Back to Journals » Journal of Inflammation Research » Volume 19
Pyroptosis, A Double-Edged Sword in Respiratory Diseases: A Narrative Review
Received 21 February 2026
Accepted for publication 30 April 2026
Published 16 May 2026 Volume 2026:19 601946
DOI https://doi.org/10.2147/JIR.S601946
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Chen Yu,1,* Caijun Dai,1,* Lujian Zhu2
1Department of Respiratory and Critical Care Medicine, Affiliated Jinhua Hospital, Zhejiang University School of Medicine, Jinhua, People’s Republic of China; 2Department of Infectious Disease, Affiliated Jinhua Hospital, Zhejiang University School of Medicine, Jinhua, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lujian Zhu, Department of Infectious Disease, Affiliated Jinhua Hospital, Zhejiang University School of Medicine, Jinhua, People’s Republic of China, Email [email protected]
Abstract: Pyroptosis is an inflammatory form of programmed cell death (PCD) driven by the gasdermin protein family. This process begins when danger- or damage-associated molecular patterns (DAMPs) are recognized, triggering either the canonical Caspase-1 or non-canonical Caspase-4/5/11 pathways. These pathways activate the cleavage of gasdermin D (GSDMD), releasing N-terminal fragments that form pores in cell membranes. This pore formation increases membrane permeability, leading to cell swelling and eventual lytic rupture. Pyroptosis serves as a double-edged sword in innate immunity: it defends against pathogens but can cause excessive inflammation and tissue damage if dysregulated. Evidence suggests that pyroptosis is involved in various respiratory diseases. The roles of pyroptosis and the Gasdermin family are complex, often exhibiting dual effects that complicate clinical interpretations. This review explores the molecular mechanisms and regulatory networks of pyroptosis, focusing on recent findings related to its role in pulmonary infections, acute lung injury (ALI)/ acute respiratory distress syndrome (ARDS), allergic asthma, chronic obstructive pulmonary disease (COPD), pulmonary fibrosis (PF), and lung cancer. We also examine therapeutic strategies targeting pyroptotic pathways and recent advancements in their development, aiming to provide a theoretical foundation and new directions for mechanistic research and clinical translation in respiratory immune-inflammatory diseases.
Keywords: pyroptosis, inflammasome, immuno-inflammation, respiratory diseases, therapeutic targets
Introduction
The respiratory system, as the primary interface between the body and the environment, is continually exposed to pathogens, smoke, particulates, and other irritants.1 This exposure can lead to persistent immune-inflammatory responses and tissue damage.2 Dysregulated inflammation may cause airway epithelial damage, lung structure remodeling, and altered immune function, resulting in respiratory diseases such as ARDS, COPD, allergic asthma, PF, and lung cancer.3 According to recent epidemiological data, respiratory diseases remain a major cause of global mortality worldwide, with chronic obstructive pulmonary disease and lower respiratory infections continuing to rank among the leading causes of death.4 These findings highlight the substantial and persistent global burden of respiratory diseases. In addition, the high mortality associated with severe conditions such as pulmonary fibrosis, particularly in the post-COVID-19 era, further underscores the importance of advancing research in this field.5 Pyroptosis, first described by Cookson et al in 2001 during Salmonella infection, contrasts with the immunologically silent nature of apoptosis. It is characterized by cell swelling, lysis, pore formation in the plasma membrane, and the massive release of proinflammatory mediators, thereby serving as a mechanism that links pathogen clearance with inflammatory injury.6 Gasdermin proteins, which are central executors of pyroptosis, are crucial in forming these pores and facilitating pyroptosis. Initially linked to immune defense against infections, pyroptosis is now recognized as an important contributor to the pathogenesis of multiple diseases, including respiratory disorders. As research has progressed, other cell death modes have been explored in the context of respiratory diseases. Early studies focused on pyroptosis in immune cells like macrophages, but recent work highlights its occurrence in nonimmune cells, such as airway epithelial cells.7 This broader cellular distribution is particularly relevant to respiratory diseases involving epithelial barrier disruption and chronic inflammation. Moreover, members of the gasdermin family have been implicated in pyroptotic processes in both immune and nonimmune cells, including pulmonary and airway epithelial cells, further supporting the importance of pyroptosis in respiratory pathology.8
Compared with previous reviews on pyroptosis, the present narrative review places greater emphasis on recent mechanistic advances, disease-specific context across major respiratory disorders, and the translational implications of targeting pyroptosis-related pathways. In addition to summarizing the dual roles of pyroptosis in respiratory diseases, we particularly focus on emerging therapeutic strategies and the major bottlenecks that currently limit clinical translation. In this narrative review, we summarize the molecular mechanisms and regulatory networks of pyroptosis, discuss its context-dependent roles in major respiratory diseases, and highlight recent advances, therapeutic implications, and future research directions in this field.
Literature Search Strategy
For this narrative review, the literature was primarily retrieved from PubMed, Web of Science, and Scopus. The main search terms included “pyroptosis”, “gasdermin”, “inflammasome”, “respiratory diseases”, “pulmonary infection”, “acute lung injury”, “acute respiratory distress syndrome”, “asthma”, “COPD”, “pulmonary fibrosis”, and “lung cancer”. We mainly focused on English-language studies published in recent years, while also including representative earlier studies of particular mechanistic importance. The final selection of references was based on their relevance to the scope of this review and the authors’ judgment.
Molecular Mechanisms and Regulatory Networks of Pyroptosis
Pyroptosis is a form of programmed necrosis distinct from apoptosis, playing roles in development, homeostasis, and aging. It occurs in various cell types in vivo and is activated to eliminate pathogen-infected cells by releasing inflammatory cytokines that recruit monocytes to sites of injury.9 Characterized as a caspase-mediated PCD, pyroptosis defends cells against extracellular pathogens. Morphologically, pyroptotic cells exhibit swelling and blister-like protrusions, followed by plasma membrane rupture and the release of cytoplasmic contents.10 This contrasts with apoptosis, where the plasma membrane remains intact and apoptotic bodies form. Excessive pyroptosis can lead to inflammation and immune dysregulation. Caspase-1 dependence has been considered a key biological effector of pyroptosis, with membrane pore formation by the gasdermin family proteins being central to its mechanism.11 Depending on the activation pathway, pyroptosis can be classified into canonical and non-canonical routes, involving the release of inflammatory cytokines and cross-talk between signaling pathways.12
Two Principal Molecular Pathways of Pyroptosis
Pyroptosis involves two molecular pathways (Figure 1): the Caspase-1-dependent canonical pathway and the Caspase-1-independent non-canonical pathway. Both pathways release the proinflammatory cytokines IL-1 and IL-18, which stimulate local or systemic inflammation.13 Despite significant differences in activating triggers, key molecules, signaling mechanisms, and physiological and pathological roles, both pathways converge on a common endpoint: the activation and proteolytic cleavage of GSDM, leading to the formation of membrane pores.14
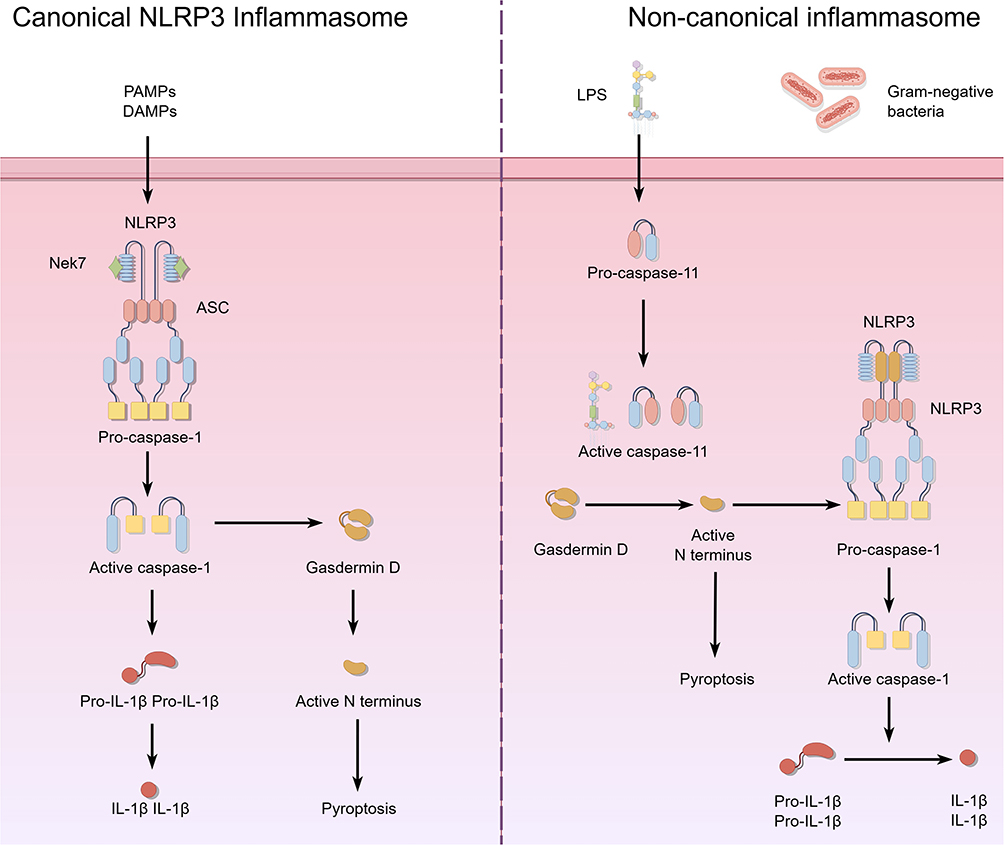 |
Figure 1 Two Principal Molecular Pathways of Pyroptosis. |
Canonical NLRP3 Inflammasome Pathway‐induced Pyroptosis
The canonical NLRP3 inflammation pathway activates pyroptosis by detecting danger signals and forming inflammasome complexes. These complexes subsequently activate inflammatory Caspase-1, leading to the cleavage of GSDM proteins and the execution of pyroptosis.13 The pathway’s main components include members of the PRR family, adaptor proteins, Caspase-1, and GSDMD. It unfolds in four stages: signal sensing, inflammasome assembly, Caspase-1 activation, and pyroptosis execution.15 Activation begins when pattern recognition receptors (PRRs) detect pathogen-associated molecular patterns (PAMPs) or DAMPs. The PRRs involved, such as NLRP1, NLRP3, AIM2, and Pyrin, belong to the NOD-like receptor family.16 Upon recognizing stimuli, PRRs interact with adaptor proteins through their domains to form inflammasome complexes. Most PRRs, like NLRP3, AIM2, and Pyrin, bind the PYD domain of the adaptor ASC (Apoptosis-associated Speck-like Protein Containing a CARD) and recruit pro-Caspase-1 via its CARD domain, assembling the complex independently of ASC.17 In contrast, NLRP1 directly recruits pro-Caspase-1 through its CARD domain, forming the complex without ASC.18 The inflammasome’s formation facilitates the autoproteolytic activation of pro-Caspase-1, resulting in the proteolytically active mature Caspase-1 (p20/p10) heterodimer. As the pathway’s main effector, activated Caspase-1 plays a crucial role in this process.15
Non-Canonical Inflammasome Pathway‐induced Pyroptosis
The non-canonical inflammasome pathway is directly activated by Gram-negative bacterial lipopolysaccharide (LPS). Unlike the Caspase-1 activation pathway, it does not require PRRs or the assembly of canonical inflammasome complexes, allowing for direct and rapid activation.19 This pathway plays a crucial role in host defense against cytosolic bacterial infections. Its core components include Caspase-4/5/11 and GSDMD. The activating signal, cytosolic LPS, originates from LPS released during the lysis of intracellular Gram-negative bacteria and LPS delivered into the cytosol by outer membrane vesicles (OMVs).20 Recent studies indicate that guanylate-binding proteins (GBPs) enhance the release of LPS from membranes, increasing the recognition efficiency of caspase-4/5/11 for LPS.21 Unlike caspase-1 in the canonical pathway, caspase-4/5/11 can directly bind the lipid A moiety of LPS through their CARD domains, leading to their oligomerization and autocatalytic activation without the need for adaptor proteins or inflammasome assembly.22 This direct activation enables cells to respond to cytosolic bacterial invasion by initiating pyroptosis to eliminate pathogens. Activated caspase-4/5/11 cleaves GSDMD at the same site as caspase-1 in the canonical pathway, releasing the GSDMD-N fragment.23 This fragment can form membrane pores on the plasma membrane through the same mechanism.
Core Differences and Synergistic Regulation Between the Two Main Pathways
The canonical and non-canonical pyroptotic pathways differ significantly in their activation triggers, key molecules, and signaling properties (Table 1). However, both pathways converge on the GSDMD execution step, a feature preserved through evolution. The canonical pathway relies on PRR-mediated inflammasome assembly and is activated by a wide array of signals, including PAMPs and DAMPs, which are involved in various physiological and pathological processes such as anti-infective immunity, autoinflammatory diseases, and tumor immunity.24 In contrast, the non-canonical pathway is directly and rapidly activated by cytosolic LPS, playing a crucial role in the swift defense against Gram-negative bacteria. However, excessive activation of this pathway can lead to severe complications like sepsis.25
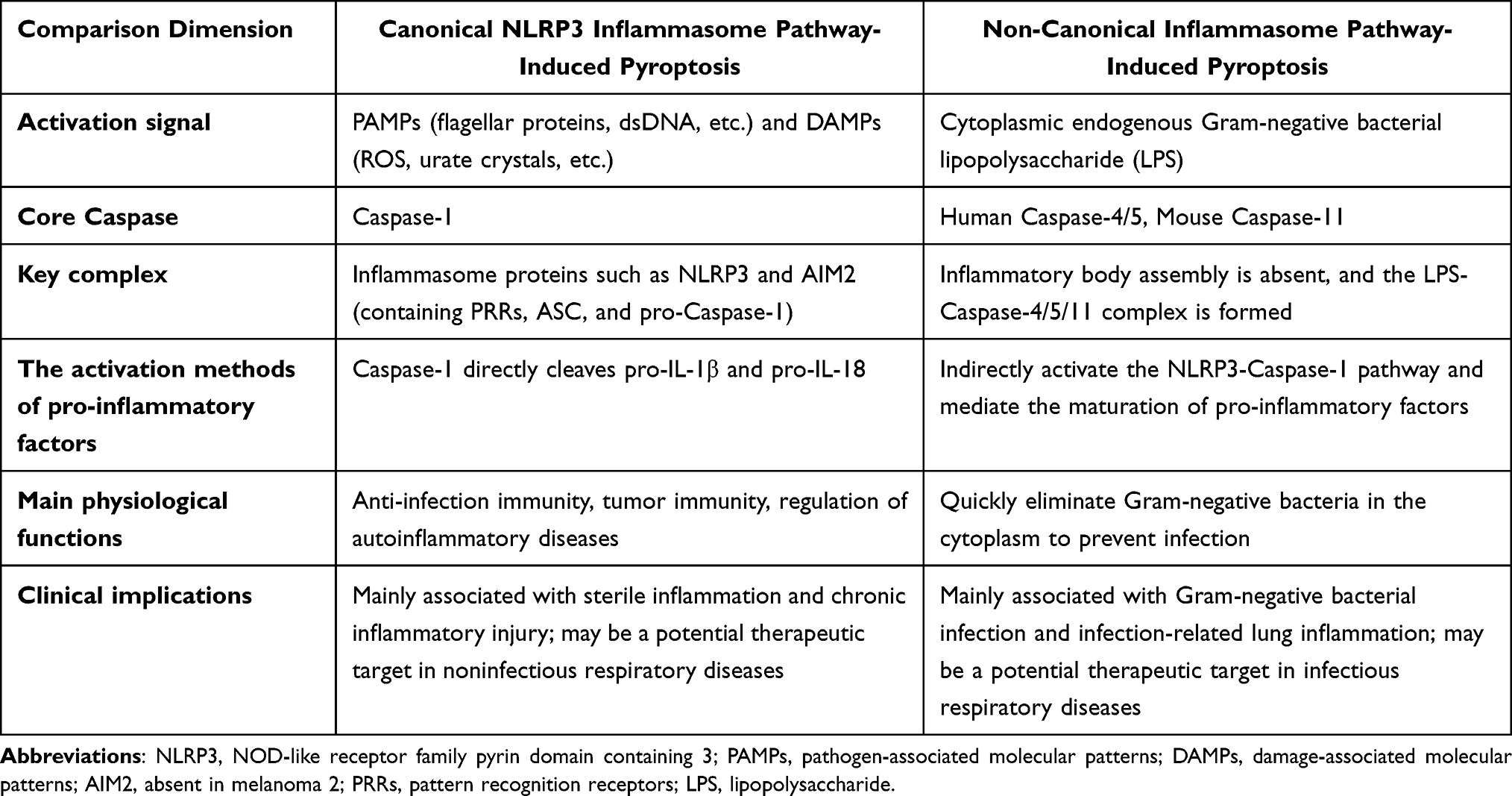 |
Table 1 Core Differences and Synergistic Regulation Between the Two Main Pathways |
Recent findings indicate that these two pathways are not entirely independent but rather work in tandem. For example, inflammation triggered by the activation of the non-canonical pathway can subsequently activate the NLRP3 inflammasome within the canonical pathway, further amplifying the inflammatory response.26 Additionally, caspase-1, activated by the canonical pathway, can cleave GSDMD, thereby enhancing the pyroptotic effects of the non-canonical pathway.27 This interaction enables the organism to modulate the intensity and spatial distribution of pyroptosis according to various pathological conditions, ensuring effective pathogen clearance while maintaining internal homeostasis.28
In summary, both canonical and non-canonical inflammasome-mediated pyroptotic pathways selectively respond to danger signals through distinct activation strategies.19 However, they converge on the activation and cleavage of GSDMD, forming a crucial component of the host’s immune defense network.29 Recent discoveries, such as the regulation of the non-canonical pathway by GBPs, the role of Ninjurin-1 in membrane rupture, and the cooperative interactions between pathways, have offered new insights into the physiological and pathological functions of pyroptosis.30 These findings lay the theoretical groundwork for developing therapeutics targeting pyroptotic pathways in respiratory diseases.
The Role of Pyroptosis in Respiratory System Diseases
The respiratory system serves as the body’s direct link to the external environment, making it particularly vulnerable to infections, physicochemical irritants, and dysregulated autoimmunity. Pyroptosis-mediated inflammation is crucial in the development and progression of various respiratory diseases.1 This inflammatory response often triggers a surge of proinflammatory agents, known as an “inflammatory storm,” which exacerbates pulmonary tissue damage. Consequently, the pyroptosis-associated regulatory axis presents potential therapeutic targets.31 The following sections outline the mechanistic role of pyroptosis in major respiratory diseases. For clarity, the key findings, major pyroptosis-related mechanisms, and potential clinical implications across these disease contexts are summarized in Table 2.
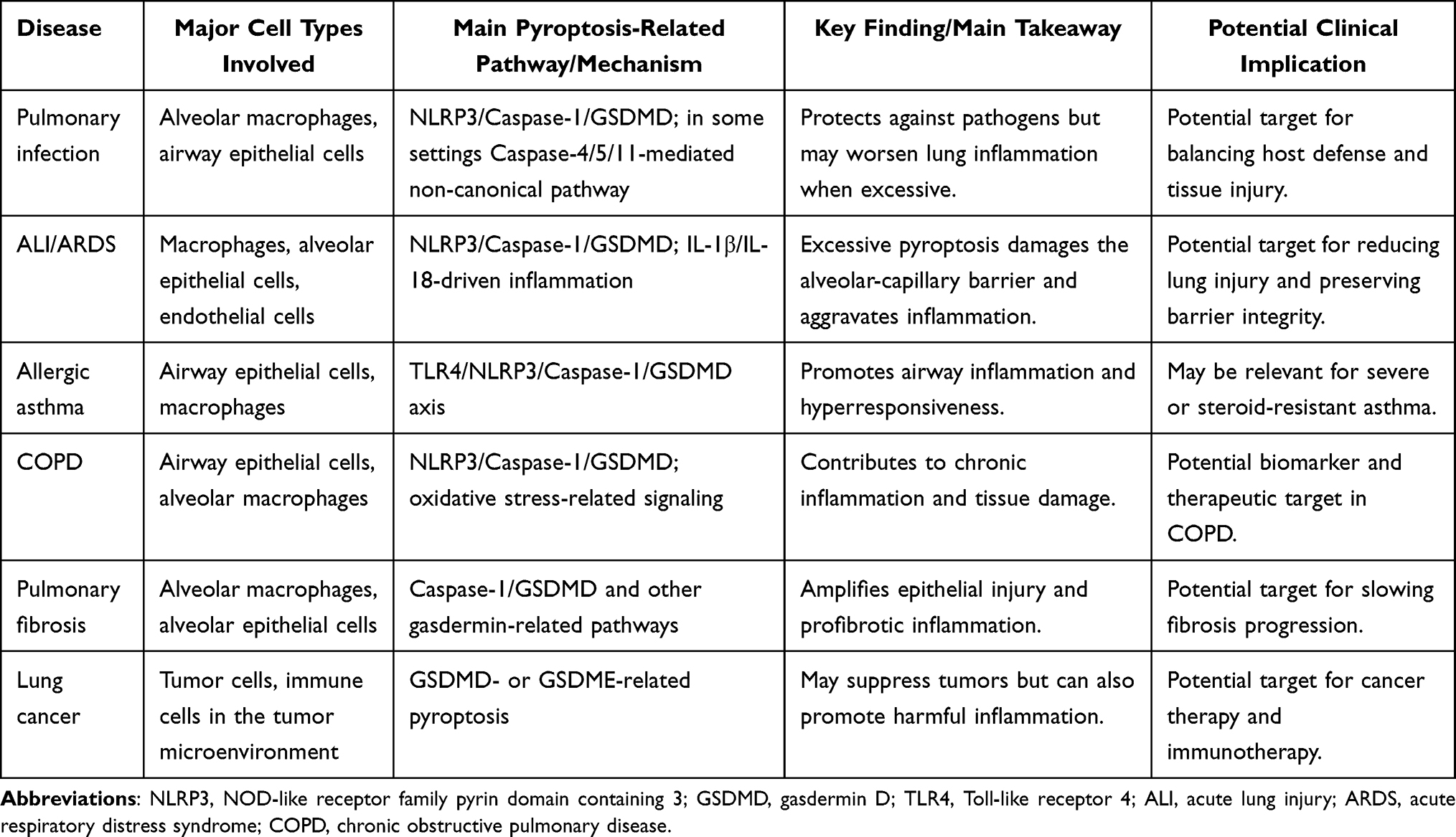 |
Table 2 Main Takeaways of Pyroptosis in Major Respiratory Diseases |
Pyroptosis and Pulmonary Infection
The lung serves as a barrier between organisms and the external environment, making it highly vulnerable to pathogen invasion, including bacteria and viruses.32 Pyroptosis, an inflammatory form of PCD mediated by the gasdermin family proteins, creates pores and releases proinflammatory mediators. This process acts as a “double-edged sword” in immune responses to pulmonary infections, where its dysregulation can directly affect infection outcomes.33 On the defensive side, pyroptosis is crucial for the innate immune system’s ability to clear pathogens. For instance, during Streptococcus pneumoniae invasion, macrophages recognize PAMPs, activating the NLRP3 inflammasome and the caspase-1–dependent canonical pyroptosis pathway.28 The lysis of infected cells eliminates bacteria from their replication niche, while IL-1, IL-18, and other proinflammatory cytokines recruit neutrophils, enhancing the anti-infective response.34 Similarly, Mycobacterium tuberculosis employs the ESX-1 secretion system to release PAMPs, activating NLRP3 and pyroptosis to limit pathogen spread.35 In viral infections like SARS-CoV-2, pyroptosis exposes viruses by lysing cells, facilitating clearance by extracellular immune effectors and forming an antiviral barrier.28 Additionally, caspase-1-mediated pyroptosis can modulate neutrophil necroptosis, further strengthening host defense against Escherichia coli pneumonia.36
Excessive or abnormal activation of pyroptosis exacerbates pulmonary injury and worsens infections. In SARS-CoV-2 infections, the virus activates the NLRP3 inflammasome through its nucleocapsid (N) protein or NSP6, leading to hyperactivation of pyroptosis in alveolar epithelium and macrophages.37 This results in a massive release of proinflammatory mediators, which can trigger a cytokine storm, compromise pulmonary parenchymal integrity, and aggravate COVID-19–associated ALI.38 Experimental data indicate that IL-6 deficiency enhances GSDMD/GSDME-mediated pyroptosis in S. pneumoniae infections, increasing lung inflammation and tissue damage.39 Conversely, exogenous IL-6 supplementation can mitigate infection by inhibiting pyroptosis. Pathogens can manipulate pyroptotic pathways to evade detection: SARS-CoV-2 NSP5 and ORF6 jointly suppress caspase-8 activity, blocking pyroptosis and promoting replication, while M. tuberculosis uses the PtpB protein to prevent GSDMD membrane localization, thereby inhibiting pyroptosis and sustaining infection.40 This host–pathogen tug-of-war over pyroptotic regulation amplifies the pathological impact of pyroptosis in pulmonary infections. From a clinical perspective, pyroptosis-related pathways may serve as potential biomarkers of inflammatory severity and therapeutic targets for balancing pathogen clearance with tissue injury during severe pulmonary infections. Recent studies further suggest that the role of pyroptosis in pulmonary infection is highly context-dependent, varying with pathogen type, host cell population, and the intensity of the inflammatory response, thereby providing new clues for more precise therapeutic modulation.
Pyroptosis and ALI/ARDS
ALI is a respiratory condition affecting the lungs and lung tissues, caused by various factors impacting the body and organs.41 It typically results from damage to the lung vasculature and alveolar epithelial cells due to inflammation and pulmonary edema. The most severe consequence of ALI is ARDS, characterized by life-threatening respiratory failure, skin discoloration, and hypoxemia.42 Both ALI and ARDS are severe inflammation-driven diseases, triggered by factors such as infection and trauma, with the primary pathological feature being the disruption of the alveolar-capillary barrier and uncontrolled inflammation.43 Pyroptosis, a regulator of inflammation, plays a “double-edged sword” role in the pathogenesis and progression of ALI/ARDS.44 Its activation and regulation of inflammation directly influence disease outcomes. Recent studies have elucidated its molecular mechanisms and regulatory networks.45 As a defensive mechanism, pyroptosis forms an anti-injury barrier by precisely modulating the inflammatory response.12 In the early stages of ALI/ARDS, innate immune cells like macrophages can activate pyroptotic pathways by recognizing DAMPs.44 This leads to the release of proinflammatory mediators such as IL-1 and IL-18, recruiting neutrophils and other immune effector cells to clear pathogens and necrotic tissue.46 Recent research indicates that RIOK2 is involved in the translocation of the FADD–RIPK1–Caspase-8 complex, activated by its ATPase activity.47 This activation leads to Caspase-8–mediated cleavage of GSDMD, generating pyroptotic responses that target infected cells, enhance host clearance of pathogens, and limit the spread of inflammation.48 Clinically, modulation of pyroptosis may represent a promising strategy to reduce excessive inflammatory injury and preserve alveolar-capillary barrier integrity in ALI/ARDS, although the optimal timing of intervention and appropriate patient stratification remain major challenges. Recent studies have further shown that pyroptosis in ALI/ARDS involves not only classical inflammasome activation but also extensive crosstalk with other regulated cell death pathways, suggesting that future therapeutic approaches may need to target multiple injury mechanisms rather than pyroptosis alone.
Pyroptosis and Allergic Asthma
Allergic asthma is characterized by an immune disorder involving excessive Th2 cell responses, airway hyperresponsiveness, and eosinophilic airway inflammation, with airway remodeling as its hallmark pathological feature.49 The NLRP3 inflammasome in the airway epithelium can be activated by environmental factors such as smoke, silica, and cigarettes, leading to epithelial injury by modulating immune responses.50 Moderate pyroptosis helps maintain immune homeostasis and constructs airway defense barriers by regulating immune responses.33 As a crucial component of innate immunity, pyroptosis limits excessive inflammatory activation by eliminating aberrantly activated immune cells or allergen complexes.51 Research indicates that macrophages can moderately activate caspase-1–mediated pyroptosis via the TLR4/IFN- pathway to regulate the Th1/Th2 immune balance and prevent an excess of inflammatory cells like eosinophils.52 However, excessive or aberrant activation of pyroptosis drives allergic asthma and is closely linked to steroid-resistant phenotypes. Airway epithelial pyroptosis is a key driver of allergic asthma pathologies.33 KLF5 can bind to the miR-182-5p promoter to repress its expression, thereby releasing the repression of TLR4, activating the NLRP3/caspase-1/GSDMD axis, and inducing epithelial cell pyroptosis. This process releases IL-1 and IL-18, causing airway inflammation and hyperresponsiveness. Silencing KLF5 significantly improves airway resistance and reduces histopathological injury in asthmatic mice.53 These findings suggest that pyroptosis-related signaling has important clinical relevance in asthma, not only for identifying distinct inflammatory phenotypes but also for developing targeted therapies, particularly in severe or steroid-resistant disease. Recent evidence further indicates that pyroptosis is not merely a downstream consequence of airway inflammation, but may also actively contribute to airway epithelial dysfunction and disease severity.
Pyroptosis and COPD
COPD is characterized by chronic airway inflammation, lung parenchyma destruction, and airway remodeling, with smoking as the primary cause of inflammation.54 Its pathogenesis is closely linked to dysregulated innate immune responses and imbalances in cell death.55 Moderate pyroptosis can delay COPD progression by maintaining immune homeostasis and tissue repair capacity.56 HDAC6, a gene associated with protective pyroptosis, is expressed at low levels in the lung tissues of COPD patients.57 It is inversely related to the extent of pulmonary tissue damage, suggesting its role in preserving airway epithelial barrier integrity by suppressing aberrant pyroptotic signaling pathways.58 In alveolar macrophages (AMs), pyroptosis regulation is subgroup-specific: M1-like AMs can resist oxidative stress-induced pyroptosis, thereby reducing inflammatory infiltration and maintaining phagocytic function.44 Moderately activated pyroptosis can eliminate damaged airway epithelial cells and aberrant immune complexes, preventing the persistent accumulation of inflammatory signals and forming an early immunological defense barrier in COPD.55 Chronic inflammation in inflammatory diseases, such as those triggered by inhaled noxious particles or gases (most commonly cigarette smoke), is driven by inflammation cascades in the airways.59 These cascades lead to the production of various cytokines, chemokines, and tissue-destroying enzymes that perpetuate chronic inflammation and tissue destruction.60 Smoking can trigger the release of interleukin-1 (IL-1) and interleukin-18 (IL-18) in the lungs of COPD patients, further inducing inflammation and tissue destruction.61 In COPD, pyroptosis-associated molecules may serve as potential biomarkers of disease activity and progression, as well as therapeutic targets for reducing chronic airway inflammation and tissue injury. Recent evidence further indicates that pyroptosis links oxidative stress, epithelial damage, and persistent immune activation in COPD, thereby offering additional mechanistic insight into disease progression and potential intervention strategies.
Pyroptosis and PF
PF is a chronic, progressive interstitial lung disease characterized by the disruption of the alveolar epithelium, aberrant fibroblast activation, and excessive extracellular matrix accumulation.62 It is closely linked to the dysregulation of regulated cell death (RCD) mechanisms. Pyroptosis, an inflammatory form of RCD mediated by gasdermin family proteins, plays a complex role in PF, acting as a “double-edged sword”.63 On the positive side, moderate pyroptosis helps maintain pulmonary tissue homeostasis by regulating inflammation and repair processes.12 In the early stages of PF, alveolar macrophages may activate pyroptosis pathways to clear damaged cells and abnormal complexes.64 This activation helps balance pro- and anti-inflammatory mediator release, preventing excessive inflammation.65 Research indicates that let-7a-5p can inhibit excessive pyroptosis by targeting MYD88.30 Physiologically, pyroptosis promotes efferocytosis by modulating macrophage polarization, reducing inflammatory debris, and creating a microenvironment conducive to alveolar epithelial repair, thereby delaying fibrosis onset.66 However, overly active pyroptosis and aberrant pathway signaling are key pathogenic drivers in PF, influenced by cell type specificity and pathway interactions. In silica-induced silicosis, alveolar macrophages and epithelial cells synergistically activate the canonical Caspase-1/GSDMD and non-canonical Caspase-3/8/GSDMME pyroptotic pathways.67 This activation releases proinflammatory cytokines IL-1 and IL-18, amplifying inflammatory cascades, directly damaging the alveolar epithelial barrier, and inducing fibrosis.68 From a translational perspective, targeting dysregulated pyroptosis may offer a potential strategy to attenuate epithelial injury, inflammatory amplification, and fibrotic progression in pulmonary fibrosis. Recent studies further indicate that pyroptosis in pulmonary fibrosis is highly cell-type-specific and may serve as a mechanistic link between epithelial damage, macrophage-driven inflammation, and fibrotic remodeling, thereby providing new insights for anti-fibrotic therapy.
Pyroptosis and Lung Cancer
Lung cancer is the deadliest cancer worldwide, characterized by abnormal tumor cell growth and changes in the tumor immune microenvironment (TME).69 Pyroptosis, an inflammatory form of PCD mediated by the Gasdermin family, plays a crucial role in regulating tumor cell fate and immune responses.70 Research has identified subtype-specific regulatory networks of pyroptosis and its interactions, whether synergistic or antagonistic, with radiotherapy and chemotherapy.71 When properly regulated at tumor-suppressive levels, pyroptosis can directly eliminate tumor cells and activate antitumor immunity, forming a defensive barrier.72 In non-small cell lung cancer (NSCLC), the downregulation of the long noncoding RNA XIST disrupts the sequestration of miR-335, thereby reducing superoxide dismutase 2 (SOD2) expression, increasing reactive oxygen species (ROS), and inducing GSDMD-mediated pyroptosis, which significantly inhibits tumor cell proliferation.73 Furthermore, pyroptosis can trigger antitumor immunity through immunogenic cell death (ICD).74 The DNA demethylating agent decitabine and doxorubicin reverse the methylation-dependent silencing of GSDME in lung cancer cells, where caspase-3 cleavage of GSDME induces pyroptosis, releasing inflammatory mediators that recruit effector cells and promote immune memory.75 Conversely, aberrant activation or dysregulation of pyroptosis can drive lung cancer progression and reduce therapeutic effectiveness.76 Cells releasing circulating cell-free DNA (CCFs) activate the ubiquitin-editing enzyme A20, which stabilizes the NLRP3 inflammasome; proinflammatory cytokines such as IL-1 can enhance stem-like properties in neighboring senescent cells.77 In lung cancer, pyroptosis may have important clinical relevance not only as a mechanism of tumor suppression, but also as a potential means of enhancing antitumor immunity and improving the efficacy of existing therapies. Recent studies further suggest that pyroptosis can influence both tumor cell death and the immune landscape of the tumor microenvironment, thereby affecting responsiveness to antitumor treatment and underscoring its complex translational potential.
Clinical Challenges and Limitations of Targeting Pyroptosis for the Treatment of Respiratory Diseases
Recent progress has identified several potential therapeutic strategies for modulating pyroptosis in respiratory diseases, including inhibition of inflammasome activation, suppression of caspase-mediated gasdermin cleavage, and regulation of upstream inflammatory signaling pathways. Experimental studies have suggested that these approaches may reduce inflammatory injury, limit tissue damage, and improve disease outcomes in different respiratory contexts. However, despite these promising advances, the clinical application of pyroptosis-targeted therapy remains in an early stage.
While targeting pyroptosis offers a novel approach for the precise treatment of respiratory diseases, its widespread application is significantly constrained by its “double-edged sword” nature, complex regulatory pathways, and translational challenges.78 Recent studies have elucidated the molecular mechanisms underlying these limitations and clinical challenges, aiming to pinpoint the bottlenecks that must be addressed in future research.79
Firstly, addressing pathway crosstalk presents significant challenges. Pyroptosis is not isolated; it forms intricate networks with ferroptosis, necroptosis, and NET formation.80 Consequently, interventions targeting a single pathway often result in compensatory inflammatory activation or functional disruption.81 In ALI, double-stranded DNA from NETs can engage the cGAS–STING axis, collaborate with the AIM2 inflammasome, and trigger an “inflammation–pyroptosis” positive-feedback loop while simultaneously promoting ferroptosis and necroptosis.82 Although blocking GSDMD can suppress pyroptosis, it fails to halt the combined tissue-death effects of multiple cell-death mechanisms.83 Secondly, maintaining immune homeostasis is challenging. Targeting pyroptosis can enhance the efficacy of anti–PD-L1 antibodies, but excessive modulation may result in CIP, while inhibiting pyroptosis could reduce immunotherapy sensitivity, creating regulatory issues.84 Lastly, clinical translation evidence is limited, and disease heterogeneity is pronounced. Current research is confined to cell and animal models, such as FMN@BSA NPs validated only in a bleomycin-induced PF model, and lacks support from large-scale multicenter clinical trials.79 Pyroptosis regulation is closely linked to xCT-mediated lipid peroxidation, while asthma regulation is primarily influenced by the KLF5/miR-182-5p/TLR4 axis, indicating that a single intervention strategy may not suit all disease subtypes.7,10
Limitations
As a narrative review, this review has several inherent limitations. Unlike a systematic review, it did not apply predefined inclusion and exclusion criteria or a formal quality assessment framework. The selection and interpretation of the literature were based on the authors’ judgment and the relevance of individual studies to the scope of this review.
Conclusion
Pyroptosis has gained attention as a potential therapeutic target for respiratory diseases, yet it is aptly described as a “double-edged sword.” On one hand, the inflammatory mediators released during pyroptosis boost the immune system’s ability to combat pathogens and inhibit the proliferation and motility of lung cancer cells by inducing pyroptotic cell death.70 On the other hand, excessive activation of pyroptosis can result in inflammation, lung injury, and even lung fibrosis and tumor progression.33 To date, two signaling pathways have been identified: the canonical caspase-1–dependent pathway and the non-canonical caspase-4/5/11–mediated alternative pathway. Recent studies indicate that pyroptosis influences the progression of pulmonary infections,13 ALI/ARDS,85 allergic asthma,33 COPD,86 PF,55 and lung cancer.70 Taken together, these findings highlight pyroptosis as an important mechanistic link between inflammation, immune defense, and tissue injury in respiratory diseases.
Future Perspectives
Despite these advances, our understanding of pyroptosis in respiratory diseases remains incomplete. First, the molecular regulation of pyroptosis requires further clarification. Although the canonical and non-canonical pathways have been identified, the upstream triggers, cell type-specific regulators, and context-dependent differences in pyroptotic signaling remain poorly understood. In addition, the interactions between pyroptosis and other forms of regulated cell death, such as apoptosis, necroptosis, and ferroptosis, deserve further investigation, as these processes may jointly influence the severity and outcome of respiratory diseases.33,55
Second, the identification of novel and reliable biomarkers is an important direction for future research. Molecules related to inflammasome activation, caspase cleavage, gasdermin family proteins, and pyroptosis-associated cytokines may serve as candidate biomarkers for disease diagnosis, severity stratification, prognosis evaluation, and therapeutic monitoring.13,85 However, their specificity, sensitivity, and clinical applicability still need to be validated in large-scale human studies.
Third, there is an urgent need to develop targeted interventions that modulate pyroptosis in a balanced and disease-specific manner. Because pyroptosis can exert both protective and detrimental effects, therapeutic strategies should not aim for indiscriminate suppression, but rather for precise regulation according to disease stage and pathological context. Potential approaches may include selective inhibition of inflammasome activation, modulation of caspase activity, targeting gasdermin-mediated membrane pore formation, and combining pyroptosis-related therapies with anti-inflammatory, anti-fibrotic, or anti-tumor treatments.70,85 In parallel, improving targeted drug delivery and reducing off-target effects will also be critical for future clinical translation.
Finally, more translational and clinical studies are needed to bridge the gap between experimental findings and practical application. Future research should aim to determine when and how modulation of pyroptosis can prevent lung injury, delay disease progression, and improve outcomes in patients with respiratory disorders. Therefore, achieving a therapeutic balance in regulating pyroptosis should remain a central goal in this field.
Data Sharing Statement
There are no data and no material associated with this paper.
Ethics Approval and Consent to Participate
There is no human subject, and this is a review, so there is no need for ethical approval and consent.
Author Contributions
Chen Yu and Caijun Dai; Conceptualization, Writing - original draft. Lujian Zhu; Conceptualization, Review - original draft. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This review was supported by Jinhua Municipal Science and Technology Bureau Key Project(No. 2023-3-091) and Jinhua Municipal Central Hospital Science and Technology Project (No. JY2023-1-04).
Disclosure
The authors declare no competing interest.
References
1. Basil MC, Alysandratos K-D, Kotton DN, Morrisey EE. Lung repair and regeneration: advanced models and insights into human disease. Cell Stem Cell. 2024;31(4):439–12. doi:10.1016/j.stem.2024.02.009
2. Radhouani M, Farhat A, Hakobyan A, et al. Eosinophil innate immune memory after bacterial skin infection promotes allergic lung inflammation. Sci Immunol. 2025;10(106). doi:10.1126/sciimmunol.adp6231
3. Saki N, Javan M, Moghimian-Boroujeni B, Kast RE. Interesting effects of interleukins and immune cells on acute respiratory distress syndrome. Clin Exp Med. 2023;23(7):2979–2996. doi:10.1007/s10238-023-01118-w
4. Zhai Y, Zhu C, Zhu T, et al. Global, regional, and national burden of chronic respiratory diseases,1990-2021 and predictions to 2035: analysis of data from the global burden of disease study 2021. Ann Med. 2025;57(1). doi:10.1080/07853890.2025.2530225
5. Jamora RDG, Albay AB, Ditching MBDF. Clinical outcomes of COVID-19 infection among patients with chronic obstructive pulmonary disease: findings from the philippine corona study. Clin Pract. 2023;13(6):1383–1392. doi:10.3390/clinpract13060124
6. Zhu L, Hu M, Xu H. The key players of inflammasomes and pyroptosis in sepsis-induced pathogenesis and organ dysfunction. Front Pharmacol. 2025;16. doi:10.3389/fphar.2025.1586364
7. Wu H, Wang H, Gao P, Ouyang S. Pyroptosis: induction and inhibition strategies for immunotherapy of diseases. Acta Pharm Sin B. 2024;14(10):4195–4227. doi:10.1016/j.apsb.2024.06.026
8. Liang X, Qin Y, Wu D, Wang Q, Wu H. Pyroptosis: a double-edged sword in lung cancer and other respiratory diseases. Cell Commun Signal. 2024;22(1). doi:10.1186/s12964-023-01458-w
9. Yang S, Zou Y, Zhong C, Zhou Z, Peng X, Tang C. Dual role of pyroptosis in liver diseases: mechanisms, implications, and therapeutic perspectives. Front Cell Dev Biol. 2025;13:1522206.
10. Broz P. Pyroptosis: molecular mechanisms and roles in disease. Cell Res. 2025;35(5):334–344. doi:10.1038/s41422-025-01107-6
11. Pan Y, Cai W, Huang J. Pyroptosis in development, inflammation and disease. Front Immunol. 2022;13. doi:10.3389/fimmu.2022.991044
12. Chai R, Li Y, Shui L, Ni L, Zhang A. The role of pyroptosis in inflammatory diseases. Front Cell Dev Biol. 2023;11. doi:10.3389/fcell.2023.1173235
13. Shang P, Gan M, Wei Z, et al. Advances in research on the impact and mechanisms of pathogenic microorganism infections on pyroptosis. Front Microbiol. 2024;15. doi:10.3389/fmicb.2024.1503130
14. Kappelhoff S, Margheritis EG, Cosentino K. New insights into Gasdermin D pore formation. Biochem Soc Trans. 2024;52(2):681–692. doi:10.1042/BST20230549
15. Nadella V, Kanneganti T-D. Inflammasomes and their role in PANoptosomes. Curr Opin Immunol. 2024;91:102489. doi:10.1016/j.coi.2024.102489
16. Navuluri N, Yata VK, Dachani SR, Rachakonda K, Kolliputi N. Heme unleashed: NLRP12 orchestrates PANoptosis in a symphony of cell fate. Cell Biochem Biophys. 2025;83(3):2727–2733. doi:10.1007/s12013-025-01712-7
17. Fu J, Wu H. Structural Mechanisms of NLRP3 Inflammasome Assembly and Activation. Annu Rev Immunol. 2023;41(1):301–316. doi:10.1146/annurev-immunol-081022-021207
18. Xu Z, Zhou Y, Liu M, et al. Homotypic CARD-CARD interaction is critical for the activation of NLRP1 inflammasome. Cell Death Dis. 2021;12(1):57.
19. Shi Y, Blander JM. Patterns of bacterial viability governing noncanonical inflammasome activation. Curr Opin Immunol. 2024;92. doi:10.1016/j.coi.2024.102512
20. Wright SS, Vasudevan SO, Rathinam VA. Mechanisms and consequences of noncanonical inflammasome-mediated pyroptosis. J Mol Biol. 2021;434(4). doi:10.1016/j.jmb.2021.167245
21. Kumar R, Kushawaha PK. Interferon inducible guanylate-binding protein 1 modulates the lipopolysaccharide-induced cytokines/chemokines and mitogen-activated protein kinases in macrophages. Microbiol Immunol. 2024;68(5):185–195. doi:10.1111/1348-0421.13123
22. Elkayam E, Gervais FG, Wu H, Crackower MA, Lieberman J. New insights into the noncanonical inflammasome point to caspase-4 as a druggable target. Nat Rev Immunol. 2025;25(8):558–568. doi:10.1038/s41577-025-01142-9
23. Zhang N, Zhang J, Yang Y. A palmitoylation-depalmitoylation relay spatiotemporally controls GSDMD activation in pyroptosis. Nat Cell Biol. 2024;26(5):757–769. doi:10.1038/s41556-024-01397-9
24. Vajjhala PR, Ve T, Bentham A, Stacey KJ, Kobe B. The molecular mechanisms of signaling by cooperative assembly formation in innate immunity pathways. Mol Immunol. 2017;86:23–37. doi:10.1016/j.molimm.2017.02.012
25. BaSAK B, Sachiko A-T. IRF3 function and immunological gaps in sepsis. Front Immunol. 2024;15. doi:10.3389/fimmu.2024.1336813
26. Chen M, Yu S, Gao Y. TRAF6-TAK1-IKKβ pathway mediates TLR2 agonists activating “one-step” NLRP3 inflammasome in human monocytes. Cytokine. 2023;169:156302. doi:10.1016/j.cyto.2023.156302
27. Li X, Zhang T, Kang L. Apoptotic caspase-7 activation inhibits non-canonical pyroptosis by GSDMB cleavage. Cell Death Differ. 2023;30(9):2120–2134. doi:10.1038/s41418-023-01211-3
28. Zhang Y, Zhao D, Wang T. Pyroptosis, a double-edged sword during pathogen infection: a review. Cell Death Discov. 2025;11(1). doi:10.1038/s41420-025-02579-6
29. Li Y, Gao W, Qiu Y, et al. RING1 dictates GSDMD-mediated inflammatory response and host susceptibility to pathogen infection. Cell Death Differ. 2025;32(11):2066–2077. doi:10.1038/s41418-025-01527-2
30. Zhou YR, Dang -J-J, Yang Q-C, Sun Z-J. The regulation of pyroptosis by post-translational modifications: molecular mechanisms and therapeutic targets. EBioMedicine. 2024;109:105420. doi:10.1016/j.ebiom.2024.105420
31. Albright JM, Davis CS, Bird MD. The acute pulmonary inflammatory response to the graded severity of smoke inhalation injury. Crit Care Med. 2011;40(4).
32. Kageyama T, Ito T, Tanaka S, Nakajima H. Physiological and immunological barriers in the lung. Semin Immunopathol. 2024;45(4–6):533–547. doi:10.1007/s00281-024-01003-y
33. Hao Y, Wang W, Zhang L, Li W. Pyroptosis in asthma: inflammatory phenotypes, immune and non-immune cells, and novel treatment approaches. Front Pharmacol. 2024;15. doi:10.3389/fphar.2024.1452845
34. Jiang M, Chen Z-G, Li H. Succinate and inosine coordinate innate immune response to bacterial infection. PLoS Pathog. 2022;18(8):e1010796. doi:10.1371/journal.ppat.1010796
35. Pattanaik KP, Ganguli G, Naik SK, Sonawane A. Mycobacterium tuberculosis EsxL induces TNF-α secretion through activation of TLR2 dependent MAPK and NF-κB pathways. Mol Immunol. 2021;130:133–141. doi:10.1016/j.molimm.2020.11.020
36. Luo Q, Shen L, Yang S. Caspase-1-licensed pyroptosis drives dsRNA-mediated necroptosis and dampens host defense against bacterial pneumonia. PLoS Pathog. 2025;21(5):e1013167. doi:10.1371/journal.ppat.1013167
37. Huang X, Zhu W, Zhang H, Qiu S, Shao H. SARS-CoV-2 N protein induces alveolar epithelial apoptosis via NLRP3 pathway in ARDS. Int Immunopharmacol. 2024;144:113503.
38. Guan T, Zhou X, Zhou W, Lin H. Regulatory T cell and macrophage crosstalk in acute lung injury: future perspectives. Cell Death Discov. 2023;9(1). doi:10.1038/s41420-023-01310-7
39. Gou X, Xu W, Liu Y. IL-6 prevents lung macrophage death and lung inflammation injury by inhibiting. Microbiol Spectr. 2022;10(2). doi:10.1128/spectrum.02049-21
40. Bader SM, Scherer L, Schaefer J, et al. IL-1β drives SARS-CoV-2-induced disease independently of the inflammasome and pyroptosis signalling. Cell Death Differ. 2025;32(7):1353–1366. doi:10.1038/s41418-025-01459-x
41. Liu H, Dong J, Xu C, et al. Acute lung injury: pathogenesis and treatment. J Transl Med. 2025;23(1):926.
42. Jones JH, Minshall RD. Endothelial transcytosis in acute lung injury: emerging mechanisms and therapeutic approaches. Front Physiol. 2022;13. doi:10.3389/fphys.2022.828093
43. Zhu Z, Zhang Y, Chen H, Zhang H. Cell-cell crosstalk in the pathogenesis of acute lung injury and acute respiratory distress syndrome. Tissue Barriers. 2025;13(3). doi:10.1080/21688370.2025.2452082
44. Cai Y, Shang L, Zhou F, et al. Macrophage pyroptosis and its crucial role in ALI/ARDS. Front Immunol. 2025;16:1530849.
45. Saleh HA, Yousef MH, Abdelnaser A. The anti-inflammatory properties of phytochemicals and their effects on epigenetic mechanisms involved in TLR4/NF-κB-mediated inflammation. Front Immunol. 2021;12. doi:10.3389/fimmu.2021.606069
46. Ginsburg I, Korem M, Koren E, Varani J. Pro-inflammatory agents released by pathogens, dying host cells, and neutrophils act synergistically to destroy host tissues: a working hypothesis. J Inflamm Res. 2019;12:35–47. doi:10.2147/JIR.S190007
47. Shen S, Lu C, Ling T, Zheng Y. Current advances on RIPK2 and its inhibitors in pathological processes: a comprehensive review. Front Mol Neurosci. 2025;18. doi:10.3389/fnmol.2025.1492807
48. Zhang N, Xu D. Controlling pyroptosis through post-translational modifications of gasdermin D. Dev Cell. 2025;60(7):994–1007. doi:10.1016/j.devcel.2025.02.005
49. Huang C, Fengqi L, Jian W, Zhigang T. Innate-like Lymphocytes and Innate Lymphoid Cells in Asthma. Clin Rev Allergy Immunol. 2019;59(3):359–70.
50. Zhou H, Zhang Q, Huang W. NLRP3 inflammasome mediates silica-induced lung epithelial injury and aberrant regeneration in lung stem/progenitor cell-derived organotypic models. Int J Biol Sci. 2023;19(6):1875–1893. doi:10.7150/ijbs.80605
51. Oh C, Spears TJ, Aachoui Y. Inflammasome-mediated pyroptosis in defense against pathogenic bacteria. Immunol Rev. 2024;329(1). doi:10.1111/imr.13408
52. Sordi MB, Cabral da Cruz AC, Panahipour L, Gruber R. Enamel matrix derivative decreases pyroptosis-related genes in macrophages. Int J Mol Sci. 2022;23(9):5078. doi:10.3390/ijms23095078
53. Lin Z, Bao R, Niu Y, Kong X. KLF5-mediated pyroptosis of airway epithelial cells leads to airway inflammation in asthmatic mice through the miR-182-5p/TLR4 axis. Mol Immunol. 2024;170:9–18. doi:10.1016/j.molimm.2024.03.007
54. Xu J, Zeng Q, Li S, Su Q, Fan H. Inflammation mechanism and research progress of COPD. Front Immunol. 2024;15:1404615.
55. Liu Y, Wang D, Liu X. Biological and pharmacological roles of pyroptosis in pulmonary inflammation and fibrosis: recent advances and future directions. Cell Commun Signal. 2024;22(1). doi:10.1186/s12964-024-01966-3
56. Liu Y, Pan R, Ouyang Y, et al. Pyroptosis in health and disease: mechanisms, regulation and clinical perspective. Signal Transduct Target Ther. 2024;9(1):245.
57. Xie T, Yang Z, Xian S, Lin Q, Huang L, Ding Y. Hsa_circ_0008833 promotes COPD progression via inducing pyroptosis in bronchial epithelial cells. Exp Lung Res. 2024;50(1):1–14. doi:10.1080/01902148.2024.2303474
58. Ren C, Wang Q, Fan S, Mi T, Zhang Z, He D. Toll-like receptor 9 aggravates pulmonary fibrosis by promoting NLRP3-mediated pyroptosis of alveolar epithelial cells. Inflammation. 2024;47(5):1744–1761. doi:10.1007/s10753-024-02006-5
59. Brusselle GG, Joos GF, Bracke KR. New insights into the immunology of chronic obstructive pulmonary disease. Lancet. 2011;378(9795):1015–1026. doi:10.1016/S0140-6736(11)60988-4
60. Sprague AH, Khalil RA. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem Pharmacol. 2009;78(6):539–552. doi:10.1016/j.bcp.2009.04.029
61. Le Rouzic O, Pichavant M, Frealle E, Guillon A, Si-Tahar M, Gosset P. Th17 cytokines: novel potential therapeutic targets for COPD pathogenesis and exacerbations. Eur Respir J. 2017;50(4):1602434. doi:10.1183/13993003.02434-2016
62. Huang J, Qin J, Zhu Y, Shen A. The role of epigenetics in pulmonary fibrosis: recent advances in mechanistic insights and therapeutic implications. Front Mol Biosci. 2025;12. doi:10.3389/fmolb.2025.1647300
63. Min R, Bai Y, Wang N-R, Liu X. Gasdermins in pyroptosis, inflammation, and cancer. Trends Mol Med. 2025;31(9):860–875. doi:10.1016/j.molmed.2025.04.003
64. Yang R, Yang Y, Yiping L, Xiaoshu L. Regulation of alveolar macrophage death in pulmonary fibrosis: a review. Apoptosis. 2023;28:1505–1519. doi:10.1007/s10495-023-01888-4
65. Gallo CG, Fiorino S, Posabella G, et al. The function of specialized pro-resolving endogenous lipid mediators, vitamins, and other micronutrients in the control of the inflammatory processes: possible role in patients with SARS-CoV-2 related infection. Prostaglandins Other Lipid Mediat. 2022;159:106619. doi:10.1016/j.prostaglandins.2022.106619
66. Hu Y, Huang Y, Guo J, Liu X, Liu Y. Targeting pyroptosis in myocardial inflammation and fibrosis: molecular mechanisms and therapeutic strategies. Apoptosis. 2025;30(9–10):1989–2007. doi:10.1007/s10495-025-02151-8
67. Song M, Wang J, Sun Y, et al. Inhibition of gasdermin D-dependent pyroptosis attenuates the progression of silica-induced pulmonary inflammation and fibrosis. Acta Pharm Sin B. 2022;12(3):1213–1224. doi:10.1016/j.apsb.2021.10.006
68. Ciminieri C, Woest ME, Reynaert NL. IL-1β induces a proinflammatory fibroblast microenvironment that impairs lung progenitors’ function. Am J Respir Cell Mol Biol. 2023;68(4):444–455. doi:10.1165/rcmb.2022-0209OC
69. Dan A, Burtavel L-M, Coman M-C, et al. Genetic blueprints in lung cancer: foundations for targeted therapies. Cancers. 2024;16(23):4048. doi:10.3390/cancers16234048
70. Wang J, Su H, Wang M, Ward R, An S, Xu TR. Pyroptosis and the fight against lung cancer. Med Res Rev. 2024;45(1):5–28.
71. Zhang RN, Jing Z-Q, Zhang L, Sun Z-J. Epigenetic regulation of pyroptosis in cancer: molecular pathogenesis and targeting strategies. Cancer Lett. 2023;575:216413. doi:10.1016/j.canlet.2023.216413
72. Bao X, Sun M, Meng L, Zhang H, Yi X, Zhang P. Applications of pyroptosis activators in tumor immunotherapy. Mater Today Bio. 2024;28. doi:10.1016/j.mtbio.2024.101191
73. Li J, Che L, Xu C. XIST/miR-34a-5p/PDL1 axis regulated the development of lung cancer cells and the immune function of CD8 + T cells. J Recept Signal Transduction Res. 2022;42(5):469–478. doi:10.1080/10799893.2021.2019274
74. Huang Y, Wang C, Chen Y, Wang D, Yao D. Nanomedicine-induced pyroptosis for anti-tumor immunotherapy: mechanism analysis and application prospects. Acta Pharm Sin B. 2025;15(7):3487–3510. doi:10.1016/j.apsb.2025.05.021
75. Pan J, Li Y, Gao W. Transcription factor Sp1 transcriptionally enhances GSDME expression for pyroptosis. Cell Death Dis. 2024;15(1). doi:10.1038/s41419-024-06455-6
76. Meybodi SM, Ejlalidiz M, Manshadi MR. Crosstalk between hypoxia-induced pyroptosis and immune escape in cancer: from mechanisms to therapy. Crit Rev Oncol Hematol. 2024;197:104340. doi:10.1016/j.critrevonc.2024.104340
77. Salminen A. Cooperation between inhibitory immune checkpoints of senescent cells with immunosuppressive network to promote immunosenescence and the aging process. Ageing Res Rev. 2025;106:102694. doi:10.1016/j.arr.2025.102694
78. Miao R. Manipulation of cancer cell pyroptosis for therapeutic approaches: challenges and opportunities. Biomark Res. 2025;13(1).
79. Khan S, Gull A, Akhtar M. Explicit analysis of in vivo, meterological and statistical hurdles in successful clinical translation of targeted nanomedicines and plausible remedial strategies. Expert Opin Drug Deliv. 2025;22(11):1769–1791. doi:10.1080/17425247.2025.2556979
80. Li P, Gao Y, Tao Z, Mu Z, Du S, Zhao X. PANoptosis: cross-Talk Among Apoptosis, Necroptosis, and Pyroptosis in Neurological Disorders. J Inflamm Res. 2025;18:8131–40.
81. Heng CKM, Gilad N, Darlyuk-Saadon I, Wong WSF, Engelberg D. Targeting the p38α pathway in chronic inflammatory diseases: could activation, not inhibition, be the appropriate therapeutic strategy? Pharmacol Ther. 2022;235:108153. doi:10.1016/j.pharmthera.2022.108153
82. Gu W, Zeng Q, Wang X, Jasem H, Ma L. Acute lung injury and the NLRP3 inflammasome. J Inflamm Res. 2024;17:3801–3813. doi:10.2147/JIR.S464838
83. Li Y, Guo B. GSDMD-mediated pyroptosis: molecular mechanisms, diseases and therapeutic targets. Mol Biomed. 2025;6(1). doi:10.1186/s43556-025-00249-8
84. Hou Y, Li W, Yang J. Is pyroptosis a brake or an accelerator in the fate of the tumor? Cell Death Dis. 2025;16(1). doi:10.1038/s41419-025-07866-9
85. Xiao J, Wang L, Zhang B, Hou A. Cell death in acute lung injury: caspase-regulated apoptosis, pyroptosis, necroptosis, and PANoptosis. Front Pharmacol. 2025;16. doi:10.3389/fphar.2025.1559659
86. Shu H-M, Lin C-Q, He B. Pyroptosis-related genes as diagnostic markers in chronic obstructive pulmonary disease and its correlation with immune infiltration. Int J Chron Obstruct Pulmon Dis. 2024;19:1491–1513. doi:10.2147/COPD.S438686
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Extracellular Histones Activate Endothelial NLRP3 Inflammasome and are Associated with a Severe Sepsis Phenotype

Beltrán-García J, Osca-Verdegal R, Pérez-Cremades D, Novella S, Hermenegildo C, Pallardó FV, García-Giménez JL
Journal of Inflammation Research 2022, 15:4217-4238
Published Date: 25 July 2022
Pyroptosis and Intervertebral Disc Degeneration: Mechanistic Insights and Therapeutic Implications
Ge Y, Chen Y, Guo C, Luo H, Fu F, Ji W, Wu C, Ruan H
Journal of Inflammation Research 2022, 15:5857-5871
Published Date: 17 October 2022
Pyroptosis and Inflammasome-Related Genes-NLRP3, NLRC4 and NLRP7 Polymorphisms Were Associated with Risk of Lung Cancer
Jing X, Yun Y, Ji X, Yang E, Li P
Pharmacogenomics and Personalized Medicine 2023, 16:795-804
Published Date: 25 August 2023
The Mechanism of Pyroptosis and Its Application Prospect in Diabetic Wound Healing
Al Mamun A, Shao C, Geng P, Wang S, Xiao J
Journal of Inflammation Research 2024, 17:1481-1501
Published Date: 6 March 2024
A Topical Chinese Herbal Alleviates Psoriasis by Regulating Keratinocytes Pyroptosis Through Inhibition of NLRP3 Inflammasome Activation
Wang X, Deng Y, Ren X, Li Y, Liu T, Hu B, Li Y
Clinical, Cosmetic and Investigational Dermatology 2026, 19:559712
Published Date: 15 January 2026