Back to Journals » OncoTargets and Therapy » Volume 19
Research Progress on the Molecular Mechanism of Metformin Regulating AMPK Signaling Pathway in Inhibiting Liver Cancer
Authors Zhang HB, Yan Y, Pang JP, Kong MT, Zhu QK
Received 1 November 2025
Accepted for publication 4 February 2026
Published 26 February 2026 Volume 2026:19 572872
DOI https://doi.org/10.2147/OTT.S572872
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sanjay Singh
Hong Bin Zhang, Yan Yan, Jin Peng Pang, Meng Ting Kong, Qian kun Zhu
Interventional Radiology Department, Affiliated Fourth Hospital, Harbin Medical University, Harbin, Heilongjiang, People’s Republic of China
Correspondence: Yan Yan, Interventional Radiology Department, Affiliated Fourth Hospital, Harbin Medical University, Harbin, Heilongjiang, People’s Republic of China, Email [email protected]
Abstract: Liver cancer is one of the common malignant tumors worldwide and ranks among the top in cancer mortality statistics. In recent years, metformin, as a first-line drug with significant efficacy and low cost, has been widely used in clinical practice for the treatment of diabetes. Now, due to its potential inhibitory effect on cancer cells, it has received extensive attention. In view of this, a large number of studies have focused on the mechanisms by which metformin affects Liver carcinoma, with the AMPK pathway being particularly favored. This is because the AMPK pathway plays a significant role in autophagy, cell cycle arrest, inhibition of tumor angiogenesis, and suppression of epithelial-mesenchymal transition. To this end, we reviewed relevant research literature and systematically explored the mechanism of metformin’s treatment of liver cancer through the AMPK pathway and its potential clinical applications. These studies provide an important basis for understanding the potential role of metformin in the treatment of liver cancer and provide important reference for future research on its new mechanisms and its clinical application.
Keywords: liver cancer, AMPK, cell cycle arrest, drug synergy, autophagy
Introduction
Despite the widely reported antitumor effects of metformin, its specific molecular mechanisms in liver cancer are complex and not yet fully elucidated. However, liver cancer is a common malignant tumor worldwide, including hepatocellular carcinoma, mixed cell carcinoma, and intrahepatic cholangiocarcinoma.1–3 It is the sixth most common cancer globally and ranks third in cancer-related human deaths.4,5 In China, the incidence of liver cancer is high, making it the third leading cause of cancer-related death.6 The main triggering factors for liver cancer include: 1) biological factors: hepatitisB, hepatitisC, liver cirrhosis, aflatoxin, etc; 2) non-biological factors: alcohol, obesity, diabetes, hyperlipidemia, metabolic syndrome, etc.7
Diabetes is one of the most concerning factors among the causes of liver cancer mentioned above. Diabetes is an endocrine disease characterized by reduced insulin production, insulin resistance, or both. In recent decades, the incidence of type 2 diabetes has increased year by year and has become an epidemic worldwide.8–10 Studies in different populations have found a significant correlation between diabetes and the incidence of liver cancer.11
Nowadays, there are various treatment options for liver cancer.12 Although short-term prognosis can be improved, the long-term prognosis of liver cancer remains bleak, making new treatment methods an important focus of clinical research.13,14 Metformin is an oral biguanide drug that has become a common medication for type 2 diabetes due to its significant efficacy and relatively few side effects (only gastrointestinal adverse reactions and rare lactic acidosis).15–20 The mechanism of metformin in lowering blood sugar mainly involves reducing the production of liver glycogen, as well as reducing the absorption of glucose by the small intestine or reducing insulin resistance.15,21–23 Many published studies have found that metformin can be used to treat or assist in the treatment of various cancers, such as: prostate cancer,24 breast cancer,25,26 colorectal cancer,27 gastric cancer,28 esophageal cancer,29 glioma,30 pancreatic cancer,31 and liver cancer.32,33 The latest retrospective studies also indicate the inhibitory effect of metformin on liver cancer.34,35
There are many pathways involved in the mechanism of metformin in the treatment of liver cancer. Among them, AMPK-related pathways have attracted special attention. It has been reported that the antitumor effect of metformin is mainly achieved by AMP activated protein kinase (AMPK).36,37 Recent related studies have found that through the AMPK-mTOR pathway, autophagy of liver cancer cells can be induced and lipid accumulation in liver cancer cells can be reduced. Additionally,38,39 the AMPK-P53 pathway can reduce epithelial-mesenchymal transition, thereby lowering liver cancer migration and invasion.40 In models simulating glucose deficiency, the AMPK pathway can increase the lifespan of mice with liver cancer, allowing them to live to normal mature age.41 It can promote oxidative phosphorylation through the AMPK-HIF1α axis, thereby disrupting glycolysis, not only inhibiting tumor cell growth but also causing DNA damage and apoptosis in hepatocellular carcinoma cells.42 Given this, this article aims to systematically review how the AMPK pathway, as a central hub, integrates various downstream effects, and to focus on the challenges and opportunities in its clinical translation.
Mechanism of Metformin Acting on Liver Cancer Through AMPK
Metformin and AMPK
AMPK is an AMP-activated protein kinase and is a highly conserved trimer complex,43 including an α subunit, a β subunit, and a γ subunit. Each subunit is encoded by 2–3 different genes,44 the α subunit has catalytic activity, and the β and γ subunits help maintain protein structure.45,46 Metformin activates AMPK by affecting mitochondrial respiratory complex I and AMP/ATP ratio, as well as upstream phosphorylation of Thr-172 of the catalytic α subunit.47–50 Phosphorylation of Thr-172 in the α subunit is achieved through the activation of LKB1 (serine/threonine kinase).51 It has also been reported in the literature that metformin can indirectly activate AMPK by inhibiting PKA (Protein Kinase A).52
AMPK acts as an energy sensor in eukaryotes43 and plays an important role in the occurrence and development of metabolism and cancer.53 In terms of energy and metabolism, AMPK can regulate the energy metabolism balance of cells, promote energy production, and limit energy use to maintain cell survival.45,46 Its main functions include: enhancing the oxidation of fatty acids and glucose, inhibiting protein biosynthesis, thus achieving a dynamic balance of ATP levels.54 In terms of cancer treatment, AMPK mediates antitumor effects through various mechanisms,55 and the activation of AMPK may be involved in the tumor cell cycle arrest induced by metformin.56 In addition, AMPK not only participates in inhibiting cancer cell proliferation, but also further enhances its anti-tumor activity by regulating p53 mediated tumor suppression (p53- tumor suppressor gene).57
Among the many AMPK-activated pathways, the mTOR pathway has been the most extensively studied. After AMPK activation, it participates in translation initiation and protein synthesis control by inhibiting the mammalian rapamycin target (mTOR), thereby regulating the growth and proliferation of tumor cells.58 mTOR is a serine/threonine kinase, which is at least two multi-protein complexes, these complexes are called mTOR complexes 1 and 2, which play a key role in cell growth and differentiation.58,59 mTOR is abnormally activated in HCC,60 while metformin can inhibit the mTOR pathway33,61,62 and inhibit the growth and metastasis of liver tumors.63 In addition, a number of clinical and preclinical studies have shown that inhibiting mTOR can be used to treat many types of solid tumors, including esophageal squamous cell carcinoma, lung cancer, renal cell carcinoma and prostate cancer. Therefore, mTOR may be a potential target for HCC treatment.64–67
In short, as a potential metabolic tumor suppressor, AMPK has shown great potential in the prevention and treatment of cancer and has become a promising therapeutic target.68–72 It is worth noting that in the following text, liver cancer refers to hepatocellular carcinoma, and cases of non-hepatocellular carcinoma are specially noted.
Autophagy
Autophagy refers to the process by which the cytoplasm is dissolved by lysosomes. Autophagy exists in three primary forms: microautophagy, macroautophagy, and chaperone-mediated autophagy (CMA).73 At present, there are two views on autophagy of tumor cells: 1) It is believed that autophagy can increase the resistance of tumors and help remove necrotic cells, thereby promoting the progress of tumors;74,75 2) It is believed that autophagy is beneficial to cells. It can be used as a new way of treating tumors.76,77 Studies have shown that autophagy can promote the apoptosis of cervical cancer,78 breast cancer,79 leukemia,80 melanoma and lymphoma.81,82 Although the role of autophagy remains controversial, studies have shown that autophagy plays a crucial role in liver diseases.83 It is well known that metformin is an activator of AMPK and can also activate cellular autophagy.81,84,85 Literature reports indicate that metformin activates AMPK to promote autophagy in pancreatic β-cells,86 cerebral arteries,87 breast cancer cells,88 colorectal cancer cells,89 and endometrial cancer cells.90 Similarly, a large amount of literature confirms that metformin can also mediate autophagy in liver cancer.
Autophagy is a complex network woven by many factors such as TFEB,91 E2F1,92 FoxO1,93 ATF4,94 CEBPβ,95 CHOP,96 etc.97 Among them, CEBPD is considered to be a tumor suppressor because its overexpression can cause cancer cells to die.98 Metformin can induce increased expression of CEBPD through the AMPK pathway, thereby promoting tumor autophagy.97 Src is a non-receptor tyrosine kinase that is highly expressed in a variety of tumors and is believed to play an important role in the occurrence and development of tumors.99 Metformin can maintain the stability of CEBPD by inhibiting the expression of Src.97 In addition, the viability of MHCC97H hepatocellular carcinoma cells significantly decreased after treatment with metformin (10mM), and the effect became more pronounced over time. Western blot analysis showed increased levels of autophagy-related protein LC3-II (a marker of autophagy initiation) and decreased levels of p62, which is caused by reduced phosphorylation of mTOR and p70 S6 kinase under AMPK activation.97,100,101 Metformin can also be combined with sorafenib to increase the expression of microtubule-associated protein light chain 3 (LC3), leading to autophagy.100 However, a few studies suggest that metformin inhibits autophagy. The article found that metformin (0.016–2 mM) treatment of serum-starved H4IIE cells dose-dependently reduced the expression of six autophagy-related proteins (Atg3, Atg5, Atg7, Atg12, LC3B, beclin-1), thereby inhibiting autophagy, and this effect could be blocked by an AMPK inhibitor. This phenomenon only occurs under glucose deprivation conditions (in glucose-free dishes), but not in dishes lacking only amino acids (with 5.5 mM glucose).102 We believe this is related to the nutritional status of the cells, and the study may have been limited to macroautophagy while neglecting chaperone-mediated autophagy(CMA). The mechanism of metformin-mediated autophagy is shown in Figure 1.
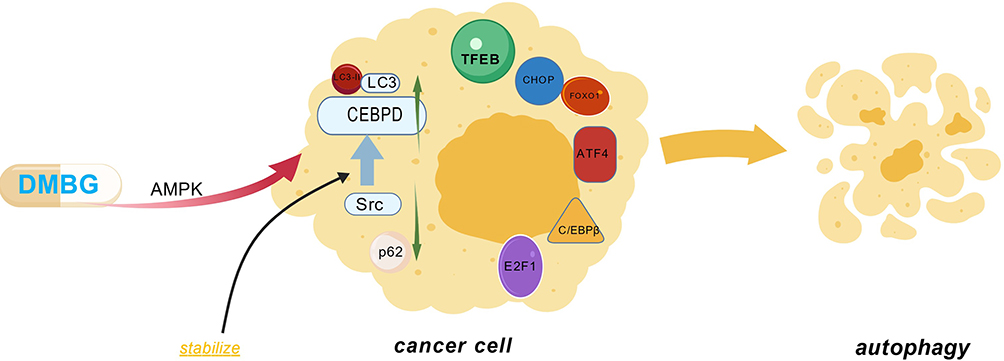 |
Figure 1 Mechanisms by which metformin induces autophagy through the AMPK pathway. On one hand: metformin can promote tumor cell autophagy by inducing CEBPD and maintain the stability of CEBPD by inhibiting Src; on the other hand: metformin can cause autophagy by reducing p70S6 and increasing LC3-II and LC3. Figure Created with BioGDP.com. Abbreviations: DMBG, metformin; AMPK, AMP-activated protein kinase; CEBPD, transcription factor (CCAAT) enhancer-binding protein delta; Src, non-receptor tyrosine kinase; LC3, microtubule-associated protein light chain 3; LC3-II, a form of LC3; p62, selective autophagy receptor; TFEB, transcription factor EB; FoxO1, transcription factor Forkhead family member; E2F1, transcription factor E2F family member; ATF4, activating transcription factor 4; CEBPβ, transcription factor (CCAAT) enhancer-binding protein beta; CHOP, C/EBP homologous protein. |
Recent studies have confirmed this view, showing that the dual role of autophagy is related to the cellular environment and associated regulatory pathways. It acts as a tumor suppressor in the early stages of cancer cells but promotes cell survival and progression in later stages.103 Under conditions of nutrient deprivation, AMPK activates ULK1, while the inhibition of mTORC1 relieves its suppression of ULK1, ultimately leading to the phosphorylation of downstream autophagy-related proteins and the initiation of autophagy.104 As liver cancer progresses, accumulated stress and metabolic pressure lead to the upregulation of chaperone-mediated autophagy(CMA) as a compensatory mechanism. At this stage, macroautophagy activity decreases, activating NRF2 and promoting mTOR signaling pathways, which drive tumor cell proliferation and growth.105
Cell Cycle Arrest
Blocking the cell cycle can promote apoptosis of tumor cells. Studies have shown that one of the effects of metformin treatment for liver cancer lies in cell cycle arrest.33 Subsequent more in-depth research has indicated that metformin can cause liver cancer cells to arrest at the G0/G1 phase of the cell cycle.79 CAI et al believe that the transition from G1 to S phase of the cell cycle is regulated by cyclins and cyclin-dependent kinase inhibitors (CDKIs), including cyclin D1, cyclin E, p21CIP, and p27KIP. Cyclin D1 and cyclin E promote cell DNA synthesis and cell growth, and the overexpression of cyclin D1 and cyclin E promotes cancer progression.106,107 Conversely, the downregulation of cyclin D1 and cyclin E expression limits the progression of the cell cycle to the G0/G1 phase and inhibits tumor cell proliferation.56,108 Metformin can reduce the levels of cyclin D1 and cyclin E through the AMPK pathway, thereby blocking the cell cycle and inhibiting the growth of tumor cells.32,56,79,109 Chen et al found that metformin inhibits the proliferation of liver cells through AMPK and its upstream kinase LKB1, thereby causing cell cycle arrest at the G0/G1 phase and upregulating P21clp1 and P27kip1 in a dose-dependent manner, while downregulating cyclin D1.32 Another study has shown that metformin reduces BrdU (Thymidine analogues) incorporation in a AMPK-dependent manner, leading to cell cycle arrest at G0/G1, with the possible mechanism being the reduction of DNA production or induction of cell cycle arrest or apoptosis.71 The Department of Medicine at Japan’s Kagawa University confirmed that metformin induces cell cycle arrest both in vitro and in vivo. After treating Huh7 cells with 10 mM metformin for 24 to 72 hours, it was found that the expression of cell cycle regulators (cyclin D1, Cdk4, and cyclin E) decreased, and Huh7 cells accumulated in the G0/G1 phase with a reduction in the proportion of cells in the S phase and G2/M phase. Subsequently, 30 nude mice were subcutaneously inoculated with 5×106 Huh7 cells, and after the transplanted tumors reached a maximum diameter of over 6 mm, the mice were randomly divided into three groups and treated with metformin at doses of 1 mg per mouse, 2 mg per mouse, and phosphate-buffered saline (PBS), respectively. The results still showed a decrease in the expression of cell cycle regulators. This confirmed that metformin can arrest tumor cells in the G0/G1 phase.110 The molecular mechanisms of metformin exerting cell cycle arrest and EMT are shown in Figure 2.
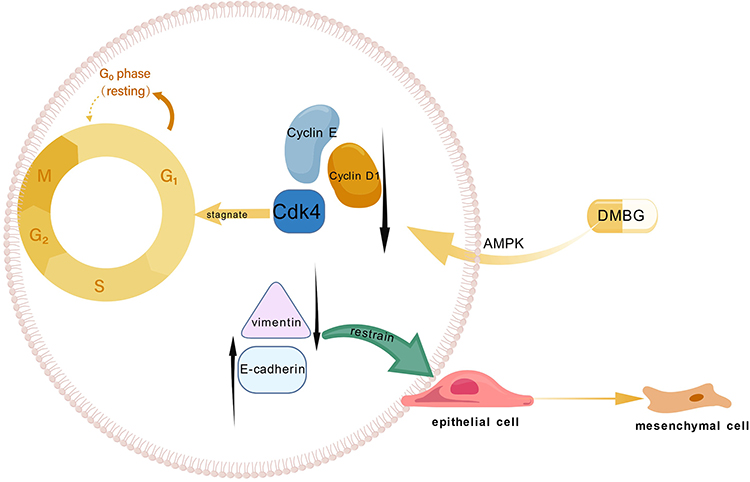 |
Figure 2 The mechanism by which metformin acts on the AMPK pathway leading to cell cycle arrest and EMT. Figure Created with BioGDP.com. Abbreviations: DMBG, metformin; AMPK, AMP-activated protein kinase; Cyclin D1, cell cycle protein D1; Cyclin E, cell cycle protein E; Cdk4, cyclin-dependent kinase (a cell cycle regulator); E-cadherin, one of the markers of epithelial-mesenchymal transition; vimentin, intermediate filament protein (one of the markers of epithelial-mesenchymal transition); p21CIP and p27KIP, cell cycle regulators. |
EMT
Epithelial-interstitial transformation (EMT) can cause significant changes in the morphology and phenotypes of epithelial cells, including cytoskeletal reorganization and changes in cell polarity, which increase the aggressiveness of cells.111 It is an ongoing problem in the study of liver cancer. Therefore, EMT-Regulation has become one of the key targets for controlling the development of HCC.112 Metformin activation of AMPK can not only increase cell differentiation, but also increase E-cadherin and reduce the expression of vimentin in HCC cells to inhibit EMT. These findings show that metformin partially inhibits the metastasis and invasion of HCC cells by reversing EMT.113–116
Similarly, in intrahepatic cholangiocarcinoma (including large duct type and small duct type), metformin can prevent EMT by activating the AMPK-FoxO3 pathway, increasing CK19 and E-Cadherin, and decreasing Vimentin, SNAIL1/2, and TWIST1 (EMT mesenchymal markers). In experiments, metformin at concentrations of 10–100 μM was applied to intrahepatic cholangiocarcinoma cells (derived from surgical specimens) for 48–96 hours, and its inhibitory effect was observed, showing a positive correlation with time and dose. When metformin-treated (metformin pretreatment for 57 days at 10 μM) and untreated intrahepatic cholangiocarcinoma cells were injected into mice, tumor growth was observed in the control group after 10 weeks, while no tumor growth was seen in the experimental group, thereby validating metformin’s inhibition of EMT both in vitro and in vivo.117 However, research regarding metformin’s inhibition of EMT remains limited and requires further verification.
Glucose Metabolism
The antidiabetic effect of metformin depends on the activation of AMPK, which lowers blood sugar levels by reducing liver gluconeogenesis and increasing glucose uptake by skeletal muscles.46,118 It is worth noting that metformin leads to a decrease in the expression of the glucose transporter Glut1, which can also lead to a decrease in glucose uptake to lower blood sugar, and this is also essential to regulate the fate of CD4+T cells. Metformin can also cause reduced differentiation of T cells through the AMPK-mTOR pathway,119 therefore, the potential connection between AMPK and Glut1 deserves our attention. As mentioned above, metformin can reverse EMT, thereby partially inhibiting the metastasis and invasion of HCC cells.113–116 The study found that glucose starvation combined with metformin can increase metformin’s ability to reverse EMT, antiproliferation, and migration.120
The relationship between AMPK and reactive oxygen species (ROS) is complicated. Although studies have shown that metformin can inhibit the accumulation of reactive ROS in an AMPK-dependent manner, studies have also reported that metformin can promote the accumulation of cytotoxic reactive oxygen species in a non AMPK-dependent manner.121–123 This may be determined by specific signaling networks under different conditions, and it aligns with glucose’s dual regulation of ROS—both glucose deficiency and high glucose concentration can induce intracellular ROS production through different signaling pathways that affect cellular metabolism and gene regulation. It has been shown that metformin reduces ROS production induced by high glucose, while simultaneously triggering and exacerbating ROS generation caused by low glucose.124
Studies on metformin’s effect on glucose metabolism via AMPK are limited, and research has found that the lysosomal pathway can activate AMPK under low glucose conditions.41,125 Since metformin can lower blood sugar and create a low-glucose environment, this suggests a direction for our future research.
Drug Combination
The treatment methods for liver cancer are diverse, with surgical resection being the preferred treatment for liver cancer.126 In recent years, immunotherapy and targeted therapy have also played a crucial role in the treatment of advanced liver cancer. However, the existing treatments still face challenges such as drug resistance and the recurrence and metastasis of liver cancer.127–129 Therefore, there is an urgent need for new drugs or combination therapies to overcome the existing limitations.
Sorafenib
Sorafenib is a first-line oral multitargeted kinase inhibitor for the treatment of advanced hepatocellular carcinoma, with evidence showing that it can block the Raf/MEK/ERK signaling pathway to inhibit tumor cell proliferation and also target tyrosine kinase receptors such as VEGFR-2 or PDGFR to produce anti-angiogenic effects.130–133 However, long-term use of sorafenib can lead to sorafenib resistance, resulting in a decline in treatment efficacy and a lack of significant improvement in patient overall survival.130,134 Therefore, there is an urgent need for new drugs to replace sorafenib or to be used in combination to counteract its limitations. Studies have shown that sorafenib can induce a shift from HIF-1α to HIF-2α, leading to an increase in HIF-2α, which is one of the reasons for sorafenib resistance.135,136 HIF-2α can also lead to a decrease in TIP30, where TIP30, as a tumor suppressor factor,137 is significantly associated with the invasion and metastasis of hepatocellular carcinoma. It not only regulates cell proliferation and apoptosis genes but also participates in cellular glucose tolerance, cancer metastasis, and invasion.137–141 You et al found that combining metformin with sorafenib can inhibit the induction of HIF-2α by sorafenib, thereby reducing resistance to sorafenib and reducing cancer invasion and metastasis.142 Guo et al also found that downregulating TIP30 can increase thioredoxin (TXN) expression, and increased TXN expression can accelerate tumor metastasis and increase tumor invasiveness.143,144 Combining metformin with sorafenib can activate the AMPK pathway, leading to upregulation of TIP30 and downregulation of TXN.145 Additionally, Ling et al believe that combining metformin with sorafenib can inhibit the activation of mTORC2 by metformin and increase the inhibition of mTORC1 by metformin, achieving synergistic effects. Compared to monotherapy, the combination not only blocks the cell cycle at the G0/G1 phase but also reduces key factors such as CyclinD1, CDK4, and Ki-67, and induces autophagy, significantly promoting the antiproliferative ability of hepatocellular carcinoma cells.100 This process is achieved through the AMPK/mTORC1/2 pathway. In summary, the combination of metformin and sorafenib exerts synergistic effects through multiple pathways, effectively reducing HIF-2α levels, upregulating TIP30, and downregulating TXN, inducing cell cycle arrest and autophagy mechanisms, thus significantly inhibiting the progression and metastasis of hepatocellular carcinoma. Mechanism of metformin combined with sorafenib is shown in Figure 3.
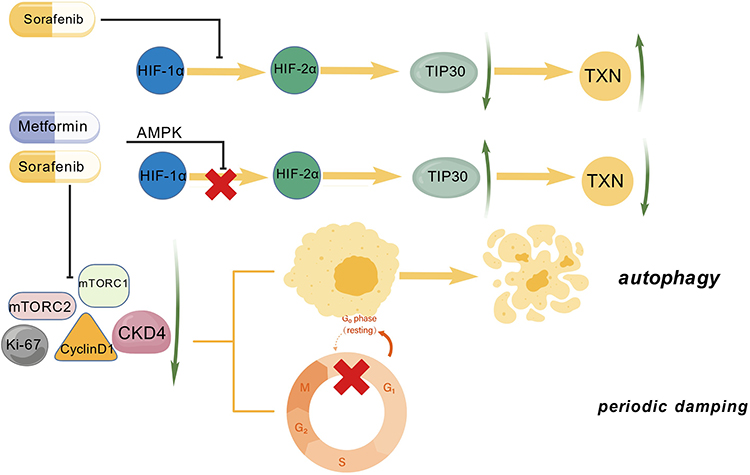 |
Figure 3 The mechanism of combined action of metformin and sorafenib on the AMPK pathway for hepatocellular carcinoma treatment. Figure Created with BioGDP.com. Abbreviations: AMPK, AMP-activated protein kinase; HIF-1α, one of the hypoxia-inducible factors; HIF-2α, one of the hypoxia-inducible factors; TIP30, tumor suppressor factor; TXN, thioredoxin; Cdk4, cyclin-dependent kinase (a cell cycle regulator); Cyclin D1, cell cycle protein D1; Ki-67, cell proliferation marker; mTORC1, one of the rapamycin target protein complexes; mTORC2, another of the rapamycin target protein complexes. |
5-Fu
5-Fu as an adenine synthase inhibitor has been approved as a chemotherapeutic drug for hepatocellular carcinoma and is widely used clinically.146 HIF-1 is a basic transcription factor that regulates the transcription of various target genes in hypoxic responses.147 HIF-1α is an oxygen-regulated subunit that mediates the basic function of HIF-1. Previous studies have shown that overexpression of HIF-1α may be involved in the pathogenesis of tumor chemoresistance,148 and HIF-1α can increase the expression of multidrug resistance-related proteins P-gp and MRP1, thereby playing an important role in drug multidrug resistance.149–151 Studies have found that metformin can reduce the expression of HIF-1α through the p-AMPK/mTOR pathway,152,153 thereby reducing the expression of P-gp and MRP1, and consequently reducing hepatocellular carcinoma resistance to 5-Fu.154
Other Hypoglycemic Drugs
Metformin and thiazolidinediones are currently commonly used oral hypoglycemic drugs.155–157 Rosiglitazone (Ros) is a representative of thiazolidinediones, which can improve insulin resistance and stimulate insulin secretion. Metformin is also a representative drug of biguanides. However, they have different side effects. Metformin can cause minor gastrointestinal adverse reactions and rare lactic acidosis, while rosiglitazone increases the risk of cardiovascular and fracture events.158 Therefore, these two drugs are often used in combination in clinical practice to improve efficacy and reduce side effects.156,159 Both drugs have been found to have anti-tumor effects. Metformin, as mentioned above. Rosiglitazone has been found to have anti-tumor capabilities in patients with lung cancer, prostate cancer, and colorectal cancer.160 Therefore, some studies have found that a 1:1 mixture of the two drugs can inhibit hepatocellular carcinoma proliferation through the AMPK/p21 pathway with no obvious side effects.161
The latest research has found that empagliflozin, as an antidiabetic drug, works by inhibiting the sodium-glucose cotransporter-2 (SGLT-2) in the proximal tubules of the kidney.162 When used in combination with metformin, the two drugs complement each other to reduce the expression of liver injury indicators such as ALT, AST, and AFP, as well as tumor-related indicators such as ERK1/2, VEGF, and Ki67. This combination therapy also upregulates the ratio of p-AMPKα1/AMPKα1 (a subunit of AMPK) and Bax/Bcl-2, thereby inhibiting the progression of liver cancer. The mechanism of this combination treatment lies in the inactivation of the NF-κB (nuclear transcription factor) pathway via the AMPK pathway.163 The mechanism of metformin combined with other drugs for liver cancer treatment is shown Figure 4.
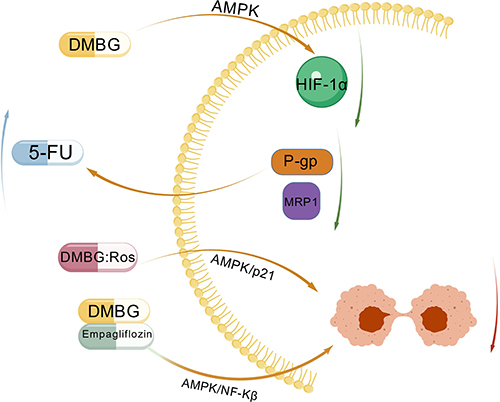 |
Figure 4 Mechanisms of Combined Treatment with Metformin and Other Drugs for Hepat-ocellular Carcinoma. Figure Created with BioGDP.com. Abbreviations: DMBG, metformin; 5-Fu, fluorouracil; Ros, rosiglitazone; A-MPK, AMP-activated protein kinase; HIF-1α, one of the hypoxia-inducible factors; MRP1 and P-gp, multidrug resistance-associated protein; P21, cyclin-dependent kinase inhibitor; NF-κB, nu-clear factor-kappa B. |
Multiple studies have confirmed that metformin is a known antidiabetic drug with good antiproliferative activity, which can be used alone or in combination with other antiproliferative drugs to control tumors and is applicable to various cell types.164–166 The mechanism of metformin combined drug therapy for liver cancer is shown in Table 1.
 |
Table 1 Mechanism of Metformin in Combination with Other Drugs in the Treatment of Liver Cancer |
Combination with Cytokines
IL
Interleukin has a variety of important functions in the human body, including positive effects such as improving the body’s immunity, but it also has side effects such as causing inflammation.167 In the study by Zhao et al changes in interleukin in the plasma of 137 patients were monitored. The results showed that IL-6 and IL-8 were elevated in patients with liver cancer and were associated with liver cancer.168 Among them, IL-8 as a chemokine in the human body has been shown to regulate the self-renewal and angiogenesis of tumor promoter cells.169,170 In addition, IL-8 is positively correlated with the expression of PDL1 and PDL2. PD-L1 is a major immune checkpoint molecule that has a negative regulatory effect on T cell activity. High expression in a variety of cancers is associated with poor prognosis.171,172 Therefore, IL-8 can be used as a checkpoint for immunosuppression.168 The study by Chen et al found that metformin can reduce the expression of IL-8 through the p-AMPK/p-JNK pathway,168 thereby achieving the purpose of controlling tumor growth. As a member of the endopeptidase protein family, MMP9 has been found to promote EMT, resulting in increased aggressiveness and metastasis of HCC cells.173 In the latest research, metformin can reduce IL-8-mediated MMP9 elevation through the AMPK/JNK/MMP9 pathway, thereby reducing the aggressiveness of liver cancer.168
IL-22 is considered an inducible factor derived from T cells, mainly produced by Th1 and Th17 cells.174 Its primary physiological functions include wound healing and innate antibacterial responses, while its receptor is mainly expressed by non-hematopoietic cells such as lung, gastrointestinal epithelial cells, and keratinocytes.175 Pan et al found that IL-22 can induce the expression of anti-apoptotic and mitotic genes in hepatocytes.176 Additionally, the presence of IL-22 allows damaged hepatocytes to survive, and these damaged hepatocytes are often precursors to HCC, thus participating to some extent in the occurrence and progression of cancer.177 Finally, the expression of IL-22 in HCC tissue is higher in stages III–IV than in stages I–II patients, suggesting that IL-22 may be associated with enhanced proliferative capacity and increased malignancy of tumors.178 Zhao et al further found that metformin inhibits the phosphorylation of STAT3/STAT4 through the p-AMPK/p-mTOR pathway, thereby blocking the derivation of Th1 and Th17 cells into IL-22, ultimately reducing the phosphorylation level of STAT3, Bcl-2 expression, and CyclinD1 expression.119
miRNA
miRNAs is a small non-coding RNA molecule that contains about 22–25 nucleotides. They are widely present in all kinds of organisms. Its main functional mechanism lies in regulating the transcriptional activity of the target gene, thereby achieving gene silencing or activation, and playing a key regulatory role in various cell behaviors, functions and diseases.179–181 Studies have found that high expression levels of miRNA-122 can enhance the sensitivity of liver cells to metformin. On the contrary, low expression can cause liver cells to be insensitive to metformin. The reason is that when miRNA is low, the AMPK pathway can be activated in advance, thereby inhibiting the activity of metformin.182 Other studies have shown that the expression level of miRNA-23a can be significantly induced after metformin treatment. When the expression of miRNA-23a is suppressed, the apoptosis process of liver malignancies induced by metformin can be eliminated.183 Through the AMPK/P53/miRNA-23a pathway, metformin can regulate the expression of miRNA-23a, thereby regulating the cell cycle regulation, apoptosis, EMT and other key processes involved in miRNA.184,185
P53
In addition to inducing the expression of miRNA-23a, P53 also plays an important role in the occurrence and metastasis of cancer. Studies have shown that when P53 is lacking, the inhibitory effect of metformin on liver cancer cells is weakened.120 In fact, metformin regulates the expression of P53 through the AMPK pathway.37 Specifically, AMPK activates the activity of P53 by phosphorylating the 15th serine. The mechanism of metformin combined with cytokines is shown in Figure 5 and Table 2.
 |
Table 2 Mechanism of Action of Metformin Combined with Cytokines in the Treatment of Liver Cancer |
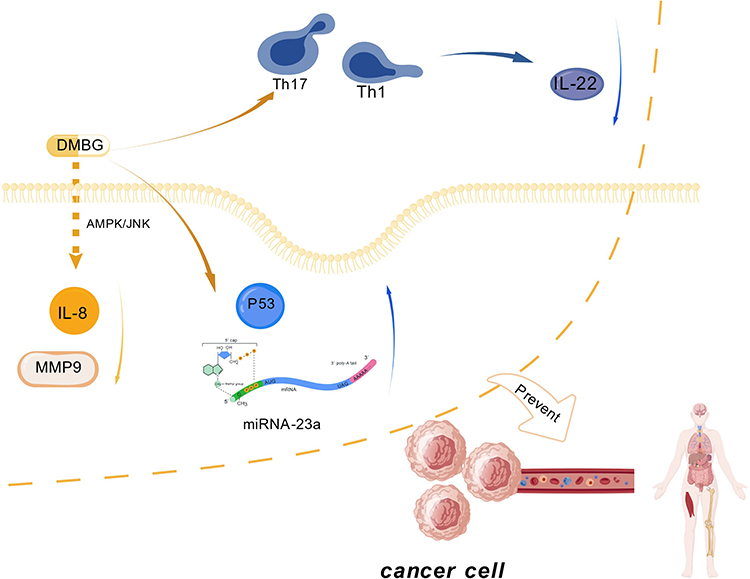 |
Figure 5 Mechanism of action of metformin in combination with other factors. Figure Created with BioGDP.com. Abbreviations: DMBG, metformin; JNK, Amino-terminal kinase; Th1 and Th17, subtypes of T lymphocytes; IL22, interleukin 22; IL8, interleukin 8; MMP9, a member of the endopeptidase protein family; P53, p53 gene; miRNA-23a, microRNA 23a. |
Tumor-Initiating Cells and Aging
Tumor stem cells (CSCs) or tumor initiation cells (TICs) are a class of cell populations with significant tumor-causing characteristics. These cells are similar to the human body’s ancestor stem cells and have the ability to differentiate and renew themselves. Therefore, they play a role in the entire cycle of tumor occurrence, development, metastasis, invasion and recurrence.186,187 The study found that metformin can directly act on the starting cells of the tumor and exert antitumor effects by impairing the self-renewal ability of the starting tumor cells. At least part of the mechanism is related to the increase in the number of AMPK caused by metformin, which leads to a decrease in the level of mTOR and p70S6 kinases.99 In addition, CD133 is one of the identification markers of CSCs.188–190 Metformin reduces CD133 expression through two mechanisms: by regulating the CD133-P1 promoter region or by inactivating STAT3,191,192 thereby targeting CSCs to achieve the treatment of liver cancer. The AMPK/CEBPβ pathway is a key mechanism by which metformin regulates the CD133 promoter.193 CEBPβ consists of three isomers, namely LAP*LAP, and LIP,194 where LAP can increase CD133 expression, while LIP can cause CD133 expression to decrease, so the ratio of LAP/LIP is the key to regulating CD133.193 For the cutting-edge field of therapeutic aging, metformin can induce the aging of liver cancer cells through the AMPK/SIRT1 pathway to achieve the purpose of treatment. The specific mechanism is AMPK activation, which leads to a decrease in SIRT1 phosphorylation and inactivation of SIRT1 acetylase.195 SIRT1 is a histone deacetylase, a member of the sirtuin family, which has important biological functions in regulating cell metabolism, aging, and stress response.196 Its potential effects include regulating the acetylation of a variety of substrates, such as p53 and FoxO.197–199 SIRT3 is also a member of the sirtuin family. As a key effector of the AMPK/mTOR pathway,200 it can reverse regulate p-mTOR and downstream HIF-1α, resulting in cancer-suppressing effects.201 At the same time, SIRT3 is also involved in the regulation of metabolism and aging,202 so SIRT-3 is regarded as a key predictor of liver cancer.203
Combined with Radiofrequency Ablation
Zhang et al research found that metformin can significantly weaken the proliferation and invasion ability of liver cancer in patients with insufficient radiofrequency ablation.204 Specifically, when radiofrequency ablation (RFA) is insufficient, the expression level of P-AKT in HCC cells increases and plays an important role in EMT.205 Studies have shown that metformin can reduce the expression of P-AKT by up-regulating AMPK and PTEN, and further down-regulate PCNA and VEGF to overcome the increased invasiveness caused by insufficient radiofrequency ablation.204
Microenvironment and Inhibition of Angiogenesis
It is worth noting that the production of tumor blood vessels is essential for the growth of tumors, and this mechanism is also applicable in liver cancer.206 In addition, the study of tumor microenvironment has become a hot topic in research. In liver cells, the microenvironment is mainly composed of hepatic stellate cells (HSC), fibroblasts, myofibroblasts, immune cells, and endothelial cells.207 The study found that the interaction between HCC and HSC can promote tumor angiogenesis, while metformin targets hepatic stellate cells through the AMPK pathway to inhibit the activity of hepatic stellate cells, thereby effectively reducing the ability of tumor blood vessels to generate.208
Immunotherapy
Immunotherapy has become a hot topic in current research and has emerged as a novel approach for cancer treatment. Metformin can induce T cell differentiation through the AMPK-mTOR pathway to participate in immune regulation.119 In patients with primary hepatocellular carcinoma and colorectal cancer liver metastasis, metformin treatment can upregulate the expression of metabolism-related genes (Mpc1, Pck1, Adh4) in NASH (non-alcoholic steatohepatitis) mice, increase CD8+ T cell motility, and restore the efficacy of anti-PD-1 therapy. In combination therapy with anti-PD-L1 and anti-VEGFR2 (simulating the clinical first-line regimen of atezolizumab plus bevacizumab), NASH also impairs its effectiveness, while metformin can reverse this inhibitory effect.209,210 However, the lack of clinical data on the combination of metformin with immunotherapeutic drugs provides us with new insights for future research.
Non-AMPK Pathway
As shown above, metformin can trigger ROS accumulation in an AMPK-independent manner. Even when using an AMPK inhibitor to block AMPK phosphorylation, it cannot suppress the reduction of mTOR expression, indicating that metformin affects mTOR through pathways other than AMPK.211 Previous reports have shown that metformin induces REDD1 through both AMPK-independent and p53-dependent mechanisms.212 Numerous findings suggest that AMPK-dependent and AMPK-independent pathways may coexist.79
Future Directions and Challenges
In recent years, metformin has emerged as a potential liver cancer treatment drug, demonstrating multiple anti-liver cancer mechanisms by activating the AMPK pathway. Studies show that its mechanisms of action are complex and multi-dimensional, but many issues remain unexplained. HIF-1α and HIF-2α have both been confirmed to be associated with drug resistance. It is worth further research to explore their relationship, whether they act synergistically or antagonistically under certain conditions. Regarding the application of metformin in liver cancer treatment, clear standard guidelines are currently lacking. Studies have indicated that the specific effects of metformin in clinical settings are closely related to its usage concentration: high concentrations (5–30 mM) may induce cell apoptosis and block the cell cycle; while low concentrations (0.1–0.2 mM) tend to inhibit the proliferation of tumor stem cells.192,213 And the dosage of metformin varies in different articles, with no unified standard. Additionally, although multiple clinical studies have confirmed the potential value of metformin as a liver cancer treatment drug, its safety issues still need further exploration. Current evidence is insufficient, and some studies even suggest that metformin may be ineffective or harmful for certain types of liver cancer, which to some extent increases its controversial application in the future clinical setting. Therefore, future research should focus on the following directions: first, draw on research from multi-omics and causal inference to validate the causal association between key downstream effect molecules of metformin (such as CEBPD, TIP30) and the risk or prognosis of liver cancer in large clinical cohorts, providing higher-level evidence-based medical evidence for the clinical application of metformin;214 second, exploring more precise dosing regimens to optimize its therapeutic effects and reduce side effects. Only with a comprehensive understanding of its mechanisms can more accurate safety and efficacy evidence be provided for its clinical application.
Limitation
This review has attempted to collect as many literature reports as possible on the mechanisms related to metformin and AMPK, and explores its potential multitarget effects. However, although metformin has been preliminarily revealed in existing studies to regulate the metabolism of liver cancer cells by activating the AMPK pathway, there may still be limitations in the scope of literature collection. In addition, the mechanism of action of metformin is not limited to the AMPK pathway, and the synergistic effects of the signal conduction pathway are not described in detail in this review. It is worth noting that although current studies have shown that metformin has antitumor effects, its application in clinical practice still faces many challenges, including the lack of clear safety and conclusive evidence of effectiveness. The results of this part of the research suggest that its true clinical efficacy still needs to be further verified by more in-depth and detailed research.
Conclusion
This article provides a systematic review of metformin’s effects on the AMPK pathway in the treatment of liver cancer, summarized as 1) inducing autophagy, 2) causing cell cycle arrest, 3) affecting glucose metabolism, 4) drug synergy, 5) inhibiting angiogenesis, 6) suppressing EMT, 7) combination with cytokines, and 8) immunotherapy combination. In addition, metformin has been widely used in clinical practice for the treatment of diabetes. Study results suggest it may have potential therapeutic effects on liver cancer, but its true efficacy still needs further validation. This finding provides us with a new research direction: first, in the clinical aspect, exploring through large-scale clinical models. The efficacy and safety of metformin, and to identify the appropriate dose range and verify key downstream effector molecules; second, in terms of molecular mechanisms, to explore in detail the specific molecular mechanisms through which metformin acts via the AMPK pathway and its relationship with other pathways, such as the interaction between AMPK and non-AMPK pathways, whether they antagonize or synergize under specific conditions, and the coordination or antagonism between HIF-1α and HIF-2α in drug resistance; third, research on metformin in specific areas is limited, such as epithelial-mesenchymal transition, and more experiments are needed to support these studies.
Abbreviations
HCC, hepatocellular carcinoma; 5-Fu, 5-Fluorouracil; AMPK, AMP-activated protein kinase; DMBG, metformin; Ros, Rosiglitazone.
Funding
This study was supported by the Provincial Medical and Health Research Project and the Hospital’s Technological Innovation Talent Project of the Fourth Affiliated Hospital of Harbin Medical University (respectively, ‘A Study on the Safety and Efficacy of AngioJet Mechanical Thrombectomy for the Treatment of Acute Pulmonary Embolism,’ project number: 20250909040005; and ‘Determination of Sensitivity and Efficacy Monitoring of Targeted Lung Cancer Therapy Using a Mutant EGFR High-Selectivity Probe,’ project number: HYDSYKJCXRC202117).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sia D, Villanueva A, Friedman SL, Llovet JM. Liver cancer cell of origin, molecular class, and effects on patient prognosis. Gastroenterology. 2017;152(4):745–19. doi:10.1053/j.gastro.2016.11.048
2. Kim E, Viatour P. Hepatocellular carcinoma: old friends and new tricks. Exp Mol Med. 2020;52(12):1898–1907. doi:10.1038/s12276-020-00527-1
3. Llovet JM, Kelley RK, Villanueva A, et al. Hepatocellular carcinoma. Nat Rev Dis Primers. 2021;7(1). doi:10.1038/s41572-020-00240-3.
4. Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. Ca A Cancer J Clin. 2022;72(1):7–33. doi:10.3322/caac.21708
5. Bray F, Laversanne M, Sung HYA, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca A Cancer J Clin. 2024;74(3):229–263. doi:10.3322/caac.21834
6. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. Ca A Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
7. Petrick JL, McGlynn KA. The changing epidemiology of primary liver cancer. Current Epidemiol Rep. 2019;6(2):104–111. doi:10.1007/s40471-019-00188-3
8. Chen L, Pei JH, Kuang J, et al. Effect of lifestyle intervention in patients with type 2 diabetes: a meta-analysis. Metab-Clin Exp. 2015;64(2):338–347. doi:10.1016/j.metabol.2014.10.018
9. Ardisson Korat AV, Willett WC, Hu FB. Diet, lifestyle, and genetic risk factors for type 2 diabetes: a review from the nurses’ health study, nurses’ health study 2, and health professionals’ follow-up study. Current Nutr Rep. 2014;3(4):345–354. doi:10.1007/s13668-014-0103-5
10. Weisman A, Fazli GS, Johns A, Booth GL. Evolving trends in the epidemiology, risk factors, and prevention of type 2 diabetes: a review. Canad J Cardiol. 2018;34(5):552–564. doi:10.1016/j.cjca.2018.03.002
11. El-Ashmawy NE, Khedr EG, El-Bahrawy HA, Abd El-Fattah EE. Sorafenib effect on liver neoplastic changes in rats: more than a kinase inhibitor. Clin Exp Med. 2017;17(2):185–191. doi:10.1007/s10238-016-0416-3
12. Durand F, Buffet C, Pelletier G, Hagege H, Ink O, Etienne JP. Hepatocellular carcinoma. New Engl J Med. 1993;328(1):65–64; authorreply65. doi:10.1056/NEJM199301073280114
13. Villanueva A, Hernandez-Gea V, Llovet JM. Medical therapies for hepatocellular carcinoma: a critical view of the evidence. Nat Rev Gastroenterol Hepatol. 2013;10(1):34–42. doi:10.1038/nrgastro.2012.199
14. Venook AP, Papandreou C, Furuse J, Ladrón de guevara L. The incidence and epidemiology of hepatocellular carcinoma: a global and regional perspective. Oncologist. 2010;15(S4):5–13. doi:10.1634/theoncologist.2010-S4-05
15. Rena G, Hardie DG, Pearson ER. The mechanisms of action of metformin. Diabetologia. 2017;60(9):1577–1585. doi:10.1007/s00125-017-4342-z
16. He L, Wondisford FE. Metformin action: concentrations matter. Cell Metab. 2015;21(2):159–162. doi:10.1016/j.cmet.2015.01.003
17. Fujita Y, Inagaki N. Metformin: clinical topics and new mechanisms of action. Diabetol Int. 2016;8(1):4–6. doi:10.1007/s13340-016-0300-0
18. Foretz M, Guigas B, Bertrand L, Pollak M, Viollet B. Metformin: from mechanisms of action to therapies. Cell Metabol. 2014;20(6):953–966. doi:10.1016/j.cmet.2014.09.018
19. Zheng Y, Ley SH, Hu FB. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat Rev Endocrinol. 2018;14(2):88–98. doi:10.1038/nrendo.2017.151
20. Dujic T, Causevic A, Bego T, et al. Organic cation transporter 1 variants and gastrointestinal side effects of metformin in patients with type 2 diabetes. Diabet Med. 2016;33(4):511–514. doi:10.1111/dme.13040
21. Maruthur NM, Tseng E, Hutfless S, et al. Diabetes medications as monotherapy or metformin-based combination therapy for type 2 diabetes a systematic review and meta-analysis. Annals Internal Med. 2016;164(11):740–+. doi:10.7326/M15-2650
22. Shaw RJ, Lamia KA, Vasquez D, et al. The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science. 2005;310(5754):1642–1646. doi:10.1126/science.1120781
23. Hundal RS, Krssak M, Dufour S, et al. Mechanism by which metformin reduces glucose production in type 2 diabetes. Diabetes. 2000;49(12):2063–2069. doi:10.2337/diabetes.49.12.2063
24. Ben Sahra I, Laurent K, Loubat A, et al. The antidiabetic drug metformin exerts an antitumoral effect in vitro and in vivo through a decrease of cyclin D1 level. Oncogene. 2008;27(25):3576–3586. doi:10.1038/sj.onc.1211024
25. Hadad SM, Appleyard V, Thompson AM. Therapeutic metformin/AMPK activation promotes the angiogenic phenotype in the ERalpha negative MDA-MB-435 breast cancer model. Breast Cancer Res Treatment. 2009;114(2):391. doi:10.1007/s10549-008-0016-3
26. Brown KA, Hunger NI, Docanto M, Simpson ER. Metformin inhibits aromatase expression in human breast adipose stromal cells via stimulation of AMP-activated protein kinase. Breast Cancer Res Treatment. 2010;123(2):591–596. doi:10.1007/s10549-010-0834-y
27. Hosono K, Endo H, Takahashi H, et al. Metformin suppresses azoxymethane-induced colorectal aberrant crypt foci by activating AMP-activated protein kinase. Mol Carcinogenesis. 2010;49(7):662–671. doi:10.1002/mc.20637
28. Kato K, Gong J, Iwama H, et al. The antidiabetic drug metformin inhibits gastric cancer cell proliferation in vitro and in vivo. Mol Cancer Therapeut. 2012;11(3):549–560. doi:10.1158/1535-7163.MCT-11-0594
29. Kobayashi M, Kato K, Iwama H, et al. Antitumor effect of metformin in esophageal cancer: in vitro study. Int J Oncol. 2013;42(2):517–524. doi:10.3892/ijo.2012.1722
30. Isakovic A, Harhaji L, Stevanovic D, et al. Dual antiglioma action of metformin: cell cycle arrest and mitochondria-dependent apoptosis. Cell Mol Life Sci. 2007;64(10):1290–1302. doi:10.1007/s00018-007-7080-4
31. Li D, Yeung S-CJ, Hassan MM, Konopleva M, Abbruzzese JL. Antidiabetic therapies affect risk of pancreatic cancer. Gastroenterology. 2009;137(2):482–488. doi:10.1053/j.gastro.2009.04.013
32. Wu C-Y, Chen H-P, Chen -T-T, Lin J-T, Wu M-S. Metformin decreases hepatocellular carcinoma risk in a dose-dependent manner: population-based and in vitro studies. Gastroenterology. 2012;142(5):S920–S920. doi:10.1016/S0016-5085(12)63571-9
33. Qu Z, Zhang Y, Liao M, Chen Y, Zhao J, Pan Y. In vitro and in vivo antitumoral action of metformin on hepatocellular carcinoma. Hepatol Res. 2012;42(9):922–933. doi:10.1111/j.1872-034X.2012.01007.x
34. Tsai P-C, Huang C-F, Yeh M-L, et al. Metformin and statins reduce hepatocellular carcinoma risk in chronic hepatitis C patients with failed antiviral therapy. Clin Mol Hepatol. 2024;30(3):468–486. doi:10.3350/cmh.2024.0038
35. Tsai P-C, Kuo H-T, Hung C-H, et al. Metformin reduces hepatocellular carcinoma incidence after successful antiviral therapy in patients with diabetes and chronic hepatitis C in Taiwan. J Hepatol. 2023;78(2):281–292. doi:10.1016/j.jhep.2022.09.019
36. Yi G, He ZM, Zhou XK, et al. Low concentration of metformin induces a p53-dependent senescence in hepatoma cells via activation of the AMPK pathway. Int J Oncol. 2013;43(5):1503–1510. doi:10.3892/ijo.2013.2077
37. Zheng LY, Yang W, Wu FQ, et al. Prognostic significance of AMPK activation and therapeutic effects of metformin in hepatocellular carcinoma. Clin Cancer Res. 2013;19(19):5372–5380. doi:10.1158/1078-0432.CCR-13-0203
38. Ma X, Aibaidula Y, Zhang C, et al. Lactucin alleviates lipid accumulation via AMPK-mediated autophagy and fatty acid β-oxidation in FFA-induced HepG2 cells. J Pharmacol Sci. 2025;159(2):116–127. doi:10.1016/j.jphs.2025.08.001
39. Zhang Y, Zhou J, Yang L, Xiao H, Liu D, Kang X. Ganoderma lucidum spore powder alleviates metabolic-associated fatty liver disease by improving lipid accumulation and oxidative stress via autophagy. Antioxidants. 2024;13(12):1501. doi:10.3390/antiox13121501
40. Hidalgo F, Ferretti AC, Etichetti CB, et al. Alpha lipoic acid diminishes migration and invasion in hepatocellular carcinoma cells through an AMPK-p53 axis. Scient Rep. 2024;14(1). doi:10.1038/s41598-024-72309-y.
41. Hu -H-H, Wang X, Lan B, et al. Glucose starvation mimetic aldometanib removes immune barriers permitting mice with hepatocellular carcinoma to live to normal ages. Cell Res. 2025;35(12):934–953. doi:10.1038/s41422-025-01195-4
42. Liu J, Zhang X, Fan X, et al. Liensinine reshapes the immune microenvironment and enhances immunotherapy by reprogramming metabolism through the AMPK-HIF-1α axis in hepatocellular carcinoma. J Exp Clin Cancer Res. 2025;44(1). doi:10.1186/s13046-025-03477-6.
43. Hardie DG, Carling D, Carlson M. The AMP-activated/SNF1 protein kinase subfamily: metabolic sensors of the eukaryotic cell? Annual Rev Biochem. 1998;67:821–855. doi:10.1146/annurev.biochem.67.1.821
44. Hardie DG. The AMP-activated protein kinase pathway--new players upstream and downstream. J Cell Sci. 2004;117(Pt 23):5479–5487. doi:10.1242/jcs.01540
45. Mihaylova MM, Shaw RJ. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat Cell Biol. 2011;13(9):1016–1023. doi:10.1038/ncb2329
46. Zhou G, Myers R, Li Y, et al. Role of AMP-activated protein kinase in mechanism of metformin action. J Clin Investig. 2001;108(8):1167–1174. doi:10.1172/JCI13505
47. Slattery ML, Fitzpatrick FA. Convergence of hormones, inflammation, and energy-related factors: a novel pathway of cancer etiology. Cancer Prevention Res. 2009;2(11):922–930. doi:10.1158/1940-6207.CAPR-08-0191
48. Corradetti MN, Inoki K, Bardeesy N, DePinho RA, Guan KL. Regulation of the TSC pathway by LKB1: evidence of a molecular link between tuberous sclerosis complex and Peutz-Jeghers syndrome (vol 18, pg 1533, 2004). Genes Dev. 2019;33(7–8):477. doi:10.1101/gad.324970.119
49. Baas AF, Kuipers J, van der Wel NN, et al. Complete polarization of single intestinal epithelial cells upon activation of LKB1 by STRAD. Cell. 2004;116(3):457–466. doi:10.1016/S0092-8674(04)00114-X
50. Hardie DG. Minireview: the AMP-activated protein kinase cascade: the key sensor of cellular energy status. Endocrinology. 2003;144(12):5179–5183. doi:10.1210/en.2003-0982
51. Xie Z, Dong Y, Scholz R, Neumann D, Zou M-H. Phosphorylation of LKB1 at serine 428 by protein kinase C-zeta is required for metformin-enhanced activation of the AMP-activated protein kinase in endothelial cells. Circulation. 2008;117(7):952–962. doi:10.1161/CIRCULATIONAHA.107.744490
52. Aw DKL, Sinha RA, Xie SY, Yen PM. Differential AMPK phosphorylation by glucagon and metformin regulates insulin signaling in human hepatic cells. Biochem Biophys Res Commun. 2014;447(4):569–573. doi:10.1016/j.bbrc.2014.04.031
53. Jones RG, Thompson CB. Tumor suppressors and cell metabolism: a recipe for cancer growth. Genes Dev. 2009;23(5):537–548. doi:10.1101/gad.1756509
54. Hardie DG, Ross FA, Hawley SA. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol. 2012;13(4):251–262. doi:10.1038/nrm3311
55. Motoshima H, Goldstein BJ, Igata M, Araki E. AMPK and cell proliferation--AMPK as a therapeutic target for atherosclerosis and cancer. J Physiol. 2006;574(Pt 1):63–71. doi:10.1113/jphysiol.2006.108324
56. Zhuang Y, Miskimins WK. Cell cycle arrest in Metformin treated breast cancer cells involves activation of AMPK, downregulation of cyclin D1, and requires p27Kip1 or p21Cip1. J Mol Signal. 2008;3:18. doi:10.1186/1750-2187-3-18
57. Choi YK, Park K-G. Metabolic roles of AMPK and metformin in cancer cells. Mol Cells. 2013;36(4):279–287. doi:10.1007/s10059-013-0169-8
58. Zoncu R, Efeyan A, Sabatini DM. mTOR: from growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol. 2011;12(1):21–35. doi:10.1038/nrm3025
59. Bhat M, Sonenberg N, Gores GJ. The mTOR pathway in hepatic malignancies. Hepatology. 2013;58(2):810–818. doi:10.1002/hep.26323
60. Wang N, Pan W, Zhu M, et al. Fangchinoline induces autophagic cell death via p53/sestrin2/AMPK signalling in human hepatocellular carcinoma cells. Brit J Pharmacol. 2011;164(2b):731–742. doi:10.1111/j.1476-5381.2011.01349.x
61. Hirsch HA, Iliopoulos D, Tsichlis PN, Struhl K. Metformin selectively targets cancer stem cells, and acts together with chemotherapy to block tumor growth and prolong remission. Cancer Res. 2009;69(19):7507–7511. doi:10.1158/0008-5472.CAN-09-2994
62. Shank J, Buckanovich R. Metformin targets ovarian cancer stem cells in vitro and in vivo. Gynecolog Oncol. 2012;125:S129–S130. doi:10.1016/j.ygyno.2011.12.315
63. Zhang D-M, Liu J-S, Deng L-J, et al. Arenobufagin, a natural bufadienolide from toad venom, induces apoptosis and autophagy in human hepatocellular carcinoma cells through inhibition of PI3K/Akt/mTOR pathway. Carcinogenesis. 2013;34(6):1331–1342. doi:10.1093/carcin/bgt060
64. Nakabayashi M, Werner L, Courtney KD, et al. Phase II trial of RAD001 and bicalutamide for castration-resistant prostate cancer. BJU Int. 2012;110(11):1729–1735. doi:10.1111/j.1464-410X.2012.11456.x
65. Grgic T, Mis L, Hammond JM. Everolimus: a new mammalian target of rapamycin inhibitor for the treatment of advanced renal cell carcinoma. Annals Pharmacother. 2011;45(1):78–83. doi:10.1345/aph.1M288
66. Reungwetwattana T, Molina JR, Mandrekar SJ, et al. Brief report: a phase II “Window-of-Opportunity” frontline study of the mTOR inhibitor, temsirolimus given as a single agent in patients with advanced NSCLC, an NCCTG study. J Thoracic Oncol. 2012;7(5):919–922. doi:10.1097/JTO.0b013e31824de0d6
67. Nishikawa T, Takaoka M, Ohara T, et al. Antiproliferative effect of a novel mTOR inhibitor temsirolimus contributes to the prolonged survival of orthotopic esophageal cancer-bearing mice. Cancer Biol Ther. 2013;14(3):230–236. doi:10.4161/cbt.23294
68. Vara D, Salazar M, Olea-Herrero N, Guzman M, Velasco G, Diaz-Laviada I. Anti-tumoral action of cannabinoids on hepatocellular carcinoma: role of AMPK-dependent activation of autophagy. Cell Death Diff. 2011;18(7):1099–1111. doi:10.1038/cdd.2011.32
69. Lee CW, Wong LLY, Tse EYT, et al. AMPK promotes p53 acetylation via phosphorylation and inactivation of SIRT1 in liver cancer cells. Cancer Res. 2012;72(17):4394–4404. doi:10.1158/0008-5472.CAN-12-0429
70. Yu R, Z-q Z, Wang B, H-x J, Cheng L, L-m S. Berberine-induced apoptotic and autophagic death of HepG2 cells requires AMPK activation. Cancer Cell Int. 2014;14:14. doi:10.1186/1475-2867-14-14
71. Cheng JD, Huang TL, Li YF, et al. AMP-activated protein kinase suppresses the in vitro and in vivo proliferation of hepatocellular carcinoma. PLoS One. 2014;9(4):e93256.
72. Li WD, Saud SM, Young MR, Chen GH, Hua BJ. Targeting AMPK for cancer prevention and treatment. Oncotarget. 2015;6(10):7365–7378. doi:10.18632/oncotarget.3629
73. Yun CW, Jeon J, Go G, Lee JH, Lee SH. The dual role of autophagy in cancer development and a therapeutic strategy for cancer by targeting autophagy. Int J Mol Sci. 2021;22(1):179.
74. Quan W, Lee M-S. Role of autophagy in the control of body metabolism. Endocrinol Metab. 2013;28(1):6–11. doi:10.3803/EnM.2013.28.1.6
75. Amaravadi RK. Autophagy-induced tumor dormancy in ovarian cancer. J Clin Investig. 2008;118(12):3837–3840. doi:10.1172/JCI37667
76. Gonzalez CD, Lee M-S, Marchetti P, et al. The emerging role of autophagy in the pathophysiology of diabetes mellitus. Autophagy. 2011;7(1):2–11. doi:10.4161/auto.7.1.13044
77. Tsujimoto Y, Shimizu S. Another way to die: autophagic programmed cell death. Cell Death Diff. 2005;12 Suppl 2:1528–1534. doi:10.1038/sj.cdd.4401777
78. Harris K, Smith L. Safety and efficacy of metformin in patients with type 2 diabetes mellitus and chronic hepatitis C. Annals Pharmacother. 2013;47(10):1348–1352. doi:10.1177/1060028013503108
79. Cai XB, Hu X, Cai BZ, et al. Metformin suppresses hepatocellular carcinoma cell growth through induction of cell cycle G1/G0 phase arrest and p21<SUP>CIP</SUP> and p27KIP expression and downregulation of cyclin D1 in vitro and in vivo. Oncol Rep. 2013;30(5):2449–2457. doi:10.3892/or.2013.2718
80. Kim J-H, Alam MM, Park DB, et al. The effect of metformin treatment on CRBP-I level and cancer development in the liver of HBx transgenic mice. Kor J Physiol Pharmacol. 2013;17(5):455–461. doi:10.4196/kjpp.2013.17.5.455
81. Tomic T, Botton T, Cerezo M, et al. Metformin inhibits melanoma development through autophagy and apoptosis mechanisms. Cell Death Dis. 2011;2:e199–e199. doi:10.1038/cddis.2011.86
82. Shi WY, Xiao D, Wang L, et al. Therapeutic metformin/AMPK activation blocked lymphoma cell growth via inhibition of mTOR pathway and induction of autophagy. Cell Death Dis. 2012;3:e275.
83. Cui JZ, Gong ZY, Shen HM. The role of autophagy in liver cancer: molecular mechanisms and potential therapeutic targets. Biochimica Et Biophysica Acta. 2013;1836(1):15–26. doi:10.1016/j.bbcan.2013.02.003
84. Chuang CH, Wang WJ, Li CF, et al. The combination of the prodrugs perforin-CEBPD and perforin-granzyme B efficiently enhances the activation of caspase signaling and kills prostate cancer. Cell Death Dis. 2014;5:e1220.
85. Ko C-Y, Hsu H-C, Shen M-R, Chang W-C, Wang J-M. Epigenetic silencing of CCAAT/enhancer-binding protein delta activity by YY1/polycomb group/DNA methyltransferase complex. J Biol Chem. 2008;283(45):30919–30932. doi:10.1074/jbc.M804029200
86. Jiang Y, Huang W, Wang J, et al. Metformin plays a dual role in MIN6 pancreatic β cell function through AMPK-dependent autophagy. Int J Biol Sci. 2014;10(3):268–277. doi:10.7150/ijbs.7929
87. Jiang T, Yu J-T, Zhu X-C, et al. Acute metformin preconditioning confers neuroprotection against focal cerebral ischaemia by pre-activation of AMPK-dependent autophagy. Brit J Pharmacol. 2014;171(13):3146–3157. doi:10.1111/bph.12655
88. Sanchez-Alvarez R, Martinez-Outschoorn UE, Lamb R, et al. Mitochondrial dysfunction in breast cancer cells prevents tumor growth Understanding chemoprevention with metformin. Cell Cycle. 2013;12(1):172–182. doi:10.4161/cc.23058
89. Din FVN, Valanciute A, Houde VP, et al. Aspirin inhibits mTOR signaling, activates AMP-activated protein kinase, and induces autophagy in colorectal cancer cells. Gastroenterology. 2012;142(7):1504–+. doi:10.1053/j.gastro.2012.02.050
90. Takahashi A, Kimura F, Yamanaka A, et al. Metformin impairs growth of endometrial cancer cells via cell cycle arrest and concomitant autophagy and apoptosis. Cancer Cell Int. 2014;14:14.
91. Settembre C, De Cegli R, Mansueto G, et al. TFEB controls cellular lipid metabolism through a starvation-induced autoregulatory loop (vol 15, pg 647, 2013). Nat Cell Biol. 2013;15(8):1016. doi:10.1038/ncb2814
92. Polager S, Ofir M, Ginsberg D. E2F1 regulates autophagy and the transcription of autophagy genes. Oncogene. 2008;27(35):4860–4864. doi:10.1038/onc.2008.117
93. Xu P, Das M, Reilly J, Davis RJ. JNK regulates FoxO-dependent autophagy in neurons. Genes Dev. 2011;25(4):310–322. doi:10.1101/gad.1984311
94. Milani M, Rzymski T, Mellor HR, et al. The role of ATF4 stabilization and autophagy in resistance of breast cancer cells treated with Bortezomib. Cancer Res. 2009;69(10):4415–4423. doi:10.1158/0008-5472.CAN-08-2839
95. Ma D, Panda S, Lin JD. Temporal orchestration of circadian autophagy rhythm by C/EBPbeta. EMBO J. 2011;30(22):4642–4651. doi:10.1038/emboj.2011.322
96. Rouschop KMA, van den Beucken T, Dubois L, et al. The unfolded protein response protects human tumor cells during hypoxia through regulation of the autophagy genes MAP1LC3B and ATG5. J Clin Investig. 2010;120(1):127–141. doi:10.1172/JCI40027
97. Tsai -H-H, Lai H-Y, Chen Y-C, et al. Metformin promotes apoptosis in hepatocellular carcinoma through the CEBPD-induced autophagy pathway. Oncotarget. 2017;8(8):13832–13845. doi:10.18632/oncotarget.14640
98. Pan Y-C, Li C-F, Ko C-Y, et al. CEBPD reverses RB/E2F1-mediated gene repression and participates in HMDB-induced apoptosis of cancer cells. Clin Cancer Res. 2010;16(23):5770–5780. doi:10.1158/1078-0432.CCR-10-1025
99. Saito T, Chiba T, Yuki K, et al. Metformin, a diabetes drug, eliminates tumor-initiating hepatocellular carcinoma cells. PLoS One. 2013;8(7):e70010. doi:10.1371/journal.pone.0070010
100. Ling S, Song L, Fan N, et al. Combination of metformin and sorafenib suppresses proliferation and induces autophagy of hepatocellular carcinoma via targeting the mTOR pathway. Int J Oncol. 2017;50(1):297–309. doi:10.3892/ijo.2016.3799
101. Gao C, Fang L, Zhang H, Zhang W-S, Li X-O, Du S-Y. Metformin induces autophagy via the AMPK-mTOR signaling pathway in human hepatocellular carcinoma cells. Cancer Manag Res. 2020;12:5803–5811. doi:10.2147/CMAR.S257966
102. Park D-B. Metformin promotes apoptosis but suppresses autophagy in glucose-deprived H4IIE hepatocellular carcinoma cells. Diab Metabol J. 2015;39(6):518–527. doi:10.4093/dmj.2015.39.6.518
103. Goyal H, Kaur J. Long non-coding RNAs and autophagy: dual drivers of Hepatocellular carcinoma progression. Cell Death Discovery. 2025;11(1). doi:10.1038/s41420-025-02667-7
104. Feng Y, He D, Yao Z, Klionsky DJ. The machinery of macroautophagy. Cell Res. 2014;24(1):24–41. doi:10.1038/cr.2013.168
105. Umemura A, He F, Taniguchi K, et al. p62, upregulated during preneoplasia, induces hepatocellular carcinogenesis by maintaining survival of stressed HCC-initiating cells. Cancer Cell. 2016;29(6):935–948. doi:10.1016/j.ccell.2016.04.006
106. Biliran H Jr, Wang Y, Banerjee S, et al. Overexpression of cyclin D1 promotes tumor cell growth and confers resistance to cisplatin-mediated apoptosis in an elastase-myc transgene-expressing pancreatic tumor cell line. Clin Cancer Res. 2005;11(16):6075–6086. doi:10.1158/1078-0432.CCR-04-2419
107. Nielsen NH, Arnerlov C, Emdin SO, Landberg G. Cyclin E overexpression, a negative prognostic factor in breast cancer with strong correlation to oestrogen receptor status. Brit J Cancer. 1996;74(6):874–880. doi:10.1038/bjc.1996.451
108. Polyak K, Kato JY, Solomon MJ, et al. p27Kip1, a cyclin-Cdk inhibitor, links transforming growth factor-beta and contact inhibition to cell cycle arrest. Genes Dev. 1994;8(1):9–22. doi:10.1101/gad.8.1.9
109. Ouyang J, Parakhia RA, Ochs RS. Metformin activates AMP kinase through inhibition of AMP deaminase. J Biol Chem. 2011;286(1):1–11. doi:10.1074/jbc.M110.121806
110. Miyoshi H, Kato K, Iwama H, et al. Effect of the anti-diabetic drug metformin in hepatocellular carcinoma in vitro and in vivo. Int J Oncol. 2014;45(1):322–332. doi:10.3892/ijo.2014.2419
111. Goncharov AP, Vashakidze N, Kharaishvili G. Epithelial-mesenchymal transition: a fundamental cellular and microenvironmental process in benign and malignant prostate pathologies. Biomedicines. 2024;12(2):418. doi:10.3390/biomedicines12020418
112. Giannelli G, Koudelkova P, Dituri F, Mikulits W. Role of epithelial to mesenchymal transition in hepatocellular carcinoma. J Hepatol. 2016;65(4):798–808. doi:10.1016/j.jhep.2016.05.007
113. Park -J-J, Seo S-M, Kim EJ, et al. Berberine inhibits human colon cancer cell migration via AMP-activated protein kinase-mediated downregulation of integrin β1 signaling. Biochem Biophys Res Commun. 2012;426(4):461–467. doi:10.1016/j.bbrc.2012.08.091
114. Kim H-S, Kim M-J, Kim EJ, Yang Y, Lee M-S, Lim J-S. Berberine-induced AMPK activation inhibits the metastatic potential of melanoma cells via reduction of ERK activity and COX-2 protein expression. Biochem Pharmacol. 2012;83(3):385–394. doi:10.1016/j.bcp.2011.11.008
115. Qu C, Zhang W, Zheng G, Zhang Z, Yin J, He Z. Metformin reverses multidrug resistance and epithelial-mesenchymal transition (EMT) via activating AMP-activated protein kinase (AMPK) in human breast cancer cells. Mol Cell Biochem. 2014;386(1–2):63–71. doi:10.1007/s11010-013-1845-x
116. Cerezo M, Tichet M, Abbe P, et al. Metformin blocks melanoma invasion and metastasis development in ampk/p53-dependent manner. Mol Cancer Therapeut. 2013;12(8):1605–1615. doi:10.1158/1535-7163.MCT-12-1226-T
117. Di Matteo S, Nevi L, Overi D, et al. Metformin exerts anti-cancerogenic effects and reverses epithelial-to-mesenchymal transition trait in primary human intrahepatic cholangiocarcinoma cells. Scient Rep. 2021;11(1). doi:10.1038/s41598-021-81172-0.
118. Musi N. AMP-activated protein kinase and type 2 diabetes. Current Med Chem. 2006;13(5):583–589. doi:10.2174/092986706776055724
119. Zhao D, Long X-D, Lu T-F, et al. Metformin decreases IL-22 secretion to suppress tumor growth in an orthotopic mouse model of hepatocellular carcinoma. Int J Cancer. 2015;136(11):2556–2565. doi:10.1002/ijc.29305
120. Ferretti AC, Hidalgo F, Tonucci FM, et al. Metformin and glucose starvation decrease the migratory ability of hepatocellular carcinoma cells: targeting AMPK activation to control migration. Scient Rep. 2019;9. doi:10.1038/s41598-019-39556-w
121. Liu X-D, Li Y-G, Wang G-Y, et al. Metformin protects high glucose-cultured cardiomyocytes from oxidative stress by promoting NDUFA13 expression and mitochondrial biogenesis via the AMPK signaling pathway. Mol Med Rep. 2020;22(6):5262–5270. doi:10.3892/mmr.2020.11599
122. Minh Truong D, Kim HG, Khanal T, et al. Metformin inhibits heme oxygenase-1 expression in cancer cells through inactivation of Raf-ERK-Nrf2 signaling and AMPK-independent pathways. Toxicol Appl Pharmacol. 2013;271(2):229–238. doi:10.1016/j.taap.2013.05.010
123. Hou X, Song J, Li X-N, et al. Metformin reduces intracellular reactive oxygen species levels by upregulating expression of the antioxidant thioredoxin via the AMPK-FOXO3 pathway. Biochem Biophys Res Commun. 2010;396(2):199–205. doi:10.1016/j.bbrc.2010.04.017
124. Gou S, Qiu L, Yang Q, et al. Metformin leads to accumulation of reactive oxygen species by inhibiting the NFE2L1 expression in human hepatocellular carcinoma cells. Toxicol Appl Pharmacol. 2021;420:115523. doi:10.1016/j.taap.2021.115523
125. Zhang C-S, Jiang B, Li M, et al. The lysosomal v-ATPase-ragulator complex is a common activator for AMPK and mTORC1, acting as a switch between catabolism and anabolism. Cell Metabol. 2014;20(3):526–540. doi:10.1016/j.cmet.2014.06.014
126. Qi XS, Wang D, Su CP, Li HY, Guo XZ. Hepatic resection versus transarterial chemoembolization for the initial treatment of hepatocellular carcinoma: a systematic review and meta-analysis. Oncotarget. 2015;6(21):18715–18733. doi:10.18632/oncotarget.4134
127. Shah SA, Cleary SP, Wei AC, et al. Recurrence after liver resection for hepatocellular carcinoma: risk factors, treatment, and outcomes. Surgery. 2007;141(3):330–339. doi:10.1016/j.surg.2006.06.028
128. Chen WT, Chau GY, Lui WY, et al. Recurrent hepatocellular carcinoma after hepatic resection: prognostic factors and long-term outcome. Eur J Surg Oncol. 2004;30(4):414–420. doi:10.1016/j.ejso.2004.01.013
129. Zhang Y, Liu G, Lin C, Liao G, Tang B. Silencing the EZH2 gene by RNA interference reverses the drug resistance of human hepatic multidrug-resistant cancer cells to 5-Fu. Life Sci. 2013;92(17–19):896–902. doi:10.1016/j.lfs.2013.03.010
130. Cheng A-L, Kang Y-K, Chen Z, et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: a Phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009;10(1):25–34. doi:10.1016/S1470-2045(08)70285-7
131. Gollob JA, Wilhelm S, Carter C, Kelley SL. Role of Raf kinase in cancer: therapeutic potential of targeting the Raf/MEK/ERK signal transduction pathway. Seminars Oncology. 2006;33(4):392–406. doi:10.1053/j.seminoncol.2006.04.002
132. Chang YS, Adnane J, Trail PA, et al. Sorafenib (BAY 43-9006) inhibits tumor growth and vascularization and induces tumor apoptosis and hypoxia in RCC xenograft models. Cancer Chemother Pharmacol. 2007;59(5):561–574. doi:10.1007/s00280-006-0393-4
133. Wilhelm SM, Carter C, Tang L, et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004;64(19):7099–7109. doi:10.1158/0008-5472.CAN-04-1443
134. Llovet JM, Ricci S, Mazzaferro V, et al. Sorafenib in advanced hepatocellular carcinoma. New Engl J Med. 2008;359(4):378–390. doi:10.1056/NEJMoa0708857
135. Zhao D, Zhai B, He C, et al. Upregulation of HIF-2α induced by sorafenib contributes to the resistance by activating the TGF-α/EGFR pathway in hepatocellular carcinoma cells. Cell Signal. 2014;26(5):1030–1039. doi:10.1016/j.cellsig.2014.01.026
136. Menrad H, Werno C, Schmid T, et al. Roles of hypoxia-inducible factor-1alpha (HIF-1alpha) versus HIF-2alpha in the survival of hepatocellular tumor spheroids. Hepatology. 2010;51(6):2183–2192. doi:10.1002/hep.23597
137. Zhao J, Lu B, Xu H, et al. Thirty-kilodalton Tat-interacting protein suppresses tumor metastasis by inhibition of osteopontin transcription in human hepatocellular carcinoma. Hepatology. 2008;48(1):265–275. doi:10.1002/hep.22280
138. Chen V, Shtivelman E. CC3/TIP30 regulates metabolic adaptation of tumor cells to glucose limitation. Cell Cycle. 2010;9(24):4941–4953. doi:10.4161/cc.9.24.14230
139. Gordan JD, Bertout JA, Hu C-J, Diehl JA, Simon MC. HIF-2alpha promotes hypoxic cell proliferation by enhancing c-myc transcriptional activity. Cancer Cell. 2007;11(4):335–347. doi:10.1016/j.ccr.2007.02.006
140. Patel SA, Simon MC. Biology of hypoxia-inducible factor-2alpha in development and disease. Cell Death Diff. 2008;15(4):628–634. doi:10.1038/cdd.2008.17
141. Zhu M, Yin F, Fan X, et al. Decreased TIP30 promotes Snail-mediated epithelial-mesenchymal transition and tumor-initiating properties in hepatocellular carcinoma. Oncogene. 2015;34(11):1420–1431. doi:10.1038/onc.2014.73
142. You A, Cao M, Guo Z, et al. Metformin sensitizes sorafenib to inhibit postoperative recurrence and metastasis of hepatocellular carcinoma in orthotopic mouse models. J Hematol Oncol. 2016;9. doi:10.1186/s13045-016-0253-6
143. Ceccarelli J, Delfino L, Zappia E, et al. The redox state of the lung cancer microenvironment depends on the levels of thioredoxin expressed by tumor cells and affects tumor progression and response to prooxidants. Int J Cancer. 2008;123(8):1770–1778. doi:10.1002/ijc.23709
144. Noike T, Miwa S, Soeda J, Kobayashi A, S-i M. Increased expression of thioredoxin-1, vascular endothelial growth factor, and redox factor-1 is associated with poor prognosis in patients with liver metastasis from colorectal cancer. Human Pathol. 2008;39(2):201–208. doi:10.1016/j.humpath.2007.04.024
145. Guo Z, Cao M, You A, et al. Metformin inhibits the prometastatic effect of sorafenib in hepatocellular carcinoma by upregulating the expression of TIP30. Cancer Sci. 2016;107(4):507–513. doi:10.1111/cas.12885
146. Alsowmely AM, Hodgson HJF. Non-surgical treatment of hepatocellular carcinoma. Alimentary Pharmacol Therapeut. 2002;16(1):1–15. doi:10.1046/j.1365-2036.2002.01149.x
147. Li Y, Ye D. Cancer therapy by targeting hypoxia-inducible factor-1. Current Cancer Drug Targets. 2010;10(7):782–796. doi:10.2174/156800910793605857
148. Huang C, Xu D, Xia Q, Wang P, Rong C, Su Y. Reversal of P-glycoprotein-mediated multidrug resistance of human hepatic cancer cells by Astragaloside II. J Pharm Pharmacol. 2012;64(12):1741–1750. doi:10.1111/j.2042-7158.2012.01549.x
149. W-s J, Z-l K, Z-f S, Y-z J, W-k Z, G-f C. Regulation of hypoxia inducible factor-1alpha expression by the alteration of redox status in HepG2 cells. J Exp Clin Cancer Res. 2011;30:61. doi:10.1186/1756-9966-30-61
150. Zhu H, S-f L, Wang J, et al. Effect of environmental factors on chemoresistance of HepG2 cells by regulating hypoxia-inducible factor-1α. Chin Med J. 2012;125(6):1095–1103.
151. Zhu MM, Tong JL, Xu Q, et al. Increased JNK1 signaling pathway is responsible for ABCG2-mediated multidrug resistance in human colon cancer. PLoS One. 2012;7(8). doi:10.1371/journal.pone.0041763.
152. Zannella VE, Dal Pra A, Muaddi H, et al. Reprogramming metabolism with metformin improves tumor oxygenation and radiotherapy response. Clin Cancer Res. 2013;19(24):6741–6750. doi:10.1158/1078-0432.CCR-13-1787
153. Takiyama Y, Harumi T, Watanabe J, et al. Tubular injury in a rat model of type 2 diabetes is prevented by metformin: a possible role of HIF-1alpha expression and oxygen metabolism. Diabetes. 2011;60(3):981–992. doi:10.2337/db10-0655
154. Ling SB, Tian Y, Zhang HQ, et al. Metformin reverses multidrug resistance in human hepatocellular carcinoma Bel-7402/5-fluorouracil cells. Mol Med Rep. 2014;10(6):2891–2897. doi:10.3892/mmr.2014.2614
155. Da Ros R, Assaloni R, Ceriello A. The preventive anti-oxidant action of thiazolidinediones: a new therapeutic prospect in diabetes and insulin resistance. Diab Med. 2004;21(11):1249–1252. doi:10.1111/j.1464-5491.2004.01312.x
156. Zinman B, Harris SB, Neuman J, et al. Low-dose combination therapy with rosiglitazone and metformin to prevent type 2 diabetes mellitus (CANOE trial): a double-blind randomised controlled study. Lancet. 2010;376(9735):103–111. doi:10.1016/S0140-6736(10)60746-5
157. Phung OJ, Sood NA, Sill BE, Coleman CI. Oral anti-diabetic drugs for the prevention of Type 2 diabetes. Diab Med. 2011;28(8):948–964. doi:10.1111/j.1464-5491.2011.03303.x
158. Motola D, Piccinni C, Biagi C, et al. Cardiovascular, ocular and bone adverse reactions associated with thiazolidinediones a disproportionality analysis of the US FDA adverse event reporting system database. Drug Safety. 2012;35(4):315–323. doi:10.2165/11596510-000000000-00000
159. Rosenstock J, Rood J, Cobitz A, Huang C, Garber A. Improvement in glycaemic control with rosiglitazone/metformin fixed-dose combination therapy in patients with type 2 diabetes with very poor glycaemic control. Diab Obes Metabol. 2006;8(6):643–649. doi:10.1111/j.1463-1326.2006.00648.x
160. Govindarajan R, Ratnasinghe L, Simmons DL, et al. Thiazolidinediones and the risk of lung, prostate, and colon cancer in patients with diabetes. J Clin Oncol. 2007;25(12):1476–1481. doi:10.1200/JCO.2006.07.2777
161. Liu Y, Hu X, Shan X, Chen K, Tang H. Rosiglitazone metformin adduct inhibits hepatocellular carcinoma proliferation via activation of AMPK/p21 pathway. Cancer Cell Int. 2019;19:19. doi:10.1186/s12935-019-0731-3
162. Zhou H, Wang S, Zhu P, Hu S, Chen Y, Ren J. Empagliflozin rescues diabetic myocardial microvascular injury via AMPK-mediated inhibition of mitochondrial fission (vol 15, pg 335, 2018). Redox Biol. 2024;71.:335–346
163. Abdelhamid AM, Saber S, Youssef ME, et al. Empagliflozin adjunct with metformin for the inhibition of hepatocellular carcinoma progression: emerging approach for new application. Biomed Pharmacother. 2022;145:112455.
164. Wang L-W, Li Z-S, Zou D-W, Jin Z-D, Gao J, Xu G-M. Metformin induces apoptosis of pancreatic cancer cells. World J Gastroenterol. 2008;14(47):7192–7198. doi:10.3748/wjg.14.7192
165. Silvestri A, Palumbo F, Rasi I, et al. Metformin induces apoptosis and downregulates pyruvate kinase M2 in breast cancer cells only when grown in nutrient-poor conditions. PLoS One. 2015;10(8):e0136250. doi:10.1371/journal.pone.0136250
166. Ben Sahra I, Laurent K, Giuliano S, et al. Targeting cancer cell metabolism: the combination of metformin and 2-deoxyglucose induces p53-dependent apoptosis in prostate cancer cells. Cancer Res. 2010;70(6):2465–2475. doi:10.1158/0008-5472.CAN-09-2782
167. Dinarello CA. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunolog Rev. 2018;281(1):8–27. doi:10.1111/imr.12621
168. Zhao C, Zheng L, Ma Y, et al. Low-dose metformin suppresses hepatocellular carcinoma metastasis via the AMPK/JNK/IL-8 pathway. Int J Immunopathol Pharmacol. 2024;38:03946320241249445.
169. Choi SH, Kwon O-J, Park JY, et al. Inhibition of tumour angiogenesis and growth by small hairpin HIF-1α and IL-8 in hepatocellular carcinoma. Liver Int. 2014;34(4):632–642. doi:10.1111/liv.12375
170. Tang KH, Ma S, Lee TK, et al. CD133+liver tumor-initiating cells promote tumor angiogenesis, growth, and self-renewal through neurotensin/interleukin-8/CXCL1 signaling. Hepatology. 2012;55(3):807–820. doi:10.1002/hep.24739
171. Zhao T, Wei P, Zhang C, et al. Nifuroxazide suppresses PD-L1 expression and enhances the efficacy of radiotherapy in hepatocellular carcinoma. ELife. 2024;12:RP90911.
172. Vranic S, Cyprian FS, Gatalica Z, Palazzo J. PD-L1 status in breast cancer: current view and perspectives. Seminars Cancer Biol. 2021;72:146–154. doi:10.1016/j.semcancer.2019.12.003
173. Liu G, Yin L, Ouyang X, Zeng K, Xiao Y, Li Y. M2 macrophages promote HCC cells invasion and migration via miR-149-5p/MMP9 signaling. J Cancer. 2020;11(5):1277–1287. doi:10.7150/jca.35444
174. Dumoutier L, Louahed J, Renauld JC. Cloning and characterization of IL-10-related T cell-derived inducible factor (IL-TIF), a novel cytokine structurally related to IL-10 and inducible by IL-9. J Immunol. 2000;164(4):1814–1819. doi:10.4049/jimmunol.164.4.1814
175. Renauld J-C. Class II cytokine receptors and their ligands: key antiviral and inflammatory modulators. Nat Rev Immunol. 2003;3(8):667–676. doi:10.1038/nri1153
176. Radaeva S, Sun R, Pan H-N, Hong F, Gao B. Interleukin 22 (IL-22) plays a protective role in T cell-mediated murine hepatitis: IL-22 is a survival factor for hepatocytes via STAT3 activation. Hepatology. 2004;39(5):1332–1342. doi:10.1002/hep.20184
177. Park O, Wang H, Weng H, et al. In vivo consequences of liver-specific interleukin-22 expression in mice: implications for human liver disease progression. Hepatology. 2011;54(1):252–261. doi:10.1002/hep.24339
178. Jiang R, Tan Z, Deng L, et al. Interleukin-22 promotes human hepatocellular carcinoma by activation of STAT3. Hepatology. 2011;54(3):900–909. doi:10.1002/hep.24486
179. Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120(1):15–20. doi:10.1016/j.cell.2004.12.035
180. Brodersen P, Voinnet O. Revisiting the principles of microRNA target recognition and mode of action. Nat Rev Mol Cell Biol. 2009;10(2):141–148. doi:10.1038/nrm2619
181. Bartel DP. Metazoan MicroRNAs. Cell. 2018;173(1):20–51. doi:10.1016/j.cell.2018.03.006
182. Peng H, Hou M, Wu Z, et al. Plasma exosomal miR-122 regulates the efficacy of metformin via AMPK in type 2 diabetes and hepatocellular carcinoma. Heliyon. 2022;8(11):e11503. doi:10.1016/j.heliyon.2022.e11503
183. Sun Y, Tao C, Huang X, et al. Metformin induces apoptosis of human hepatocellular carcinoma HepG2 cells by activating an AMPK/p53/miR-23a/FOXA1 pathway. Oncotargets Ther. 2016;9:2845–2853.
184. Nie M, Yu S, Peng S, Fang Y, Wang H, Yang X. miR-23a and miR-27a promote human granulosa cell apoptosis by targeting SMAD5. Biol Reprod. 2015;93(4). doi:10.1095/biolreprod.115.130690
185. Zheng H, Li W, Wang Y, et al. miR-23a inhibits E-cadherin expression and is regulated by AP-1 and NFAT4 complex during Fas-induced EMT in gastrointestinal cancer. Carcinogenesis. 2014;35(1):173–183. doi:10.1093/carcin/bgt274
186. Visvader JE, Lindeman GJ. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer. 2008;8(10):755–768. doi:10.1038/nrc2499
187. Jordan CT, Guzman ML, Noble M. Cancer stem cells. New Engl J Med. 2006;355(12):1253–1261. doi:10.1056/NEJMra061808
188. Hermann PC, Huber SL, Herrler T, et al. Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer. Cell Stem Cell. 2007;1(3):313–323. doi:10.1016/j.stem.2007.06.002
189. Ricci-Vitiani L, Lombardi DG, Pilozzi E, et al. Identification and expansion of human colon-cancer-initiating cells. Nature. 2007;445(7123):111–115. doi:10.1038/nature05384
190. Yin S, Li J, Hu C, et al. CD133 positive hepatocellular carcinoma cells possess high capacity for tumorigenicity. Int J Cancer. 2007;120(7):1444–1450. doi:10.1002/ijc.22476
191. Chai X, Chu H, Yang X, Meng Y, Shi P, Gou S. Metformin Increases sensitivity of pancreatic cancer cells to gemcitabine by reducing CD133+ cell populations and suppressing ERK/P70S6K signaling. Scient Rep. 2015;5:14404.
192. Gou S, Cui P, Li X, Shi P, Liu T, Wang C. Low concentrations of metformin selectively inhibit CD133<SUP>+ cell proliferation in pancreatic cancer and have anticancer action. PLoS One. 2013;8(5):e63969. doi:10.1371/journal.pone.0063969
193. Maehara O, Ohnishi S, Asano A, et al. Metformin regulates the expression of CD133 through the AMPK-CEBPβ pathway in hepatocellular carcinoma cell lines. Neoplasia. 2019;21(6):545–556. doi:10.1016/j.neo.2019.03.007
194. Won C, Kim B-H, Yi EH, et al. Signal transducer and activator of transcription 3-mediated CD133 up-regulation contributes to promotion of hepatocellular carcinoma. Hepatology. 2015;62(4):1160–1173. doi:10.1002/hep.27968
195. Zhou X, Chen J, Chen L, et al. Negative regulation of Sirtuin 1 by AMP-activated protein kinase promotes metformin-induced senescence in hepatocellular carcinoma xenografts. Cancer Lett. 2017;411:1–11. doi:10.1016/j.canlet.2017.09.027
196. Brooks CL, Gu W. How does SIRT1 affect metabolism, senescence and cancer? Nat Rev Cancer. 2009;9(2):123–128. doi:10.1038/nrc2562
197. Brunet A, Sweeney LB, Sturgill JF, et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004;303(5666):2011–2015. doi:10.1126/science.1094637
198. Vaziri H, Dessain SK, Ng Eaton E, et al. hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase. Cell. 2001;107(2):149–159. doi:10.1016/S0092-8674(01)00527-X
199. Luo J, Nikolaev AY, Imai S, et al. Negative control of p53 by Sir2alpha promotes cell survival under stress. Cell. 2001;107(2):137–148. doi:10.1016/S0092-8674(01)00524-4
200. Bell EL, Emerling BM, Ricoult SJH, Guarente L. SirT3 suppresses hypoxia inducible factor 1alpha and tumor growth by inhibiting mitochondrial ROS production. Oncogene. 2011;30(26):2986–2996. doi:10.1038/onc.2011.37
201. Yang S-L, Liu L-P, Jiang J-X, Xiong Z-F, He Q-J, Wu C. The correlation of expression levels of HIF-1 and HIF-2 in hepatocellular carcinoma with capsular invasion, portal vein tumor thrombi and patients clinical outcome. Jap J Clin Oncol. 2014;44(2):159–167. doi:10.1093/jjco/hyt194
202. Teodoro JS, Duarte FV, Gomes AP, et al. Berberine reverts hepatic mitochondrial dysfunction in high-fat fed rats: a possible role for SirT3 activation. Mitochondrion. 2013;13(6):637–646. doi:10.1016/j.mito.2013.09.002
203. De Matteis S, Scarpi E, Granato AM, et al. Role of SIRT-3, p-mTOR and HIF-1 in hepatocellular carcinoma patients affected by metabolic dysfunctions and in chronic treatment with metformin. Int J Mol Sci. 2019;20(6):1503. doi:10.3390/ijms20061503
204. Zhang Q, Kong J, Dong S, Xu W, Sun W. Metformin exhibits the anti-proliferation and anti-invasion effects in hepatocellular carcinoma cells after insufficient radiofrequency ablation. Cancer Cell Int. 2017;17:17. doi:10.1186/s12935-016-0380-8
205. Dong S, Kong J, Kong F, et al. Insufficient radiofrequency ablation promotes epithelial-mesenchymal transition of hepatocellular carcinoma cells through Akt and ERK signaling pathways. J Transl Med. 2013;11. doi:10.1186/1479-5876-11-273
206. Yang ZF, Poon RTP. Vascular changes in hepatocellular carcinoma. Anat Record. 2008;291(6):721–734. doi:10.1002/ar.20668
207. Hernandez-Gea V, Toffanin S, Friedman SL, Llovet JM. Role of the microenvironment in the pathogenesis and treatment of hepatocellular carcinoma. Gastroenterology. 2013;144(3):512–527. doi:10.1053/j.gastro.2013.01.002
208. Qu H, Yang X. Metformin inhibits angiogenesis induced by interaction of hepatocellular carcinoma with hepatic stellate cells. Cell Biochem Biophys. 2015;71(2):931–936. doi:10.1007/s12013-014-0287-8
209. Wabitsch S, McCallen JD, Kamenyeva O, et al. Metformin treatment rescues CD8+T-cell response to immune checkpoint inhibitor therapy in mice with NAFLD. J Hepatol. 2022;77(3):748–760. doi:10.1016/j.jhep.2022.03.010
210. Papadakos SP, Ferraro D, Carbone G, et al. The emerging role of metformin in the treatment of hepatocellular carcinoma: is there any value in repurposing metformin for HCC immunotherapy? Cancers. 2023;15(12):3161. doi:10.3390/cancers15123161
211. Xiong Y, Lu Q-J, Zhao J, Wu G-Y. Metformin inhibits growth of hepatocellular carcinoma cells by inducing apoptosis via mitochondrion-mediated pathway. Asian Pacific J Cancer Prevent. 2012;13(7):3275–3279. doi:10.7314/APJCP.2012.13.7.3275
212. DeWaal D, Nogueira V, Terry AR, et al. Hexokinase-2 depletion inhibits glycolysis and induces oxidative phosphorylation in hepatocellular carcinoma and sensitizes to metformin. Nat Commun. 2018;9:9. doi:10.1038/s41467-017-01881-x
213. Tong D, Liu Q, Liu G, et al. Metformin inhibits castration-induced EMT in prostate cancer by repressing COX2/PGE2/STAT3 axis. Cancer Lett. 2017;389:23–32. doi:10.1016/j.canlet.2016.12.031
214. Zhu W, Fan C, Liu B, et al. Therapeutic targets for hepatocellular carcinoma identified using proteomics and Mendelian randomization. J Gastroenterol Hepatol. 2025;40(1):282–293. doi:10.1111/jgh.16785
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Ginsenoside Rg1 Ameliorates Pancreatic Injuries via the AMPK/mTOR Pathway in vivo and in vitro
Chen J, Zhu G, Xiao W, Huang X, Wang K, Zong Y
Diabetes, Metabolic Syndrome and Obesity 2023, 16:779-794
Published Date: 15 March 2023
STK11 Alleviates Pulmonary Inflammation During Acute Lung Injury by Phosphorylating AMPK to Activate Autophagy in A549 Cell Model
Xiao R, Qi Z, Chen T, Wei F
Journal of Inflammation Research 2025, 18:17789-17802
Published Date: 19 December 2025