Back to Journals » Drug Design, Development and Therapy » Volume 20
Sacubitril/Valsartan Fails to Ameliorate Cisplatin-Induced Cognitive Impairment
Authors Alshammari AO, Aldubayan MA, Alwashmi AH ![]() , Alhowail AH
, Alhowail AH ![]()
Received 13 February 2026
Accepted for publication 27 May 2026
Published 3 June 2026 Volume 2026:20 561133
DOI https://doi.org/10.2147/DDDT.S561133
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Abdullah Obaid Alshammari,1 Maha A Aldubayan,1 Ahmad H Alwashmi,2 Ahmad H Alhowail1
1Department of Pharmacology and Toxicology, College of Pharmacy, Qassim University, Buraydah, Kingdom of Saudi Arabia; 2Department of Orthopedic Surgery, College of Medicine, Qassim University, Buraydah, Kingdom of Saudi Arabia
Correspondence: Ahmad H Alhowail, Email [email protected]
Background: Chemotherapy-induced cognitive impairment (“chemobrain”) is mechanistically linked to oxidative stress, neuroinflammation, and apoptotic pathway dysregulation. Angiotensin receptor–neprilysin inhibitors and angiotensin II type-1 receptor blockers, such as sacubitril/valsartan (VS), have demonstrated neuroprotective potential; however, their effects in cisplatin (CIS)-induced neurotoxicity models remain poorly characterized.
Methods: Forty male Wistar rats were randomized into four groups: Control, CIS (5 mg/kg, i.p). VS alone (≈40 mg/kg/day in drinking water), and CIS+VS, receiving three doses over 7 days. Cognitive performance was assessed 24 hours after the final dose via the Y-maze and novel object recognition test (NORT). Survival and body weight were monitored throughout. Hippocampal levels of reactive oxygen species (ROS), malondialdehyde (MDA), TNF-α, IL-1β, IL-6, total NF-κB, BAX, and total caspase-3 were quantified by ELISA, complemented by histopathological analysis.
Results: CIS produced dose-limiting toxicity, with 40% mortality and approximately 13% body weight loss over 8 days. The CIS+VS group exhibited greater toxicity, with 50% mortality and approximately 16% weight loss, while the control and VS animals gained weight. Both CIS and CIS+VS groups demonstrated significant impairment in spatial and recognition memory, as reflected by reduced novel-arm entries and time spent in the Y-maze, and diminished novel object exploration in NORT relative to controls. Hippocampal oxidative stress, neuroinflammatory, and apoptosis-related signaling markers were markedly elevated following CIS and remained comparably elevated in the CIS+VS group, consistent with persistent neuroinflammation. Histopathological examination confirmed severe hippocampal tissue damage in both CIS-treated groups, with no attenuation observed in the CIS+VS group.
Conclusion: CIS induces significant cognitive deficits accompanied by elevated neurotoxicity markers. Under the tested dosing schedule, adjunctive VS failed to rescue cognitive function or attenuate hippocampal neuro-injury. These findings reinforce oxidative stress, neuroinflammation, and apoptotic signaling as key mechanistic drivers of chemobrain and indicate that VS confers no neuroprotective benefit in this model. The experiment involves 40 Wistar rats divided into four groups: Control, CIS (5 mg/kg, i.p., 3 doses), VS alone (40 mg/kg/day, water) and CIS+VS. Dosing occurs from Day 0 to Day 5. Assessments include Y-Maze for spatial memory on Day 8, NORT for recognition memory on Day 9 and hippocampal tissue extraction on Day 10. Results show systemic toxicity with survival rates and body weight changes. Control and VS groups gain weight, while CIS and CIS+VS groups lose 13-16% weight. Cognitive tests reveal impairment in CIS and CIS+VS groups. Hippocampal markers indicate oxidative stress, neuroinflammation and apoptotic signaling, with severe tissue damage. Conclusion: CIS-induced neurotoxicity (’Chemobrain’) is driven by oxidative stress, neuroinflammation and apoptosis. Sacubitril/valsartan (VS) did not improve cognitive function or reduce neuro-injury in this dosing schedule.A flowchart detailing an experimental design with results on CIS-induced neurotoxicity in Wistar rats.
Keywords: cisplatin, sacubitril, valsartan, cognitive impairment, oxidative stress, inflammation, apoptosis-related signaling
Introduction
Chemotherapy-induced cognitive impairment (“chemobrain”) is a frequent and clinically significant adverse effect of cancer treatment, particularly with CIS. Affected patients report deficits in memory, attention, executive function, and processing speed that can persist long after therapy and diminish their quality of life.1,2 The pathophysiology of CIS-related cognitive dysfunction is multifactorial and involves neuroinflammation, oxidative stress, mitochondrial dysfunction, and disruption of neuronal–glial homeostasis.3,4 Despite increasing recognition of this syndrome, effective pharmacological strategies to prevent or mitigate these deficits remain limited.1
Cisplatin (CIS) is a cornerstone chemotherapeutic agent for ovarian, testicular, lung, and head-and-neck cancers; however, its clinical utility is constrained by neurotoxicity.5 Preclinical and clinical investigations have suggested that CIS can cross the blood–brain barrier, accumulate within the central nervous system, and perturb neuronal function.6 Proposed mechanisms include excessive reactive oxygen species (ROS) production, malondialdehyde (MDA) and mitochondrial injury leading to neuronal apoptosis and release apoptosis-related signaling markers such as BAX and Caspase-3, activation of microglia and astrocytes that sustain a pro-inflammatory milieu by elevated expression of IL-1β, IL-6, and TNF-α, alongside changes in the NF-κB signaling pathway, which is known to drive transcription of multiple pro-inflammatory mediators, and impairment of hippocampal synaptic plasticity and long-term potentiation, processes essential for learning and memory.5,7 These converging pathways offer a biologically plausible explanation for persistent cognitive decline after CIS exposure, while underscoring the lack of proven neuroprotective interventions.3
Sacubitril/valsartan (VS), an angiotensin receptor–neprilysin inhibitor approved for the treatment of heart failure with reduced ejection fraction, integrates neprilysin inhibition (sacubitril) with angiotensin II type-1 receptor blockade (valsartan).8 In addition to its hemodynamic effects, this dual mechanism enhances natriuretic peptide signaling, which possesses vasodilatory, antioxidative, and anti-inflammatory properties, while simultaneously mitigating angiotensin II–induced oxidative stress, vascular dysfunction, and inflammation.9 VS is rapidly metabolized into its active neprilysin-inhibiting form, LBQ657, which has been observed in cerebrospinal fluid in both animal and human studies. This finding implies limited blood-brain barrier (BBB) permeability.10 While VS is predominantly prescribed for cardiovascular conditions, emerging evidence suggests that it may indirectly modulate neuroinflammatory pathways relevant to cognitive function. VS has been shown to mechanistically inhibit NF-κB signaling, a crucial pathway that regulates pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6. This inhibition consequently diminishes neuroinflammation and glial activation.11 Furthermore, VS demonstrates significant antioxidant properties, mitigating the oxidative stress that frequently accompanies inflammatory pathways and contributes to neuronal damage.12 By integrating neprilysin inhibition with angiotensin II type-1 receptor blockade, VS has the potential to enhance nitric oxide bioavailability and improve microvascular function, thereby promoting neurovascular health and cognitive function.13 Reflecting these attributes, VS has been shown to alleviate pathological tissue remodeling in extra-neuronal organs, including hepatic fibrosis, by suppressing oxidative stress, inflammatory mediators, and NF-κB activation.14
While VS exhibits limited direct permeability across the BBB, emerging evidence indicates that its neuroprotective effects may be mediated through indirect systemic mechanisms rather than direct CNS entry. By simultaneously inhibiting the renin–angiotensin system and neprilysin, VS mitigates peripheral oxidative stress and the release of inflammatory cytokines, thereby reducing systemic signals that could exacerbate central neuroinflammation.15 Several preclinical studies have notably demonstrated that VS enhances cognitive performance and reduces neuronal damage in models of Alzheimer’s disease and cerebral ischemia, despite its limited ability to penetrate the BBB.16,17 These findings collectively substantiate the rationale for utilizing VS in CNS injury models, as its modulation of systemic inflammatory, vascular, and mitochondrial pathways may indirectly facilitate neuroprotection and enhance cognitive outcomes.
Therefore, this study aimed to evaluate the neuroprotective potential of sacubitril/valsartan against CIS-induced neurotoxicity by examining its ability to downregulate peripheral and central oxidative and inflammatory markers and modulate the apoptotic signaling pathway.
Materials and Methods
Chemicals
Cisplatin (1 mg/mL) was purchased from EBEWE Pharma Ges mbH Nfg.KG (Austria). Sacubitril/valsartan (Entresto®, 100 mg film-coated tablets) were purchased from Novartis Pharma AG, Basel, Switzerland. Phosphate-buffered saline (PBS), CO2 for euthanasia, N-PER™ tissue protein extraction reagent (Thermo Scientific, Madison, WI, USA), and bicinchoninic acid (BCA) protein assay kits were obtained from standard suppliers. ELISA kits were obtained from ELK Biotechnology, Denver, CO, USA: MDA [Cat. No. RK15281], ROS [Cat. No. RK15283], (IL-1β [Cat. No. ELK1272], IL-6 [Cat. No. ELK1158], TNF-α [Cat. No. ELK1396], and NF-κB [Cat. No. ELK1693], caspase-3 [Cat. No. RK03549], and BAX [Cat. No. RK03522].
Animals and Ethics
Male Wistar rats (12 weeks old; 170–215 g) were obtained from the College of Pharmacy, Qassim University Animal Facility. Rats were housed individually under specific pathogen-free conditions (22 ± 2 °C, 50 ± 10% humidity, 12 h light/dark cycle, lights on 06:00) with ad libitum access to chow and water. The animals were acclimated for ≥7 d before experimentation. The procedures conformed to the institutional guidelines, adhered to ARRIVE 2.0, and were approved by the Qassim University IACUC (protocol ID: 25–19-21). Humane endpoints included >20% body weight loss, persistent inability to eat or drink, severe lethargy, and moribund state. At the end of the experimental procedures, or upon reaching a humane endpoint, animals were euthanized by CO2 inhalation using a dedicated chamber at a gradual displacement rate of 10–30% of the chamber volume per minute, followed by cervical dislocation as a secondary method to confirm death. All euthanasia procedures were performed in strict accordance with the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals (2020 Edition).
Drug Preparation and Administration
CIS was administered intraperitoneally (i.p.) at 5 mg/kg per injection on days 1, 4, and 7 (cumulative dose of 24 mg/kg).3,5,18,19 The VS group received Entresto tablets containing 100 mg each, freshly prepared daily before administration. This specific dosage (60 mg/kg/day) was selected based on previous preclinical investigations demonstrating that it is sufficient to achieve systemic anti-inflammatory, antioxidant, and potential neuroprotective effects in rodent models without inducing severe hypotension. The tablets were finely ground and suspended in 10 mL of distilled water, yielding a concentration of 100 mg/10 mL. The dosage for each rat was determined by body weight, with each animal receiving 60 mg/kg/day via oral gavage.20 VS dosing began on Day 1 (prophylaxis) and continued through Day 7. On CIS injection days (Days 1, 4, and 7), VS was administered approximately 120 minutes before CIS. The control animals received matched volumes of vehicle. The mortality rate and body weight were observed and recorded daily. A total of five animals per group were included in the final analysis, accounting for any treatment-related mortality. Following completion of behavioral assessments, animals underwent hippocampal tissue collection for histopathological evaluation and enzyme-linked immunosorbent assay (ELISA)-based biochemical analyses (Figure 1).
 |
Figure 1 Experimental timeline of sacubitril/valsartan (VS) and cisplatin (CIS) treatment protocol. |
Evaluation of Mortality and Body Weight Changes
Mortality and body weight were monitored daily throughout the experimental period. All animals were weighed at the same time each day (between 08:00 and 09:00 h) using a calibrated digital balance, and the values were recorded. Body weight changes were calculated as the percentage difference between the initial body weight (Day 1) and the final body weight (Day 8) using the following formula:
Body weight change (%) = [(Final body weight − Initial body weight) / Initial body weight] × 100
Mortality was recorded daily, and the survival rate for each group was calculated as the percentage of animals surviving at the end of the experimental period relative to the initial number of animals per group (n = 10). The Kaplan–Meier survival curve was generated to illustrate the survival rate across all experimental groups.
Experimental Design, Randomization, and Blinding
Forty male Wistar rats were randomly allocated to four groups (n = 10/group) using a random number table The sample size of 10 animals per group was determined based on previous behavioral and biochemical studies in rodent models of chemobrain, which indicated this number provides sufficient statistical power (80% at α=0.05) to detect meaningful differences in cognitive and biochemical outcomes while accounting for potential CIS-induced mortality. Allocation concealment was ensured through sealed opaque envelopes prepared by an independent investigator who was not involved in the study conduct or data analysis:
- Control (saline)
- VS (sacubitril/valsartan alone)
- CIS (cisplatin alone)
- CIS+VS (cisplatin + sacubitril/valsartan)
Experimenters were blinded to group allocation for behavioral scoring and ELISA analyses, and an independent technician coded the cages. Pre-specified exclusion criteria were as follows: (<) 3 total arm entries in the Y-maze test session (hypoactivity), exploration time <10 s across both objects in NORT, or technical assay failure.
Behavioral Testing
Behavior was assessed during the light phase by a blinded assessor in a quiet room under uniform illumination. The apparatus was cleaned with 70% ethanol between trials to minimize olfactory cues. Tests were conducted in the following order, with ≥24 h between paradigms: Y-maze (training/test on Day 8) → NORT (familiarization and test on Day 9). Behavioral assessments were performed on Days 8 and 9. On these days, VS was administered after the completion of behavioral testing to minimize potential acute pharmacological effects on cognitive performance. Tissue collection was performed 24 hours after the final treatment (Day 7).
Y-Maze
A three-armed maze (50 × 10×15 cm; 120° between arms) with a painted matte brown was used. Visual cues (circles, triangles, and Xs) were placed on the walls. Training (15 min): one arm was blocked, and the rats explored the start arm and one open arm—inter-trial interval: 3 h in the home cage. In the test session, rats were confined to the start arm for 5 min, then allowed to freely explore all three arms for 5 min. The trials were video-recorded and scored offline by two blinded raters (inter-rater ICC > 0.9). Arm entry was defined as entry of >50% of body length. Outcomes: (i) novel-arm entries, (ii) time spent in novel arm (s), and (iii) total arm entries (locomotor control) (Figure 2A).21
 |
Figure 2 Schematic representation of the behavioral assessment procedures used in this study. (A) Y-maze spontaneous alternation test. (B) Novel Object Recognition (NOR) test. |
Novel Object Recognition Test (NORT)
Testing was performed in a wooden open field (41 cm × 41 cm × 41 cm). Habituation: 24 h free exploration (no objects). Familiarization (15 min): Two identical objects (ceramic teacups) were placed equidistant from the walls; object identity and positions were counterbalanced across rats. Inter-trial interval: 3 h. Test (5 min): one familiar object was replaced with a novel rectangular box of similar volume; positions were counterbalanced. Exploration was defined as sniffing or directing the nose to an object ≤2 cm; sitting on or leaning against an object was not scored. Outcomes: Time with novel objects, and discrimination index (DI) = (T_novel − T_familiar)/(T_novel + T_familiar). Animals with a total exploration time of <10 s were excluded per protocol. All trials were video-recorded and scored offline by two independent assessors who were blinded to the treatment conditions, ensuring unbiased evaluation. An inter-rater reliability analysis was performed to ensure consistency between scorers (Figure 2B).22
Tissue Collection and Processing
Following behavioral testing, five animals per group (n = 5) that survived to the predetermined experimental endpoint were included in the final analyses. For each animal, one hippocampus (one hemisphere) was fixed in 10% neutral-buffered formalin for histopathological processing, which included paraffin embedding and hematoxylin and eosin (H&E) staining. The contralateral hippocampus was rapidly snap-frozen in liquid nitrogen and stored at −80 °C for subsequent biochemical analyses. This design ensured that both histological and biochemical outcomes were obtained from the same animals. Frozen hippocampal tissues were homogenized on ice using a Qsonica probe homogenizer (30 Hz; Qsonica LLC, Newtown, CT, USA) in N-PER extraction buffer supplemented with protease and phosphatase inhibitors (1:100). Homogenates were centrifuged at 12,000 × g for 10 min at 4 °C, and the resulting supernatants were collected. Total protein concentration was determined using the BCA assay, and all ELISA results were normalized to total protein content and expressed as pg or ng per mg protein.
ELISA for Neuroinflammation, Oxidative Stress, and a Supportive Indicator of Apoptotic Signaling Measurements
Hippocampal levels of oxidative stress markers, inflammatory cytokines, and apoptosis-related proteins were quantified using commercially available sandwich ELISA kits (ELK Biotechnology, Denver, CO, USA) according to the manufacturer’s instructions.
Oxidative Stress Markers
Reactive oxygen species (ROS) levels were measured using an ELISA kit (Cat. No. RK15283). This assay is based on the principle of competitive enzyme immunoassay, where ROS in the sample competes with a fixed amount of ROS on the solid phase for binding sites of the anti-ROS antibody. ROS concentrations were expressed as U/mL.3,23 Malondialdehyde (MDA), a biomarker of lipid peroxidation, was quantified using an ELISA kit (Cat. No. RK15281). The assay utilizes a double-antibody sandwich technique, where MDA in the sample binds to a pre-coated capture antibody and is subsequently detected by a biotinylated secondary antibody. MDA levels were normalized to total protein content and expressed as pg/mg protein.3,5
Inflammatory Markers
Pro-inflammatory cytokines, including tumor necrosis factor-alpha (TNF-α; Cat. No. ELK1396), interleukin-1 beta (IL-1β; Cat. No. ELK1272), interleukin-6 (IL-6; Cat. No. ELK1158), and total nuclear factor kappa B (NF-κB; Cat. No. ELK1693) protein levels were measured using their respective ELISA kits.
Apoptotic Markers
Total caspase-3 protein (Cat. No. RK03549) and Bcl-2-associated X protein (BAX; Cat. No. RK03522) levels were quantified as indicators of apoptotic signaling.
For all assays, 50 µL of hippocampal homogenate supernatant or standard was added to the appropriate wells and incubated at 37°C. Following washing steps, biotinylated detection antibody and streptavidin-horseradish peroxidase (HRP) conjugate were added sequentially. Color development was achieved using 3,3′,5,5′-tetramethylbenzidine (TMB) substrate, and the reaction was terminated with stop solution. The optical density was measured at 450 nm using an ELx800 microplate reader (BioTek, Winooski, VT, USA). Concentrations were calculated from standard curves, and all results were normalized to total protein content determined by bicinchoninic acid (BCA) assay.
Histopathology Evaluation and Scoring of Hippocampus Injury
Hippocampal samples (n=5) were immediately fixed in 10% neutral buffered formalin for a duration of 24–48 hours. Following fixation, the tissues were dehydrated through a graded ethanol series, cleared in xylene, and embedded in paraffin wax. Serial coronal sections, with a thickness of 4–5 μm, were prepared using a rotary microtome. These sections were mounted on glass slides, deparaffinized, rehydrated, and stained with hematoxylin and eosin (H&E) according to standard histological procedures. Semi-quantitative assessment of hippocampal neuronal injury was performed using ImageJ software (Fiji distribution, National Institutes of Health, USA). Digitized H&E-stained images were captured under identical magnification and illumination settings and calibrated using the scale bar. A standardized region of interest (ROI) was placed over the CA1 pyramidal layer, and neurons exhibiting morphological features of injury, such as nuclear pyknosis, cytoplasmic eosinophilia, and shrunken cell bodies, were quantified using the Cell Counter plugin. Injury scores were assigned based on the severity and extent of neuronal degeneration. The scale ranges from 0 (none) to 4 (severe). All slides were examined in a blinded manner, and the mean score was used for analysis. The stained slides were examined under a light microscope at 400X magnification, and representative images were captured.
Outcomes
The primary outcomes were survival, body weight changes, cognitive performance (Y-maze novel-arm time and entries; NORT-DI), and hippocampal inflammatory marker levels (TNF-α, IL-1β, IL-6, and total NF-κB). The secondary outcomes included mitochondrial, ROS, MDA, caspase-3, and Bax levels.
Data Analysis
All data are expressed as the mean ± standard error of the mean (SEM). Prior to analysis, the normality of the data distribution was assessed using the Shapiro–Wilk test. Following confirmation of normal distribution, one-way analysis of variance (ANOVA) was applied, followed by Tukey’s post-hoc test. Statistical analyses were performed using GraphPad Prism 10.6. One-way analysis of variance (ANOVA) was applied, followed by Tukey’s post-hoc test. Statistical significance was set at p < 0.05. The group comparisons of the histopathological injury scores were performed using the Kruskal–Wallis test, followed by Dunn’s post hoc test.
Result
Effect of CIS and VS Treatments on the Survival Rate
CIS treatment affected the survival rates, with a 50% mortality rate observed in the CIS-treated rat cohort. Conversely, the CIS + VS group exhibited a 60% mortality rate following the 8-day treatment period (Figure 3).
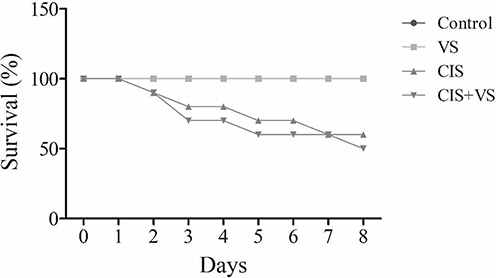 |
Figure 3 Effects of CIS and VS on survival. The survival rate of the rats was determined after 10 days of treatment with CIS, VS, CIS+VS, and the control. Data represent mean ± SEM (n = 10 for each group). CIS, Cisplatin; VS, Sacubitril/Valsartan. |
Effect of VS Treatment on CIS-Induced Loss in Body Weight
The CIS and CIS+VS treatments resulted in a gradual reduction in body weight throughout the 10-day study period (approximately 16% and 11%, respectively). In contrast, the control and VS-alone groups showed an increase in body weight (approximately 11% and 7%, respectively) (Figure 4).
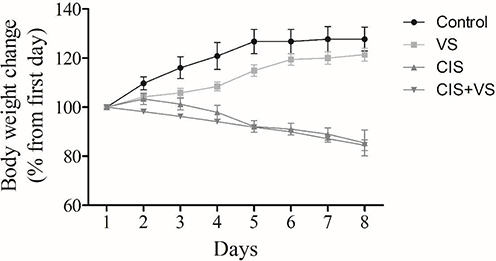 |
Figure 4 Effects of CIS and VS on body weight. The body weights of the rats were determined during the study period, before and after treatment with CIS, VS, and CIS+VS, and the control. CIS, Cisplatin; VS, Sacubitril/Valsartan. |
Effect of VS Treatment on CIS-Induced Cognitive Impairment in the Y-Maze
Analysis of entries into the novel arm of the Y-maze revealed significant differences between groups (Figure 5A). The control group comprised approximately five entries. The VS group had significantly fewer entries (2.5, p < 0.01 compared to the control. The CIS group also showed a reduction in entries (1.8), although this was not statistically significant compared with the control group. The CIS + VS group had the fewest entries, approximately 0.5, which was significantly lower than the control group (p < 0.01).
 |
Figure 5 Y-maze performance across experimental groups. (A) Number of entries to the novel arm. (B) Time spent in the novel arm (seconds). (C) Total number of entries into all arms. Data are presented as mean ± SEM (n=5) for each group: Control, VS (Sacubitril/Valsartan), CIS (cisplatin), and CIS+VS (combined treatment). Statistical significance: * p < 0.05, ** p < 0.01 compared to control; ns, not significant. |
The time spent in the novel arm also varied significantly between groups (Figure 5B). The control group spent approximately 75s in the novel arm. This was significantly higher than that in the VS group (25 s, p < 0.05) and CIS group (20 s, p < 0.05). The CIS+VS group also spent less time in the novel arm (~25 s), which was comparable to the VS and CIS groups but did not differ significantly from the control group in the analysis.
Finally, the total number of entries into all the arms of the maze was examined as a measure of general locomotor activity (Figure 5C). The control group had approximately 11.5 entries. While the VS (7.5 entries) and CIS (6 entries) groups showed a trend toward reduced activity, neither group differed significantly from the control group. However, the CIS+VS group made significantly fewer total entries (~3 entries, p < 0.01 compared to the control), indicating a substantial reduction in overall locomotor activity.
Effect of VS Treatment on the CIS Novel Object Recognition Test
There was a significant effect of treatment on the time spent exploring novel objects (Figure 6). The control group spent approximately 34 seconds engaging in this behavior. In contrast, the CIS group showed a significant reduction in exploration time to approximately 10s (p < 0.01 compared to the control). An even greater reduction was observed in the combined CIS+VS group, which spent only approximately 5 s exploring the novel objects (p < 0.01 compared to the control group). The VS group also showed a trend towards reduced exploration (~24 s), but this was not statistically significant compared to the control group. No significant differences were found between the VS, CIS, and CIS + VS groups (Figure 6A). To further assess recognition memory, the Discrimination Index (DI) was calculated. The analysis revealed a significant effect of treatment on the DI. The CIS and CIS+VS groups showed a significantly lower DI compared to the control group (p < 0.01), indicating impaired discrimination between the novel and familiar objects. There was no significant difference in the DI between the CIS and CIS+VS groups, suggesting that sacubitril/valsartan did not rescue the cognitive deficit induced by CIS (Figure 6B).
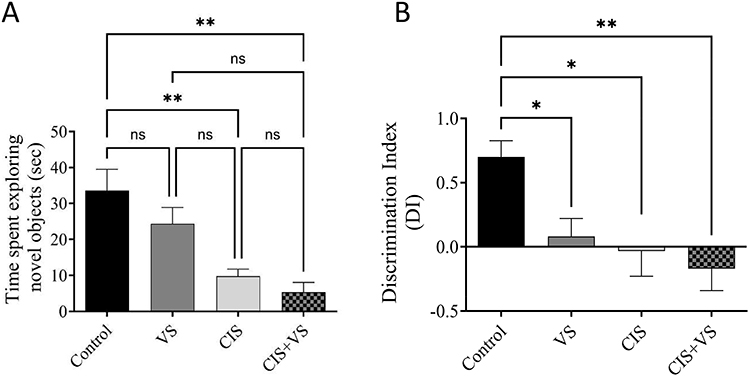 |
Figure 6 Novel Object Recognition Test Performance. (A) Time spent exploring the novel object and (B) the Discrimination Index (DI) across the four treatment groups: Control, VS (Sacubitril/Valsartan), CIS (cisplatin), and CIS+VS (combined treatment). Data are presented as mean ± SEM (n=5 per group). Statistical significance was determined by one-way ANOVA with Tukey’s post-hoc test. *p < 0.05, **p < 0.01 compared to the control group; ns, not significant. |
Effect of VS in Conjunction with CIS on Oxidative Stress Markers in Rat Hippocampal Tissues
In the CIS-treated group, ROS and MDA levels were significantly increased compared with those in the VS alone and the control groups. Furthermore, the CIS+VS-treated groups showed no significant difference in ROS and MDA levels when compared with the CIS group (Figure 7A and B).
 |
Figure 7 Effect of VS administration on CIS-induced alterations in oxidative stress markers. (A) ROS, and (B) MDA in rat hippocampal tissue. Data are expressed as the mean ± SEM (n=5) and analyzed using one-way ANOVA followed by Tukey’s post hoc test, with statistical significance set at ***p < 0.001, ****p < 0.0001, compared with the control group; ns, not significant. |
Effect of VS Treatment on CIS-Induced Neuroinflammation
The results demonstrated significantly increased total NF-κB, TNF-α, IL-1β, and IL-6 protein levels in both CIS and CIS + VS treatment groups, relative to the control; however, the levels of these proteins were comparably elevated in the CIS + VS group. Protein levels were normalized to the total protein content and expressed as a percentage of the control group (set at 100%) (Figures 8A and 7B).
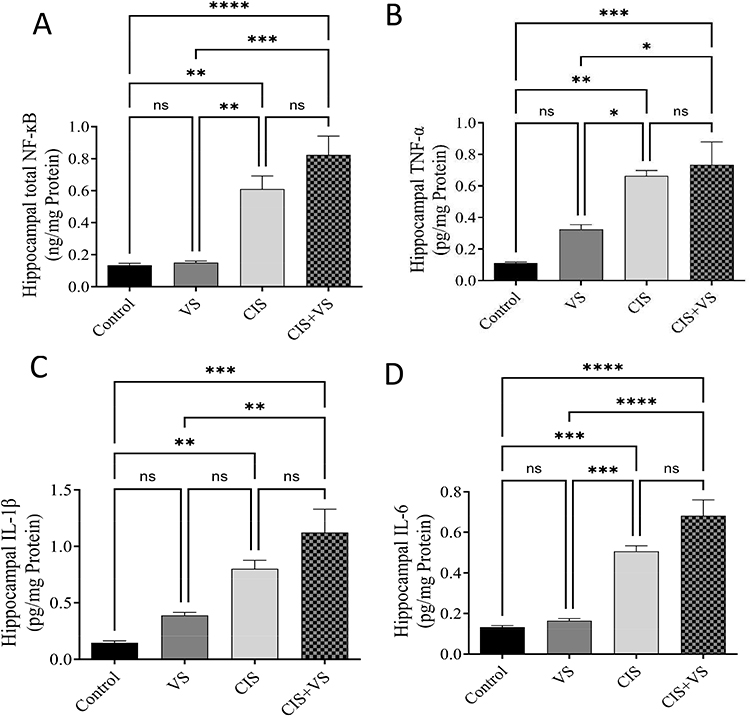 |
Figure 8 Effects of CIS and VS on TNF-α and IL-1β expression in the rat hippocampus. The effects of CIS and VS on (A) total NF-κB, (B) TNF-α, (C) IL-1β, and (D) IL-6 levels in the rat brain were evaluated by ELISA after 10 days of treatment with CIS, VS, and CIS+VS (the control group did not receive any treatment). Data are expressed as the mean ± SEM (n=5) and analyzed using one-way ANOVA followed by Tukey’s post hoc test, with statistical significance set at ****p < 0.0001, ***p < 0.001, **p < 0.01, and *p < 0.05 compared with the control group; ns, not significant. |
Effect of VS in Conjunction with CIS on Apoptotic Markers in Rat Hippocampal Tissues
In the CIS-treated group, caspase-3 and BAX levels were significantly increased compared with those in the VS-alone or control groups. However, CIS+VS administration resulted in a significant elevation in caspase-3 compared with all other groups. Further, no significant difference was observed in BAX when compared with the CIS group (Figure 9A and B).
 |
Figure 9 Effect of VS administration on CIS-induced alterations in apoptotic markers. (A) BAX and (B) caspase-3 in rat hippocampal tissue. Data are expressed as the mean ± SEM (n=5) and analyzed using one-way ANOVA followed by Tukey’s post hoc test, with statistical significance set at ***p < 0.001, ****p < 0.0001, and *p < 0.05 compared with the control group; ns, not significant. |
Hippocampal Histopathology (CA1 Region)
Hematoxylin and eosin (H&E)-stained sections of the hippocampus demonstrated distinct structural changes in the CA1 pyramidal layer that were dependent on the treatment administered. In Panels A and B, the pyramidal cell layer was intact, compact, and well-organized, with neurons densely packed and exhibiting normal morphology. The neuronal soma appeared round with clear nuclei, and there was no evidence of pyknosis, shrinkage, or dropout. The surrounding neuropil displayed a uniform texture, devoid of vacuolation or edema. The findings suggest that the CA1 region exhibits normal cytoarchitecture with no observable neuronal damage. Conversely, the group treated with CIS (Panel C) displayed significant neurodegenerative alterations, characterized by neuronal pyknosis, cytoplasmic eosinophilia, reduced compactness and organization, diminished pyramidal cell density, and extensive neuropil rarefaction. The VS+CIS group (Panel D) showed similarly pronounced structural damage, including extensive neuronal degeneration, disorganization of the CA1 layer, and widespread neuropil rarefaction, indicating that VS did not provide structural protection against CIS-induced neurotoxicity (Figure 10A). The combination of VS with CIS showed no difference in injury scores compared with those seen with CIS treatment alone (Figure 10B).
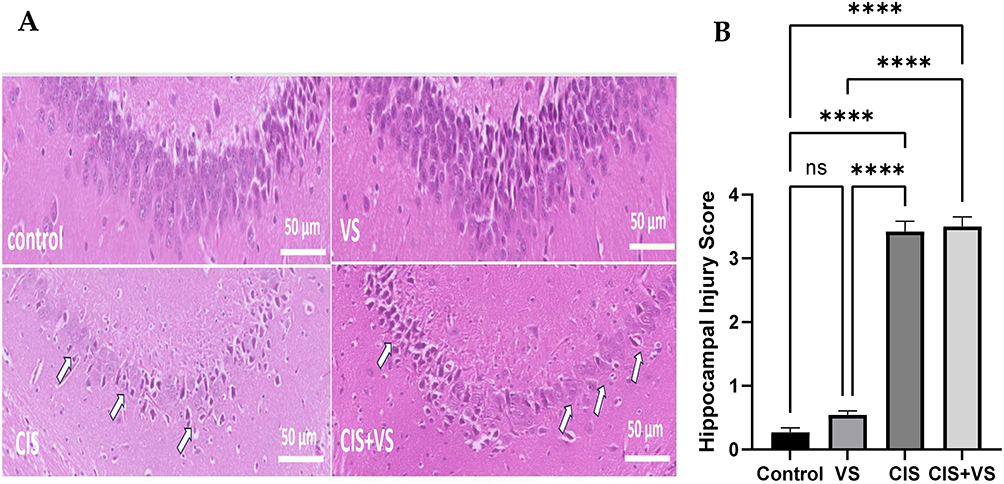 |
Figure 10 Representative histological images of CA1 hippocampal sections stained with hematoxylin and eosin (scale bar = 50 µm) are presented. (A) The control group displays normal histoarchitecture of the pyramidal cell layer with intact neuronal morphology. The VS-treated group exhibits preserved neuronal density and architecture. The CIS-treated group demonstrates significant neuronal injury (indicated by arrows), characterized by shrunken eosinophilic neurons, nuclear pyknosis, widespread vacuolated neuropil, and disorganization of the pyramidal cell layer. The CIS+VS-treated group shows severe neuronal degeneration and loss of pyramidal cell organization (indicated by arrows), with no significant improvement compared to the CIS group. (B) The hippocampal injury score. The quantitative hippocampal injury score was obtained using ImageJ-based analysis, and data are presented as mean ± SEM (n=5). The histopathological injury scores were evaluated using the Kruskal–Wallis test, followed by Dunn’s post hoc test at ****p < 0.0001, compared with the control group; ns, not significant. |
Discussion
Cisplatin remains the cornerstone of chemotherapy for various solid tumors; however, its clinical utility is frequently hampered by severe dose-limiting side effects, including debilitating neurotoxicity and cognitive impairment, often termed “chemobrain”.1,2 The present study was designed to investigate whether sacubitril/valsartan (VS) could offer a potential neuroprotective effect against CIS-induced neurotoxicity in a rat model. Our findings demonstrate that CIS administration resulted in a marked reduction in body weight throughout the experiment, likely due to anorexia, gastrointestinal toxicity, and systemic inflammation associated with its use. The survival rate was approximately 50% in the CIS group and 60% in the CIS+VS group, whereas all animals in the control and VS groups survived (100%). These findings indicate the systemic toxicity of CIS and confirm the validity of this model for studying chemotherapy-induced complications.21,24
To evaluate the effects of cisplatin (CIS) and sacubitril/valsartan (VS) on cognitive performance, a battery of hippocampal-dependent behavioral tasks was conducted. In the Y-maze test, a well-established paradigm for assessing spatial working memory,25 the neurotoxic impact of CIS was clearly demonstrated. Specifically, rats administered CIS exhibited a statistically significant reduction in both the number of entries into the novel arm and the time spent within it, as compared to the vehicle-treated control group. This outcome signifies a pronounced cognitive deficit, whereas the control group displayed intact cognitive function. The concurrent administration of VS with CIS failed to reverse these behavioral impairments. To rule out the possibility that these deficits were a mere consequence of reduced physical activity, locomotor function was assessed by quantifying the total number of arm entries. Interestingly, the group receiving the combined VS and CIS treatment was the only group to show a significant decrease in total arm entries when compared to the control group, with no significant differences observed among the other experimental groups. This reduction in locomotor activity in the VS+CIS group is hypothesized to be a consequence of the observed synergistic elevation in oxidative stress and apoptosis.
To further investigate the cognitive repercussions, the Novel Object Recognition (NOR) test was employed to assess short-term recognition memory. The results of this test corroborated the findings from the Y-maze, revealing that rats in both the CIS and the CIS+VS treatment groups spent significantly less time exploring the novel object in comparison to the control group. To provide a more robust measure of recognition memory, a Discrimination Index (DI) was calculated. This analysis revealed that both the CIS and CIS+VS groups had a significantly lower DI than the control group, indicating a profound impairment in the ability to discriminate between novel and familiar objects. Critically, there was no statistically significant difference in the DI between the CIS and CIS+VS groups, which strongly suggests that the co-administration of VS did not confer any therapeutic benefit or alleviate the cognitive deficits induced by CIS. These behavioral findings collectively confirm the presence of significant cognitive impairment resulting from CIS administration, a conclusion that is in alignment with previous research in the field.5
Our biochemical findings provide a plausible mechanistic basis for the observed behavioral deficits, particularly the decreased locomotor activity in the VS+CIS-treated animals. In our investigation, the administration of CIS markedly increased ROS production and MDA levels within the hippocampus, supporting the recognized role of oxidative damage in CIS-induced neurotoxicity.26 However, the simultaneous administration of VS failed to mitigate these oxidative changes; in fact, the VS+CIS group showed even higher ROS production than the CIS group alone, demonstrating that VS was ineffective in influencing CIS-induced oxidative stress pathways and potentially aggravated them. This exacerbation of oxidative stress is critical, as excessive ROS generation is known to trigger mitochondrial dysfunction and activate intrinsic apoptotic pathways, ultimately increasing caspase-3 activity.27,28 Consistent with this, our findings demonstrated that caspase-3 levels were significantly higher in the VS+CIS group than in the CIS group, indicating enhanced neuronal apoptosis. This synergistic rise in both oxidative stress and apoptosis can impair neuronal circuits involved in locomotion, motivation, and exploratory behavior.29 Given that CIS neurotoxicity is strongly driven by ROS-mediated apoptotic mechanisms,25 which appear to be amplified when combined with VS, animals in the VS+CIS group likely experience greater neurotoxicity. This is evidenced by reduced spontaneous activity and explains their significantly lower number of arm entries compared with the CIS group. Thus, the combined elevation of ROS and caspase-3 offers a coherent explanation for the heightened behavioral impairment observed in the VS+CIS-treated animals.
Beyond the oxidative stress profile, the rise in inflammatory biomarkers, including IL-6, IL-1β, TNF-α, and NF-κB, is linked to the CNS injury and cognitive behavioral deficits.30 Our study observed a significant increase in these biomarkers within the CIS group compared to the control group, corroborating previous findings of elevated neuroinflammatory markers.23 The production of TNF-α further stimulated the synthesis of additional inflammatory mediators via NF-κB activation, leading to elevated IL-1β and IL-6 levels. This neuroinflammatory cascade is closely associated with the onset of cognitive impairment and suggests that effective neuroprotective strategies must address this complex cytokine network rather than targeting a single pathway.31 Furthermore, the CIS+VS group did not exhibit a significant effect on inflammatory cytokine markers compared with the CIS group alone. This observation indicates that the drug did not protect by reducing levels of inflammatory markers, which contrasts with previous studies on cardiac tissue that reported reductions in inflammation.32
It is important to acknowledge a key limitation in our assessment of the NF-κB pathway. In this study, we quantified total NF-κB protein levels. This measurement does not distinguish between the inactive cytosolic and active nuclear forms of NF-κB. To definitively characterize the activation state of this pathway, future studies must determine the phosphorylated forms of the p65 subunit and its inhibitor IκBα, which would provide direct evidence of transcriptional activation. The activation of NF-κB is a complex process involving the phosphorylation and subsequent degradation of its inhibitory subunit, IκBα, which permits the translocation of the active p65 subunit to the nucleus. Therefore, while our findings indicate an upregulation of total NF-κB protein, which may contribute to a heightened inflammatory state, they do not provide conclusive evidence of increased NF-κB-mediated transcription. Future studies should incorporate more specific methods, such as Western blot analysis of phosphorylated p65 and IκBα levels or electrophoretic mobility shift assays (EMSA), to definitively characterize the activation state of the NF-κB signaling pathway in this model.
Our findings summarize that while VS provides robust anti-inflammatory, antioxidant, and anti-apoptotic protection in the heart,32–34 it fails to confer similar benefits in the brain during CIS-induced neurotoxicity. This is further elucidated in the context of apoptosis-related signaling, where VS did not protect the hippocampus against CIS-induced cell death. Although total caspase-3 levels were elevated in the CIS+VS group compared with CIS alone, this finding should be interpreted as reflecting an altered apoptotic signaling potential rather than a definitive enhancement of executioner caspase activation, as cleaved caspase-3 was not specifically assessed. This suggests a dysregulation of apoptosis-related pathways without conclusive evidence of an additive or synergistic pro-apoptotic interaction. Several organ-specific pharmacokinetic and mechanistic factors may account for this discrepancy. Firstly, VS exhibits limited penetration across the BBB, leading to reduced CNS drug exposure and inadequate modulation of neuroinflammatory, oxidative, and apoptotic pathways.35 Secondly, CIS-induced neurotoxicity is predominantly mediated by microglial activation, mitochondrial oxidative stress, and DNA-damage–driven apoptotic signaling, which are mainly independent of the renin-angiotensin-aldosterone system (RAAS) and thus exhibit limited responsiveness to VS.33,36 Thirdly, the sacubitril-mediated enhancement of natriuretic peptide signaling, a crucial anti-inflammatory and anti-fibrotic mechanism in the myocardium, is significantly weaker in the CNS due to lower receptor density and restricted peptide access.37
Histopathological examination of the hippocampal CA1 region revealed a distinct, graded pattern of neuronal vulnerability across the analyzed panels, indicative of progressive structural deterioration due to neurotoxic insult. In the control and VS treated groups exhibited preserved cytoarchitecture, characterized by a compact, uninterrupted pyramidal cell layer, indicating the integrity of hippocampal neurons under non-injurious conditions. This normal morphology aligns with existing literature on healthy CA1 neurons. Conversely, CIS treated group showed degenerative changes, including reduced neuronal density and focal pyknosis. These alterations indicate stages of neuronal distress, often associated with oxidative stress, mitochondrial dysfunction, or excitotoxic signaling. The observed loosening of the neuropil in this panel may further suggest edema or synaptic compromise, a phenomenon identified as a precursor to more extensive neuronal degeneration in hippocampal injury models. The most severe alterations were observed in VS + CIS group, where pronounced neuronal dropout, shrunken hyperchromatic nuclei, and disruption of the pyramidal layer confirmed moderate CA1 neurodegeneration. The combination of architectural disorganization and neuropil rarefaction suggests ongoing apoptotic and necrotic processes. These findings are consistent with the present study, demonstrating that the VS+CIS treatment resulted in significantly higher caspase-3 and oxidative ROS production compared to CIS treatment alone, highlighting the particular vulnerability of this region to metabolic stress and chemotherapeutic neurotoxicity due to its high energy demand and limited antioxidant defenses.
This study has several significant strengths. One of the primary strengths is the uniform application of the same animal strain, age range, and sex, which effectively reduces biological variability. Additionally, while the cardioprotective effects of VS have been extensively documented in the literature, this study is unique in its focus on evaluating VS’s effects on oxidative stress, neuroinflammation, apoptotic pathways, and cognitive function, specifically in rats treated with CIS. This study notably integrated behavioral assessments with biochemical analyses of the hippocampus, thereby providing a comprehensive understanding of functional changes in the brain. However, the study faced certain limitations. The lack of a treatment group receiving only valsartan restricts the ability to discern whether the absence of a neuroprotective effect was due to neprilysin inhibition, angiotensin receptor blockade, or a possible pharmacodynamic interaction between these mechanisms. Another methodological limitation is the use of commercially available Entresto® tablets rather than a mixture of raw sacubitril/valsartan active pharmaceutical ingredients. While this approach mimics clinical administration, we cannot definitively rule out the possibility that tablet excipients may have influenced the physiological responses in the treated animals. Future mechanistic studies should utilize pure compounds to ensure all observed effects are strictly related to the active mixture. The primary aim of this study was to evaluate the combined effects of sacubitril/valsartan administered as a fixed-dose combination, reflecting its clinical application. However, future studies that include only a valsartan treatment group would yield important mechanistic insights. The findings revealed that VS did not provide neuroprotection, underscoring the significant tissue-specific limitations of neprilysin inhibition and informing the development of future neuroprotective therapies.
In addition to the limitations discussed above, it is important to acknowledge that the cumulative CIS exposure in this acute experimental model was associated with notable mortality, which may limit its suitability for modeling chronic chemotherapy-related cognitive impairment. Accordingly, future studies aiming to investigate long-term chemobrain should consider lower-dose, longer-duration CIS regimens that more closely reflect chronic clinical chemotherapy exposure while minimizing systemic toxicity. Furthermore, NF-κB was assessed using an ELISA-based approach that quantifies total protein levels but does not directly assess nuclear translocation, DNA-binding activity, or transcriptional activation of downstream target genes. Consequently, the NF-κB findings should be interpreted as reflecting changes in the potential for inflammatory signaling rather than definitive pathway activation. Future studies incorporating nuclear fractionation, Western blot analysis, immunolocalization, or gene expression profiling of NF-κB–regulated targets would provide more conclusive mechanistic insight. Similarly, apoptosis was assessed by measuring total caspase-3; evaluating the cleaved (active) form would further strengthen conclusions regarding executioner caspase activation. Another limitation of this study is the absence of neuromodulatory markers such as BDNF and CREB, which are key regulators of hippocampal synaptic plasticity and memory; future studies incorporating these markers would provide additional insight into the neurocognitive consequences of CIS treatment.
In summary, this research establishes that CIS induces significant behavioral impairments, accompanied by marked oxidative stress, neuroinflammation, and alterations in apoptosis-related signaling. Notably, the study reveals that co-administration of sacubitril/valsartan does not alleviate these toxic effects across the assessed parameters. These findings highlight that the neuroprotective potential of neprilysin inhibition differs from its well-recognized cardioprotective benefits. Further investigation using more precise molecular and cellular approaches is warranted to clarify the underlying mechanisms.
Data Access Statement
Data available upon request for reasonable reasons from the corresponding author.
Acknowledgments
The researchers would like to thank the Deanship of Graduate Studies and Scientific Research at Qassim University for financial support (QU-APC-2026).
Funding
There was no funding for the correct work.
Disclosure
The authors declare that they have no conflict of Interest.
References
1. Borchelt DR, Zhou W, Kavelaars A, Heijnen CJ. Metformin prevents cisplatin-induced cognitive impairment and brain damage in mice. PLoS One. 2016;11. doi:10.1371/journal.pone.0151890
2. Ongnok B, Khuanjing T, Chunchai T, et al. Donepezil protects against doxorubicin-induced chemobrain in rats via attenuation of inflammation and oxidative stress without interfering with doxorubicin efficacy. Neurotherapeutics. 2021;18:2107–16. doi:10.1007/s13311-021-01092-9
3. Alhowail AH. Cisplatin induces hippocampal neurotoxicity and cognitive impairment in rats through neuroinflammation, oxidative stress, and overexpression of glutamatergic receptors mRNA. Front Pharmacol. 2025;16. doi:10.3389/fphar.2025.1592511
4. Kim H-G, Rashid MA, Poleschuk M, et al. Cognitive dysfunction in chemobrain: molecular mechanisms and therapeutic implications. Biomed Pharmacother. 2025;192. doi:10.1016/j.biopha.2025.118581
5. Aldubayan MA. Avenanthramide C mitigates cisplatin-induced hippocampal neurotoxicity and cognitive impairment in rats via suppression of neuroinflammation and neuronal apoptosis. Front Pharmacol. 2025;16. doi:10.3389/fphar.2025.1706224
6. Patai R, Csik B, Nyul-Toth A, et al. Persisting blood–brain barrier disruption following cisplatin treatment in a mouse model of chemotherapy-associated cognitive impairment. GeroScience. 2025;47:3835–3847. doi:10.1007/s11357-025-01569-x
7. Alotayk LI, Aldubayan MA, Alenezi SK, Anwar MJ, Alhowail AH. Comparative evaluation of doxorubicin, cyclophosphamide, 5-fluorouracil, and cisplatin on cognitive dysfunction in rats: delineating the role of inflammation of hippocampal neurons and hypothyroidism. Biomed Pharmacother. 2023;165:115245. doi:10.1016/j.biopha.2023.115245
8. Dargad RR, Prajapati MR, Dargad RR, Parekh JD. Sacubitril/valsartan: a novel angiotensin receptor-neprilysin inhibitor. Indian Heart J. 2018;70:S102–S110. doi:10.1016/j.ihj.2018.01.002
9. Rajzer P, Biegus J. Sacubitril/valsartan in a wide spectrum of heart failure patients (from mechanisms of action to outcomes in specific populations). Heart Fail Rev. 2025;30:387–405. doi:10.1007/s10741-024-10471-1
10. Poorgolizadeh E, Homayouni Moghadam F, Dormiani K, Rezaei N, Nasr-Esfahani MH. Do neprilysin inhibitors walk the line? Heart ameliorative but brain threatening! Eur J Pharmacol. 2021;894:173851. doi:10.1016/j.ejphar.2021.173851
11. Bai W, Huo T, Chen X, et al. Sacubitril/valsartan inhibits ox‑LDL‑induced MALAT1 expression, inflammation and apoptosis by suppressing the TLR4/NF‑κB signaling pathway in HUVECs. Mol Med Rep. 2021;23(6). doi:10.3892/mmr.2021.12041
12. Yu C, Li D, Li Z, Yu D, Zhai G. Effect of sacubitril/valsartan on inflammation and oxidative stress in doxorubicin-induced heart failure model in rabbits. Acta Pharm. 2021;71(3):473–484. doi:10.2478/acph-2021-0030
13. Trivedi RK, Polhemus DJ, Li Z, et al. Combined angiotensin receptor–neprilysin inhibitors improve cardiac and vascular function via increased NO bioavailability in heart failure. J Am Heart Assoc. 2018;7(5). doi:10.1161/jaha.117.008268
14. Alqahtani F, Mohany M, Alasmari AF, et al. Angiotensin II receptor neprilysin inhibitor (LCZ696) compared to valsartan attenuates hepatotoxicity in STZ-induced hyperglycemic rats. Int J Med Sci. 2020;17(18):3098–3106. doi:10.7150/ijms.49373
15. Shi H, Lu H, Zheng Y, et al. Bioinformatics and experimental studies jointly reveal that Sacubitril Valsartan improves myocardial oxidative stress and inflammation by regulating the MAPK signaling pathway to treat chemotherapy related cardiotoxicity. Biochem Biophys Res Commun. 2024;690:149244. doi:10.1016/j.bbrc.2023.149244
16. Zhang M, Zou Y, Li Y, Wang H, Sun W, Liu B. The history and mystery of sacubitril/valsartan: from clinical trial to the real world. Front Cardiovascul Med. 2023;10. doi:10.3389/fcvm.2023.1102521
17. Alresheedi MM, Aldubayan MA. Sacubitril/valsartan fails to prevent doxorubicin-induced cognitive impairment and hippocampal oxidative, inflammatory, and apoptotic alterations in rats. Front Pharmacol. 2026;16. doi:10.3389/fphar.2025.1732842
18. Alaqeel N. Protective role of apigenin nanoparticles in cisplatin induced cardiotoxicity in rats. Nutr Food Sci. 2024;55:1–15. doi:10.1108/nfs-03-2024-0095
19. Alharbi M, Aldubayan M. Ameliorative effect of empagliflozin and linagliptin on cisplatin-induced nephrotoxicity and cardiotoxicity by reducing oxidative stress. Drug Des Devel Ther. 2025;19:11483–11497. doi:10.2147/dddt.S561268
20. Burke RM, Lighthouse JK, Mickelsen DM, Small EM. Sacubitril/Valsartan decreases cardiac fibrosis in left ventricle pressure overload by restoring PKG signaling in cardiac fibroblasts. Circulation. 2019;12. doi:10.1161/circheartfailure.118.005565
21. Alsaud MM, Alhowail AH, Aldubayan MA, Almami IS. The ameliorative effect of pioglitazone against neuroinflammation caused by doxorubicin in rats. Molecules. 2023;28:4775. doi:10.3390/molecules28124775
22. Alsikhan RS, Aldubayan MA, Almami IS, Alhowail AH. Protective effect of galantamine against doxorubicin-induced neurotoxicity. Brain Sci. 2023;13(6):971. doi:10.3390/brainsci13060971
23. Cicek B, Danisman B, Bolat I, et al. Effect of tangeretin on cisplatin-induced oxido-inflammatory brain damage in rats. J Cell Mol Med. 2024;28. doi:10.1111/jcmm.18565
24. Aldubayan MA. Protective effects of avenanthramide-C against cisplatin-induced cardiotoxicity in rats by attenuating oxidative stress, inflammatory cytokines, and modulating p62–Keap1–Nrf2 pathway. Front Pharmacol. 2025;16. doi:10.3389/fphar.2025.1694060
25. Alhowail AH, Aldubayan MA. Doxorubicin impairs cognitive function by upregulating AMPAR and NMDAR subunit expression and increasing neuroinflammation, oxidative stress, and apoptosis in the brain. Front Pharmacol. 2023;14:1251917. doi:10.3389/fphar.2023.1251917
26. Mostafa F, Mantawy EM, Said RS, Azab SS, El-Demerdash E. Captopril attenuates oxidative stress and neuroinflammation implicated in cisplatin-induced cognitive deficits in rats. Psychopharmacology. 2025;242:563–578. doi:10.1007/s00213-024-06706-6
27. Redza-Dutordoir M, Averill-Bates DA. Activation of apoptosis signalling pathways by reactive oxygen species. BBA. 2016;1863:2977–2992. doi:10.1016/j.bbamcr.2016.09.012
28. Dho SH, Cho M, Woo W, Jeong S, Kim LK. Caspases as master regulators of programmed cell death: apoptosis, pyroptosis and beyond. Exp Mol Med. 2025;57:1121–1132. doi:10.1038/s12276-025-01470-9
29. Wu D, Wang D, Tan -S-S, et al. Synergistic effects of lead and copper co-exposure on promoting oxidative stress and apoptosis in the neuronal cells. Toxicology. 2025;513. doi:10.1016/j.tox.2025.154103
30. Lawrence KA, Gloger EM, Pinheiro CN, Schmitt FA, Segerstrom SC. Associations between IL-1β, IL-6, and TNFα polymorphisms and longitudinal trajectories of cognitive function in non-demented older adults. Brain Behav Immun Health. 2024;39:100816. doi:10.1016/j.bbih.2024.100816
31. Du J, Zhang A, Li J, et al. Doxorubicin-Induced cognitive impairment: the mechanistic insights. Front Oncol. 2021;11. doi:10.3389/fonc.2021.673340
32. Hu F, Yan S, Lin L, Qiu X, Lin X, Wang W. Sacubitril/valsartan attenuated myocardial inflammation, fibrosis, apoptosis and promoted autophagy in doxorubicin-induced cardiotoxicity mice via regulating the AMPKα–mTORC1 signaling pathway. Mol Cell Biochem. 2024;480:1891–1908. doi:10.1007/s11010-024-05117-7
33. Pascual-Figal D, Bayés-Genis A, Beltrán-Troncoso P, et al. Sacubitril-Valsartan, clinical benefits and related mechanisms of action in heart failure with reduced ejection fraction. a review. Front Cardiovascul Med. 2021;8. doi:10.3389/fcvm.2021.754499
34. Zhang H, Huang T, Shen W, et al. Efficacy and safety of sacubitril-valsartan in heart failure: a meta-analysis of randomized controlled trials. ESC Heart Failure. 2020;7:3841–3850. doi:10.1002/ehf2.12974
35. Boyette-Davis JA, Hou S, Abdi S, Dougherty PM. An updated understanding of the mechanisms involved in cHemotherapy-Induced neuropathy. Pain Manage. 2018;8:363–375. doi:10.2217/pmt-2018-0020
36. Evbayekha E, Idowu AB, LaRue S. Sacubitril/Valsartan vs ACE inhibitors or ARBs. JACC. 2025;4. doi:10.1016/j.jacadv.2025.101598
37. Iwata N, Tsubuki S, Takaki Y, et al. Metabolic regulation of brain aβ by neprilysin. Science. 2001;292:1550–1552. doi:10.1126/science.1059946
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Platelet-Activating Factor Promotes the Development of Non-Alcoholic Fatty Liver Disease
Yin H, Shi A, Wu J
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2003-2030
Published Date: 8 July 2022
The Effects and Pathogenesis of PM2.5 and Its Components on Chronic Obstructive Pulmonary Disease

Wang Q, Liu S
International Journal of Chronic Obstructive Pulmonary Disease 2023, 18:493-506
Published Date: 6 April 2023
Quercetin: A Flavonoid with Potential for Treating Acute Lung Injury

Huang M, Liu X, Ren Y, Huang Q, Shi Y, Yuan P, Chen M
Drug Design, Development and Therapy 2024, 18:5709-5728
Published Date: 6 December 2024
Exercise Prescription Training in Chronic Obstructive Pulmonary Disease: Benefits and Mechanisms
Liu S, Yang A, Yu Y, Xu B, Yu G, Wang H
International Journal of Chronic Obstructive Pulmonary Disease 2025, 20:1071-1082
Published Date: 15 April 2025
Rhein Alleviates Cisplatin-Induced Acute Kidney Injury via Downregulation of NOX4-COX2/PGFS Signaling Pathway

Yuan X, Long L, Wang M, Chen W, Liang B, Xu L, Wang W, Li C
Drug Design, Development and Therapy 2025, 19:4641-4664
Published Date: 31 May 2025