacterial cells from host immune responses. It supports the formation of microenvironmental gradients of oxygen, nutrients, and signaling molecules, allowing subpopulations of bacteria to persist in metabolically inactive or stress-tolerant states. Transcriptomic evidence suggests that biofilm-embedded C. acnes cells may exhibit reduced metabolic activity and adaptive shifts in gene expression that are consistent with quasi-dormant or persister-like phenotypes. However, direct evidence linking these phenotypes to clinically relevant follicular biofilms remains limited.36 These features collectively enhance bacterial survival under adverse conditions and may contribute to the chronic and recurrent nature of acne, thereby reducing the effectiveness of conventional antimicrobial therapies. This protective effect is strongly associated with key structural components such as poly-N-acetylglucosamine (PNAG), which plays a central role in maintaining biofilm integrity and antimicrobial resistance. Disruption of PNAG has been shown to significantly enhance the efficacy of conventional agents such as benzoyl peroxide, leading to markedly improved bacterial eradication compared to monotherapy.29 Notably, acne severity is not solely determined by the abundance of C. acnes, but rather by strain-specific characteristics, including virulence factor expression and biofilm-forming capacity, highlighting the importance of strain-level and microbiome-based perspectives in acne pathogenesis.37 This heterogeneity is further supported by comparative analyses demonstrating that biofilm formation capacity varies significantly across C. acnes phylotypes, with IA1 strains exhibiting the highest biofilm-forming potential and strongest association with acne severity.38 These observations are further supported by studies suggesting that biofilm-associated C. acnes may contribute to reduced treatment responsiveness, as biofilm-embedded bacteria often exhibit greater tolerance to antimicrobial agents than their planktonic counterparts.21,23,25 Emerging evidence suggests that QS mechanisms may contribute to the regulation of biofilm development, virulence expression, and bacterial persistence in C. acnes.25,26 However, the complexity of these regulatory networks and their specific roles within human follicular biofilms remain incompletely understood.25 In addition, extracellular DNA and protein components are key determinants of biofilm structure and initial surface adhesion, further emphasizing the functional complexity of the matrix.38
Collectively, available evidence suggests that biofilm formation is an important contributor to acne persistence and therapeutic resistance. Biofilm-associated mechanisms may help explain the limited effectiveness of conventional treatment approaches that primarily target planktonic bacteria while inadequately addressing biofilm-associated tolerance and persistence. However, much of the current understanding is derived from in vitro studies and experimental biofilm models, whereas direct evidence from clinically relevant human follicular systems remains limited. Consequently, further investigation is needed to clarify the precise contribution of biofilm-associated mechanisms to acne pathogenesis and treatment outcomes. Nevertheless, these findings support continued exploration of therapeutic strategies that target biofilm architecture and virulence-associated pathways as complementary approaches to conventional acne management.
Critical Appraisal of Conventional Therapies
Current therapeutic approaches for acne, including topical and systemic antibiotics, retinoids, benzoyl peroxide, and adjunctive physical therapies, remain the cornerstone of clinical management.15 These treatments primarily aim to reduce bacterial load, normalize keratinization, and suppress inflammation. However, despite their widespread use, they frequently fail to achieve sustained remission, with many patients experiencing relapse following treatment discontinuation.15,16
A major limitation of conventional therapies lies in their inability to effectively target biofilm-associated C. acnes. Most antimicrobial agents are designed to act against planktonic bacterial populations, whereas C. acnes predominantly exists as biofilms in vivo.21,25 The EPS matrix acts as a pharmacokinetic barrier that restricts drug penetration, sequesters antimicrobial molecules, and reduces local drug concentrations at the site of bacterial cells.32 Consequently, concentrations effective against planktonic bacteria are often insufficient to eradicate biofilm-embedded populations.
Beyond this physical barrier, biofilm-associated C. acnes may exhibit metabolic and phenotypic adaptations that further compromise treatment efficacy. Transcriptomic evidence suggests that biofilm-embedded C. acnes cells exhibit reduced metabolic activity and adaptive shifts in gene expression consistent with quasi-dormant or persister-like phenotypes.36 However, direct experimental evidence demonstrating a causal relationship between these persister-like subpopulations and reduced antibiotic susceptibility in clinically relevant follicular C. acnes biofilms remains limited. Therefore, these phenotypes should be interpreted as potential contributors to antimicrobial tolerance rather than as directly proven mechanisms of treatment failure in acne.
Another critical limitation is the non-selective mode of action of conventional therapies. Broad-spectrum antimicrobial agents not only target pathogenic C. acnes strains but also disrupt commensal microbiota, leading to dysbiosis and impaired skin barrier function.17,18 Importantly, disruption of commensal microbial communities may reduce microbial competition and alter follicular niche conditions, which could theoretically favor the persistence or expansion of acne-associated C. acnes phylotypes. However, direct evidence demonstrating that conventional therapies specifically promote dominance of virulent C. acnes phylotypes remains limited.14,37 Consequently, indiscriminate microbial eradication may exacerbate microbial imbalance rather than restore homeostasis. Furthermore, the extensive and prolonged use of antibiotics has contributed to the global emergence of antimicrobial resistance in C. acnes.18,19 Resistance mechanisms include target site modification, efflux pump activation, and adaptive stress responses that reduce antibiotic susceptibility. Biofilm formation may further contribute to reduced antimicrobial susceptibility by limiting drug penetration, supporting stress-adapted subpopulations, and creating protective microenvironments.25
Collectively, these limitations reflect a fundamental mismatch between the mechanisms of conventional therapies and the biofilm-driven, virulence-mediated nature of C. acnes in vivo. The continued reliance on broad-spectrum approaches that primarily target bacterial viability appears increasingly insufficient in the context of biofilm-associated persistence and strain-specific pathogenicity. This disconnect underscores an urgent need to shift toward mechanism-based therapeutic strategies that specifically target biofilm integrity and bacterial behavior, rather than bacterial eradication alone.
Selective Modulation: An Emerging Therapeutic Framework
The growing recognition of biofilm-mediated persistence and strain-specific pathogenicity in acne has stimulated interest in therapeutic strategies that move beyond conventional broad-spectrum antimicrobial approaches toward more targeted, mechanism-based interventions. Notably, it is estimated that 60–80% of microbial infections are associated with biofilm formation, underscoring the clinical relevance of developing biofilm-targeted therapeutic strategies.28 Unlike traditional treatments that indiscriminately reduce bacterial load, emerging approaches focus on selectively modulating C. acnes behavior, disrupting biofilm integrity, and attenuating virulence while minimizing broad-spectrum disruption of commensal skin microbial communities.14,25
However, because C. acnes is a dominant inhabitant of sebaceous skin sites, interventions that alter its abundance, biofilm architecture, metabolic activity, or virulence-associated behavior may also influence the composition, spatial organization, and ecological dynamics of the surrounding skin microbiome. Therefore, the extent to which selective modulation strategies preserve microbiome balance remains to be established in clinically relevant human studies. This concept aligns with the broader framework of precision dermatology and microbiome-informed therapeutics, but its clinical implementation should be considered emerging rather than established.13,14,25
Anti-Virulence Strategies: Disarming Pathogenic Behavior
As a therapeutic concept, anti-virulence strategies aim to attenuate pathogenicity by targeting virulence-associated enzymes, regulatory pathways, or host–microbe interactions, rather than relying primarily on bacterial eradication.26,39 These approaches may reduce virulence factor expression, biofilm-associated activity, or inflammatory stimulation, and are therefore considered promising adjunctive strategies for biofilm-associated infections.
In the context of C. acnes, however, many investigated compounds exhibit both antimicrobial and virulence-associated inhibitory effects, making it difficult to classify them as purely anti-virulence agents. For example, Quercus salicina extract and its active fraction have been reported to inhibit C. acnes growth, biofilm formation, lipase activity, and virulence-associated gene expression.22 Notably, because the strongest biofilm inhibition was reported at 1000 μg/mL, whereas the MIC of the E1 fraction was 16 μg/mL, it is difficult to distinguish biofilm or virulence-associated inhibition from growth inhibition or reduced bacterial viability.22 Therefore, these effects should be interpreted as combined antimicrobial and virulence-associated inhibitory activity rather than definitive evidence of selective anti-virulence activity independent of growth inhibition. Accordingly, in this review, anti-virulence strategies are discussed as an emerging therapeutic concept, while individual agents are evaluated based on available evidence of their effects on bacterial growth, viability, biofilm formation, and virulence-associated pathways.
Experimental evidence also suggests that some skin microbiota-derived metabolites may modulate biofilm formation without substantially affecting bacterial viability in specific experimental settings. For instance, short-chain fatty acids produced by C. acnes have been shown to inhibit biofilm formation by Staphylococcus epidermidis at concentrations that did not significantly affect bacterial viability.40 Mechanistically, this inhibition was associated with interference in biofilm maturation rather than initial adhesion, including reduced polysaccharide and extracellular DNA accumulation during later stages of biofilm development.40 These findings support the possibility that certain biofilm-modulating effects may occur independently of growth inhibition in specific experimental models, although their direct relevance to C. acnes virulence control in acne requires further validation.
Key targets of anti-virulence therapy include adhesins, toxins, QS systems, and regulatory pathways such as two-component systems, many of which are involved in biofilm formation and persistence.39 This approach is particularly relevant in biofilm-associated infections, where microbial communities exhibit enhanced resilience and reduced antimicrobial susceptibility. Polymicrobial biofilms, for example, have been reported to be more resistant than monospecies biofilms due to cooperative interactions, increased expression of virulence factors, and the protective role of the EPS matrix. In addition, QS-mediated communication may coordinate virulence expression and biofilm maturation, contributing to bacterial persistence under therapeutic pressure.27
In the context of C. acnes, inhibition of lipase activity is particularly relevant, as lipase-mediated hydrolysis of sebum triglycerides generates free fatty acids that contribute to comedogenesis and inflammation. Clinically, inflammatory lesions, such as papules, have been identified as significant predictors of both acne scarring and post-inflammatory hyperpigmentation (OR = 5.65), underscoring the importance of early intervention targeting inflammatory pathways.41 By attenuating virulence-associated pathways, anti-virulence approaches may help reduce inflammatory burden while potentially limiting the selective pressure associated with bactericidal strategies. However, the efficacy of anti-virulence strategies may depend on the complexity of host–microbe interactions, the stability of virulence suppression in vivo, and whether the observed effects occur independently of bacterial growth inhibition. The major anti-virulence strategies discussed in this review are summarized in Table 2.
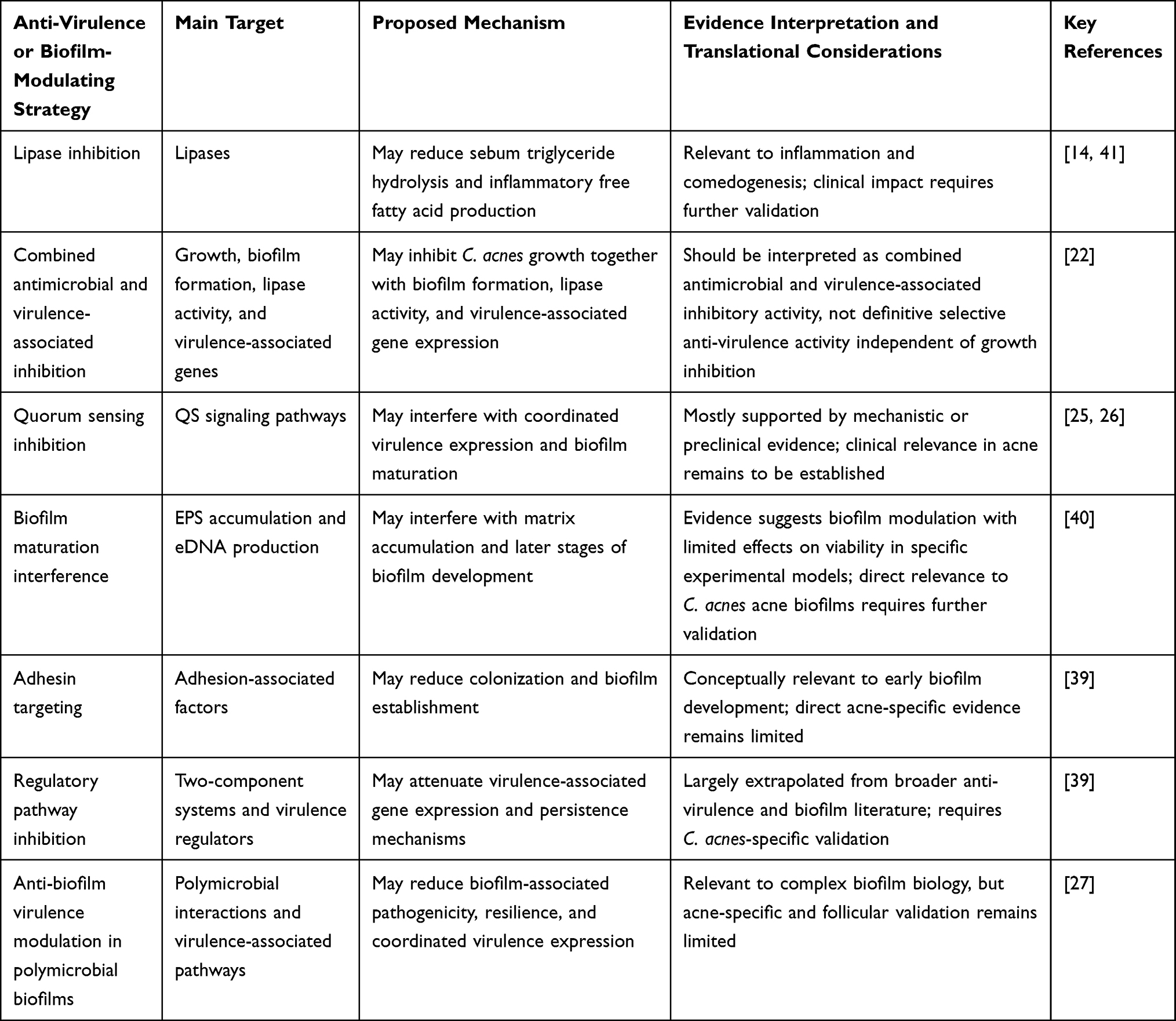 |
Table 2 Potential Anti-Virulence and Biofilm-Modulating Strategies Relevant to Cutibacterium acnes Biofilm-Associated Acne |
Quorum-Sensing Inhibition: Disrupting Bacterial Communication
Quorum sensing (QS) is a bacterial communication mechanism that enables bacteria to coordinate population density-dependent behaviors through the production, release, and detection of signaling molecules known as autoinducers. In many bacterial species, QS can regulate collective phenotypes, including virulence expression, biofilm development, and stress adaptation.26 In C. acnes, however, the specific contribution of QS-related signaling to biofilm regulation, virulence expression, and antimicrobial tolerance remains less clearly defined than in classical QS model organisms.21,25,26 Therefore, QS-related mechanisms in C. acnes should be interpreted cautiously and distinguished from mechanisms inferred from broader bacterial biofilm literature.25,26
At the biofilm level, QS-related signaling has been proposed to influence biofilm maturation, matrix-associated processes, and adaptive bacterial behavior in several experimental biofilm systems.25,26,28 However, direct evidence demonstrating that QS specifically controls EPS production, mature biofilm architecture, persister-like phenotypes, or antibiotic tolerance in human follicular C. acnes biofilms remains limited. Therefore, these mechanisms should be interpreted as hypothesis-generating or model-derived rather than established features of C. acnes biofilm regulation in acne.25,26,42
The currently available acne-relevant evidence is more closely related to autoinducer-2 (AI-2)-mediated signaling than to an accessory gene regulator (agr)-like peptide-based QS system.42,43 Accordingly, the previous statement referring to peptide-based signaling systems analogous to the agr system was removed. Several QS-modulating strategies, including natural compounds and signal-interfering agents, have been investigated as potential approaches to reduce C. acnes biofilm-associated behavior without relying solely on broad-spectrum bacterial eradication.42,43 For example, Cañellas-Santos et al reported that Camellia sinensis callus lysate reduced AI-2 production, biofilm formation, lipase activity, and keratinocyte inflammatory responses, including IL-6, IL-8, TNF-α, and CXCL1 expression, without significantly affecting bacterial viability.43 These findings support a potential AI-2-related biofilm-modulating and anti-inflammatory effect, although they should not be interpreted as definitive evidence that QS inhibition preserves the broader skin microbiome in vivo. Similarly, Hamada et al reported that heparinoid inhibited AI-2-mediated signaling associated with C. acnes, suppressed C. acnes biofilm formation without reducing viable bacterial counts, and enhanced the bactericidal efficacy of 4-isopropyl-3-methylphenol (IPMP) against C. acnes biofilms.42 These findings suggest that AI-2-related QS inhibition may represent a potential biofilm-modulating strategy for acne-related C. acnes control.42,43
Nevertheless, current evidence does not yet establish QS as a fully defined regulatory system controlling EPS production, biofilm architecture, persister-like phenotypes, follicular accumulation of signaling molecules, or antibiotic tolerance in human acne lesions.21,25,26,42 Several of these mechanisms are better regarded as hypothesis-generating or extrapolated from broader bacterial QS and biofilm models unless directly demonstrated in C. acnes.25,26 Therefore, statements regarding QS-mediated regulation of EPS production, structural biofilm organization, stress-adaptive subpopulations, and antimicrobial tolerance should be interpreted cautiously.21,25,26,42
Beyond direct QS interference, other anti-biofilm or virulence-associated modulators may affect C. acnes biofilm development through mechanisms that are not necessarily QS-specific.44 For example, Kim et al demonstrated that shikonin inhibited mono- and multispecies biofilms involving C. acnes, reduced EPS-associated biofilm features, and downregulated several biofilm- and virulence-related genes, including those associated with adhesion, lipase activity, and hyaluronate lyase production.44 However, because this effect was not presented as direct QS inhibition and shikonin also showed concentration-dependent antimicrobial activity, these findings should be interpreted as biofilm- and virulence-associated modulation rather than definitive QS-specific anti-virulence activity.44
Overall, QS-targeted and virulence-associated modulation strategies may offer a complementary approach to conventional antimicrobial therapy by interfering with coordinated biofilm-associated behavior rather than relying solely on bacterial eradication.25,26,42,43 However, most evidence remains experimental, and the effectiveness of QS-targeted approaches may be influenced by the complexity of signaling networks, strain-level variation, and redundancy in regulatory pathways.25,26,42,43 Further studies using clinically relevant follicular models and human acne lesions are required to clarify whether AI-2-related QS inhibition can meaningfully reduce C. acnes biofilm persistence, improve antimicrobial susceptibility, and preserve microbiome balance in vivo.42,43
Biofilm Disruption Strategies: Targeting the Matrix
Although QS-related signaling may contribute to biofilm-associated behavior, the structural integrity of mature biofilms is largely supported by the EPS matrix, making the matrix a potential therapeutic target. In experimental biofilm systems, this matrix may contain polysaccharides, proteins, extracellular DNA (eDNA), and lipids, and can facilitate bacterial cohesion, restrict antimicrobial penetration, and create protective microenvironments.21,28 However, direct evidence linking specific EPS-mediated mechanisms to human follicular C. acnes biofilms remains limited, and extrapolation from broader biofilm models should be interpreted cautiously. Biofilm-associated bacteria may exhibit reduced antimicrobial susceptibility due to restricted drug diffusion, altered metabolic activity, putative tolerant or persister-like subpopulations, and adaptive stress responses.21,27,28 Spatial gradients within biofilms may further create metabolically heterogeneous microenvironments that contribute to reduced antimicrobial efficacy.27,28
Targeting the biofilm matrix has therefore emerged as a potential strategy to enhance antimicrobial penetration and destabilize biofilm-associated communities. Enzymatic approaches, including DNases, proteases, and polysaccharide-degrading enzymes, may disrupt biofilm structure and improve antimicrobial access to embedded bacterial cells.29,38 For instance, dispersin B-mediated degradation of PNAG has been shown to destabilize biofilm matrices and enhance the activity of conventional agents such as benzoyl peroxide against C. acnes/Staphylococcus epidermidis dual-species biofilms.29 Similarly, microbiota-derived small molecules such as short-chain fatty acids (SCFAs) have been reported to interfere with biofilm maturation in specific experimental models, including reduced polysaccharide and eDNA accumulation in S. epidermidis biofilms.40 However, the direct relevance of these matrix-disrupting or biofilm-modulating mechanisms to follicular C. acnes biofilms in acne remains to be further validated.
Importantly, combining biofilm-disrupting agents with antimicrobial therapies may improve antibiofilm activity compared with antimicrobial treatment alone.28,29 However, the magnitude of benefit may vary depending on the biofilm model, microbial composition, treatment timing, and formulation strategy. Challenges such as enzyme instability, delivery limitations, and potential off-target effects remain, highlighting the need for optimized formulations and further translational validation.
Advanced Drug Delivery Systems: Enhancing Penetration and Targeting
In addition to biofilm disruption strategies, advanced drug delivery systems offer innovative solutions to overcome the limitations of conventional therapies. The biofilm matrix acts as both a physical and biochemical barrier that restricts antimicrobial diffusion and alters drug distribution within infected sites.45 As a result, biofilm-associated bacteria can exhibit resistance levels up to 10–1,000-fold higher than planktonic cells, underscoring the need for delivery systems capable of penetrating this barrier.30
Nanoparticle-based carriers have emerged as promising strategies to enhance drug delivery, follicular targeting, and sustained release, with the potential to improve therapeutic outcomes in acne management. For example, Arooj et al developed adapalene-loaded liposomes with a particle size of approximately 172 nm, zeta potential of −35 mV, and high encapsulation efficiency of 89.7%. When incorporated into Carbopol gel, this system showed sustained drug release, improved dermal deposition, and significant improvement of acne lesions in a testosterone-induced acne model without visible irritation or inflammation.46 Ansari et al developed karanjin-loaded ethosomal gel with a particle size of approximately 141 nm and entrapment efficiency of 71.4%. This formulation increased transdermal flux by 1.9-fold and skin deposition by 2.4-fold compared with conventional karanjin solution, while also showing antibacterial activity against C. acnes, antioxidant activity, anti-inflammatory effects, and reduced sebaceous gland enlargement.47 Ogunjimi et al further demonstrated that isotretinoin-loaded Delonix polymeric nanoparticles, with a particle size of approximately 230 nm, preferentially accumulated in hair follicles and increased follicular isotretinoin deposition by approximately three-fold compared with isotretinoin solution. These nanoparticles also provided sustained release, reduced phototoxicity, and suppressed IL-6 expression.48 Collectively, these examples indicate that nanocarrier-based systems can enhance acne therapy by improving follicular localization, prolonging skin retention, controlling release, reducing irritation, and exerting anti-inflammatory activity, rather than merely increasing drug loading. Beyond improving drug delivery, these examples illustrate that nanocarrier platforms can provide multifunctional therapeutic benefits. In addition to enhancing follicular targeting and skin retention, these systems may simultaneously exert antibacterial, anti-inflammatory, antioxidant, and photoprotective effects. Such multifunctionality is particularly attractive for acne management, where therapeutic success often depends on simultaneously addressing multiple pathogenic processes. Nevertheless, most of the currently available evidence is derived from preclinical studies, and well-designed clinical trials remain limited, underscoring the need for further validation before widespread clinical implementation.
Stimuli-responsive nanocarriers further enhance therapeutic precision by enabling site-specific drug release in response to microenvironmental cues such as pH changes, enzymatic activity, and inflammatory signals.49 In acne lesions, where pH and inflammatory conditions are altered, these systems can improve local drug bioavailability while reducing off-target effects. Notably, deformable systems such as transfersomes have demonstrated enhanced skin penetration in experimental studies, further supporting their potential utility in topical therapy.50
Beyond their role as delivery vehicles, certain nanomaterials exhibit intrinsic antibacterial activity through mechanisms such as reactive oxygen species (ROS) generation, membrane disruption, and interference with intracellular processes. These multimodal actions can bypass conventional resistance mechanisms and enhance the efficacy of co-administered antibiotics through synergistic effects.51
Nanoparticles can also function as dynamic sensor–actuator systems, capable of detecting local pathological signals and triggering targeted drug release within the biofilm microenvironment.52 Their effectiveness is influenced by physicochemical properties such as particle size, surface charge, and hydrophobicity, which determine interactions with the EPS matrix and biofilm penetration dynamics.45 In addition, surface functionalization strategies enable targeted delivery to specific bacterial populations or biofilm components, further enhancing therapeutic precision.30
Experimental evidence supports these advantages, with systems such as calcium fluoride nanoparticles that suppress virulence gene expression and gold nanoparticles functionalized with hordenine that demonstrate enhanced antibiofilm activity. Nitric oxide-releasing nanoparticles have also shown superior antimicrobial efficacy compared to conventional antibiotics in in vivo models.45 Furthermore, nanoparticles can facilitate the delivery of matrix-degrading enzymes, such as DNase and proteases, thereby enabling simultaneous biofilm disruption and antimicrobial action.30 However, despite their significant potential, nanocarrier-based systems face important challenges, including formulation stability, high production costs, and limited long-term safety data, which may constrain their clinical translation.
Toward Precision Dermatology: Integrating Microbiome and Strain-Specific Targeting
Emerging evidence suggests that acne is not solely driven by bacterial abundance but by the dominance of specific virulent C. acnes phylotypes. This is clinically relevant, considering that acne vulgaris affects approximately 85% of adolescents and a significant proportion of adults, emphasizing the need for more targeted and effective therapeutic strategies.53 The human skin harbors approximately 1000 bacterial species, forming a highly complex and dynamic microbial ecosystem that plays a fundamental role in maintaining skin homeostasis.53 Actinobacteria constitute approximately 51.8% of the skin microbiota, highlighting the dominance of specific microbial groups within distinct skin niches.54
C. acnes exhibits substantial genetic heterogeneity, with over 100 strains classified into eight phylotypes, among which IA1 and IA2 are strongly associated with severe acne phenotypes, underscoring the need for strain-specific therapeutic strategies.31 This strain-specific pathogenicity is partly driven by metabolic adaptations, particularly lipase-mediated hydrolysis of sebum triglycerides, which generates free fatty acids, thereby promoting bacterial adhesion and follicular colonization.54
This concept is further supported by experimental host–microbe co-culture models, which demonstrate that different skin-associated bacteria exert distinct effects on epidermal structure and inflammatory responses. For instance, Staphylococcus aureus induces pronounced pro-inflammatory signaling, including elevated secretion of IL-8 and CXCL1, whereas commensal species such as C. acnes and S. epidermidis exhibit more limited or context-dependent effects. These findings highlight that microbial impact on skin health is highly species- and strain-dependent, reinforcing the importance of targeted and microbiome-preserving therapeutic strategies.55
Importantly, resistance in polymicrobial biofilms is not merely additive but rather a synergistic, emergent property driven by complex interspecies interactions. Propionibacterium-derived metabolites have been shown to induce aggregation and biofilm formation in S. aureus, further illustrating how interspecies interactions can enhance pathogenicity in polymicrobial environments.54 Moreover, microbe–microbe and host–microbe interactions collectively regulate disease outcomes through multiple coordinated mechanisms. Commensal microorganisms contribute to microbial homeostasis via competitive exclusion, antimicrobial peptide (AMP) production, and immune modulation, thereby limiting pathogen colonization and maintaining ecological balance.54,56 In parallel, keratinocytes play a central role in microbiome regulation by expressing pattern recognition receptors (PRRs), including Toll-like receptors (TLRs) and NOD-like receptors (NLRs), and by producing AMPs that shape microbial composition and inflammatory responses.54 Microbial metabolites such as SCFAs further support skin barrier integrity by regulating keratinocyte differentiation, lipid synthesis, and pH balance. Disruption of this tightly regulated system leads to microbial dysbiosis, which can drive chronic low-grade inflammation, barrier dysfunction, and increased susceptibility to inflammatory skin conditions.22
Supporting this concept, experimental studies have demonstrated that certain bioactive metabolites exhibit selective antibiofilm activity that varies by bacterial species, with pronounced effects against specific commensal or opportunistic strains while sparing others. This selective inhibition profile suggests that targeted modulation of microbial communities may be achievable without broad-spectrum disruption. However, the long-term effects of such approaches on the skin microbiome and their clinical relevance require further investigation.40
This growing body of evidence highlights a critical limitation of conventional broad-spectrum antibiotics, which fail to discriminate between pathogenic and commensal strains, often leading to microbial imbalance and dysbiosis.14 Consequently, increasing attention has been directed toward microbiome-based and strain-selective therapeutic approaches that aim to preserve beneficial microbial communities while selectively targeting pathogenic strains.14,37 Recent advances highlight multiple microbiome-targeted strategies, including the application of specific commensal bacteria, bacteriophage therapy, prebiotic and probiotic metabolites, and microbiome transplantation, all of which aim to selectively modulate microbial communities rather than indiscriminately eliminate them.13
Such strategies may include targeted antimicrobial peptides, bacteriophage therapy, or microbiome-modulation approaches to restore microbial balance. In particular, nanocarrier-based systems have shown potential to enhance follicular drug delivery, thereby improving local drug accumulation at sites of C. acnes colonization while reducing off-target exposure.53 Clinical studies have demonstrated that topical administration of probiotic strains (~108 CFU) significantly reduces inflammatory acne lesions within 4 weeks, with sustained therapeutic effects observed even after treatment cessation, suggesting potential involvement of adaptive immune modulation.54 Although these findings are encouraging, available clinical studies remain limited in size and duration, and further validation is required before routine clinical implementation can be recommended.
By integrating insights from microbiome research, biofilm biology, and host–microbe interactions, these approaches represent a shift toward precision dermatology, where treatment is tailored to the specific microbial and molecular characteristics of the disease. This emerging precision-oriented framework challenges the long-standing reliance on broad-spectrum antimicrobial eradication, which appears increasingly inadequate in the context of biofilm-mediated persistence and strain-specific virulence. Instead, precision dermatology emphasizes selective interventions to restore microbial balance and attenuate pathogenic activity while minimizing disruption of beneficial microbial communities. However, the extent to which these objectives can be consistently achieved in clinical settings remains an active area of investigation.14,25
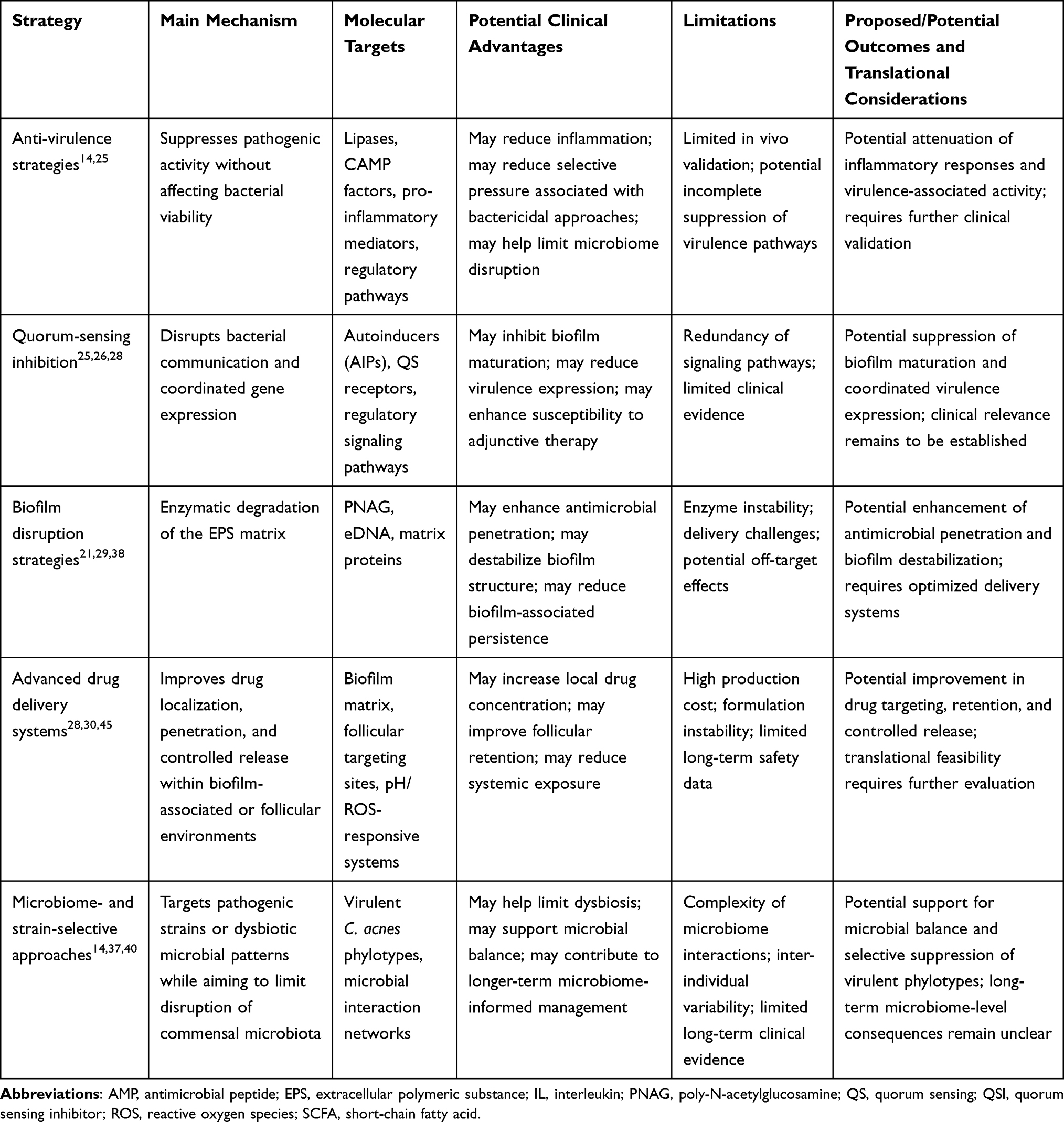 |
Table 3 Emerging Selective Modulation Strategies Targeting Cutibacterium acnes Biofilms |
Taken together, these strategies underscore the need for integrated therapeutic approaches that simultaneously target biofilm architecture, virulence regulation, and microbial ecology. By shifting from broad-spectrum antimicrobial eradication toward selective, mechanism-based interventions, these approaches may offer a more targeted framework for acne management. The key mechanisms and therapeutic targets of selective modulation strategies are summarized in Figure 1 and Table 3. Despite their mechanistic diversity, these approaches converge on a shared therapeutic goal of reducing biofilm-associated resilience while minimizing unnecessary broad-spectrum disruption of microbial communities.21,25,28,29 Anti-virulence strategies aim to attenuate pathogenic activity, although the extent to which individual agents act independently of bacterial growth inhibition varies and requires careful interpretation.22,26,39,40,44 QS inhibitors may interfere with AI-2-related signaling and biofilm-associated behaviors, whereas biofilm-disruption strategies directly target the extracellular matrix to enhance antimicrobial penetration.29,42,43,45 Advanced drug delivery systems may improve drug localization and controlled release within biofilm-associated or follicular environments.30,45–48 In parallel, microbiome- and strain-selective approaches aim to modulate acne-associated microbial ecology, although their long-term microbiome-level consequences remain incompletely understood.13,14,37,53 Despite growing interest in microbiome-informed therapies, much of the current evidence remains preclinical or limited to small clinical studies. Furthermore, the long-term consequences of selectively manipulating skin microbial communities require further investigation.
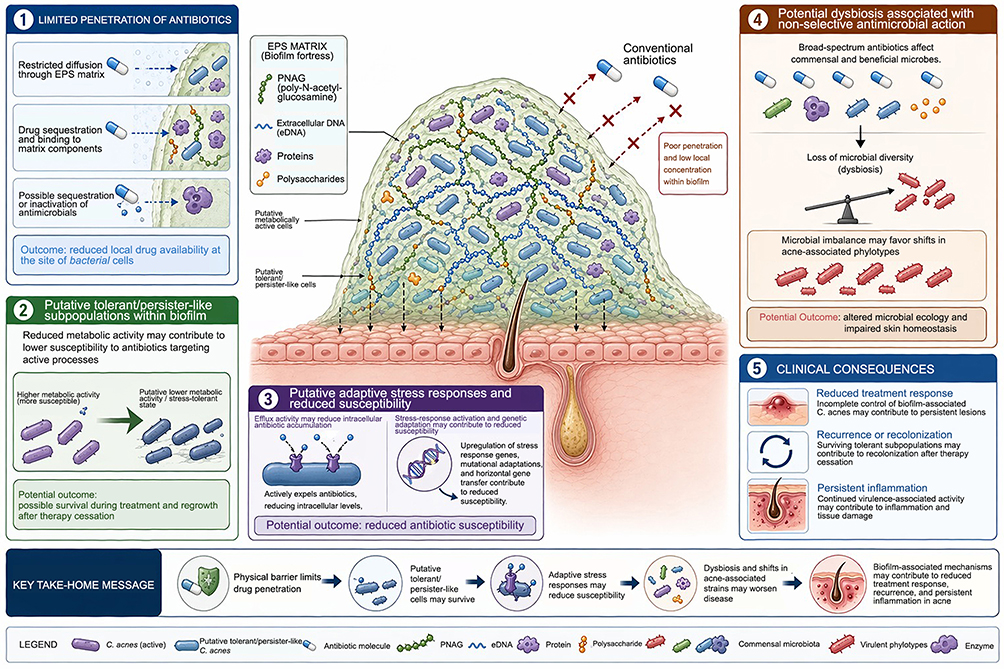 |
Figure 1 Conceptual model of mechanisms underlying reduced antimicrobial susceptibility in Cutibacterium acnes biofilm-associated acne. This figure presents a conceptual representation of biofilm-associated mechanisms that may contribute to reduced treatment response in acne, based on experimental C. acnes biofilm studies and broader biofilm biology. The model illustrates how the EPS matrix may restrict antimicrobial diffusion, reduce local drug availability, and support the presence of putative tolerant or persister-like subpopulations. Additional contributing mechanisms may include adaptive stress responses, altered metabolic states, and reduced susceptibility to antimicrobial therapy. The figure also highlights the potential consequences of non-selective antimicrobial treatment, including dysbiosis and shifts in acne-associated phylotypes. Collectively, these mechanisms may contribute to persistent inflammation, recurrence, and reduced therapeutic efficacy. This illustration is intended as a conceptual model and should not be interpreted as a definitive structural representation of C. acnes biofilms within the human pilosebaceous follicle. |
Future Clinical Integration
Despite promising in vitro findings, the clinical translation of selective modulation strategies remains limited by challenges related to formulation stability, delivery efficiency, and regulatory approval.27,28 A major constraint is the lack of robust in vivo validation and standardized comparative studies, as most current evidence is derived from small-scale experimental models that do not fully reflect human skin physiology.23,50
Conventional therapies, including topical antibiotics and retinoids, are further limited by adverse effects such as irritation, photosensitivity, and antimicrobial resistance.49 Clinical outcomes remain suboptimal, with up to 36.2% of patients developing acne scarring and post-inflammatory hyperpigmentation, while only 18.7% remain free of long-term complications.41 These limitations are compounded by intrinsic delivery barriers, including poor stratum corneum penetration, follicular obstruction, and reduced drug deposition at pilosebaceous sites.57
In addition, the structural complexity of polymicrobial biofilms—comprising C. acnes, Staphylococcus spp., and Malassezia—creates a dense EPS matrix that restricts drug diffusion and establishes microenvironmental gradients, contributing to discrepancies between in vitro and in vivo efficacy.21,45,58 Biofilm-associated microorganisms can exhibit antibiotic tolerance levels up to 10–1000 times higher than planktonic counterparts due to restricted diffusion, metabolic dormancy, and adaptive stress responses.58 Although emerging evidence supports the translational potential of biofilm-targeted therapies, including topical formulations with efficacy comparable to benzoyl peroxide and reduced adverse effects, their broader clinical application remains limited.59 Additional challenges include instability of bioactive compounds, variability in plant-derived therapeutics, and lack of standardization, which hinder reproducibility and scalability.23,30
Regulatory and economic barriers further complicate clinical integration. Nanomaterial-based therapies require rigorous safety evaluation due to concerns regarding cytotoxicity, systemic toxicity, and environmental impact.49 The regulatory landscape remains fragmented, with insufficient harmonization and limited long-term safety data.60 These challenges are consistent with broader observations in nanomedicine, where the absence of harmonized global regulatory frameworks, variability in manufacturing processes, and complexity in physicochemical characterization continue to hinder clinical translation and commercialization of nanoparticle-based therapeutics.61 In addition, manufacturing challenges such as batch variability and scalability constraints further limit clinical implementation.50,57 Nanomaterials may also induce oxidative stress through ROS, potentially disrupting skin homeostasis.23,53,60
Another critical challenge lies in inter-individual variability of the skin microbiome, which contributes to heterogeneous treatment responses and limits the effectiveness of uniform therapeutic approaches.14 Microbiome-based interventions, therefore, require standardized protocols and careful regulatory consideration to ensure safety and efficacy.13
Despite growing interest in selective modulation strategies, several important limitations remain within the current evidence base. Much of the available literature is derived from in vitro biofilm models, ex vivo systems, or preclinical investigations, whereas direct evidence from clinically relevant human follicular models and large-scale randomized clinical trials remains limited.23,50,57 In addition, substantial heterogeneity exists in experimental designs, biofilm models, outcome measures, and formulation platforms, making cross-study comparisons challenging.23,57 Consequently, the long-term efficacy, safety, and translational relevance of strategies targeting microbial communities, QS pathways, and biofilm-associated mechanisms remain incompletely understood and require further validation in clinically relevant settings.50,57
Future clinical integration will depend on the development of physiologically relevant in vivo models, large-scale clinical trials, and microbiome-informed therapeutic strategies that enable personalized intervention.13,14,23,50,57 Advances in multi-omics technologies and real-time microbiome profiling may facilitate the identification of patient-specific microbial signatures and guide targeted therapy.25 In parallel, improvements in formulation strategies, including stimuli-responsive and biofilm-penetrating systems, will be essential to enhance therapeutic efficacy.30,52
Ultimately, bridging the gap between experimental findings and clinical application will require an integrated, multidisciplinary approach combining microbiology, materials science, dermatology, and regulatory science to advance precision dermatology. This highlights that the clinical translation of advanced delivery systems is not solely a scientific challenge, but also a regulatory and manufacturing bottleneck.61
Conclusion
Selective modulation strategies targeting biofilm integrity, virulence pathways, and microbial ecology represent a promising emerging direction in acne research, particularly because conventional approaches primarily focus on bacterial eradication and may insufficiently address biofilm-associated tolerance, strain-level virulence, and microbial dysbiosis. By integrating anti-virulence therapy, QS inhibition, biofilm disruption, advanced drug delivery systems, and microbiome-informed interventions, these strategies may provide a more targeted framework for controlling C. acnes–associated pathology. However, the current level of evidence remains uneven, and many of these approaches are still supported mainly by in vitro, ex vivo, or preclinical findings rather than robust clinical validation.
Significant challenges remain in translating selective modulation strategies into clinical practice, including formulation stability, delivery efficiency, regulatory standardization, long-term safety, and manufacturing scalability. In addition, the structural complexity of follicular biofilms and inter-individual variability in the skin microbiome may influence therapeutic responses and complicate clinical outcomes. Important unresolved questions remain, including whether selective modulation can improve acne outcomes in controlled clinical trials, whether microbiome-informed interventions can truly preserve or restore microbial balance, how targeting a dominant commensal species such as C. acnes affects broader skin microbiome ecology, and whether follicular C. acnes biofilms in human acne lesions share the same structural and physiological properties observed in experimental biofilm models.
Future research should prioritize physiologically relevant in vivo models, standardized outcome measures, large-scale clinical validation, and personalized therapeutic frameworks that integrate microbiome profiling with advanced delivery technologies. Ultimately, the successful clinical integration of these strategies will require a multidisciplinary approach bridging microbiology, materials science, dermatology, and regulatory science. Rather than replacing established acne therapies at present, selective modulation may serve as a complementary and more targeted strategy for future acne management, provided that its efficacy, safety, and microbiome-level consequences are validated in clinically relevant settings.
Declaration of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this work, the author used AI-assisted tools to support language editing. The author developed the content, reviewed, and revised the manuscript, and takes full responsibility for the final version.
Acknowledgments
This study is funded by Universitas Padjadjaran through the Indonesian Endowment Fund for Education (LPDP) on behalf of the Indonesian Ministry of Higher Education, Science and Technology and managed under the EQUITY Program (Contract No. 4303/83/DT.03.08/2025 and 3927/UN6.RKT/HK.07.00/2025).
Disclosure
The author reports funding support for this work from Universitas Padjadjaran through the Indonesian Endowment Fund for Education (LPDP), as stated in the Acknowledgments section. The author reports no conflicts of interest in this work.
References
1. Wolkenstein P, Machovcová A, Szepietowski JC. et al. Acne prevalence and associations with lifestyle: a cross-sectional online survey of adolescents/young adults in 7 European countries. J Eur Acad Dermatol Venereol. 2018;32(2):298–17. doi:10.1111/jdv.14475
2. Maddiboyina B, Jhawat V, Ramesh K, et al. A Review on nanoparticle based herbal remedies for the management of alopecia. Der Pharmacia Lettre. 2020;12(5):1–17.
3. Hazarika N. Acne vulgaris: new evidence in pathogenesis and future modalities of treatment. J Dermatol Treat. 2021;32(3):277–285. doi:10.1080/09546634.2019.1654075
4. Heath C. Acne vulgaris. Cutis. 2021;108(3):167. doi:10.12788/cutis.0339
5. Escamilla R, Verónica D, Pineda A, et al. From Comedones to Scars: understanding acne pathogenesis, clinical types, and contemporary management. SAS J Med. 2025;11(07):702–705. doi:10.36347/sasjm.2025.v11i07.011
6. Tan AU, Schlosser BJ, Paller AS. A review of diagnosis and treatment of acne in adult female patients. Int J Womens Dermatol. 2018;4(2):56–71. doi:10.1016/j.ijwd.2017.10.006
7. Bungau AF, Marin RC, Tit DM, et al. Multifactorial refractory acne in women: insights from a case series involving hormonal-, metabolic-, and corticosteroid-related triggers. Life. 2025;15(8):1196. doi:10.3390/life15081196
8. Vasam M, Korutla S, Bohara RA. Acne vulgaris: a review of the pathophysiology, treatment, and recent nanotechnology based advances. Biochem Biophys Rep. 2023;36:101578. doi:10.1016/j.bbrep.2023.101578
9. Jisa TA, MdTI R, MostSA S, et al. Prevalence of acne and its impact on quality of life, social appearance anxiety and treatment practices among young adults. J Cosmet Dermatol. 2026;25(1):e70654. doi:10.1111/jocd.70654
10. Meixiong J, Ricco C, Vasavda C, et al. Diet and acne: a systematic review. JAAD International. 2022;7:95–112. doi:10.1016/j.jdin.2022.02.012
11. Li Z, Zhang J, Zhang Y, et al. Skin microbiome in health and disease: mechanisms and emerging therapeutic strategies. Clin Cosmet Investig Dermatol. 2025;18:3443–3455. doi:10.2147/CCID.S571984
12. Cong TX, Hao D, Wen X, et al. From pathogenesis of acne vulgaris to anti-acne agents. Arch Dermatol Res. 2019;311(5):337–349. doi:10.1007/s00403-019-01908-x
13. Suri H, Suri H, Nagda N, et al. Current perspectives on the human skin microbiome: functional insights and strategies for therapeutic modulation. Biomed Pharmacother. 2025;193:118655. doi:10.1016/j.biopha.2025.118655
14. Dreno B, Dekio I, Baldwin H, et al. Acne microbiome: from phyla to phylotypes. J Eur Acad Dermatol Venereol. 2024;38(4):657–664. doi:10.1111/jdv.19540
15. Drake L, Reyes-Hadsall S, Barbieri JS, et al. New developments in topical acne therapy. Am J Clin Dermatol. 2022;23(2):125–136. doi:10.1007/s40257-021-00666-9
16. Li Y, Hu X, Dong G, et al. Acne treatment: research progress and new perspectives. Front Med. 2024;11:1425675. doi:10.3389/fmed.2024.1425675
17. Sheffer-Levi S, Rimon A, Lerer V, et al. Antibiotic susceptibility of Cutibacterium acnes strains isolated from Israeli acne patients. Acta Derm Venereol. 2020;100(17):1–8. doi:10.2340/00015555-3654
18. Dessinioti C, Katsambas A. Antibiotics and antimicrobial resistance in acne: epidemiological trends and clinical practice considerations. Yale J Biol Med. 2022;95(4):429–443. PubMed PMID: 36568833.
19. Beig M, Shirazi O, Ebrahimi E, et al. Prevalence of antibiotic-resistant Cutibacterium acnes (formerly Propionibacterium acnes) isolates, a systematic review and meta-analysis. J Glob Antimicrob Resist. 2024;39:82–91. doi:10.1016/j.jgar.2024.07.005
20. Paul NA, Bhat MR, Antony B, et al. Antibiotic resistance and biofilm formation in Cutibacterium acnes: a descriptive cross-sectional study. Indian J Dermatol Venereol Leprol. 2025;91:315–320. doi:10.25259/IJDVL_539_2024
21. Ruchiatan K, Rizqandaru T, Satjamanggala PR, et al. Characteristics of biofilm-forming ability and antibiotic resistance of Cutibacterium acnes and Staphylococcus epidermidis from acne vulgaris patients. Clin Cosmet Investig Dermatol. 2023;16:2457–2465. doi:10.2147/CCID.S422486
22. Hong S, Park S, Park M, et al. Anti-acne potential of Quercus salicina extract: inhibition of Cutibacterium acnes growth and virulence. J Microbiol Biotechnol. 2026;36:e2510052. doi:10.4014/jmb.2510.10052
23. Karar W, Mohamed SA, Subramaniam G, et al. Battling biofilms: evaluating selected agents against Cutibacterium acnes —a review. PeerJ. 2026;14:e20652. doi:10.7717/peerj.20652
24. Ruffier d’Epenoux L, Fayoux E, Veziers J, et al. Biofilm of Cutibacterium acnes: a target of different active substances. Int J Dermatol. 2024;63(11):1541–1550. doi:10.1111/ijd.17194
25. Coenye T, Spittaels KJ, Achermann Y. The role of biofilm formation in the pathogenesis and antimicrobial susceptibility of Cutibacterium acnes. Biofilm. 2022;4:100063. doi:10.1016/j.bioflm.2021.100063
26. Fleitas Martínez O, Cardoso MH, Ribeiro SM, et al. Recent advances in anti-virulence therapeutic strategies with a focus on dismantling bacterial membrane microdomains, toxin neutralization, quorum-sensing interference and biofilm inhibition. Front Cell Infect Microbiol. 2019;9:74. doi:10.3389/fcimb.2019.00074
27. Nithyanand P, Boya BR, Lee J, et al. Polymicrobial biofilms: interkingdom interactions, resistance and therapeutic strategies. Microb Biotechnol. 2025;18(8):e70218. doi:10.1111/1751-7915.70218
28. Yang S, Li X, Cang W, et al. Biofilm tolerance, resistance and infections increasing threat of public health. Microb Cell. 2023;10(11):233–247. doi:10.15698/mic2023.11.807
29. Kaplan JB, Muzaleva A, Sailer M, et al. Synergistic activity of dispersin B and benzoyl peroxide against Cutibacterium acnes/Staphylococcus epidermidis dual-species biofilms. Abdeltawab NF editor. PLoS One. 2025;20(3):e0320662. doi:10.1371/journal.pone.0320662
30. Yeh YC, Huang TH, Yang SC, et al. Nano-based drug delivery or targeting to eradicate bacteria for infection mitigation: a review of recent advances. Front Chem. 2020;8:286. doi:10.3389/fchem.2020.00286
31. Burkhart CG. Assessment of Cutibacterium acnes: acne biofilm, comedones, and future treatments for acne. Open Dermatol J. 2024;18(1):e18743722279314. doi:10.2174/0118743722279314240219091938
32. Gannesen AV, Zdorovenko EL, Botchkova EA, et al. Composition of the biofilm matrix of Cutibacterium acnes Acneic strain RT5. Front Microbiol. 2019;10:1284. doi:10.3389/fmicb.2019.01284
33. Gowda A, Burkhart CG. Virulent acne biofilms offer insight into novel therapeutic options. Open Dermatol J. 2018;12(1):80–85. doi:10.2174/1874372201812010080
34. Mayslich C, Grange PA, Dupin N. Cutibacterium acnes as an opportunistic pathogen: an update of its virulence-associated factors. Microorganisms. 2021;9(2):303. doi:10.3390/microorganisms9020303
35. Di Francesco AM, Pasciuto G, Verrecchia E, et al. The role of Cutibacterium acnes in the etiopathogenesis of sarcoidosis: current insights and future study directions. Int J Mol Sci. 2025;26(14):6652. doi:10.3390/ijms26146652
36. Bjerg CSB, Poehlein A, Bömeke M, et al. Increased biofilm formation in dual-strain compared to single-strain communities of Cutibacterium acnes. Sci Rep. 2024;14(1):14547. doi:10.1038/s41598-024-65348-y
37. Kasmal N, Kartika AV, Roem NR. Acne infection (acne vulgaris) and the relationship between Cutibacterium acnes colonization and severity. J Community Health Provis. 2026;6(1):68–77. doi:10.55885/jchp.v6i1.928
38. Kuehnast T, Cakar F, Weinhäupl T, et al. Comparative analyses of biofilm formation among different Cutibacterium acnes isolates. Int J Med Microbiol. 2018;308(8):1027–1035. doi:10.1016/j.ijmm.2018.09.005
39. Dehbanipour R, Ghalavand Z. Anti-virulence therapeutic strategies against bacterial infections: recent advances. Germs. 2022;12(2):262–275. doi:10.18683/germs.2022.1328
40. Nakamura K, O’Neill AM, Williams MR, et al. Short chain fatty acids produced by Cutibacterium acnes inhibit biofilm formation by Staphylococcus epidermidis. Sci Rep. 2020;10(1):21237. doi:10.1038/s41598-020-77790-9
41. Alamri A, Alzahrani D, Alharbi G, et al. The predictive factors of acne scarring and post-inflammatory hyperpigmentation: a retrospective cohort study. Clin Cosmet Investig Dermatol. 2025;18:143–150. doi:10.2147/CCID.S504281
42. Hamada S, Minami S, Gomi M. Heparinoid enhances the efficacy of a bactericidal agent by preventing Cutibacterium acnes biofilm formation via quorum sensing inhibition. J Microorg Control. 2024;29(1):27–31. doi:10.4265/jmc.29.1_27
43. Cañellas-Santos M, Rosell-Vives E, Montell L, et al. Anti-Inflammatory and anti-quorum sensing effect of Camellia sinensis callus lysate for treatment of acne. Curr Issues Mol Biol. 2023;45(5):3997–4016. doi:10.3390/cimb45050255
44. Kim YG, Lee JH, Kim S, et al. Inhibition of biofilm formation in Cutibacterium acnes, Staphylococcus aureus, and Candida albicans by the phytopigment shikonin. Int J Mol Sci. 2024;25(4):2426. doi:10.3390/ijms25042426
45. Shrestha L, Fan HM, Tao HR, et al. Recent strategies to combat biofilms using antimicrobial agents and therapeutic approaches. Pathogens. 2022;11(3):292. doi:10.3390/pathogens11030292
46. Arooj A, Rehman AU, Iqbal M, et al. Development of Adapalene loaded liposome based gel for acne. Gels. 2023;9(2):135. doi:10.3390/gels9020135
47. Ansari SA, Qadir A, Warsi MH, et al. Ethosomes-based gel formulation of karanjin for treatment of acne vulgaris: in vitro investigations and preclinical assessment. 3 Biotech. 2021;11(11):456. doi:10.1007/s13205-021-02978-3
48. Ogunjimi AT, Chahud F, Lopez RFV. Isotretinoin-Delonix polymeric nanoparticles: potentials for skin follicular targeting in acne treatment. Int J Pharm. 2021;610:121217. doi:10.1016/j.ijpharm.2021.121217
49. Chen R, Tang Y, Duan L, et al. Global trends and hotspot analysis of nanomaterials in the field of acne: a bibliometric perspective. Clin Cosmet Investig Dermatol. 2026;19:586477. doi:10.2147/CCID.S586477
50. Badshah SM, Mdlovu NV, Wu CM, et al. Transdermal nanomedicine: emerging horizons, unresolved challenges, and the path forward. Curr Opin Biomed Eng. 2026;37:100648. doi:10.1016/j.cobme.2025.100648
51. Parvin N, Joo SW, Mandal TK. Nanomaterial-based strategies to combat antibiotic resistance: mechanisms and applications. Antibiotics. 2025;14(2):207. doi:10.3390/antibiotics14020207
52. Shah RM, Kaur IP, Jadhav SR. The surface-first paradigm: engineering nanocarriers for infection-responsive drug delivery. Adv Colloid Interface Sci. 2026;349:103761. doi:10.1016/j.cis.2025.103761
53. Lalit DM, Pooja RH, Ravindra LB, et al. Nanotechnology-based approaches for acne treatment: a comprehensive review. GSC Biol Pharm Sci. 2025;31(1):156–162. doi:10.30574/gscbps.2025.31.1.0150
54. Madaan T, Doan K, Hartman A, et al. Advances in microbiome-based therapeutics for dermatological disorders: current insights and future directions. Exp Dermatol. 2024;33(12):e70019. doi:10.1111/exd.70019
55. Mieremet A, Rietveld M, Van Leijden B, et al. Advancing human skin models by integrating skin microbes for next-generation research. Sci Rep. 2026;16(1):13182. doi:10.1038/s41598-026-44005-6
56. Hong JY, Kwon D, Park KY. Microbiome-based interventions for skin aging and barrier function: a comprehensive review. Ann Dermatol. 2025;37(5):259. doi:10.5021/ad.25.009
57. Mustaffa MF, Mohd Ghazali NSM, Khalid SH, et al. Nanoformulations in acne therapy: translational opportunities and barriers for next-generation topical treatments. AAPS PharmSciTech. 2026;27(3):143. doi:10.1208/s12249-026-03345-2
58. Hosthota A, Khunger N, Tampi AS, et al. The impact of biofilms in dermatology: the fortified enemies. Our Dermatol Online. 2025;16(2). doi:10.7241/ourd.20252.17
59. Marshall-Hudson A, Tuley M, Damstra M, et al. A 6-month, multi-center, double-blind, controlled study to evaluate the effect of a biofilm disrupting acne cream on mild-to-moderate facial acne in female volunteer subjects. J Clin Aesthetic Dermatol. 2023;16(4):43–52. PubMed PMID: 37077927.
60. Yan Z, Zhang S, Wu G, et al. Advances in Nanotechnology-based topical delivery systems for skincare applications. Pharmaceutics. 2026;18(1):63. doi:10.3390/pharmaceutics18010063
61. Desai N, Rana D, Patel M, et al. Nanoparticle therapeutics in clinical perspective: classification, marketed products, and regulatory landscape. Small. 2025;21(29):e2502315. doi:10.1002/smll.202502315. PubMed PMID: 40454890.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Characteristics of Biofilm-Forming Ability and Antibiotic Resistance of Cutibacterium acnes and Staphylococcus epidermidis from Acne Vulgaris Patients
Ruchiatan K, Rizqandaru T, Satjamanggala PR, Tache N, Cahyadi AI, Rezano A, Gunawan H, Sutedja EK, Dwiyana RF, Hidayah RMN, Achdiat PA, Sutedja E, Suwarsa O, Hindritiani R
Clinical, Cosmetic and Investigational Dermatology 2023, 16:2457-2465
Published Date: 11 September 2023
AI-2 Signaling: A Potential Driver of Bacteremia in Non-Typhoidal Salmonella Infections
Li Y, Lu B, Qiang X, Lin Y, He J, Cai Y
Infection and Drug Resistance 2025, 18:1521-1537
Published Date: 19 March 2025
Scopoletin and Scoparone from Viola philippica Attenuate the Virulence of Pseudomonas aeruginosa by Inhibiting Quorum Sensing-Related Genes
Luo Y, Zhang S, Ou S, Shen C, Xiao Q, Chen W, Pu J, Zeng J, Zheng D, Huang B, Chen C
Infection and Drug Resistance 2025, 18:3891-3906
Published Date: 5 August 2025
Biofilm–Host Immune Crosstalk at the Diabetic Foot Ulcer Interface: Molecular Mechanisms, Immune Evasion, and Next-Generation Anti-Biofilm Strategies
Han Y, Yang Y
Diabetes, Metabolic Syndrome and Obesity 2026, 19:608789
Published Date: 22 May 2026