Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
A Topical Chinese Herbal Alleviates Psoriasis by Regulating Keratinocytes Pyroptosis Through Inhibition of NLRP3 Inflammasome Activation
Authors Wang X, Deng Y, Ren X, Li Y, Liu T, Hu B, Li Y
Received 10 August 2025
Accepted for publication 15 December 2025
Published 15 January 2026 Volume 2026:19 559712
DOI https://doi.org/10.2147/CCID.S559712
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Xuewan Wang,1 Yutong Deng,1 Xuewen Ren,2 Yatong Li,1 Tangyunni Liu,1 Bo Hu,1 Yuanwen Li1
1Department of Dermatology, Dongfang Hospital, Beijing University of Chinese Medicine, Beijing, People’s Republic of China; 2Department of Dermatology, Shanxi Provincial Hospital of Traditional Chinese Medicine, Shanxi, People’s Republic of China
Correspondence: Yuanwen Li, Department of Dermatology, Dongfang Hospital, Beijing University of Chinese Medicine, Fangzhuang, Fengtai District, Beijing, 100078, People’s Republic of China, Email [email protected]
Purpose: Psoriasis is a chronic inflammatory skin disease involving complex immune dysregulation, where NLRP3 inflammasome-mediated pyroptosis—a pro-inflammatory programmed cell death—has been identified as a key driver of disease pathogenesis. Qingshi anti-itch ointment (QS), a traditional Chinese medicine used for psoriasis, has demonstrated clinical efficacy; however, its specific impact on the NLRP3-pyroptosis pathway remains unclear. This study therefore aims to elucidate the role and underlying mechanisms of QS in regulating NLRP3 inflammasome activation in keratinocytes.
Methods: Psoriasis models were established using both in vivo and in vitro approaches: (1) a mouse model induced by topical application of Imiquimod (IMQ) cream, and (2) a cellular model of keratinocyte pyroptosis stimulated with Lipopolysaccharide (LPS) and Adenosine triphosphate (ATP) in vitro. Therapeutic effects were evaluated through Psoriasis area and severity index (PASI) scoring and Hematoxylin-Eosin staining method (HE). NLRP3 inflammasome-mediated pyroptosis was assessed by IHC, RT-qPCR, and Western blot. Levels of IL-18, IL-1β, IL-17, and IL-22 were measured. Transmission electron microscopy was employed to examine NHEK cellular ultrastructure and pyroptotic status. MCC950, a specific NLRP3 inhibitor, was used to determine whether QS modulates keratinocyte pyroptosis through NLRP3 inflammasome regulation.
Results: QS effectively ameliorated IMQ-induced psoriasiform lesions by suppressing keratinocyte pyroptosis and maintaining cellular integrity. Both in vivo and in vitro experiments demonstrated QS’s ability to inhibit the NLRP3 signaling pathway and selectively regulate key pyroptotic molecules (Caspase-1/GSDMD), thereby reducing proinflammatory cytokine release. Notably, QS combined with MCC950 exhibited significant synergistic effects in suppressing NLRP3 inflammasome activation in both IMQ-induced and LPS/ATP-stimulated models.
Conclusion: QS primarily alleviates psoriasis by modulating keratinocyte pyroptosis through mitochondrial protection, inhibition of GSDMD-mediated membrane perforation, and downregulation of Caspase-1 activity, collectively attenuating inflammatory responses. These findings provide novel mechanistic insights into QS’s anti-psoriatic effects and may facilitate the development of innovative therapeutic strategies for psoriasis.
Keywords: ointment, psoriasis, pyroptosis, NLRP3, inflammasome
Introduction
As the primary cellular barrier of the body, keratinocytes continuously monitor extracellular danger signals by recognizing pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). Upon stimulation, these cells activate inflammasomes and caspase-1, resulting in the release of IL-1β and IL-18, which amplify immune-inflammatory cascades.1,2 Keratinocytes, the predominant cell type in the epidermis, play essential roles in skin renewal, barrier function, and immune defense.
Psoriasis is a chronic immune-mediated inflammatory skin disorder characterized by erythema and silvery-white scales as its primary clinical manifestations. Psoriasis is characterized by keratinocyte hyperproliferation, abnormal differentiation, and infiltration of diverse inflammatory cells, creating a complex network of interactions among keratinocytes, immune cells, and other skin-resident cells. Both domestic and international studies estimate the global prevalence of psoriasis to be approximately 2–3%.3 Psoriasis, along with its associated comorbidities, imposes significant physical and psychological distress on patients, as well as a substantial economic burden on families.
During the progression of psoriasis, dysregulated programmed cell death in keratinocytes disrupts proliferation and differentiation processes, impairing epidermal renewal and facilitating the infiltration of inflammatory cells. This disruption contributes to the intricate interplay between keratinocytes, immune cells, and other skin-resident cells, fueling the pathogenesis of psoriasis.4,5 Intriguingly, keratinocyte pyroptosis has been established as a key player in psoriatic immunoinflammatory responses. Recent studies1,2,6 have identified cellular pyroptosis as a novel pro-inflammatory form of programmed cell death,7 playing a pivotal role in the initiation and amplification of inflammatory responses in psoriasis. Pyroptosis is triggered by inflammasome activation,8 with the NLRP3 inflammasome being the most extensively studied. The NLRP3 inflammasome comprises NOD-like receptor protein 3 (NLRP3), ASC, and pro-caspase-1, the precursor of caspase-1,9,10 and has been closely linked to psoriasis pathogenesis. Under resting conditions, NLRP3 expression in cells remains minimal, but microbial infections or endogenous danger signals stimulate the assembly of the inflammasome, driving inflammatory responses. While abnormal keratinocyte differentiation and keratinization are central to psoriasis pathology, normal human keratinocytes produce pro-IL-1β but do not process it into mature IL-1β. In contrast, psoriatic lesions exhibit significantly elevated caspase-1 activity, enabling the processing and secretion of mature IL-1β, with keratinocytes serving as the primary cellular source of IL-1β.6
Consequently, NLRP3 has emerged as a promising drug target for psoriasis treatment. Among the most studied inhibitors, MCC950 (also known as CP-456773) directly binds to the NACHT domain of NLRP3, maintaining it in an inactive state.11–13 Given that psoriasis and its comorbidities impose substantial physical, psychological, and economic burdens on patients and families, and considering that current mainstay treatments like oral medications and biologics still face challenges including safety concerns and non-response or diminished efficacy in a subset of patients,14–17 it is therefore imperative to investigate therapeutic strategies targeting the NLRP3 pathway for the development of novel psoriasis treatments.
Qingshi anti-itch ointment (QS) Comprises six traditional Chinese medicine: Indigo Naturalis [Asteraceae; Baphicacanthus cusia (Nees) Bremek. extract], Calamina, Gypsum Ustum, Phellodendri [Amurensis Cortex [Rutaceae; Phellodendron amurense Rupr. bark], Sophorae Flavescentis Radix [Fabaceae; Sophora flavescens Ait. dry roots], Borneolum syntheticum. The plant names were verified at http://mpns.kew.org/mpns-portal, and the name of Calamina, Gypsum Ustum, Borneolum syntheticum was authenticated using the Pharmacopoeia of the People’s Republic of China. QS has the effect of clearing heat and removing toxins, removing dampness and relieving itching. Topical treatment with QS (Beijing Drug Preparation Z20160001) can significantly reduce the area of lesions in patients with psoriasis, and alleviate the erythema, hypertrophic infiltration, and scales. Its active ingredient, cynoderma, has anti-inflammatory, anti-proliferating, and anti-angiogenic effects on keratinocytes and human vascular endothelial cells.18–20 Network pharmacology and molecular docking studies have identified the aryl hydrocarbon receptor (AHR) as a key target of indigo, indirubin, and tryptophan—representative active components of Qingdai. Notably, activation of AHR has been shown to suppress both the activation of the NLRP3 inflammasome and its gene transcription.21,22 We have checked the quality standard of the raw and auxiliary materials of Qingshi anti-itch Ointment in the previous period, and used thin-layer chromatography to identify its active ingredients Qingdai, Bitter Ginseng and Phellodendron Bark, and high performance liquid chromatography to determine the content of indigo and indirubin. The results showed that the thin-layer chromatograms of Qingdai, Bitter Ginseng, and Phellodendron Bark had obvious spots, and there was no interference from the negative control, and that the concentration of indigo was in the range of 2.35–4.7 μg (r = 0.9998), and indigofera in the range of 0.00832–0.2912 μg (r=0.9996), which showed a good linear relationship with the peak area (Supplementary S1).23 This study seeks to explore the molecular mechanisms and therapeutic targets of Qingshi anti-itch ointment in two experimental models: IMQ-induced psoriasiform dermatitis in BALB/c mice and LPS/ATP-stimulated keratinocytes.
Materials and Methods
Materials
The reagents used in this study included: Qingshi anti-itch ointment (Beijing Pharmaceutical Preparation Z20160001) as the experimental drug; its base ointment (beeswax:olive oil = 1:10) as the negative control; Calcipotriol ointment (Licence No. HJ20160070, Leo Pharma) as the positive control; and MCC950 (batch no. HY-12815, MCE) as the NLRP3 inhibitor. MCC950 was prepared as a stock solution by dissolving 1 mg of powder in 1 mL of saline (1 mg/mL). This solution was warmed to 37°C and administered to mice via daily intraperitoneal injection at a dose of 10 mg/kg body weight.
IMQ-Induced Psoriasis-Like Mouse Model
The back experimental parts of 70 mice were dehairing with electric pusher combined with depilatory cream, the area was about 2cm*3cm, after 24 hours of dehairing, except for the blank group, the rest of the groups of mice were externally coated with 5% imiquimod (National Pharmaceutical Standard H20031230, Tianfang Pharmaceutical Co., Ltd). 62.5 mg at the dehairing parts to make psoriasis-like mouse model, once a day for 7 consecutive days.
Animals
A well-established mouse model was utilized in this study. Specific Pathogen-Free (SPF) male BALB/c mice (8 weeks old, weighing 20–27 g) were procured from the Centre of Laboratory Animals of SiPeiFu (Beijing) Co., Ltd. (experimental animal quality certificate No. 110324231105899934; experimental animal production license No. SCXK [Beijing] 2019–0010). The mice were housed in standard cages under controlled conditions (22 ± 2°C, 45–55% relative humidity, 12-hour light/dark cycle) and provided unrestricted access to water and feed. All animals were maintained in a specific pathogen-free environment for 7 days prior to the initiation of the experiment.
Seventy mice were acclamatized and fed for 1 week, then given a back dehairing treatment and randomly divided into 7 groups of 10 mice each. In the Control group, 0.5 g of petroleum jelly was applied topically. In the model group, imiquimod was applied topically to induce psoriatic-like dermatitis. The mice were divided into the following groups: the Control group received topical application of 0.5 g of petroleum jelly; the Model group was topically treated with imiquimod to induce psoriasiform dermatitis. Beginning at 6 hours post-modeling each day, the QS, Calcipotriol, and Base groups were administered a daily intraperitoneal injection of saline at 10 mg/(kg·d). In contrast, the MCC950 and QS+MCC950 groups received a daily intraperitoneal injection of MCC950 at the same dosage. Concurrently, the aforementioned five groups (QS, Calcipotriol, Base, MCC950, and QS+MCC950) were treated with topical applications of 0.5 g of Qingshi anti-itch ointment, Calcipotriol ointment, base ointment, petroleum jelly, and Qingshi anti-itch ointment, respectively, using the “one fingertip unit” method for standardized application. This entire regimen was conducted once daily for 7 consecutive days, after which all mice were weighed and tissues were collected. Body mass index = [final body weight (g) - initial body weight (g)]/initial body weight (g) * 100%.
Skin Severity Observation
Observation and recording of daily skin lesion changes in mice, using a digital camera to take pictures of the skin lesion area of the mice, and to ensure that the mice were placed in the same position, at the same height, and with the same background when taking pictures. According to the corrected PASI score as a standard,24 the severity of erythema, infiltration and scaling was scored, and divided into five grades from light to heavy, in the order of 0 (none), 1 (light), 2 (medium), 3 (heavy), 4 (very heavy). The mean values of the three scores were calculated to indicate the severity of the skin lesions, and the mean values of the scores of each group were taken daily to draw a trend curve, thus reflecting the changes of the skin lesions in the mice.
Cell Culture and Treatment
NHEK cells were purchased from Cyagen Biosciences Inc. of USA. The complete medium composition was as follows: Cells were cultured in DMEM supplemented with 10% fetal bovine serum (FBS; Cyagen Biosciences Inc., USA) and 1% penicillin/streptomycin in a humidified incubator maintained at 37°C with 5% CO2. The model was stimulated with a “two-step approach” to activate NLRP3 inflammasome: the first step was to give LPS as an initiating signal for NLRP3 inflammasome activation; the second step was to give ATP as an activating signal for NLRP3 inflammasome activation.25–30
Alcoholic extract of celadonite anti-itching ointment was prepared by the laboratory of the Institute of Traditional Chinese Medicine, China Academy of Chinese Medical Sciences, consisting of 60g of calcined glycerite, 30g of calcined gypsum, 20g of qingdai, 30g of bitter ginseng, 30g of cyperus rotundus, and 10g of icicle. For use, 500 mg of celadonite anti-itching ointment powder was dissolved in 20 mL of DMEM culture solution (without fetal bovine serum), stirred, and then passed through a 20 μmol/L cell sieve. Different dose groups (0.25 mg/mL, 0.5 mg/mL, 1 mg/mL, 2.5 mg/mL, 5 mg/mL, 10 mg/mL) were set up, and the maximum non-toxicity concentration of lapis lazuli itch-relieving ointment extract on NHEK cells was determined by the CCK8 method. The concentration of NLRP3 inhibitor MCC950 was set at 10 umol/L.29
Hematoxylin-Eosin Staining Method (HE)
Mouse skin tissues were fixed in 4% paraformaldehyde for 24 hours and subsequently embedded in paraffin. Tissue sections (5 μm thick) were prepared and stained with HE. Pathological and morphological changes in the mouse skin tissues were examined under a microscope (Ningbo Sunyu Instrument, China), and images were captured using CaseViewer software.
Immunohistochemistry Assay (IHC)
Mouse skin tissues were fixed in 4% paraformaldehyde for 24 hours and processed for IHC on paraffin-embedded tissue sections (5 μm thickness). Antigen retrieval was performed using citrate buffer solution (pH 6.0), followed by incubation with 3% H2O2 to quench endogenous peroxidase activity. The sections were then incubated overnight at 4°C with the following primary antibodies: NF-κB p65 Monoclonal antibody (66535-1-Ig, Proteintech, China), NLRP3 Monoclonal antibody (68102-1-Ig-100, Proteintech, China), Caspase-1 antibody (sc-56036, Santa Cruz, USA), and GSDMD Polyclonal antibody (20770-1-AP, Proteintech, China). Images of the stained sections were captured using a microscope (Ningbo Sunyu Instrument, China), and the expression levels of the target proteins were quantified using Image-Pro Plus 6.0 image analysis software. 3 fields of view were randomly selected from each section without repeating, and the integrated optical density (IOD) and area were measured respectively. The average optical density (AOD) was calculated based on IOD/area, reflecting the difference in protein expression per unit area.
Transmission Electron Microscopy (TEM)
After intervention on NHEK cells, cell suspension was made, supernatant was discarded, washed twice with PBS and centrifuged. The cells were added to glutaraldehyde overnight at 4°C, fixed, embedded and stained. The cell structure and pyroptosis status were observed and analysed by TEM (jeol, Japan) and images were captured.
RT-qPCR Analysis
NHEK cells and frozen mouse skin tissue specimens stored at −80°C were utilized for RNA extraction. Total RNA from mouse skin tissues was isolated using the SuperScript III RT reverse transcription kit (ABI-Invitrogen, USA), while RNA from cells was extracted using TRIZOL reagent (Invitrogen, USA). The isolated RNA was reverse-transcribed into cDNA using specific primers. The mRNA expression levels of cytokines, normalized to β-actin expression, were quantified using a fluorescence quantitative PCR instrument (Applied Biosystems, USA). Details of the primers used for each target gene are provided in Table 1.
 |
Table 1 Real-Time PCR Primer Sequences |
Western Blot (WB)
NHEK cells and mouse skin tissue specimens frozen in −80°C freezer were taken. Cells were lysed with 1×RIPA lysis solution (Biyun Tian, China), and proteins were extracted by centrifugation at 4°C and 12000g for 15 min. The protein concentration was quantified by BCA protein concentration assay kit (MDL, China). Proteins from each sample were analyzed using 10% SDS-PAGE gel electrophoresis. The membranes were incubated overnight at 4°C with primary antibodies, including NF-κB p65 antibody (66535-1-Ig, Proteintech, China), NLRP3 Monoclonal antibody (68102-1-Ig-100, Proteintech, China), ASC Polyclonal antibody (BD-PT0365-100, Biodragon, China), Anti-pro Caspase-1 antibody (ab179515-40, Abcam, UK), Caspase-1 antibody (sc-56036, Santa Cruz, USA), and GSDMD Polyclonal antibody (20770-1-AP, Proteintech, China), and Beta Actin Polyclonal antibody (AF7018, Affinity, USA). Subsequently, the membranes were incubated for 1 hour with HRP-conjugated Goat Anti-Rabbit IgG (H+L) (SA00001-2, Proteintech, China) or HRP-conjugated Goat Anti-Mouse IgG (H+L) (SA00001-1, Proteintech, China) as secondary antibodies. Protein bands were visualized using the ChemiDoc MP Imaging System (ChemiScope 6100, CLINX Qinxiang, China).
Enzyme Linked Immunosorbent Assay (ELISA)
The concentrations of inflammatory cytokines including interleukin (IL)-18, IL-1β, IL-17, and IL-22 were quantified using ELISA kits according to the manufacturer’s instructions. Specifically, mouse IL-18 (Catalog No. JL20253), IL-1β (JL18442), IL-17 (JL20250), and IL-22 (JL20258) were assayed using kits (Shanghai Jianglai, China), whereas human IL-18 (MD13525), IL-1β (MD145659), IL-17 (MD15876), and IL-22 (MD16317) were measured with kits (Beijing BioSciTech, China). The absorbance at 450 nm was determined using a Labsystems Multiskan MS microplate reader (Thermo Fisher Scientific, Finland).
Statistical Analysis
GraphPad Prism 9.5 (GraphPad Software Inc., San Diego, CA) and SPSS 25.0 were used to statistically process the collected data. Data were expressed as mean ± standard deviation (mean ± SD), and one-way ANOVA with Tukey’s multiple comparisons. The criterion for significant difference was set at P<0.05.
Results
QS Ameliorated IMQ-Induced Psoriasis-Like Skin Lesions in Mice
IMQ, a Toll-like receptor 7/8 ligand and potent immune activator, is widely employed to replicate human psoriasis in mouse models.31 In this study, we investigated the potential anti-psoriatic effects of QS in vivo using an IMQ-induced mouse model of psoriasis-like skin lesions. Observations of phenotypic changes in the back skin (Figure 1) and lesion severity scores (Figure 2A–D) revealed that, following 7 days of IMQ application, the mice developed progressive erythema, infiltration, and scaling. Over time, the erythema deepened in color, infiltration thickened further, and scaling became increasingly widespread and pronounced. These lesion indices peaked on day 7, culminating in the development of psoriasis-like skin lesions. The Base control developed a psoriasiform phenotype comparable to that of the IMQ-induced mice, both exhibiting dark erythema, hypertrophic infiltrates, and scale accumulation. In contrast, mice treated with Calcipotriol or QS showed a notable alleviation of symptoms, characterized by a lighter dorsal infiltrate and significantly fewer and finer scales compared to the IMQ-induced mice. Furthermore, an analysis of the mouse body mass index revealed the most significant weight loss in the IMQ-induced mice, with the QS group exhibiting a less pronounced reduction (Figure 2E). Histopathological analysis of the dorsal skin using HE staining showed hyperkeratosis with hyperkeratosis, hypertrophy of the stratum spinosum, increase in the number of spinous cells, downward extension of the epidermal protrusion in the form of a mallet, and infiltration of inflammatory cells in the dermis accompanied by dilation of capillaries after IMQ induction, which was consistent with the pathological manifestations of psoriatic lesions. In the skin of QS-treated mice the degree of pathological damage was lower, the stratum corneum was thinner, there was partial restoration of the epidermis, the keratosis was attenuated, occasional hyperkeratosis was seen, the granular layer was preserved, the spinous layer was thinned, the vasodilatation was insignificant, the inflammatory cell infiltration was reduced, and the downward extension of the epidermal protuberance was attenuated (Figure 2F).
 |
Figure 1 Effect of each group on IMQ-induced psoriasis-like mouse skin. Abbreviation: D, Day. |
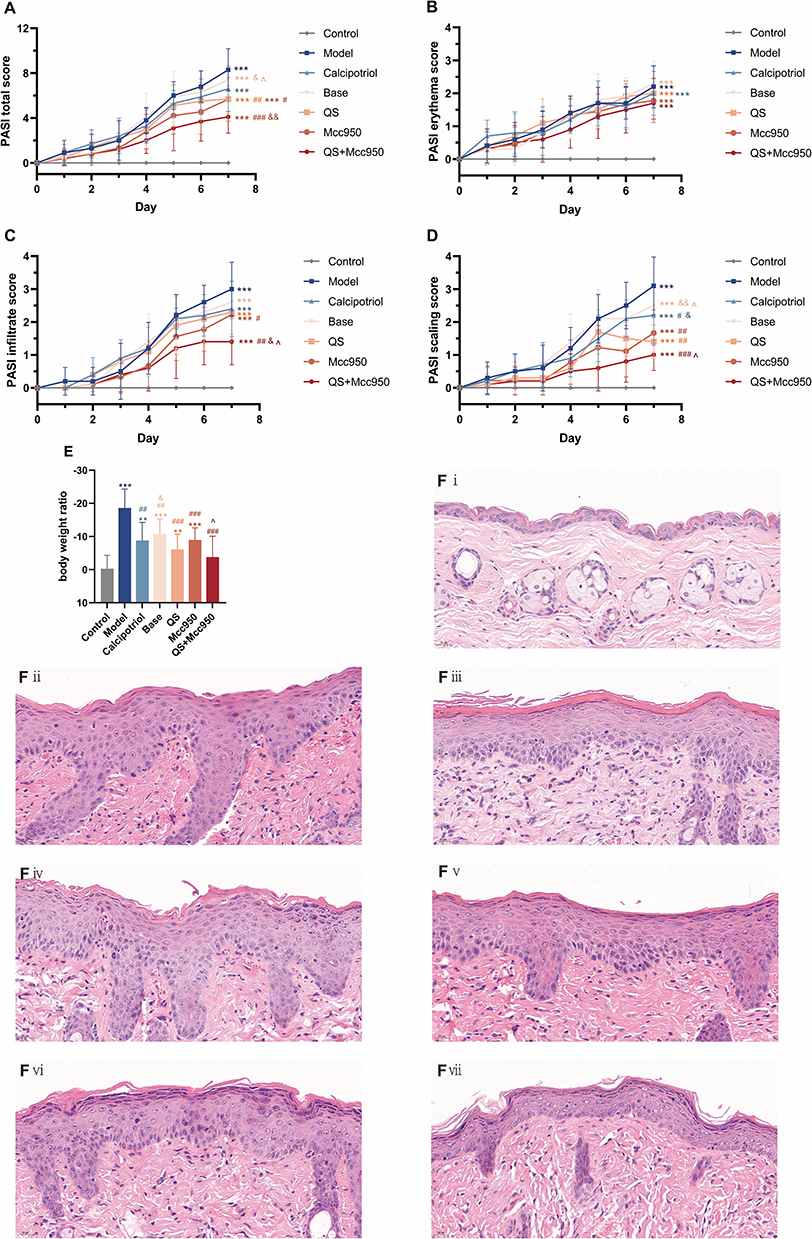 |
Figure 2 (A–D) Trend plot of PASI score, n = 10; (E) Body weight ratio, n = 10. **P < 0.01, ***P < 0.001 compared with Control; #P < 0.05, ##P < 0.01, ###P < 0.001 compared with Model &P < 0.05, &&P < 0.01 compared with QS; ^P < 0.05 compared with Mcc950; (F) HE staining of skin tissue, (i) Control; (ii) Model; (iii) Calcipotriol; (iv) Base; (v) QS; (vi) Mcc950; (vii) QS+Mcc950. |
QS Alleviates LPS+ATP-Induced Pyroptosis and Ultrastructural Damage in NHEK Cells
Cell viability was detected by CCK-8 assay (Figure 3A), RNA was extracted for qPCR to detect GSDMD, Caspase-1, IL-1β (Figure 3B–D), and LPS (0.5 ug/mL 12h) + ATP (5mM 30min) was selected to construct the focal pyroptosis model. Based on the safety range and considering the validity of the subsequent experiments, the concentration of 2.5 mg/mL drug was selected (Figure 3E).
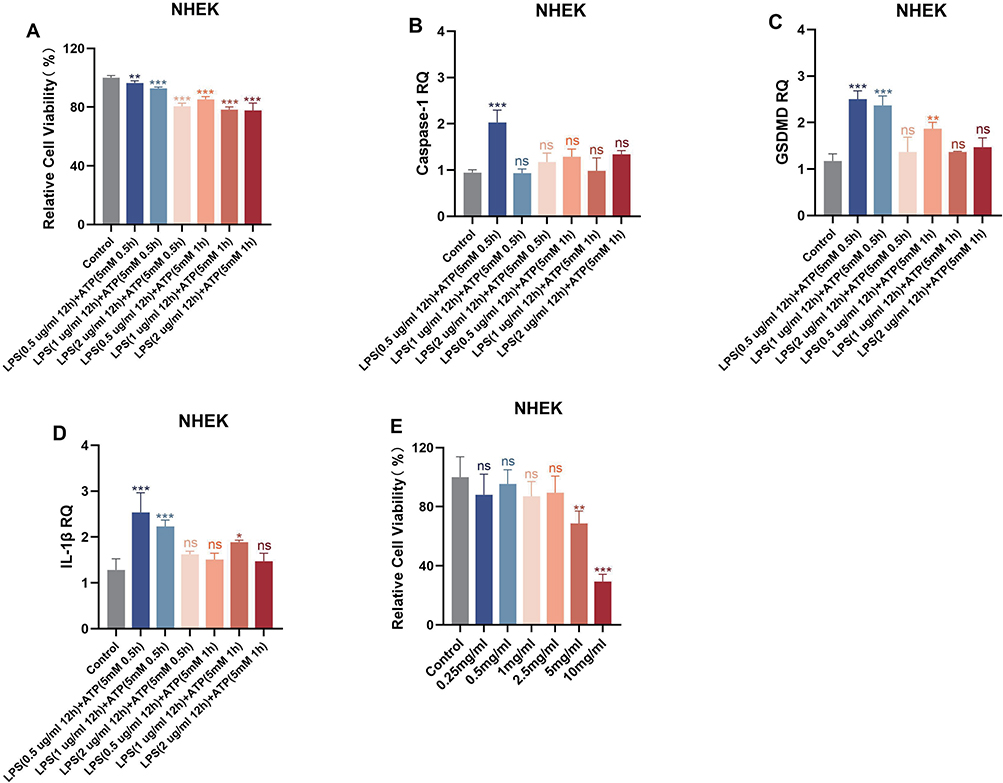 |
Figure 3 (A) Relative cell viability under LPS+ATP stimulation was analysed in NHEK cells, n=5; (B–D) mRNA levels of Caspase-1, GSDMD, IL-1β under LPS+ATP stimulation were analysed in NHEK cells, n=3; (E) The effect of QS on the viability of NHEK cells, n=5. *P< 0.05, **P < 0.01, ***P < 0.001 compared with Control. Abbreviation: ns, no significance. |
Bacterial infections are a significant contributing factor in both the onset and exacerbation of psoriasis.32 In this study, we used LPS+ATP “two-step method” to establish a keratinocyte pyroptosis model to simulate the pathological process of psoriasis induced by bacterial infection. The results showed that LPS and ATP treatments significantly disrupted mitochondrial function, which together promoted the assembly of NLRP3 inflammatory vesicles, ultimately leading to caspase-1 activation, GSDMD cleavage and IL-1β/IL-18 maturation release. During this process, LPS and ATP stimulated cells showed typical features of pyroptosis: cell membrane rupture with pyroptotic vesicle formation, mitochondrial cristae degradation and vacuolisation, endoplasmic reticulum dilatation, and other ultrastructural alterations. QS significantly improved the above pathological changes: (1) at the morphological level, it effectively protected the integrity of the cell membrane, reduced pyroptotic vesicle formation, and markedly attenuated the mitochondrial swelling and endoplasmic reticulum dilatation; (2) In terms of molecular mechanisms, the inhibition of the pyroptosis process was achieved through multi-targeted effects: (a) direct inhibition of GSDMD cleavage activation, reducing the formation of cell membrane pores; (b) maintenance of mitochondrial function, possibly through the regulation of mitochondrial membrane potential and scavenging of ROS; and (c) alleviation of endoplasmic reticulum stress, possibly through the modulation of the response to unfolded proteins. Notably, the combination of QS with the NLRP3-specific inhibitor MCC950 showed significant synergistic effects, and the cellular ultrastructure and molecular indexes were close to normal after the combined application, suggesting that QS may complement the upstream NLRP3 inhibitor by acting on the downstream link of pyroptosis (Figure 4).
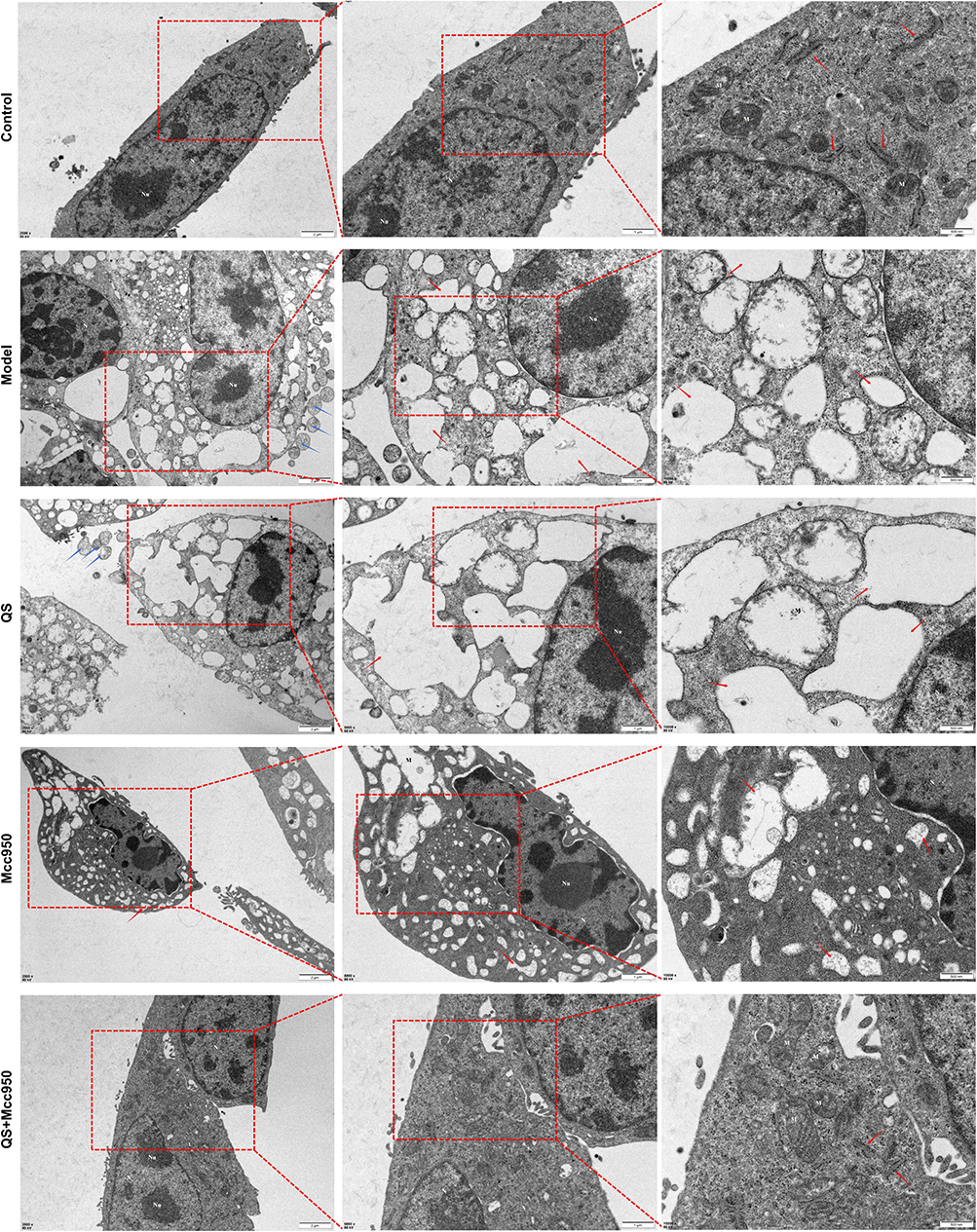 |
Figure 4 NHEK transmission electron microscopy results. Red arrow: endoplasmic reticulum; blue arrow: cytoplasm. Abbreviations: N, nucleus; Nu, nucleolus; M, mitochondria. |
QS Inhibits the NLRP3 Signalling Pathway in vivo and in vitro, Targeting a Key Molecule in the Regulation of Caspase-1/GSDMD Pyroptosis
NLRP3 serves as a critical mediator in the formation of inflammasomes, which activate innate immunity and promote the release of inflammatory factors, thereby initiating an inflammatory response.33 In psoriatic tissues, NLRP3 expression is markedly elevated compared to normal skin, with levels approximately 3.5 to 4.3 times higher.34 These findings suggest that NLRP3 may play a significant role in the pathogenesis of psoriasis. In the present study, elevated gene and protein expression of NLRP3-related upstream and downstream factors were observed in IMQ-induced mouse skin tissues. The results indicated that over-activation of NF-κB signaling could promote the assembly of the NLRP3 inflammasome and facilitate the cleavage of pro-caspase-1 into active caspase-1. In addition, increased levels of GSDMD mRNA and protein were detected, suggesting possible cleavage of GSDMD. This process may subsequently lead to the extracellular release of active inflammatory factors such as IL-1β and IL-18 through membrane pores, and further induce secretion of IL-17 and IL-22, ultimately triggering keratinocyte pyroptosis that manifested as skin inflammation and epidermal hyperproliferation (Figure 5).
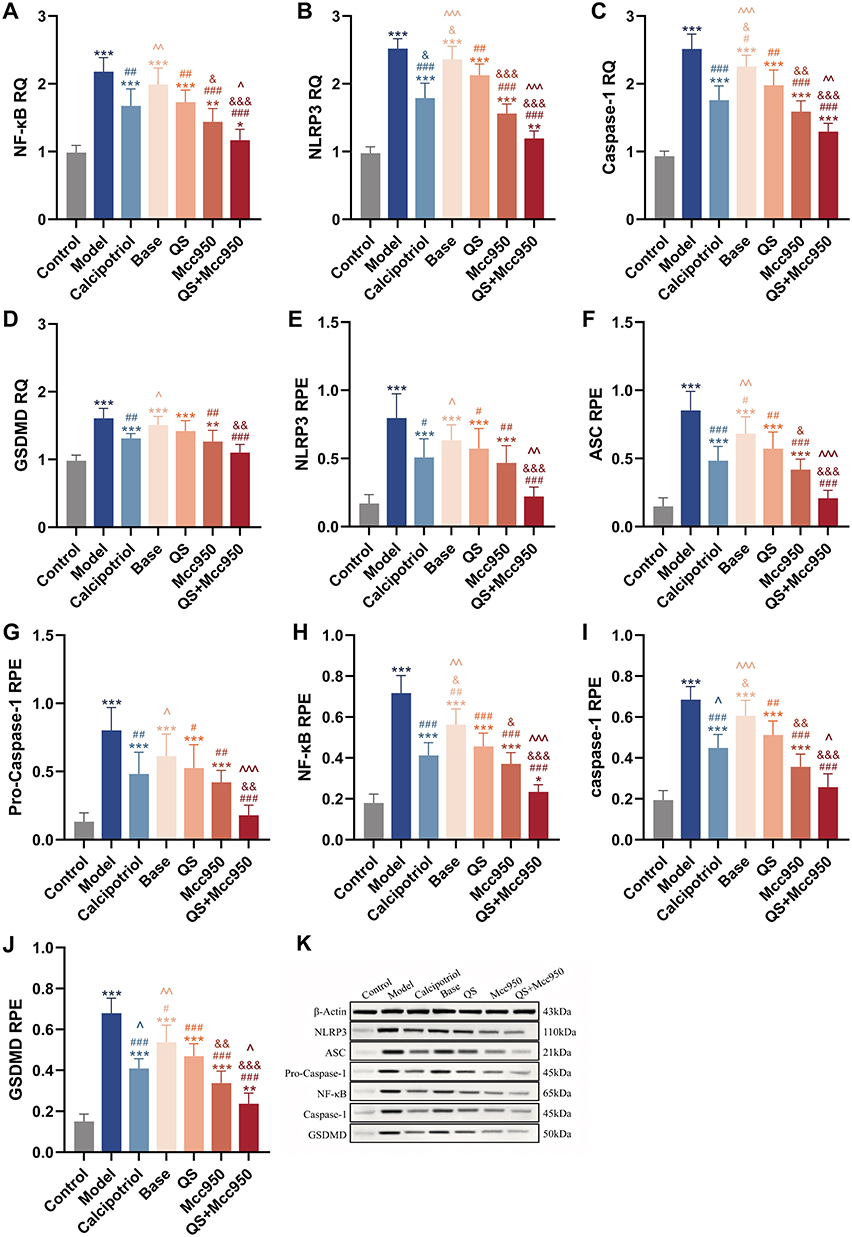 |
Figure 5 (A–D) NLRP3, NF-κB, caspase-1, GSDMD mRNA expression in mouse skin tissues of each group, n = 6. (E–K) Protein expression results of NLRP3, ASC, Pro-Caspase-1, NF-κB, caspase-1, GSDMD in mouse skin tissues of each group, n = 6. *P < 0.05, **P < 0.01, ***P < 0.001; compared with Control; #P < 0.05, ##P < 0.01, ###P < 0.001 compared with Model; &P < 0.05, &&P < 0.01, &&&P < 0.001 compared with QS; ^P < 0.05, ^^P < 0.01, ^^^P < 0.001 compared with Mcc950. |
We verified the modulation of NLRP3 signalling in psoriasis by QS. In vitro, there was inhibition of pyroptosis-related targets in keratinocytes by QS, mainly in the protein expression of NF-κB, NF-κB Pp65, NLRP3, Caspase-1, GSDMD, as well as in the gene expression of NLRP3, Caspase-1 and GSDMD. QS significantly inhibited LPS+ATP-stimulated NHEK cells (Figure 6). In vivo, QS modulated the expression of NLRP3 inflammatory vesicle-related proteins and genes equally positively, and although its overall efficacy was slightly lower than that of Calcipotriol, there was no significant difference between the two in the expression of NF-κB, Caspase-1, and GSDMD genes as well as the proteins related to NLRP3, which can be seen to show a very high degree of similarity between the two, especially at the protein level, this similarity was particularly remarkable, suggesting that they may play similar roles or be affected by similar regulatory mechanisms in the cellular pyroptosis process of keratinocytes (Figure 5). To further clarify the target of action of QS, immunohistochemical analysis was performed (Figure 7), and it was found that the inhibitory effects of QS alone versus combined with Mcc950 on Caspase-1 and GSDMD in epidermal acanthosis and basal lamina were particularly significant, suggesting that QS may not only inhibit the upstream NF-κB/NLRP3 signalling through multi-target modulation, but may also directly or indirectly intervene in the Caspase-1 activation and GSDMD shearing, thereby attenuating cellular pyroptosis. In addition, at the molecular level, Base showed no significant difference from the IMQ-induced mouse model at both protein and gene levels, and QS added the drug to Base, and the two showed no significant difference in the expression of Caspase-1 genes and proteins as well as in the downstream inflammatory factors (eg, IL-18, IL-1β, IL-17, IL-22) at the levels were significantly different, which reinforces the idea that QS may play a role in psoriasis treatment by affecting caspase-1 activity (Figures 5 and 8). Therefore, we believe that QS demonstrated superior efficacy, which may be due to its ability to act on Caspase-1 and GSDMD, which in turn regulate IL-18 and IL-1β, key factors in the immune-inflammatory response of keratinocytes, and thus significantly inhibit the onset and progression of keratinocytes juxtaposition in psoriasis, and positively affect the pathological process of psoriasis in a therapeutic manner.
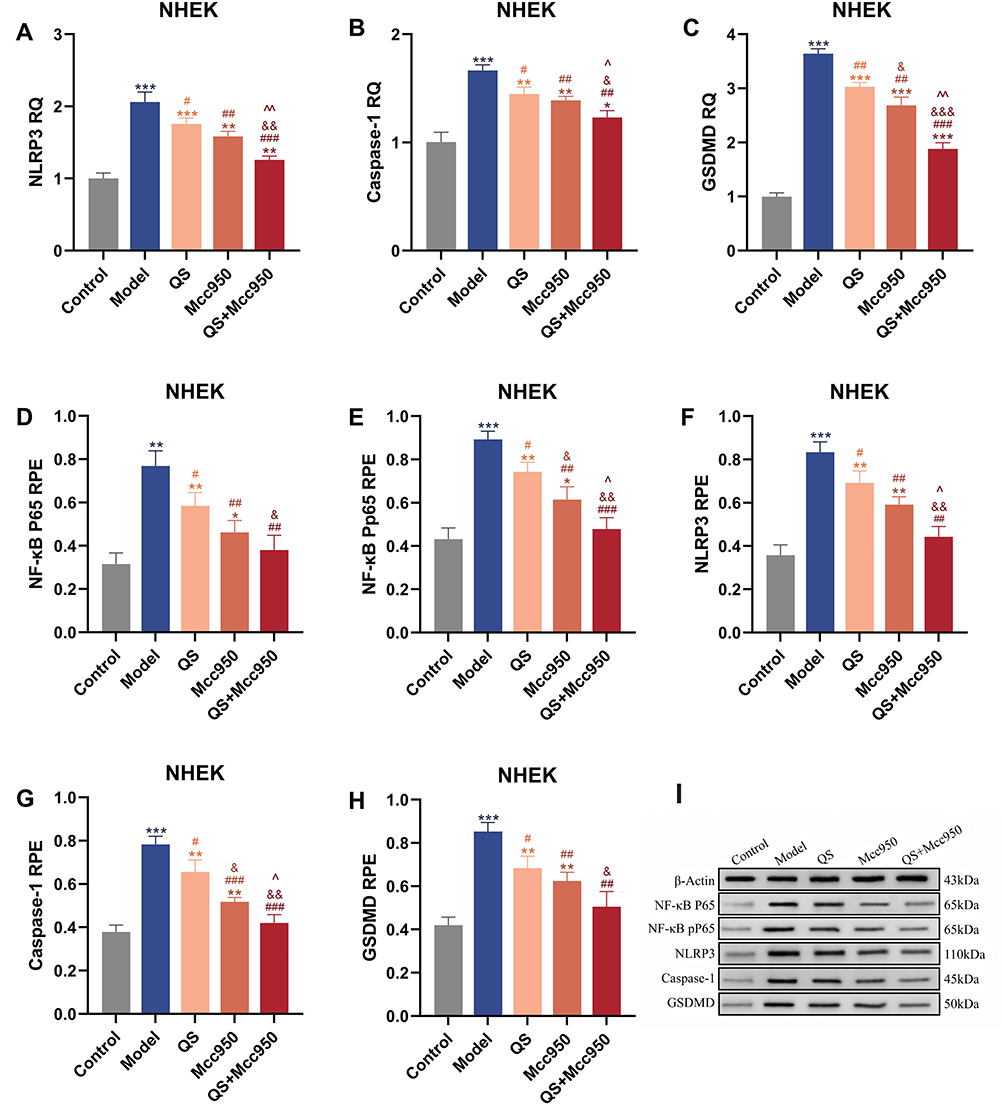 |
Figure 6 (A–C) mRNA levels of NLRP3, Caspase-1, GSDMD in NHEK cells, n = 3. (D–I) Protein levels of NF-κB, P65, NLRP3, caspase-1, GSDMD in NHEK cells, n = 3.*P < 0.05, **P < 0.01, ***P < 0.001 compared with Control; #P < 0.05, ##P < 0.01, ###P < 0.001 compared with Model; &P < 0.05, &&P < 0.01, &&&P < 0.001 compared with QS; ^P < 0.05, ^^P < 0.01 compared with Mcc950. |
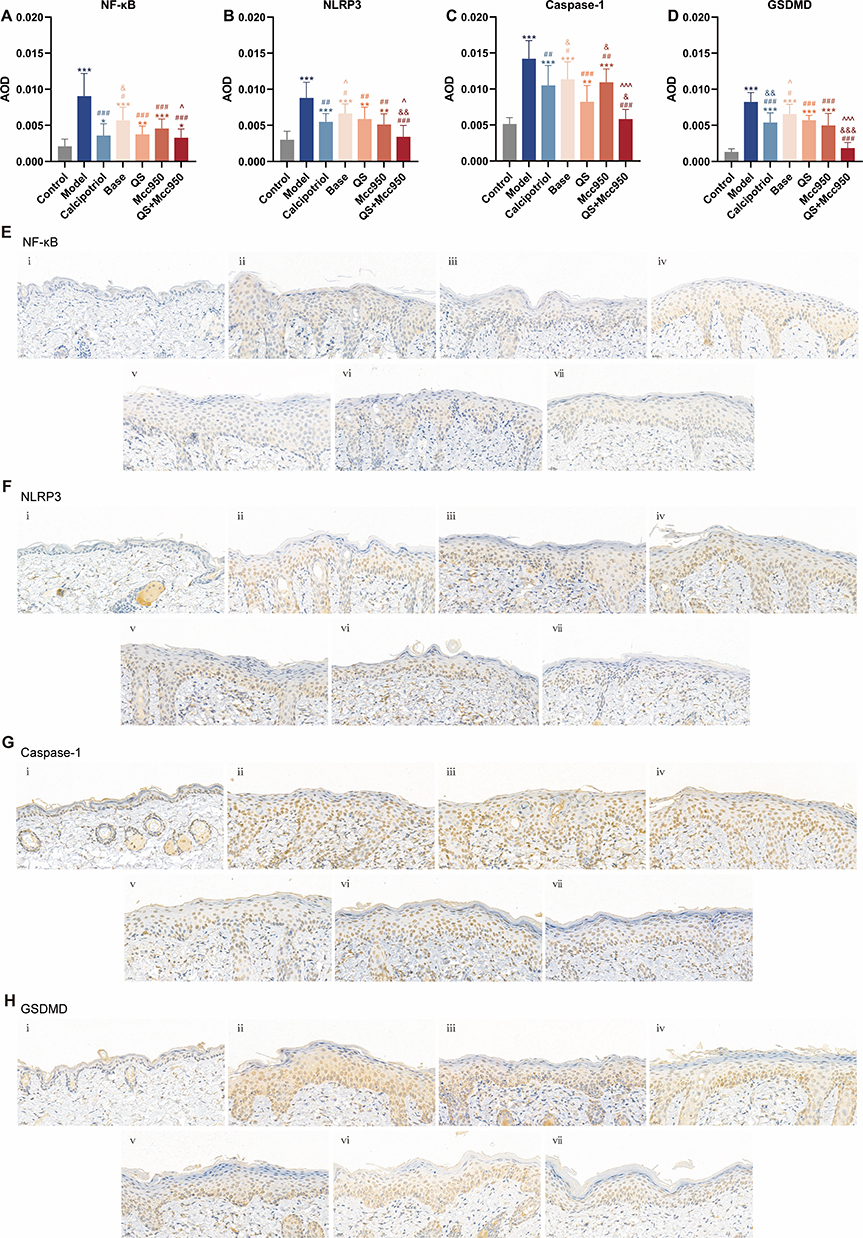 |
Figure 7 (A–D) AOD values of NF-κB, NLRP3, Caspase-1, and GSDMD immunohistochemical staining in skin tissues from each group, n =3. (E–H) Immunohistochemical Staining for NF-κB, NLRP3, Caspase-1, and GSDMD in skin tissues from each group. (i) Control; (ii) Model; (iii) Calcipotriol; (iv) Base; (v) QS; (vi) Mcc950; (vii) QS+Mcc950. *P < 0.05, **P < 0.01, ***P < 0.001 compared with Control; #P < 0.05, ##P < 0.01, ###P < 0.001 compared with Model; &P < 0.05, &&P < 0.01, &&&P < 0.001 compared with QS; ^P < 0.05, ^^^P < 0.001 compared with Mcc950. |
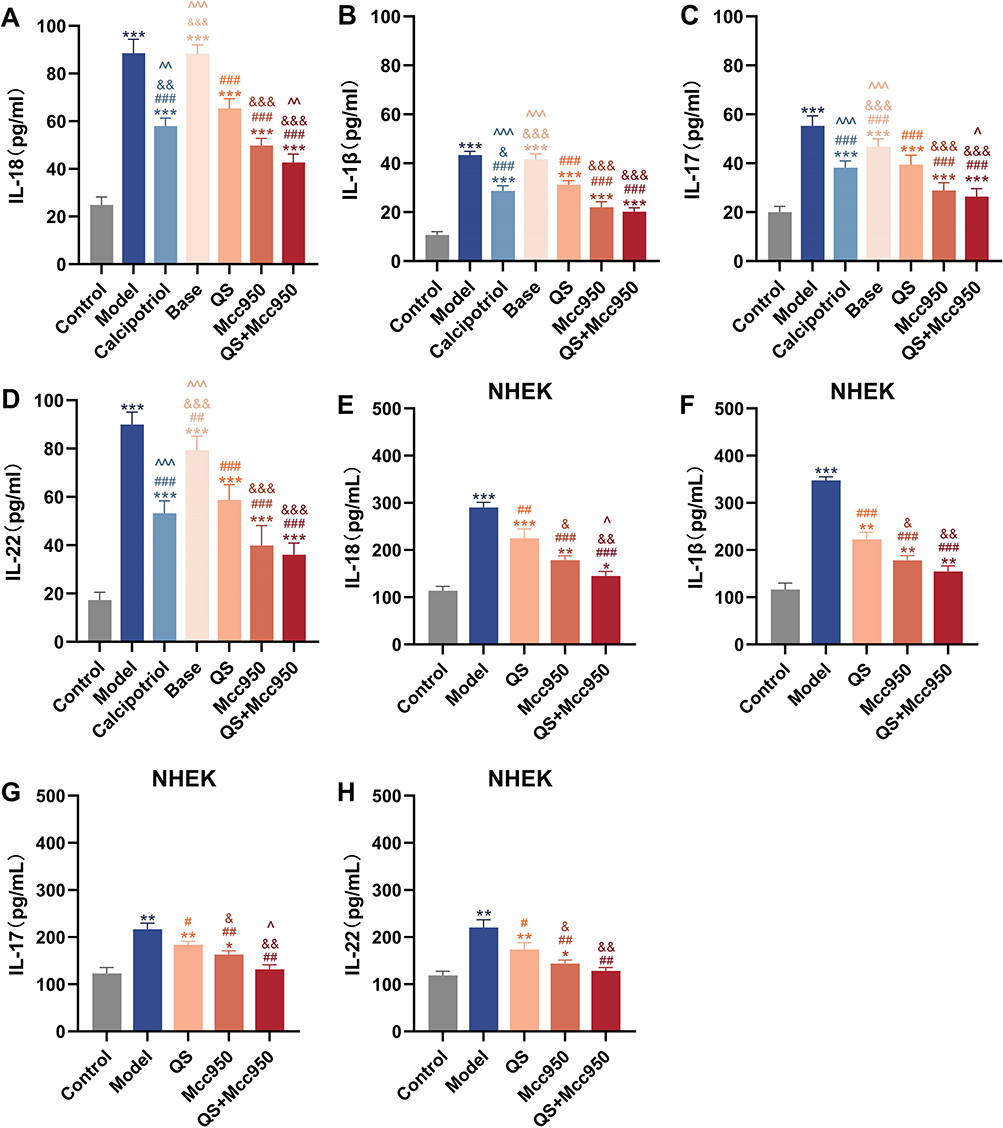 |
Figure 8 (A–D) Mouse skin tissue IL-18, IL-1β, IL-17, IL-22 levels, n = 6. (E–H) IL-18, IL-1β, IL-17, IL-22 levels in NHEK cells, n = 3. *P < 0.05, **P < 0.01, ***P < 0.001 compared with Control; #P < 0.05, ##P < 0.01, ###P < 0.001 compared with Model; &P < 0.05, &&P < 0.01, &&&P < 0.001 compared with QS; ^P < 0.05, ^^P < 0.01, ^^^P < 0.001 compared with Mcc950. |
QS Enhances Synergistic Inhibition of MCC950 Both in vivo and in vitro
MCC950 is a highly potent and selective NLRP3 inhibitor capable of effectively suppressing inflammasome activation.35 Based on these findings, we aimed to further elucidate the potential mechanisms underlying the anti-psoriatic effects of QS. In vivo studies demonstrated that the combination of QS with Mcc950 achieved optimal improvement in erythema, infiltration, and scaling - manifested as lighter erythema, smoother skin texture, and finely fragmented or even absent scaling (Figure 1). This combination showed particularly outstanding performance in PASI scores, especially for infiltration and scaling parameters (Figure 2A–D). Regarding body weight index, while QS or Mcc950 monotherapy had minimal impact on IMQ-treated mice, their combined use showed the least effect, forming a marked contrast with both untreated IMQ mice and those receiving Base. This suggests that QS can effectively counteract psoriasis-induced systemic depletion while avoiding drug toxicity, with combination therapy causing even less physiological burden (Figure 2E). IHC analysis revealed significantly enhanced NLRP3 expression following IMQ stimulation, which was markedly reduced in the basal layer after Mcc950 inhibition. This suppression was further potentiated by QS co-administration, indicating that both Mcc950 (as an NLRP3-specific inhibitor) and its combination with QS may directly target the NLRP3 inflammasome to block its activation (Figure 7). Further integrated analysis at both gene and protein levels (Figures 5 and 8A–D) demonstrated superior therapeutic efficacy of the MCC950+QS combination in mitigating psoriasis progression. Notably, compared to Mcc950 monotherapy, the combined regimen showed significantly stronger inhibition of NF-κB, NLRP3, and Caspase-1 at both transcriptional and translational levels, along with more pronounced suppression of ASC, Pro-Caspase-1, IL-18, and IL-17 protein expression. These findings suggest that when pyroptosis pathway is inhibited, QS may exert enhanced anti-psoriatic effects by modulating NF-κB expression, interfering with inflammasome assembly, reducing Caspase-1 levels, and consequently diminishing downstream inflammatory cytokine production. This mechanism likely represents a crucial therapeutic target through which QS treats psoriasis via pyroptosis-related pathways, offering novel perspectives and potential therapeutic strategies for psoriasis management.
Consistent with the in vivo results, our in vitro experiments (Figure 6) systematically compared the characteristics of QS and MCC950, revealing complementary regulatory sites and potential synergistic mechanisms between the two within the pyroptosis pathway:
(1) In targeting the upstream NLRP3 component, MCC950 demonstrated a more direct inhibitory trend. At both mRNA and protein levels, NLRP3 expression in the MCC950-treated group was lower than in the QS group. Although this difference did not reach statistical significance, it aligns with MCC950’s molecular mechanism of specifically binding to the NACHT domain, suggesting its potential advantage in directly blocking inflammasome assembly. In contrast, QS exhibited a relatively limited direct impact on NLRP3. (2) In regulating the downstream pyroptosis execution phase, QS displayed a crucial enhancing effect. Firstly, regarding the inhibition of the pore-forming protein GSDMD, the combination of QS and MCC950 showed significantly stronger suppression of GSDMD mRNA than MCC950 alone, and restored GSDMD protein levels to near-normal. This indicates that QS may effectively reinforce the blockade of the final execution step of pyroptosis. Secondly, although the two monotherapies showed comparable inhibition of Caspase-1, their combination further reduced Caspase-1 expression at both transcriptional and translational levels. This suggests that QS might enhance the suppression of Caspase-1 activity across the entire pathway via synergistic or parallel pathways, thereby more thoroughly curbing the amplification of inflammatory signals. (3) The combination therapy demonstrated remarkable synergistic effects: In NHEK cells, where all measured pyroptosis pathway markers showed no significant difference from normal cells. These comprehensive results demonstrate that QS primarily targets downstream execution events mediated by GSDMD (membrane pore formation) and Caspase-1 activation, creating perfect cascade blockade when combined with MCC950’s upstream NLRP3 inflammasome inhibition. This complementary action achieves complete pathway suppression from initial activation to final effector steps.
QS Inhibits the Release of Inflammatory Cytokines
Deficiency in IL-1 receptor antagonists is closely associated with cutaneous autoimmune diseases, particularly as IL-1β and IL-18 - key effector molecules of pyroptosis - play pivotal roles in psoriasis pathogenesis. IMQ treatment significantly elevated cutaneous levels of inflammatory mediators including IL-18, IL-1β, IL-17, and IL-22. QS treatment markedly reduced the production of these inflammatory cytokines, an effect not observed with Base. Notably, the QS+MCC950 combination demonstrated superior suppression of IL-18 and IL-17 levels compared to MCC950 monotherapy (Figure 8A–D). In vitro experiments (Figure 8E–H) confirmed that LPS and ATP stimulation significantly elevated levels of IL-1β, IL-18, IL-17, and IL-22, validating the central role of pyroptosis in driving a robust inflammatory cascade. Although QS markedly reduced these cytokine levels, its efficacy when used alone was weaker than that of MCC950, which directly targets NLRP3. This suggests that QS has a limited direct inhibitory effect on the upstream NLRP3 inflammasome. However, the combination of QS and MCC950 demonstrated optimal global suppressive efficacy, restoring all cytokine levels to near-normal ranges. Most notably, in inhibiting IL-18 and IL-17, the combined application showed significantly superior effects compared to MCC950 alone, indicating that this combination strategy may possess a unique synergistic advantage in modulating the specific inflammatory axis (IL-18/IL-17). These results not only confirm the synergistic effect of the drug combination from multiple perspectives but also reveal that QS may possess a multi-targeted mode of action beyond a single pyroptosis pathway in alleviating psoriatic inflammation, thereby laying a critical theoretical foundation for the development of novel therapeutic strategies.
Discussion
Psoriasis currently affects approximately 125 million people worldwide.36 Its pathological mechanisms are highly complex but primarily involve chronic inflammation, autoimmune dysregulation, and genetic predisposition.37 Among these factors, chronic inflammation plays a pivotal role in the pathogenesis of psoriasis. In recent years, the NLRP3 inflammasome has gained increasing attention as a trigger of inflammatory responses.38 Elevated levels of NLRP3-mediated pyroptosis have been observed in psoriatic skin lesions, particularly within keratinocytes.39 Activation of the NLRP3 inflammasome through the NF-κB signaling pathway initiates caspase-1 activation, triggering inflammatory responses and pyroptosis, which has progressively become a breakthrough point in psoriasis research. Traditional Chinese medicine formulations have demonstrated considerable potential in alleviating psoriasis due to their multi-component composition, multi-target mechanisms, and favorable safety profile, making them a viable alternative therapeutic approach. The clinical application of QS in psoriasis treatment has confirmed its efficacy, with its active components capable of suppressing NLRP3 inflammasome activation and gene transcription.21,22 Therefore, QS represents an effective therapeutic strategy for psoriasis by targeting the NLRP3 inflammasome signaling pathway to minimize detrimental effects.
In this study, we initially evaluated the effect of QS on IMQ-treated mice with psoriasis. The results demonstrated that QS effectively alleviated the psoriatic condition. IMQ, an agonist of Toll-like receptor 7/8, is known to induce dermatitis in mice that closely resembles human psoriasis40 and is widely used to establish psoriatic-like mouse models. Interestingly, IMQ-treated mice developed psoriatic-like skin conditions, including erythema, epidermal thickening, and increased scaling severity scores. Furthermore, the base ointment alone had no therapeutic effect, whereas QS treatment ameliorated both the phenotypic and histological changes induced by IMQ. The body mass index (BMI) serves as an important parameter reflecting the general health status of mice and may correlate with disease severity and inflammatory responses. QS administration significantly reduced the IMQ-induced increase in BMI. In addition, recent studies have shown that biologics targeting IL-17 exhibit remarkable efficacy in clinical treatment.41,42 IL-1β promotes IL-17 secretion by activating dermal T cells, while IL-18 expression levels not only positively correlate with PASI scores but are also significantly higher in active lesions compared to stable plaques.43 Similarly, our results indicated that QS intervention markedly suppressed the release of inflammatory cytokines in IMQ-treated mice. These findings collectively demonstrate the anti-inflammatory properties of QS.
Pyroptosis, a significant form of programmed cell death, is primarily mediated through the canonical caspase-1-dependent pathway and the non-canonical caspase-4/5/11-dependent pathway.44 In the canonical pathway, following recognition of PAMPs/DAMPs by pattern recognition receptors, the ASC adapter protein recruits pro-caspase-1 to form the inflammasome complex.1 Activated caspase-1 subsequently exerts dual pro-inflammatory effects: (1) it cleaves GSDMD to generate the pore-forming N-terminal domain, which induces plasma membrane perforation and osmotic imbalance45; (2) it facilitates the maturation and release of pro-IL-1β and pro-IL-18,46,47 thereby amplifying inflammatory responses. Currently, the role of pyroptosis in the pathogenesis of psoriasis is predominantly attributed to cellular pyroptosis driven by the activation of the NLRP3 and AIM2 inflammasome pathways. In this process, NF-κB signaling activates inflammasomes, which subsequently trigger caspase-1 activation. This leads to the cleavage of GSDMD and the maturation of pro-IL-1β and pro-IL-18 into their active forms, IL-1β and IL-18. These active cytokines further stimulate the IL-23/Th17 pathway, promoting the release of large quantities of inflammatory cytokines and chemokines. This cascade ultimately results in skin inflammation and epidermal hyperproliferation.48 Therefore, we further investigated the effects of QS on the NLRP3 signaling pathway in psoriasis. In addition to the IMQ-induced psoriatic mouse model, we established an in vitro pyroptosis model by treating keratinocytes with LPS and ATP to simulate bacterial infection-related pathological processes. NLRP3 inflammasome activation involves a complex cascade comprising two critical phases: priming and assembly. In the priming phase, microbial components or endogenous danger signals activate the TLRs/NF-κB pathway, leading to the upregulation of NLRP3, pro-IL-1β, and pro-IL-18 transcription.49 The assembly phase is triggered by ATP-induced P2X7 receptor-mediated K+ efflux and Ca2+ influx, with additional contributions from mitochondrial dysfunction and lysosomal rupture mechanisms.33,50–53 In LPS/ATP-stimulated NHEK cells, QS intervention effectively counteracted the ultrastructural damage induced by LPS/ATP in keratinocytes. The preservation of plasma membrane integrity and alleviation of mitochondrial swelling and endoplasmic reticulum dilation functionally confirmed the anti-pyroptotic activity of QS. At the molecular level, this cytoprotective effect can be attributed to its inhibition of the NLRP3 inflammasome signaling pathway and subsequent regulation of both Caspase-1 and GSDMD. The consistency between these in vitro mechanisms and our in vivo results strengthens the conclusion that QS alleviates psoriatic inflammation through this specific pathway. These findings offer new experimental insights into the therapeutic mechanism of QS in psoriasis and lay a theoretical groundwork for the development of combined treatment strategies aimed at the pyroptosis pathway.
MCC950 specifically binds to the NACHT domain of NLRP3, effectively stabilizing it in an inactive state.11,13 This inhibitory effect is crucial for controlling NLRP3 inflammasome-mediated inflammatory responses and demonstrates significant therapeutic potential for inflammatory diseases. Our study revealed that QS can potentiate the synergistic inhibitory effects of MCC950, a finding consistently validated in both in vivo and in vitro experiments. Specifically, QS exerts its therapeutic action by targeting downstream pyroptotic execution events — including GSDMD-mediated plasma membrane pore formation and Caspase-1 activation — while MCC950 inhibits upstream NLRP3 inflammasome activation. This combination creates a comprehensive cascade blockade throughout the pyroptotic pathway. These pivotal findings not only precisely delineate QS’s primary therapeutic targets within the pyroptosis pathway localized to downstream execution phases, but more importantly, establish a robust theoretical and experimental foundation for developing innovative combination therapies based on coordinated “upstream suppression coupled with downstream interception”.
Conclusion
The NLRP3 inflammasome serves as a pivotal therapeutic target for chronic inflammatory diseases.54 This study investigated NLRP3-related mechanisms to elucidate the psoriasis-pyroptosis relationship and systematically clarify QS’s multi-target regulation of keratinocyte pyroptosis. Results indicate QS exerts therapeutic effects primarily by preserving mitochondrial function, inhibiting GSDMD-mediated membrane pore formation, and downregulating Caspase-1 activity to block pyroptosis execution, though its direct inhibition of upstream NLRP3 remains limited. Significantly, QS demonstrates remarkable synergy with MCC950 (NLRP3 inhibitor), enhancing efficacy while enabling dosage reduction and minimized side effects. These findings substantiate QS as a promising psoriasis therapeutic that alleviates disease-related systemic impairment without drug-induced toxicity. Building on its defined role in blocking downstream pyroptosis, future investigations should assess QS’s potential to concurrently target upstream regulatory events (eg, mitochondrial ROS, mtDNA release) and non-canonical pathways, which could explain its enhanced efficacy in combination therapy.
Abbreviations
AHR, Aryl hydrocarbon receptor; AOD, Average optical density; ATP, Adenosine triphosphate; DAMPs, Damage-associated molecular patterns; HE, Hematoxylin-Eosin staining method; IHC, Immunohistochemistry assay; IL-17, Interleukin-17; IL-18, Interleukin-18; IL-1β, Interleukin-1β; IL-22, Interleukin-22; IMQ, Imiquimod cream; IOD, Integrated optical density; LPS, Lipopolysaccharide; NF-κB, Nuclear factor-kappa B; NLRP3, NOD-like receptor protein 3; PAMPs, Pathogen-associated molecular patterns; PASI, Psoriasis Area and severity index; QS, Qingshi anti-itch Ointment; TEM, Transmission electron microscopy.
Data Sharing Statement
All relevant data are contained within the article.
Ethics Approval and Consent to Participate
Animal experiments were approved by the Ethics Committee of the Animal House of Beijing University of Chinese Medicine (approval number: BUCM-2023100703-4007). The care and use of animals were conducted according to the ethical guidelines stipulated by the Chinese National Standard GB/T 35892-2018.
Acknowledgments
The authors would like to thank all the laboratory staff and assistants for their technical assistance.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Beijing’s “14th Five-Year Plan” Key Specialty Project in Traditional Chinese Medicine (BJZKBC0015); the Beijing Natural Science Foundation Project (7232295).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Strowig T, Henao-Mejia J, Elinav E, et al. Inflammasomes in health and disease. Nature. 2012;481(7381):278–19. doi:10.1038/nature10759
2. Zhang C, Xiao C, Dang E, et al. CD100-plexin-B2 promotes the inflammation in psoriasis by activating NF-κB and the inflammasome in keratinocytes. J Invest Dermatol. 2018;138(2):375–383. doi:10.1016/j.jid.2017.09.005
3. Dan D, Srivastava N. Psoriasis: striving for potential biomarkers. Assay Drug Dev Technol. 2023;21(6):235–257. doi:10.1089/adt.2023.014
4. Vieyra-Garcia PA, Wolf P. A deep dive into UV-based phototherapy: mechanisms of action and emerging molecular targets in inflammation and cancer. Pharmacol Ther. 2021;222:107784. doi:10.1016/j.pharmthera.2020.107784
5. Zhou X, Chen Y, Cui L, et al. Advances in the pathogenesis of psoriasis: from keratinocyte perspective. Cell Death Dis. 2022;13(1):81. doi:10.1038/s41419-022-04523-3
6. Mizutani H, Black R, Kupper TS. Human keratinocytes produce but do not process pro-interleukin-1 (IL-1) beta. Different strategies of IL-1 production and processing in monocytes and keratinocytes. J Clin Invest. 1991;87(3):1066–1071. doi:10.1172/JCI115067
7. Eckhart L, Tschachler E. Control of cell death-associated danger signals during cornification prevents autoinflammation of the skin. Exp Dermatol. 2018;27(8):884–891. doi:10.1111/exd.13700
8. Burdette BE, Esparza AN, Zhu H, et al. Gasdermin D in pyroptosis. Acta Pharm Sin B. 2021;11(9):2768–2782. doi:10.1016/j.apsb.2021.02.006
9. Zhou C, Zheng J, Fan Y, et al. TI: NLRP3 inflammasome-dependent pyroptosis in CNS trauma: a potential therapeutic target. Front Cell Dev Biol. 2022;10:821225. doi:10.3389/fcell.2022.821225
10. Schroder K, Zhou R, Tschopp J. The NLRP3 inflammasome: a sensor for metabolic danger? Science. 2010;327(5963):296–300. doi:10.1126/science.1184003
11. Coll RC, Robertson AA, Chae JJ, et al. A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nat Med. 2015;21(3):248–255. doi:10.1038/nm.3806
12. Coll RC, Hill JR, Day CJ, et al. MCC950 directly targets the NLRP3 ATP-hydrolysis motif for inflammasome inhibition. Nat Chem Biol. 2019;15(6):556–559. doi:10.1038/s41589-019-0277-7
13. Tapia-Abellán A, Angosto-Bazarra D, Martínez-Banaclocha H, et al. MCC950 closes the active conformation of NLRP3 to an inactive state. Nat Chem Biol. 2019;15(6):560–564. doi:10.1038/s41589-019-0278-6
14. Griffiths CEM, Armstrong AW, Gudjonsson JE, et al. Psoriasis. Lancet. 2021;397(10281):1301–1315. doi:10.1016/S0140-6736(20)32549-6
15. Boutet MA, Nerviani A, Gallo Afflitto G, et al. Role of the IL-23/IL-17 axis in psoriasis and psoriatic arthritis: the clinical importance of its divergence in skin and joints. Int J Mol Sci. 2018;19(2):530. doi:10.3390/ijms19020530
16. Krueger J, Puig L, Thaçi D. Treatment options and goals for patients with generalized pustular psoriasis. Am J Clin Dermatol. 2022;23(Suppl 1):51–64. doi:10.1007/s40257-021-00658-9
17. Palakornkitti P, Nimmannitya K, Rattanakaemakorn P. Biological therapy in psoriasis: an emphasis on its dermatologic adverse events. Asian Pac J Allergy Immunol. 2021;39(4):215–230. doi:10.12932/AP-110521-1129
18. Lee CL, Wang CM, Hu HC, et al. Indole alkaloids indigodoles A-C from aerial parts of strobilanthes cusia in the traditional Chinese medicine Qing Dai have anti-IL-17 properties. Phytochemistry. 2019;162:39–46. doi:10.1016/j.phytochem.2019.02.016
19. Cheng HM, Kuo YZ, Chang CY, et al. The anti-TH17 polarization effect of Indigo naturalis and tryptanthrin by differentially inhibiting cytokine expression. J Ethnopharmacol. 2020;255:112760. doi:10.1016/j.jep.2020.112760
20. Chang H-N, Huang S-T, Yeh Y-C, et al. Indigo naturalis and its component tryptanthrin exert anti-angiogenic effect by arresting cell cycle and inhibiting Akt and FAK signaling in human vascular endothelial cells. J Ethnopharmacol. 2015;174:474–481. doi:10.1016/j.jep.2015.08.050
21. Gu S, Xue Y, Gao Y, et al. Mechanisms of indigo naturalis on treating ulcerative colitis explored by GEO gene chips combined with network pharmacology and molecular docking. Sci Rep. 2020;10(1):15204. doi:10.1038/s41598-020-71030-w
22. Huai W, Zhao R, Song H, et al. Aryl hydrocarbon receptor negatively regulates NLRP3 inflammasome activity by inhibiting NLRP3 transcription. Nat Commun. 2014;5(1):4738. doi:10.1038/ncomms5738
23. Jiang YJ, Zhang FC, Cai LL, et al. A study on the quality standard of Qingshi antipruritic ointment. World TCM. 2018;13(11):2672–2676+2681.
24. Guo J, Liu Y, Guo X, et al. Depressive-like behaviors in mice with Imiquimod-induced psoriasis. Int Immunopharmacol. 2020;89(Pt B):107057. doi:10.1016/j.intimp.2020.107057
25. Song X, Chen G, Li C, et al. Tadalafil alleviates LPS-induced inflammation and oxidative stress of RWPE-1 cell by regulating the Akt/Nrf2 signaling pathway. Inflammation. 2021;44(3):890–898. doi:10.1007/s10753-020-01384-w
26. Liu Y, Zhou J, Luo Y, et al. Honokiol alleviates LPS-induced acute lung injury by inhibiting NLRP3 inflammasome-mediated pyroptosis via Nrf2 activation in vitro and in vivo. Chin Med. 2021;16(1):127. doi:10.1186/s13020-021-00541-z
27. Li Y, Song W, Tong Y, et al. Isoliquiritin ameliorates depression by suppressing NLRP3-mediated pyroptosis via miRNA-27a/SYK/NF-κB axis. J Neuroinflammation. 2021;18(1):1. doi:10.1186/s12974-020-02040-8
28. Liang Q, Cai W, Zhao Y, et al. Lycorine ameliorates bleomycin-induced pulmonary fibrosis via inhibiting NLRP3 inflammasome activation and pyroptosis. Pharmacol Res. 2020;158:104884. doi:10.1016/j.phrs.2020.104884
29. Zhang C, Tang B, Zheng X, et al. Analysis of the potential pyroptosis mechanism in psoriasis and experimental validation of NLRP3 in vitro and in vivo. Int Immunopharmacol. 2023;124(Pt A):110811. doi:10.1016/j.intimp.2023.110811
30. Chen XY, Xu F, Chen JQ, et al. UBE2L3 reduces TRIM21 expression and IL-1β secretion in epidermal keratinocytes and improves psoriasis-like skin. J Invest Dermatol. 2023;143(5):822–831.e4. doi:10.1016/j.jid.2022.10.016
31. Li X, Xi B, Miao Y, et al. Nintedanib ameliorates imiquimod-induced psoriasis in mice by inhibiting NF-κB and VEGFR2 signaling. Int Immunopharmacol. 2021;100:108129. doi:10.1016/j.intimp.2021.108129
32. Tu C, Wang S, Hu X, et al. Lipopolysaccharide induces TREM-1-dependent HIF-1α expression in human keratinocyte cell line. Cell Biol Int. 2016;40(12):1357–1365. doi:10.1002/cbin.10693
33. Elliott EI, Sutterwala FS. Initiation and perpetuation of NLRP3 inflammasome activation and assembly. Immunol Rev. 2015;265(1):35–52. doi:10.1111/imr.12286
34. Su F, Xia Y, Huang M, et al. Expression of NLPR3 in psoriasis is associated with enhancement of interleukin-1β and caspase-1. Med Sci Monit. 2018;24:7909–7913. doi:10.12659/MSM.911347
35. Ma H, Shu Q, Li Z, et al. Formaldehyde aggravates allergic contact dermatitis by facilitating NLRP3 inflammasome activation in macrophages. Int Immunopharmacol. 2023;117:109904. doi:10.1016/j.intimp.2023.109904
36. Griffiths CEM, Van der Walt JM, Ashcroft DM, et al. The global state of psoriasis disease epidemiology: a workshop report. Br J Dermatol. 2017;177(1):e4–e7. doi:10.1111/bjd.15610
37. Rendon A, Schäkel K. Psoriasis pathogenesis and treatment. Int J Mol Sci. 2019;20(6):1475. doi:10.3390/ijms20061475
38. Kong X, Gao M, Liu Y, et al. GSDMD-miR-223-NLRP3 axis involved in B(a)P-induced inflammatory injury of alveolar epithelial cells. Ecotoxicol Environ Saf. 2022;232:113286. doi:10.1016/j.ecoenv.2022.113286
39. Lachner J, Mlitz V, Tschachler E, et al. Epidermal cornification is preceded by the expression of a keratinocyte-specific set of pyroptosis-related genes. Sci Rep. 2017;7(1):17446. doi:10.1038/s41598-017-17782-4
40. Van der Fits L, Mourits S, Voerman JS, et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J Immunol. 2009;182(9):5836–5845. doi:10.4049/jimmunol.0802999
41. Wang WM, Jin HZ. Biologics in the treatment of pustular psoriasis. Expert Opin Drug Saf. 2020;19(8):969–980. doi:10.1080/14740338.2020.1785427
42. Amatore F, Villani AP, Tauber M, et al. French guidelines on the use of systemic treatments for moderate-to-severe psoriasis in adults. J Eur Acad Dermatol Venereol. 2019;33(3):464–483. doi:10.1111/jdv.15340
43. Cai Y, Xue F, Quan C, et al. A critical role of the IL-1β-IL-1R signaling pathway in skin inflammation and psoriasis pathogenesis. J Invest Dermatol. 2019;139(1):146–156. doi:10.1016/j.jid.2018.07.025
44. Zhu L, Han Z, He Y, et al. Caspase-1-dependent pyroptosis mediates adjuvant activity of platycodin D as an adjuvant for intramuscular vaccines. Cells. 2022;11(1):134. doi:10.3390/cells11010134
45. Sborgi L, Rühl S, Mulvihill E, et al. GSDMD membrane pore formation constitutes the mechanism of pyroptotic cell death. EMBO J. 2016;35(16):1766–1778. doi:10.15252/embj.201694696
46. Liu X, Zhang Z, Ruan J, et al. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature. 2016;535(7610):153–158. doi:10.1038/nature18629
47. He WT, Wan H, Hu L, et al. Gasdermin D is an executor of pyroptosis and required for interleukin-1β secretion. Cell Res. 2015;25(12):1285–1298. doi:10.1038/cr.2015.139
48. Ciążyńska M, Olejniczak-Staruch I, Sobolewska-Sztychny D, et al. The role of NLRP1, NLRP3, and AIM2 inflammasomes in psoriasis: review. Int J Mol Sci. 2021;22(11):5898. doi:10.3390/ijms22115898
49. Sreejit G, Nooti SK, Jaggers RM, et al. Retention of the NLRP3 inflammasome-primed neutrophils in the bone marrow is essential for myocardial infarction-induced granulopoiesis. Circulation. 2022;145(1):31–44. doi:10.1161/CIRCULATIONAHA.121.056019
50. Kwak SB, Koppula S, In EJ, et al. Artemisia extract suppresses NLRP3 and AIM2 inflammasome activation by inhibition of ASC phosphorylation. Mediators Inflamm. 2018;2018:6054069. doi:10.1155/2018/6054069
51. Rossol M, Pierer M, Raulien N, et al. Extracellular Ca2+ is a danger signal activating the NLRP3 inflammasome through G protein-coupled calcium sensing receptors. Nat Commun. 2012;3(1):1329. doi:10.1038/ncomms2339
52. Katsnelson MA, Rucker LG, Russo HM, et al. K+ efflux agonists induce NLRP3 inflammasome activation independently of Ca2+ signaling. J Immunol. 2015;194(8):3937–3952. doi:10.4049/jimmunol.1402658
53. Lawlor KE, Vince JE. Ambiguities in NLRP3 inflammasome regulation: is there a role for mitochondria? Biochim Biophys Acta. 2014;1840(4):1433–1440. doi:10.1016/j.bbagen.2013.08.014
54. Mangan MSJ, Olhava EJ, Roush WR, et al. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat Rev Drug Discov. 2018;17(9):688. doi:10.1038/nrd.2018.149
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Extracellular Histones Activate Endothelial NLRP3 Inflammasome and are Associated with a Severe Sepsis Phenotype

Beltrán-García J, Osca-Verdegal R, Pérez-Cremades D, Novella S, Hermenegildo C, Pallardó FV, García-Giménez JL
Journal of Inflammation Research 2022, 15:4217-4238
Published Date: 25 July 2022
Pyroptosis and Intervertebral Disc Degeneration: Mechanistic Insights and Therapeutic Implications
Ge Y, Chen Y, Guo C, Luo H, Fu F, Ji W, Wu C, Ruan H
Journal of Inflammation Research 2022, 15:5857-5871
Published Date: 17 October 2022
The Role of NLRP3 Inflammasome Signaling on Arrhythmias in Diabetes
Zhang L, Liu HH, Li F, Yang F, Qian LL, Wang RX
Journal of Inflammation Research 2022, 15:6883-6889
Published Date: 29 December 2022
The Mechanism of Pyroptosis and Its Application Prospect in Diabetic Wound Healing
Al Mamun A, Shao C, Geng P, Wang S, Xiao J
Journal of Inflammation Research 2024, 17:1481-1501
Published Date: 6 March 2024
NLRP3 Inflammasome Targeting Offers a Novel Therapeutic Paradigm for Sepsis-Induced Myocardial Injury
Jin Y, Fleishman JS, Ma Y, Jing X, Guo Q, Shang W, Wang H
Drug Design, Development and Therapy 2025, 19:1025-1041
Published Date: 14 February 2025